 Mélissa Peignier1,2,3
Mélissa Peignier1,2,3 Tamara Pokorny4
Tamara Pokorny4 Jürgen Heinze4Rosanna Lindgren1,2
Jürgen Heinze4Rosanna Lindgren1,2 Heikki Helanterä1,2,5*†
Heikki Helanterä1,2,5*† Eva Schultner4*†
Eva Schultner4*†- 1Organismal and Evolutionary Biology Research Programme, University of Helsinki, Helsinki, Finland
- 2Tvärminne Zoological Station, University of Helsinki, Hanko, Finland
- 3Comparative Cognition, Messerli Research Institute, Vetmeduni Vienna, Medical University of Vienna, University of Vienna, Vienna, Austria
- 4Zoology and Evolutionary Biology, Biologie I, University of Regensburg, Regensburg, Germany
- 5Ecology and Genetics Research Unit, University of Oulu, Oulu, Finland
Social insects live in highly complex societies with efficient communication systems. Begging is one display commonly used by offspring to signal their nutritional state, however begging behavior has received very little attention in social insects. Theory predicts that begging can be either an honest (i.e., honest-signaling strategy) or a dishonest (i.e., scrambling competition) signal of need, with dishonest signals expected to be more likely when relatedness within the group is low. To investigate the presence and honesty of begging, as well as the nature of the involved signals, we used a comparative approach with four species of the ant genus Formica known to differ in the degree of intra-colony relatedness. We investigated the behavior of starved and non-starved larvae of F. aquilonia, F. pressilabris (both low intra-colony relatedness), F. exsecta (intermediate relatedness), and F. fusca (high relatedness). In addition, we assessed the attraction of conspecific workers toward odors extracted from these two classes of larvae and analyzed the larval cuticular hydrocarbon profiles. We found that in F. fusca and F. exsecta, larvae signaled significantly more when starved. In contrast, larvae of F. aquilonia signaled significantly more when they were non-starved, while there was no significant difference in the behavior of starved vs. non-starved larvae in F. pressilabris. Our results show that workers were not preferentially attracted to the odor of starved larvae, and we also did not detect any differences between the cuticular hydrocarbon profiles of starved and non-starved larvae. Overall, this study demonstrates among species variation in larval hunger signaling in Formica ants, and encourages further studies to confirm the link between kin structure variation and the honesty of begging signals.
Introduction
Communication systems allow individuals to base their adaptive decisions on the behavior, morphology, or physiology of others (Endler, 1993). Signals have evolved to convey information to receivers, so that the information extracted from the signal elicits a response and results, on average, in positive fitness consequences for both the sender and the receiver (Laidre and Johnstone, 2013). To be efficient these interactions must depend on a repertoire of signs and rules shared between the participants of the interaction (Maynard Smith and Harper, 2003; Witzany, 2010). Communication and signaling systems thus allow individuals to affect the plastic decisions of others, possibly generating coevolution (Font and Carazo, 2010).
Despite the social nature of communicative signals, the adaptive function of signals from the receiver's perspective may be fundamentally different from that of the sender (Marler, 1961). Traditionally, communication has been seen as a voluntary transfer of information from the sender to the receiver, when the inclusive fitness interests of the sender and the receiver converge. However, when the inclusive fitness interests of the sender and the receiver diverge, the ultimate aim of the sender is not the sharing of mutual beneficial information, and conflicts over information use arise (Owren and Rendall, 1997, 2001; Seyfarth and Cheney, 2003). In such cases, selection should favor senders, whose signals affect the behavior of the receiver at their own advantage, and receivers, who are able to extract information from the signals (Endler, 1993; Font and Carazo, 2010). Kinship and the inclusive fitness benefits gained through helping relatives (Hamilton, 1964) should influence the type of communication. Closely related individuals should be more likely to communicate honestly in order to cooperate, whereas the opposite would be expected for non-relatives.
Begging is commonly used by offspring to influence the quantity and quality of received food, with consequences for survival (Hodar et al., 2002) and other fitness-related traits (Breteler and Gonzalez, 1988; Chapman, 1998; Shafiei et al., 2001; Engels and Sauer, 2007). Godfray (1991, 1995) developed the idea that begging should be an honest signal of need. Food provisioning by parents is obviously a cooperative behavior but contains an element of conflict as well. This is because the existence of relatedness asymmetries leads offspring to value themselves more than they value their siblings, and thus, to selfishly beg for a larger part of parental investment. However, for a parent, providing one offspring with a larger investment influences the trade-off between the expected fitness returns from this offspring vs. the others (Trivers, 1972, 1974). Therefore, parents would benefit from accurately assessing the level of need expressed through the begging signal. Other models, called scramble competition models, predict that this conflict between parent and offspring over the distribution of resources is increased by competition among siblings (Parker and Macnair, 1979). In such a case, each offspring is assumed to beg more than other offspring even when already fed, leading to an exaggeration of need (Godfray, 1995; Royle et al., 2002; Johnstone, 2004; Akçay, 2012). Scramble competition, and consequently dishonest signaling, should thus be more prevalent when competition occurs among a greater number of siblings or when the relatedness among brood is low (Parker et al., 2002; Johnstone, 2004). This is because the inclusive fitness costs of dishonestly taking food from other brood members are lower under low relatedness. In contrast, signals should be honest indicators of need under high relatedness.
Both honest and dishonest begging has been demonstrated previously. Food-deprived pigeon squabs (Columba livia) spend more time begging than recently fed ones (Mondloch, 1995), giving support to honest signaling models. Conversely, begging calls of yellow-headed blackbird chicks varied with the intensity of sibling competition, independent of need (Price et al., 1996). Empirical studies on begging behavior are taxonomically highly biased toward birds and consequently also heavily focused on vocal and visual signals (Kedar et al., 2000; Leonard and Horn, 2005; Marques et al., 2008; Klenova, 2015; Caro et al., 2016). However, offspring of insects with maternal care also display various begging behaviors, mainly in order to solicit protection and food (reviewed in Mas and Kölliker, 2008). For instance, burying beetle larvae (Nicrophorus) beg by approaching their parents' mouthparts, pushing their heads against them and touching them with their legs (Milne and Milne, 1976; Rauter and Mooref, 1999).
Colonies of social insects—ants, bees, wasps, and termites—are especially interesting to study from a signaling perspective because they form large, complex societies, which function based on chemical, visual, acoustic, vibrational, and tactile signals (Leonhardt et al., 2016). In ants, kinship among colony members varies widely due to variation in queen number and queen mating frequency. This makes them particularly suitable for testing predictions concerning signaling honesty. Recently, selfish brood behaviors were shown to be correlated with low relatedness, raising the possibility that brood might indeed respond to kin structure variation in adaptive ways (Schultner et al., 2013, 2014). However, offspring begging as a classic conflict-related trait has been largely overlooked in social insects. The few empirical studies that have characterized begging in social Hymenopteran larvae show that various behavioral hunger signals exist, from scraping mandibles against cell walls (vespine wasp larvae: Ishay and Schwartz, 1973) to flexing the head or whole body to attract workers (ant larvae: Brian, 1977; Creemers et al., 2003; Kaptein et al., 2005). For example, Gnamptogenys striatula ant larvae perform two types of movement to signal their hunger toward workers after a 24 h starvation period: “swaying,” which corresponds to a repositioning of the larval body to reach food or workers and “stretching,” which corresponds to a rhythmic stretching of the body (Kaptein et al., 2005). Hungry larvae perform these movements more often, indicating that begging signals honestly reflect need in this species. Similar behavioral patterns have been detected in Myrmica ant larvae (Creemers et al., 2003). Additionally, larvae may solicit food from workers using chemical hunger signals in bees (den Boer and Duchateau, 2006; He et al., 2016), and fire ants (Cassill and Tschinkel, 1995). While these studies provide first evidence for the ability of social insect larvae to influence their food intake via begging, comparative studies are needed to test how begging in social insects is influenced by colony kin structure.
We used a comparative approach to investigate larval hunger signaling in ants. Our aim was to assess the existence of hunger signals and the effect of kinship on the honesty of these signals. We focused on Formica ants because their colony kin structures range from single queen colonies with high intra-colony relatedness to multiple queen colonies with extremely low relatedness among nestmates (Rosengren et al., 1993; Sundström et al., 2005). For four species of Formica ants, representing a range of intra-colonial relatedness, we assessed the behavior of starved and non-starved larvae. We furthermore analyzed the cuticular odors of starved and non-starved larvae and experimentally tested for potential chemical hunger signals by presenting workers with larval cuticular extracts. In line with inclusive fitness predictions, we find that starved larvae signaled significantly more than non-starved larvae in Formica fusca and F. exsecta, i.e., species with high and intermediate levels of intra-colony relatedness. In contrast in F. aquilonia, a low relatedness species, larvae signaled significantly more when they were not starved, while there was no significant difference in the behavior of starved vs. non-starved larvae in the other low relatedness species. Workers were not preferentially attracted to the odor of starved larvae, and we also did not detect any differences between the cuticular hydrocarbon profiles of starved and non-starved larvae. Our study demonstrates that larval hunger signaling varies among species in ants and suggests links between kinship and the honesty of hunger signaling in social insects.
Materials and Methods
Study Species and Experimental Setup
Formica ants are characterized by remarkable intra- and interspecific variability in within-nest relatedness, largely due to variation in queen numbers (Rosengren et al., 1993; Sundström et al., 2005). We studied four Formica species covering the full range of relatedness values: F. aquilonia forms large networks of interconnected nests (so-called polydomy), where each nest contains hundreds of queens (Rosengren et al., 1993; personal observation), and relatedness among nestmates is very low (r = 0.06 ± 0.09 CI, Schultner et al., 2014, 2016). F. pressilabris is also typically polydomous and nests contain multiple queens, usually some dozens (r = 0.21 ± 0.09 CI, Schultner et al., 2014). These species are therefore classified as “low-relatedness” species. In contrast, F. fusca nests typically contain ~1–10 queens (Hannonen and Sundström, 2003; Helanterä et al., 2011), and each colony comprises one nest only (so-called monodomy). This species is thus considered a “high-relatedness” species (r = 0.46 ± 0.14 CI, Schultner et al., 2014). F. exsecta can form both monodomous colonies containing single queens and polydomous colonies in which each nest contains dozens of queens (Rosengren et al., 1993). The F. exsecta colonies used in this study were collected from a polydomous population (r = 0.09 ± 0.08 CI, Schultner et al., 2014). However, in our study area most nests of this species form monodomous populations, in which colonies have only one queen and very high relatedness (r = 0.71 ± 0.03 SE; Sundström et al., 2003). Compared to F. aquilonia and F. pressilabris, which predominantly live in polydomous, low relatedness populations, and F. fusca, which always live in monodomous, high relatedness populations, F. exsecta is therefore considered an “intermediate relatedness” species. Note that of the four species, only F. fusca founds colonies independently, while the other three species are so-called temporary social parasites, which found colonies in the nests of other ants, including F. fusca (Buschinger, 2009), or dependently with the help of conspecific workers. All colonies used in this study were collected from established populations in which the host species have already been eradicated, and none contained any host workers.
