 Francisco Valente-Neto1*
Francisco Valente-Neto1* Fabio de Oliveira Roque1,2
Fabio de Oliveira Roque1,2 Carolina Ferreira Pauliquevis3Ademir Kleber Morbeck de Oliveira3
Carolina Ferreira Pauliquevis3Ademir Kleber Morbeck de Oliveira3 Diogo B. Provete1,4
Diogo B. Provete1,4 Judit K. Szabo5,6Franco Leandro Souza1
Judit K. Szabo5,6Franco Leandro Souza1- 1Instituto de Biociências, Universidade Federal de Mato Grosso do Sul, Campo Grande, Brazil
- 2Centre for Tropical Environmental and Sustainability Science and College of Science and Engineering, James Cook University, Cairns, QLD, Australia
- 3Programa de Pós-Graduação em Meio Ambiente e Desenvolvimento Regional, Universidade Anhanguera-Uniderp, Campo Grande, Brazil
- 4Gothenburg Global Biodiversity Centre, Gothenburg, Sweden
- 5Instituto de Biologia, Universidade Federal da Bahia, Salvador, Brazil
- 6College of Engineering, IT and Environment, Charles Darwin University, Darwin, NT, Australia
Birds provide many ecosystem services to people, including provisioning, regulating and cultural services. People attribute multiple cultural values to ecosystems and biodiversity and the diversity of these cultural values can be considered as cultural diversity. While human-nature interactions occur more frequently in cities and urbanization negatively affects different facets of avian biodiversity, little is known about its consequence for cultural diversity. Here, we assess how the urbanization gradient in Campo Grande, a Brazilian city in the Cerrado biodiversity hotspot, affects functional and cultural diversity associated with birds and if functional and cultural diversity are congruent. We also investigate the relation between urbanization gradient with functional traits and cultural values, weighted by species abundance. We used a dataset based on bird surveyed in 61 landscapes along a gradient of impervious surface cover. To estimate functional and cultural diversity, we used indices that estimate richness and divergence of functional traits and cultural values. We found that urbanization affected functional and cultural richness negatively, while there was no effect on functional and cultural divergence. Functional and cultural richness and functional and cultural divergence were weakly, but significantly correlated. Bird species that nest on trees decreased and those that nest in artificial structures and on the ground increased along the impervious surface gradient. Body size, diet, habitat, mating system, flock behavior, and all cultural values (number of times the species was mentioned by football teams, music or poetry, city flags and anthems, and folklore tales) were not significantly affected by impervious surface. The negative relationship between impervious surface and bird cultural richness may indicate that people living in more urbanized areas experience nature less compared to people in less urbanized areas, which can affect their psychological well-being. In these highly urbanized areas, contact with culturally valued birds and cultural services provided by birds may also diminish. The negative relationship between functional richness and urbanization also indicate that highly urbanized areas may be losing important ecosystems services provided by birds.
Introduction
By 2050, an additional of 2.5 billion people are expected to be living in cities around the world (United Nations, 2018). The increase in urban areas will exert a high pressure on natural systems, diminishing biodiversity and associated ecosystem services (Aronson et al., 2017), even though the emergence of Covid-19 may alter this scenario and drive a more sustainable urbanization process (United Nations Human Settlements Programme, 2020). Urban biodiversity provides provisioning services, such as pollination and seed dispersal, regulates climate and water and provides non-material or cultural services (Millenium Ecosystem Assessment, 2005; Daniel et al., 2012; Elmqvist et al., 2013; Cox and Gaston, 2016). These cultural ecosystem services are associated with multiple cultural values that people attribute to ecosystems and biodiversity, including aesthetic, inspirational and cultural heritage values through particular species or landscape that are strongly associated with human cultural identity (Millenium Ecosystem Assessment, 2005; Daniel et al., 2012; Tribot et al., 2018). The diversity of these cultural values attributed to ecosystems and biodiversity can be considered as cultural diversity (Garibaldi and Turner, 2004; Heyd, 2010). However, the cultural dimension of biodiversity and its relation with urbanization are poorly known (Cox et al., 2018; Schuetz and Johnston, 2019).
Humans may act as selective agents in cities, both indirectly by altering the set of species that occur in cities through the urbanization processes (Alberti, 2015; Goodness et al., 2016) and directly by selecting species (for instance by gardening) with particular traits (Niinemets and Peñuelas, 2008). Humans also appreciate esthetic aspects of biodiversity in urban ecosystems (Belaire et al., 2015; Goodness et al., 2016) and prioritize visually pleasant species (Conway and Vander Vecht, 2015; Leveau, 2019; Schuetz and Johnston, 2019). For instance, humans usually value birds with brighter and more contrasting plumage (Belaire et al., 2015; Schuetz and Johnston, 2019), larger body size (Murgui and Hedblom, 2017; Schuetz and Johnston, 2019) and certain feeding habits (Souza et al., 2019). Thereby, birds connect humans with nature, shaping cultural identity (Cox et al., 2018; Soga and Gaston, 2020) and improve human mental health (Cox et al., 2017). They also play a prominent functional role in cities, providing provisioning and regulation services, such as pollination, seed dispersal and insect control (Wenny et al., 2011; Gaston et al., 2018a). Therefore, the cultural and functional diversity of urban bird communities may be biased by anthropogenic effects and human perception of nature (Zmihorski et al., 2013; Pena et al., 2017; Schuetz and Johnston, 2019).
Studying birds as indicators of cultural services (Cox et al., 2018) is challenging in tropical regions, given the high socio-cultural diversity (Burnside et al., 2012). Brazil holds several thousand Important Bird and Biodiversity Areas, many of them near cities (Donald et al., 2019). It is a biologically megadiverse country harboring around 1,920 bird species (Piacentini et al., 2015), and it has a rich human culture related to ethnicities and languages (i.e., 305 indigenous ethnicities and 274 languages; IBGE, 2010). Human–nature interactions occur in different systems, including rural areas, indigenous lands and cities, and people from these systems interact with nature differently based on their social and cultural perspectives (Tryjanowski et al., 2015; Cámara-Leret et al., 2019). Considering that cities have denser human populations, they offer opportunities for more people to interact with nature (Soga and Gaston, 2020). For example, birds are important in Brazilian culture and are frequently represented in music, poetry, and sports (Straube, 2010; Dias et al., 2016; CEO, 2018).
A growing number of studies have called attention to changes in bird diversity in tropical cities and the associated ecological services, across a gradient of land use intensification and urbanization (Cox et al., 2018; Maruyama et al., 2019; Sol et al., 2020). The functional diversity of bird assemblages—measured by functional quadratic entropy—was 20% lower in highly urbanized areas compared to the surrounding natural habitats, driven by changes in species richness and evenness, leading to declines in functional redundancy (Sol et al., 2020). Even though frugivores and nectarivores provide essential ecosystem services, both are negatively affected by urbanization (Souza et al., 2019). If negative changes in functional diversity with increasing urbanization are also accompanied by the loss of cultural diversity (i.e., diversity of cultural values), extinctions of interaction between people and nature can accelerate a negative feedback loop and lead to further biodiversity loss.
Here we assess how the urbanization gradient in Campo Grande, a Brazilian city in the Cerrado biodiversity hotspot, affects functional and cultural diversity of birds and if functional and cultural diversity are congruent. To better understand which functional traits (i.e., measurable characteristics of individuals) and cultural values that people attribute to birds are affected by urbanization, we also investigate the correlation between functional traits and cultural values along an urbanization gradient, weighted by species abundance. Urbanization affects abundance and behavior of native urban bird communities negatively and only a few urban exploiters thrive (e.g., Souza et al., 2019; Tryjanowski et al., 2020). Birds are also under artificial selection, which favors bird traits valued by humans. Therefore, we expect that both estimates of cultural and functional diversity would be affected negatively by urbanization. However, as birds associated with cultural values differ in abundance, distribution, frequency of contact with humans, feeding habit and other characteristics (Zmihorski et al., 2013; Goodness et al., 2016; Gaston et al., 2018b; Schuetz and Johnston, 2019), we expect that the urbanization gradient would affect functional more than cultural diversity.
