 Pauline Ducouret
Pauline Ducouret Andrea Romano
Andrea Romano Amélie N. Dreiss1
Amélie N. Dreiss1 Patrick Marmaroli
Patrick Marmaroli Xavier Falourd
Xavier Falourd Alexandre Roulin
Alexandre Roulin- 1Department of Ecology and Evolution, University of Lausanne, Lausanne, Switzerland
- 2PRONA, Yverdon-les-Bains, Switzerland
To resolve conflicts over limited resources, animals often communicate about their motivation to compete. When signals are transient, the resolution of conflicts may be achieved after an interactive process, with each contestant adjusting its signaling level according to the rival's behavior. Unfortunately, the importance of the real-time signal adjustment in conflict resolution remains understudied, especially using experimental approaches. Here we developed a novel “automatic interactive playback” that interacts real-time with a live individual. It allowed us to experimentally test the efficacy of different behavioral strategies to dominate conflicts in nestling barn owls (Tyto alba). In this species, nestlings vocally negotiate for priority access to the impending food item in the absence of parents. Two opposite vocal strategies were tested for their prospects of success: under the “matching” vs. “mismatching” strategy, the playback behaves in the same vs. opposed way as the nestling, respectively. We evaluated how these two strategies affected the two main negotiation parameters: call duration and call rate. We found that the best strategies to reduce the nestling's vocalizations and hence dominate the negotiation are to match the call duration of the opponent and to mismatch its call rate. However, the latter strategy is the only one that allowed the playback to dominate the vocal interaction by inducing the opponent to become totally silent. Therefore, to prevail in a negotiation session, barn owl nestlings should delay the transmission of signals rather than simultaneously escalate vocalizations as commonly observed in animal competitive interactions. In addition, we showed that matching call duration and mismatching call rate require a larger investment by the playback, in terms of number and duration of calls, than the less effective strategies. Assuming that vocalizations are costly, this suggests that such behavioral strategies are honest. Our results highlight the importance of real-time signaling adjustment in communication processes over resource competition and emphasize the power of using interactive playback settings to investigate conflict resolution in animals.
Introduction
In nature, conspecifics compete over limited resources, such as territories, mates, or food (McGregor, 2005) by either fighting or communicating (Parker, 1974; Maynard Smith, 1982). Such communication processes may involve “negotiation” which is defined as two or more individuals trying to reach an agreement about how to share contended resources or how to invest in collaborative activities (Johnstone and Roulin, 2003; Johnstone and Hinde, 2006; Hinde and Kilner, 2007; Sirot, 2012). The central goal of negotiation is thus to limit aggressive behaviors, which may lead to serious or lethal injuries. Although such a term has been mostly used in the context of humans that bargain for resources (Binmore, 2010), this concept has been applied to animals especially in a context of long-term relationship between opponents (McNamara et al., 1999; Johnstone and Roulin, 2003; Johnstone and Hinde, 2006; Patricelli et al., 2011; Sirot, 2012). While humans use various negotiation tactics by modulating gestures, words, and voice, it is not fully clear whether this process is common in animals and how it exactly occurs (Pika and Frohlich, 2019). However, it is well-known that animals use different signals, often in a specific sequence (Searcy and Beecher, 2009; Akcay et al., 2013), to inform rivals about their resource holding potential (i.e., fighting ability) and motivation to compete. Such information is crucial for an individual to adjust its effort in competitive interactions, and thus to decide whether to engage in, keep competing or retreat from a contest according to its chance of success (Parker, 1974; Maynard Smith, 1982; Enquist et al., 1990; Briffa et al., 1998). When interests among the competitors are incompatible, like non-familiar adults competing for limiting resources such as mates or territories, an escalation in competitive behaviors is expected until the weakest or the less motivated individual withdraws from the contest (e.g., Keil and Watson, 2010; Reddon et al., 2011). However, competition also occurs among individuals that partly share interest in each other's fitness (Roberts, 2005). Indeed, when contestants are genetically related, losing a contest in favor of a kin still rewards the looser with indirect fitness benefits (Hamilton, 1964). Moreover, when living in stable social groups, individuals repeatedly interact and benefit from groupmates' survival. In such cases, for the same given amount of resources, a de-escalation in competitive interactions could be instead expected (Johnstone and Roulin, 2003).
A key element of negotiation is the use of transient signals for which several parameters (e.g., number, duration) can quickly vary irrespectively of a change in individual condition (Greenfield et al., 1997; Briffa et al., 1998), and can be fine-tuned according to signals previously emitted by the opponent(s) (Enquist and Leimar, 1983; Enquist et al., 1990; Payne and Pagel, 1996; Briffa et al., 1998). The resolution of conflicts through negotiation is thus an interactive process achieved after repeated interactions which leads to a progressive variation in the signaling level. Under such a scenario, a real-time adjustment in signaling level may be as important as the average/maximum signal strength to outcompete a rival (Payne and Pagel, 1997; Briffa et al., 1998; McNamara et al., 1999; Patricelli et al., 2002; Van Dyk et al., 2007; Dreiss et al., 2015). Unfortunately to date, no experimental study has investigated the importance of real-time adjustment strategies for conflict resolution through vocal signaling.
In this study, we aimed at identifying which vocal adjustment strategies best dominate an opponent during a negotiation session, and therefore prevail in competition for food, in nestling barn owls (Tyto alba). Although possessing well-designed weapons, sharp claws, and bills, young barn owls behave surprisingly peacefully. Siblings frequently preen and feed each other (Roulin et al., 2016) and they socially huddle to keep warm (Dreiss et al., 2016). Most remarkably, while waiting for an indivisible prey brought by parents to the nest, siblings vocally negotiate to decide which individual will obtain the impending food item without engaging in physical fights (Roulin et al., 2000; Johnstone and Roulin, 2003). By emitting many long calls, nestlings demonstrate that they are hungry, which deters their less needy siblings from negotiating and begging once parents return to the nest with food (Roulin et al., 2009; Dreiss et al., 2010b; Ruppli et al., 2013a). Importantly, in barn owl broods, negotiation invariably occurs before the arrival of each prey, and it is intimately linked with parental food allocation, with the vocal dominant nestling having the largest probability to receive the impending prey, as repeatedly shown by previous studies (Roulin, 2002; Roulin et al., 2009; Dreiss et al., 2010b). The less needy individuals withdraw from the competition and invest in negotiation only once the likelihood to monopolize a prey increases.
The negotiation in barn owl nestlings is an interactive process through progressive step-like series of variations in call parameters ending when an individual becomes vocally dominant by emitting more and longer calls or even silencing the opponents. Barn owl nestlings are in fact able to assess opponents' acoustic changes at a fine temporal scale and use this information both to decide when it will resume calling in a negotiation session and how it fine-tunes its signal level according to sibling's signal level (Dreiss et al., 2014, 2015). Whenever the temporary vocally dominant nestling progressively emits shorter calls at a lower rate, silent siblings attempt to take the floor (Dreiss et al., 2015). In addition, when siblings are exchanging calls, their call duration and call rate are, respectively, positively and negatively adjusted (Dreiss et al., 2014, 2015). Thus, during a negotiation session nestlings tend to match siblings' change in call duration and mismatch siblings' change in call rate. We hence predicted that these strategies are most effective to dominate the negotiation by inducing a sibling to progressively emit shorter calls at a lower rate. These strategies could be considered as signals by themselves and should hence entail costs in order to prevent dishonesty and be evolutionary stable (Zahavi, 1974; Grafen, 1990; Maynard Smith and Harper, 2003; Searcy and Nowicki, 2005). If matching duration and mismatching call rate are more effective to deter a sibling from competing, these strategies should be costly by inducing individuals to produce more and longer calls than by using a less successful strategy.
