 Balázs Deák1,2*
Balázs Deák1,2* Zoltán Rádai1
Zoltán Rádai1 Zoltán Bátori3
Zoltán Bátori3 András Kelemen1Katalin Lukács1,4
András Kelemen1Katalin Lukács1,4 Réka Kiss1István Elek Maák3,5
Réka Kiss1István Elek Maák3,5 Orsolya Valkó1
Orsolya Valkó1- 1Lendület Seed Ecology Research Group, Centre for Ecological Research, Institute of Ecology and Botany, Vácrátót, Hungary
- 2‘Lendület' Landscape and Conservation Ecology, Centre for Ecological Research, Institute of Ecology and Botany, Vácrátót, Hungary
- 3Department of Ecology, University of Szeged, Szeged, Hungary
- 4Juhász Nagy Pál Doctoral School, University of Debrecen, Debrecen, Hungary
- 5Museum and Institute of Zoology, Polish Academy of Sciences, Warsaw, Poland
Due to the intensified land use in transformed landscapes, grassland biodiversity is often restricted to habitat fragments inadequate for arable use or for urban development. In continental parts of Eurasia, the ~600,000 ancient burial mounds (called “kurgans”) built by nomadic tribes of the steppes are amongst the most widespread landmarks providing refuge for dry grassland species. In our study by using plant functional groups and functional traits, we aimed at gaining insight into the ecological and evolutionary processes shaping the structure and the composition of assemblages of grassland specialist plant species on kurgans embedded in the agricultural landscapes of East-Hungary. As a comparison, we also studied roadside verges and pristine extensive grasslands in the same region. We found that despite their small size, due to the lack of human disturbances and high microhabitat diversity kurgans can maintain a high species richness and percentage cover of specialists, especially when compared to verges. We revealed that assemblages of specialist plants on kurgans are characterized by traits typical to terrestrial habitat islands such as self-compatibility, large seed mass and tall stature. Kurgans and extensive grasslands were characterized by higher functional diversity (both at the level of single traits and multi-trait based functional dispersion) which is probably due to the higher level of environmental heterogeneity compared to the homogeneous environment in verges.
Introduction
Land use intensification leading to the loss and fragmentation of natural habitats is considered one of the major threats to biodiversity on local, regional and global scales. In intensively used landscapes, the remnants of formerly widespread natural habitats could persist within the few protected areas, and in small fragments surrounded by intensive croplands, forest plantations or urban areas (Auffret et al., 2015; Batáry et al., 2020; Deák et al., 2020a). This phenomenon is especially valid for dry grasslands often harboring extremely high herbaceous plant diversity and a high number of endemic species (Dengler et al., 2014). Due to the land-use changes in the past centuries, vast grassland areas with fertile soils were transformed into arable lands, while grasslands with less fertile soils were afforested or destroyed by the expansion of settlements and urban infrastructure (Biró et al., 2018). In such landscapes, remaining grassland fragments generally exist as “terrestrial islands” which, similarly to “true islands,” are often isolated from similar habitats by a matrix inadequate for the existence of dry grassland specialists (Deák et al., 2020b; Ottaviani et al., 2020).
In many landscapes, dry grassland fragments are represented in verges, field margins, rocky outcrops, or steep slopes of ravines that are unsuitable for agricultural production (Lindborg et al., 2014; Hunter et al., 2017; Fekete et al., 2019). Recently it has been realized that sites with sacred, historical and cultural values, such as old cemeteries, churchyards, shrines and sacred groves, can also act as important safe havens for biodiversity (Bhagwat and Rutte, 2006; Löki et al., 2019; Rath and Ormsby, 2020). These sites are often protected and managed by local people and less affected by negative land-use changes such as tilling or management intensification (Bhagwat and Rutte, 2006). In continental parts of Eurasia, ancient earthen burial mounds, the so-called “kurgans,” are one of the most widespread sacred natural sites harboring dry grassland vegetation (Deák et al., 2016). Nowadays, there are ~400–600,000 kurgans in Eurasia that were built by steppic cultures predominantly during the Bronze and Iron ages (Deák et al., 2016). Their typical height is 0.5–15 meters and their area is generally smaller than one hectare (Dembicz et al., 2020). Due to their cultural significance and the fact that their steep slopes hinder tilling, kurgans often preserve the last remnants of dry grassland vegetation and maintain a high diversity of grassland specialist species even in intensively used landscapes (Sudnik-Wójcikowska et al., 2011; Deák et al., 2020a,b; Dembicz et al., 2020).