Materials and Methods
Location
Campo Grande is a city in central Brazil with 840,000 inhabitants and 104 inhabitants/km2. It has a marked seasonality, characterized by a dry (April–September) and a rainy (October–March) season, with a mean of 1,500 mm annual precipitation. The city harbors 572 bird species mostly found in natural habitats, such as Cerrado remnants and aquatic environments (Wikiaves, 2020). About a fifth (107) of the species have been reported in urban areas, including streets, gardens, public and private land (Souza et al., 2019).
Bird Dataset
We used assemblage composition data collected by Souza et al. (2019). They conducted point counts in February and March 2016. They divided the city into 61 hexagonal sampling sites, 16-ha each, along a gradient of urbanization, from non-urbanized (0% impervious surfaces) to highly urbanized (88.9%) land cover. Impervious surface was estimated using RapidEye base satellite images with a 5-m resolution. Each hexagon was surveyed once, at four points at least 200 m from one another (for more details see Souza et al., 2019).
Functional Traits and Cultural Values
We selected functional traits that represented phenotypic characteristics and may be affected by urbanization and functional traits that can influence ecosystem services in cities (effect traits), such as feeding habits (Petchey and Gaston, 2006; Luck et al., 2012; Pena et al., 2017). These functional traits included five categorical (diet, habitat, nesting substrate, mating system and flock behavior) and one continuous variable (body size) and were obtained from Sick (1997) and were complemented by del Hoyo et al. (2004) and Wilman et al. (2014) (Table 1 and Supplementary Table 1). Functional traits were organized into a species by traits matrix (Supplementary Table 1).
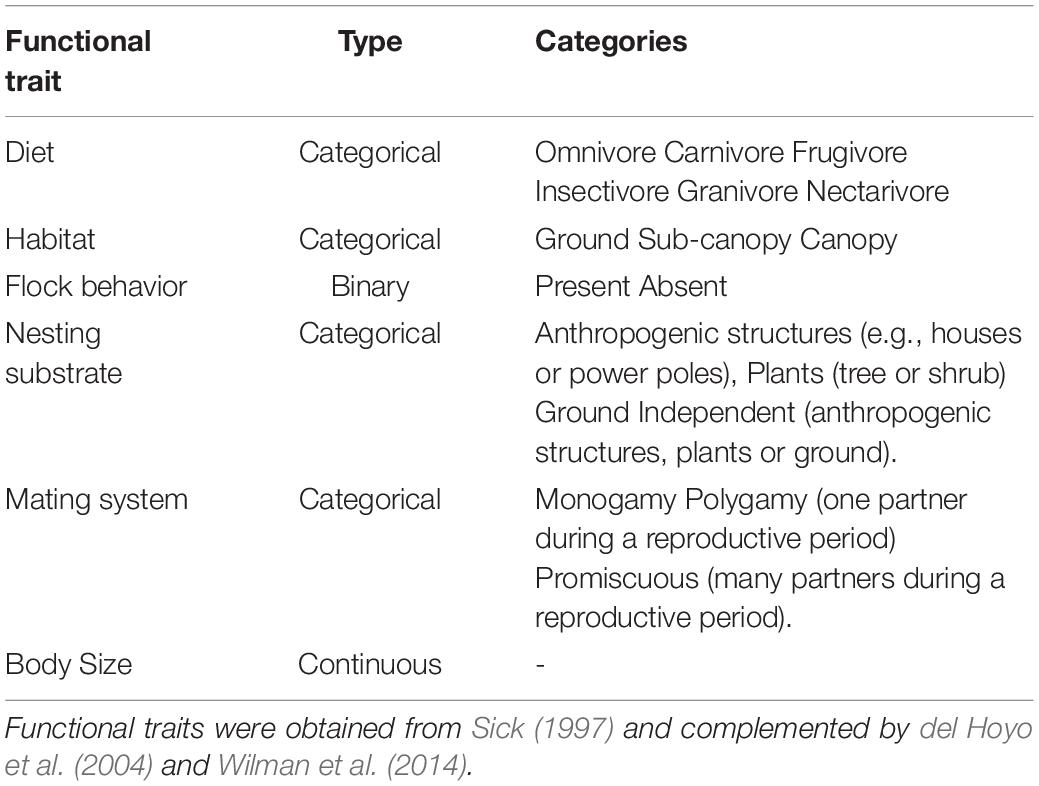
Table 1. Functional traits of 107 bird species sampled by Souza et al. (2019) in Campo Grande that were used in this study.
Cultural values included counts of birds used by football teams (badges and mascots), mentioned in music or poetry, present on city flags and mentioned in anthems and in folklore tales around Brazil (Table 2 and Supplementary Table 2). Following the Millenium Ecosystem Assessment (2005), cultural values of birds include those related to inspiration (e.g., mentioned in music, poetry or folklore), esthetic (e.g., used by football teams) and cultural heritage (e.g., used on city flags or in anthems). It is important to highlight that these cultural value categories can overlap (Daniel et al., 2012). For example, birds used by football teams also have a cultural heritage value, i.e., an experience shared across generations.
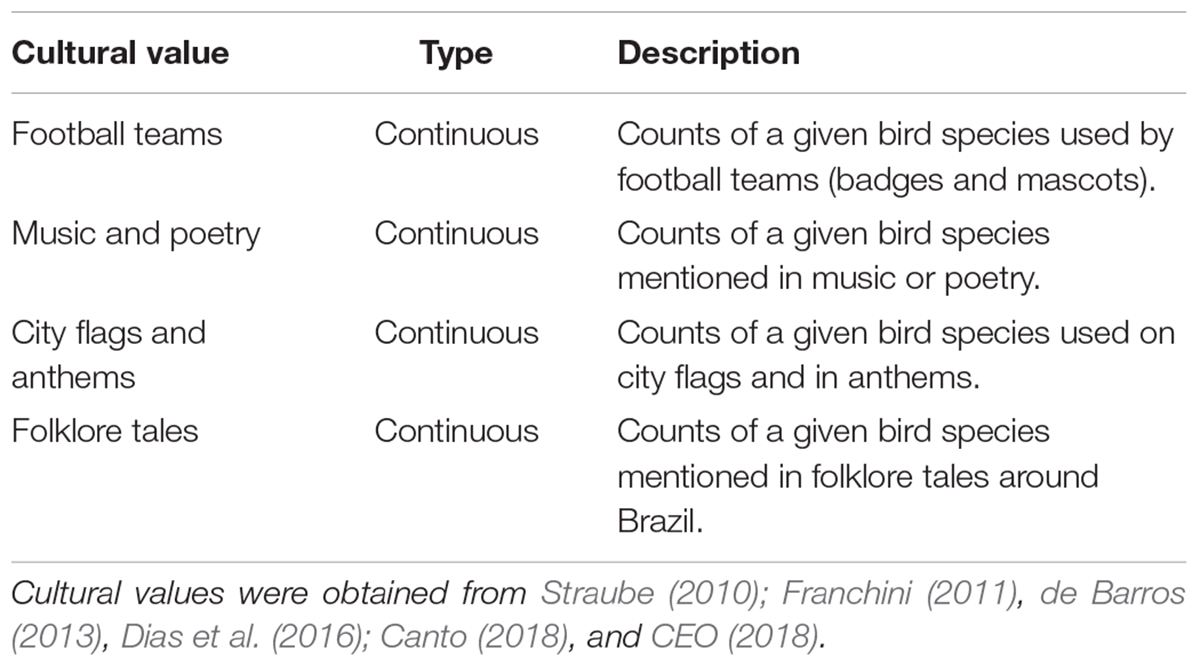
Table 2. Cultural values of 107 bird species sampled by Souza et al. (2019) in Campo Grande that were used in this study.
