 Angelika L. Kurthen1,2,3*
Angelika L. Kurthen1,2,3* Fengzhi He1,2,4
Fengzhi He1,2,4 Xiaoyu Dong4
Xiaoyu Dong4 Alain Maasri1,5
Alain Maasri1,5 Naicheng Wu6Qinghua Cai4*
Naicheng Wu6Qinghua Cai4* Sonja C. Jähnig1,7
Sonja C. Jähnig1,7- 1Leibniz-Institute of Freshwater Ecology and Inland Fisheries, Berlin, Germany
- 2Institute of Biology, Freie Universität Berlin, Berlin, Germany
- 3Department of Integrative Biology, Oregon State University, Corvallis, OR, United States
- 4State Key Laboratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, China
- 5The Academy of Natural Sciences of Drexel University, Philadelphia, PA, United States
- 6Department of Health and Environmental Sciences, Xi’an Jiaotong-Liverpool University, Suzhou, China
- 7Geography Department, Faculty of Mathematics and Natural Sciences, Humboldt-Universität zu Berlin, Berlin, Germany
The metacommunity concept has received increasing interest in the past two decades. However, there has been limited research examining metacommunity structure of communities in high mountain streams. These ecosystems are often physically constrained and can display large environmental gradients within a relatively small spatial extent. Here, we examined metacommunity structures of stream organisms in a high mountain region, which is part of the Hengduan Mountains region in Southwest China. Macroinvertebrates and diatoms were collected from six streams in two opposite aspects of the same mountain with different connectivity between streams. On the west aspect, streams are tributaries of a river (i.e., river-connected) while streams flow into a lake (i.e., lake-connected) on the east aspect. We used Elements of Metacommunity Structure analysis to explore the metacommunity structuring of these two biological models. We also compared the contribution of dispersal and environmental filtering in structuring metacommunities by looking at Euclidean, network, topographic, and environmental distances. Communities of diatoms and macroinvertebrates were structured with clear turnover on both aspects. Further, diatom communities exhibited Clementsian structure on both aspects. Macroinvertebrates exhibited different metacommunity structures on the river-connected aspect (Quasi-Clementsian) and lake-connected aspect (Clementsian). Our results indicated that on the lake-connected aspect, environmental filtering had a stronger association with community dissimilarity than on the river-connected aspect for both macroinvertebrate and diatom communities. Diatom communities were more influenced by environmental filtering on the east aspect with weakened network connectivity compared with those on the west aspect. Our results also emphasized the potential effects of biotic interactions between macroinvertebrates and diatoms on shaping community structures of one other. Our study provides substantial elements to further understand metacommunity structure and highlights the necessity of future research to reveal the underlying mechanisms of community structuring in these remote ecosystems.
Introduction
Patterns and variation in biodiversity and their underlying mechanisms are central topics in biogeographical and ecological research (Gaston, 2000; Willig et al., 2003; Sanders and Rahbek, 2012). The metacommunity concept provides a theoretical framework for understanding biodiversity patterns and their variations and has received increasing attention from researchers in the last two decades (Leibold et al., 2004; Logue et al., 2011; Heino, 2013). Four different paradigms have been proposed to explain how spatial and environmental factors influence ecological patterns within the metacommunity, including neutral, patch dynamics, species sorting, and mass effects (Leibold et al., 2004; Logue et al., 2011). All four metacommunity paradigms are defined by dispersal, whether it is high, low, constant, or species-dependent as individual communities do not exist in isolation and are often linked to one another (Leibold et al., 2004; Logue et al., 2011).
The exchange of species between river communities is promoted by overland dispersal, dispersal along the river network, or a combination of both (Thompson and Townsend, 2006). Studies focusing on river ecosystems have emphasized the important role of river networks in influencing the dispersal of organisms (Brown and Swan, 2010; Altermatt, 2013; Maasri et al., 2018; Tonkin et al., 2018). For example, diatoms often disperse passively overland through wind or animal vectors or passively within rivers, following the flow direction, while fishes disperse actively within the river networks and are restricted to the stream corridor (Heino et al., 2015; Dong et al., 2016). Macroinvertebrates disperse through a variety of pathways including actively dispersing overland and instream, dispersing passively overland, and drifting downstream along the river network (Grönroos et al., 2013; Heino et al., 2015).
Network connectivity can therefore influence how communities are linked to one another, in turn influencing the roles that dispersal, the local environment, and local biotic interactions play in structuring them (Schmera et al., 2017; Tonkin et al., 2018), as well as the relative strength of source-sink dynamics between communities (Gundersen et al., 2001). Compared to lowland streams, streams in high mountain regions are more spatially isolated from one another by topographic barriers (Finn and Adler, 2006; Brown and Swan, 2010). Therefore, the dispersal of species in high mountain streams is often dominated by network dispersal, as opposed to overland dispersal (Wang et al., 2012; Altermatt et al., 2013; Dong et al., 2016). In addition to dispersal limitation by mountain ridges and steep valleys, the steep elevational gradient of high mountain streams can be a strong driver of change in local environmental conditions including solar radiation, riparian vegetation, water temperature, or dissolved oxygen levels (Jacobsen, 2008; Presley and Willig, 2010; Altermatt et al., 2013), in turn influencing the interaction between dispersal and environmental filtering. Several studies have demonstrated the effects of elevation-driven environment on stream communities (Wang et al., 2012; Dong et al., 2016; Tonkin et al., 2016; He S. et al., 2020). However, the role of network connectivity in structuring metacommunities of macroinvertebrates and diatoms in high mountain streams is rarely explored.
Here we aim to examine the metacommunity structures of macroinvertebrates and diatoms in high-mountain streams located in the Cangshan Mountain, a part of the Hengduan Mountains, and to understand how they are affected by network connectivity and environmental factors. Previous studies have suggested that communities in streams with large elevational gradients witness significant turnover rates (Shah et al., 2015; Tonkin et al., 2016). Turnover is therefore considered a prominent characteristic of high-mountain stream metacommunities (Presley et al., 2010; e.g., Wang et al., 2012; Shah et al., 2015; Tonkin et al., 2016). Given the large elevational gradient in our study streams, we hypothesize that turnover exists in both macroinvertebrate and diatom communities (H1).
Additionally, on the west aspect of the Cangshan Mountain, streams are connected by the Heihui River while streams on the east aspect are connected by Lake Erhai (Figure 1). The distance-decay relationships (DDRs) in stream communities, i.e., associations between community dissimilarity and environmental or physical distances (Nekola and White, 1999), could be different between two aspects due to distinct stream connectivity. We assume that macroinvertebrates flushed into the lake via drift are likely to face mortality because of environmental change (i.e., from lotic to lentic environment) or predation by fish in the lake, limiting the in-stream dispersal between streams (Brittain and Eikeland, 1988). In comparison, local environmental similarities between the Heihui River and its tributary streams, such as similar flow velocities, water depths and substrates, would allow within-river upstream dispersal into other streams for macroinvertebrates. Therefore, we expect the river-connected aspect to provide a better in-stream, network connection for dispersal of macroinvertebrates than the lake-connected aspect. Hence, we hypothesize that the contribution of physical distance, particularly network distance, to dissimilarities among macroinvertebrate communities on the lake-connected aspect to be lower than the contribution of these same distances on the river-connected aspect (H2). Diatoms, like most passively dispersing microbial organisms, do not move actively against the flow within stream and their dispersal is strongly influenced by the flow direction of the stream (Finlay, 2002; Wang et al., 2012; Dong et al., 2016). We therefore expect that their within-stream dispersal is similar between the lake-connected and river-connected aspects (i.e., diatoms are mainly flushed to downstream reaches from upstream reaches by flow). Hence, we further hypothesize that diatom communities will be structured similarly on both aspects, regardless of the weakened connectivity between streams on the lake-connected aspect (H3). Finally, emerging research has emphasized the influence of biotic interactions in structuring communities (Ohlmann et al., 2018; García-Girón et al., 2020). Considering the links between macroinvertebrates and diatoms (dispersal vector, consumer-resource relationship), we hypothesize, biotic interactions between macroinvertebrate and diatom communities have influence on the metacommunity structuring of them (H4).