Besides the relatively undisturbed conditions, the special dome shape of kurgans has the potential of maintaining high biodiversity by providing high environmental heterogeneity at very fine scales (Lisetskii et al., 2014; Deák et al., 2020c). The slopes with different aspects and the top of kurgans are characterized by different microclimatic and soil conditions, resulting in contrasting microhabitats (north-facing slope: cool and humid microclimates, low amount of heat load, and high soil moisture; top and south-facing slope: hot and dry microclimates, high amount of heat load and low soil moisture; east- and west-facing slopes are characterized by dynamic and large-scale daily temporal changes regarding microclimates) within the small area of a mound (Deák et al., 2020c). Like on inselbergs (Ottaviani et al., 2016; De Smedt et al., 2018; de Paula et al., 2019; Yates et al., 2019) and in topographic depressions such as dolines (Bátori et al., 2017, 2019, 2020a), high environmental heterogeneity on kurgans also supports the co-existence of several species with different environmental preferences, resulting in high species richness within a small area (Deák et al., 2020c). The special environmental conditions provided by the kurgan microhabitats also support the development of unique vegetation (Deák et al., 2017). The high biodiversity value of kurgans covered by dry grasslands has been demonstrated by Sudnik-Wójcikowska et al. (2011) and Deák et al. (2020a). For instance, Sudnik-Wójcikowska et al. (2011) found 721 plant species (including 71 species of special conservation interest) on 106 Ukrainian kurgans (total area: approximately 107 ha), which represent 14% of the whole Ukrainian flora. Similar results were obtained by Deák et al. (2020a), who surveyed 138 kurgans (total area: 38.9 ha) in Hungary and found 446 plant species (including 23 species of special conservation interest) in the kurgan microhabitats, which represent 17% of the whole Hungarian flora.
Due to the marked changes in the landscape composition and configuration of fertile lowland areas, kurgans often function as terrestrial habitat islands (Deák et al., 2016, 2020a; Dembicz et al., 2020; Kuli-Révész et al., 2021). The colonization and persistence of grassland specialists in grassland islands are considerably influenced by the special environmental characteristics (such as area, management, microclimate, soil fertility) and the degree of isolation providing several abiotic and biotic filters shaping the local species assemblages (Szabó et al., 2012; Heinken and Weber, 2013; Deák et al., 2018; Dembicz et al., 2020; Ottaviani et al., 2020). For revealing mechanisms affecting species occurrences and driving patterns of local community assembly in insular ecosystems and fragmented landscapes, trait-based approaches may provide a powerful tool (Janečková et al., 2017; Keppel et al., 2018).
