 David Giralt
David Giralt Javiera Pantoja
Javiera Pantoja Manuel B. Morales
Manuel B. Morales Juan Traba
Juan Traba Gerard Bota
Gerard Bota- 1Biodiversity and Animal Conservation Laboratory Landscape Dynamics and Biodiversity Program, Forest Science and Technology Centre of Catalonia (CTFC), Solsona, Spain
- 2Terrestrial Ecology Group (TEG-UAM), Department of Ecology, Universidad Autónoma de Madrid, Madrid, Spain
- 3Centro de Investigación en Biodiversidad y Cambio Global, Universidad Autónoma de Madrid (CIBC-UAM), Madrid, Spain
Landscape modification and habitat fragmentation are one of the major current threats to biodiversity. The main source of habitat fragmentation is the loss of focal habitat area, but changes in the composition of the surrounding landscape also have a direct effect on biodiversity. These changes may lead to the loss of some species but also may favor species replacement. Farmland birds in Europe are affected by landscape changes due to farmland intensification, such as the spread of irrigation, which may occur at different spatial scales. As irrigation is expected to increase in the coming years, which may affect protected areas, it is necessary to evaluate its potential consequences over focal biodiversity. In this study we assess the relationship between the increase of irrigated land at different spatial scales and changes in a dry cereal farmland bird community, bird abundance and species richness, using generalized linear models. We used a dry cereal farmland affinity index to describe the level of community specificity for dry cereal farmland. The increase in irrigated tree orchards produced an increase in species richness up to 500 m away from the irrigated area, which had a negative effect on the dry cereal farmland bird community, by triggering a replacement of specialist by generalist species. Our results show the importance of landscape-scale effects of irrigation occurring outside protected areas on the farmland bird community inside Natura 2000 sites, as well as how these effects are detected even at long distances from the disturbance source.
Introduction
Local species diversity and abundance often depend on spatial scales larger than the focal habitat patch (Fahrig, 2003; Villard and Metzger, 2014; De Camargo et al., 2018). Apart from quality and total surface of focal habitats at the patch (local) scale, surrounding landscape composition and configuration have effects on species diversity (Jules and Shahani, 2003), as it may modify animal movements and dispersal (Moilanen and Hanski, 1998), provide or limit alternative resources (Norton et al., 2000) and alter survival and reproduction rates (Prevedello and Vieira, 2010). In general, the more similar the matrix is to the focal habitat, the higher is its functional connectivity for those species occupying each individual patch (Prevedello and Vieira, 2010; Fahrig et al., 2011). Therefore, ecologists and conservationists need to be aware of the importance of linking landscape scale patterns and ecological processes when estimating basic structural parameters (e.g., abundance and species richness) of natural communities (Fahrig, 2003; Fischer and Lindenmayer, 2007).
Habitat loss is a main threat to biodiversity across the world (Pimm and Raven, 2000; IPBES, 2019). In Europe, farmland intensification is one of the main current drivers of habitat loss as it often results in landscape homogenization (Emmerson et al., 2016), and has been clearly linked to the widespread decline of different taxa, including farmland birds in recent decades (Donald et al., 2001; Benton et al., 2003; Geiger et al., 2010). Intensification through irrigation, for example, has been favored in many dry regions, and particularly in southern Europe, to increase crop yield or to allow new crop types in low productivity areas (Paracchini et al., 2007). In the Iberian Peninsula, extensive areas of dry cereal farmland (pseudo-steppes, sensu Suárez et al., 1997) have been replaced or fragmented by other crop systems such as irrigated fruit-tree orchards and herbaceous crops (e.g., maize and alfalfa). Thus, irrigation represents a drastic, quick, and large-scale change of dry cereal farmland landscapes, with predicted and demonstrated negative effects on biodiversity, and specifically on farmland and steppe birds (Brotons et al., 2004; González-Estébanez et al., 2011; Traba et al., 2013; Cardador et al., 2015; De Frutos et al., 2015). Because irrigated land is expected to continue increasing in the next years and given that landscape changes may affect protected areas or biodiversity hotspots, even if changes occur outside their borders, it becomes necessary to evaluate its potential consequences over high nature value areas (e.g., Natura 2000 sites) that have been designed to preserve dry cereal bird communities.
Birds are widely recognized as indicators of environmental changes acting at different scales (Fraixedas et al., 2020). However, not all the species respond equally to these changes because some of them (e.g., habitat specialists) are more susceptible than others to habitat degradation (Rooney et al., 2004; Julliard et al., 2006), contributing to a decrease of species diversity. On the contrary, certain changes in the landscape matrix could benefit widespread and habitat generalist species, increasing bird richness and diversity in focal habitats (Devictor et al., 2008). Specific indices for particular communities that may respond to particular ecological process and anthropogenic changes, such as measures of species specialization (Julliard et al., 2006; Devictor et al., 2008) may help to understand changes not occurring at specific but at the community level. Whether and how communities change in species composition because of the landscape context is thus crucial for predicting the consequences of habitat loss, planning conservation strategies, and managing protected areas.
In this study we evaluated the response of bird richness and abundance of a dry cereal farmland bird community within protected areas in Catalonia region (NE Spain) to the amount and type of irrigated land (herbaceous and arboreal irrigated crops) surrounding those protected sites. To evaluate in a more holistic way how the dry farmland bird community responded to nearby agrarian intensification, we calculated a dry cereal affinity index for the bird assemblage based on species-specific habitat selection indices that were derived from the Catalan Breeding Bird Atlas (Estrada et al., 2004). We expected that increases in the amount of irrigated land in the nearby landscape would negatively affect the dry cereal bird community decreasing the abundance of most representative species inside protected areas, independently of the effect on total species richness and abundance. Furthermore, a different response to irrigated herbaceous crops and tree orchards was also expected given the adaptation of dry farmland birds to open and treeless environments.