In order to experimentally test these two predictions, we developed a novel “automated interactive playback” that interacts with a live nestling. A computer records the calls emitted by the nestling and instantly measures call rate and call duration. Then it immediately starts to broadcast vocalizations depending on the animal's fine-tuning signal level (i.e., call rate and call duration), simulating a barn owl nestling that follows one of two negotiation vocal strategies: under the “matching” strategy, the playback behaves in the same way as the nestling, and under the “mismatching” strategy, the playback responses in the reverse way as the nestling. For example, if the nestling increases call rate or call duration, the playback emits respectively more or longer calls under the matching strategy vs. fewer or shorter calls under the mismatching strategy. The effects of real time vocal adjustment were tested in two distinct experiments: one testing the adjustment of the call rate while the call duration was kept constant and the other one the call duration while the call rate was kept constant. Finally, this setting allowed us to determine whether the most effective strategies to prevail in negotiation are also the costliest in the sense that the playback emits more and/or longer calls.
Methods
General Procedures
The study was performed on a barn owl population breeding in western Switzerland (46°4′N, 6°5′E). Between April and September 2015, 114 nestlings (52 males, 57 females, and 5 of unknown sex) from 41 broods were brought to the laboratory for 3 days and 2 nights (mean ± SE age: 34 ± 0.5 days, range: 22–41 days). We already showed on several occasions that nestlings vocalize in captivity as in the lab and in natural conditions. In particular, they are not physiologically stressed (Dreiss et al., 2010a), and similarly vocalize in captivity conditions as in the wild (Roulin et al., 2009; Dreiss et al., 2010b). The first night was an acclimation night during which siblings stayed together in a wooden nest-box identical to the one where they were raised. They were fed ad libitum (67 g of mice per nestling, Durant and Handrich, 1998). At 08:00 the next morning, the remaining food was removed. Then, at 12:00 all nestlings were isolated in separated experimental nest-boxes in order to allow nestlings to get used to the new environment before 22:00, when the experiment started. The “call duration” and the “call rate” interactive playback experiments were performed respectively between 22:00 and 01:00, and between 1:30 and 4:30. On the following morning, nestlings were fed and brought back to their original nest.
The experimental nest-box was divided into two equal parts by a wooden wall pierced with five holes. A loudspeaker was placed in one of the partitions, while the nestling occupied the other. Two microphones per nest-box were fixed on the roof and were oriented toward the nestling. One microphone was connected to a pre-amplifier PreSonusDigimax FS and a computer with the interactive playback algorithm developed in Matlab R2012b 8.0.0.783 (MathWorks. Natick, MA, U.S.A.). To increase the computing capacity, the recorded calls were deleted as soon as analyzed. This is why we used a second microphone to record all calls produced during the experiments. This microphone was connected to a pre-amplifier Steinberg UR44 and to a second computer.
Playback Experiment Design
The program detected in real time the calls emitted by the nestling and determined their duration (for detail on acoustic criteria used see Supplementary Material S1 and Ducouret et al., 2016). Two adjustment strategies were programmed and the playback followed a unique strategy randomly selected during 15-min period (hereafter “period”), before changing to the other one. Each strategy was repeated four times per experiment.
Playback Call Rate Experiment
In order to assess the variation of the call rate of focal nestlings, at the end of each 10 s lapse, the computer program compared the number of calls emitted by the nestling with the number of calls emitted during the previous 10 s lapse. The 10 s duration of the time lapse was chosen because the mean call rate observed in two-nestling broods when food-deprived is 6 calls/min, hence on average 1 call every 10 s, although around 50% of the 1-min interval of free vocal interactions contains more than 6 calls (Ruppli et al., 2013a). The call rate measured by the algorithm was hence in number of calls per 10 s. During the first 10 s lapse of each 15-min period, the playback emitted the same number of calls as the nestling during the first 10 s lapse. Next, according to the two successive 10 s lapses comparison of number of calls, the playback modified its number of broadcasted calls following two different playback strategies (Figure 1, for an example see Supplementary Material S2). In the matching strategy (hereafter “Match-Call Rate”), if the nestling emitted a number equal to “X” more (or respectively fewer) calls than during the previous 10 s lapse, the playback broadcasted X more (or respectively fewer) calls than during the previous 10 s lapse. In the mismatching strategy (hereafter “Mismatch-Call Rate”), if the nestling emitted X more (or respectively fewer) calls than during the previous 10 s lapse, the playback broadcasted X less (or respectively more) calls than during the previous 10 s lapse. In both cases, playback calls were equally distributed along the next 10 s lapse (e.g., if the playback had to emit one call, it was broadcasted 5 s later; in case of 2 calls, the playback broadcasted them 3.3 and 6.6 s later). In case the number of calls comparison between the two consecutive 10 s lapses would have led the playback to broadcast a negative number of calls, the algorithm reset the playback number of calls to 0 calls. We fixed the broadcast call duration at 800 ms on average (SE = 50 ms), which was the average value obtained from interacting pairs of nestlings (Ruppli et al., 2013a).
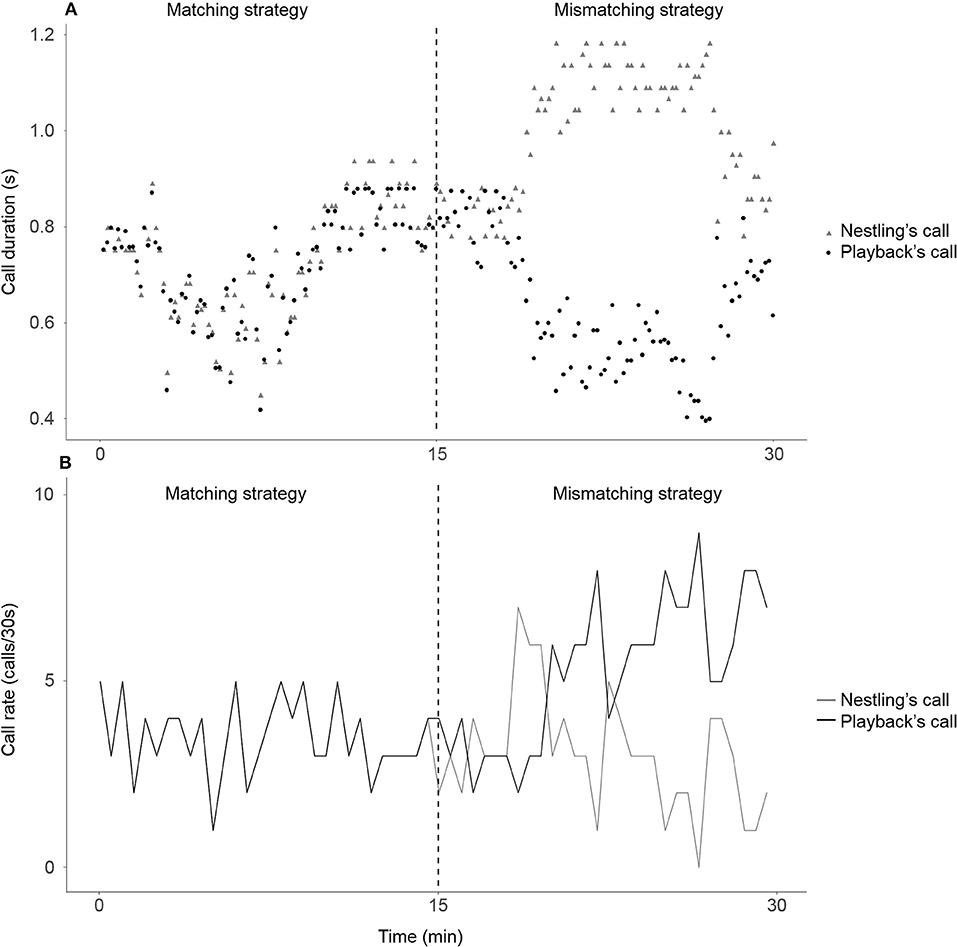
Figure 1. An example of two periods of interactive playback experiment on call duration (A) and on call rate (B). The computer program automatically detects the calls produced by a nestling and measures its duration. Then, the program compares this duration (or respectively rate) to that recorded in the previous 10 s period. According to this change, it chooses to broadcast a call of a duration (or respectively rate) based on one of two pre-programmed calling strategies: under the matching strategy (left panels), the playback changes its call duration (or respectively rate) similarly to the nestling, while under the mismatching strategy (right panels), changes its call duration (or respectively rate) reversely to the nestling. The strategies were repeated four times each for 15 min and were randomly ordered across the experiment.