Given the special environmental conditions within, and the high proportion of unfavorable matrix around terrestrial habitat islands, both the taxonomic and functional composition of their vegetation may differ from those of a similar habitat with a larger area and less affected by land-use changes (Ottaviani et al., 2020). Thus, plants in terrestrial habitat islands (such as in true islands) are likely characterized by unique functional trait values, trait combinations (trait syndromes) and functional diversity. As suggested by Ottaviani et al. (2020) for insular ecosystems, colonization is strongly affected by dispersal-related traits, while persistence is influenced by a combination of traits related to on-spot persistence, resource economy, space occupancy, stress- and disturbance avoidance, resistance and tolerance. Colonization of terrestrial habitat islands is supported by small seed mass which allows propagules to disperse farther than heavy seeds (Lindborg et al., 2012; Helsen et al., 2013). Possessing persistent seed bank, long life-span, ability for clonal growth and reproduction are traits that potentially increase the chance of species for persisting in isolated habitats (Auffret et al., 2015; Janečková et al., 2017). Due to the limited availability of pollinator species, autonomous plants may have an advantage in isolated habitats, because they can exist even in small populations of a few individuals (Ottaviani et al., 2020). Vegetation of small terrestrial habitat islands is also influenced by the consequences of lacking management and increasing competition. These factors might influence trait composition such as seed mass (heavy seeds support vigorous seedling growth; Bossuyt and Honnay, 2008), plant height (tall growth supports enhanced competition ability for light; Marini et al., 2012), and specific leaf area (decreased SLA in undisturbed, stable habitats; Auffret et al., 2015). As species developed a large variety of adaptations to a certain set of biotic and abiotic filters, both positive and negative correlations and trade-offs can be observed among the strategies. Understanding these patterns can provide deeper insights into the mechanisms shaping the species composition of terrestrial habitat islands (Götzenberger et al., 2012; Marini et al., 2012; Mouillot et al., 2013).
By using plant functional traits, we aim at gaining insights into the ecological and evolutionary processes shaping the composition of assemblages of dry grassland specialist species on kurgans embedded in agricultural landscapes in East-Hungary. For comparison, we also studied linear grassland fragments represented by road verges and extensive pristine grasslands in the same region. In the present study, we considered only dry grassland specialist species because, compared to generalists, these species are more sensitive to the composition of the adjacent landscape matrix (Auffret et al., 2015; Deák et al., 2018). For specialist species confined only to dry grassland habitats both non-grassland natural and anthropogenic habitats should act as an effective dispersal barrier (Chetcuti et al., 2020; Ottaviani et al., 2020). However, fragmented dry grasslands do not necessarily serve as habitat islands for generalists, because these species might establish in a wide variety of non-grassland habitats and therefore obscure the patterns related to insularity (Horsák et al., 2012).
We hypothesized that: (i) Assemblages of grassland specialist plants in secondary grassland fragments (kurgans, verges) are characterized by different cover of functional groups and different trait distributions compared to assemblages in extensive pristine grasslands; (ii) Functional traits of grassland specialist plants vary between isolated patch-like secondary grasslands (represented by kurgans) and less isolated linear secondary grasslands (represented by verges) differing in their main characteristics such as area, shape, degree of spatial and temporal isolation and management intensity. We believe that our study will contribute to a deeper understanding of how these safe havens function as habitats for grassland specialist species in agricultural landscapes.
Materials and Methods
Study Area
Our study area covering ~5,800 km2 is situated in the Hungarian Great Plain, which is a representative example of lowland agricultural landscapes in Europe. The area is characterized by a continental climate; the mean annual temperature is 10.4°C and the mean annual precipitation is 538 mm (Fick and Hijmans, 2017). Although in historical times the region was covered by extensive loess and alkaline grasslands and wetlands, due to the land-use changes since the eighteenth century a huge proportion of grasslands have been transformed into ploughlands and forest plantations (Biró et al., 2018). As a consequence of the severe habitat loss, these formerly continuous grasslands had been fragmented. Given their fertile chernozemic soils, loess grasslands (Salvio nemorosae—Festucetum rupicolae) were the most severely affected by land-use intensification, ~90% of their stands has been converted into ploughlands (Biró et al., 2018). In the modern agricultural landscape, remnants of loess grasslands generally occur in small fragments, such as in road verges and on kurgans (for photos of the studied habitats see Supplementary Figure 1). Despite the great reduction in their area, remaining loess grasslands still hold high biodiversity with several dry grassland specialist plant species (such as the red-listed Anchusa barrelieri, Ajuga laxmannii, and Carduus hamulosus). Due to their high conservation values, the Habitats Directive of the European Union (Council Directive 92/43/EEC) acknowledges loess grasslands of the Carpathian Basin as habitats of European importance (Pannonic loess steppic grasslands; code: 6250).