Materials and Methods
Study area
The study was carried out in the Lleida plain (1,000 km2), on the north-eastern edge of the Ebro Valley (Catalonia, Spain) (Figure 1). Climate is continental Mediterranean, with annual rainfall between 300 and 450 mm and an average annual temperature of 14.5°C (Cantero and Moncunill, 2012). The landscape is flat and mostly devoted to agriculture with a central area occupied by irrigated lands and an outer belt with remaining dry cereal farmland (i.e., pseudo-steppe habitat) (Figure 1). The irrigated area was first established at the beginning of the 20th century and later consolidated with new irrigation projects still currently ongoing. Irrigated land is characterized by herbaceous crops like alfalfa, maize, and winter cereal, or tree orchards like apple, pear, and peach. The remaining 600 km2 pseudo-stepped area is dominated by dry winter cereal crops, mainly wheat and barley, representing 70% of the area, and to a lesser extent, fallows and permanent woody crops like olive and almond trees, and vineyards. Interspersed in the agricultural matrix, there are still small patches of natural vegetation areas, mainly sparse scrubland. Due to its value for steppe birds (Traba et al., 2007), most of this pseudo-steppe area is included in Special Protection Area (SPA) sites (ES5130038, ES0000021, ES0000479, ES0000478, ES5130035, ES0000477, ES5130021, and ES0000321), and included in the Natura 2000 network. The aim of this network is to ensure the long-term survival of Europe’s most valuable and threatened species and habitats, listed under both the 2009/147/EC Birds Directive and the 92/43/EEC Habitats Directive.

Figure 1. Location of the study area in NE Spain and distribution of the 500 m transects (points) across the studied dry cereal farmland areas (lighter colored areas). Intermediate and dark gray areas represent irrigated herbaceous crops and orchards and white areas represent other land uses (urban, etc.). Polygons represent the Special Protected Areas included in Natura 2000 network.
Bird and Land-Use Data Collection
Bird data were collected on 178 linear transects (Järvinen and Väisänen, 1975) of ∼500 m of length, spaced more than 1 km apart and repeated yearly in 2010, 2011, and 2013 (mean number of transects/year was 155.7 ± 11.0). All transects were in dry cereal farmland areas and at 0.1 km to about 10 km from the irrigated edge. Three distance bands (<25, 25–50, and 50–100 m) were defined on both sides of the transect and each bird observed or heard was assigned to one of them. For the present work we considered only breeding species in the area with reference to the Breeding Bird Atlas of Catalonia (Estrada et al., 2004). All birds seen actively migrating or flying over the transect were excluded. To prevent potential detectability errors (Sanz-Pérez et al., 2020), we estimated species richness and abundance including only the individuals detected up to the 50 m band from both sides of the transect line, except for Little bustard (Tetrax tetrax) males, Eurasian stone-curlew (Burhinus oedicnemus), Red-legged partridge (Alectoris rufa), Pin-tailed sandgrouse (Pterocles alchata) and Cattle egret (Bubulcus ibis). These are large sized species that remain highly visible or audible up to 100 m and thus estimation of their abundance is not significantly affected by detection up to that distance (Sanz-Pérez et al., 2020). Bird surveys were conducted by hired observers with previous demonstrated experience in the identification of bird species and with similar skills, so we reasonably assumed no significant differences in detectability due to observer identity. Censuses were carried out by foot and performed once per season, in May from sunrise to 4 h later, and only with good weather conditions (with no rain nor wind). Census period matches the breeding season of the target species, as well as their peak daily activity.
For each transect and year, we calculated species richness and total bird abundance, as well as the abundance of three species highly specialized in dry cereal farmland and of conservation interest: Calandra lark (Melanocorypha calandra), Little bustard and Eurasian stone-curlew. Crested (Galerida cristata) and Thekla’s (Galerida theklae) larks, as well as Common (Sturnus vulgaris) and Spotless (Sturnus unicolor) starlings were considered as unique species (i.e., Galerida larks and Starlings), because they could not always be differentiated in the field. Because transect length was not always exactly 500 m (mean ± SD was 524 ± 46.5 m), all abundances and species richness were expressed as the number of individuals or species per hectare, respectively. Within-transect crops (100 m band) were mapped just after bird censuses, in order to calculate the proportion of area occupied by the following land-use categories: dry orchards (almond and olive trees), dry herbaceous crops (mostly cereal but also fallows), natural vegetation (mainly shrubland or open woodland) and other non-cropped areas (urban and watercourses).
Bird Community Dry Land Affinity Index
Although the presence of some bird species in certain ecosystems may be an indicator of each community’s degree of habitat specialization (Julliard et al., 2006), not all species in an assemblage usually have the same degree of affinity for that particular habitat. Thus, indices incorporating and combining that species-specific degree of habitat affinity may provide a more integrated way to evaluate the degree of habitat specialization of the community. Steppe birds are good indicators of irrigation-driven impacts (Brotons et al., 2004; De Frutos et al., 2015), since their presence and abundance are affected by irrigation. For this reason, we devised a Dryland Affinity Index (DAI hereafter), which measures bird community affinity for dry cereal habitats, and calculated its value per each transect and year (Equation 1). To do so, we used existing habitat affinity indices (K in Equation 1) for dry cereal areas estimated for each bird species in Estrada et al. (2004).