Playback Call Duration Experiment
The second experiment was developed with the same approach as the one described above but focusing on the mean nestling call duration (Figure 1, for an example of calculation see Supplementary Material S3). At the end of the first 10 s lapse of each 15-min period, the playback emitted a call with the same duration as the nestling's mean call duration. Afterward, in the matching strategy (hereafter “Match-Call Duration”), the playback modified its call duration in the same way as the sibling. In the mismatching strategy (hereafter “Mismatch-Call Duration”), the playback modified its call duration in the opposite way as the sibling. The playback always broadcasted one call every 10 s so the playback call rate was 6 calls/min corresponding to the mean call rate observed in two-nestling broods when food deprived (Ruppli et al., 2013a). In case the nestling did not emit any call during the 10 s lapse, no comparison of call duration could be made with the call duration measured during the previous 10 s lapse. However, the playback had to emit a call to keep its call rate constant. Therefore, the playback's call duration remained constant which is the most parsimonious solution.
Construction of Playback Soundtracks
Calls broadcasted by the playback were isolated from four different individuals (age mean ± SE: 32.5 ± 2.25 days, two males and two females) recorded in three-nestling brood experiments conducted in 2011 (for experimental setup details see Dreiss et al., 2017). At the beginning of the experiment, a single individual was selected randomly by the computer to be broadcasted to one focal nestling. Therefore, a focal nestling faced a unique playback individual. In total, 120 calls were isolated, 15 in each eight call-duration groups: 300–400, 400–500, 500–600, 600–700, 700–800, 750–850, 800–900, 900–1,000, and 1,000–1,100 ms. The computer randomly picked up one call within the relevant group. For the call rate experiment, only calls from the 750–850 ms group were picked up by the playback. Each call was first normalized to have the same loudness using Matlab R2012b.
Acoustic Analyses
In order to detect a call emitted by the nestling in the pseudo real-time, the audio record was analyzed each 46 ms time windows. The choice of this time frame was motivated by a trade-off between reactivity (we wanted to know as fast as possible if an acoustic event occurred) and reliability (the longer the signal is, the better the frequency resolution is and in turn the more reliable the detection is). Here, 46 ms was empirically found to be a good candidate for combining both reliability and reactivity. The algorithm determined first if there was an acoustic event or not by calculating the sound level (dB) and compared it to a threshold empirically determined. Then, to determine if this call was a negotiation call, we used two frequency descriptors and a temporal descriptor. For further technical details see Ducouret et al. (2016) and Supplementary Material S1. We tested the accuracy of this detection in the real time by comparing 180 h of a recording including 55,247 negotiation calls of interacting pairs of nestlings recorded in similar condition with the same acoustic tools. We achieved an accuracy of 97% of true detection (i.e., in only 3% of cases the software wrongly identified another type of call or a noise as a negotiation call).
Statistical Analyses
In the Call Rate experiment, we removed from the analyses one individual which did not call during the experiment. For the Call Duration experiment, we removed from the analyses 55 nestlings which did not emit any call or called only during one 15-min period. This was done because, if the nestling remained silent, it was not possible to analyse any vocal interaction between nestling and playback. The absence of calls for many individuals in the call duration experiment may be explained by a playback call rate representing a more competitive individual than the tested nestling. Indeed, the playback call rate corresponded to a highly motivated nestling that was experimentally food-deprived (Ruppli et al., 2013a). Overall nestlings emitted on average 2.9 calls/min (range 0 to 31 calls/min) during the call rate experiment and emitted on average 4 calls/min (range 0 to 34 calls/min) during the call duration experiment. Nestlings emitted hence slightly fewer calls than during free vocal exchange in laboratory and nature (i.e., 6 calls/min) but with a similar range (Roulin, 2001a; Ruppli et al., 2013a). This might be explained by a playback representing a highly motivated individual.
Statistical analyses were performed using linear mixed models implemented with the “lme4” package (Bates et al., 2014) in the R software (R Core Team, 2013). Residuals were checked for homoscedasticity. In each model presented below, the order in which playback strategies appeared, the nestling's age and sex were included as covariates and cofactor. Nestling identity, nested in brood identity, was included as a random factor to control for data pseudoreplication and possible family effect. The identity of the nestling broadcasted by the playback was also included as a random factor to control for potential difference in the stimulatory capacity.
The effect on nestling's call parameters of the playback strategies (Match coded 0 and Mismatch coded 1) was tested by considering the mean call rate (i.e., the mean number of calls per min) and the mean call duration emitted by the nestling in each 15-min period for both experiments. We investigated the effect of the two strategies on both the call parameter which was adjusted by the playback (i.e., the nestling's call rate for the Call Rate experiment and the nestling's call duration for the Call Duration experiment) and the cross effect on the other call parameter (i.e., the call rate for the Call Duration experiment and reversely). We hence ran two independent but similar linear mixed models for the focal parameter and two other ones for the cross parameter for both experiments. Since call rate and call duration can be traded-off, we also investigated whether the playback strategies have different effects on the overall nestling call effort by calculating the duty cycle (product of call duration and call rate). The duty cycle measures the total amount of time spent calling by an individual and thus it is a reliable proxy of the overall call effort (e.g., Reichert and Gerhardt, 2012). The mean call rate and the duty cycle were Box-Cox transformed in order to approximate them as a Gaussian variables.