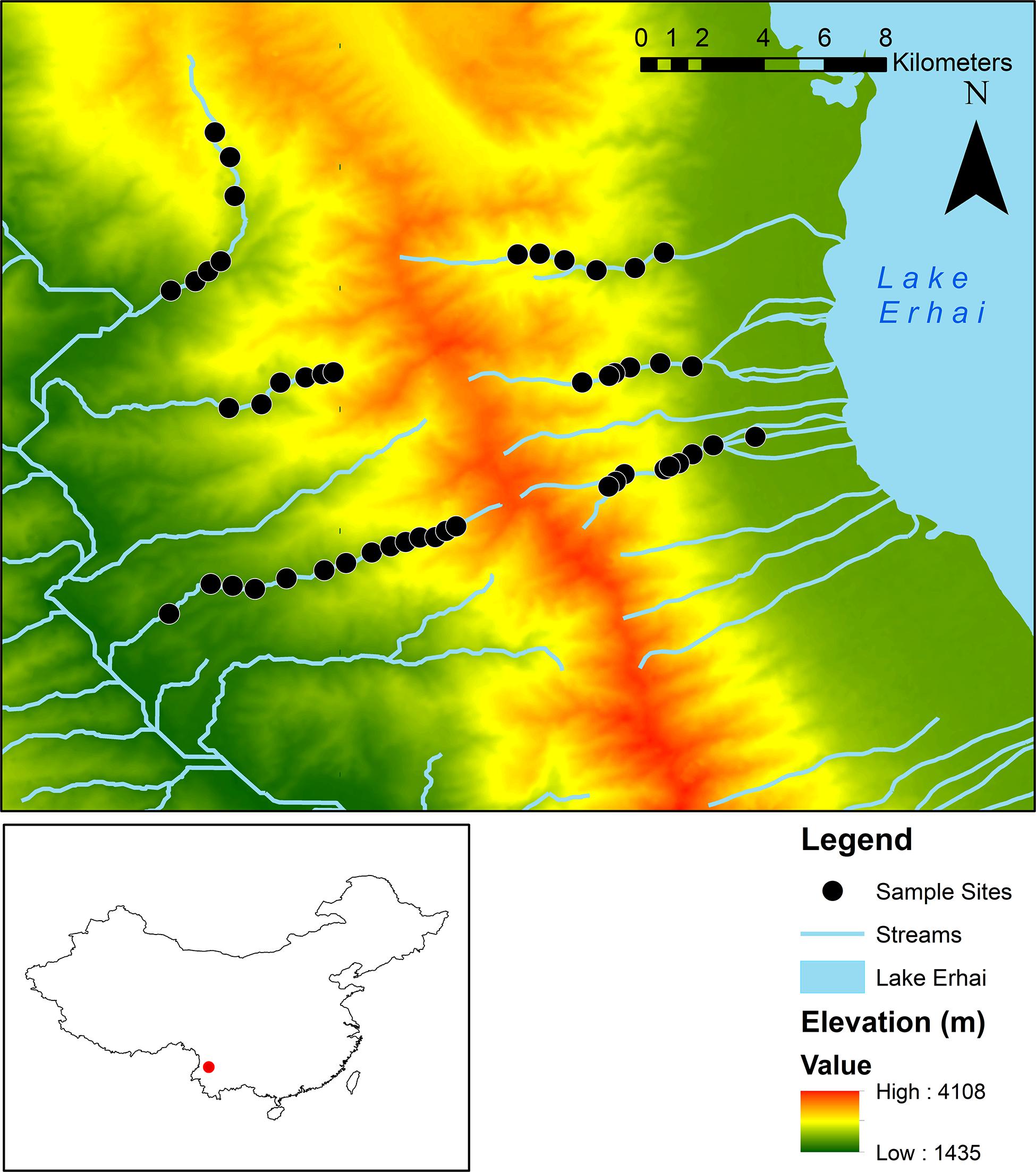
Figure 1. Map of Cangshan Mountain and sampled sites (n = 48). Study sites on the west or river-connected aspect (n = 27) had an elevational range of 1,623–2,905 m a.s.l, while study sites on the east or lake-connected aspect (n = 21) had an elevational range of 2,042–2,825 m a.s.l. The inset map depicts the study area in red, in the context of mainland China.
Materials and Methods
Field Sampling and Lab Analysis
The study area is located in the Cangshan-Erhai National Nature Reserve (25.64–25.85°N, 99.95–100.20°E), Northwest Yunnan, China. The Cangshan Mountain, which is part of the Hengduan Mountains, is characterized by a series of 19 mountain ridges with almost parallel streams running between them (Figure 1). The highest of these ridges reaches 4,122 m a.s.l. On the east aspect, streams flow into Lake Erhai, while on the west aspect, streams are connected by the Yangbi River. In addition, vegetation exhibits different elevational patterns between the east and the west aspects due to the influence of solar radiation (Sun, 2008). From here on, the east aspect will be referred to as the lake-connected aspect and the west as the river-connected aspect. The region is located in the Mekong River basin and characterized by a monsoon season from May to October and a dry season from November to April.
From November to December 2012, 48 sites were sampled from six streams (i.e., three on each aspects), ranging from 1,623 to 2,905 m a.s.l. (He F. et al., 2020). At each site, conductivity (Cond), dissolved oxygen (DO), pH, salinity (Sal), total dissolved solids (TDS), water temperature (Wtemp), and oxidation reduction potential (ORP) were measured in situ with a multiparameter probe (YSI Professional plus, US). Stream width was measured as the average of three cross sections. Depth and velocity were measured along a transect every 50 cm using a water flow probe and averaged (Global Water FP201, US). Geographical coordinates and elevations of sampling sites were determined using a portable GPS device (Magellan 500E, US). Water samples were collected and preserved using sulfuric acid to keep pH under 2. In the lab, total phosphorus (TP), orthophosphate (PO43–), total nitrogen (TN), nitrate (NO3–), ammonium (NH4+), and dissolved silica (SiO2) were measured using a segmented flow analyzer (Skalar San ++, Netherlands).
Macroinvertebrate samples were collected using a Surber net (30 × 30 cm, 500 μm). Five sub-samples were taken at each site to cover multiple habitat types. Macroinvertebrates were identified following Morse et al. (1994) and literature in the lab. All macroinvertebrates were identified to genus level, except for Chironomidae, which was classified to the subfamily, and Oligochaeta, Turbellaria, and Nematoda, to the class level.
Benthic diatom samples were collected from pebble or cobble substrates. A cap with a radius of 2.7 cm was placed over the substrate. The periphyton around the cap was removed with a nylon brush and flushed away. Then the periphyton under the cap was collected. In the lab, samples were digested using acid and examined at a magnification of 1,000 using oil immersion (Olympus CX21, Japan). Diatoms were classified to the species level using Krammer and Lange-Bertalot (1986, 1988, 1991a, 1991b) and Qi et al. (1995, 2004a,b, 2009, 2013). More details are described in Dong et al. (2016).
Data Analysis
Sites were grouped by aspect (i.e., a river-connected network on the west aspect and a lake-connected network on the east aspect). Collinearity between variables was tested using a pairwise Spearman correlation. The threshold of r = | 0.7| (Dormann et al., 2012) were used to exclude highly correlated variables (e.g., TDS, pH, PO43–, NO3–, NH4+, and SiO2) for further analysis. Taxon richness and dominant diatom and macroinvertebrate taxa were calculated for each aspect, as well as each individual stream sampled.
In order to determine the metacommunity structure on different aspects, an Elements of Metacommunity Structure (EMS) analysis was performed using the function Metacommunity in the package metacom (Dallas, 2018) in R (R Core Team, 2017). The metacom function runs through a three-step analysis with species-site data, in which coherence, species turnover, and boundary clumping are tested to determine the idealized metacommunity structure. Coherence is measured by comparing species absences in the data to a null-model checkerboard presence-absence matrix using a z-score. A significant negative coherence indicates a checkerboard structure, a non-significant coherence represents random coherence, and a significant positive coherence suggests a structure other than checkerboard or random. Turnover, or species replacement, is tested against a null model without species replacement. A significant positive z-score suggests significant species turnover, while a significant negative z-score indicates a nested metacommunity structure. Non-significant z-scores are treated ambiguously and are considered quasi-structures. Boundary clumping measures whether the metacommunity exhibits individual or grouped turnover and is measured using Morista’s Index (MI). A mean MI value significantly greater than one represents clumped or Clementsian metacommunity structure while a mean MI value significantly smaller than one indicates an evenly-spaced metacommunity structure. Metacommunities with a mean MI value not significantly different from one are thought to have Gleasonian structure (Dallas, 2018). The site-scores were extracted from the primary EMS axis and Spearman correlations were run to identify which environmental variables provide the structuring force behind the EMS. Although it has limitations, the use of EMS to fit community data to a theoretical pattern is widespread and of interest due to the small study area we sampled (approximately 100 km2 on the river-connected aspect, approximately 50 km2 on the lake-connected aspect).
Four different matrices were calculated to measure distances between sites, including environmental, Euclidean, topographic, and network distances. The environmental distances between sites were determined with log10-transformed environmental variables with the package vegan (Oksanen et al., 2019). Euclidean distance between sites was calculated using the earth.dist function in the package fossil (Vavrek, 2011) while topographic distance was calculated using CIRCUITSCAPE v. 4.0 with a 30 m digital elevation model (DEM). Detailed methods can be found in Dong et al. (2016). The network distance was calculated in QGIS (version 2.8; QGIS Development Team, 2015). On the lake-connected aspect, streams are connected by a lake and not a river. Because some macroinvertebrates can live in the littoral zones and flying insects can still actively disperse along the lake’s edge, some degree of connectivity could still be maintained (Bagge, 1995). Although we realize that stream-lake connection provides a weaker dispersal path for macroinvertebrates than the connection between a stream and river, for the sake of comparison, the edge of the lake was still regarded as a network path for both macroinvertebrates and diatoms. With abundance data, the community dissimilarity between sites was calculated with Bray-Curtis index with the package vegan. To determine the correlation between each of the four distances and community dissimilarity, we performed Mantel tests using Spearman’s rank-order correlation with 999 permutations.
We further analyzed the association between community dissimilarity and all measured distances using a distance-decay relationships (DDRs). We plotted the site-values of community dissimilarity against the site-values of physical (Euclidean, Topographic, Network) distances and environmental distances, conducting a logarithmic regression to determine which model best described the relationship between community dissimilarities and distances. To disentangle contributions of different distance matrices to community dissimilarity, partial Mantel tests were conducted using Spearman’s rank-order correlation also with 999 permutations. Partial Mantel tests allow for the further analysis of Mantel tests and can be used to unmask the effects of different distances matrices that could be auto-correlated (Moritz et al., 2013). The associations between community dissimilarity and the three physical distances were adjusted while controlling environmental distance. The association between community dissimilarity and environmental distance was also adjusted to control for the influence of each physical distance. Partial Mantel tests were conducted in using the package ecodist (Goslee and Urban, 2007).