Where n is the number of species present in the transect; K is the index of dry cereal farmland habitat affinity for species j estimated by Estrada et al. (2004); and Abundance j is the abundance of species j per hectare in the transect. The K index was calculated in Estrada et al. (2004) for each species and habitat as the ratio between habitat occurrence in the 1 × 1 km Universal Transverse Mercator squares where that species was present and habitat occurrence in the whole set of squares in Catalonia (Spain). To make its interpretation easier, Estrada et al. (2004) subtracted one from this ratio. Therefore, K represents an index of affinity of any given species for a given habitat, with positive and negative values reflecting positive and negative affinity for that habitat, respectively. To calculate the DAI we only used the K index of species for dryland cereal, and ignored K values for the rest of habitats. Since the range values of K varied between species, we previously standardized this parameter in a range of values between −1 and +1 in two steps. We first subtracted the minimum value and divided it by the range, and then multiplied the resulting value by two and subtracted one. The higher the DAI value, the stronger the community’s affinity for dry cereal farmland is.
Irrigated Farmland Area in the Landscape
To assess the effect of irrigated land on our six response variables (DAI, total abundance, species richness, Little bustard abundance, Eurasian stone-curlew abundance, and Calandra lark abundance), we calculated and log-transformed the total area of irrigated herbaceous crops and orchards included in sequentially greater buffers around each transect, as two different variables (see Table 1). Using the centroid of each transect and based on the regional Geographic Information System of Farming Land (SIGPAC), total irrigated land of each type was calculated at three different buffers (radius of 500, 1,000, and 2,000 m). These buffers cover the home ranges of all species in our bird community, except for Marsh harrier (Circus aeruginosus) whose mean ranging distance is 3,070 ± 1,487 m during the nesting period in the study area (Cardador et al., 2009). Because in preliminary analyses we observed very low inter-annual variability across the 3-year study period in the area covered by irrigated orchards and herbaceous crops around transects, we extracted these variables from 1 year only (2011). All calculations were performed with the gvSIG 1.12.0 software.

Table 1. Habitat variables measured within and around each linear transect.
Statistical Analysis
We first checked collinearity between the six predictor variables (irrigated herbaceous crops and irrigated tree orchards at 500, 1,000, and 2,000 m) and discarded total area of irrigated herbaceous crops and tree orchards between 0–1,000 m from the transect (HC-1000 and TO-1000), due to high correlation (above 0.55) with the other variables (Dormann et al., 2013). To perform the analyses, the six response variables were averaged across years to cancel annual variation. In order to control for the local effect of transect habitat composition and to avoid correlation between proportion values, we performed a Principal Component Analysis (PCA) on the proportion of each habitat type within a 100 m wide buffer around each transect and included PCA factors as model covariates.
After checking normality assumptions prior to the analysis, we used general linear models to study the linear response of DAI index, species richness and total bird abundance to landscape composition in terms of area occupied by both irrigated herbaceous crops and irrigated orchards. To analyze the effects of irrigation on species-specific abundance (Calandra lark, Little bustard, and Eurasian stone-curlew) we used Compound Poisson Generalized Linear Models, which can handle extra zero inflation and be applied to continuous data (Zhang, 2013).
To assess the linear models, we used a multimodel inference approach based on Akaike’s Information Criterion (AIC). Model-averaged parameter estimates were calculated using the natural average method when more than one model was selected based on an ΔAIC ≤ 2 (Burnham and Anderson, 2002). Predictor variables were standardized in order to achieve comparability in the averaged models. Significance of explanatory variables was evaluated based on whether the confidence interval (CI) at 95% of its estimator excluded the value 0 (Burnham and Anderson, 2002).
Model fit was checked by exploring residuals normality and homoscedasticity. As Moran’s index estimation revealed significant spatial autocorrelation in the residuals, and in order to control for it, we included x and y coordinates of the transect centroids in all models. All analyses were performed using the software R (v. 3.5.3) and packages Stats (v. 3.6.2.), MuMin (v. 1.43.17), and cplm (v.0.7-9).
Results
A total of 6,662 individuals of 85 different bird species were detected across all transects and years. Mean DAI, total bird abundance and species richness across transects (n = 178) was −0.53 ± 0.93 (range −3.57–3.21), 2.55 ± 1.18 birds/ha (range 0.39–7.52 and 0.69 ± 0.36 species/ha (range 0.22–2.39), respectively. Both total bird abundance and species richness were lower in the 10 transects with highest DAI value (Table 2). Little bustard and Calandra lark were more abundant in transects with a high DAI value, but the opposite occurred for Eurasian stone-curlew (Table 2). Indeed, transects with high DAI values were mainly dominated by typical dry cereal farmland species such as Calandra lark and Little bustard, as well as other dry farmland species such as Corn bunting (Emberiza calandra) and Galerida larks (Supplementary Table 1). On the other hand, transects with low DAI values were dominated by more generalist species such as House sparrow (Passer domesticus), Common/Spotless starling and Feral pigeon (Columba livia var. domestica) (Supplementary Table 2).

Table 2. Mean and standard deviation (SD) of dryland affinity index (DAI), species richness (species/ha) and total and specific species abundance (birds/ha) of the 10 transects with highest and lowest DAI values.
