 Nina Gerber
Nina Gerber Yannick Auclair
Yannick Auclair Barbara König1
Barbara König1 Anna K. Lindholm
Anna K. Lindholm- 1Department of Evolutionary Biology and Environmental Studies, University of Zurich, Zurich, Switzerland
- 2Wildlife Sciences, Faculty of Forest Sciences and Forest Ecology, Univ of Goettingen, Goettingen, Germany
In mammals, reproduction is influenced by sexual competition, temperature and food availability and these factors might be crucial already during early life. Favorable early life environment and high maternal investment are expected to improve survival and reproduction. For example, in mammals, maternal investment via lactation predicts offspring growth. As body mass is often associated with fitness consequences, females have the potential to influence offspring fitness through their level of investment, which might interact with effects of population density and temperature. Here, we investigate the relationship between house mouse (Mus musculus domesticus) pup body mass at day 13 (used as approximation for weaning mass) and individual reproductive parameters, as well as longevity, under natural variation in population density and temperature (as approximation for season). Further, we assessed the extent to which mothers influence the body mass of their offspring until weaning. To do so, we analyzed life data of 384 house mice from a free-living wild commensal population that was not food limited. The mother’s contribution accounted for 49% of the variance in pup body mass. Further, we found a complex effect of population density, temperature and maternal investment on life-history traits related to fitness: shorter longevity with increasing pup body mass at day 13, delayed first reproduction of heavier pups when raised at warmer temperatures, and increased lifetime reproductive success for heavier pups at high densities. Our study shows that the effects of maternal investment are not independent of the effects of the environment. It thus highlights the importance of considering ecological conditions in combination with maternal effects to unravel the complexity of pup body mass on fitness measures.
Introduction
Organisms adopt different life-history strategies to cope with changing environments, trading off growth, survival and reproduction. The life-history response can be plastic in response to environmental condition. Early life is a critical period to investigate as conditions during early life can shape future life-history strategies (Lindström, 1999; Metcalfe and Monaghan, 2001; Maestripieri and Mateo, 2009). The experienced environment during early life is not only determined by abiotic conditions but can also be shaped by parental investment. Such non-genetic parental effects can be key components of reproductive strategies later in life (Badyaev and Uller, 2009; Bonduriansky and Crean, 2017).
In mammals, females typically provide most if not all of the parental care, including lactation which represents the most essential post-natal parental investment component determining food availability and thus the condition of offspring (Gittleman, 1985). Previous laboratory studies have shown that maternal investment through lactation can explain up to 65% of the variation observed in body mass at weaning in the house mouse Mus musculus (Cox et al., 1959; El Oksh et al., 1967; Atchley and Zhu, 1997). This maternal source of variance in offspring phenotype can be partitioned into prenatal maternal effects like resources allocated to an egg, and postnatal maternal effects such as maternal behavior. In species with a prolonged period of maternal dependence, the contribution of postnatal maternal effects on offspring body size can outweigh the effects of prenatal maternal effects (Reinhold, 2002; Steiger, 2013).
There are many examples of the benefits of being relatively heavier from small mammals to large herbivores. Heavier individuals usually have a higher probability of settlement in their population (Wauters et al., 1993), can achieve higher dominance ranks (e.g., Krackow, 1993; Klemme et al., 2006), reproduce earlier and/or produce more offspring (e.g., Fuchs, 1982; Anderson and Fedak, 1985; Dobson and Michener, 1995; Festa-Bianchet et al., 2000), or have increased survival (e.g., Murie and Boag, 1984; Wauters and Dhondt, 1989; Millar and Hickling, 1990). Moreover, body mass is considered a reliable proxy for the quality or health of an individual (Peters, 1986; Oli and Dobson, 2003). The influence of body mass on performance is not only observed during adulthood but can also be detected at earlier life stages (Dias and Marshall, 2010). The positive relationship between offspring and adult body mass of an individual (Birgersson and Ekvall, 1997; Festa-Bianchet et al., 2000) and the increased offspring quality at weaning observed whenever offspring received extended maternal care (Dahle and Swenson, 2003) suggest that higher maternal investment improves offspring fitness.
However, investment in bigger offspring might not always be favorable as under certain conditions smaller offspring size might be beneficial through alternative breeding strategies (Oakeshott, 1974; Gross, 1985; Feh, 1999) or because investing in larger offspring is costly for mothers (Dantzer et al., 2013). Thus, in fluctuating environments, mothers might adjust their investment to environmental conditions. Maternal effects where mothers adjust the phenotype of their offspring to the predicted offspring environment have been termed “anticipatory maternal effects” (Marshall and Uller, 2007). For such maternal effects to be adaptive, mothers need to be able to predict relevant aspects of the future environments of their offspring (Dantzer et al., 2013). Depending on environmental conditions, adjustment of milk quality or availability could improve offspring fitness. Mothers may invest differently in offspring depending on litter size, condition or offspring sex. In house mice, for example, investment into individual offspring (and their weaning weight) decreases with increasing litter size (König et al., 1988), including a reduction in milk lipid content (Ferrari et al., 2015).
Different maternal investment depending on offspring sex has been demonstrated in mammals as sexes can benefit differently from maternal care (Landete-Castillejos et al., 2005; Ceacero et al., 2018; Quesnel et al., 2018). Likewise, other factors can influence the effect of maternal investment. For example, the timing of birth (within a season) can influence growth, survival, and fitness (Clutton-Brock et al., 1982), and thus the condition at birth might influence the effect of maternal investment. In terms of lactation, laboratory experiments have shown that warmer temperatures reduce milk production and offspring growth (Barnett and Dickson, 1984; Król and Speakman, 2003). Further, population density is strongly related to competition over resources (Saitoh, 1981; Clutton-Brock, 1988; Gilbert and Krebs, 1991; Gaillard et al., 1997), and population density and/or climatic conditions, such as temperature during gestation and at birth of a litter, might influence physiological needs of the mother as well as the offspring, and affect future reproductive behavior. Modification of maternal care according to the social and seasonal conditions experienced when offspring are born might allow the adaptive adjustment of offspring mass to the conditions expected once sexually mature (Dantzer et al., 2013).
Measuring maternal investment can be challenging, especially under natural conditions. Since offspring body mass at weaning correlates with maternal investment (McDowell et al., 1930; Falconer, 1947; Don Bowen et al., 2001; Mateo, 2009; Ferrari et al., 2015), weaning represents the best time point to assess cumulative maternal energy allocation (pre- and postnatal). Body mass at or near weaning represents a good alternative to direct metabolic measurements of parental investment that are often too complicated or invasive to be used on wild populations (König et al., 1988; Sadowska et al., 2013).
Here, we aim to investigate the influence of social cues (population density) and the physical environment (temperature at birth) in combination with maternal investment on reproductive success using long-term data from an intensively monitored wild population of house mice (Mus musculus domesticus) in middle Europe that is not limited by food. This reflects a natural situation for commensal house mice in Europe and North America, as they normally breed in man-made structures with non-limited food sources (Bronson, 1979). House mice are non-hibernating and are not strictly seasonal breeders, although breeding is higher in summer than in winter. They produce several litters over an extended breeding period that may cover even up to 12 months per year (König and Lindholm, 2012), as they can breed at freezing temperatures as long as enough food is available (Bronson and Pryor, 1983). Since house mice mature within a few weeks after weaning (wild-derived males from the same study population reach sexual maturity at an age of 35 days, females at 54 days of age; Carlitz et al., 2019), environmental conditions experienced by the mother at birth of a litter might well allow to anticipate conditions experienced by offspring at the onset of reproduction. First, we test whether pup body mass at day 13 (which we used as an approximation for weaning weight) has an effect on the age at first reproduction, longevity and lifetime reproductive success (LRS), while accounting for population density and temperature at birth as well as sex differences. We predict increasing LRS and longevity with pup body mass. Further, we expect that at unfavorable conditions (low temperature/high population density) reproduction is reduced and that under these conditions, first reproduction is delayed. Second, we assessed the maternal influence on pup body mass while controlling for offspring sex as well as litter size and litter sex ratio. Last, we assessed the relationship between the estimated weaning mass and adult body mass. This study offers the opportunity to analyze the flexibility and the evolutionary consequences of maternal care dependent on environmental conditions, given a natural annual cycle in temperature and fluctuation in local population density.
Materials and Methods
Study Population
Data were collected from a free-living house mouse population in a 72 m2 agricultural building near Zurich, Switzerland (for more details see König and Lindholm, 2012). Although mice could easily exit the barn through numerous gaps, none of the large mammalian and avian predators that occur outside could enter. This reflects a normal situation for commensal house mice since they typically breed out of reach of nest predators (Latham and Mason, 2004) and large predators as cats, dogs or foxes are usually not efficient enough to control a population (Timm, 1994). The high permeability of the building toward mice did not allow us to directly monitor exits and entrances (but see Runge and Lindholm, 2018 for estimation of migration propensity). Water and food, a 50/50 mixture of oats and hamster food (Landi AG, Switzerland), were provided ad libitum to match conditions under which natural commensal house mouse populations are typically observed in Western Europe (Berry, 1970). The entire population inside the building was captured on average every 7 weeks to estimate adult population density and to examine animals.
Reproductive Activity
Reproduction occurred in 40 artificial nest boxes. We searched for new litters approximately every 10 days between January 2007 and December 2009. Each new litter was given an identification number. At this stage it was not possible to identify and record body mass of mothers, since females typically share nest boxes and identification of mothers is only possible through genetic analysis. Pups were sexed according to their anogenital distance and genital morphology (Hotchkiss and Vandenbergh, 2005) and aged according to a combination of skin pigmentation, ear development, fur development, teeth eruption, and eye development to assess the age of pups. In Supplementary Figure S1 we show the developmental stages at every day of age. Experimenters gained experience with sex and age estimates in the laboratory where births of litters were precisely documented. Because birth in house mice can last several hours, an uncertainty of about 1 day remains in our age estimation. Besides these regular 10 days checks, we also checked nest boxes whenever pups raised inside were forecast to be 13 days of age.