From each species, colony fragments including queens, workers, and nest material were collected in the field and brought back to the laboratory. Colony fragments were then placed in containers lined with Fluon, together with a ceramic plate, some compost as nest material and Sphagnum moss to maintain humidity. The colonies were fed with Bhatkar-Whitcomb diet (Bhatkar and Whitcomb, 1970) and watered daily. Colony fragments were checked daily for the presence of eggs. When the first eggs were observed, up to 10 queens were taken from each fragment, placed individually in Petri dishes with two pieces of 1 cm × 1 cm humid sponge and kept in the dark at room temperature. After 2 days, queens were removed and put back into their colony fragments. Eggs deposited on the Petri dishes were then counted and checked every 2 days to remove fungal spores, re-humidify sponges and remove dead eggs. Eggs from all species hatched within 9–15 days.
We created two groups of larvae differing in regard to their state of hunger for behavioral observations and chemical assays. Freshly hatched larvae (0–3 h old) were considered not to be hungry (“non-starved” hereafter), and were either moved to a new Petri dish, in which their behavior was observed immediately (see “Quantification of larval behavior”), or directly collected in glass vials for chemical analyses (Figure 1). To obtain hungry (“starved” hereafter) larvae, freshly hatched larvae were moved to a new Petri dish containing a piece of 1 cm × 1 cm sponge, where they remained for 21–24 h. Larvae were thereafter subjected to behavioral or chemical analyses as described for non-starved larvae (Figure 1). Larvae did not have the opportunity to cannibalize one another on the Petri dish because they were physically separated, and the total number of larvae on each Petri dish was documented. No larvae disappeared from any of the Petri dishes during the starvation period, indicating that cannibalism did not occur. The number of replicates conducted for behavioral assays and the number of samples used in chemical analyses are summarized in Table 1.
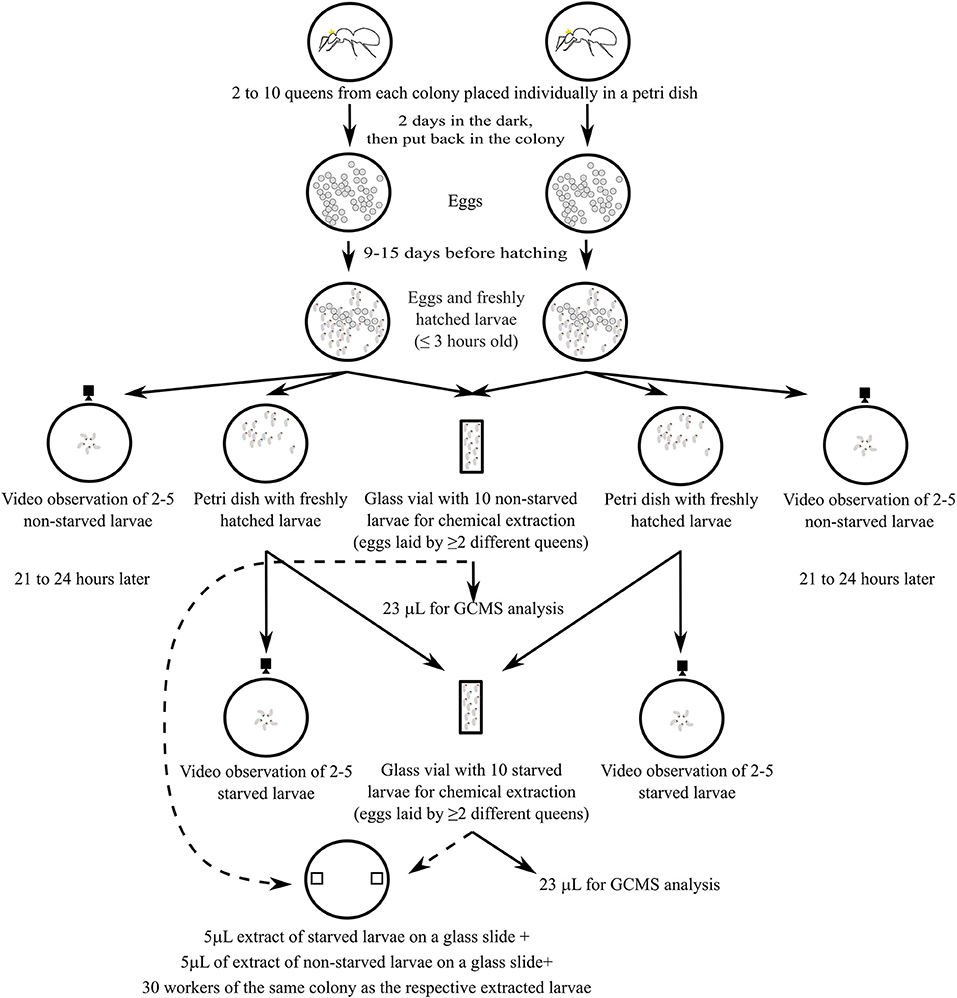
Figure 1. Experimental setup for rearing of larvae of one species, the same setup was used for all the species. Eggs are represented by gray dots. Non-starved larvae hatched between 0 and 3 h after the first observation. Starved larvae hatched between 0 and 3 h after the first observation and were placed on a separate Petri dish for an additional 21–24 h. All the video observations have been carried out with larvae from a single matriline.
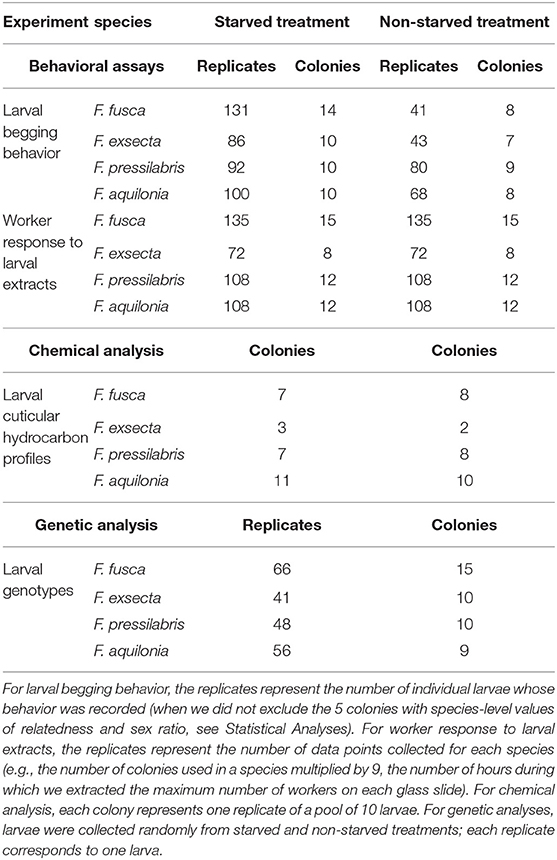
Table 1. Number of larvae, workers, and colonies used in behavioral assays and for chemical and genetic analyses.
Quantification of Larval Behavior
To quantify begging behavior, groups of 2–5 starved or non-starved larvae were placed on their backs in a Petri dish (Figure 1). The Petri dish was put under a stereomicroscope onto which a camera (DigiMicroScale) was installed and behaviors were recorded for 10 min. Each group consisted only of larvae produced by the same queen, thus groups were genetically homogenous. To avoid any matriline bias, several groups of larvae produced by different queens from each colony were used (at least two different queens). To avoid observer bias, videos were renamed by someone unfamiliar with the experimental setup prior to analysis. We observed two different kind of behaviors performed by the larvae. The first one, termed “swaying” in reference to Kaptein et al. (2005), consisted of the larva moving its body upwards, remaining in this position for some time and/or rhythmically repeating the movement. The second recorded behavior was a rhythmic opening and closing of the mandibles. This behavior has been implicated in larval begging in attine ants (Wheeler and Bailey, 1920), and preliminary observations suggest it may correlate with nutritional status in Formica ants. For each larva, the number and duration (in seconds) of swaying movements and the number and duration of mandible movements were recorded with the software Boris (version 6.3.1, Friard and Gamba, 2016).
Intracolony Relatedness and Larval Sex Ratios
Levels of selfishness–in the form of egg cannibalism–exhibited by Formica ant larvae depend on intra-colony relatedness and larval sex (Schultner et al., 2013, 2014). While female larvae behave less selfishly when within-colony relatedness is high, male larvae do not adjust their levels of selfishness to variation in colony kin structure. In order to estimate the effect of intra-colony relatedness and larval sex on begging behavior, we determined relatedness and sex of larvae from genotype data using six microsatellite loci previously tested in our focal species (FE 19, FE 42, FE 51, FY 4, FY7, FL20; Schultner et al., 2013, 2014, 2016). To this end, we collected 2–7 larvae from 10 to 15 colonies of each species in 70% EtOH (Table 1). For DNA extraction, each larva was placed in an individual well with a 2.5 μl: 100 μl Proteinase K–Chelex solution (6%), and incubated overnight at 56°C. After inhibition of Proteinase K activity at 100°C for 10 min, 1 μl of DNA was used in simplex PCR reactions, together with 5 μl of TAQ polymerase, 3 μl of H2O, and 1 μl primer (0.5 μl forward primer, 0.5 μl reverse primer). PCRs were run with protocols optimized for each primer and PCR products analyzed in a 310 ABI PRISM sequencer (Applied Biosystems). Each microsatellite peak was scored manually with GeneScan 3.1 software (Applied Biosystems).
Intra-colony relatedness was estimated with Relatedness 5.0.8 software (Queller and Goodnight, 1989) using background allele frequencies calculated from the genotypes generated for this study. For all relatedness estimates, standard errors were calculated by jackknifing over colonies. To determine larval sex, individuals were scored as females if they were heterozygous at one or more loci and as males if they were homozygous at all six loci. In some cases, not all loci amplified successfully; for putative males, which were homozygous at <6 loci (15 out of 29 homozygous individuals), we calculated the probability of wrongly assigning a larva as a male using previously established methods (Schultner et al., 2014), and scored larvae with a misclassification probability below 10% as males (0 putative male larvae excluded). For each colony, the average larval sex ratio was calculated from sexed individuals, with 0 = all females and 1 = all males.