In addition, we assessed the efficacy of different playback strategies to induce the focal nestling to stop calling by analyzing the time needed to silence a nestling. Because of the high number of individuals that did not stop vocalizing, the distribution of the time needed to silence a nestling was neither Gaussian nor Poisson and no transformation was suitable. We hence performed the analysis in two steps. First, we investigated if the probability that a nestling retreated was different depending on the playback strategy using a generalized mixed model assuming a binomial distribution. We considered that a nestling had withdrawn (coded as 1) when it stopped emitting calls at least 1 min before the end of each 15-min period (similar results were obtained using thresholds of 30 s and 2 min; details not shown for brevity). Second, we analyzed whether the time needed to momentarily silence a nestling differed between playback strategies using a linear mixed model. This latter analysis only included periods when nestlings stopped calling.
We finally investigated the impact of following a particular strategy on the call parameters emitted by the playback. To this end, we considered the mean call rate (i.e., the mean number of calls per min) during the Call Rate experiment and the mean call duration during the Call Duration experiment emitted by the playback in each 15-min period. The playback's call rate was Box-Cox transformed in order to analyse it as Gaussian variable. We ran two independent but similar linear mixed models for the call rate and the call duration. Because one of the two playback call parameters was maintained constant, analyzing the duty cycle of the playback corresponds to the analysis of the focal call parameter multiplied by a constant. Hence, an increase of the playback call rate in the call rate experiment, and an increase of the playback call duration in the call duration experiment, leads inevitably to an increase of the duty cycle in both cases. Therefore, the analysis of the duty cycle of the playback is redundant.
Since analyzing the global mean value of call rate and call duration might not reflect the effect of the playback strategy on different stages of a period, we replicated the same analyses presented above by using the mean value of call parameters calculated on different time windows (each 1.5, 3, 5, and 7.5 min) rather than the global mean. Different windows were chosen because in natural conditions, a single negotiation session is of variable duration, and we could not isolate different negotiation cycles independently for each nestling. Models were exactly the same as described above with the exception that we added the window number as a random factor to account for data collected in the same part of each period. Results were always qualitatively similar. Thus, in the main text we only present the results with the global mean value of call parameters for brevity. Tables reporting the results of these collateral analyses can be found in Supplementary Material S4.
Ethical Notes
Experiments were done in the second part of the rearing period (mean nestling age: 34 ± 0.5 days; fledging age: ca. 55 days) when it did not disturb parental care as parents stay outside the nest and only enter briefly to bring food. At least two nestlings were left in their nest to still stimulate parental care and prevent brood abandonment, which was not observed during the experiment. It was already shown that this type of experiment neither stresses nestlings nor reduces nestling body condition at fledgling (Dreiss et al., 2010b). Experiments were carried out within University of Lausanne's facilities, under all required permits from the veterinary services (authorization 2109.2).
Results
Impact of Playback Strategies on Nestling's Call Parameters
Playback Call Rate Experiment
The most effective playback strategy for inducing the nestling to emit fewer calls was the Mismatch-Call Rate strategy, i.e., staying almost silent when the nestling is calling but increasing call rate when the opponent decreases the number of calls (Table 1A, Figure 2A, Figure S4). The Mismatch-Call Rate strategy also more often induced a nestling to become totally silent before the end of the 15-min period than the Matching-Call Rate strategy (Table 2A, mean probability of becoming silent was 0.68 ± 0.23 SE and 0.15 ± 0.23 SE for the Mismatch- and Match-Call Rate strategy, respectively). In addition, the Mismatch-Call Rate strategy took less time to induce the nestling to stop calling than the Match-Call Rate strategy (Table 2B, Figure 3). Nestlings that listened to a playback that mismatched call rate also emitted shorter calls than when they listened to a playback that matched call rate (Table 3). Finally, the Mismatch-Call Rate strategy induced a nestling to have a lower duty cycle (i.e., call effort) than the Match-Call Rate strategy (Table 4).
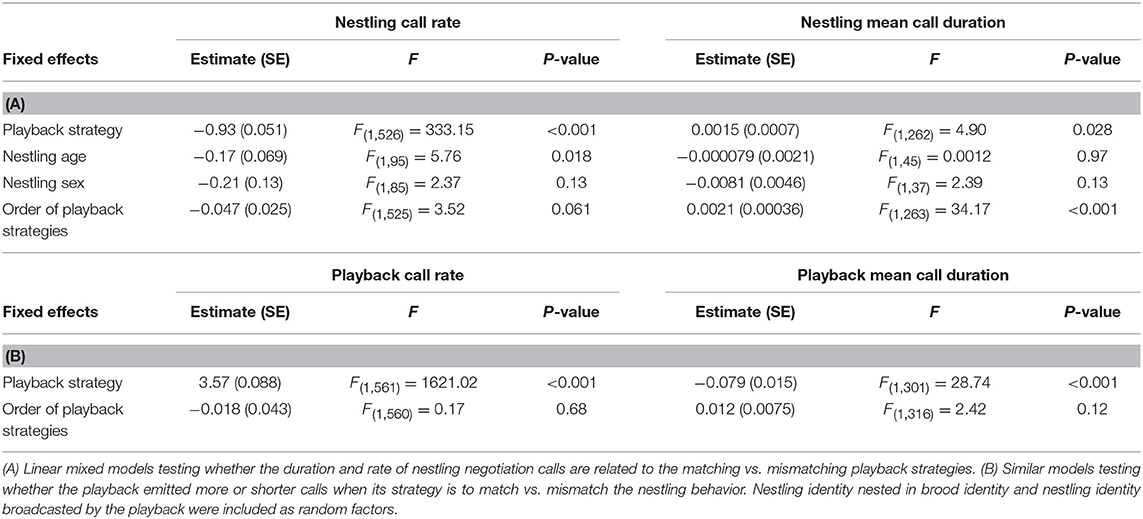
Table 1. Effect of the playback strategies on the vocal behavior of nestling barn owls (A) and of the playback (B).
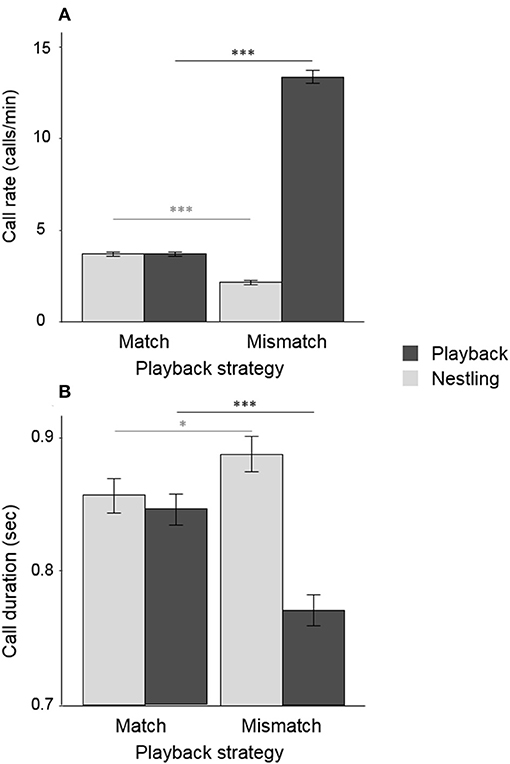
Figure 2. Mean call duration (A) and mean call rate (B) of nestlings and playback adopting the matching and mismatching strategies. The error bars represent the standard errors. Asterisks indicate significant differences between groups (*P < 0.05; ***P < 0.001), calculated from the models reported in Table 1. Each playback strategy was repeated for four 15-min periods, randomly ordered across the experiment.