Body Mass Measurements
Pups were weighed to the nearest 0.1 g when they were 13 days old (±1 day). This age is the last day before weaning when they can be reliably captured as they are still blind, largely unreactive and entirely dependent on milk (actual weaning starts at day 17 and is completed at approximately 21–23 days old; König and Markl, 1987; König, 1993). Pup body mass is expected to increase linearly between day 13 and day 17 (Bronson, 1979; König and Markl, 1987). This increase, however, is likely to stop between day 17 and day 21 as females encourage their pups to eat solid food until they are independent (König and Markl, 1987). Using data collected in a laboratory study using descendants from our house mouse population (Ferrari et al., 2015), we found that body mass at day 13 was a strong predictor of body mass at the onset of weaning at day 17 (r207 = 0.85, p < 0.001; Supplementary Figure S2). We therefore consider mass at day 13 as a “weaning mass estimate.”
In captures of the entire population, each individual was weighed to the nearest 0.1 g, and those weighing at least 17.5 g were considered adults (Pelikán, 1981; Auclair et al., 2014). At that age, adults were individually marked with RFID tags (Trovan® ID-100A implantable micro transponder: 0.1 g weight, 11.5 mm length, 2.1 mm diameter; implanter Trovan® IID100E; Euro ID Identifikationssysteme GmbH and Co, Germany) for other research projects (e.g., Auclair et al., 2014; Harrison et al., 2018; Ferrari et al., 2019; Evans et al., 2020). Because the population is captured every 7 weeks and some resident animals may have been outside of the building, individuals differ in the age at which they were first captured as adults.
Genotyping and Parentage Analysis
An ear tissue sample was collected from every pup that was weighed at 13 days of age, every adult that was tagged, and from all corpses found inside of the barn. Following the same procedure as in Auclair et al. (2014), DNA was amplified using 25 microsatellite loci and a parentage analysis allowed assignment of the mother and the father of each individual to a 95% level of confidence using Cervus 3.0 (Marshall et al., 1998). Only fully assigned offspring and corpses that gave good quality DNA were kept in the analysis (n = 2,631; number of unassigned offspring = 629). Litter sizes were estimated by counting the number of pups with the same estimated day of birth (±1 day) that were assigned to the same mother. This was necessary as litters often share nest boxes, and are sometimes relocated to other nest boxes (Ferrari et al., 2019).
Life-History Traits
Individual reproductive success was assessed using genetic parentage analyses and separately defined as age at first reproduction, and the total number of offspring weaned (defined as surviving to 13 days of age) throughout life. Lifetime reproductive success (LRS, total number of offspring weaned) was monitored for 384 house mice (167 females and 217 males) born in the barn between January 2007 and December 2009 originating from 178 litters/120 mothers. Among these 384 individuals, 219 (100 females and 119 males) reproduced. They produced a total of 2,631 pups. The collection of corpses also allowed us to calculate longevity for 147 (56 females and 91 males) of these 384 individuals. We included LRS and life expectancy measures of these mice until February 2012, the last time a living focal mouse was recorded. The adult population density was estimated using an algorithm developed by Runge and Lindholm (2018) that allowed us to calculate the density of adults at any given date based on the number of sampled individuals in combination with their birth and death dates. The temperature was measured by a thermologger (HOBO U12-013) installed on an inside wall of the barn and mean monthly temperatures were used for the analyses.
Statistical Analyses
Statistical tests were carried out using R 3.5.1 (R Development Core Team, 2013). We estimated the influence of mother identity, population density, litter size, sex, and sex ratio on pup and adult body mass fitting two separate linear mixed-effects models (Bates et al., 2012) using the lme4 package. Mother identity was defined as a random factor while population density, sex ratio, litter size, sex and their interactions were defined as fixed factors. Final models were selected by dropping non-significant (p > 0.05) two-way interactions from the full model (model including fixed effects with all two-way interactions) as long as the model fit did not decrease. We proceeded with this approach until the AIC of the model no longer decreased (a ΔAIC of 2 was considered a model improvement) or no non-significant interactions were left. Continuous predictors were centered and model assumptions were checked visually using diagnostic plots and they were met for all models. We also did not detect any under- or overdispersion of the GLMM. The importance of mother identity was estimated as the proportion of the total variance explained by the random factor “mother identity” and tested by a likelihood ratio test comparing this model to a model having the same fixed factors structure but no random effect (Zuur et al., 2009). To investigate significant interaction effects with continuous predictors, we checked whether the 95% confidence intervals of the slopes predicted by the model overlapped (Aiken and West, 1991). For the illustration and slope analysis of continuous by continuous variable interactions, one of the predictors was categorized with the 0.5 quantile for sex ratio or the 0.25 and 0.75 quantile for population density, weaning weight and temperature at birth.
The influence of pup body mass at day 13 on longevity was analyzed with a linear model accounting for pup body mass, sex, population density, temperature at birth and their interactions. The number of offspring weaned was analyzed with a zero-inflated model with the same fixed effects structure (Zeileis et al., 2008). For the age at first reproduction, we performed a Cox proportional hazard regression (Kaplan and Meier, 1958), again with the same structure for fixed effects. This analysis considers that the timing of reproduction varies among groups and whether the event of reproduction happens or not.
Results
Life-History Consequences of Pup Body Mass at Day 13
The age at first reproduction was influenced by the temperature at birth (mean temperature during the month of birth) and its interaction with pup body mass at day 13 (Table 1 and Figure 1). When raised at cold temperatures, mice reproduced earlier in their life compared to offspring born at warmer temperatures. Temperature was, however, strongly associated with population density, with population density being generally higher in summer (April to September) when mean monthly temperatures were higher (Pearson correlation, r = 0.685, Figure 2). Despite the significant interaction between the temperature at birth and pup body mass, the effect of pup body mass alone was not significantly different from zero, neither at high nor low temperatures considering that the 95% confidence intervals are overlapping (Figure 1).
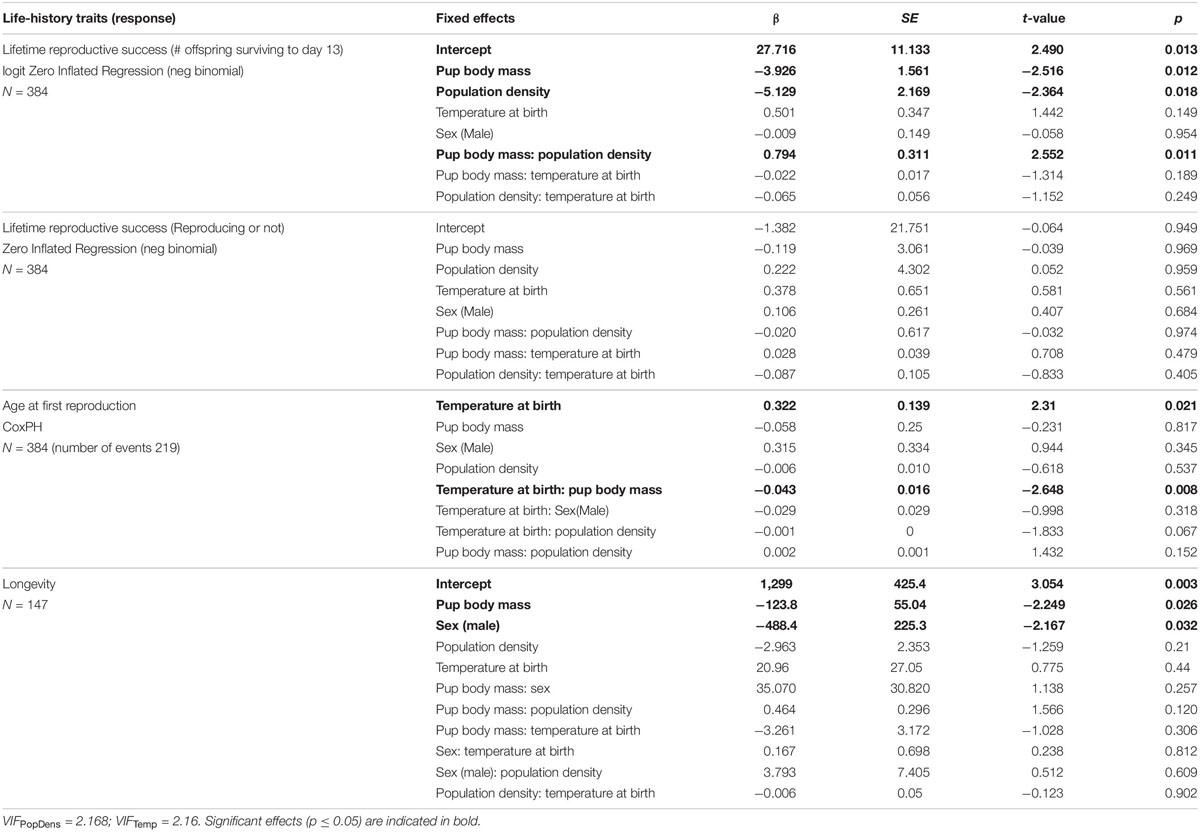
Table 1. Influence of pup body mass at day 13, sex, population density (number of individuals) and temperature at birth and their interactions on individuals’ life-history traits (measured by the age at first reproduction), lifetime reproductive success (analyzing total number of offspring surviving to day 13), the probability to reproduce at all, using the binomial and logit part of zero-inflated regression model and longevity.