The PCA describing land use variability within transects (local scale) yielded two factors explaining 74.84% of variance in the data (Table 3). The first factor (Habitat-F1) explained the 39.50% of variance and can be interpreted as a gradient from only-crop areas to more heterogeneous transects with natural vegetation patches (mainly open scrubland). The second factor (Habitat-F2) explained 35.34% of variance and it segregated dry cereal monocultures from mosaic transects where both dry herbaceous and woody crops (almond and olive trees) co-dominated (Table 3).

Table 3. Results of the principal component analysis (PCA) carried out to synthesize variation in habitat composition within transects.
The multi-model inference procedure for DAI selected six models with ΔAIC ≤ 2 (Supplementary Table 3). The area occupied by irrigated orchards (TO-500) within 500 m around transects had a negative effect on DAI (Table 4 and Figure 2).
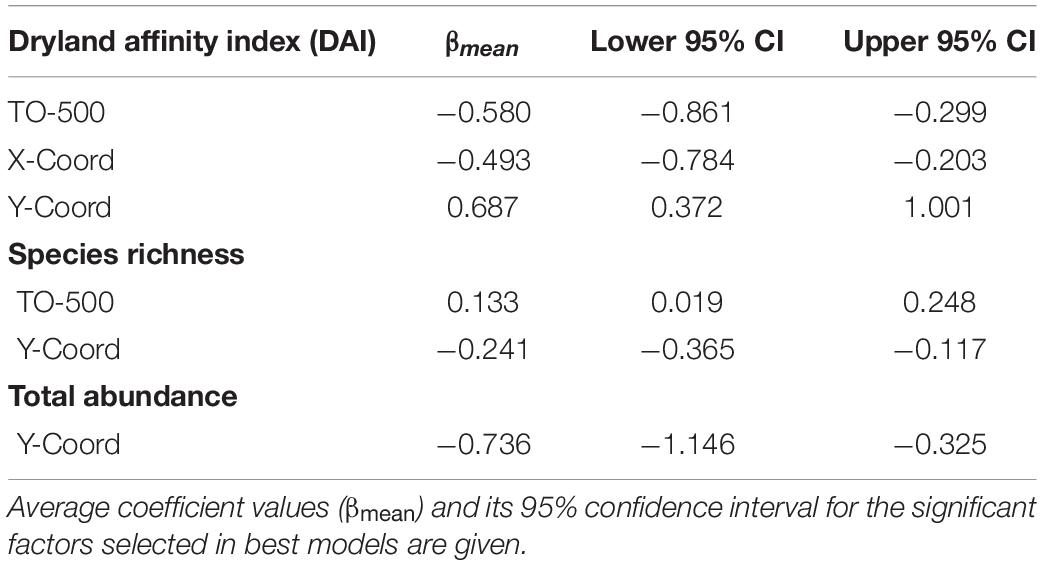
Table 4. Results of the multi-model inference analyses and model averaging for the effects of local habitat within transects (Habitat-F1 and F2) and landscape context (around transects) on dryland affinity index (DAI), species richness and total abundance of birds (n = 178 transects).
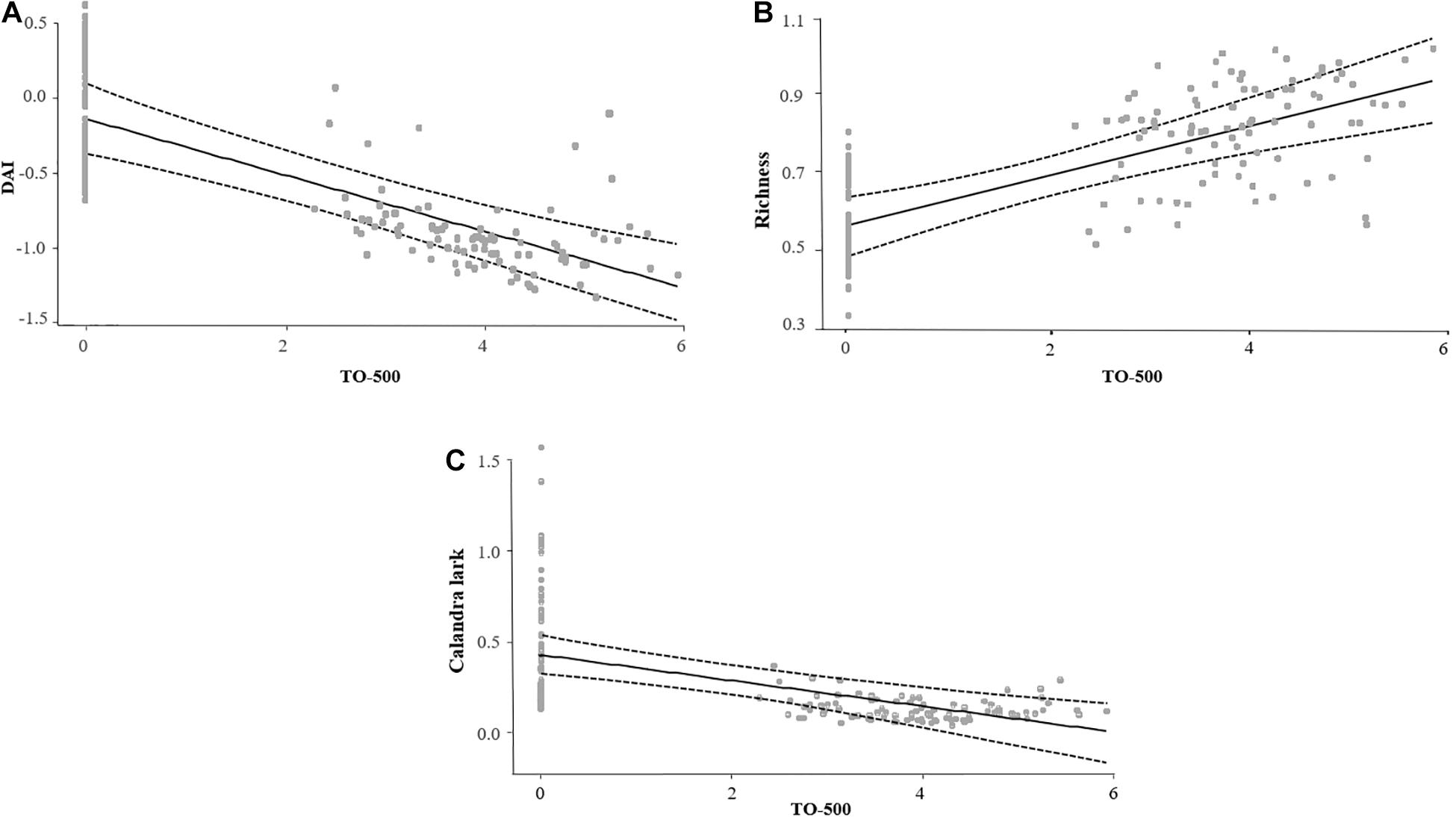
Figure 2. Relationship between model’s predicted values of (A) the dryland affinity index (DAI), (B) species richness, and (C) Calandra lark abundance and the log transformed area (ha) of irrigated orchards up to 500 m (TO-500) around transects. Mean (solid line) and CI (dashed line) of predicted values are shown.
For total bird abundance and species richness, 10 and 6 models were selected and averaged, respectively (Supplementary Table 3). Contrary to DAI, species richness presented a positive relationship with TO-500, while bird abundance did not respond to any landscape variable related to the amount of irrigated area. None of the local habitat variables had any effect on DAI, species richness or total bird abundance (Table 4).