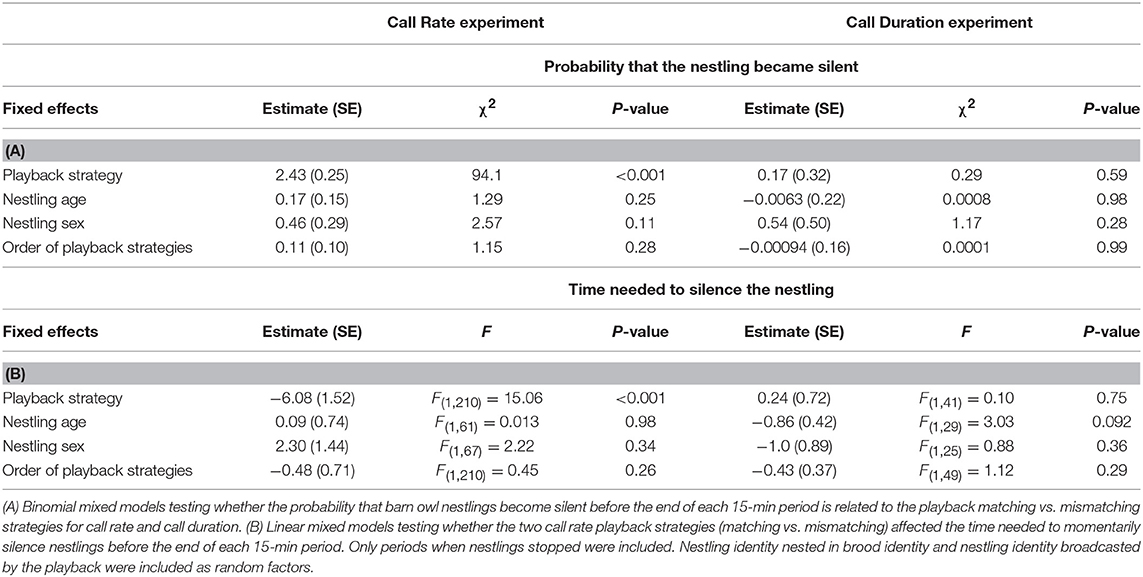
Table 2. Effect of playback strategies on the probability that nestlings became silent (A) and on the time needed to silence nestlings (B).
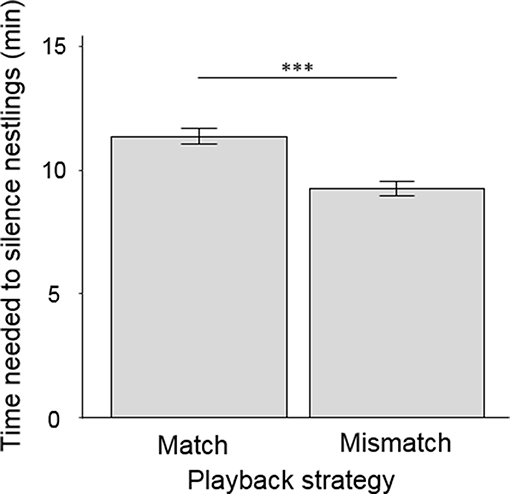
Figure 3. Mean time taken by the playback to silence nestlings depending on the call rate playback strategy (Match vs. Mismatch nestling Call Rate). Only 15-min periods when nestlings stopped calling is included (i.e., N = 63 for Matching and N = 150 for Mismatching strategy). The error bars represent the standard errors. Asterisks indicate significant differences between groups (***P < 0.001), calculated from the models reported in Table in Supplementary Material S5.
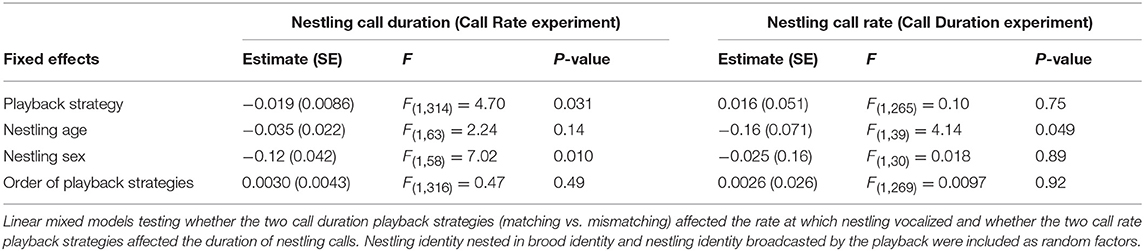
Table 3. Cross-effects of the playback strategies on the vocal behavior of nestling barn owls.
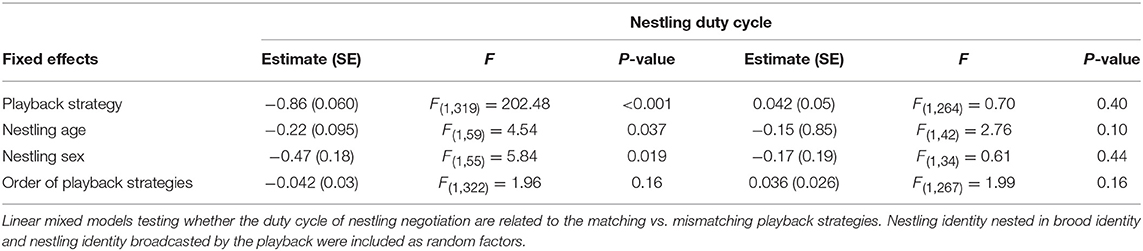
Table 4. Effect of the playback strategies on the vocal duty cycle of nestling barn owls.
Playback Call Duration Experiment
The most effective negotiation strategy to induce the nestling to reduce call duration was exactly the opposite as the one for call rate. Playbacks had to emit calls that matched rather than mismatched the duration of the nestling calls (Table 1A, Figure 2B, Figure S4). Match- and Mismatch-Call Duration strategies did not have different effects on nestling call rate and duty cycle (Tables 3, 4) and did not differ in the efficacy to momentarily silence nestlings (mean probability of becoming silent was 0.14 ± 0.34 SE and 0.17 ± 0.33 SE for the Match- and Mismatch-Call Duration strategy, respectively; Table 2A), and similar time needed (Table 2B). Therefore, a modulation in call duration only affects nestling call duration. The significant effect of playback strategy order indicates that nestlings produce longer calls with time, which reflects the increase of hunger level and was repeatedly found in previous studies (e.g., Ruppli et al., 2013a,b; Dreiss et al., 2017).
Impact of Playback Strategies on Playback's Call Parameters
Mismatch-Call Rate strategy led the playback to broadcast more calls than Match-Call Rate strategy (Table 1B, Figure 2A, Figure S4). Similarly, Match-Call Duration strategy led the playback to broadcast longer calls than Mismatch-Call Duration strategy (Table 1B, Figure 2B). By following the strategies that are the most effective to restrain a nestling from vocalizing (i.e., Match-Call Duration and Mismatch-Call Rate), the playback vocalized more intensely, emitting more calls during the Call Rate experiment and longer calls during the Call Duration experiment which resulted in an increase of the playback duty cycle.