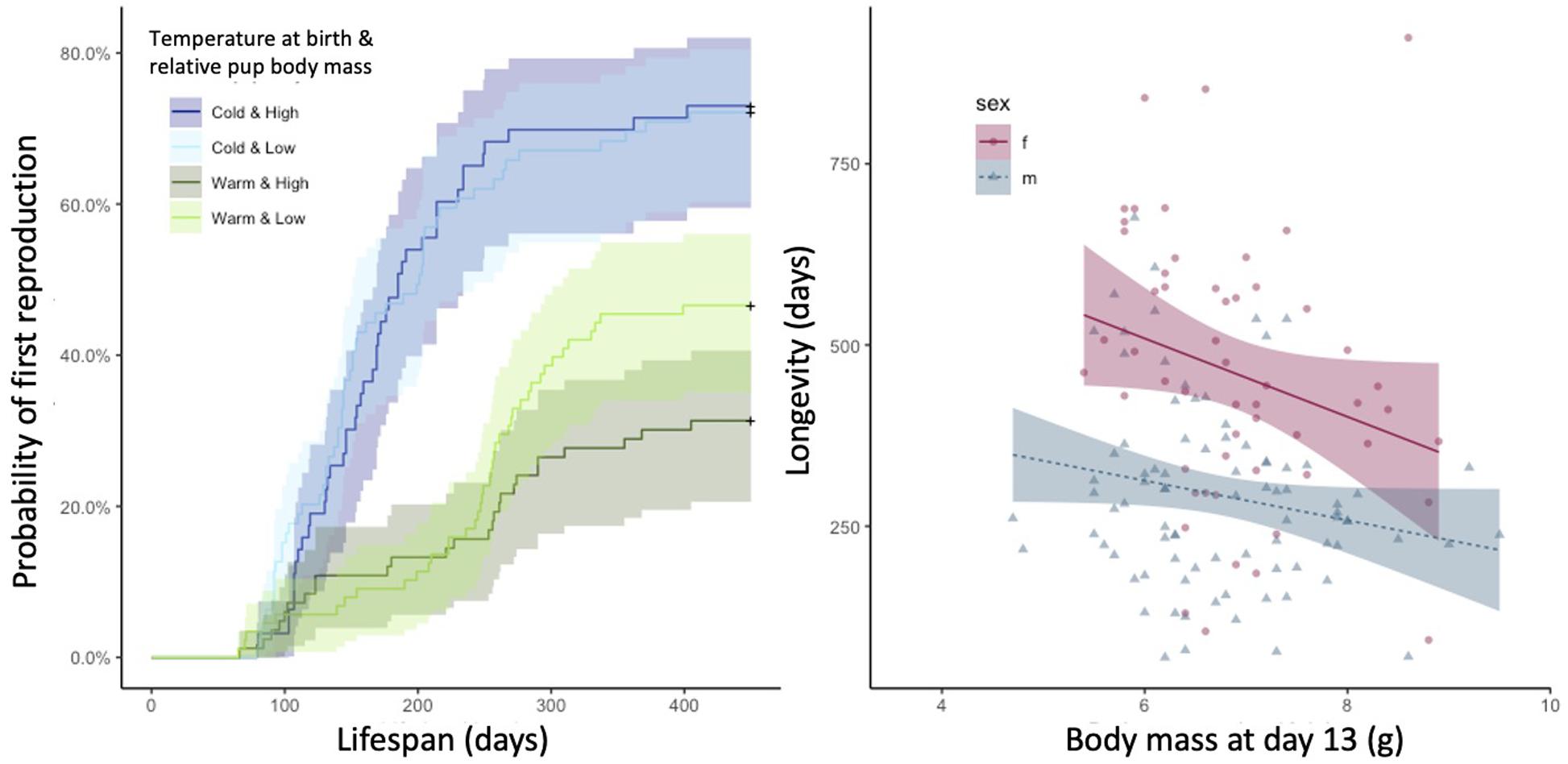
Figure 1. Probability of onset of reproduction (left) and longevity (right) with pup body mass at day 13 and relevant interaction. The shaded areas indicate the 95% confidence intervals. Temperature at birth and pup body mass at day 13 were categorized for illustration. Values above the median were considered as high, respectively warm, and values below the median as low, respectively cold.
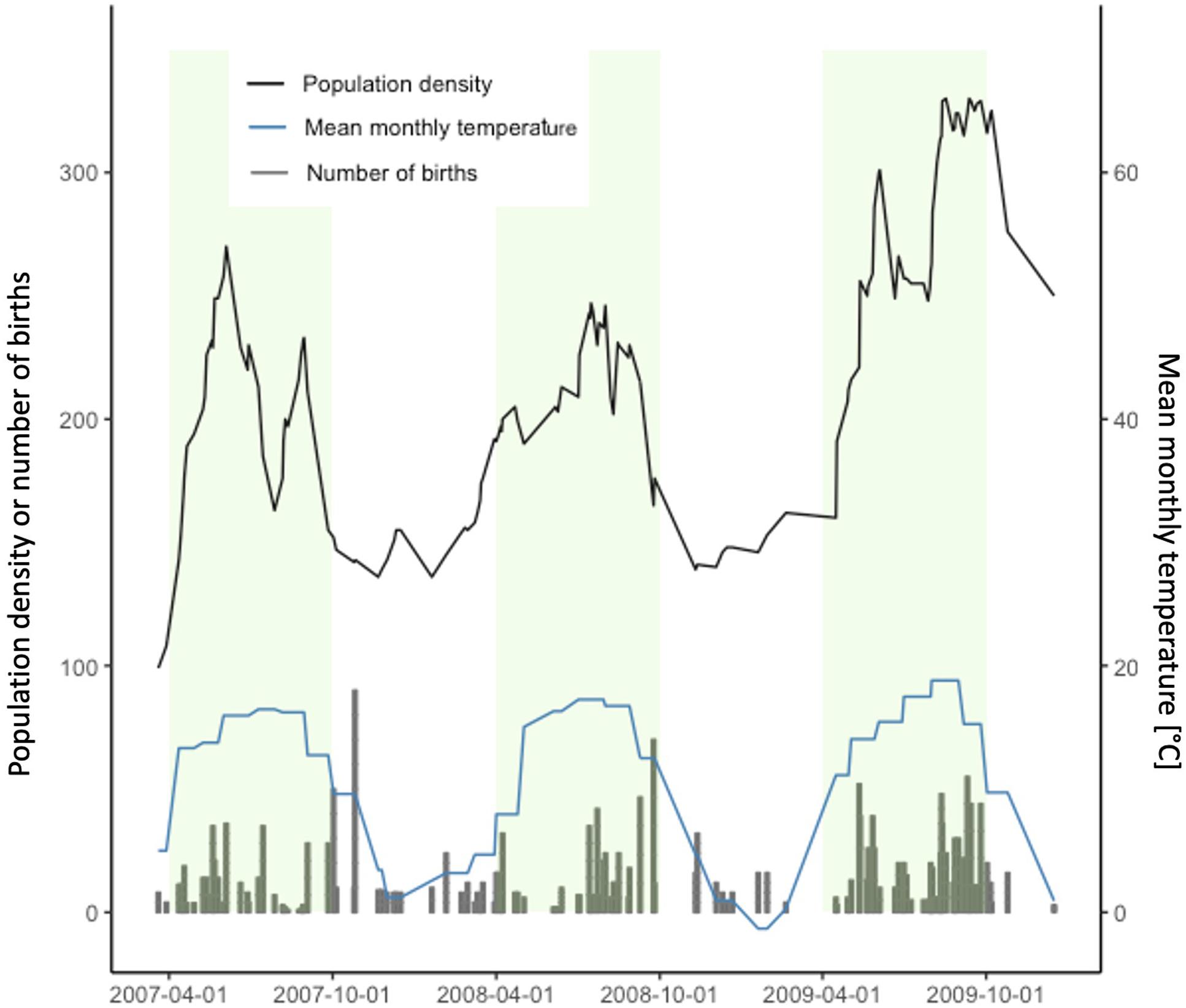
Figure 2. Population density (number of individuals), mean monthly temperature in the barn and number of births over time. Green shaded areas represent the summer months (April–September) and white areas refer to winter months (October–March).
Individual lifetime reproductive success (LRS, measured by the number of offspring surviving until day 13 over a lifetime) was influenced by the interaction of pup body mass at day 13 with population density at birth (Figure 3 and Table 1). There was a positive effect of pup body mass on LRS at high population densities, but the slopes did not differ significantly from zero at low densities (Figure 3). As 165/384 individuals did not breed at all, we also tested for factors predicting breeding. When looking at the binomial part of the zero-inflated GLM, it seems that the effect of pup body mass and density on LRS stem from differences in the number of offspring, rather than differences in the probability of reproducing (Table 1).