Cultural values were compiled from the gray literature and peer-reviewed articles, all of them published in Portuguese, using Google and Google Scholar, respectively. Searches were conducted in 2018 using a combination of keyword in Portuguese, including “birds and music,” “birds and football teams,” “birds and city flags,” “birds and city anthems” and “birds and folklore.” This search resulted in one article that assessed and summarized birds used on city flags and mentioned in anthems (Canto, 2018). We also located a website (Center for Brazilian Ornithological Studies) that recorded this sort of information (CEO, 2018). For football teams, we found two articles with information on the use of birds on badges and as mascots by Brazilian football teams (Straube, 2010; Dias et al., 2016). For music and poetry, we also included in our search the common names of birds, obtained from Wikiaves (2020), adding to our search the keywords in Portuguese “common name and music.” We found CEO (2018) as a source of information of birds mentioned in music and poetry. For poetry, we restricted our search to Manoel de Barros (1916–2014), a local poet widely known in Brazil for describing nature in his poems. Specifically, we searched his “Poesia Completa” (Complete Poetry) collection for mentions of birds (de Barros, 2013). For folklore tales, our search resulted in one book that compiled 100 Brazilian folklore stories (Franchini, 2011). As the same popular name can refer to various species (Straube et al., 2007), we recorded each mention under every possible species. Cultural values were organized into a species by cultural values matrix (Supplementary Table 2).
Statistical Analyses
Because the functional traits matrix contained both qualitative and quantitative traits, we converted it into a distance matrix using Gower distance (Podani, 1999). As all cultural values were quantitative, we applied Euclidean distance to convert them into a distance matrix. We then performed a principal coordinate analysis (PCoA) on the distance matrices (functional and cultural), and used the resulting PCoA axes as variables to compute functional and cultural diversity indices. To estimate functional and cultural diversity, we used indices that estimate richness and divergence of functional traits and cultural values. Functional and cultural richness were estimated by functional richness index (FRic) and Rao’s quadratic entropy (RaoQ) was used to estimate functional and cultural divergence. We estimated richness and divergence indices using the dbFD function in the FD package and the R version 4.0.0 (Laliberté et al., 2014; R Core Team, 2020). Using this approach, we obtained values of cultural and functional richness and divergence for each hexagon. FRic captures the amount of functional (or cultural) space occupied by species in a given community (Villéger et al., 2008), independently of the species abundance (Mason et al., 2005). RaoQ is calculated using the sum of pairwise distances between species weighted by their relative abundance (Botta-Dukát, 2005). RaoQ increases as the most abundant species exhibits increasingly extreme trait (or cultural values). While FRic is a measure of richness, i.e., the amount of functional or cultural space available, RaoQ is a measure of divergence, i.e., how the species are distributed in the functional or cultural space (more spread out or more clumped). The use of different indices is important, because each captures one aspect of functional and cultural diversity (Mouchet et al., 2010). Previous studies have recommended the use of a combination of richness and divergence indices to test community assembly mechanisms along stress gradients (Mason et al., 2013).
We used linear regression to relate the percentage of impervious surface (predictor variable) to richness and divergence (response variables) obtained from cultural values and functional traits. Although other predictors may be important in shaping urban bird communities, Souza et al. (2019) showed that impervious surface was the main driver of species richness and feeding groups for our study area. We visually inspected residuals for homogeneity of variance and normality (Supplementary Figures 1–4). We used Pearson correlation to test the congruence between functional and cultural richness and functional and cultural divergence. We applied the fourth-corner analysis (Dray and Legendre, 2008) using the ade4 package in R (Dray and Dufour, 2007) to better understand the relationships between cultural value and functional trait composition and impervious surface. The fourth-corner analysis estimates the correlation between traits (or cultural values) and environmental predictors and its significance is tested by permuting the value of sites and the species (Dray and Legendre, 2008).
Results
Cultural richness and divergence had larger values and variation compared to functional richness and divergence. Cultural richness and functional richness varied from 0.15 to 41.42 and from 3e–9 to 4e–5, respectively. Cultural divergence and functional divergence varied from 0.84 to 9.12 and from 0.08 to 0.16, respectively. Both functional and cultural richness were negatively affected by impervious surface (Figures 1A,C). Overall, functional and cultural richness had higher values in the peripheric regions of Campo Grande (Figures 2A,C). Impervious surface explained 21% of cultural richness variation (slope = – 0.15, F = 15.85, DF = 59; P = 0.0001) and 29% of functional richness variation (slope = – 5.4e–8, F = 21.71, DF = 59; P = 0.00001). However, impervious surface did not affect cultural (slope = – 0.003, F = 0.09, P = 0.75) nor functional divergence (slope = 4.9e–05, F = 1.58, P = 0.21; Figures 1B,D, 2B,D). Cultural and functional richness were weakly, but significantly correlated (Pearson r = 0.45, P = 0.0002; Figure 3A), similar to cultural and functional divergence (Pearson r = 0.30, P = 0.01) (Figure 3B).
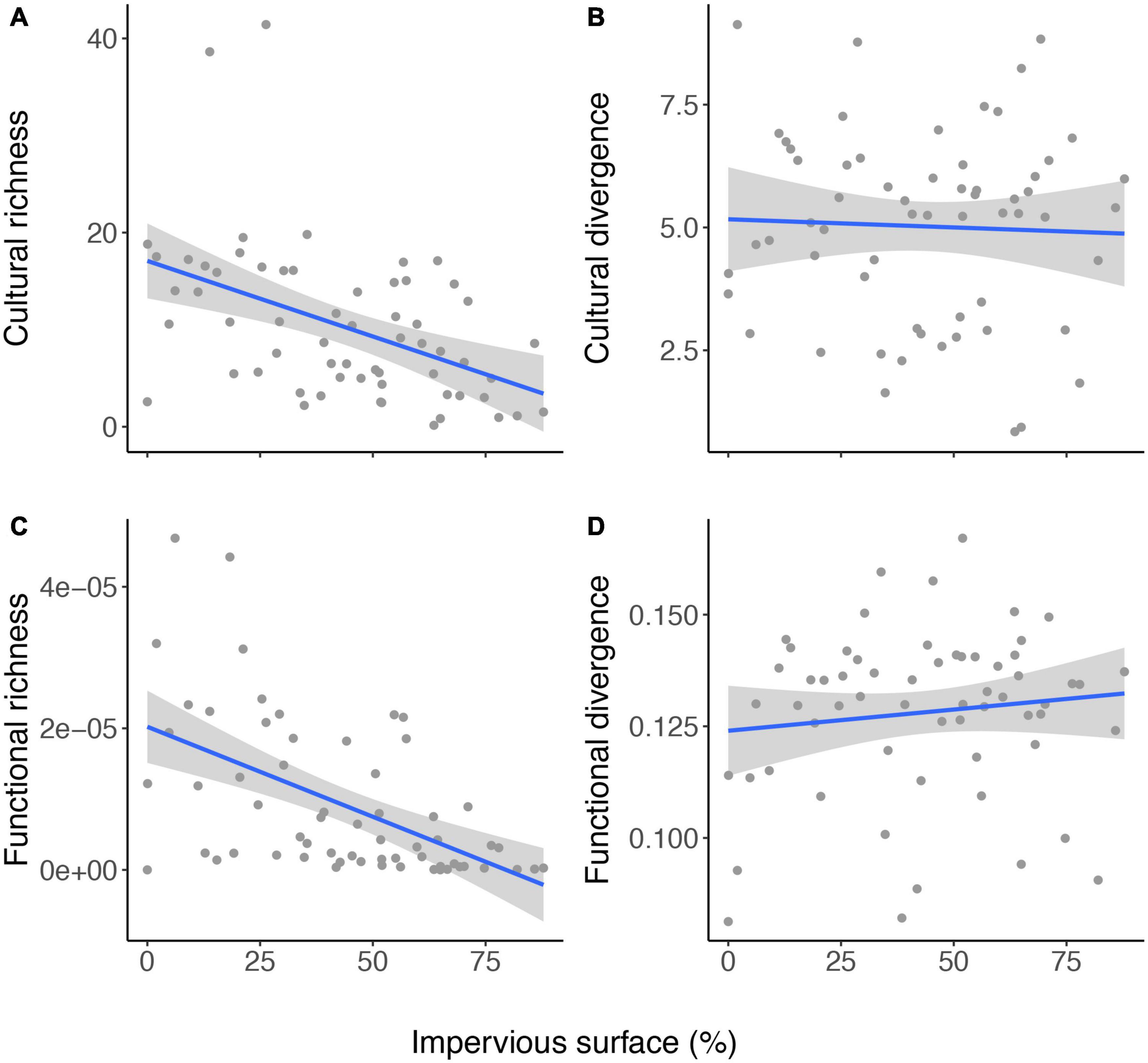
Figure 1. Relationships between the percentage of impervious surface and cultural (A) and functional (B) richness, and cultural (C) and functional (D) divergence for bird sampled in 61 hexagons in Campo Grande, central Brazil.