We decided to keep the playback's call parameters constant when the nestling did not emit any call in the previous 10 s lapse. This choice does not fully reflect natural vocal exchanges, because nestlings usually decrease call duration and rate after having deterred a sibling from negotiating (Dreiss et al., 2015). The decision not to decrease playback's calls after the nestling was silenced could explain why the playback emitted so many calls under the Mismatch-Call Rate strategy. However, we note that if we restrict the analyses to the data recorded until the moment when the live nestling stopped calling (i.e., removing the calls emitted by the playback afterwards) the results were qualitatively similar (Supplementary Material S5).
Discussion
In the present experimental study, we first identified the most effective real-time vocal strategies to dominate social interactions in barn owl broods, and then we showed that the successful strategies impose more vocal investment to the sender (i.e., the playback). This was feasible by using a procedure similar to artificial intelligence, with a computer being programmed to “behave” depending on the behavior of a live animal that in turn listens to what the computer broadcasts. Using such an innovative approach, which was seldom used previously (see Goutte et al., 2010; King, 2015), we demonstrated that mismatching the opponent's call rate and matching its call duration lead to a de-escalation in the opponent's vocalization (shorter and fewer calls respectively), as already shown in natural conditions using a correlative approach (Dreiss et al., 2014, 2015). This de-escalation in vocalization is particularly strong in the Mismatch-Call Rate experiment, which leads also to an overall decrease in call effort (i.e., duty cycle). Therefore, by following these strategies during the sibling negotiation process, nestlings reach the dominant position, and consequently have higher chances to be fed at the next parental visit (Roulin, 2002; Roulin et al., 2009; Dreiss et al., 2010b), without provoking a vocal escalation (Roulin et al., 2009; Dreiss et al., 2010b; Ruppli et al., 2013a).
Interestingly, the best strategy to reduce opponent's call duration is exactly the opposite as the most effective strategy to reduce opponent's call rate. Call rate and call duration may thus have different functions in a negotiation process. On the one hand, call duration seems to be used as a challenging signal, as the most effective strategy is to match the opponent's behavior, possibly to test its willingness to engage in a vocal duel. On the other hand, call rate is used as a deterring signal, as the most effective strategy is to escalate but only when the opponent relaxes which is also the ultimate way to induce opponents to retreat from the contest. Indeed, the Mismatch-Call Rate is the most successful strategy in momentarily silencing siblings. Our findings therefore suggest that vocal negotiation in nestling barn owls might be a hierarchical signaling system, where different signals are used in sequence from the weakest to the strongest, as for example consistently observed in conflict resolution over territory in other bird species (Searcy and Beecher, 2009; Akcay et al., 2013). Although the Match-Call Duration strategy was more effective to induce a nestling to decrease its call duration than Mismatch-Call Duration strategy, this call duration decrease was not sufficient to induce a significant decrease of the overall call effort (i.e., duty cycle). Moreover, the effect of the playback call duration was weaker than the effect of the playback call rate on vocal parameters of the nestling, as already found in a previous study (Ruppli et al., 2013a). As in other biological systems where a matching strategy is typically used as a conventional signal of intent to compete (e.g., Akcay et al., 2013), nestling barn owls could initially challenge siblings by keeping a low call rate but by matching the duration of sibling calls, the weakest and potentially the least expensive signal. Only if this strategy is not sufficient to vocally dominate, individuals increase call rate and try to prevail in the negotiation. By gradually emitting different signals, individuals give the opponents the possibility to give up the contest at an early stage without having invested too much in a negotiation session (Akcay et al., 2013).
Another interesting finding of our study was the evidence that by following the most effective strategies, and mainly for Mismatch-Call Rate strategy, the playback gradually intensified its signal level. Such strategies therefore require a higher overall call effort (i.e., duty cycle) by the sender—number of calls and call duration—than the less effective ones. This result is coherent with the theory of honest signaling, postulating that signals conveying reliable information about the sender should impose a cost to prevent cheating and thus to be evolutionary stable (Zahavi, 1974; Grafen, 1990; Maynard Smith and Harper, 2003; Searcy and Nowicki, 2005). Admittedly, we have no information about the energetic costs of producing many long calls in our model system, and our results should be therefore considered with this caveat in mind. However, although the cost of begging is still a controversial issue (Leech and Leonard, 1996; Moreno-Rueda, 2007), several studies showed that begging vocalizations impose a variety of metabolic, immunological, and growth costs to nestling birds (e.g., McCarty, 1996; Kilner, 2001; Moreno-Rueda, 2010; Noguera et al., 2010). In addition, we note that demonstrating high motivation by following the most effective strategies requires not only a high investment in terms of vocalization production, and therefore a reduction in time devoted to other activities (Roulin, 2001b), but also a particular attention to opponents' fine-tune behavior. The signal processing and vigilance to an opponent's signal change is likely to entail additional costs (Benton et al., 1994; Moss et al., 1998).
By waiting until its opponents relax to increase their call rate, nestlings give the opportunity to a sibling to vocalize, thus favoring the exchange of information. This finding corroborates those of previous research showing that barn owl nestlings seek to improve the exchange of information by first avoiding sibling's call overlaps to limit signal interference (Dreiss et al., 2013; Ducouret et al., 2018) and second, by favoring alternation of monologs (Dreiss et al., 2015). Indeed, during free vocal interactions between two siblings, 67% of calls are transmitted in monologs (i.e., 10 calls produced by one nestling without being interrupted by a sibling; Dreiss et al., 2015). An intrinsic risk of this behavior is that a parent could arrive at the nest with food which should be consequently given to the most vocal sibling. However, we note that in the barn owl parental absence is unpredictable (on average 30 min, range: 30 s to 3 h 40; Roulin and Bersier, 2007) which differs from many bird species where parental feeding visits are regular and quick (see e.g., Wright and Leonard, 2002). Negotiation can therefore be prolonged, and nestlings may not be able to maintain a dominant position during the entire period of parental absence. In addition, barn owl nestlings negotiate with full siblings only (Roulin et al., 2004) and hence the food is consumed by a genetically related individual and provides inclusive fitness benefits to individuals that give up (Hamilton, 1964). Moreover, negotiation occurs every night before the arrival of each prey and involves the same participants during the entire rearing period. In a social group composed of relatives where time is not a restricted parameter, the lack of escalation could thus be beneficial because it limits the costs of the interactions and increases the trust between negotiators for future sessions, which could counterbalance the risk of missing a food item. Therefore, the best strategy is to listen and take the floor once the opponent becomes less vocal. This fits with the quote of François de Callieres “one of the most necessary qualities in good negotiator is to be an apt listener” (De Callieres, 1738).
In summary, we have pinpointed the importance of real-time signaling adjustment during the resolution of conflicts over indivisible food in barn owl broods by showing that the adjustment strategies to opponent's vocalizations are signals by themselves as they trigger different behavioral responses. More interestingly, although the most effective strategies to dominate the negotiation are different for call rate and duration, these opposite strategies impose a large investment in vocalizations by the sender, thus suggesting the honesty of the negotiation process. Finally, our study emphasizes the power of using interactive playback to uncover the importance of social strategies evolved to resolve conflicts of interest in animals.
Data Availability
The datasets supporting this article have been uploaded as part of the Supplementary Material.
Ethics Statement
The protocol was approved by, and the study carried out in accordance with, the recommendations of the veterinary service of the canton de Vaud (authorization 2109.2).