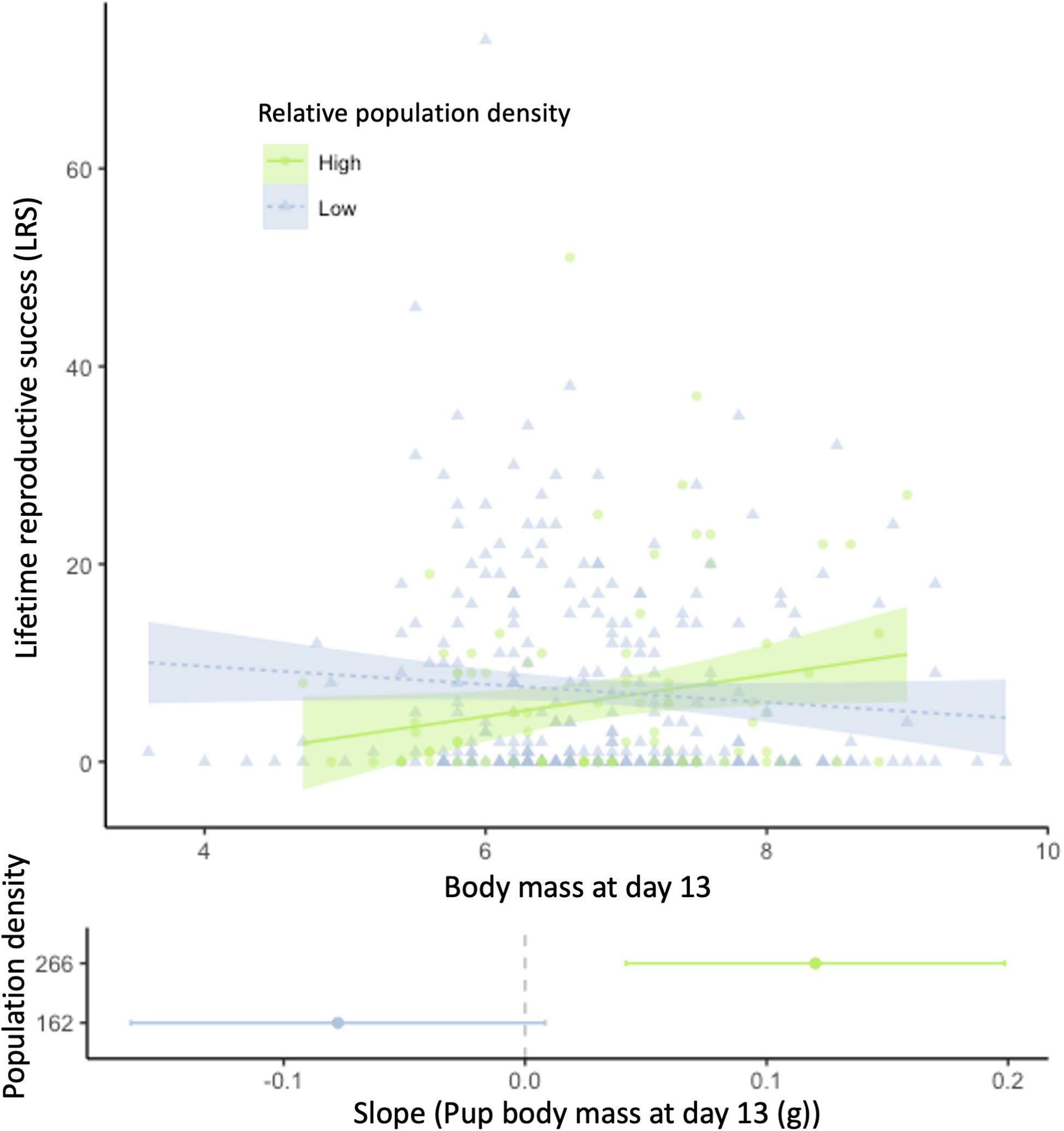
Figure 3. Relationship of lifetime reproductive success (LRS; number of offspring surviving until day 13) with body mass as pup at day 13. The shaded areas indicate the 95% confidence intervals. Population density was categorized for the figure and the slope analysis with high population density above the 0.75 quantile and low population density below the 0.25 quantile.
The mean life expectancy of mice was 357.5 days and was influenced by sex, with females living longer than males (Table 1 and Figure 1). Additionally, pup body mass had a significant negative influence on longevity (Table 1 and Figure 1). Neither population density nor the time in the year (analyzed as average temperature during the month of birth) nor any interactions had a significant effect on longevity (Table 1). There was no bias in the sex ratio of the 237 individuals for which we found no dead bodies (and thus we were not able to analyze their longevity) and assumed they died outside of the building (111 females vs. 126 males; χ2 = 0.95, df = 1, p = 0.330).
Influence of Mother Identity, Sex, Sex Ratio, Litter Size, and Population Density on Pup Body Mass
Differences between mothers accounted for 48% of the total variance observed in pup body mass at day 13 when corrected for litter size (Table 2). When not corrected for litter size, the mother’s contribution accounted for 49% of the variance in pup body mass (Supplementary Table S1). Pup body mass was significantly influenced by the interaction of sex with population density (Table 2). Female pup body mass was positively related to population density, whereas the slope for male body mass did not differ from zero, considering 95% confidence intervals (Figure 4). Pup body mass at day 13 was positively related to the temperature at birth (Figure 4 and Table 2). Pup body mass was also influenced by the interaction between litter size and sex ratio (Table 2), such that slopes of male- and female-biased sex ratios differed from each other, but the slopes did not differ from zero at male- or female-biased sex ratio (Figure 4), thus, this effect may not be biologically relevant.
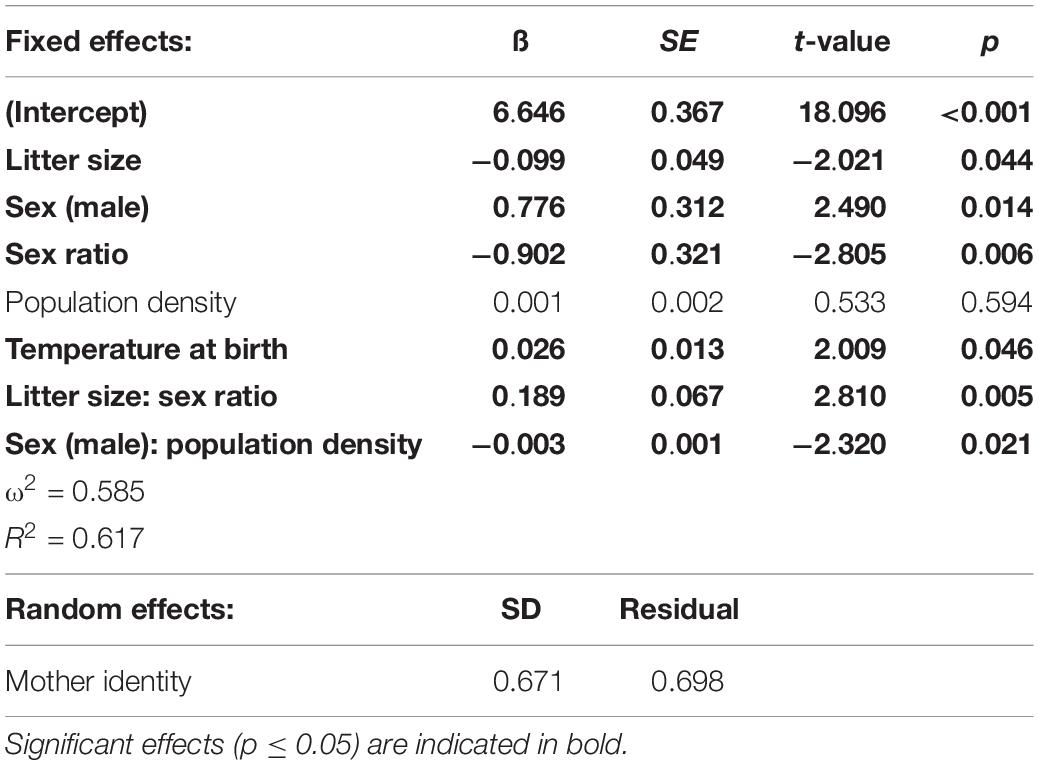
Table 2. Influence of mother identity, sex, sex ratio, and litter size on pup body mass at day 13 (N = 368); estimated by linear mixed model with “ML.”
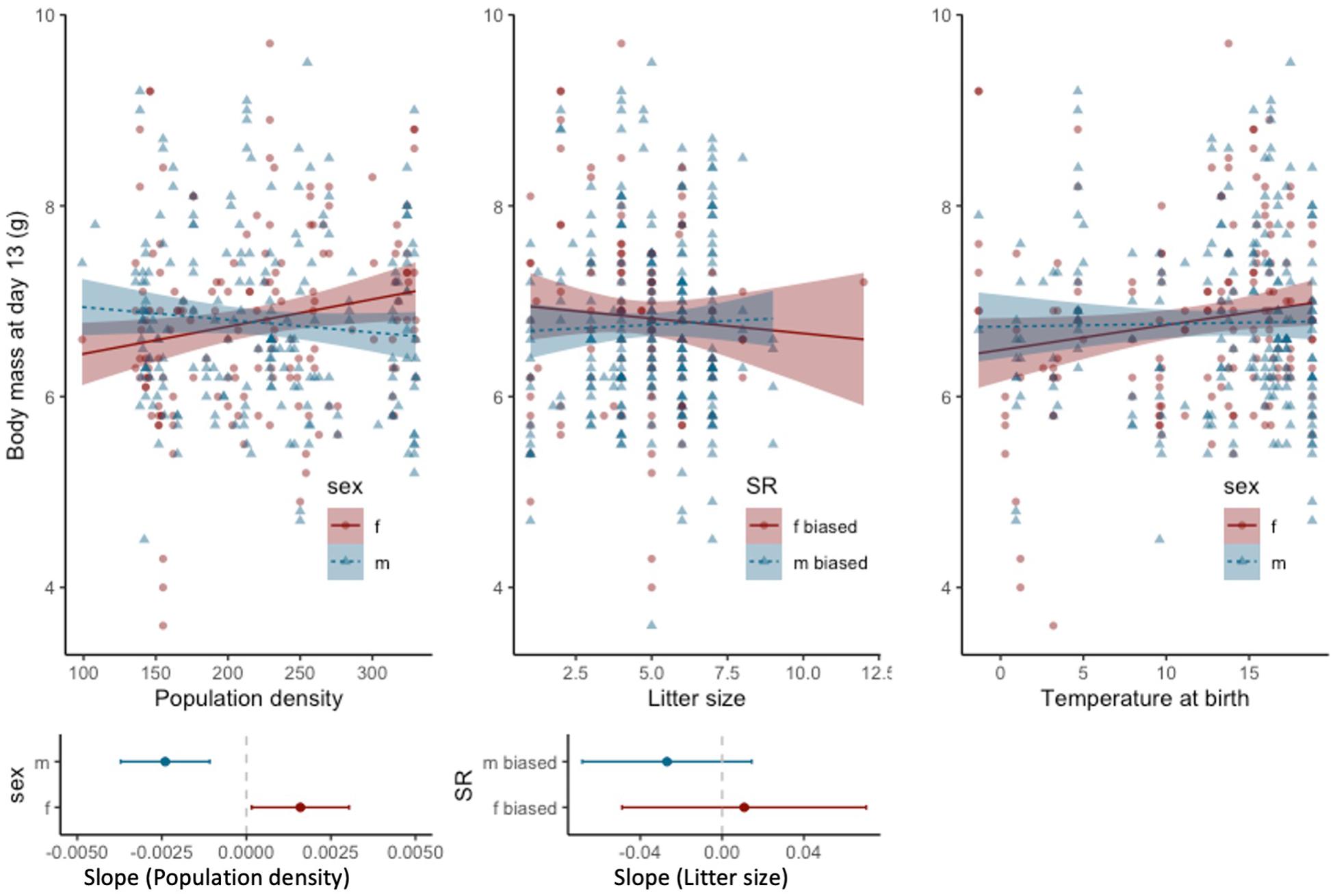
Figure 4. Relationship of population density (number of individuals, left), litter size at day 13 (middle) and temperature at birth (right) with pup body mass at day 13; the shaded area indicates the 95% confidence intervals. Sex ratio (SR) was categorized and was considered male-biased when greater than 0.5 and female-biased when below 0.5. Figures below show slope analysis with 95% confidence intervals.