Moreover, we investigated the potential influence of biotic interactions between macroinvertebrates and diatoms on community structuring of these two organisms by conducting partial Mantel tests. We examined the associations between community dissimilarity and physical distance controlling for environmental distance and the other taxa community dissimilarity, as well as a partial Mantel test to analyze the association between community dissimilarity physical distance controlling for environmental distance and the other taxa community dissimilarity. We also included a partial Mantel test to determine the association between macroinvertebrate community dissimilarity and diatom community dissimilarity, controlling for all physical and environmental distances.
The elevational range of sampling sites on the river-connected aspect (1,623–2,905 m a.s.l.) is almost twice that of the lake-connected aspect (2,042–2,825 m a.s.l.). To test whether there was any bias caused by difference in elevational range, the EMS, Mantel tests, and partial Mantel tests were also conducted on sites within the common zone (sites with an elevation between 2,042 and 2,825 m a.s.l. on both aspects).
Results
In total, 70 macroinvertebrates and 74 diatom taxa were identified on the lake-connected aspect. On the river-connected aspect, 79 macroinvertebrate taxa and 119 diatom taxa were identified. Dominant taxa were similar on both aspects. For macroinvertebrates the most abundant taxa were Baetis spp., Baetiella spp., and Orthocladiinae spp. For diatoms, the most abundant taxa were Achnanthidium minutissimum (Kützing) Czarnecki, Anchnathidium rivulare Potapova & Ponader, and Cocconeis placentula Ehrenberg.
Diatom communities on both aspects, as well as macroinvertebrate communities on the lake-connected aspect exhibited Clementsian metacommunity structure (significant positive coherence, significant turnover, significant boundary clumping; Table 1). The macroinvertebrate community on the river-connected aspect had a Quasi-Clementsian metacommunity structure (significant positive coherence, non-significant turnover, significant boundary clumping). In the macroinvertebrate communities on the lake-connected aspect, EMS site scores were significantly correlated with conductivity (r = −0.66, p < 0.01), velocity (r = −0.62, p < 0.01), DO (r = 0.55, p = 0.01), and water temperature (r = −0.51, p = 0.02). All the aforementioned environmental variables are significantly correlated with elevation on the lake-connected aspect. Water temperature (r = 0.77, p < 0.01), conductivity (r = 0.55, p < 0.01, TN (r = 0.55, p < 0.01), TP (r = 0.46, p = 0.02), and velocity (r = 0.45, p = 0.02) were all significantly correlated with macroinvertebrate community EMS site scores on the river-connected aspect. Elevation was significantly correlated with conductivity, water temperature, and TN on the river-connected aspect. The EMS site scores of the diatom communities on the lake-connected aspect were not significantly correlated with any environmental variables or elevation. On the river-connected aspect, diatom community EMS site scores were significantly correlated with conductivity (r = −0.50, p < 0.01). In the common zone analysis, EMS results were similar with the whole range sites, with diatom and macroinvertebrate communities exhibited Clementsian metacommunity structure on both aspects (Supplementary Table S1).
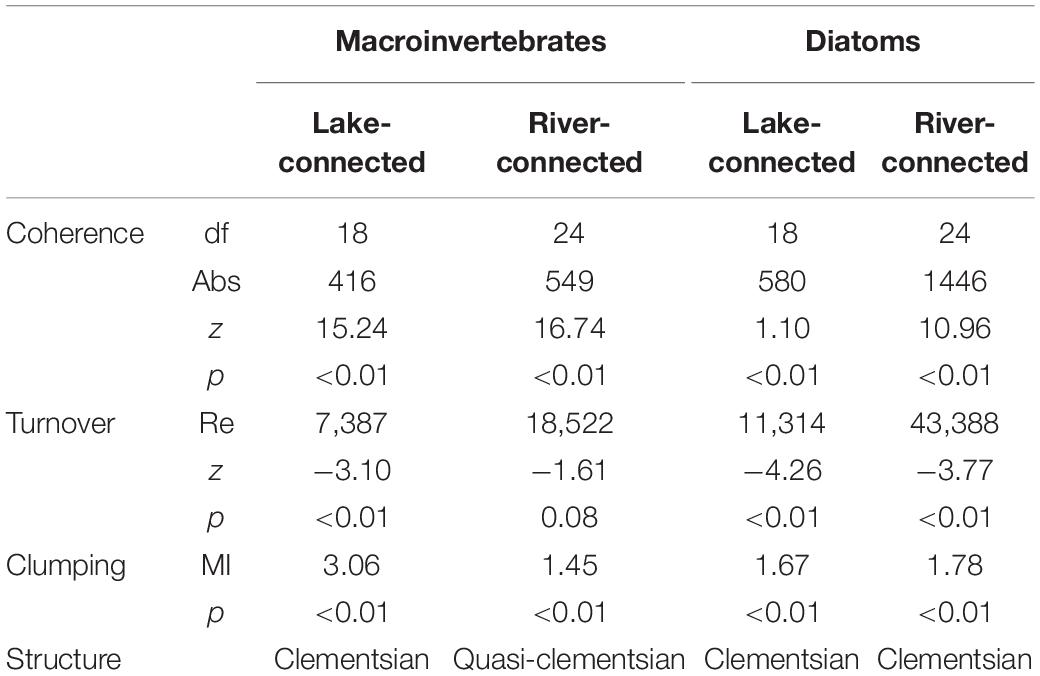
Table 1. Elements of metacommunity structure (EMS) data for macroinvertebrates and diatoms on both aspects.
For both diatom and macroinvertebrate communities, there was were significant positive Distance Decay Relationships (DDRs) between Bray-Curtis dissimilarity and all distances based on Mantel tests (Figure 2 and Table 2), except for topographic distance on the lake-connected aspect. The DDRs in common zone showed the same patterns as in the whole range for both the diatom and macroinvertebrate communities (Supplementary Figure S1 and Supplementary Table S2).
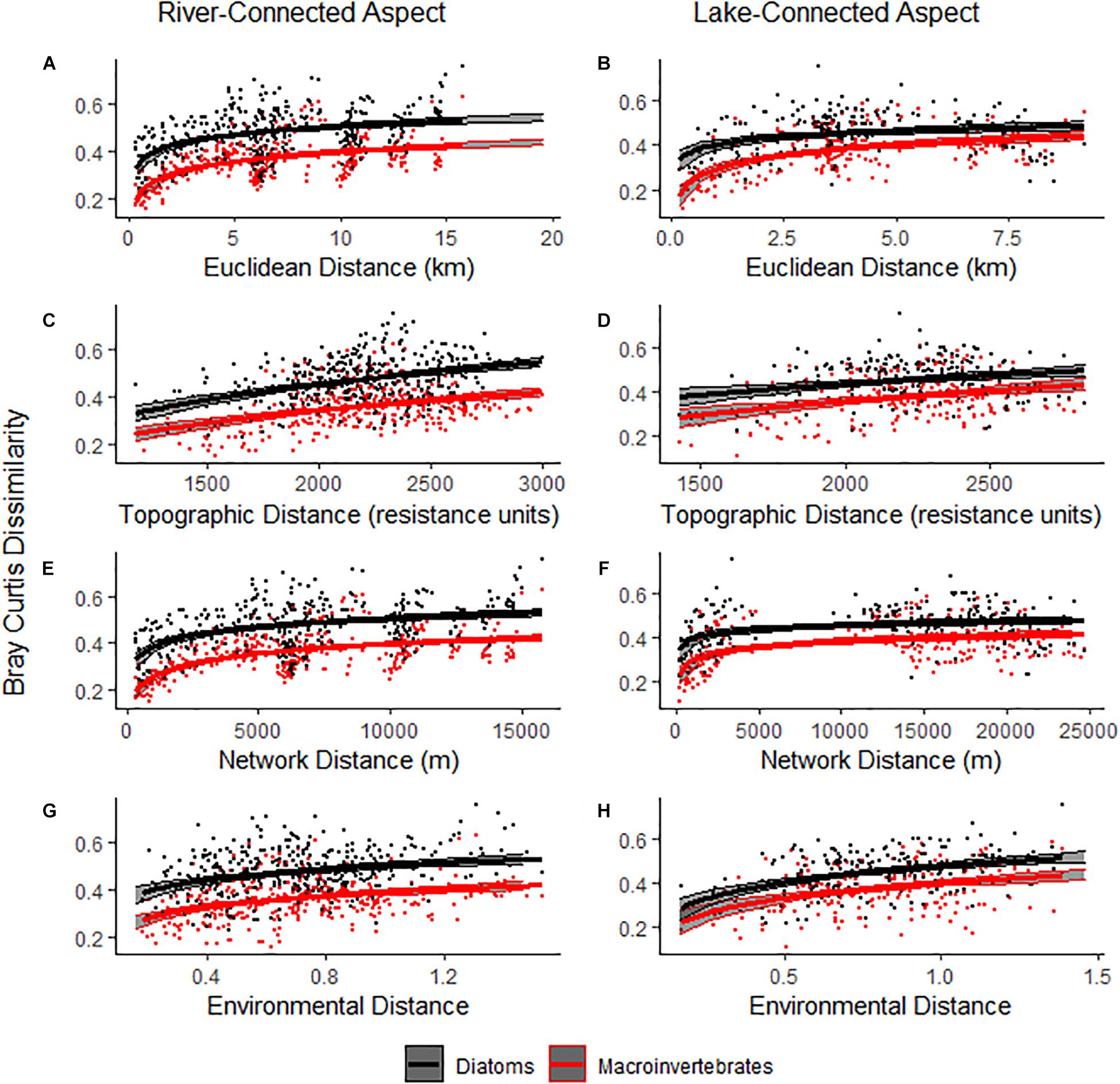
Figure 2. Distance Decay Relationships (DDR) of macroinvertebrates and diatoms. Points represent individual distances between two given sites, while lines represent the logarithmic regression for each group of points. Macroinvertebrates are represented by red points and lines, while diatoms are represented by black points and lines. The lines shown in the figures are based on logarithmic models. Solid lines represent significant Mantel tests, while dashed lines represent insignificant Mantel tests. The gray area surrounding the logarithmic regressions represents a 95% CI. Formulas and R2 values for each regression can be found in Supplementary Table S3.