Author Contributions
PD, AD, and ARou conceived and designed the experiment. PD collected the data. PD, PM, and XF developed the algorithm of the interactive playbacks. PD and ARom carried out the statistical analyses and wrote the manuscript. All authors gave final approval for publication.
Funding
Funding for this work was provided by the Swiss National Science Foundation 31003A_153467.
Conflict of Interest Statement
PM and XF were employed by company PRONA.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the Swiss National Science Foundation (grant no. 31003A_173178) for financial support and are grateful to two referees who gave useful comments on a previous version of the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2019.00351/full#supplementary-material
References
Akcay, C., Tom, M. E., Campbell, S. E., and Beecher, M. D. (2013). Song type matching is an honest early threat signal in a hierarchical animal communication system. Proc. R. Soc. B Biol. Sci. 280:20122517. doi: 10.1098/rspb.2012.2517
Bates, D., Mächler, M., Bolker, B., and Walker, S. (2014). Fitting linear mixed-effects models using lme4. arXiv preprint arXiv:1406.5823.
Benton, D., Owens, D. S., and Parker, P. Y. (1994). Blood-glucose influences memory and attention in young-adults. Neuropsychologia 32, 595–607. doi: 10.1016/0028-3932(94)90147-3
Binmore, K. (2010). Bargaining in biology? J. Evol. Biol. 23, 1351–1363. doi: 10.1111/j.1420-9101.2010.02011.x
Briffa, M., Elwood, R. W., and Dick, J. T. A. (1998). Analysis of repeated signals during shell fights in the hermit crab Pagurus bernhardus. Proc. R. Soc. B Biol. Sci. 265, 1467–1474. doi: 10.1098/rspb.1998.0459
De Callieres, F. (1738). The Art of Negotiating with Sovereign Princes. London: George Strahan; Bernard Lintott and John Graves.
Dreiss, A. N., Ducouret, P., Ruppli, C. A., Rossier, V., Hernandez, L., Falourd, X., et al. (2017). No need to shout: effect of signal loudness on sibling communication in barn owls Tyto alba. Ethology 123, 419–424. doi: 10.1111/eth.12612
Dreiss, A. N., Henry, I., Ruppli, C., Almasi, B., and Roulin, A. (2010a). Darker eumelanic barn owls better withstand food depletion through resistance to food deprivation and lower appetite. Oecologia 164, 65–71. doi: 10.1007/s00442-010-1680-7
Dreiss, A. N., Lahlah, N., and Roulin, A. (2010b). How siblings adjust sib–sib communication and begging signals to each other. Anim. Behav. 80, 1049–1055. doi: 10.1016/j.anbehav.2010.09.012
Dreiss, A. N., Ruppli, C. A., Antille, S., and Roulin, A. (2014). Information retention during competitive interactions: siblings need to constantly repeat vocal displays. Evol. Biol. 42, 63–74. doi: 10.1007/s11692-014-9299-y
Dreiss, A. N., Ruppli, C. A., Faller, C., and Roulin, A. (2015). Social rules govern vocal competition in the barn owl. Anim. Behav. 102, 95–107. doi: 10.1016/j.anbehav.2014.12.021
Dreiss, A. N., Ruppli, C. A., Oberli, F., Antoniazza, S., Henry, I., and Roulin, A. (2013). Barn owls do not interrupt their siblings. Anim. Behav. 86, 119–126. doi: 10.1016/j.anbehav.2013.04.019
Dreiss, A. N., Sechaud, R., Beziers, P., Villain, N., Genoud, M., Almasi, B., et al. (2016). Social huddling and physiological thermoregulation are related to melanism in the nocturnal barn owl. Oecologia 180, 371–381. doi: 10.1007/s00442-015-3491-3
Ducouret, P., Dreiss, A. N., Gemard, C., Falourd, X., and Roulin, A. (2018). Barn owl nestlings vocally escalate when interrupted by a sibling: evidence from an interactive playback experiment. Anim. Behav. 145, 51–57. doi: 10.1016/j.anbehav.2018.09.003
Ducouret, P., Dreiss, A. N., Marmaroli, P., Falourd, X., and Roulin, A. (2016). “Quelle est la stratégie optimale pour remporter la négociation? Expériences de playbacks interactifs avec les poussins de Chouette effraies,” in Congrès Français d'Acoustique, CFA 2016, 20e colloque VIbrations, SHocks and NOise (Le Mans), 2329–2334.
Durant, J. M., and Handrich, Y. (1998). Growth and food requirement flexibility in captive chicks of the European barn owl (Tyto alba). J. Zool. 245, 137–145. doi: 10.1111/j.1469-7998.1998.tb00083.x
Enquist, M., and Leimar, O. (1983). Evolution of fighting behavior - decision rules and assessment of relative strength. J. Theor. Biol. 102, 387–410. doi: 10.1016/0022-5193(83)90376-4
Enquist, M., Leimar, O., Ljungberg, T., Mallner, Y., and Segerdahl, N. (1990). A test of the sequential assessment game - fighting in the cichlid fish Nannacara-Anomala. Anim. Behav. 40, 1–14. doi: 10.1016/S0003-3472(05)80660-8
Goutte, S., Kime, N. M., Argo, T. F., and Ryan, M. J. (2010). Calling strategies of male tungara frogs in response to dynamic playback. Behaviour 147, 65–83. doi: 10.1163/000579509x12483520922205
Grafen, A. (1990). Biological signals as handicaps. J. Theor. Biol. 144, 517–546. doi: 10.1016/S0022-5193(05)80088-8
Greenfield, M. D., Tourtellot, M. K., and Snedden, W. A. (1997). Precedence effects and the evolution of chorusing. Proc. R. Soc. B Biol. Sci. 264, 1355–1361. doi: 10.1098/rspb.1997.0188
Hinde, C. A., and Kilner, R. M. (2007). Negotiations within the family over the supply of parental care. Proc. R. Soc. B Biol. Sci. 274, 53–60. doi: 10.1098/rspb.2006.3692
Johnstone, R. A., and Hinde, C. A. (2006). Negotiation over offspring care - how should parents respond to each other's efforts? Behav. Ecol. 17, 818–827. doi: 10.1093/beheco/arl009
Johnstone, R. A., and Roulin, A. (2003). Sibling negotiation. Behav. Ecol. 14, 780–786. doi: 10.1093/beheco/arg024
Keil, P. L., and Watson, P. J. (2010). Assessment of self, opponent and resource during male-male contests in the sierra dome spider, Neriene litigiosa: Linyphiidae. Anim. Behav. 80, 809–820. doi: 10.1016/j.anbehav.2010.07.013
Kilner, R. M. (2001). A growth cost of begging in captive canary chicks. Proc. Natl. Acad. Sci. U.S.A. 98, 11394–11398. doi: 10.1073/pnas.191221798
King, S. L. (2015). You talkin' to me? Interactive playback is a powerful yet underused tool in animal communication research. Biol. Lett. 11:20150403. doi: 10.1098/rsbl.2015.0403.