Influences of Mother Identity, Pup Body Mass, and Sex on Adult Body Mass
As adult body mass increased with age, the measure of body mass had to be corrected for the age when adults were first captured and tagged (minimal body mass of 17.5 g), which varied (see Supplementary Figure S3). A negative exponential function was fitted to the data for both sexes, and the residuals were used as an index for adult body mass in further analyses.
Once adult, the influence of mother identity decreased to 12% of the total variance observed in the body mass when first captured as adults (Supplementary Table S2). The effect of pup body mass at day 13 on adult body mass depended on the population density at birth, as well as the sex of the offspring (Supplementary Figure S4 and Supplementary Table S2). Thus, at high population density, pup body mass in females was positively related to adult body mass, whereas there was no effect at low population density. In males, pup body mass was positively related to adult mass regardless of density (Supplementary Figure S4). There was a significant interaction of the litter size with pup body mass at day 13. However, the slopes at different weaning weights were not different from zero (Supplementary Figure S5). We have also found a significant interaction between the temperature at birth and the offspring sex, but again, simple slope analysis revealed that neither of the slopes differed from zero.
Discussion
This study aimed to examine the fitness consequences of variation in offspring body mass at day 13 (used as an approximation for weaning mass) under varying social and climatic conditions during the month of birth, with a particular focus on the extent to which lactating mothers might influence the body mass of their offspring. Here, we report delayed reproduction when pups were raised at warmer temperatures, shorter longevity with increasing pup body mass at day 13, and a positive effect of body mass on lifetime reproductive success for pups born at high population densities. Furthermore, we observed that mothers had a significant influence on offspring body mass accounting for nearly half of all residual variation.
Effect of Pup Body Mass, Population Density, and Temperature on Life-History Traits
Heavier pups developed into heavier adults, with the exception of females born into low density. In many taxa, larger individuals have higher reproductive success (Dias and Marshall, 2010). In mammals, bigger males typically have higher chances to acquire a breeding territory, become dominant and reproduce (Oakeshott, 1974; Clutton-Brock et al., 1979; Anderson and Fedak, 1985; Bouteiller-Reuter and Perrin, 2005). Similarly, female competition may occur to control territories housing the best nest sites or to control access to food (Reimer and Petras, 1967; Bujalska, 1973; Ostfeld, 1985; Wolff, 1993). As in males, reproductive success can be skewed toward dominant females (Clutton-Brock et al., 1984; Rusu and Krackow, 2004). Laboratory studies of mice have shown that a higher weaning weight in females results in larger litters and shorter inter-birth intervals during their first two reproductive events (Fuchs, 1982).
We therefore expected that a heavier weaning mass would allow an earlier onset of reproduction hence leading to a higher reproductive success if individuals manage to reproduce regularly (Roff, 2002). We found a more complicated result involving temperature during the month of birth, with individuals born at high temperatures delaying reproduction. Summer (when temperatures were high) was the main breeding season and also was when population density was highest (see Figure 2). Thus, delayed reproduction when raised during warmer months could indicate that pups encountered a few weeks later a situation with high reproductive competition, known to inhibit sexual maturity (Van Zegeren, 1980; Kruczek et al., 1989; Gilbert and Krebs, 1991; Manzano Nieves et al., 2019) or cause reproduction to cease (Yasukawa et al., 1985). When born at high population densities, high pup body mass at day 13 was associated with higher lifetime reproductive success (LRS), suggesting that high mass offspring have higher chances to breed under high reproductive competition. Poor competitors, of low mass, might more likely defer their reproduction and queue to acquire breeding or social positions at later age (Kokko and Johnstone, 1999; van de Pol et al., 2007). Low temperatures are associated with delayed puberty and reduced reproduction according to previous studies (Biggers et al., 1958; Barnett, 1965) and breeding in house mice is greatly reduced in winter (Bronson, 2009; König and Lindholm, 2012; Runge and Lindholm, 2018). For pups raised in our study population in winter, nevertheless, the delay to the next warmer period that also has low density (spring) is short and could explain why they bred early.
Heavier pups also died younger in our population, in both males and females. Large body size and its associated slower metabolic rate is linked to increased longevity across mammal species (Speakman, 2005). Metabolism is also influenced by temperature (Speakman, 2005), but we found no effect of temperature at birth on lifespan. Thus, basic energetic considerations do not easily account for these results. Moreover, Duarte and Speakman (2014) found no general effect of body size on longevity in a laboratory population of mice. The shorter lifespan of pups with a higher body mass might be due to strong intrasexual competition for breeding opportunities and/or nesting sites acting in both sexes (Miller et al., 2002). In female house mice such competition can prevent up to half of the females from reproducing (Ferrari et al., 2019). Heavy offspring may experience more frequent agonistic interactions that may compromise their survival compared to their smaller counterparts (Oakeshott, 1974). In females, the evolution of physiological suppression may help to decrease the rate of intrasexual aggression (Drickamer, 1977; Kruczek et al., 1989; Stockley et al., 2013; Clutton-Brock, 2016) and may contribute to their survival advantage over males (Clutton-Brock et al., 1979; Clutton-Brock and Isvaran, 2007; Clutton-Brock, 2009). Low mass offspring at high density may benefit from higher longevity, that allows them to survive until the next low-density period, coinciding with a warmer breeding period in spring. This is supported by the interaction between body mass and temperature on the probability of reproduction (Table 1) and the negative relationship between mass and longevity (Figure 1).
A potential bias in our study is that we only had data for the reproductive success and longevity of non-dispersing individuals, which could influence the interpretation of the results if body mass influences the propensity to disperse (see Massot et al., 2002; Bonte and De La Peña, 2009). In our study population, few individuals migrated among groups within the building, and pup body mass at day 13 did not predict emigration from the building (Runge and Lindholm, 2018). We, therefore, do not expect a bias related to a missing size class. Furthermore, house mice mainly disperse as subadults, before the onset of reproduction (Gerlach, 1996), and our study only includes mice that were recorded as adults. Thus, we assume that individuals in our data set were non-dispersers.
Early Life Population Density and Sex-Specific Effects of Maternal Care
In mammals, milk production is energetically expensive, increasing females’ food consumption and basal metabolic rate by up to 7.2 times (König et al., 1988; Hammond and Diamond, 1992). Reducing investment in milk, leading to smaller pups, could thus benefit mothers if their fitness benefits outweigh fitness losses due to lighter offspring. Females in poor condition would be expected to have higher fitness costs than good condition females from equivalent milk production. Unfortunately, we were unable to measure individual maternal condition at birth, or proxies for it such as dominance status. However, larger trends in condition related to temperature or the social environment affecting most females would be captured in our data. In house mice, high population densities affect local competition for resources (Bronson, 1979), and increased competition might lead to decreased maternal investment. We found that at high densities, male pups were on average smaller, whereas female pups were larger than at low densities. Increased maternal investment at high densities could be adaptive, as heavier pups born under these conditions had higher reproductive success, perhaps due to competitive advantages when reaching maturity as discussed before. This would be consistent with the idea that mothers alter their investment in offspring according to the social environment, which has been shown to influence offspring fitness (e.g., Siracusa et al., 2017). In our population, the social environment fluctuated within an individual’s life, and thus mothers may have enhanced the fitness of their offspring by adjusting offspring phenotypes to match the environment they will experience when breeding. Our study suggests, however, that only female pups born under high densities experienced this potential benefit.
Sex differences in pup mass under the same social environment and temperature could potentially be due to differences in either maternal care or in offspring feeding behavior. Sex-specific resource allocation in house mice is unlikely because the immobile arched posture of nursing females limits their control of offspring access to the nipples (König, 1989). However, mothers might not increase investment to single offspring, but they might increase their investment in female-biased litters at high population densities. Furthermore, male and female pups may differ in their behavior or metabolism (Garel et al., 2009) as seen in spotted hyenas Crocuta crocuta (Golla et al., 1999) and sea lions Zalophus californianus (Ono and Boness, 1996), leading to different growth potential under competition. An inter-sibling competition favoring the access to milk in one sex or a sex differential digesting efficiency has, however, never been reported in house mice.