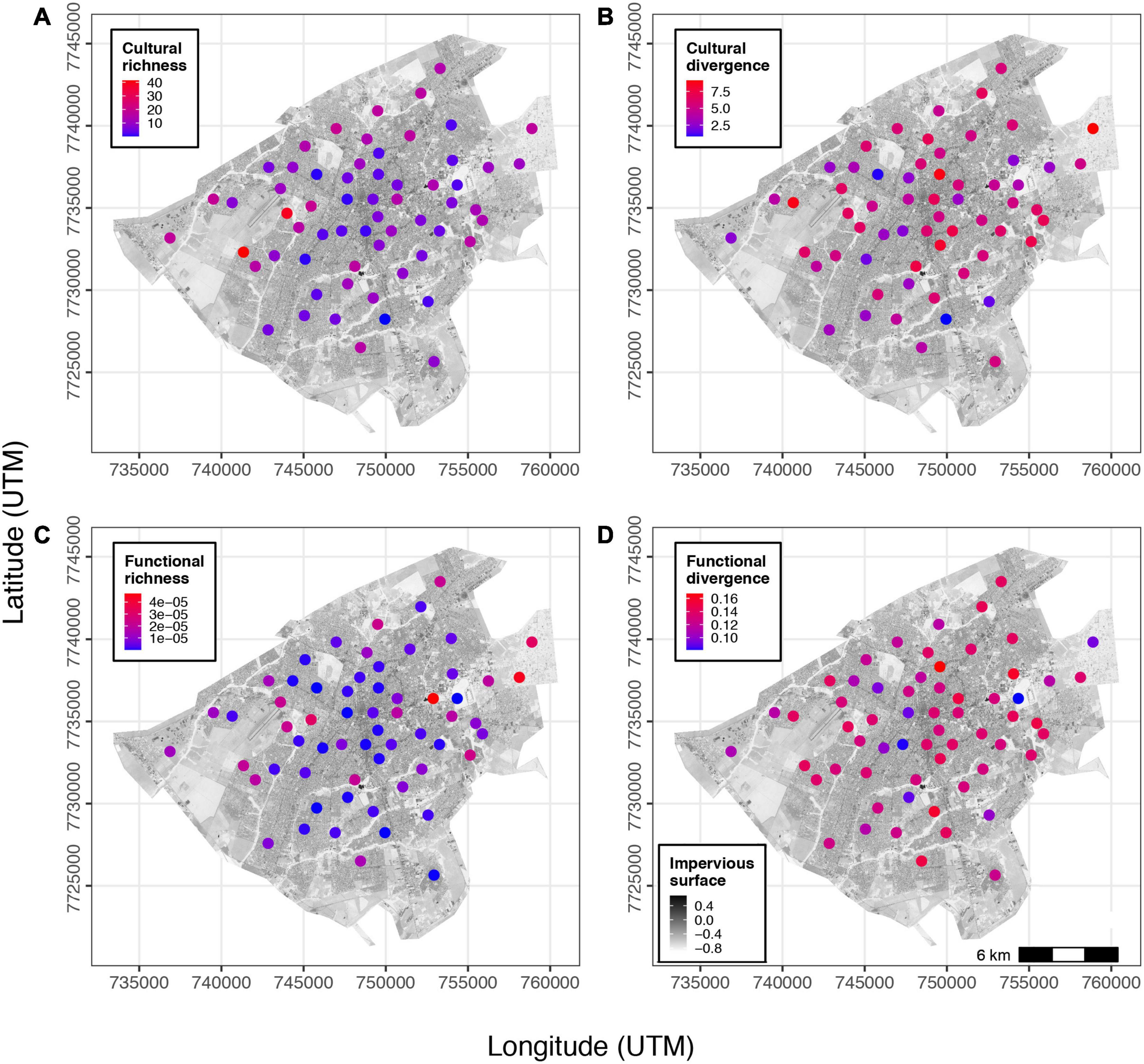
Figure 2. Spatial distribution of cultural (A) and functional richness (B), and cultural (C) and functional divergence (D) along the gradient of impervious surface cover in Campo Grande, central Brazil.
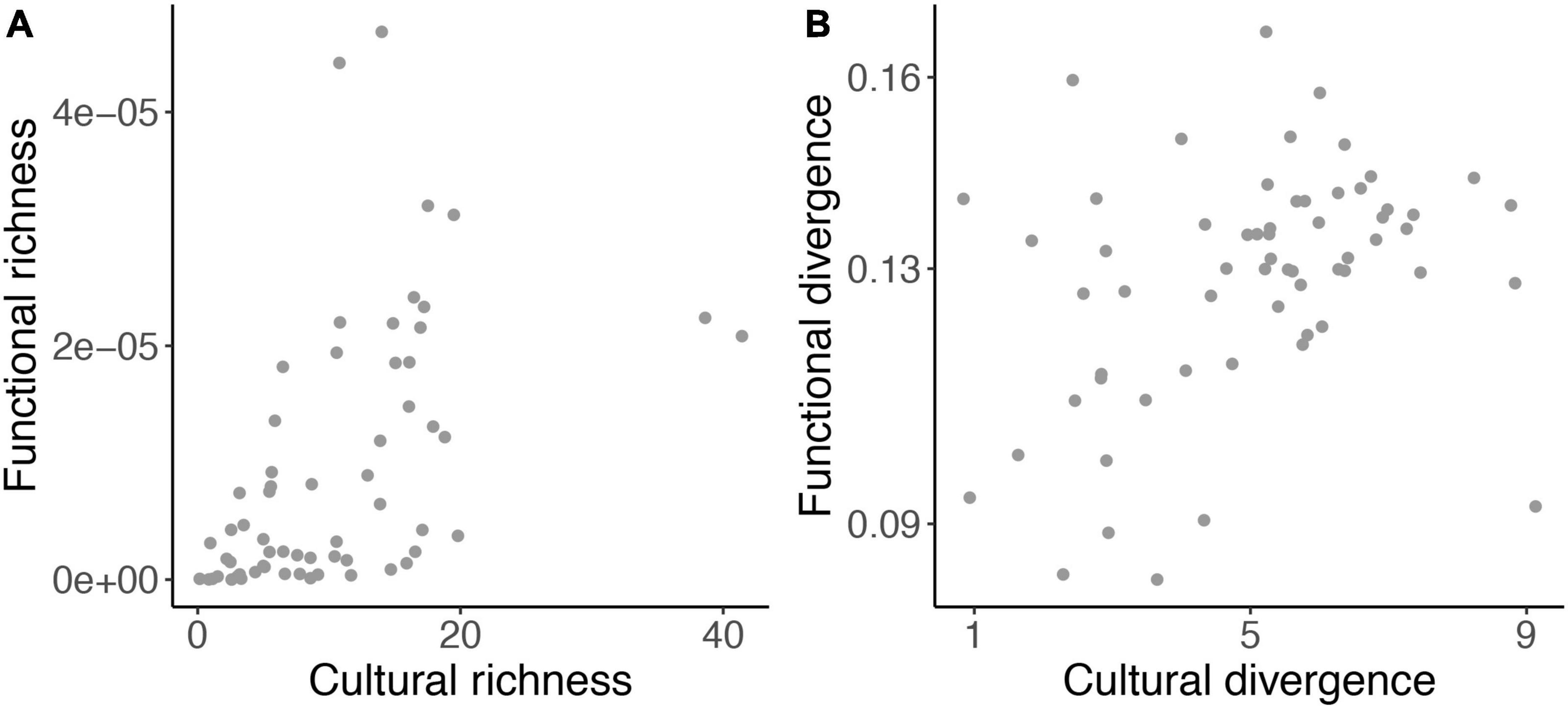
Figure 3. Relationships between cultural and functional richness (A), and cultural and functional divergence (B) for bird sampled in 61 hexagons in Campo Grande, central Brazil.