Worker Response to Larval Odors
From several (at least two) queens from each colony fragment, we collected and froze pools of 10 starved and 10 non-starved larvae in individual glass vials (Sigma Aldrich) (Table 1). Cuticular compounds were extracted from each group of 10 larvae for 1 min with 30 μL of hexane. Thereafter, 28 μl of the solvent were transferred to a new, clean vial. From each pooled sample (e.g., each group of 10 larvae), 5 μl were used to investigate chemical hunger signaling. The test setup consisted of two 0.5 cm × 0.5 cm glass slides that were placed in a round container (7 cm diameter). The containers' sides were covered with black paper and the containers themselves were covered with red transparent paper and placed in a cardboard box to minimize luminosity and disturbance. In each container, one glass slide was covered with 5 μL of cuticular extract from non-starved larvae (hereafter, “non-starved odor”) while the other one was covered with 5 μL of cuticular extract from starved larvae (hereafter, “starved odor”) (Figure 1). After the solvent had evaporated (30 min), 30 workers were added from the same colony fragment as the respective extracted larvae. After 30 min of adaptation, 10 pictures were taken at 5 s intervals at every full hour for 9 h, for a total of 90 pictures per container. Each camera (two in total, camera model: SONY HDR-SR8E) was able to capture 12 containers at the same time. The number of workers with at least one body part (legs, antennae) on the glass slide was recorded in each picture for each glass slide. To avoid any side bias, the position of the cuticular extracts was randomized among replicates for each species. Prior to analyses, the pictures were renamed by someone unfamiliar with the experimental setup to avoid any observer bias. We predict that if a chemical signal is produced by larvae to signal their state, workers should be more attracted toward the odors of starved larvae than to the odors of non-starved larvae, irrespective of the species.
Characterization of Larval Chemical Profiles
Chemical analyses were conducted on the remaining 23 μl of each sample. Samples were fractionated on silica columns (Chromabond SiOH, 1 mL/100 mg) to remove fatty acids (likely originating internally, as the larval cuticle is thin and not all larvae remained intact throughout the extraction). The non-polar hexane fraction was carefully evaporated under a stream of nitrogen, and then re-dissolved in 23 μl of hexane. Two μl per sample were then subjected to gas chromatography with coupled mass spectrometry (GCMS, Shimadzu GCMS-QP2010 Plus) fitted with a SGE BPX-5 column (31 m × 0.25 μm × 0.25 mm). Helium gas (1 mL/min) served as carrier gas, and injection was splitless. The temperature program started at 70°C isothermal for 1 min, after which the temperature was raised to 200° C at 30°C/min and then from 200 to 320°C at 5°C/min, where it was held for 5 min. As larval samples showed a number of unusual peaks, we also ran control samples of Petri dishes, which were considered a likely cause of contaminations. For that purpose, four clean Petri dishes were separately rinsed with 30 μL of hexane for 1 min each and the resulting samples were analyzed following the protocol above. The chromatograms of each sample were analyzed using ChemStation software (E.02.01.1177). Compounds detected in samples of the clean petri dishes were also found in larval samples and were thus excluded from further analyses. Tentative identification of the remaining peaks was based on their retention indices and mass spectra. We used letters to indicate different but unidentifiable peaks, and “x” and “y” to indicate different positions of double-bonds or methyl groups (Supplement Table 1). Relative amounts for each of the remaining peaks were calculated following Aitchison (1982):
Statistical Analyses
We conducted all statistical analyses with R software 3.5.1 (R Core Team, 2018). To investigate larval begging behavior, we ran generalized linear mixed models that account for zero-inflated data following a Poisson distribution (function glmmTMB in R package glmmTMB, Brooks et al., 2017). Each of the four measures of begging intensity (number of swaying movements, swaying duration, number of mandible movements, mandible movement duration) were treated as dependent variables and investigated separately. To test for overall differences in begging intensity between species, we constructed two global models. The first model included the effects of species and the interaction between mean intra-colony relatedness and larval state as fixed terms, and the group of larvae observed at the same time as a random term:
The second global model tested the effects of species and the interaction between mean intra-colony sex ratio and larval state as fixed terms, and the group of larvae observed at the same time as a random term:
For global models, overall p-values for each term were obtained using Type II Wald X2 tests (R function anova). For each species, we furthermore constructed two species-specific models. The first species-specific model tested the effect of larval state and its interaction with mean intra-colony relatedness (fixed terms), while the group of larvae observed at the same time was used as a random term:
The second species-specific model tested the effect of larval state and its interaction with mean intra-colony sex ratio (fixed terms), while the group of larvae observed at the same time was used as a random term.
The effects of mean intra-colony relatedness and mean intra-colony sex ratios were tested separately because these factors covary for each individual colony as they are based on one set of genotypes per colony. For five colonies, larval genotypes were not available (F. aquilonia = 1 colony, F. exsecta = 1, F. fusca = 2, F. pressilabris = 1); here we substituted species-level mean relatedness and sex ratio values. We ran the two global models and the eight species-specific models without these five colonies to verify whether the results stayed the same. As there were no major differences in the statistical results between the two data sets (compare Tables 2, 3, Supplement Tables 2–4 and Supplement Tables 5–9), we retained the entire data set for all analyses given in the main text.
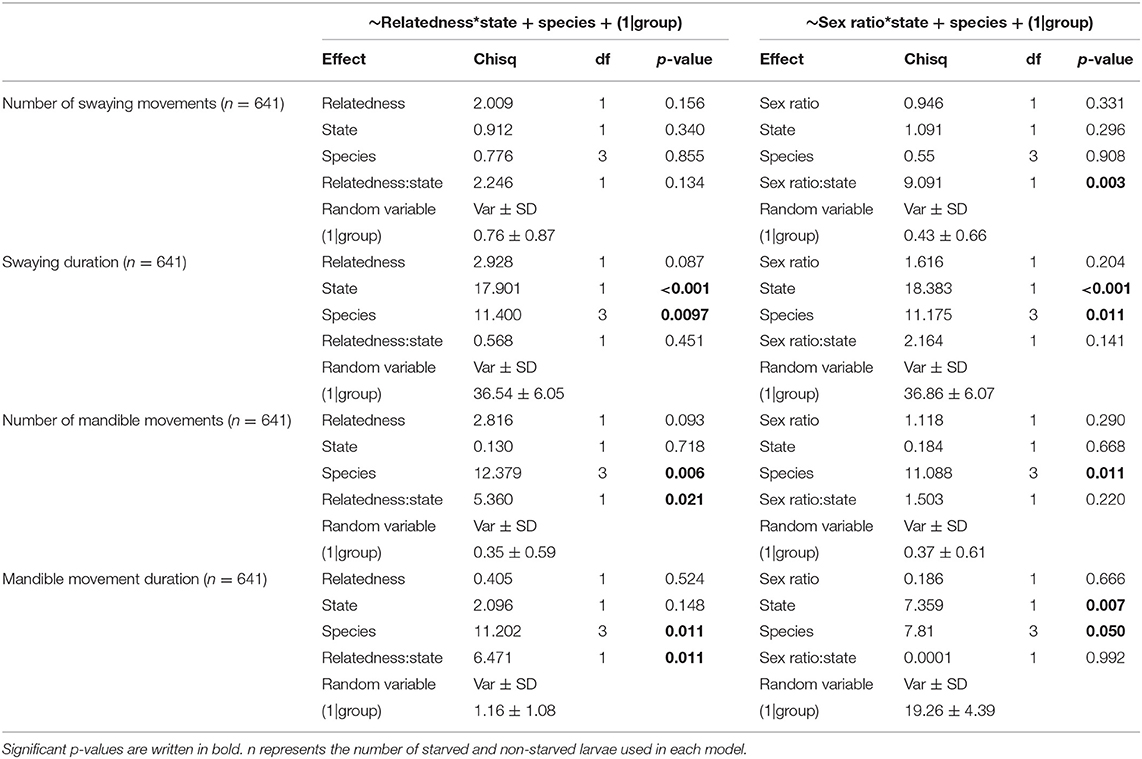
Table 2. Results of Generalized Linear Mixed Models using Template Model Builder linking the number of swaying movements, swaying duration, number of mandible movements, or mandible movement duration to the state and species of the larvae, intra-colony relatedness, and intra-colony sex ratio.
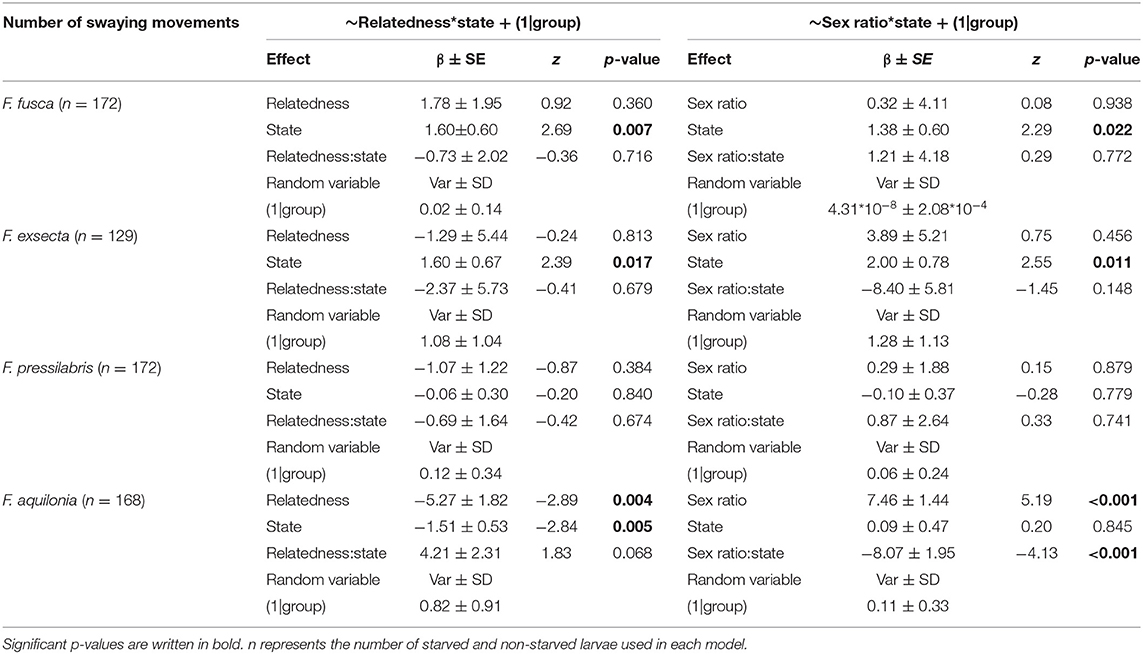
Table 3. Results of Generalized Linear Mixed Models using Template Model Builder linking the number of swaying movements to the state of the larvae, intra-colony relatedness, and intra-colony sex ratio for each species separately.