Compensatory growth is likely to be important, as at low population density we did not find an effect of pup body mass on adult body mass in females, or of pup body mass on lifetime reproductive success. Low competition for resources might increase the potential for compensatory growth (Sundström et al., 2013). This could be influenced by density-dependent maternal effects, such as have been reported in avian and insect systems, where mothers adapt hormone allocation to the current social environment (Mazuc et al., 2003; Remeš et al., 2011; Crocker and Hunter, 2018). The evolution of plastic maternal investment might be favored in house mice, whereby increased offspring growth coincides with conditions under which it enhances fitness (Dantzer et al., 2013), namely, high population density. For maternal effects to be adaptive, mothers would need to be able to predict relevant aspects of the future environments of their offspring (Schwabl et al., 1997; Dantzer et al., 2013). When population growth is density-dependent, population density can act as an indicator of conditions and reproductive output of the offspring.
Conclusion
Our study might seem unusual in that food was not limiting. The ready access to food in the study population might have substantially lowered the costs of maternal investment. However, our study reflects natural conditions for house mice in Western Europe, as house mice usually live commensally with humans. Populations grow wherever food is easily accessible and available in good quantity, feral populations, that live independent of humans all year round, are restricted to islands (Berry, 1970; Latham and Mason, 2004; Pocock et al., 2004). Thus, the food availability in our study reflects normal conditions, and we generally expect commensal populations to experience similar resource conditions as our study population.
Our study is observational, and does not allow us to identify causal factors. Temperature and population density are correlated, thus an abiotic variable that has physiological effects varies together with a social variable, making interpretation difficult. We are unable to take into account the energy budgets of mothers, which will affect their ability to invest in offspring. However, our results do not corroborate clear predictions based on energetics of the effect of temperature on pup mass (Król and Speakman, 2003), suggesting that this is a highly complex system, in which females are able to buffer energetic constraints to some degree. The effects that population density and temperature had on life-history traits of pups might further allow to hypothesize on anticipatory maternal effects. Given that current density and temperature predict environmental conditions a few weeks later, such cues might allow a mother to adjust her investment according to the environment in which offspring will breed in order to maximize fitness. This is the first study that investigates maternal investment in house mice under natural conditions, and we see a need for an experimental approach to further understand biotic and abiotic effects.
Our results show a strong effect of population density, temperature at birth and maternal investment on life-history traits related to fitness. Similar to our results, other studies also find a more complex effect of pup body mass on fitness measures than often predicted (Ylönen et al., 2004; Pigeon and Pelletier, 2018). We show that the effects of maternal investment are not independent of the effects of the environment. Future experiments with controlled conditions along with the analysis of data from different years will help to unravel the complexity we find.
Data Availability Statement
The data that support the findings of this study are openly available on figshare at https://figshare.com/articles/dataset/Rdata_Weight_Fitness_LastAnalyses_new_txt/13507197 (Auclair et al., 2020b).
Ethics Statement
The animal study was reviewed and approved by the Veterinäramt der Gesundheitsdirektion Kanton Zürich.
Author Contributions
AKL and BK conceived the ideas and designed methodology. AKL, BK, and YA contributed to the data collection. YA did a preliminary analysis and wrote an initial version. NG revised and advanced the statistical analysis of this initial version. AKL, BK, and NG contributed to the interpretation of the new results and made major changes in the whole manuscript. All authors contributed to reviewing previous drafts and gave final approval for publication.
Funding
This study was funded by the Swiss National Science Foundation (3100A0-120444 and 31003A_176114), the Claraz Foundation, Promotor, Baugarten Stiftung, Julius Klaus Stiftung, and the University of Zurich.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Gabi Stichel who transpondered the mice, Jari Garbely who performed the genetic analyses, and Corinne Ackermann for the parentage analysis and everyone who contributed to the data collection. We also thank Pawel Koteja and Marco Festa-Bianchet, as well as two reviewers for helpful comments on the manuscript. The content of this manuscript has been published in Chapter 3 as part of the thesis of YA (Auclair, 2014). This manuscript has been released as a pre-print at bioRxiv (Auclair et al., 2020a).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2020.602359/full#supplementary-material
References
Aiken, L. S., and West, S. G. (1991). Multiple Regression: Testing and Interpreting Interactions. Thousand Oaks, CA: Sage.
Anderson, S. S., and Fedak, M. A. (1985). Grey seal males: energetic and behavioral links between size and sexual success. Anim. Behav. 33, 829–838. doi: 10.1016/s0003-3472(85)80017-8
Atchley, W. R., and Zhu, J. (1997). Developmental quantitative genetics, conditional epigenetic variability and growth in mice. Genetics 147, 765–776.
Auclair, Y. (2014). Short and Long-Term Fitness Consequences of Maternal Care in Wild House Mice Mus musculus domesticus. PhD thesis, University of Zurich, Zürich. doi: 10.5167/uzh-101084
Auclair, Y., Gerber, N., König, B., and Lindholm, A. (2020a). Population density and temperature influence the return on maternal investment in wild house mice. bioRxiv [Preprint]. doi: 10.1101/2020.06.30.177089
Auclair, Y., Gerber, N., König, B., and Lindholm, A. K. (2020b) LifehistoryDataHouseMice. figshare. Dataset. doi: 10.6084/m9.figshare.13507197.v3
Auclair, Y., König, B., Ferrari, M., Perony, N., and Lindholm, A. K. (2014). Nest attendance of lactating females in a wild house mouse population: benefits associated with communal nesting. Anim. Behav. 92, 143–149. doi: 10.1016/j.anbehav.2014.03.008
Badyaev, A. V., and Uller, T. (2009). Parental effects in ecology and evolution: mechanisms, processes and implications. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 364, 1169–1177. doi: 10.1098/rstb.2008.0302
Barnett, S. A. (1965). Adaptation of mice to cold. Biol. Rev. 40, 5–51. doi: 10.1111/j.1469-185x.1965.tb00794.x
Barnett, S. A., and Dickson, R. G. (1984). Milk production and consumption and growth of young of wild mice after ten generations in a cold environment. J. Physiol. 346, 409–417. doi: 10.1113/jphysiol.1984.sp015031
Biggers, J. D., Ashoub, M. R., McLaren, A., and Michie, D. (1958). The effect of the environment of phenotypic variability. J. Exp. Biol. 35, 144–155.
Birgersson, B., and Ekvall, K. (1997). Early growth in male and female fallow deer fawns. Behav. Ecol. 8, 495–499.
Bonduriansky, R., and Crean, A. J. (2017). What are parental condition transfer effects and how can they be detected? Methods Ecol. Evol. 9, 450–456. doi: 10.1111/2041-210X.12848
Bonte, D., and De La Peña, E. (2009). Evolution of body condition-dependent dispersal in metapopulations. J. Evol. Biol. 22, 1242–1251. doi: 10.1111/j.1420-9101.2009.01737.x
Bouteiller-Reuter, C., and Perrin, N. (2005). Sex-specific selective pressures on body mass in the greater white-toothed shrew. Crocidura russula. J. Evol. Biol. 18, 290–300. doi: 10.1111/j.1420-9101.2004.00836.x
Bronson, F. H. (1979). The reproductive ecology of the house mouse. Q. Rev. Biol. 54, 265–299. doi: 10.1086/411295
Bronson, F. H. (2009). Climate change and seasonal reproduction in mammals. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 364, 3331–3340. doi: 10.1098/rstb.2009.0140
Bronson, F. H., and Pryor, S. (1983). Ambient temperature and reproductive success in rodents living at different latitudes. Biol. Reprod. 29, 72–80. doi: 10.1095/biolreprod29.1.72
Bujalska, G. (1973). The role of spacing behavior among females in the regulation of reproduction in the bank vole. J. Reprod. Fertil. 19, 465–474.
Carlitz, E. H. D., Runge, J.-H., König, B., Winkler, L., Kirschbaum, C., Gao, W., et al. (2019). Steroid hormones in hair reveal sexual maturity and competition in wild house mice (Mus musculus domesticus). Sci. Rep. 9:16925. doi: 10.1038/s41598-019-53362-4
Ceacero, F., Komárková, M., García, A., and Gallego, L. (2018). Different maternal investment strategies for male and female calves in a polygynous mammal. Curr. Zool. 65, 269–277. doi: 10.1093/cz/zoy049
Clutton-Brock, T., Albon, S., and Guinness, F. (1982). Competition between female relatives in a matrilocal mammal. Nature 300, 178–180. doi: 10.1038/300178a0
Clutton-Brock, T. H. (1988). Reproductive Success: Studies of Individual Variation in Contrasting Breeding Systems. Chicago, IL: University of Chicago Press.