Pristine loess grasslands of the study area were formed during the Holocene, and function as the grassland components of the forest-steppe vegetation (Biró et al., 2018). The remaining extensive grasslands are managed by extensive grazing and they are occasionally mown (Table 1). Kurgans involved in our study represented isolated patch-like grasslands with a small area and a mean height of 5.1 m (SD = 2.0). As typical representatives of these kinds of habitats, all surveyed kurgans were isolated from other grasslands by habitats (e.g., ploughlands) inadequate for the establishment of grassland specialist species at least since the Second World War. Due to their isolated state, the management of grassland fragments on kurgans has been ceased for decades or centuries. As they have been built using the topsoil of the neighboring areas, their soil is rich in nutrients and has a good physical structure (Tóth et al., 2019). Verges represented grassland fragments with a linear shape. Given the fact that verges have a long linear shape stretching along the landscape, at some points, they may have direct contact with other grassland stands. They are affected by a high level of human disturbance such as trampling, mowing and application of herbicides. The selected verges were not sown or overseeded neither by native or alien plant species. During the selection of the study sites, we made an effort to select typical representatives of kurgans and verges regarding the level of isolation, habitat area and management (see Table 1). Given the surveyed sites had very similar attributes within each habitat type, we did not aim to analyze the separate effects of the above factors but focused on the general patterns typical to these habitats. Please note that this approach does not allow a separate evaluation of the particular effects of isolation and management, but aims to capture the general patterns typical to the studied habitats.
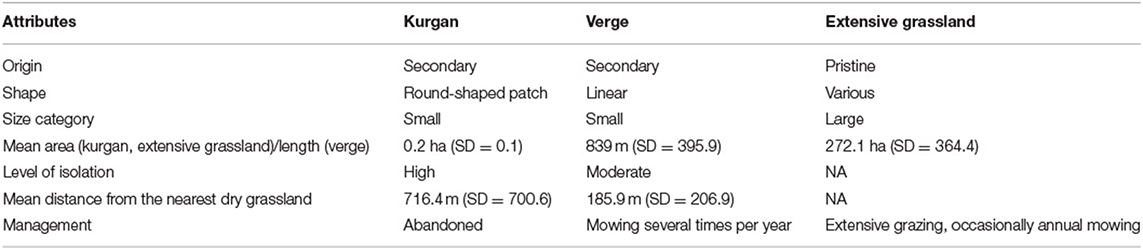
Table 1. Main attributes of the studied grassland habitats.
Field Sampling
We selected eight sites of each habitat type (i.e., kurgans, verges, and extensive grasslands) covered by loess grassland vegetation (in total 24 sites). At each site, we recorded the percentage cover of vascular plants in 25 plots of 1 m × 1 m in July 2019 (in total 600 plots: 3 habitat types × 8 sites × 25 plots), within an area of 0.1 ha. We chose 0.1 ha as a standard sample area as this was the size of the smallest kurgan. To obtain representative samples covering the microsite heterogeneity typical to kurgans (Deák et al., 2020c), we randomly placed five plots in each kurgan microhabitat (north-, east-, south- and west-facing slopes and the top). In the case of verges and extensive grasslands, we placed all plots randomly without any spatial preferences.