Calandra lark abundance decreased with the amount of TO-500 and increased with the amount of natural vegetation within transects (Habitat-F1) (Table 5). Eurasian stone-curlew abundance positively responded to the area of dry cereal at local scale but no effect of irrigation was detected on this species, nor on Little bustard abundance (Supplementary Table 4).
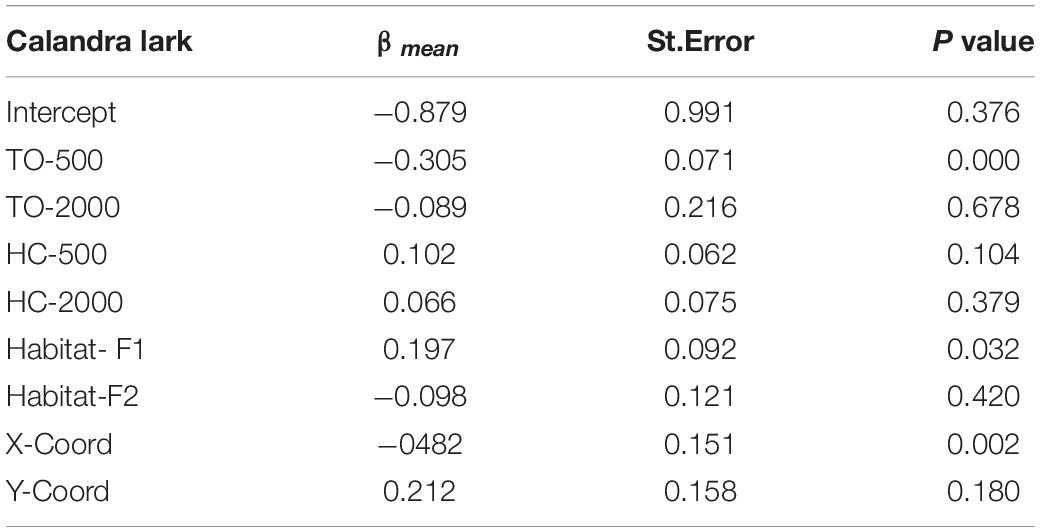
Table 5. Results of the Compound Poisson Generalized Linear Model for the effects of local habitat within transects (Habitat-F1 and F2) and landscape context (around transects) on Calandra lark abundance (n = 178).
Discussion
An extensive literature argues about the importance of landscape change and habitat fragmentation on biodiversity (Fisher and Lindermayer, 2007; Fahrig, 2013; Villard and Metzger, 2014; De Camargo et al., 2018). At the same time, the irrigation effects on biodiversity are still poorly known, though they are acknowledged as one of the main drivers of agrarian intensification and sources of bird-farmland decrease (e.g., Emmerson et al., 2016). Our results show the effects of changes in landscape composition for dry cereal farmland bird communities, and specifically how the amount of irrigated areas located at up to 500 m can alter species richness and community composition inside protected areas of the Natura 2000 network.
On one hand, irrigated orchards increased species richness present in dry cereal farmland areas up to 500 m away from the irrigated area. This seems to be due both to an increase of widespread or more generalist species (i.e., Common/Spotless starling and House sparrow) that positively selected both dry cereal farmland and irrigated areas (Estrada et al., 2004), and to the entry of new species such as the Great tit (Parus major) and the Mistle thrush (Turdus viscivorus), probably because the woody component of orchards increases landscape heterogeneity and thus the number and type of available resources (Benton et al., 2003; Fahrig et al., 2011) (see Supplementary Tables 1, 2).