Leech, S. M., and Leonard, M. L. (1996). Is there an energetic cost to begging in nestling tree swallows (Tachycineta bicolor)? Proc. R. Soc. B Biol. Sci. 263, 983–987. doi: 10.1098/rspb.1996.0145
Maynard Smith, J. (1982). Do animals convey information about their intentions? J. Theor. Biol. 97, 1–5. doi: 10.1016/0022-5193(82)90271-5
McCarty, J. P. (1996). The energetic cost of begging in nestling passerines. Auk 113, 178–188. doi: 10.2307/4088944
McNamara, J. M., Gasson, C. E., and Houston, A. I. (1999). Incorporating rules for responding into evolutionary games. Nature 401, 368–371. doi: 10.1038/43872
Moreno-Rueda, G. (2007). Is there empirical evidence for the cost of begging? J. Ethol. 25, 215–222. doi: 10.1007/s10164-006-0020-1
Moreno-Rueda, G. (2010). An immunological cost of begging in house sparrow nestlings. Proc. R. Soc. B Biol. Sci. 277, 2083–2088. doi: 10.1098/rspb.2010.0109
Moss, M. C., Scholey, A. B., and Wesnes, K. (1998). Oxygen administration selectively enhances cognitive performance in healthy young adults: a placebo controlled double blind crossover study. Psychopharmacology 138, 27–33. doi: 10.1007/s002130050641
Noguera, J. C., Morales, J., Perez, C., and Velando, A. (2010). On the oxidative cost of begging: antioxidants enhance vocalizations in gull chicks. Behav. Ecol. 21, 479–484. doi: 10.1093/beheco/arq005
Parker, G. A. (1974). Assessment strategy and evolution of fighting behavior. J. Theor. Biol. 47, 223–243. doi: 10.1016/0022-5193(74)90111-8
Patricelli, G. L., Krakauer, A. H., and McElreath, R. (2011). Assets and tactics in a mating market: economic models of negotiation offer insights into animal courtship dynamics on the lek. Curr. Zool. 57, 225–236. doi: 10.1093/czoolo/57.2.225
Patricelli, G. L., Uy, J. A. C., Walsh, G., and Borgia, G. (2002). Male displays adjusted to female's response - macho courtship by the satin bowerbird is tempered to avoid frightening the female. Nature 415, 279–280. doi: 10.1038/415279a
Payne, R. J. H., and Pagel, M. (1996). Escalation and time costs in displays of endurance. J. Theor. Biol. 183, 185–193. doi: 10.1006/jtbi.1996.0212
Payne, R. J. H., and Pagel, M. (1997). Why do animals repeat displays? Anim. Behav. 54, 109–119. doi: 10.1006/anbe.1996.0391
Pika, S., and Frohlich, M. (2019). Gestural acquisition in great apes: the social negotiation hypothesis. Anim. Cogn. 22, 551–565. doi: 10.1007/s10071-017-1159-6
R Core Team (2013). R: A Language And Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available online at: https://www.R-project.org/
Reddon, A. R., Voisin, M. R., Menon, N., Marsh-Rollo, S. E., Wong, M. Y., and Balshine, S. (2011). Rules of engagement for resource contests in a social fish. Anim. Behav. 82, 93–99. doi: 10.1016/j.anbehav.2011.04.003
Reichert, M. S., and Gerhardt, H. C. (2012). Trade-Offs and upper limits to signal performance during close-range vocal competition in gray tree frogs Hyla versicolor. Am. Nat. 180, 425–437. doi: 10.1086/667575
Roberts, G. (2005). Cooperation through interdependence. Anim. Behav. 70, 901–908. doi: 10.1016/j.anbehav.2005.02.006
Roulin, A. (2001a). Food supply differentially affects sibling negotiation and competition in the barn owl (Tyto alba). Behav. Ecol. Sociobiol. 49, 514–519. doi: 10.1007/s002650100322
Roulin, A. (2001b). On the cost of begging vocalization: implications of vigilance. Behav. Ecol. 12, 506–511. doi: 10.1093/beheco/12.4.506
Roulin, A. (2002). “The sibling negotiation hypothesis,” in The Evolution of Begging (Heidelberg: Springer), 107–126.
Roulin, A., and Bersier, L. F. (2007). Nestling barn owls beg more intensely in the presence of their mother than in the presence of their father. Anim. Behav. 74, 1099–1106. doi: 10.1016/j.anbehav.2007.01.027
Roulin, A., Des Monstiers, B., Ifrid, E., Da Silva, A., Genzoni, E., and Dreiss, A. N. (2016). Reciprocal preening and food sharing in colour polymorphic nestling barn owls. J. Evol. Biol. 29, 380–394. doi: 10.1111/jeb.12793
Roulin, A., Dreiss, A. N., Fioravanti, C., and Bize, P. (2009). Vocal sib–sib interactions: how siblings adjust signalling level to each other. Anim. Behav. 77, 717–725. doi: 10.1016/j.anbehav.2008.12.004
Roulin, A., Kolliker, M., and Richner, H. (2000). Barn owl (Tyto alba) siblings vocally negotiate resources. Proc. R. Soc. B Biol. Sci. 267, 459–463. doi: 10.1098/rspb.2000.1022
Roulin, A., Muller, W., Sasvari, L., Dijkstra, C., Ducrest, A. L., Riols, C., et al. (2004). Extra-pair paternity, testes size and testosterone level in relation to colour polymorphism in the barn owl Tyto alba. J. Avian Biol. 35, 492–500. doi: 10.1111/j.0908-8857.2004.03294.x
Ruppli, C. A., Dreiss, A. N., and Roulin, A. (2013a). Efficiency and significance of multiple vocal signals in sibling competition. Evol. Biol. 40, 579–588. doi: 10.1007/s11692-013-9233-8
Ruppli, C. A., Dreiss, A. N., and Roulin, A. (2013b). Nestling barn owls assess short-term variation in the amount of vocally competing siblings. Anim. Cogn. 16, 993–1000. doi: 10.1007/s10071-013-0634-y
Searcy, W. A., and Beecher, M. D. (2009). Song as an aggressive signal in songbirds. Anim. Behav. 78, 1281–1292. doi: 10.1016/j.anbehav.2009.08.011
Searcy, W. A., and Nowicki, S. (2005). The Evolution of Animal Communication: Reliability and Deception in Signaling Systems. Princeton, NJ: Princeton University Press.
Sirot, E. (2012). Negotiation may lead selfish individuals to cooperate: the example of the collective vigilance game. Proc. R. Soc. B Biol. Sci. 279, 2862–2867. doi: 10.1098/rspb.2012.0097
Van Dyk, D. A., Taylor, A. J., and Evans, C. S. (2007). Assessment of repeated displays: a test of possible mechanisms. J. Exp. Biol. 210, 3027–3035. doi: 10.1242/jeb.007492
Keywords: communication, interactive playback, sibling competition, sibling negotiation, temporal dynamics, Tyto alba
Citation: Ducouret P, Romano A, Dreiss AN, Marmaroli P, Falourd X and Roulin A (2019) The Art of Diplomacy in Vocally Negotiating Barn Owl Siblings. Front. Ecol. Evol. 7:351. doi: 10.3389/fevo.2019.00351
Received: 13 May 2019; Accepted: 02 September 2019;
Published: 20 September 2019.
Edited by:
James Luke Savage, University of Sheffield, United KingdomReviewed by:
Wendt Müller, University of Antwerp, BelgiumMichael Reichert, Oklahoma State University, United States
Copyright © 2019 Ducouret, Romano, Dreiss, Marmaroli, Falourd and Roulin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pauline Ducouret, cGF1bGluZS5kdWNvdXJldEBnbWFpbC5jb20=