Impervious surface negatively affected bird nesting (r = – 0.23; P = 0.01), showing that urbanization decreased the number of birds that nested on trees and increased those that nested on artificial structures and on the ground. Impervious surface did not affect any of the remaining functional traits and or cultural values significantly (Table 3).
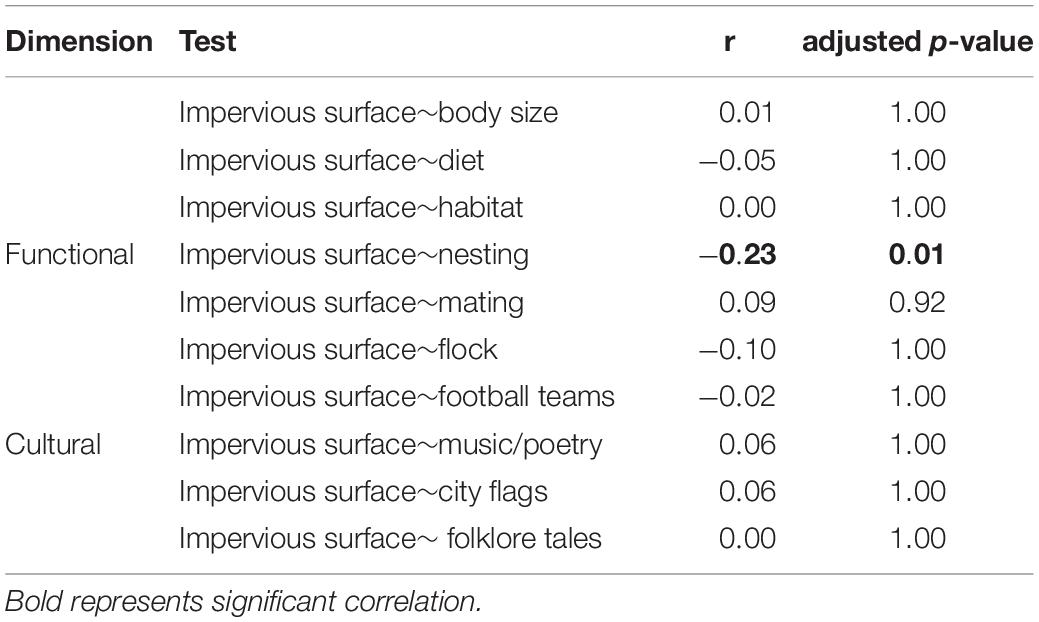
Table 3. Relationships between bird functional traits and cultural values and impervious surface in Campo Grande obtained from fourth corner analysis.
Discussion
We found that bird functional and cultural richness (i.e., the amount of functional or cultural space occupied by species) decreased along a gradient of impervious surface in a city located in the Cerrado hotspot. This pattern means that people who live in highly urbanized areas may have less contact with culturally and functionally important bird species compared to those in less urbanized areas. However, functional and cultural divergence (i.e., how the species are distributed in the functional or cultural space) were not affected by impervious surface, which could be related to the sensitivity of these indexes to the abundance of some species. Both functional and cultural richness and divergence were weakly correlated, demonstrating that the loss of functional diversity implies in a certain loss of cultural diversity and vice-versa. In highly urbanized areas, people may experience less contact with birds that are culturally valued and cultural services provided by birds may be diminished. Among individual traits, we found that impervious surface negatively affected bird nesting.
Impervious surface affected functional and cultural richness negatively. This negative effect of urbanization on bird functional diversity was also found by Sol et al. (2020) using a global dataset. Overall, urbanization represents a drastic land use change, converting natural habitats into impervious surface and thereby reducing the amount of resources (Chace and Walsh, 2006). In Brazil, exposure to noise was the main variable responsible for variation in bird functional diversity in Belo Horizonte (Pena et al., 2017) and bird richness was affected by noise level and distance to water in São Paulo (Barbosa et al., 2020). As only a few species can cope with these novel habitats, the functional diversity will decrease (Schütz and Schulze, 2015; Concepción et al., 2017).
We found that urbanization decreased the number of tree-nesting birds, but increased the number of birds nesting in or on artificial surface and on the ground. Our results partially agreed with Concepción et al. (2017), who found that urbanization mostly affected nesting traits by reducing the number of tree-, and ground-nesting species. Therefore, people have fewer opportunities to observe tree-nesting birds and nestlings in highly urbanized areas. The loss of this interaction, particularly between humans and fledglings, may be concerning, because young animals in general stimulate positive human attitudes, such as compassion, empathy and prosocial and environmental behavior toward animals (Young et al., 2018; Castillo-Huitrón et al., 2020). Despite this, gardens play an important role to maintain bird species richness in Campo Grande, including habitat for parrots, parakeets, macaws, and other tree-nesting species (Souza et al., 2019). The negative relationship between functional diversity and nesting traits with urbanization indicates that increasing green surface should be a priority to maintain tree-nesting birds, as well as bird functional diversity, ensuring essential ecosystem services in cities.
The effects of urbanization on bird cultural diversity and cultural values are less known. Most studies rely on assessing how humans value birds and which traits increased the value of a particular species (Zmihorski et al., 2013; Cox and Gaston, 2016; Cox et al., 2018; Schuetz and Johnston, 2019; Echeverri et al., 2020). Despite the many publications on the topic, to our knowledge, no studies have related human well-being with cultural diversity associated with birds. Watching birds can improve the well-being and metal health of urban residents (Belaire et al., 2015; Cox and Gaston, 2016; Hedblom et al., 2017). The negative relationship between bird cultural richness and the amount of impervious surface may indicate that people lose the experience of nature in more urbanized areas and this can affect human psychological well-being (Cox and Gaston, 2016; Hedblom et al., 2017; Cox et al., 2018). Nevertheless, urbanization did not affect any specific cultural value. Unlike functional traits that can be selected by environmental gradients, as we showed here, cultural values are attributed to biodiversity by people and these values may not be strongly correlated with functional traits or biological characteristics (Figure 3) and may vary depending on the culture (Cámara-Leret et al., 2019; Schuetz and Johnston, 2019; Echeverri et al., 2020). For example, in our study, the most culturally important birds, such as turkey vulture (Cathartes aura), certain passerines (e.g., rufous-bellied thrush Turdus rufiventris, chalk-browed mockingbird Mimus saturninus, and southern rough-winged swallow Stelgidopteryx ruficollis) and parrots and parakeets (e.g., turquoise-fronted amazon Amazona aestiva, yellow-chevroned parakeet Brotogeris chiriri, and white-eyed conure Psittacara leucophthalmus) have different feeding habits, size, habitat preference, and nesting, which can explain the lack of relationship between urbanization and cultural values (Goodness et al., 2016).