As expected, not only species richness was altered by surrounding irrigation, but the whole dryland bird community composition was negatively affected when the area occupied by irrigated orchards increased at up to 500 m away. A decrease of DAI value means a decline in dryland specialization of the community (De Frutos et al., 2015), even if the total number of species and bird abundance increases. These results are also consistent with those of Devictor et al. (2008) who found that specialists are more vulnerable to habitat degradation, while less specialized species would win competitive interactions. Furthermore, in our case, a decrease of dryland specialists can be translated into a decrease of conservation values since these SPA sites are devoted to these group species. For example, Calandra lark, the most abundant species in dryland protected areas, decreased when irrigated orchard area increased at up to 500 m away, which suggests that this species can be a good indicator of landscape fragmentation (Morgado et al., 2010) and conservation status of dryland communities. On the contrary, we did not find any landscape effect of irrigated crops on Eurasian stone-curlew and Little bustard. While the former species is much more flexible in habitat selection, Little bustard is known to be negatively affected by irrigation (Brotons et al., 2004; Cardador et al., 2015). Variability in Little bustard abundance might be more sensitive to other key resources such as food availability or some habitat feature not captured by our predictor variables (Morales et al., 2008; Traba et al., 2008s).
Although the habitat loss and fragmentation processes themselves are the greatest threats associated with species conservation in landscape ecology (Fischer and Lindenmayer, 2007), several studies showed that the type of change in landscape composition is key to understanding positive or negative effects of habitat loss on biodiversity at focal sites (Vandermeer and Carvajal, 2001; Jules and Shahani, 2003; Fahrig, 2013). For example, less negative effects of habitat loss and fragmentation should be expected if low structural contrast exists between the original and the new habitat (Prevedello and Vieira, 2010). This is because species’ habitat associations are largely dictated by the availability of key resources in such habitats, rather than the habitat per se (Morales et al., 2008; Fahrig et al., 2011). Our study partially supports this idea since we find no effect of irrigated herbaceous crops on the dryland cereal bird community. Unlike irrigated herbaceous crops, irrigated orchards further involve a sharp change in vegetation structure and landscape visibility, which are key parameters for the survival of the dry cereal bird community (Cardador et al., 2014).
The change from a dry cereal farmland community to a more generalist one has important conservation implications, as changes within protected areas devoted to conservation of dryland specialists (e.g., steppe birds) may be triggered by factors occurring outside their limits, where no conservation measures are implemented. An increase of irrigated areas around the cereal steppes currently included in the Natura 2000 network can be expected in the future, considering the increase of irrigated area observed in recent years in Spain (MAPA, 2019). Our study shows that this negative effect would be relevant if irrigation allows the increase of irrigated orchards in the vicinity of the dryland protected areas, while no effects at the community level would be expected with a less drastic transition dominated by irrigated herbaceous crops. Therefore, from a conservation planning point of view, it would be advisable to consider a buffer zone around protected areas where irrigated orchards should be avoided in favor of irrigated herbaceous crops (i.e., low structural contrast between the original and the new habitat). The purpose of a buffer zone is to insulate protected areas from potentially damaging external influences, such as those caused by unsuitable land uses (Bennett and Mulongoy, 2006).
Our results offer an opportunity to optimize conservation in future irrigation schemes that are implemented close to dryland farmland protected areas while keeping the irrigated area constant, by avoiding some crops in contrast to others. However, more detailed studies (e.g., on vital rates such as breeding success and survival) should be conducted to understand the exact mechanisms (demographic, behavioral, etc.) by which the species of conservation concern respond to those landscape changes.
Data Availability Statement
The datasets generated for this study will be available on request to the corresponding author (DG) and with the permission of “Infraestructures de la Generalitat de Catalunya SAU”, who funded most part of data acquisition.
Author Contributions
DG, GB, MM, and JT conceived and designed the study. JP and DG implemented the analysis and wrote the manuscript. DG coordinated fieldwork. GB secured funding. All authors contributed to subsequent drafts and gave final approval for publication.
Funding
Departament d’Agricultura, Ramaderia, Pesca i Alimentació and Infraestructures de la Generalitat de Catalunya S.A.U. have funded a relevant part of the field work and data acquisition.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We acknowledge the field assistance of Joan Estrada, Sergi Sales, Joan Castelló, Arnau Bonan, Xavi Larruy, Albert Petit, Ferran González, Juan Bécares, Ferran Broto, Xavier Riera, Marc Anton, and David Guixé. We also acknowledge Núria Pou for the extensive support in data handling. This paper is a contribution to the REMEDINAL 3 (S2013/MAE-2719) network.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2021.611563/full#supplementary-material
References
Bennett, G., and Mulongoy, K. J. (2006). Review of Experience with Ecological Networks, Corridors and Buffer Zones. Technical Series No. 23. Montreal: Secretariat of the Convention on Biological Diversity.
Benton, T. G., Vickery, J. A., and Wilson, J. D. (2003). Farmland biodiversity: is habitat heterogeneity the key? Trends Ecol. Evol. 18, 182–188. doi: 10.1016/S0169-5347(03)00011-9
Brotons, L., Mañosa, S., and Estrada, J. (2004). Modelling the effects of irrigation schemes on the distribution of steppe birds in Mediterranean farmland. Biodivers. Conserv. 13, 1039–1058. doi: 10.1023/B:BIOC.0000014468.71368.35
Burnham, K. P., and Anderson, D. R. (2002). Model Selection and Multimodel Inference: a Practical Information-Theoretic Approach, 2nd Edn. New York, NY: Springer.