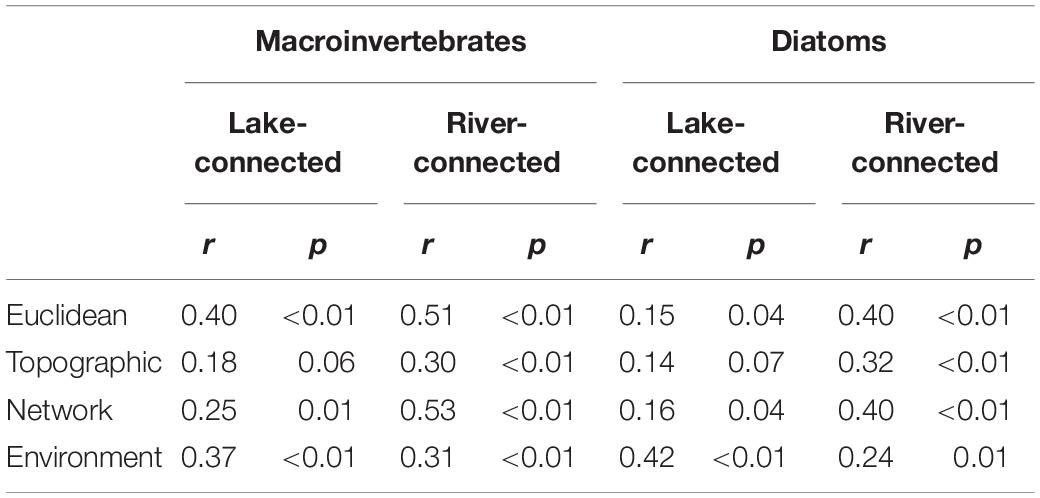
Table 2. Mantel test between community dissimilarity (Bray-Curtis dissimilarity) and physical and environmental distances for macroinvertebrates and diatoms.
In the whole range analysis, Euclidean DDR and network DDRs had higher R2 values that topographic DDRs for diatoms and macroinvertebrates on both aspects. When only analyzing sites in the common zone, the R2 values of topographic DDRs, Euclidean DDRs and network DDRs on the river-connected aspect were similar (Supplementary Table S4).
Partial Mantel tests showed that macroinvertebrate community dissimilarity on the lake-connected aspect had a significant association with Euclidean distance when adjusted for environmental distance (r = 0.40, p < 0.01, Table 3), as well as significant associations with environmental distance when adjusted for all three physical distances (Euclidean distance, r = 0.37, p < 0.01; topographic distance, r = 0.35, p < 0.01; network distance, r = 0.36, p < 0.01). On the river-connected aspect, macroinvertebrate communities had significant associations with all three physical distances when adjusted for environmental distance, with Euclidean distance (r = 0.44, p < 0.01) having the greatest correlation coefficient, followed by the network distance (r = 0.43, p < 0.01) and topographic distance (r = 0.21, p = 0.01). Additionally, there was a significant association between macroinvertebrate community dissimilarity on the river-connected aspect and environmental distance adjusted for topographic distance (r = 0.22, p = 0.02, Table 3). On the river-connected aspect of the common zone analysis, macroinvertebrate communities were significantly associated with all adjusted distances except for environmental distance adjusted for Euclidean distance and environmental distance adjusted for network distance (Supplementary Table S5).
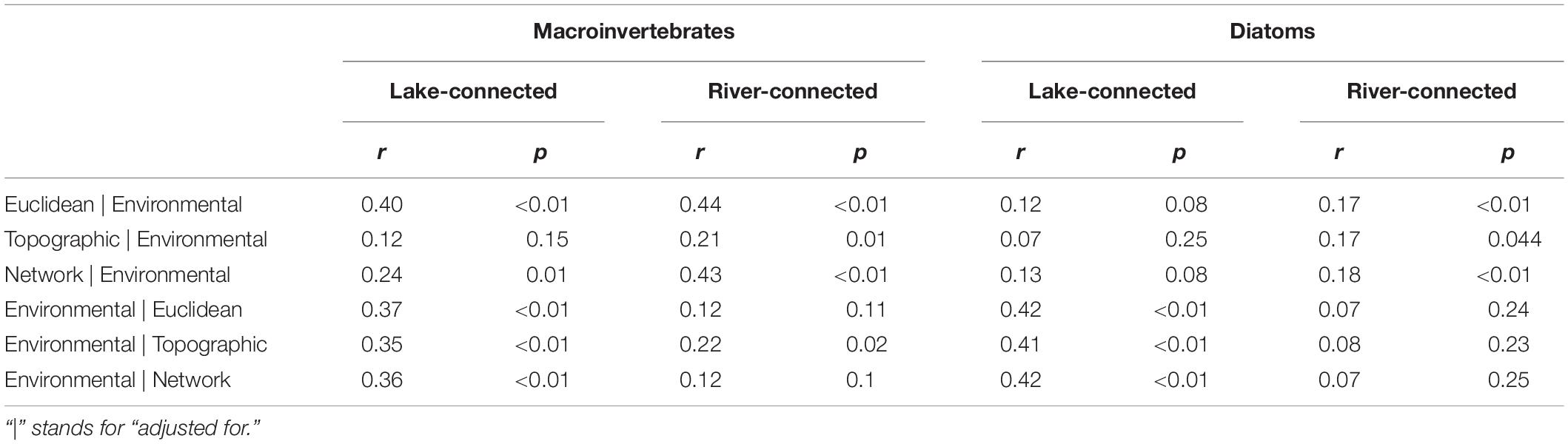
Table 3. Partial mantel tests of Bray-Curtis dissimilarity using Spearman’s rank correlation for macroinvertebrate communities on the lake-connected and river-connected aspects and diatom communities on the east and west aspects.
Compared to macroinvertebrates, major differences in the results of the partial Mantel test for diatoms were observed between the lake- and river-connected aspects. Diatom community dissimilarity on the lake-connected aspect was significantly associated with environmental distances adjusted for all physical distances (adjusted for Euclidean distance, r = 0.42, p < 0.01; topographic distance, r = 0.41, p < 0.01, network distance, r = 0.42, p < 0.01, Table 3). On the river-connected aspect, diatom community dissimilarity was significantly associated with all physical distances when adjusted for environmental distance (Euclidean distance, r = 0.17, p < 0.01; topographic distance, r = 0.17, p < 0.01; network distance, r = 0.18, p < 0.01). The common zone analysis of diatom communities on the lake-connected aspect was similar to that of the whole range analysis (Supplementary Table S1). On the river-connected aspect of the common zone analysis, diatom community dissimilarity was significantly associated with Euclidean distance adjusted for environmental distance (r = 0.35, p < 0.01), topographic distance adjusted for environmental distance (r = 0.42, p < 0.01) and network distance adjusted for environmental distance (r = 0.34, p < 0.01).
We found a significant association between macroinvertebrate and diatom community dissimilarity on both the lake-connected (r = 0.31, p < 0.01) and river-connected aspect (r = 0.33, p < 0.01, Tables 4, 5), when corrected for all other physical distances and environmental distance. In the common zone analysis, macroinvertebrate and diatom dissimilarities were significant on the lake-connected aspect (r = 0.31, p < 0.01) but not on the river-connected aspect (r = 0.2, p = 0.08, Supplementary Tables S6, S7).
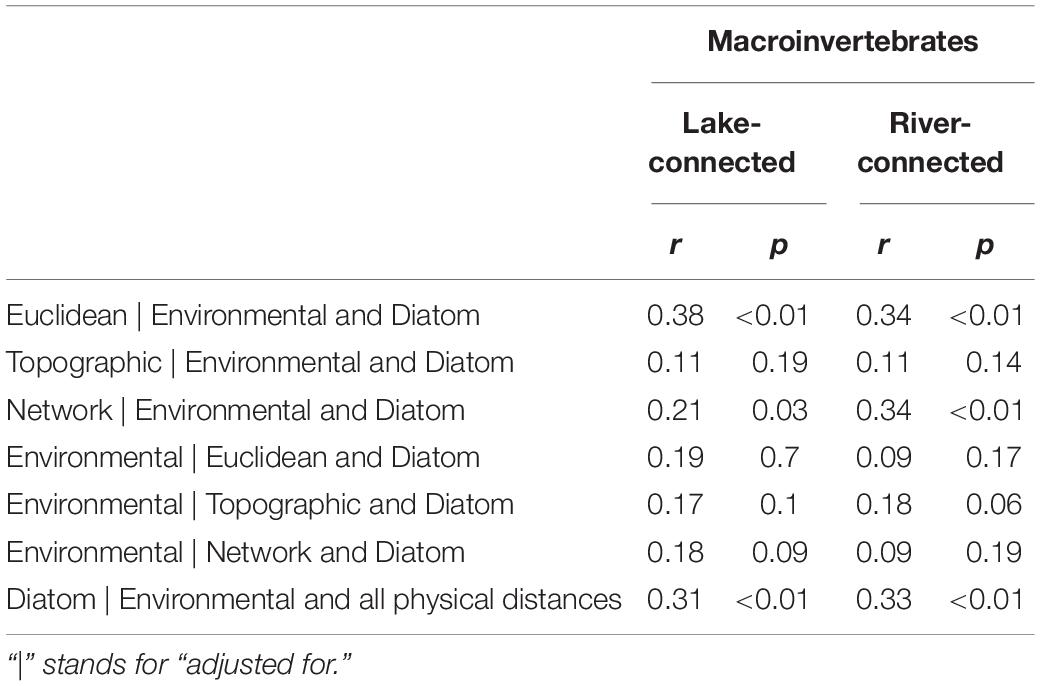
Table 4. Partial mantel tests of Bray-Curtis dissimilarity using Spearman’s rank correlation for macroinvertebrate communities on the lake-connected and river-connected aspects of the whole range.