For species associated with less urbanized areas, such as greater thornbird (Phacellodomus ruber), violaceous euphonia (Euphonia violacea), and yellow-chinned spinetail (Certhiaxis cinnamomeus), we found no or few mentions of cultural significance. Nevertheless, our findings may not reflect trends across other societies, considering that indigenous people, rural and urban dwellers have different views of nature, which can impact cultural values of biodiversity (Tryjanowski et al., 2015; Cámara-Leret et al., 2019). For example, people from rural and urban areas differ in the amount and type of food they supplied to birds, as a consequence of social and cultural differences (Tryjanowski et al., 2015). In addition, cultural values are not static and can be evaluated and interpreted in different ways by different actors (Tengberg et al., 2012). The lack of cultural values of some species found in Campo Grande could potentially result in low importance attributed to the local extinction of these birds in urban landscapes. This can create a negative feedback, as people in urban areas can easily adapt to impoverished landscapes (Cox and Gaston, 2016; Soga and Gaston, 2016). As a result, the biodiversity of these landscape can further deteriorate, thus exacerbating the “extinction of experience” phenomenon (Pyle, 1993; Soga and Gaston, 2016). Moreover, bird conservation efforts are tied to social values (Ainsworth et al., 2016, 2018), directing decision-making processes toward more valued species. Environmental awareness and citizen-science programs are useful to expand the perception of these species with low or no associated cultural value (Trumbull et al., 2000). Therefore, birdwatching, environmental education, citizen science (including monitoring) and other ongoing initiatives should be improved and expanded in Campo Grande (Mamede and Benites, 2018).
Using aggregated measures, such as richness and divergence, we showed that cultural and functional measures were weakly correlated and had a similar relationship with impervious surface. These results provide some clear messages. First, either functional or cultural indices can be used to monitor the effects of urbanization on bird assemblages. Second, the correlation between functional and cultural richness with urbanization indicates that in city centers and highly urbanized areas, people may be losing positive nature experiences, thereby negatively affecting human well-being (Cox and Gaston, 2016; Hedblom et al., 2017; Cox et al., 2018). The loss may also affect ecosystem services, including pest control and pollination (Wenny et al., 2011; Gaston et al., 2018a). Urbanization also drastically decreases bird richness (Souza et al., 2019; Barbosa et al., 2020) and induces homogenization of plumage color, characteristics that attract people’s attention (Schuetz and Johnston, 2019). Highly urbanized areas may also increase undesirable human-bird experiences (Cox et al., 2018). On the other hand, parks, lakes and other less urbanized areas can serve as locations for positive interactions between birds and people, including the possibility to see native passerines, parrots, parakeets and other culturally valued species.
Contrary to our prior hypothesis and the results of Sol et al. (2020), we found that cultural and functional divergence were not correlated with impervious surface cover, i.e., impervious cover did not decrease functional or cultural dissimilarity between co-occurring species. In fact, most sites contained species with intermediate abundance and trait values, indicating intermediate to low levels of divergence. For functional traits, some generalist may be responsible for this pattern. For example, the most abundant and widespread species (house sparrow Passer domesticus, feral pigeon Columba livia, yellow-chevroned parakeet, saffron finch Sicalis flaveola, and rufous hornero Furnarius rufus) are of small to intermediate size, with varied feeding and habitat requirements, nesting and mating strategies and flocking behavior. In Belo Horizonte, another major inland city in Brazil, exposure to noise negatively affected bird divergence (RaoQ) estimated from similar traits, but the average distance to a green surface had a positive effect on divergence (Pena et al., 2017). Concepción et al. (2017) reported that divergence (RaoQ) of nesting and diet traits were positively related to urban areas, indicating trait divergence along a gradient of urban use. For cultural values, some culturally important species, such as the turkey vulture and the chalk-browed mockingbird were not necessarily the most abundant. Other bird species (rufous-bellied thrush, saffron finch, and yellow-chevroned parakeet) were widespread and abundant and also of high cultural importance. These patterns can explain the lack of relationship between cultural RaoQ and impervious surface cover.
In urban areas, people usually interact with bird species that are common, diurnal, and easy to see or hear (Gaston et al., 2018b). The increasing familiarity with these birds may lead to more positive perceptions and interactions (Echeverri et al., 2020). Therefore, we would expect these common species to be the most represented in human culture. While Echeverri et al. (2020) reported that in Costa Rica people preferred species associated with forests, we found that most species of cultural importance were not dependent on forests. This difference can result from the different methods used to assess cultural services. Unlike us, Echeverri et al. (2020) used online surveys to study cultural services, while we used common cultural representations (football teams, music, poetry, city flags, anthems, and folklore tales), we acknowledge the existence of many others. Similarly, different functional traits could be used to estimate functional diversity. We used traits available for the species in our dataset and in a published study assessing the effects of urbanization on birds in Brazil (Pena et al., 2017).
The loss of functional and cultural bird diversity due to urbanization suggests that people have fewer opportunities to experience biodiversity in highly urbanized areas and, consequently, their well-being might suffer (Cox and Gaston, 2016; Cox et al., 2017). It is not only about losing personal benefits, but this loss of functional and cultural bird diversity creates a negative cycle of disaffection. This can have disastrous consequences, since such experiences with nature are irreplaceable and direct personal contact with the natural environment is vital to weave a person’s emotional intimacy with nature (Pyle, 1993; Soga and Gaston, 2016).
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethics Statement
Ethical review and approval were not required for the animal study because the dataset we used–Souza et al. (2019) (https://doi.org/10.1016/j.landurbplan.2019.103643) – recorded all bird species seen and/or heard, so the authors did not sample or manipulate birds.
Author Contributions
FV-N, FR, CP, and FS conceived and designed the study and wrote the original draft of the manuscript. CP and AO gathered cultural traits. FV-N carried out all the formal analysis. FV-N, FR, CP, AO, DP, JS, and FS contributed to interpretation of data, writing, and reviewed the manuscript.
Funding
FV-N was supported by the grant no. 88882.317337/2019-01, Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). We thank CNPq that has continuously supported this project with productivity grants (grant no. 301306/2018-4 to FS, grant no. 302755/2018-7 to FR, and grant no. 304946/2017-6 to AO). This study was partly financed by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior–Brasil (CAPES)–Finance Code 001.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful to Berinaldo Bueno, Francisco Severo-Neto, and José Manoel Ochoa-Quintero for surveying birds and for contributing to the original sampling design as described in Souza et al. (2019). We thank the two reviewers for their useful contribution.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2021.615797/full#supplementary-material
References
Ainsworth, G. B., Aslin, H. J., Weston, M. A., and Garnett, S. T. (2016). Do social values influence levels of conservation effort in threatened species? The case of two Australian chats. Oryx 50, 636–645. doi: 10.1017/S0030605315000538
Ainsworth, G. B., Fitzsimons, J. A., Weston, M. A., and Garnett, S. T. (2018). The culture of bird conservation: Australian stakeholder values regarding iconic, flagship and rare birds. Biodivers. Conserv. 27, 345–363. doi: 10.1007/s10531-017-1438-1
Alberti, M. (2015). Eco-evolutionary dynamics in an urbanizing planet. Trends Ecol. Evol. 30, 114–126. doi: 10.1016/j.tree.2014.11.007
Aronson, M. F. J., Lepczyk, C. A., Evans, K. L., Goddard, M. A., Lerman, S. B., MacIvor, J. S., et al. (2017). Biodiversity in the city: key challenges for urban green space management. Front. Ecol. Environ. 15, 189–196. doi: 10.1002/fee.1480
Barbosa, K. V., de, C., Rodewald, A. D., Ribeiro, M. C., and Jahn, A. E. (2020). Noise level and water distance drive resident and migratory bird species richness within a Neotropical megacity. Landsc. Urban Plan. 197:103769. doi: 10.1016/j.landurbplan.2020.103769
Belaire, J. A., Westphal, L. M., Whelan, C. J., and Minor, E. S. (2015). Urban residents’ perceptions of birds in the neighborhood: biodiversity, cultural ecosystem services, and disservices. Condor 117, 192–202. doi: 10.1650/CONDOR-14-128.1
Botta-Dukát, Z. (2005). Rao’s quadratic entropy as a measure of functional diversity based on multiple traits. J. Veg. Sci. 16, 533–540. doi: 10.1111/j.1654-1103.2005.tb02393.x
Burnside, W. R., Brown, J. H., Burger, O., Hamilton, M. J., Moses, M., and Bettencourt, L. M. A. (2012). Human macroecology: linking pattern and process in big-picture human ecology. Biol. Rev. 87, 194–208. doi: 10.1111/j.1469-185X.2011.00192.x
Cámara-Leret, R., Fortuna, M. A., and Bascompte, J. (2019). Indigenous knowledge networks in the face of global change. Proc. Natl. Acad. Sci. U.S.A. 116, 9913–9918. doi: 10.1073/pnas.1821843116
Canto, J. B. D. M. (2018). Representação da avifauna nos brasões, bandeiras e hinos dos estados e de municípios brasileiros. Atualidades Ornitol. 202, 67–81.