To compare intra-colony relatedness and sex ratio levels across species, we used Kruskal-Wallis rank sum tests for non-parametric data (R function kruskal.test).
To investigate worker responses to larval odors, we performed generalized linear mixed effects models following a Poisson distribution (function glmer in R package lme4, Bates et al., 2014). We determined the maximum number of workers on a glass slide in each treatment (i.e., starved or non-starved larvae odor) in the 10 pictures from each hour and used it as a response variable. For all species combined, we constructed a model where treatment and species were included as fixed terms while the hour when the picture was taken and the colony from which larvae and workers were sampled were used as random terms:
We also constructed a separate model for each species, where treatment was used as a fixed term and the hour when the picture was taken and the colony from which larvae and workers were sampled were used as random terms:
To characterize the chemical profiles of starved and non-starved larvae we performed principal component analyses (PCA) on transformed proportions of compounds (function fviz_pca in R package factoextra). We performed four separate PCAs using the proportions of all compounds, only alkanes, only methylated alkanes or only alkenes. Each of these PCAs was performed on the combined data from all the species, and for each species separately. The 95% confidence ellipses for each species and treatment were calculated by bootstrapping based on the function coord.ellipse in R package FactoMineR (Lê et al., 2008).
Results
Quantification of Larval Behavior
Overall, larvae swayed slightly less often when they were starved, with the number of swaying events averaging 0.47 ± 0.05 SE in starved larvae and 0.58 ± 0.14 SE in non-starved larvae. Swaying duration was also shorter in starved larvae (12.05 s ± 14.55 SE) than in non-starved larvae (12.85 ± 32.29 SE). Similarly, starved larvae moved their mandibles less often (starved: 1.01 ± 0.08 SE; non-starved: 1.06 ± 0.11 SE) and for shorter periods of time (starved: 24.93 s ± 29.67 SE; non-starved: 25.94 ± 46.70 SE). However, only swaying duration differed significantly between starved and non-starved larvae (Table 2, fixed term state in global model “Swaying duration,” χ2 = 17.90, p < 0.001). Swaying duration, the number of mandible movements and mandible movement duration also differed significantly between species, with F. aquilonia larvae signaling significantly more than larvae of the other three species (Table 2, Global models “Swaying duration,” “Number of mandible movements,” “Mandible movement duration”).
In contrast, there were no significant effects of intra-colony relatedness or sex ratio on the number of swaying events, swaying duration, number of mandible movements or mandible movement duration (Table 2). However, we did find a significant negative effect of the interaction between intra-colony relatedness and larval state on the number and duration of mandible movements (Figure 2, Table 2, fixed term relatedness*state in global models “Number of mandible movements”: χ2 = 5.36, p = 0.021, “Mandible movement duration”; χ2 = 6.47, p = 0.011), indicating that the number and duration of mandible movements decreased significantly with increasing intra-colony relatedness in non-starved but not starved larvae. Additionally, the interaction between intra-colony sex ratio and state had a significant positive effect on the number of swaying events (Figure 2, Table 2, fixed term state*sex ratio in global model “Number of swaying movements,” χ2 = 9.09, p = 0.003). This suggests that the number of swaying movements increased significantly faster with increasing male bias in non-starved compared to starved larvae.
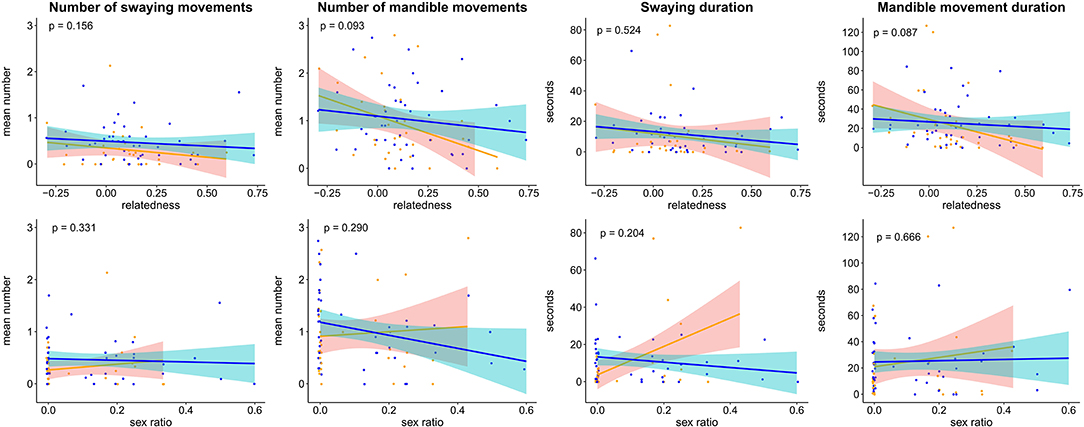
Figure 2. Influence of intra-colony relatedness (top row) and intra-colony sex ratio (bottom row) on the mean number of swaying movements, mean duration of swaying movements, mean number of mandible movements, and mean duration of mandible movements performed by non-starved (orange) and starved (blue) Formica larvae. Each point represents the mean from one colony. Regression lines and confidence intervals were drawn using the R function geom_smooth (method = glm). P-values denote the results from zero-inflated GLMMs (see Table 2).
Within-species analyses showed that F. fusca and F. exsecta larvae swayed more often when starved (Figure 3, Table 3, fixed term state, F. fusca: z = 2.69, p = 0.007; F. exsecta: z = 2.39, p = 0.017), while F. aquilonia larvae swayed more when not starved (Figure 3, Table 3, fixed term state, F. aquilonia: z = −2.84, p = 0.005). The number of swaying movements performed by F. pressilabris larvae did not vary depending on nutritional state (Figure 3, Table 3, fixed term state, F. pressilabris: z = −0.20, p = 0.840). There was no difference between starved and non-starved larvae in the other three measures of begging intensity, with the exception of swaying duration in F. exsecta (Supplement Table 2, fixed term state, F. exsecta: z = 4.72, p < 0.001) and number of mandible movements in F. aquilonia (Supplement Table 3, fixed term state, F. aquilonia: z = −2.90, p = 0.004). The effect of larval state on these behaviors was the same as for the number of swaying movements, with F. exsecta larvae swaying longer when starved and F. aquilonia larvae exhibiting more mandible movements when non-starved.
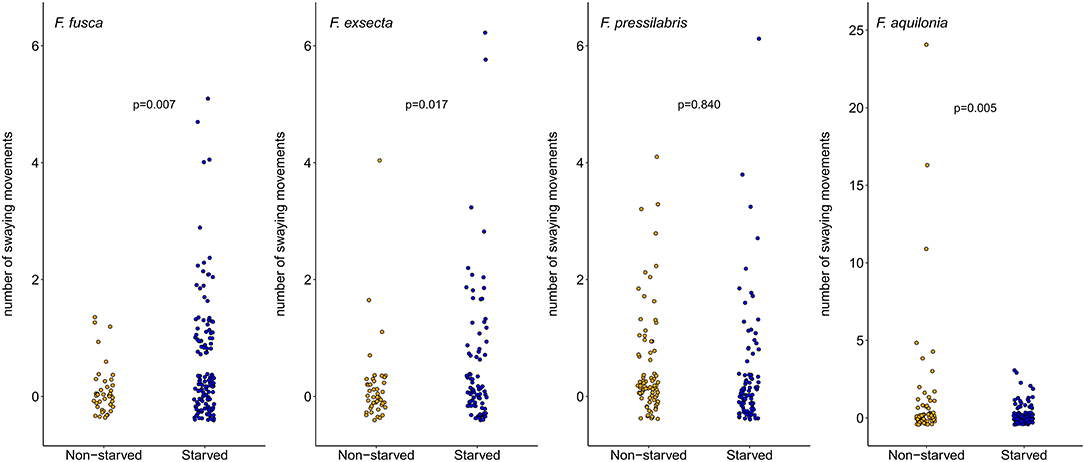
Figure 3. Number of swaying movements performed by non-starved and starved larvae of F. fusca, F. exsecta, F. pressilabris, and F. aquilonia. Each dot represents the number of swaying movements performed by one larva. P-values denote the results from zero-inflated GLMMs (see Table 3). Note different scales on the y-axes.
Analyses of begging behavior within species furthermore revealed that variation in intra-colony relatedness and sex ratio do not influence begging signals to the same degree in all species (Figure 4, Table 3, Supplement Tables 2–4). For example, increasing relatedness had significant negative effects while increasing male-bias in sex ratio had significant positive effects on the number of swaying movements in F. aquilonia (Figure 4, Table 3, fixed term relatedness, F. aquilonia: z = −2.89, p = 0.004; fixed term sex ratio, F. aquilonia: z = 5.19, p < 0.001) but not the other three species. Conversely, increasing relatedness had significant negative effects on the number of mandible movements performed by larvae in F. fusca, F. pressilabris, and F. aquilonia, but not F. exsecta (Supplement Table 3, fixed term relatedness, F. fusca: z = −2.14, p = 0.033; F. pressilabris: z = −2.31, p = 0.021; F. aquilonia: z = −2.42, p = 0.007; F. exsecta: z = 4.02, p = 0.091).
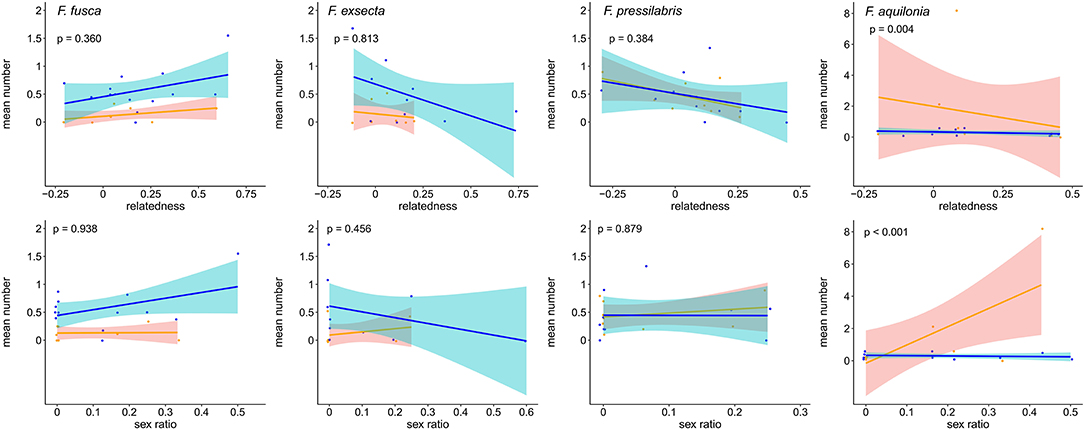
Figure 4. Influence of intra-colony relatedness (top row) and intra-colony sex ratio (bottom row) on the mean number of swaying movements performed by non-starved (orange) and starved (blue) larvae of F. fusca, F. exsecta, F. pressilabris, and F. aquilonia. Each point represents the mean from one colony. Regression lines and confidence intervals were drawn using the R function geom_smooth (method = glm). P-values denote the results from zero-inflated GLMMs (see Table 3). Note different scales on the y-axes.