Cantero, C., and Moncunill, J. (2012). Sistemas Agrícolas de la Plana de Lleida: Descripción Y Evaluación de los Sistemas de Producción en el Área del Canal Segarra-Garrigues Antes de su Puesta en Funcionamiento. Lleida: Universitat de Lleida.
Cardador, L., De Cáceres, M., Bota, G., Giralt, D., Casas, F., Arroyo, B., et al. (2014). A resource-based modelling framework to assess habitat suitability for steppe birds in semiarid Mediterranean agricultural systems. PLoS One 9:e92790. doi: 10.1371/journal.pone.0104319
Cardador, L., De Cáceres, M., Giralt, D., Bota, G., Aquilué, N., Arroyo, B., et al. (2015). Tools for exploring habitat suitability for steppe birds under land use change scenarios. Agric. Ecosys. Environ. 200, 119–125. doi: 10.1016/j.agee.2014.11.013
Cardador, L., Mañosa, S., Varea, A., and Bertolero, A. (2009). Ranging behaviour of Marsh Harriers Circus aeruginosus in agricultural landscapes. IBIS 151, 766–770. doi: 10.1111/j.1474-919X.2009.00963.x
De Camargo, R. X., Boucher-Lalonde, V., and Currie, D. J. (2018). At the landscape level, birds respond strongly to habitat amount but weakly to fragmentation. Divers. Distrib. 24, 629–639. doi: 10.1111/ddi.12706
De Frutos, A., Olea, P. P., and Mateo-Tomás, P. (2015). Responses of medium-and large-sized bird diversity to irrigation in dry cereal agroecosystems across spatial scales. Agric. Ecosys. Environ. 207, 141–152. doi: 10.1016/j.agee.2015.04.009
Devictor, V., Julliard, R., and Jiguet, F. (2008). Distribution of specialist and generalist species along spatial gradients of habitat disturbance and fragmentation. Oikos 117, 507–514. doi: 10.1111/j.0030-1299.2008.16215.x
Donald, P. F., Green, R. E., and Heath, M. F. (2001). Agricultural intensification and the collapse of Europe’s farmland bird populations. Proc. Roy. Soc. Lond. B. 268, 25–29. doi: 10.1098/rspb.2000.1325
Dormann, C. F., Elith, J., Bacher, S., Buchmann, C., Carl, G., Carré, G., et al. (2013). Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36, 27–46. doi: 10.1111/j.1600-0587.2012.07348.x
Emmerson, M., Morales, M. B., Oñate, J. J., Batáry, P., Berendse, F., Liira, J., et al. (2016). “How agricultural intensification affects biodiversity and ecosystem services,” in Advances in Ecological Research, Vol. 55, eds A. J. Dumbrell, R. L. Kordas, and G. Woodward (Cambridge, MA: Academic Press).
Estrada, J., Pedrocchi, V., and Brotons, L. (2004). Atles Dels Ocells Nidificants de Catalunya 1999-2002. Barcelona: Institut Catalá d’Ornitologia (ICO).
Fahrig, L. (2003). Effects of habitat fragmentation on biodiversity. Ann. Rev. Ecol. Syst. 34, 487–515. doi: 10.1146/annurev.ecolsys.34.011802.132419
Fahrig, L. (2013). Rethinking patch size and isolation effects: the habitat amount hypothesis. J. Biogeogr. 40, 1649–1663. doi: 10.1111/jbi.12130
Fahrig, L., Baudry, J., Brotons, L., Burel, F. G., Crist, T. O., Fuller, R., et al. (2011). Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol. Lett. 14, 101–112. doi: 10.1111/j.1461-0248.2010.01559.x
Fischer, J., and Lindenmayer, D. B. (2007). Landscape modification and habitat fragmentation: a synthesis. Glob. Ecol. Biogeogr. 16, 265–280. doi: 10.1111/j.1466-8238.2007.00287.x
Fraixedas, S., Lindén, A., Piha, M., Cabeza, M., Gregory, R., and Lehikoinen, A. (2020). A state-of-the-art review on birds as indicators of biodiversity: advances, challenges, and future directions. Ecol. Indic. 118, 106728.
Geiger, F., Bengtsson, J., Berendse, F., Weisser, W. W., Emmerson, M., Morales, M. B., et al. (2010). Persistent negative effects of pesticides on biodiversity and biological control potential on European farmland. Basic Appl. Ecol. 11, 97–105. doi: 10.1016/j.baae.2009.12.001
González-Estébanez, F. J., García-Tejero, S., Mateo-Tomás, P., and Olea, P. P. (2011). Effects of irrigation and landscape heterogeneity on butterfly diversity in Mediterranean farmlands. Agric. Ecosys. Environ. 14, 262–270.
IPBES (2019). Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services, eds E. S. Brondizio, J. Settele, S. Díaz, and H. T. Ngo (Bonn: IPBES secretariat).