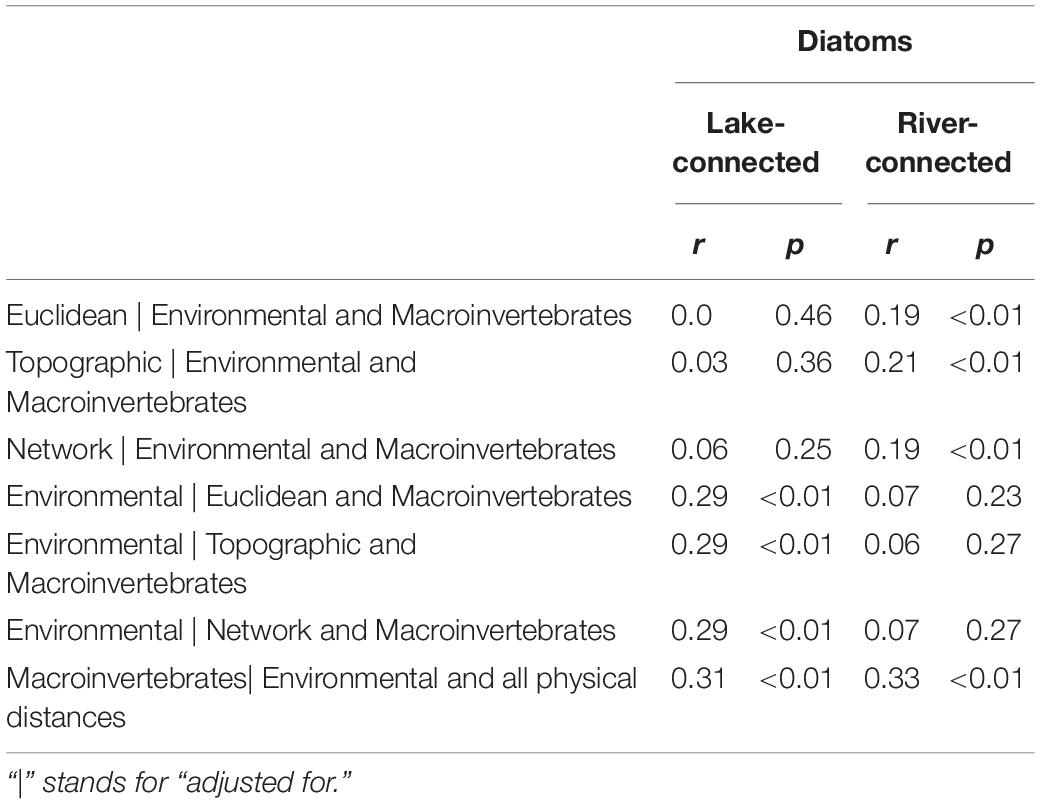
Table 5. Partial mantel tests of Bray-Curtis dissimilarity using Spearman’s rank correlation for diatoms communities on the lake-connected and river-connected aspects on the whole range.
We found significant associations between Euclidean distance and network distance and macroinvertebrate community dissimilarity on both river- and lake-connected aspects, when controlled for environmental distance and diatom dissimilarity (Table 4). However, no significant association between macroinvertebrate community dissimilarity and environmental distances was observed when effects of physical distance and diatom dissimilarity were considered (Table 4). On the river-connected aspect, diatom community dissimilarity was significantly associated with all physical distances controlled for environmental distance and macroinvertebrate community dissimilarity (Table 5). On the lake-connected aspect, diatom community dissimilarity was only significantly associated with environmental distance adjusted for physical distance and macroinvertebrate community dissimilarity (Table 5). These observed associations suggested the influence of biotic interactions on community structuring of both macroinvertebrate and diatom communities.
Discussion
Metacommunity Turnover
Turnover was observed in both stream macroinvertebrates and diatoms communities in the Cangshan Mountain, supporting our first hypothesis (H1). This finding stands in line with previous studies focusing on communities of high-mountain stream systems with large elevational gradients (Shah et al., 2015; Tonkin et al., 2016). A large elevational gradient is often coupled with changes in local environmental variables, like DO, water temperature, conductivity, riparian vegetation, wind exposure, solar radiation, and precipitation (Townsend et al., 2003; Körner, 2007; Jacobsen, 2008; Sundqvist et al., 2013; Willig and Presley, 2015; Kim and Lee, 2017). Local environmental conditions play a critical role in determining which organisms can be found at a given site through species sorting or environmental filtering. For example, macroinvertebrate species richness showed a negative association with elevation in Andes because of the sub-lethal effects of low DO levels in high-elevational areas, like lowered metabolism (Jacobsen, 2008). Kim and Lee (2017) described a change in diatom communities due to decreases in water temperature, pH, and total phosphorus associated with increases in elevation and He F. et al., 2020 also found that elevation is important in structuring diatoms species in China.
Moreover, the observed Clementsian and Quasi-Clementsian metacommunity structuring suggests that there is a grouped community response to changes in the local environmental variables (Leibold and Mikkelson, 2002). That is, species pools are thought to replace each other, suggesting either trait-related grouping based on dispersal related processes or environmental filtering or groupings related to biotic interactions between species, or some combination of forces (Presley et al., 2010; Heino et al., 2015). Other studies focused on freshwater organisms have found significant Clementsian structuring at larger scales, where Clementsian structuring is likely caused by multiple different ecotones and species pools within the study area (Tonkin et al., 2015, low mountain stream macroinvertebrates, 20–1,000 km2; Tonkin et al., 2016, high mountain macroinvertebrates, 400–650 km2; Heino et al., 2017, aquatic insects and diatoms, 63,609 km2).
The Clementsian structuring in a smaller study area like ours is likely caused by the large elevational gradient and the associated heterogeneous environmental gradients, where multiple taxon-pools can occur within a small area. Clementsian metacommunity structures indicate significant turnover and niche-based, grouped, species sorting (Leibold and Mikkelson, 2002). Instead of constant turnover, like in Clementsian structure, Quasi-Clementsian structures is thought to exhibit turnover at the ends of the environmental gradient, resulting in insignificant turnover, but still have significant grouped, niche-based sorting occurring (Presley et al., 2010). This aligns with our results, which show that for the macroinvertebrate community on the river-connected aspect, the whole range metacommunity has a Quasi-Clementsian structure while the common-zone has a Clementsian structure. The switch between Quasi-Clementsian and Clementsian could be caused by the removal of low-elevation communities, which may be more homogenous due to human impacts and have less turnover than communities in higher elevations.
Influence of Connectivity on Dispersal and Metacommunities
The significant relationships between community dissimilarity and physical distances confirm the role of dispersal in structuring metacommunities in high-mountain streams, where source-sink dynamics are limited (Göthe et al., 2013; Heino et al., 2015). Additionally, significant relationships between environmental distance and community dissimilarity often signal species sorting dynamics (Leibold et al., 2004), which was also observed in our study. Hence, it is likely that dispersal limitation and species sorting jointly shape metacommunities of stream macroinvertebrates and diatoms in the Cangshan Mountain. The metacommunity paradigms described by Leibold et al. (2004) are not mutually exclusive and combinations of more than one paradigm are common (Thompson and Townsend, 2006; Brown and Swan, 2010; Grönroos et al., 2013; Brown et al., 2017).
In stream ecosystems, connectivity of dendritic networks and dispersal ability through that network play an important role in structuring metacommunities (Altermatt and Fronhofer, 2017; Hayes and Anderson, 2017; Tonkin et al., 2018). When the connectivity between communities is weakened, dispersal pathways are disrupted and the communities become more isolated (Cañedo-Argüelles et al., 2015). Consequently, the influence of environmental filtering increases as more isolated communities are more strongly structured by local environmental factors (Brown and Swan, 2010; Cañedo-Argüelles et al., 2015). Studies focused on European high mountain lakes, which are more isolated than high mountain streams and rivers, have found that local environmental factors are more important in structuring macroinvertebrate communities than spatial factors (Kernan et al., 2009; de Mendoza and Catalan, 2010; de Mendoza et al., 2015). On the lake-connected aspect of the Cangshan Mountain, the interaction between dispersal and environmental filtering on river metacommunities has been altered compared to the river-connected aspect. The Mantel tests showed that environmental distance exhibited stronger correlation with community dissimilarity on the lake-connected aspect than on the river-connected aspect for macroinvertebrates, and the partial Mantel tests further supported this result. This verified our second hypothesis (H2) that environmental factors contribute more to the community dissimilarity of macroinvertebrates when connectivity between streams is weakened.