Intra-colony Relatedness and Larval Sex Ratios
F. fusca had a mean intra-colony relatedness of 0.18 ± 0.05 SE and an intra-colony sex ratio (proportion of males) of 0.13 ± 0.19 SD. F. exsecta had a mean intra-colony relatedness of 0.16 ± 0.06 SE and an intra-colony sex ratio of 0.11 ± 0.20 SD. F. pressilabris had a mean intra-colony relatedness of 0.14 ± 0.07 SE and an intra-colony sex ratio of 0.06 ± 0.11 SD. F. aquilonia had a mean intra-colony relatedness of 0.09 ± 0.05 SE and intra-colony sex ratio of 0.21 ± 0.22 SD. There were no differences in intra-colony sex ratios between species (Kruskal Wallis rank sum test, X2 = 3.357, p = 0.339). Similarly, intra-colony relatedness estimates showed large overlap and there were no differences in relatedness estimates between species (Kruskal Wallis rank sum test, X2 = 0.033, p = 0.998). Intra-colony relatedness was nevertheless included as an explanatory variable in the statistical analyses of begging intensity in order to test whether begging behavior is plastic, i.e., larvae adjust begging intensity to intra-colony relatedness. When interpreting results obtained from comparisons between species, we additionally relied on more robust relatedness estimates obtained from larger data sets in previous studies (Sundström et al., 2003, Schultner et al., 2014, 2016).
Worker Response to Larval Odors
On average, 1.3 (± 1.3 SD) F. fusca workers, 0.8 (± 0.9 SD) F. exsecta workers, 1.3 (± 1.4 SD) F. pressilabris workers, and 1.2 (± 1.3 SD) F. aquilonia workers approached larval odors during the observation period. Workers did not move significantly more toward the odor of starved larvae than toward the odor of non-starved larvae, neither when combining data from all species nor when analyzing each species separately (Figure 5, Table 4, all species: p = 0.61, F. fusca: p = 0.32, F. exsecta: p = 0.85, F. pressilabris: p = 0.06, F. aquilonia: p = 0.61).
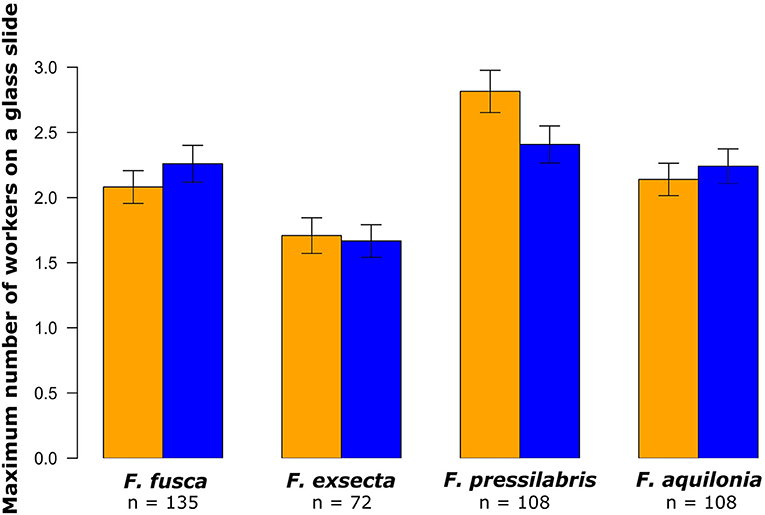
Figure 5. Maximum hourly number of F. fusca, F. exsecta, F. pressilabris, and F. aquilonia workers on glass slides covered with odors from non-starved (orange) and starved (blue) larvae. The number of replicates is noted below each species name. The error bars refer to the 95% confidence interval of the mean.
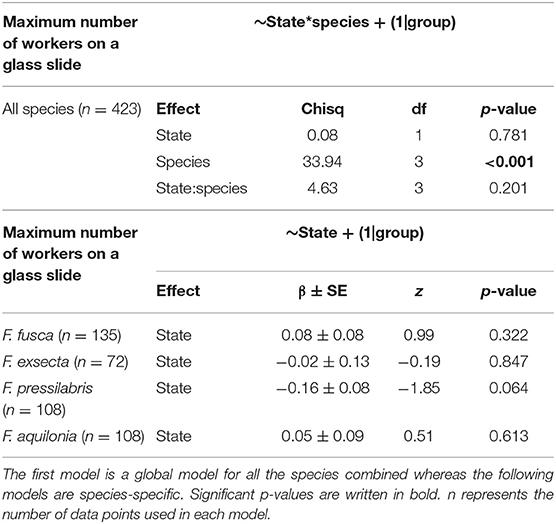
Table 4. Results of Generalized Linear Mixed effects Models linking the maximum number of workers on a glass slide to the odor of starved or non-starved larvae on this slide.
Characterization of the Larval Chemical Profiles
Overall, we found 46 peaks after exclusion of Petri dish contaminations (Supplement Table 1). When analyzing all species together, visual inspection of PCAs carried out with all compounds showed no difference in the cuticular hydrocarbons of starved and non-starved larvae (Figure 6A). Similarly, there was no difference in the cuticular hydrocarbons of starved and non-starved larvae across all species when compound groups where analyzed separately: alkanes (Figure 6B), methylated alkanes (Figure 6C), alkenes (Figure 6D). We also did not find strong differences between the cuticular hydrocarbon profiles of starved and non-starved larvae when analyzing each species separately (Supplement Figures 1–4), with the exception of F. exsecta. However, the apparent difference in the chemical compounds of starved and non-starved F. exsecta larvae may be due to the smaller number of colony replicates for this species. While we did not find any treatment effects on larval odors, the analysis did reveal differences between species (Figure 7). Species differences in larval odor profiles were most apparent when analyses included all compounds (Figure 7A) and only alkenes (Figure 7D). Additionally, the hydrocarbon profiles of F. fusca larvae differed from those of all other species in the composition of methyl-branched alkanes (Figure 7C).
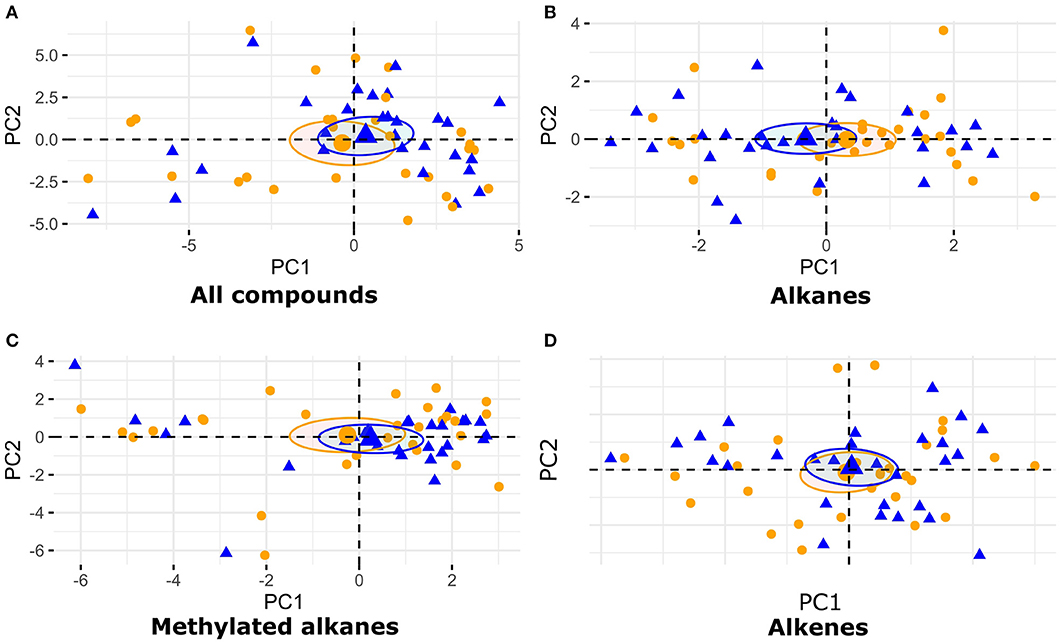
Figure 6. Representation of PCAs of (A) all cuticular hydrocarbons, (B) alkanes, (C) methylated alkanes, and (D) alkenes calculated from relative proportions of cuticular compounds extracted from larvae of four Formica species. Non-starved larvae are represented in orange and starved larvae are represented in blue. The circles represent the 95% confidence ellipse for each group.
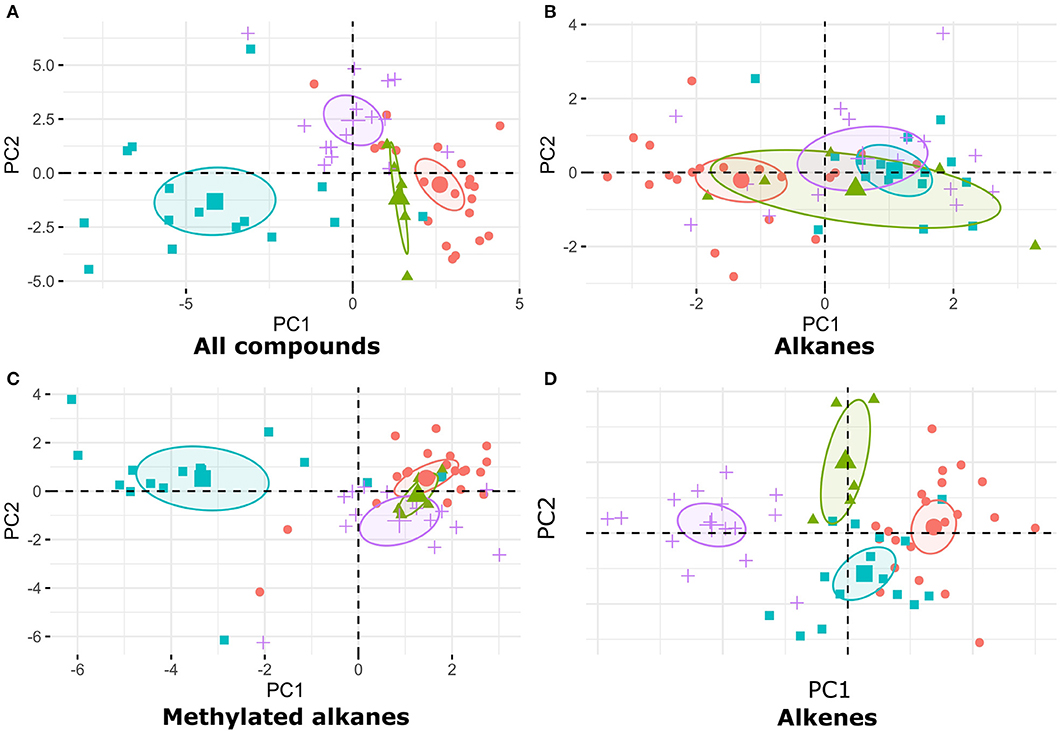
Figure 7. Representation of PCAs of (A) all cuticular hydrocarbons, (B) alkanes, (C) methylated alkanes, and (D) alkenes calculated from relative proportions of cuticular compounds extracted from larvae. F. fusca larvae are represented in blue, F. exsecta in green, F. pressilabris in violet, and F. aquilonia in red. The circles represent the 95% confidence ellipse for each group.