Clutton-Brock, T. H. (2009). Structure and function in mammalian societies. Philos. Trans. R. Soc. B Biol. Sci. 364, 3229–3242. doi: 10.1098/rstb.2009.0120
Clutton-Brock, T. H., Albon, S. D., Gibson, R. M., and Guinness, F. E. (1979). The logical stag: adaptive aspects of fighting in red deer (Cervus elaphus L.). Anim. Behav. 27, 211–225. doi: 10.1016/0003-3472(79)90141-6
Clutton-Brock, T. H., Albon, S. D., and Guinness, F. E. (1984). Maternal dominance, breeding success and birth sex ratios in red deer. Nature 308, 358–360. doi: 10.1038/308358a0
Clutton-Brock, T. H., and Isvaran, K. (2007). Sex differences in ageing in natural populations of vertebrates. Proc. R. Soc. B Biol. Sci. 274, 3097–3104. doi: 10.1098/rspb.2007.1138
Cox, D. F., Legates, J. E., and Clark Cockerham, C. (1959). Maternal influence on body weight. J. Anim. Sci. 18, 519–527. doi: 10.2527/jas1959.182519x
Crocker, K. C., and Hunter, M. D. (2018). Social density, but not sex ratio, drives ecdysteroid hormone provisioning to eggs by female house crickets (Acheta domesticus). Ecol. Evol. 8, 10257–10265. doi: 10.1002/ece3.4502
Dahle, B., and Swenson, J. E. (2003). Factors influencing length of maternal care in brown bears (Ursus arctos) and its effect on offspring. Behav. Ecol. Sociobiol. 54, 352–358. doi: 10.1007/s00265-003-0638-8
Dantzer, B., Newman, A. E. M., Boonstra, R., Palme, R., Boutin, S., Humphries, M. M., et al. (2013). Density triggers maternal hormones that increase adaptive offspring growth in a wild mammal. Science 340, 1215–1217. doi: 10.1126/science.1235765
Dias, G. M., and Marshall, D. J. (2010). Does the relationship between offspring size and performance change across the life-history? Oikos 119, 154–162. doi: 10.1111/j.1600-0706.2009.17725.x
Dobson, F. S., and Michener, G. R. (1995). Maternal traits and reproduction. in Richardson’s ground squirrels. Ecology 76, 851–862. doi: 10.2307/1939350
Don Bowen, W., Ellis, S. L., Iverson, S. J., and Boness, D. J. (2001). Maternal effects on offspring growth rate and weaning mass in harbour seals. Can. J. Zool. 79, 1088–1101. doi: 10.1139/z01-075
Drickamer, L. C. (1977). Delay of sexual maturation in female house mice by exposure to grouped females or urine from grouped females. J. Reprod. Fertil. 51, 77–81. doi: 10.1530/jrf.0.0510077
Duarte, L. C., and Speakman, J. R. (2014). Low resting metabolic rate is associated with greater lifespan because of a confounding effect of body fatness. AGE 36:9731. doi: 10.1007/s11357-014-9731-3
El Oksh, H. A., Sutherland, T. M., and Williams, J. S. (1967). Prenatal and postnatal maternal influence on growth in mice. Genetics 57, 79–94.
Evans, J. C., Liechti, J. I., Boatman, B., and König, B. (2020). A natural catastrophic turnover event: individual sociality matters despite community resilience in wild house mice. Proc. R. Soc. B Biol. Sci. 287:20192880. doi: 10.1098/rspb.2019.2880
Falconer, D. S. (1947). Milk production in mice. J. Agric. Sci. 37, 224–235. doi: 10.1017/s0021859600007474
Feh, C. (1999). Alliances and reproductive success in Camargue stallions. Anim. Behav. 57, 705–713. doi: 10.1006/anbe.1998.1009
Ferrari, M., Lindholm, A. K., and König, B. (2015). The risk of exploitation during communal nursing in house mice, Mus musculus domesticus. Anim. Behav. 110, 133–143. doi: 10.1016/j.anbehav.2015.09.018
Ferrari, M., Lindholm, A. K., and König, B. (2019). Fitness consequences of female alternative reproductive tactics in house mice (Mus musculus domesticus). Am. Nat. 193, 106–124. doi: 10.1086/700567
Festa-Bianchet, M., Jorgenson, J. T., and Réale, D. (2000). Early development, adult mass, and reproductive success in bighorn sheep. Behav. Ecol. 11, 633–639. doi: 10.1093/beheco/11.6.633
Fuchs, S. (1982). Optimality of parental investment: the influence of nursing on reproductive success of mother and female young house mouse. Behav. Ecol. Sociobiol. 10, 39–51. doi: 10.1007/bf00296394
Gaillard, J.-M., Boutin, J.-M., Delorme, D., Van Laere, G., Duncan, P., and Lebreton, J.-D. (1997). Early survival in roe deer: causes and consequences of cohort variation in two contrasted populations. Oecologia 112, 502–513. doi: 10.1007/s004420050338
Garel, M., Loison, A., Jullien, J.-M., Dubray, D., Maillard, D., and Gaillard, J.-M. (2009). Sex-specific growth in Alpine chamois. J. Mammal. 90, 954–960. doi: 10.1644/08-mamm-a-287.1
Gerlach, G. (1996). Emigration mechanisms in feral house mice: a laboratory investigation of the influence of social structure, population density, and aggression. Behav. Ecol. Sociobiol. 39, 159–170. doi: 10.1007/s002650050277
Gilbert, B. S., and Krebs, C. J. (1991). Population dynamics of Clethrionomys and Peromyscus in soutwestern Yukon 1973-1989. Holarctic Ecol. 14, 250–259. doi: 10.1111/j.1600-0587.1991.tb00659.x
Gittleman, J. L. (1985). “Functions of communal care in mammals,” in Evolution. Essays in Honour of John Maynard Smith, eds P. J. Greenwood, P. H. Harvey, and M. Slatkin (Cambridge: Cambridge University Press), 187–205.
Golla, W., Hofer, H., and East, M. L. (1999). Within-litter sibling aggression in spotted hyaenas: effect of maternal nursing, sex and age. Anim. Behav. 58, 715–726. doi: 10.1006/anbe.1999.1189
Gross, M. R. (1985). Disruptive selection for alternative life histories in salmon. Nature 313, 47–48. doi: 10.1038/313047a0
Hammond, K. A., and Diamond, J. (1992). An experimental test for a ceiling on sustained metabolic rate in lactating mice. Physiol. Zool. 65, 952–977. doi: 10.1086/physzool.65.5.30158552
Harrison, N., Lindholm, A. K., Dobay, A., Halloran, O., Manser, A., and König, B. (2018). Female nursing partner choice in a population of wild house mice (Mus musculus domesticus). Front. Zool. 15:4. doi: 10.1186/s12983-018-0251-3
Hotchkiss, A. K., and Vandenbergh, J. G. (2005). The anogenital distance index of mice (Mus domesticus): an analysis. J. Am. Assoc. Lab. Anim. Sci. 44, 46–48.
Kaplan, E. L., and Meier, P. (1958). Nonparametric-estimation from incomplete observations. J. Am. Stat. Assoc. 53, 457–481. doi: 10.1080/01621459.1958.10501452
Klemme, I., Eccard, J. A., Gerlach, G., Horne, T. J., and Ylönen, H. (2006). Does it pay to be a dominant male in a promiscuous species. ? Ann. Zool. Fennici 43, 248–257.
Kokko, H., and Johnstone, R. A. (1999). Social queuing in animal societies: a dynamic model of reproductive skew. Proc. R. Soc. B Biol. Sci. 266, 571–578. doi: 10.1098/rspb.1999.0674
König, B. (1989). Behavioural ecology of kin recognition in house mice. Ethol. Ecol. Evol. 1, 99–110. doi: 10.1080/08927014.1989.9525534
König, B. (1993). Maternal investment of communally nursing female house mice (Mus musculus domesticus). Behav. Process. 30, 61–74. doi: 10.1016/0376-6357(93)90012-g
König, B., and Lindholm, A. K. (2012). “The complex social environment of female house mice (Mus domesticus),” in Evolution of the House Mouse, eds M. Macholán, S. J. E. Baird, P. Munclinger, and J. Piálek (Cambridge: Cambridge University Press), 114–134. doi: 10.1017/cbo9781139044547.007
König, B., and Markl, H. (1987). Maternal care in house mice. Behav. Ecol. Sociobiol. 20, 1–9. doi: 10.1007/bf00292161
König, B., Riester, J., and Markl, H. (1988). Maternal care in house mice (Mus musculus): II. The energy cost of lactation as a function of litter size. J. Zool. 216, 195–210. doi: 10.1111/j.1469-7998.1988.tb02425.x
Krackow, S. (1993). The effect of weaning weight on offspring fitness in wild house mice (Mus musculus domesticus). Ethology 95, 76–82. doi: 10.1111/j.1439-0310.1993.tb00458.x
Król, E., and Speakman, J. R. (2003). Limits to sustained energy intake VI. Energetics of lactation in laboratory mice at thermoneutrality. J. Exp. Biol. 206, 4255–4266. doi: 10.1242/jeb.00674
Kruczek, M., Marchlewska-Koj, A., and Drickamer, L. C. (1989). Social inhibition of sexual maturation in female and male bank voles (Clethrionomys glareolus). Acta Theriol. 34, 479–485. doi: 10.4098/at.arch.89-47
Landete-Castillejos, T., García, A., and López-Serrano, F. R. (2005). Maternal quality and differences in milk production and composition for male and female Iberian red deer calves (Cervus elaphus hispanicus). Behav. Ecol. Sociobiol. 57, 267–274. doi: 10.1007/s00265-004-0848-8
Latham, N., and Mason, G. (2004). From house mouse to mouse house: the behavioural biology of free-living Mus musculus and its implications in the laboratory. Appl. Anim. Behav. Sci. 86, 261–289. doi: 10.1016/j.applanim.2004.02.006
Lindström, J. (1999). Early development and fitness in birds and mammals. Trends Ecol. Evol. 14, 343–348. doi: 10.1016/s0169-5347(99)01639-0
Maestripieri, D., and Mateo, J. M. (2009). “The role of maternal effects in mammalian evolution and adaptation,” in Maternal Effects in Mammals, eds D. Maestripieri and J. M. Mateo (Chicago, IL: University of Chicago Press).