Järvinen, O., and Väisänen, R. A. (1975). Estimating relative densities of breeding birds by the line transect method. Oikos 26, 316–322. doi: 10.2307/3543502
Jules, E. S., and Shahani, P. (2003). A broader ecological context to habitat fragmentation: why matrix habitat is more important than we thought. J. Veg. Sci. 14, 459–464. doi: 10.1111/j.1654-1103.2003.tb02172.x
Julliard, R., Clavel, J., Devictor, V., Jiguet, F., and Couvet, D. (2006). Spatial segregation of specialists and generalists in bird communities. Ecol. Lett. 9, 1237–1244. doi: 10.1111/j.1461-0248.2006.00977.x
MAPA (2019). Encuestas Sobre Superficies y Rendimientos de Cultivos. Resultados Nacionales y Autonómicos (ESYRCE). Madrid: Ministerio de Agricultura, Pesca y Alimentación.
Moilanen, A., and Hanski, I. (1998). Metapopulation dynamics: effects of habitat quality and landscape structure. Ecology 79, 2503–2515. doi: 10.1890/0012-9658(1998)079[2503:MDEOHQ]2.0.CO;2
Morales, M. B., Traba, J., Carriles, E., Delgado, M. P., and García de la Morena, E. (2008). Sexual differences in microhabitat selection of breeding Little Bustards Tetrax tetrax: ecological segregation based on vegetation structure. Act. Oecol. 34, 345–353. doi: 10.1016/j.actao.2008.06.009
Morgado, R., Beja, P., Reino, L., Gordinho, L., Delgado, A., Borralho, R., et al. (2010). Calandra lark habitat selection: strong fragmentation effects in a grassland specialist. Act Oecol. 36, 63–73. doi: 10.1016/j.actao.2009.10.002
Norton, M. R., Hannon, S. J., and Schmiegelow, F. K. (2000). Fragments are not islands: patch vs landscape perspectives on songbird presence and abundance in a harvested boreal forest. Ecography 23, 209–223. doi: 10.1111/j.1600-0587.2000.tb00277.x
Paracchini, M. L., Terres, J. M., Peterson, J., and Hoogeveen, Y. (2007). High Nature Value Farmland and Traditional Agricultural Landscapes: Open Opportunities in the Development of Rural Areas. Zeist: Wageningen/KNNI Publishing.
Pimm, S. L., and Raven, P. (2000). Biodiversity: extinction by numbers. Nature 403, 843–845. doi: 10.1038/35002708
Prevedello, J. A., and Vieira, M. V. (2010). Does the type of matrix matter? A quantitative review of the evidence. Biodivers. Conserv. 19, 1205–1223. doi: 10.1007/s10531-009-9750-z
Rooney, T. P., Wiegmann, S. M., Rogers, D. A., and Waller, D. M. (2004). Biotic impoverishment and homogenization in unfragmented forest understory communities. Conserv. Biol. 18, 787–798. doi: 10.1111/j.1523-1739.2004.00515.x
Sanz-Pérez, A., Sollmann, R., Sardá-palomera, F., Bota, G., and Giralt, D. (2020). The role of detectability on bird population trend estimates in an open farmland landscape. Biodivers. Conserv. 29, 1747–1765. doi: 10.1007/s10531-020-01948-0
Suárez, F., Naveso, M. A., and De Juana, E. (1997). “Farming in the drylands of Spain: birds of the pseudosteppes,” in Farming and Birds in Europe, eds D. Pain and M. W. Pienkowski (London: Academic Press), 297–330.
Traba, J., García de la Morena, E. L., Morales, M. B., and Suárez, F. (2007). Determining high value areas for steppe birds in Spain. Hot spots, complementarity and the efficiency of protected areas. Biodivers. Conserv. 16, 3255–3275. doi: 10.1007/s10531-006-9138-2
Traba, J., Morales, M. B., García de la Morena, E. L., Delgado, M. P., and Krístin, A. (2008). Selection of breeding territory by little bustard (Tetrax tetrax) males in Central Spain: the role of arthropod availability. Ecol. Res. 23, 615–622. doi: 10.1007/s11284-007-0418-4
Traba, J., Sastre, P., and Morales, M. B. (2013). “Factors determining species richness and community composition in steppe birds of peninsular Spain,” in Steppe Ecosystems: Biological Diversity, Management and Restoration, eds M. B. Morales and J. Traba (New York, NY: NOVA Science Publishers), 29–46.
Vandermeer, J., and Carvajal, R. (2001). Metapopulation dynamics and the quality of the matrix. Am. Nat. 158, 211–220. doi: 10.1016/j.biocon.2004.10.002
Villard, M. A., and Metzger, J. P. (2014). Beyond the fragmentation debate: a conceptual model to predict when habitat configuration really matters. J. Appl. Ecol. 51, 309–318. doi: 10.1111/1365-2664.12190
Keywords: habitat loss and fragmentation, irrigation, farmland intensification, protected areas, indicator, bird community
Citation: Giralt D, Pantoja J, Morales MB, Traba J and Bota G (2021) Landscape-Scale Effects of Irrigation on a Dry Cereal Farmland Bird Community. Front. Ecol. Evol. 9:611563. doi: 10.3389/fevo.2021.611563
Received: 29 September 2020; Accepted: 18 February 2021;
Published: 18 March 2021.
Edited by:
Jasmine Lee, Monash University, AustraliaReviewed by:
Ángel De Frutos, University of Jaume I, SpainNaoki Katayama, National Agriculture and Food Research Organization (NARO), Japan
Copyright © 2021 Giralt, Pantoja, Morales, Traba and Bota. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: David Giralt, ZGF2aWQuZ2lyYWx0QGN0ZmMuY2F0