Castillo-Huitrón, N. M., Naranjo, E. J., Santos-Fita, D., and Estrada-Lugo, E. (2020). The importance of human emotions for wildlife conservation. Front. Psychol. 11:1277. doi: 10.3389/fpsyg.2020.01277
CEO (2018). Centro de Estudos Ornitológicos. Available online at: http://www.ceo.org.br (accessed February 13, 2019).
Chace, J. F., and Walsh, J. J. (2006). Urban effects on native avifauna: a review. Landsc. Urban Plan. 74, 46–69. doi: 10.1016/j.landurbplan.2004.08.007
Concepción, E. D., Götzenberger, L., Nobis, M. P., de Bello, F., Obrist, M. K., and Moretti, M. (2017). Contrasting trait assembly patterns in plant and bird communities along environmental and human-induced land-use gradients. Ecography 40, 753–763. doi: 10.1111/ecog.02121
Conway, T. M., and Vander Vecht, J. (2015). Growing a diverse urban forest: species selection decisions by practitioners planting and supplying trees. Landsc. Urban Plan. 138, 1–10. doi: 10.1016/j.landurbplan.2015.01.007
Cox, D. T. C., and Gaston, K. J. (2016). Urban bird feeding: connecting people with nature. PLoS One 11:e0158717. doi: 10.1371/journal.pone.0158717
Cox, D. T. C., Hudson, H. L., Plummer, K. E., Siriwardena, G. M., Anderson, K., Hancock, S., et al. (2018). Covariation in urban birds providing cultural services or disservices and people. J. Appl. Ecol. 55, 2308–2319. doi: 10.1111/1365-2664.13146
Cox, D. T. C., Shanahan, D. F., Hudson, H. L., Plummer, K. E., Siriwardena, G. M., Fuller, R. A., et al. (2017). Doses of neighborhood nature: the benefits for mental health of living with nature. Bioscience 67, 147–155. doi: 10.1093/biosci/biw173
Daniel, T. C., Muhar, A., Arnberger, A., Aznar, O., Boyd, J. W., Chan, K. M. A., et al. (2012). Contributions of cultural services to the ecosystem services agenda. Proc. Natl. Acad. Sci. U.S.A. 109, 8812–8819. doi: 10.1073/pnas.1114773109
del Hoyo, J., Sargatal, J., and Christie, D. A. (2004). Handbook of the Birds of the World. Barcelona: Lynx Edicions.
Dias, D. V. C., Brandão, L. E. D., Rodrigues, H. T. S., Sousa, V. B. P., and Barros, M. D. M. (2016). As aves como mascotes dos times brasileiros de futebol. Trilhas Pedagógicas 6, 9–26. doi: 10.1017/CBO9781107415324.004
Donald, P. F., Fishpool, L. D. C., Ajagbe, A., Bennun, L. A., Bunting, G., Burfield, I. J., et al. (2019). Important Bird and Biodiversity Areas (IBAs): the development and characteristics of a global inventory of key sites for biodiversity. Bird Conserv. Int. 29, 177–198. doi: 10.1017/S0959270918000102
Dray, S., and Dufour, A. B. (2007). The ade4 package: implementing the duality diagram for ecologists. J. Stat. Softw. 22, 1–20.
Dray, S., and Legendre, P. (2008). Testing the species traits environment relationships: the fourth-corner problem revisited. Ecology 89, 3400–3412. doi: 10.1890/08-0349.1
Echeverri, A., Karp, D. S., Naidoo, R., Tobias, J. A., Zhao, J., and Chan, K. M. A. (2020). Can avian functional traits predict cultural ecosystem services? People Nat. 2, 138–151. doi: 10.1002/pan3.10058
Elmqvist, T., Fragkias, M., Goodness, J., Güneralp, B., Parnell, S., Schewenius, M., et al. (2013). Urbanization, Biodiversity and Ecosystem Services: Challenges and Opportunities: A Global Assessment. Dordrecht: Springer.
Garibaldi, A., and Turner, N. (2004). Cultural keystone species: implications for ecological conservation and restoration. Ecol. Soc. 9:1. doi: 10.5751/ES-00669-090301
Gaston, K. J., Cox, D. T. C., Canavelli, S. B., García, D., Hughes, B., Maas, B., et al. (2018a). Population abundance and ecosystem service provision: the case of birds. Bioscience 68, 264–272. doi: 10.1093/biosci/biy005
Gaston, K. J., Soga, M., Duffy, J. P., Garrett, J. K., Gaston, S., and Cox, D. T. C. (2018b). Personalised ecology. Trends Ecol. Evol. 33, 916–925. doi: 10.1016/j.tree.2018.09.012
Goodness, J., Andersson, E., Anderson, P. M. L., and Elmqvist, T. (2016). Exploring the links between functional traits and cultural ecosystem services to enhance urban ecosystem management. Ecol. Indic. 70, 597–605. doi: 10.1016/j.ecolind.2016.02.031
Hedblom, M., Knez, I., and Gunnarsson, B. (2017). “Bird diversity improves the well-being of city resident,” in Ecology and Conservation of Birds in Urban Environments, eds E. Murgui and M. Hedblom (Cham: Springer), 287–306. doi: 10.1007/978-3-319-43314-1_15
Heyd, D. (2010). Cultural diversity and biodiversity: a tempting analogy. Crit. Rev. Int. Soc. Polit. Philos. 13, 159–179. doi: 10.1080/13698230903326315
IBGE (2010). O Brasil Indígena. Available online at: http://www.funai.gov.br/index.981php/indios-no-brasil/o-brasil-indigena-ibge (accessed September 21, 2020)
Laliberté, E., Legendre, P., and Shipley, B. (2014). FD: Measuring Functional Diversity From Multiple Traits, and Other Tools for Functional Ecology. R Package Version 1.0-12.
Leveau, L. M. (2019). Urbanization induces bird color homogenization. Landsc. Urban Plan. 192:103645. doi: 10.1016/j.landurbplan.2019.103645
Luck, G. W., Lavorel, S., Mcintyre, S., and Lumb, K. (2012). Improving the application of vertebrate trait-based frameworks to the study of ecosystem services. J. Anim. Ecol. 81, 1065–1076. doi: 10.1111/j.1365-2656.2012.01974.x
Mamede, S., and Benites, M. (2018). Por que Campo Grande é a capital brasileira do turismo de observação de aves e propostas para o fortalecimento da cultura local em relação a esta prática. Atualidades Ornitológicas 201, 8–15.