Discussion
So far, there have been few studies on the influence of kinship on signaling honesty in insects. Caro et al. (2016) found that offspring from around 60 species of birds are generally less honest when facing competition from distantly related nestmates. Begging honesty has mostly been studied in birds and mammals, even though the large natural range of relatedness variation makes social insects great models for understanding the evolution of offspring signals. The results we obtained by comparing larval begging in ant species with varying kin structure are generally consistent with the idea that dishonesty is more prevalent under low relatedness, when inclusive fitness costs suffered through depriving nestmates of food are low. Larvae from high and intermediate relatedness species (F. fusca, F. exsecta) begged when hungry, and thus honestly signaled their need. In contrast, larvae from low-relatedness species (F. aquilonia), showed behaviors more resembling scramble competition or dishonest signaling. However, given the small number of species studied, and potential confounding factors that could explain the species differences discussed below, further studies are necessary for strong conclusions.
Begging intensity varied with species-specific traits in complex ways. Most importantly, we did not find an overall significant effect of intra-colony relatedness on begging levels. However, this does not mean that species-specific kin structures do not influence begging. Instead, in the global analyses we may not have been able to detect species-level changes in begging behavior caused by intra-colony relatedness variation due to the large variation in our relatedness estimates, which do not perfectly reflect estimates obtained in previous studies (e.g., Sundström et al., 2003; Schultner et al., 2014, 2016). There are two main reasons that could explain why our estimates deviate from those of previous studies. First, in order to obtain enough larvae for experiments, we preferentially used multiple queen colonies for F. fusca, even though these have lower relatedness levels than single queen colonies (multiple queen colonies: r = 0.2 ± 0.18 SD, single queen colonies: r = 0.68 ± 0.12, Helanterä et al., 2011). This likely biased intra-colony relatedness toward lower values in this species. Second, we were limited by small sample sizes, which generally results in imprecise allele frequency estimates both in group level allele frequencies and the population wide background allele frequencies, thus increasing the uncertainty of relatedness estimates (Queller and Goodnight, 1989). Nevertheless, the difference in kin structure is one of the most prominent traits that characterizes our focal species, as other traits such as nesting behavior show large overlap between the species. For example, F. exsecta, F. pressilabris, and F. aquilonia can found their colonies dependently in nests of other ant species, while F. fusca often acts as a host (Seifert, 2018). In terms of colony sizes, F. fusca has the smallest colonies, with F. exsecta and F. pressilabris having intermediate and F. aquilonia by far the largest colonies. Finally, adult size is largest in F. aquilonia compared to the other three species, but large intraspecific variation both within and between colonies occurs in all species.
Two measures of begging intensity were significantly affected by the interaction between state and intra-colony relatedness in the global analyses, indicating that kin structure may modulate state-dependent begging behaviors. In addition, in the species-specific analyses, increasing relatedness generally had a negative influence on begging intensity, as expected if begging were generally correlated with kin competition. Although these effects were not always statistically significant, and suffer from the uncertainty of our relatedness estimates, this suggests that kin structure may shape begging behavior both over evolutionary time (i.e., on a species level) and over shorter time spans (i.e., on a colony level). This is surprising, as begging behavior was quantified in sibling groups, and larvae presumably had no access to information about average colony kin structure. This suggests that the relatedness effect is either mediated through a maternal effect—as the mother and the workers who tend to her do have access to information on kin structure (see e.g., Helanterä and d'Ettorre, 2015)—or factors that covary with both kin structure and brood provisioning levels. Further studies that include more species with varying kin structures and, ideally, studies that experimentally manipulate relatedness and its potential covariates such as colony size, are now needed to better comprehend the impact of kin structure on larval behavior within and between species.
Within-colony sex ratios also explained some of the variation in begging intensity (but note that the same proviso about confounding variables applies here as well). Specifically, the number and duration of swaying events was higher when larval sex ratios were male-biased in F. aquilonia. Similar differences in begging intensity between males and females have been recorded in zebra finches (von Engelhardt et al., 2006). In addition, in F. aquilonia as well as over all species combined, only non-starved larvae swayed more often in response to male-biased sex ratios. Selfishness has been shown to be more common in male Formica larvae in the context of cannibalism (Schultner et al., 2013, 2014), and males may be under selection to compete more fiercely for resources, in particular if nutritional gains increase fitness-related traits such as sperm quantity, quality (Delisle and Hardy, 1997) or adult body size (Wiernasz et al., 1995; Osawa, 2002). In addition, if competition for mating opportunities among males is stronger than among females, male-biased sex ratios may result in high levels of male-male competition. Such competition may be especially intense when larvae can afford to expend energy for begging, i.e., when they are in a non-starved state. In ants, selfish larval behavior in the form of cannibalism is predicted to depend on an interaction between kinship and sex ratio (Schultner et al., 2014), and the complex patterns observed here suggest that similar models, which take into account haplodiploidy and kin structure variation, are needed for a better understanding of begging in social insects.
One factor that may potentially confound the results is the age of larvae. Indeed, it is possible that older, starved, larvae are simply more experienced and better at begging than young, non-starved larvae. Conversely, it is possible that young, non-starved, larvae had more resources to invest in begging, resulting in more intense begging. Finally, contrasting patterns of begging intensity in the focal species may stem from differences in larval size, developmental rates or metabolism, rather than from species-specific responses to nutritional status. While these are plausible alternative explanations, there are several reasons why we think begging intensity is likely more dependent on nutritional status than larval age. First, the behavior of starved and non-starved larvae in F. pressilabris was very similar, indicating that larvae know how to beg innately and can do so regardless of their age or nutritional status. Second, in F. exsecta and F. fusca older, starved larvae begged more than young, non-starved larvae, indicating that starved larvae were not too deficient in energy to prevent them from begging. Finally, like in other insects, temperature is an important factor influencing development rates in ants (e.g., Porter, 1988). As larval rearing and behavioral assays were conducted under controlled temperature conditions, it is unlikely that larvae from different species underwent development at fundamentally different rates.
While we found clear patterns when looking at the number of swaying events, the patterns of swaying duration, and the number and duration of mandible movement, were not as clear; nevertheless, the data revealed similar trends. In Gnamptogenys striatula ants, Kaptein et al. (2005) found that starved larvae swayed significantly longer than non-starved larvae in the presence of workers. It is thus possible that swaying duration depends on the presence of workers, which reflects the difficulty of interpreting behaviors of isolated larvae. It is also possible that mandible movements are not related to begging, or that the quality of the videos did not allow us to correctly count all incidents of mandible movement.
As signaling through cuticular hydrocarbons in social insects is particularly well-developed (e.g., Blomquist and Bagnères, 2010), we hypothesized that larvae emit chemical hunger signals, and that workers should preferentially respond to odors of starved larvae. This hypothesis was based on previous studies showing that Bombus terrestris workers reacted to extracts of starved larvae with increased feeding (den Boer and Duchateau, 2006), and that female burrower bugs provisioned more when exposed to extracts from nymphs reared in low food conditions (Kölliker et al., 2005). In contrast to these results, Formica ant workers did not preferentially approach the odors of starved larvae compared to the odors of non-starved larvae. In line with this, chemical analysis of larval cuticular hydrocarbon profiles did not reveal any differences between starved and non-starved larvae. Nevertheless, there are several reasons why we cannot rule out that chemical hunger signaling plays a role in Formica ants. First, we may have overlooked potential chemical signals, for instance because relevant compounds are more polar, their concentrations lay below the detection limit, or because the compounds had lower or higher molecular weights than those included in our analysis. Second, our study was not designed to test for volatile compounds such as those known to play a role in larval chemical hunger signaling in honey bees (He et al., 2016). Third, chemical signaling by larvae may be context-dependent, and, if it is costly, may only occur in the presence of workers. Finally, workers may respond to larval chemical signals only when they are accompanied by other visual, tactile or behavioral cues. In the ant Formica cunicularia for example, workers accept heterospecific brood only when it resembles conspecific brood in both size and odor (Mori and Le Moli, 1988). Further studies are thus needed to rule out the existence of chemical hunger signals in Formica larvae.
Although we did not find any hunger-related differences in larval cuticular chemistry, chemical analyses confirmed the presence of species-typical chemical profiles (Martin and Drijfhout, 2009) in Formica ant larvae. However, some inconsistencies remain. For instance, not all F. fusca samples clustered together. This suggests that larval odors are less strong, or less species-specific, than adult odors (Carlin, 1988), or that individual colonies have very clear and distinct profiles. The last hypothesis is particularly likely for high-relatedness species, where each colony is genetically distinct. Indeed, the chemical profiles of F. fusca eggs are known to be more colony-specific than those of species with lower relatedness such as F. aquilonia (Helanterä and d'Ettorre, 2015). Chemical analyses of larval odors also highlighted an interesting phenomenon. Previous studies on cuticular hydrocarbons of ants have typically focused on eggs and adults (van Zweden and d'Ettorre, 2010). In some of these studies, individuals were placed on Petri dishes before being moved to glass vials for extraction. Our samples were handled in a similar manner, and we consistently observed contamination of larval profiles, most likely originating from Petri dishes. This may be due to the fact that, unlike eggs, which are protected by a chorion, freshly hatched larvae like those used in our experiments lack thick cuticles. In natural colonies, this may allow young larvae to pick up colony odors easily. As the only available studies of larval cuticular chemistry in ants have focused on larvae in later stages of development (Brian, 1975; Hare, 1996; Akino et al., 1999; Viana et al., 2001; Villalta et al., 2016; Penick and Liebig, 2017), further studies are required to assess how development stage-dependent differences in cuticle composition may affect odor acquisition.