Our results contradicted our third hypothesis (H3) that diatom communities would be more similarly structured on the differently connected aspects than macroinvertebrates. This hypothesis was based on the differences in dispersal capabilities of macroinvertebrates and diatoms. Apart from in-stream passive dispersal via flow, many macroinvertebrates can move actively along the network corridor and fly overland as well. Flying adult insects can disperse along the lake shore from confluence to confluence, as well as along the littoral zone of the lake (Bagge, 1995), contributing, therefore, to the dispersal between streams on the lake-connected aspect. Thus, there is weakened network connectivity between the macroinvertebrate communities in streams on the lake-connected aspect compared to the river-connected aspect.
Compared to macro-organisms such as macroinvertebrates, diatoms are efficient and ubiquitous passive dispersers like other micro-organisms (Finlay, 2002). The Baas-Becking hypothesis, i.e., diatoms are ubiquitous dispersers and their biodiversity patterns are structured by local environmental conditions (Baas Becking, 1934; Finlay, 2002), has been challenged in recent research. Both lowland and high mountain stream diatom biodiversity is thought to be shaped not only by local environmental factors, but by dispersal-related spatial processes as well (Potapova and Charles, 2002; Soininen, 2004; Martiny et al., 2006; Bottin et al., 2014; Dong et al., 2016). In addition, Vilmi et al. (2020) observed more stochastic patterns in stream micro-organism (i.e., bacteria) than in macro-organism (i.e., macroinvertebrates) in the Hengduan Mountains. Our results suggest that diatom communities in high mountain streams are influenced by both spatial and environmental factors (Wang et al., 2012; Dong et al., 2016). Similar results from high mountain lakes in Europe and in the Andes have also shown that diatom assemblages are structured by both local environmental and spatial factors, although local environmental factors are more important in structuring lentic diatoms than geographical factors in Europe (Kernan et al., 2009; Benito et al., 2018). Diatoms are fundamentally limited to passive dispersal, either depending on wind or biotic vectors for overland dispersal or dispersing along the network in the direction of flow (Heino et al., 2015; Dong et al., 2016). Barriers, like the presence of a lake, may affect diatom dispersal. For example, diatoms flushed into lakes via in-stream drift may be unable to tolerate lentic conditions, as lake and stream diatom communities are known to be different from one another (Soininen and Weckström, 2009). This may limit in-stream dispersal to adjacent lotic habitats, although some dispersal may be possible via wind or other animal vectors (Kristiansen, 1996). However, topographical barriers can affect the dispersal of vectors themselves in high mountain ecosystems. For example, macroinvertebrates are important vectors for diatom dispersal (Kristiansen, 1996). If macroinvertebrate dispersal is weakened in high mountain streams, this could weaken the dispersal ability of diatoms. In the Hengduan Mountains, small-mammal communities had high turnover and were found to be dispersal limited by topographical and environmental barriers (Wen et al., 2016). Additionally, Adams et al. (2000) found that Brook Trout with a small body size in the western USA had limited upstream dispersal ability in headwater streams with steep slopes. Thus, the dispersal limitation of important vectors like macroinvertebrates, aquatic mammals, and fish could affect the dispersal of diatoms, in turn, posing impact on metacommunity structuring of diatoms.
Our results showing significant associations between macroinvertebrate and diatoms community dissimilarity when adjusted for all other distances support our hypothesis that biotic interactions significantly affect the community structure of aquatic organisms in the Cangshan Mountain (H4). Macroinvertebrate and diatom communities can interact with one another through producer-consumer relationships, as grazer macroinvertebrates consume diatoms in the periphyton, which could lead to the significant association between macroinvertebrate and diatom dissimilarity (Thompson et al., 2020). Additionally, as mentioned above, macroinvertebrates are a dispersal vector for diatoms, which could lead to co-occurrence. However, co-variation between two groups of taxa, as in our results, could also be caused by responses to abiotic filtering (Zhao et al., 2019; García-Girón et al., 2020) not examined in this study. Previous studies have found that in addition to abiotic factors, like landscape structure and local environmental conditions, biotic interactions can significantly shape metacommunities. For example, macrophyte communities were found to structure filter-feeding zooplankton and predacious macroinvertebrate communities in pond ecosystems (García-Girón et al., 2020). However, we found that when comparing the whole range to the common zone, the association between macroinvertebrate and diatom community dissimilarity is no longer significant. There could be an underlying environmental gradient to which diatoms and macroinvertebrates respond, for example, a shortened gradient of water temperature or DO gradient (Jacobsen, 2008; Kim and Lee, 2017). If part of the underlying gradient is not included, it is possible that community structure responses or biotic interactions will no longer be significant. In addition to underlying abiotic and biotic gradients, the potential spatial autocorrelation between distance matrices could also have impacts on our results. For example, the presence of spatial autocorrelation between distance matrices may cause Mantel test results to have an inflated Type I error, that is, rejecting the null hypothesis even though it is true (Guillot and Rousset, 2013). Partial Mantel tests are often used to control for an underlying spatial matrix, but research has suggested that partial Mantel tests may not be adequate for controlling for spatial autocorrelation (Guillot and Rousset, 2013; Crabot et al., 2019). Future research focused on how abiotic (physical distance, local environment) and biotic (competition, predation, symbiosis) factors interact will provide more insight to the different roles that each factor plays in shaping metacommunities (Chiu et al., 2020).
Summary and Outlook
We explored metacommunity structures of macroinvertebrate and diatom communities in the Cangshan Mountain and found that like many other high mountain stream communities, both macroinvertebrate and diatom communities exhibit a clear turnover. Furthermore, we found that weakened network connectivity resulted in environmental variables being stronger structuring forces than spatial factors. Macroinvertebrate metacommunities were jointly shaped by dispersal limitation and environmental filtering in networks with weakened connectivity while environmental filtering showed a stronger influence on diatom metacommunities than dispersal limitation. We found significant associations between diatom and macroinvertebrate community dissimilarity, however, the concrete cause of this association is speculative. Disentangling the local metacommunity structure and the influence of spatial and environmental factors on local areas provides valuable insight into the structuring forces of this high biodiversity region and its vulnerability to human impacts (Chase et al., 2020) and can help improve biological assessments and conservation in these dynamic ecosystems (Cid et al., 2020). Future research should expand metacommunity analysis to include other high mountain areas and use replicated metacommunities at multiple spatial scales to disentangle drivers of this variability for different organismal groups.
Data Availability Statement
Publicly available datasets were analyzed in this study. This data can be found at: https://doi.org/10.1016/j.scitotenv.2020.140548; https://doi.org/10.1038/srep24711.
Author Contributions
FH, AM, QC, and SJ conceived the research idea. FH and XD collected the data. AK conducted the data analysis, with assistance from FH and AM. AK, FH, and AM drafted the manuscript. All authors contributed to the manuscript revision.
Funding
This work was supported by the National Natural Science Foundation of China (U1602262) and the German Federal Ministry of Education and Research (BMBF) within the “GLANCE” project (Global Change Effects in River Ecosystems; 01 LN1320A). FH was supported by the SMART Joint Doctorate (Science for the MAnagement of Rivers and their Tidal systems), funded with the support of the Erasmus Mundus programme of the European Union. AM was additionally supported by the US NSF Macrosystems Grant No. 1442595. We acknowledge support by the Open Access Publication Initiative of Freie Universität Berlin.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Meiqing Sun, Haomiao Zhang, Bin Li, Yuan Gu, Wen Xiao, Yihao Fang, and Yun Zhou for their help with field sampling and lab analysis.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2020.571887/full#supplementary-material
References
Adams, S. B., Frissell, C. A., and Rieman, B. E. (2000). Movements of nonnative brook trout in relation to stream channel slope. Trans. Am. Fish. Soc. 129, 623–638. doi: 10.1577/1548-8659(2000)129<0623:monbti>2.3.co;2
Altermatt, F. (2013). Diversity in riverine metacommunities: a network perspective. Aquat. Ecol. 47, 365–377. doi: 10.1007/s10452-013-9450-3
Altermatt, F., and Fronhofer, E. (2017). Dispersal in dendritic networks: ecological consequences on the spatial distribution of population densities. Freshw. Biol. 63, 22–32. doi: 10.1111/fwb.12951
Altermatt, F., Seymour, M., and Martinez, N. (2013). River network properties shape α-diversity and community similarity patterns of aquatic insect communities across major drainage Basins. J. Biogeogr. 40, 2249–2260. doi: 10.1111/jbi.12178
Baas Becking, L. G. M. (1934). Geobiologie of Inleiding Tot De Milieukunde (in Dutch). The Hague: Van Stockum and Zoon.