Manzano Nieves, G., Schilit Nitenson, A., Lee, H. I., Gallo, M., Aguilar, Z., Johnsen, A., et al. (2019). Early life stress delays sexual maturation in female mice. Front. Mol. Neurosci. 12:27. doi: 10.3389/fnmol.2019.00027
Marshall, D. J., and Uller, T. (2007). When is a maternal effect adaptive? Oikos 116, 1957–1963. doi: 10.1111/j.2007.0030-1299.16203.x
Marshall, T. C., Slate, J., Kruuk, L. E. B., and Pemberton, J. M. (1998). Statistical confidence for likelihood-based paternity inference in natural populations. Mol. Ecol. 7, 639–655. doi: 10.1046/j.1365-294x.1998.00374.x
Massot, M., Clobert, J., Lorenzon, P., and Rossi, J.-M. (2002). Condition-dependent dispersal and ontogeny of the dispersal behaviour: an experimental approach. J. Anim. Ecol. 71, 253–261. doi: 10.1046/j.1365-2656.2002.00592.x
Mateo, J. M. (2009). “Maternal influence on development, social relationships, and survival behaviors,” in Maternal Effects in Mammals, eds D. Maestripieri and J. M. Mateo (Chicago, IL: University of Chicago Press).
Mazuc, J., Bonneaud, C., Chastel, O., and Sorci, G. (2003). Social environment affects female and egg testosterone levels in the house sparrow (Passer domesticus). Ecol. Lett. 6, 1084–1090. doi: 10.1046/j.1461-0248.2003.00535.x
McDowell, E. C., Gates, W. H., and McDowell, C. G. (1930). The influence of the quantity of nutrition upon the growth of the suckling mouse. J. Gen. Physiol. 13, 529–545. doi: 10.1085/jgp.13.5.529
Metcalfe, N. B., and Monaghan, P. (2001). Compensation for a bad start: grow now, pay later? Trends Ecol. Evol. 16, 254–260. doi: 10.1016/s0169-5347(01)02124-3
Millar, J., and Hickling, G. (1990). Fasting endurance and the evolution of mammalian body size. Funct. Ecol. 41, 5–12. doi: 10.2307/2389646
Miller, R. A., Harper, J. M., Galecki, A., and Burke, D. T. (2002). Big mice die young: early life body weight predicts longevity in genetically heterogeneous mice. Aging Cell 1, 22–29. doi: 10.1046/j.1474-9728.2002.00006.x
Murie, J. O., and Boag, D. A. (1984). The relationship of body weight to overwinter survival in columbian ground squirrels. J. Mammal. 65, 688–690. doi: 10.2307/1380854
Oakeshott, J. G. (1974). Social dominance, aggressiveness and mating success among male house mice (Mus musculus). Oecologia 15, 143–158. doi: 10.1007/bf00345742
Oli, M. K., and Dobson, F. S. (2003). The relative importance of life-history variables to population growth rate in mammals: cole’s prediction revisited. Am. Nat. 161, 422–440. doi: 10.1086/367591
Ono, K. A., and Boness, D. J. (1996). Sexual dimorphism in sea lion pups: differential maternal investment, or sex-specific differences in energy allocation? Behav. Ecol. Sociobiol. 38, 31–41. doi: 10.1007/s002650050214
Ostfeld, R. S. (1985). Limiting resources and territoriality in microtine rodents. Am. Nat. 126, 1–15. doi: 10.1086/284391
Pelikán, J. (1981). “Patterns of reproduction in the house mouse,” in Biology of the House Mouse, ed. R. J. Berry (London: Academic Press), 205–229.
Peters, R. H. (1986). The Ecological Implications of Body Size, Vol. 2. Cambridge: Cambridge University Press.
Pigeon, G., and Pelletier, F. (2018). Direct and indirect effects of early-life environment on lifetime fitness of bighorn ewes. Proc. R. Soc. B Biol. Sci. 285, 1935–2017.
Pocock, M. J. O., Searle, J. B., and White, P. C. L. (2004). Adaptations of animals to commensal habitats: population dynamics of house mice Mus musculus domesticus on farms. J. Anim. Ecol. 73, 878–888. doi: 10.1111/j.0021-8790.2004.00863.x
Quesnel, L., King, W. J., and Coulson, G. (2018). Tall young females get ahead: size-specific fecundity in wild kangaroos suggests a steep trade-off with growth. Oecologia 186, 59–71. doi: 10.1007/s00442-017-4003-4
R Development Core Team (2013). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Reimer, J. D., and Petras, M. L. (1967). Breeding structure of the house mouse. Mus musculus, in a population cage. J. Mammal. 48, 88–99. doi: 10.2307/1378173
Reinhold, K. (2002). Maternal effects and the evolution of behavioral and morphological characters: a literature review indicates the importance of extended maternal care. J. Heredity 93, 400–405. doi: 10.1093/jhered/93.6.400
Remeš, V., Matysioková, B., and Klejdus, B. (2011). Egg yolk antioxidant deposition as a function of parental ornamentation, age, and environment in great tits Parus major. J. Avian Biol. 42, 387–396. doi: 10.1111/j.1600-048x.2011.05402.x
Runge, J.-N., and Lindholm, A. K. (2018). Carrying a selfish genetic element predicts increased migration propensity in free-living wild house mice. Proc. R. Soc. B Biol. Sci. 285:20181333. doi: 10.1098/rspb.2018.1333
Rusu, A. S., and Krackow, S. (2004). Kin-preferential cooperation, dominance-dependent reproductive skew, and competition for mates in communally nesting female house mice. Behav. Ecol. Sociobiol. 56, 298–305.
Sadowska, J., Gebczynski, A. K., and Konarzewski, M. (2013). Basal metabolic rate is positively correlated with parental investment in laboratory mice. Proc. R. Soc. B Biol. Sci. 280:20122576. doi: 10.1098/rspb.2012.2576
Saitoh, T. (1981). Control of female maturation in high density populations of the red-backed vole, Clethrionomys rufocanus bedfordiae. J. Anim. Ecol. 50, 79–87. doi: 10.2307/4032
Schwabl, H., Mock, D. W., and Gieg, J. A. (1997). A hormonal mechanism for parental favouritism. Nature 386, 231–231. doi: 10.1038/386231a0
Siracusa, E., Boutin, S., Humphries, M. M., Gorrell, J. C., Coltman, D. W., Dantzer, B., et al. (2017). Familiarity with neighbours affects intrusion risk in territorial red squirrels. Anim. Behav. 133, 11–20. doi: 10.1016/j.anbehav.2017.08.024
Speakman, J. R. (2005). Body size, energy metabolism and lifespan. J. Exp. Biol. 208, 1717–1730. doi: 10.1242/jeb.01556
Steiger, S. (2013). Bigger mothers are better mothers: disentangling size-related prenatal and postnatal maternal effects. Proc. R. Soc. B Biol. Sci. 280:20131225. doi: 10.1098/rspb.2013.1225
Stockley, P., Bottell, L., and Hurst, J. L. (2013). Wake up and smell the conflict: odour signals in female competition. Philos. Trans. R. Soc. B Biol. Sci. 368:20130082. doi: 10.1098/rstb.2013.0082
Sundström, L. F., Kaspersson, R., Näslund, J., and Johnsson, J. I. (2013). Density-dependent compensatory growth in brown trout (Salmo trutta) in nature. PLoS One 8:e63287. doi: 10.1371/journal.pone.0063287
Timm, R. (1994). “House,” in The Handbook: Prevention and Control of Wildlife Damage. 4. Available onlinr at: http://digitalcommons.unl.edu/icwdmhandbook/4 (accessed September 20, 2019).
van de Pol, M., Pen, I., Heg, D., and Weissing, F. J. (2007). Variation in habitat choice and delayed reproduction: adaptive queuing strategies or individual quality differences? Am. Nat. 170, 530–541. doi: 10.1086/521237
Van Zegeren, K. (1980). Variation in aggressiveness and the regulation of numbers in house mouse populations. Neth. J. Zool. 30, 635–770. doi: 10.1163/002829679x00241
Wauters, L., Bijnens, L., and Dhondt, A. (1993). Body mass at weaning and juvenile recruitment in the red squirrel. J. Anim. Ecol. 62, 280–286. doi: 10.2307/5359
Wauters, L., and Dhondt, A. (1989). Body weight, longevity and reproductive success in red squirrels (Sciurus vulgaris). J. Anim. Ecol. 58, 637–651. doi: 10.2307/4853
Wolff, J. O. (1993). Why are female small mammals territorial? Oikos 68, 364–370. doi: 10.2307/3544853
Yasukawa, N. J., Monder, H., Leff, F. R., and Christian, J. J. (1985). Role of female behavior in controlling population growth in mice. Aggress. Behav. 11, 49–64. doi: 10.1002/1098-2337(1985)11:1<49::aid-ab2480110107>3.0.co;2-a
Ylönen, H., Horne, J. T., and Luukkonen, M. (2004). Effect of birth and weaning mass on growth, survival and reproduction in the bank vole. Evol. Ecol. Res. 6, 433–442.
Zeileis, A., Kleiber, C., and Jackman, S. (2008). Regression models for count data in R. J. Stat. Softw. 27, 1–25.
Keywords: house mice, life-history, lifetime reproductive success, maternal effects, offspring fitness, weaning weight
Citation: Gerber N, Auclair Y, König B and Lindholm AK (2021) Population Density and Temperature Influence the Return on Maternal Investment in Wild House Mice. Front. Ecol. Evol. 8:602359. doi: 10.3389/fevo.2020.602359
Received: 03 September 2020; Accepted: 15 December 2020;
Published: 02 February 2021.
Edited by:
Carlos Alonso Alvarez, National Museum of Natural Sciences (MNCN), SpainReviewed by:
Pablo Burraco, University of Glasgow, United KingdomStan Boutin, University of Alberta, Canada
Copyright © 2021 Gerber, Auclair, König and Lindholm. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nina Gerber, bmluYS5nZXJiZXJAdW5pLWdvZXR0aW5nZW4uZGU=; bmluYS5nZXJiZXIuZWNvbG9neUBnbWFpbC5jb20=
†These authors share first authorship