Begging as a classic conflict-related trait has been largely overlooked in social insects. In particular, the importance of chemical signals as indicators of nutritional state, and the impact of relatedness on the honesty of begging signals have not yet been studied. Our study revealed that, in F. fusca and F. exsecta, starved larvae begged more frequently than non-starved larvae. In these species with predominantly high intra-colony relatedness, this can be interpreted as an honest signal of need. This is in line with what has been found in Myrmica rubra (Creemers et al., 2003) and Gnamptogenys striatula (Kaptein et al., 2005). Accordingly, G. striatula nests exhibit relatively high levels of relatedness (r = 0.65 ± 0.25 SE, Giraud et al., 2001). In M. rubra, intra-colony relatedness varies strongly depending on the study population (r = 0–0.82, Seppä and Walin, 1996). As relatedness among M. rubra larvae was not estimated in the previous study, it is difficult to draw conclusions about the influence of kin structure on begging honesty in this species. In contrast, larvae in low-relatedness species were more inclined to convey dishonest signals about their state. Thus, as predicted by inclusive fitness theory, begging in social insects may be influenced by colony kin structure. However, it remains unclear whether dishonestly signaling larvae from low-relatedness nests actually receive more food from workers as a result of increased begging, i.e., whether the dishonest signal pays off. Furthermore, our study does not allow us to infer how individual traits such as future reproductive caste may influence a larva's propensity to beg. For example, individual begging intensities may vary with species-level traits such as the degree of queen-worker dimorphism, with more begging expected to occur when the dimorphism is more pronounced. As caste of developing females has previously been shown to affect worker behavior (e.g., Brian, 1973; Passera et al., 1995; Penick and Liebig, 2012, 2017), it is furthermore possible that larval caste affects worker responses to begging.
Overall, this study provides the first test of inclusive fitness predictions in the context of hunger signaling in social insects. By demonstrating that the honesty of larval hunger signaling varies among species with different colony kin structures, our study highlights yet another social conflict in which kinship covaries with social insect behavior, thus providing further evidence that larvae are active players in colony life (Schultner et al., 2017).
Data Availability Statement
The raw data supporting the conclusions of this manuscript will be made available by the authors, without undue reservation, to any qualified researcher.
Author Contributions
MP, HH, and ES contributed conception and design of the study, and carried out field collections. MP, RL, TP, and ES collected the data. MP, TP, ES, and HH analyzed the data. MP wrote the first draft of the manuscript. MP, TP, JH, HH, and ES contributed to manuscript revision and all authors read and approved the submitted version.
Funding
This work was funded by the Academy of Finland (Grants #251337 and #284666 to the Centre of Excellence in Biological Interactions), Kone Foundation (HH) and Ella and Georg Ehrnrooth foundation (HH).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank Melanie Schlossberger for help with GC-MS sample preparation.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2019.00398/full#supplementary-material
References
Aitchison, J. (1982). The statistical analysis of compositional data. J. R. Stat. Soc. B Met. 44, 139–160. doi: 10.1111/j.2517-6161.1982.tb01195.x
Akçay, E. (2012). Incentives in the family II: behavioral dynamics and the evolution of non-costly signaling. J. Theor. Biol. 294, 9–18. doi: 10.1016/j.jtbi.2011.10.031
Akino, T., Knapp, J. J., Thomas, J. A., and Elmes, G. W. (1999). Chemical mimicry and host specificity in the butterfly Maculinea rebeli, a social parasite of Myrmica ant colonies. Proc. R. Soc. Lond. B Biol. 266, 1419–1426. doi: 10.1098/rspb.1999.0796
Bates, D., Mächler, M., Bolker, B., and Walker, S. (2014). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 6, 1–48. arXiv:1406.5823. doi: 10.18637/jss.v067.i01
Bhatkar, A., and Whitcomb, W. H. (1970). Artificial diet for rearing various species of ants. Fla. Entomol. 53:229. doi: 10.2307/3493193
Blomquist, G. J., and Bagnères, A.-G. (2010). Insect Hydrocarbons. Biology, Biochemistry, and Chemical Ecology. Cambridge, UK: Cambridge University Press.
Breteler, W. C. M. K., and Gonzalez, S. R. (1988). Influence of temperature and food concentration on body size, weight and lipid content of two Calanoid copepod species. Hydrobiologia 167–168, 201–210. doi: 10.1007/BF00026306
Brian, M. V. (1973). Caste control through worker attack in the ant Myrmica. Insect Soc. 20, 87–102. doi: 10.1007/BF02223340
Brian, M. V. (1975). Larval recognition by workers of the ant Myrmica. Anim. Behav. 23, 745–756. doi: 10.1016/0003-3472(75)90102-5
Brooks, M. E., Kristensen, K., van Benthem, K. J., Magnusson, A., Berg, C. W., Nielsen, A., et al. (2017). glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 9, 378–400. Available online at: https://journal.r-project.org/archive/2017/RJ-2017-066/index.html
Buschinger, A. (2009). Social parasitism among ants: a review (Hymenoptera: Formicidae). Myrmecol. News 12, 219–235.
Carlin, N. F. (1988). “Species, kin and other forms of recognition in the brood discrimination behavior of ants,” in Advances in Myrmecology, ed J. C. Trager (New York, NY: E. J. Brill, 267–295.
Caro, S. M., West, S. A., and Griffin, A. S. (2016). Sibling conflict and dishonest signaling in birds. Proc. Natl. Acad. Sci. U.S.A. 113, 13803–13808. doi: 10.1073/pnas.1606378113
Cassill, D. L., and Tschinkel, W. R. (1995). Allocation of liquid food to larvae via trophallaxis in colonies of the fire ant, Solenopsis invicta. Anim. Behav. 50, 801–813. doi: 10.1016/0003-3472(95)80140-5
Chapman, R. F. (1998). The Insects: Structure and Function. Cambridge, UK: Cambridge University Press.
Creemers, B., Billen, J., and Gobin, B. (2003). Larval begging behaviour in the ant Myrmica rubra. Ethol. Ecol. Evol. 15, 261–272. doi: 10.1080/08927014.2003.9522671
Delisle, J., and Hardy, M. (1997). Male larval nutrition influences the reproductive success of both sexes of the Spruce Budworm, Choristoneura fumiferana (Lepidoptera: Tortricidae). Funct. Ecol. 11, 451–463. doi: 10.1046/j.1365-2435.1997.00114.x
den Boer, S. P. A., and Duchateau, M. J. H. M. (2006). A larval hunger signal in the bumblebee Bombus terrestris. Insect Soc. 53, 369–373. doi: 10.1007/s00040-006-0883-8
Endler, J. A. (1993). Some general comments on the evolution and design of animal communication systems. Philos. T. R. Soc. B 340, 215–225. doi: 10.1098/rstb.1993.0060
Engels, S., and Sauer, K. P. (2007). Energy beyond the pupal stage: Larval nutrition and its long-time consequences for male mating performance in a scorpionfly. J. Insect. Physiol. 53, 633–638. doi: 10.1016/j.jinsphys.2007.05.003
Font, E., and Carazo, P. (2010). Animals in translation: why there is meaning (but probably no message) in animal communication. Anim. Behav. 80, 1–6. doi: 10.1016/j.anbehav.2010.05.015
Friard, O., and Gamba, M. (2016). BORIS: a free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol. Evol. 7, 1325–1330. doi: 10.1111/2041-210X.12584
Giraud, T., Blatrix, R., Poteaux, C., Solignac, M., and Jaisson, P. (2001). High genetic relatedness among nestmate queens in the polygynous ponerine ant Gnamptogenys striatula in Brazil. Behav. Ecol. Sociobiol. 49,128–134. doi: 10.1007/s002650000284
Godfray, H. C. J. (1991). Signalling of need by offspring to their parents. Nature 352, 328–330. doi: 10.1038/352328a0
Godfray, H. C. J. (1995). Signalling of need between parents and young: parent-offspring conflict and sibling rivalry. Am. Nat. 146, 1–24. doi: 10.1086/285784
Hamilton, W. D. (1964). The genetical evolution of social behaviour. II. J. Theor. Biol. 7, 17–52. doi: 10.1016/0022-5193(64)90039-6
Hannonen, M., and Sundström, L. (2003). Reproductive sharing among queens in the ant Formica fusca. Behav. Ecol. 14, 870–875. doi: 10.1093/beheco/arg069
Hare, J. F. (1996). Discrimination of nestmate larvae by the ant Leptothorax longispinosus. Can. J. Zool. 74, 2055–2061. doi: 10.1139/z96-233
He, X. J., Zhang, X. C., Jiang, W. J., Barron, A. B., Zhang, J. H., and Zeng, Z. J. (2016). Starving honey bee (Apis mellifera) larvae signal pheromonally to worker bees. Sci. Rep. 6:22359. doi: 10.1038/srep22359
Helanterä, H., and d'Ettorre, P. (2015). A comparative study of egg recognition signature mixtures in Formica ants. Evolution 69, 520–529. doi: 10.1111/evo.12590
Helanterä, H., Lee, Y. R., Drijfhout, F. P., and Martin, S. J. (2011). Genetic diversity, colony chemical phenotype, and nest mate recognition in the ant Formica fusca. Behav. Ecol. 22, 710–716. doi: 10.1093/beheco/arr037
Hodar, J. A., Zamora, R., and Castro, J. (2002). Host utilisation by moth and larval survival of pine processionary caterpillar Thaumetopoea pityocampa in relation to food quality in three Pinus species. Ecol. Entomol. 27, 292–301. doi: 10.1046/j.1365-2311.2002.00415.x
Ishay, J., and Schwartz, A. (1973). Acoustic communication between the members of the oriental hornet (Vespa orientalis) colony. J. Acoust. Soc. Am. 53, 640–649. doi: 10.1121/1.1913368
Johnstone, R. A. (2004). Begging and sibling competition: how should offspring respond to their rivals? Am. Nat. 163, 388–406. doi: 10.1086/375541
Kaptein, N., Billen, J., and Gobin, B. (2005). Larval begging for food enhances reproductive options in the ponerine ant Gnamptogenys striatula. Anim. Behav. 69, 293–299. doi: 10.1016/j.anbehav.2004.04.012
Kedar, H., Guez–Gironés, M. A., Yedvab, S., Winkler, D. W., and Lotem, A. (2000). Experimental evidence for offspring learning in parent–offspring communication. P. R. Soc. Lond. B Biol. 267, 1723–1727. doi: 10.1098/rspb.2000.1201
Klenova, A. V. (2015). Chick begging calls reflect degree of hunger in three auk species (Charadriiformes: Alcidae). PLoS ONE 10:e0140151. doi: 10.1371/journal.pone.0140151
Kölliker, M., Chuckalovcak, J. P., and Brodie, E. D. (2005). Offspring chemical cues affect maternal food provisioning in burrower bugs, Sehirus cinctus. Anim. Behav. 69, 959–966. doi: 10.1016/j.anbehav.2004.06.031
Lê, S., Josse, J., and Husson, F. (2008). FactoMineR: an R package for multivariate analysis. J. Stat. Softw. 25, 1–18. doi: 10.18637/jss.v025.i01
Laidre, M. E., and Johnstone, R. A. (2013). Animal signals. Curr. Biol. 23, R829–R833. doi: 10.1016/j.cub.2013.07.070
Leonard, M. L., and Horn, A. G. (2005). Ambient noise and the design of begging signals. P. R. Soc. Lond. B Biol. 272, 651–656. doi: 10.1098/rspb.2004.3021
Leonhardt, S. D., Menzel, F., Nehring, V., and Schmitt, T. (2016). Ecology and evolution of communication in social insects. Cell 164, 1277–1287. doi: 10.1016/j.cell.2016.01.035
Marler, P. (1961). The logical analysis of animal communication. J. Theor. Biol. 1, 295–317. doi: 10.1016/0022-5193(61)90032-7
Marques, P. A. M., Vicente, L., and Márquez, R. (2008). Iberian azure-winged magpie Cyanopica (cyana) cooki nestlings begging calls: call characterization and hunger signaling. Bioacoustics 18, 133–149. doi: 10.1080/09524622.2008.9753596
Martin, S., and Drijfhout, F. (2009). A review of ant cuticular hydrocarbons. J. Chem. Ecol. 35, 1151–1161. doi: 10.1007/s10886-009-9695-4
Mas, F., and Kölliker, M. (2008). Maternal care and offspring begging in social insects: chemical signalling, hormonal regulation and evolution. Anim. Behav. 76, 1121–1131. doi: 10.1016/j.anbehav.2008.06.011
Milne, L. J., and Milne, M. (1976). The social behavior of burying beetles. Sci. Am. 235, 84–89. doi: 10.1038/scientificamerican0876-84
Mondloch, C. J. (1995). Chick hunger and begging affect parental allocation of feedings in pigeons. Anim. Behav. 49, 601–613. doi: 10.1016/0003-3472(95)80193-6
Mori, A., and Le Moli, F. (1988). The role of different stimuli in eliciting cocoon care behaviour in Formica cunicularia ant-workers (Hymenoptera: Formicidae). Boll. Zool. 55, 93–98. doi: 10.1080/11250008809386605
Osawa, N. (2002). Sex-dependent effects of sibling cannibalism on life history traits of the ladybird beetle Harmonia axyridis (Coleoptera: Coccinellidae). Biol. J. Linn. Soc. 76, 349–360. doi: 10.1111/j.1095-8312.2002.tb01702.x
Owren, M. J., and Rendall, D. (1997). An affect-conditioning model of nonhuman primate vocal signaling. Perspect. Ethol. 12, 299–346.