Bagge, P. (1995). Emergence and upstream flight of lotic mayflies and caddisflies. Entomol. Fenn. 6, 91–97. doi: 10.33338/ef.83844
Benito, X., Fritz, S. C., Steinitz-Kannan, M., Vélez, M. I., and McGlue, M. M. (2018). Lake regionalization and diatom metacommunity structuring in tropical South America. Ecol. Evol. 8, 7865–7878. doi: 10.1002/ece3.4305
Bottin, M., Soininen, J., Ferrol, M., and Tison-Rosebery, J. (2014). Do spatial patterns of benthic diatom assemblages vary across regions and years? Freshw. Sci. 33, 402–416. doi: 10.1086/675726
Brittain, J. E., and Eikeland, T. J. (1988). Invertebrate drift - a review. Hydrobiologia 166, 77–93. doi: 10.1007/BF00017485
Brown, B. L., Sokol, E. R., Skelton, J., and Tornwall, B. (2017). Making sense of metacommunities: dispelling the mythology of a metacommunity typology. Oecologia 183, 643–652. doi: 10.1007/s00442-016-3792-1
Brown, B. L., and Swan, C. M. (2010). Dendritic network structure constrains metacommunity properties in riverine ecosystems. J. Anim. Ecol. 79, 571–580. doi: 10.1111/j.1365-2656.2010.01668.x
Cañedo-Argüelles, M., Boersma, K. S., Bogan, M. T., Olden, J. D., Phillipsen, I., Schriever, T. A., et al. (2015). Dispersal strength determines meta-community structure in a dendritic riverine network. J. Biogeogr. 42, 778–790. doi: 10.1111/jbi.12457
Chase, J. M., Jeliazkov, A., Ladouceur, E., and Duarte, V. S. (2020). Biodiversity conservation through the lens of metacommunity ecology. Ann. N. Y. Acad. Sci. 1469, 86–104. doi: 10.1111/nyas.14378
Chiu, M. C., Ao, S., He, F., Resh, V. H., and Cai, Q. (2020). Elevation shapes biodiversity patterns through metacommunity-structuring processes. Sci. Total. Environ. 743:140548. doi: 10.1016/j.scitotenv.2020.140548
Cid, N., Bonada, N., Heino, J., Cañedo-Argüelles, M., Crabot, J., Sarremejane, R., et al. (2020). A metacommunity approach to improve biological assessments in highly dynamic freshwater ecosystems. Bioscience 70, 427–438. doi: 10.1093/biosci/biaa033
Crabot, J., Clappe, S., Dray, S., and Datry, T. (2019). Testing the mantel statistic with a spatially-constrained permutation procedure. Methods Ecol. Evol. 10, 532–540. doi: 10.1111/2041-210X.13141
Dallas, T. (2018). Metacom: Analysis of the ‘Elements of Metacommunity Structure. Available online at: https://CRAN.R-project.org/package=metacom (accessed November 1, 2018).
de Mendoza, G., and Catalan, J. (2010). Lake macroinvertebrates and the altitudinal environmental gradient in the pyrenees. Hydrobiologia 648, 51–72. doi: 10.1007/s10750-010-0261-264
de Mendoza, G., Ventura, M., and Catalan, J. (2015). Environmental factors prevail over dispersal constraints in determining the distribution and assembly of Trichoptera Species in Mountain Lakes. Ecol. Evol. 5, 2518–2532. doi: 10.1002/ece3.1522
Dong, X., Li, B., He, F., Gu, Y., Sun, M., Zhang, H., et al. (2016). Flow directionality, mountain barriers and functional traits determine diatom metacommunity structuring of high mountain streams. Sci. Rep. 6:24711. doi: 10.1038/srep24711
Dormann, C., Elith, J., Bacher, S., Buchmann, C., Carl, G., Carré, G., et al. (2012). Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36, 27–46. doi: 10.1111/j.1600-0587.2012.07348.x
Finlay, B. (2002). Hypothesis: the rate and scale of dispersal of freshwater diatom species is a function of their global abundance. Protist 153, 261–273. doi: 10.1078/1434-4610-4103
Finn, D. S., and Adler, P. H. (2006). Population genetic structure of a rare high-elevation black fly, Metacnephia Coloradensis, occupying Colorado lake outlet streams. Freshw. Biol. 51, 2240–2251. doi: 10.1111/j.1365-2427.2006.01647.x
García-Girón, J., Heino, J., García-Criado, F., Fernández-Aláez, C., and Alahuhta, J. (2020). Biotic interactions hold the key to understanding metacommunity organisation. Ecography 43, 1–11. doi: 10.1111/ecog.05032
Goslee, S. C., and Urban, D. L. (2007). The ecodist package for dissimilarity-based analysis of ecological data. J. Statis. Softw. 22, 1–19.
Göthe, E., Angeler, D. G., and Sandin, L. (2013). Metacommunity structure in a small boreal stream network. J. Anim. Ecol. 82, 449–458. doi: 10.1111/1365-2656.12004
Grönroos, M., Heino, J., Siqueira, T., Landeiro, V. L., Kotanen, J., and Bini, L. M. (2013). Metacommunity structuring in stream networks: roles of dispersal mode, distance type, and regional environmental context. Ecol. Evol. 3, 4473–4487. doi: 10.1002/ece3.834
Guillot, G., and Rousset, F. (2013). Dismanteling the mantel tests. Methods Ecol. Evol. 4, 336–344. doi: 10.1111/2041-210x.12018
Gundersen, G., Johannensen, E., Andreassen, H. P., and Ims, R. A. (2001). Source–sink dynamics: how sinks affect demography of sources. Ecol. Lett. 4, 14–21. doi: 10.1046/j.1461-0248.2001.00182.x
Hayes, S. M., and Anderson, K. E. (2017). Beyond connectivity: how the structure of dispersal influences metacommunity dynamics. Theor. Ecol. 11, 151–159. doi: 10.1007/s12080-017-0355-y
He, S., Chen, K., Soininen, J., Heino, J., Ding, N., and Wang, B. (2020). Elements of metacommunity structure of diatoms and Macroinvertebrates within stream networks differing in environmental heterogeneity. J. Biogeogr. 47, 1–10. doi: 10.1111/jbi.13859
He, F., Wu, N., Dong, X., Tang, T., Domisch, S., Cai, Q., et al. (2020). Elevation, aspect, and local environment jointly determine diatom and Macroinvertebrate diversity in the Cangshan Mountain, Southwest China. Ecol. Indic. 108:105618. doi: 10.1016/j.ecolind.2019.105618
Heino, J. (2013). The importance of metacommunity ecology for environmental assessment research in the freshwater realm. Biol. Rev. 88, 166–178. doi: 10.1111/j.1469-185X.2012.00244.x
Heino, J., Nokela, T., Soininen, J., Tolkkinen, M., Virtanen, L., and Virtanen, R. (2015). Elements of metacommunity structure and community-environment relationships in stream organisms. Freshw. Biol. 60, 973–988. doi: 10.1111/fwb.12556
Heino, J., Soininen, J., Alahuhta, J., Lappalainen, J., and Virtanan, R. (2017). Metacommunity ecology meets biogeography: effects of geographical region, spatial dynamics and environmental filtering on community structure in aquatic organisms. Oecologica 183, 121–137. doi: 10.1007/s00442-016-3750-y
Jacobsen, D. (2008). Low oxygen pressure as a driving factor for the altitudinal decline in taxon richness of stream macroinvertebrates. Oecologia 154, 795–807. doi: 10.1007/s00442-007-0877-x
Kernan, M., Ventura, M., Bitušík, P., Brancelj, A., Clarke, G., Velle, G., et al. (2009). Regionalisation of remote european mountain lake ecosystems according to their biotia: environmental versus geographical patterns. Freshw. Biol. 54, 2470–2493. doi: 10.1111/j.1365-2427.2009.02284.x
Kim, Y. J., and Lee, O. M. (2017). A study of low-temperature and mountain epilithic diatom community in mountain stream at the Han River System, Korea. J. Ecol. Environ. 41, 1–10. doi: 10.1186/s41610-017-0048-x
Körner, C. (2007). The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 22, 569–574. doi: 10.1016/j.tree.2007.09.006
Krammer, K., and Lange-Bertalot, H. (1986). “Bacillariophyceae 1. Teil: naviculaceae,” in Süsswasser Von Mitteleuropa. Bacillariophyceae, eds G. Gärtner, H. Ettl, J. Gerloff, H. Heynig, and D. Mollenhauser (Stuttgart: Gustav Fischer Verlag).
Krammer, K., and Lange-Bertalot, H. (1988). “Bacillariophyceae 2. Teil: bacillariaceae, Epithemiaceae, Surirellaceae,” in Süsswasserflore Von Mitteleuropa. Bacillariophyceae, eds G. Gärtner, H. Ettl, J. Gerloff, H. Heynig, and D. Mollenhauser (Stuttgart: Gustav Fischer Verlag).
Krammer, K., and Lange-Bertalot, H. (1991a). “Bacillariophyceae 3. Teil: centrales, fragilariaceae, eunotiaceae,” in Süsswasserflora Von Mitteleuropa. Bacillariophyceae, eds G. Gärtner, H. Ettl, J. Gerloff, H. Heynig, and D. Mollenhauser (Stuttgart: Gustav Fischer Verlag).
Krammer, K., and Lange-Bertalot, H. (1991b). “Bacillariophyceae 4. Teil: achnanthaceae,” in Süsswasserflora Von Mitteleuropa. Bacillariophyceae, eds G. Gärtner, H. Ettl, J. Gerloff, H. Heynig, and D. Mollenhauser (Stuttgart: Gustav Fischer Verlag).