Maruyama, P. K., Bonizário, C., Marcon, A. P., D’Angelo, G., da Silva, M. M., da Silva Neto, E. N., et al. (2019). Plant-hummingbird interaction networks in urban areas: generalization and the importance of trees with specialized flowers as a nectar resource for pollinator conservation. Biol. Conserv. 230, 187–194. doi: 10.1016/j.biocon.2018.12.012
Mason, N. W. H., De Bello, F., Mouillot, D., Pavoine, S., and Dray, S. (2013). A guide for using functional diversity indices to reveal changes in assembly processes along ecological gradients. J. Veg. Sci. 24, 794–806. doi: 10.1111/jvs.12013
Mason, N. W. H., Mouillot, D., Lee, W. G., and Wilson, J. B. (2005). Functional richness, functional evenness and functional divergence: the primary components of functional diversity. Oikos 111, 112–118. doi: 10.1111/j.0030-1299.2005.13886.x
Millenium Ecosystem Assessment (2005). Ecosystems and Human Well-Being: Biodiversity Synthesis. Washington, DC: World Resources Institute.
Mouchet, M. A., Villéger, S., Mason, N. W. H., and Mouillot, D. (2010). Functional diversity measures: an overview of their redundancy and their ability to discriminate community assembly rules. Funct. Ecol. 24, 867–876. doi: 10.1111/j.1365-2435.2010.01695.x
Murgui, E., and Hedblom, M. (2017). Ecology and Conservation of Birds in Urban Environments. Berlin: Springer International Publishing.
Niinemets, Ü, and Peñuelas, J. (2008). Gardening and urban landscaping: significant players in global change. Trends Plant Sci. 13, 60–65. doi: 10.1016/j.tplants.2007.11.009
Pena, J. C., de, C., Martello, F., Ribeiro, M. C., Armitage, R. A., Young, R. J., et al. (2017). Street trees reduce the negative effects of urbanization on birds. PLoS One 12:e0174484. doi: 10.1371/journal.pone.0174484
Petchey, O. L., and Gaston, K. J. (2006). Functional diversity: back to basics and looking forward. Ecol. Lett. 9, 741–758. doi: 10.1111/j.1461-0248.2006.00924.x
Piacentini, V. Q., Aleixo, A., Agne, C. E., Maurício, G. N., Pacheco, J. F., Bravo, G. A., et al. (2015). Annotated checklist of the birds of Brazil by the Brazilian ornithological records committee. Rev. Bras. Ornitol. 23, 91–298. doi: 10.1007/bf03544294
Podani, J. (1999). Extending gower’s general coefficient of similarity to ordinal characters. Taxon 48, 331–340. doi: 10.2307/1224438
R Core Team (2020). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Schuetz, J. G., and Johnston, A. (2019). Characterizing the cultural niches of North American birds. Proc. Natl. Acad. Sci. U.S.A. 166, 10868–10873. doi: 10.1073/pnas.1820670116
Schütz, C., and Schulze, C. H. (2015). Functional diversity of urban bird communities: effects of landscape composition, green space area and vegetation cover. Ecol. Evol. 5, 5230–5239. doi: 10.1002/ece3.1778
Soga, M., and Gaston, K. J. (2016). Extinction of experience: the loss of human-nature interactions. Front. Ecol. Environ. 14, 94–101. doi: 10.1002/fee.1225
Soga, M., and Gaston, K. J. (2020). The ecology of human–nature interactions. Proc. Biol. Sci. 287:20191882. doi: 10.1098/rspb.2019.1882
Sol, D., Trisos, C., Múrria, C., Jeliazkov, A., González-Lagos, C., and Pigot, A. L., et al. (2020). The worldwide impact of urbanisation on avian functional diversity. Ecol. Lett. 23, 962–972. doi: 10.1111/ele.13495
Souza, F. L., Valente-Neto, F., Severo-Neto, F., Bueno, B., Ochoa-Quintero, J. M., Laps, R. R., et al. (2019). Impervious surface and heterogeneity are opposite drivers to maintain bird richness in a Cerrado city. Landsc. Urban Plan. 192:103643. doi: 10.1016/j.landurbplan.2019.103643
Straube, F. C. (2010). As aves nos símbolos do futebol brasileiro: escudos. Atualidades Ornitológicas 158, 33–48.
Straube, F. C., Accordi, I. A., and Argel, M. (2007). Nomes populares de aves brasileiras coletados por Johann Natterer (1817-1835). Atualidades Ornitológicas 136, 1–6.
Tengberg, A., Fredholm, S., Eliasson, I., Knez, I., Saltzman, K., and Wetterberg, O. (2012). Cultural ecosystem services provided by landscapes: assessment of heritage values and identity. Ecosyst. Serv. 2, 14–26. doi: 10.1016/j.ecoser.2012.07.006
Tribot, A. S., Deter, J., and Mouquet, N. (2018). Integrating the aesthetic value of landscapes and biological diversity. Proc. R. Soc. B Biol. Sci. 285:20180971. doi: 10.1098/rspb.2018.0971
Trumbull, D. J., Bonney, R., Bascom, D., and Cabral, A. (2000). Thinking scientifically during participation in a citizen-science project. Sci. Educ. 84, 265–275. doi: 10.1002/(sici)1098-237x(200003)84:2<265::aid-sce7>3.0.co;2-5
Tryjanowski, P., Kosicki, J. Z., Hromada, M., and Mikula, P. (2020). The emergence of tolerance of human disturbance in Neotropical birds. J. Trop. Ecol. 36, 1–5. doi: 10.1017/S0266467419000282
Tryjanowski, P., Skórka, P., Sparks, T. H., Biaduń, W., Brauze, T., Hetmański, T., et al. (2015). Urban and rural habitats differ in number and type of bird feeders and in bird species consuming supplementary food. Environ. Sci. Pollut. Res. 22, 15097–15103. doi: 10.1007/s11356-015-4723-0
United Nations (2018). World Urbanization Prospects. Available online at: https://population.un.org/wup/Publications/Files/WUP2018-Report.pdf (accessed September 2, 2020).
United Nations Human Settlements Programme (2020). World Cities Report 2020: The Value Of Sustainable Urbanization. Nairobi: United Nations Human Settlements Programme.
Villéger, S., Mason, N. W. H., and Mouillot, D. (2008). New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 89, 2290–2301. doi: 10.1890/07-1206.1
Wenny, D. G., DeVault, T. L., Johnson, M. D., Kelly, D., Sekercioglu, C. H., Tomback, D. F., et al. (2011). Perspectives in ornithology the need to quantify ecosystem services provided by birds. Auk 128, 1–14. doi: 10.1525/auk.2011.10248
Wikiaves. (2020). Wikiaves. Available online at: www.wikiaves.com.br (accessed October 5, 2020).
Wilman, H., Belmaker, J., Simpson, J., de la Rosa, C., Rivadeneira, M. M., and Jetz, W. (2014). EltonTraits 1.0: species-level foraging attributes of the world’s birds and mammals. Ecology 95:2027. doi: 10.1890/13-1917.1
Young, A., Khalil, K. A., and Wharton, J. (2018). Empathy for animals: a review of the existing literature. Curator 61, 327–343. doi: 10.1111/cura.12257
Keywords: cultural services, cultural values, impervious surface, urban biodiversity, urban ecology
Citation: Valente-Neto F, Roque FO, Pauliquevis CF, Oliveira AKM, Provete DB, Szabo JK and Souza FL (2021) Loss of Cultural and Functional Diversity Associated With Birds Across the Urbanization Gradient in a Tropical City. Front. Ecol. Evol. 9:615797. doi: 10.3389/fevo.2021.615797
Received: 09 October 2020; Accepted: 03 May 2021;
Published: 28 May 2021.
Edited by:
Darryl Jones, Griffith University, AustraliaReviewed by:
Marco Moretti, Swiss Federal Institute for Forest, Snow and Landscape Research (WSL), SwitzerlandPiotr Tryjanowski, Poznań University of Life Sciences, Poland
Copyright © 2021 Valente-Neto, Roque, Pauliquevis, Oliveira, Provete, Szabo and Souza. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Francisco Valente-Neto, ZnZhbGVudGVuZXRvQGdtYWlsLmNvbQ==