Owren, M. J., and Rendall, D. (2001). Sound on the rebound: bringing form and function back to the forefront in understanding nonhuman primate vocal signaling. Evol. Anthropol. 10, 58–71. doi: 10.1002/evan.1014
Parker, G. A., and Macnair, M. R. (1979). Models of parent-offspring conflict. IV. Suppression: evolutionary retaliation by the parent. Anim. Behav. 27, 1210–1235. doi: 10.1016/0003-3472(79)90068-X
Parker, G. A., Royle, N. J., and Hartley, I. R. (2002). Begging scrambles with unequal chicks: interactions between need and competitive ability. Ecol. Lett. 5, 206–215. doi: 10.1046/j.1461-0248.2002.00301.x
Passera, L., Aron, S., and Bach, D. (1995). Elimination of sexual brood in the Argentine ant Linepithema humile: queen effect and brood recognition. Entomol. Exp. Appl. 75, 203–212. doi: 10.1111/j.1570-7458.1995.tb01928.x
Penick, C. A., and Liebig, J. (2012). Regulation of queen development through worker aggression in a predatory ant. Behav. Ecol. 23, 992–998. doi: 10.1093/beheco/ars062
Penick, C. A., and Liebig, J. (2017). A larval ‘princess pheromone' identifies future ant queens based on their juvenile hormone content. Anim. Behav. 128, 33–40. doi: 10.1016/j.anbehav.2017.03.029
Porter, S. D. (1988). Impact of temperature on colony growth and developmental rates of the ant, Solenopsis invicta. J. Insect Physiol. 34, 1127–1133.
Price, K., Harvey, H., and Ydenberg, R. (1996). Begging tactics of nestling yellow-headed blackbirds, Xanthocephalus xanthocephalus, in relation to need. Anim. Behav. 51, 421–435. doi: 10.1016/0022-1910(88)90215-6
Queller, D. C., and Goodnight, K. F. (1989). Estimating relatedness using genetic markers. Evolution 43, 258–275.
R Core Team (2018) R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. doi: 10.2307/2409206
Rauter, C. M., and Mooref, A. J. (1999). Do honest signalling models of offspring solicitation apply to insects? Proc. R. Soc. Lond. B Biol. 266, 1691–1696. doi: 10.1098/rspb.1999.0833
Rosengren, R., Sundström, L., and Fortelius, W. (1993). “Monogyny and polygyny of Formica ants: a result of alternative dispersal tactics?” in Queen Number and Sociality in Insects, ed L. Keller (New York, NY: Oxford University Press, 308–333.
Royle, N. J., Hartley, I. R., and Parker, G. A. (2002). Begging for control: when are offspring solicitation behaviours honest? Trends Ecol. Evol. 17, 434–440. doi: 10.1016/S0169-5347(02)02565-X
Schultner, E., d'Ettorre, P., and Helanterä, H. (2013). Social conflict in ant larvae: egg cannibalism occurs mainly in males and larvae prefer alien eggs. Behav. Ecol. 24, 1306–1311. doi: 10.1093/beheco/art067
Schultner, E., Gardner, A., Karhunen, M., and Helanterä, H. (2014). Ant larvae as players in social conflict: relatedness and individual identity mediate cannibalism intensity. Am. Nat. 184, E161–E174. doi: 10.1086/678459
Schultner, E., Oettler, J., and Helanterä, H. (2017). The role of brood in eusocial hymenoptera. Q. Rev. Biol. 92, 39–78. doi: 10.1086/690840
Schultner, E., Saramäki, J., and Helanterä, H. (2016). Genetic structure of native ant supercolonies varies in space and time. Mol. Ecol. 25, 6196–6213. doi: 10.1111/mec.13912
Seppä, P., and Walin, L. (1996). Sociogenetic organization of the red ant Myrmica rubra. Behav. Ecol. Sociobiol. 38, 207–217 doi: 10.1007/s002650050234
Seyfarth, R. M., and Cheney, D. L. (2003). Signalers and receivers in animal communication. Annu. Rev. Psychol. 54, 145–173. doi: 10.1146/annurev.psych.54.101601.145121
Shafiei, M., Moczek, A. P., and Nijhout, H. F. (2001). Food availability controls the onset of metamorphosis in the dung beetle Onthophagus taurus (Coleoptera: Scarabaeidae). Physiol. Entomol. 26, 173–180. doi: 10.1046/j.1365-3032.2001.00231.x
Sundström, L., Keller, L., and Chapuisat, M. (2003). Inbreeding and sex-biased gene flow in the ant Formica exsecta. Evolution 57, 1552–1561. doi: 10.1111/j.0014-3820.2003.tb00363.x
Sundström, L., Seppä, P., and Pamilo, P. (2005). Genetic population structure and dispersal patterns in Formica ants—a review. Ann. Zool. Fenn. 42, 163–177. Available online at: https://www.jstor.org/stable/23735905
Trivers, R. L. (1972). “Parental investment and sexual selection” in Sexual Selection and the Descent of Man, ed B. Campbell (Chicago, IL: Aldine-Atherton, 136–179.
van Zweden, J. S., and d'Ettorre, P. (2010). “Nestmate recognition in social insects and the role of hydrocarbons,” in Insect Hydrocarbons: Biology, Biochemistry, and Chemical Ecology, eds A.-G. Blomquist, and G.C. Bagnères (Cambridge: Cambridge University Press, 222–243.
Viana, A. M. M., Frézard, A., Malosse, C., Della Lucia, T. M. C., Errard, C., and Lenoir, A. (2001). Colonial recognition of fungus in the fungus-growing ant Acromyrmex subterraneus subterraneus (Hymenoptera: Formicidae). Chemoecology 11, 29–36. doi: 10.1007/PL00001829
Villalta, I., Amor, F., Cerdá, X., and Boulay, R. (2016). Social coercion of larval development in an ant species. Sci. Nat. 103:18. doi: 10.1007/s00114-016-1341-8
von Engelhardt, N., Carere, C., Dijkstra, C., and Groothuis, T. (2006). Sex-specific effects of yolk testosterone on survival, begging and growth of zebra finches. Proc. R. Soc. Lond. B Biol. 273, 65–70. doi: 10.1098/rspb.2005.3274
Wheeler, W. M., and Bailey, I. W. (1920). The feeding habits of Pseudomyrmine and other ants. Trans. Am. Philos. Soc. 22, 235–279. doi: 10.2307/1005485
Wiernasz, D. C., Yencharis, J., and Cole, B. J. (1995). Size and mating success in males of the western harvester ant, Pogonomyrmex occidentalis (Hymenoptera: Formicidae). J. Insect Behav. 8, 523–531. doi: 10.1007/BF01995324
Keywords: social insects, kin conflict, parent-offspring conflict, honest signal, scramble competition, relatedness, cuticular hydrocarbons
Citation: Peignier M, Pokorny T, Heinze J, Lindgren R, Helanterä H and Schultner E (2019) Honesty of Larval Begging Signals Covaries With Colony Kin Structure in Formica Ants. Front. Ecol. Evol. 7:398. doi: 10.3389/fevo.2019.00398
Received: 25 June 2019; Accepted: 08 October 2019;
Published: 23 October 2019.
Edited by:
Mark A. Elgar, The University of Melbourne, AustraliaReviewed by:
Samuel Ellis, University of Exeter, United KingdomYuko Ulrich, Université de Lausanne, Switzerland
Copyright © 2019 Peignier, Pokorny, Heinze, Lindgren, Helanterä and Schultner. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Heikki Helanterä, aGVpa2tpLmhlbGFudGVyYUBvdWx1LmZp; Eva Schultner, ZXZhLnNjaHVsdG5lckBnbWFpbC5jb20=
†These authors have contributed equally to this work