Kristiansen, J. (1996). 16. Dispersal of freshwater algae - a review. Hydrobiologia 336, 151–157. doi: 10.1007/BF00010829
Leibold, M., Holyoak, M., Mouquet, N., Amarasekare, P., Chase, J., Hoopes, M., et al. (2004). The metacommunity concept: a framework for multi-scale community ecology. Ecol. Lett. 7, 601–613. doi: 10.1111/j.1461-0248.2004.00608.x
Leibold, M. A., and Mikkelson, G. M. (2002). Coherence, species turnover, and boundary clumping: elements of meta-community structure. Oikos 97, 237–250. doi: 10.1034/j.1600-0706.2002.970210.x
Logue, J. B., Mouquet, N., Peter, H., and Hillebrand, H. (2011). Empirical approaches to metacommunities: a review and comparison with theory. Trends Ecol. Evol. 26, 482–491. doi: 10.1016/j.tree.2011.04.009
Maasri, A., Hayford, B., Erdenee, B., and Gelhaus, J. (2018). Macroscale drivers influencing the structural and functional organization of stream macroinvertebrate metacommunities: potential role of hydrological connectivity. Freshw. Sci. 37, 159–168. doi: 10.1086/695945
Martiny, J. B. H., Bohannan, B. J. M., Brown, J. H., Colwell, R. K., Fuhrman, J. A., Green, J. L., et al. (2006). Microbial biogeography: putting microorganisms on the map. Nat. Rev. Microbiol. 4, 101–112. doi: 10.1038/nrmicro1341
Moritz, C., Meynard, C., Devictor, V., Guizien, K., Labrune, C., Guarini, J., et al. (2013). Disentangling the role of connectivity, environmental filtering, and spatial structure on metacommunity dynamics. Oikos 122, 1401–1410. doi: 10.1111/j.1600-0706.2013.00377.x
Morse, J. C., Yang, L., and Tian, L. (1994). Aquatic Insects of China Useful for Monitoring Water Quality. Nanjing: Hohai University Press.
Nekola, J. C., and White, P. S. (1999). The distance decay of similarity in biogeography and ecology. J. Biogeogr. 26, 867–878. doi: 10.1046/j.1365-2699.1999.00305.x
Ohlmann, M., Maxel, F., Chalmandrier, L., Bec, S., Coissac, E., Gielly, L., et al. (2018). Mapping the imprint of biotic interactions on B-diversity. Ecol. Lett. 21, 1660–1669. doi: 10.1111/ele.13143
Oksanen, J., Guillaume Blanchet, F., Friendly, M., Kindt, R., Legendre, P., and McGlinn, D. (2019). Vegan: Community Ecology Package. : R Package Version 2.5.4. Available online at: https://CRAN.R-project.org/package=vegan (accessed May 15, 2019).
Potapova, M., and Charles, D. (2002). Benthic diatoms in USA rivers: distributions along spatial and environmental gradients. J. Biogeogr. 29, 167–187. doi: 10.1046/j.1365-2699.2002.00668.x
Presley, S., and Willig, M. (2010). Bat metacommunity structure on caribbean islands and the role of endemics. Glob. Ecol. Biogeogr. 19, 185–199. doi: 10.1111/j.1466-8238.2009.00505.x
Presley, S. J., Higgins, C. L., and Willig, M. R. (2010). A comprehensive framework for the evaluation of metacommunity structure. Oikos 199, 908–917. doi: 10.1111/j.1600-0706.2010.18544.x
QGIS Development Team (2015). QGIS Geographic Information System. Available online at: http://qgis.org (accessed November 1, 2018).
Qi, Y., Li, J., and Shi, Z. (1995). Flora Algarum Sinicarum Aquae Dulcis, Vol. Tomus IV, X, XII, XIV, XVI. Beijing: Science Press.
Qi, Y., Li, J., and Shi, Z. (2004a). Flora Algarum Sinicarum Aquae Dulcis, Vol. Tomus X. Beijing: Science Press.
Qi, Y., Li, J., and Shi, Z. (2004b). Flora Algarum Sinicarum Aquae Dulcis, Vol. Tomus XII. Beijing: Science Press.
Qi, Y., Li, J., and Shi, Z. (2009). Flora Algarum Sinicarum Aquae Dulcis, Vol. Tomus XIV. Beijing: Science Press.
Qi, Y., Li, J., and Shi, Z. (2013). Flora Algarum Sinicarum Aquae Dulcis, Vol. Tomus XVI. Beijing: Science Press.
R Core Team (2017). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Sanders, N., and Rahbek, C. (2012). The patterns and causes of elevational diversity gradients. Ecography 35, 1–3. doi: 10.1111/j.1600-0587.2011.07338.x
Schmera, D., Árva, D., Boda, P., Bódis, E., Bolgovics, Á, Borics, G., et al. (2017). Does isolation influence the relative role of environmental and dispersal-related processes in stream networks? an empirical test of the network position hypothesis using Multiple Taxa. Freshw. Biol. 63, 74–85. doi: 10.1111/fwb.12973
Shah, R. D. T., Sharma, S., Haase, P., Jähnig, S. C., and Pauls, S. U. (2015). The climate sensitive zone along an altitudinal gradient in central himalayan rivers: a useful concept to monitor climate change impacts in mountain regions. Clim. Chang. 132, 265–278. doi: 10.1007/s10584-015-1417-z
Soininen, J. (2004). Determinants of benthic diatom community structurein boreal streams: the role of environmental and spatial factors at different scales. Intern. Rev. Hydrobiol. 89, 139–150. doi: 10.1002/iroh.200310714
Soininen, J., and Weckström, J. (2009). Diatom community structure along environmental and spatial gradients in lakes and streams. Fundament. Appl. Limnol. 174, 205–213. doi: 10.1127/1863-9135/2009/0174-0205
Sundqvist, M. K., Sanders, N. J., and Wardle, D. A. (2013). Community and ecosystem responses to elevational gradients: processes, mechanisms and insights for global change. Annu. Rev. Ecol. Evol. Syst. 44, 261–280. doi: 10.1146/annurev-ecolsys-110512-135750
Thompson, P. L., Guzman, L. M., de Meester, L., Horvath, Z., Ptacnik, R., Vanschoenwinkel, B., et al. (2020). A process-based metacommunity framework linking local and regional scale community ecology. Ecol. Lett. 23, 1314–1329. doi: 10.1111/ele.13568
Thompson, R., and Townsend, C. (2006). A truce with neutral theory: local deterministic factors, species traits and dispersal limitation together determine patterns of diversity in stream invertebrates. J. Anim. Ecol. 75, 476–484. doi: 10.1111/j.1365-2656.2006.01068.x
Tonkin, J. D., Heino, J., and Altermatt, F. (2018). Metacommunities in river networks: the importance of network structure and connectivity on patterns and processes. Freshw. Biol. 63, 1–5. doi: 10.1111/fwb.13045
Tonkin, J. D., Sundermann, A., Jähnig, S. C., and Haase, P. (2015). Environmental controls on river assemblages at the regional scale: an application of the elements of Metacommunity structure framework. PLoS One 10:e0135450. doi: 10.1371/journal.pone.0135450
Tonkin, J. D., Tachamo Shah, R. D., Shah, D. N., Hoppeler, F., Jähnig, S. C., and Pauls, S. U. (2016). Metacommunity structuring in himalayan streams over large elevational gradients: the role of dispersal routes and niche characteristics. J. Biogeogr. 44, 62–74. doi: 10.1111/jbi.12895
Townsend, C. R., Dolédec, S., Norris, R., Peacock, K., and Arbuckle, C. (2003). The influence of scale and geography on relationships between stream community composition and landscape variables: description and prediction. Freshw. Biol. 48, 768–785. doi: 10.1046/j.1365-2427.2003.01043.x
Vavrek, M. J. (2011). Fossil: palaeoecological and palaeogeographical analysis tools. Palaeontol. Electron. 14:16.
Vilmi, A., Zhao, W., Picazo, F., Li, M., Heino, J., Soininen, J., et al. (2020). Ecological processes underlyfing community assembly of aquatic bacteria and Macroinvertebrates under contrasting climates on the Tibetan Plateau. Sci. Total Environ. 702:134974. doi: 10.1016/j.scitotenv.2019.134974
Wang, J., Soininen, J., Zhang, Y., Wang, B., Yang, X., and Shen, J. (2012). Patterns of elevational beta diversity in micro- and macroorganisms. Glob. Ecol. Biogeogr. 21, 743–750. doi: 10.1111/j.1466-8238.2011.00718.x
Wen, Z., Quan, Q., Du, Y., Xia, L., Ge, D., and Yang, Q. (2016). Dispersal, niche, and isolation processes jointly explain species turnover patterns of Nonvolant small mammals in a large mountainous region of China. Ecol. Evol. 6, 946–960. doi: 10.1002/ece3.1962
Willig, M., Kaufman, D., and Stevens, R. (2003). Latitudinal gradients of biodiversity: pattern, process, scale, and synthesis. Annu. Rev. Ecol. Evol. Syst. 34, 273–309. doi: 10.1146/annurev.ecolsys.34.012103.144032
Willig, M., and Presley, S. (2015). Biodiversity and metacommunity structure of animals along altitudinal gradients in tropical Montane forests. J. Trop. Ecol. 32, 421–436. doi: 10.1017/S0266467415000589
Keywords: dispersal, network, connectivity, Hengduan Mountains (Hengduanshan), elevation, distance-decay, biotic interactions
Citation: Kurthen AL, He F, Dong X, Maasri A, Wu N, Cai Q and Jähnig SC (2020) Metacommunity Structures of Macroinvertebrates and Diatoms in High Mountain Streams, Yunnan, China. Front. Ecol. Evol. 8:571887. doi: 10.3389/fevo.2020.571887
Received: 12 June 2020; Accepted: 14 September 2020;
Published: 21 October 2020.
Edited by:
Miguel Cañedo-Argüelles, University of Barcelona, SpainReviewed by:
Annika Vilmi, Finnish Environment Institute (SYKE), FinlandXavier Benito, National Socio-Environmental Synthesis Center (SESYNC), United States
Copyright © 2020 Kurthen, He, Dong, Maasri, Wu, Cai and Jähnig. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Angelika L. Kurthen, a3VydGhlbmFAb3JlZ29uc3RhdGUuZWR1; Qinghua Cai, cWhjYWlAaWhiLmFjLmNu