Functional Groups and Plant Traits
Functional traits (continuous variables such as start and length of flowering period, seed mass, plant height, and SLA) and functional groups (cover of wind-pollinated, insect-pollinated, and autonomous specialists, cover of specialists with a persistent seed bank, specialists with rosette leaves and perennial specialists) used in the analyses were selected to reveal whether functional characteristics of grassland specialist plants explain their occurrence in the studied grassland types, taking into account their ability for colonization (dispersal ability, reproduction strategy) and persistence (on-spot persistence, resource economy, space occupancy, stress- and disturbance avoidance, -resistance, and -tolerance). When available, trait data were collected from regional data sources and own trait measurements (Table 2). If regional data was not available, we used available databases (LEDA—Kleyer et al., 2008; CLO-PLA—Klimešová et al., 2017; Pladias – A Database of Czech Flora and Vegetation1) to complete the dataset. We used the mean of the indicated minimum and maximum values of plant height. If multiple records were available, we calculated the mean for specific leaf area (SLA). We considered the highest reported value for seed bank persistence. Based on the pollination type, we classified species into the following groups: (i) insect-pollinated, (ii) wind-pollinated and (iii) predominantly autonomous.
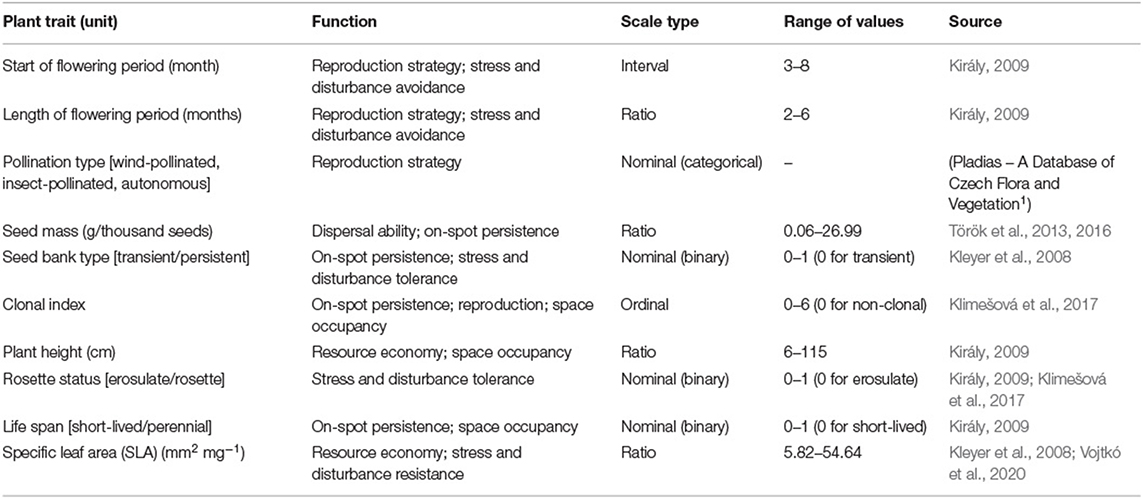
Table 2. Plant traits considered, their function, scale type and range, and the source of trait data.
Data Analyses
In this study, we focused on the species of the Festuco-Brometea phytosociological class that are confined exclusively to dry grasslands (called “specialists” hereafter) (Borhidi, 1995).
All statistical analyses were carried out in the R statistical environment (ver. 4.0.1; R Core Team, 2020).
We fitted a generalized linear mixed-effects model (GLMM) with Poisson error distribution (using the R-package “lme4”; Bates et al., 2015) on the number of specialists, and log-linked Gamma GLMM due to the strong skewness in the continuous data (with the R-package “glmmTMB”; Brooks et al., 2017) on the total cover of specialists. In these models, habitat type was the predictor variable. We also fitted log-linked Gamma GLMMs on the cover of wind-pollinated, insect-pollinated, and autonomous specialists and the cover of specialists with a persistent seed bank, specialists with rosette leaves and perennial specialists.
We calculated community-weighted means (CWMs) for numerical trait variables (start and length of the flowering period, seed mass, plant height, SLA, and clonal index) on the plot level (Ricotta and Moretti, 2011). The acquired variables were then used in log-linked Gamma GLMMs, in which CWMs were response variables, and habitat type was the predictor factor variable.
We estimated functional diversity as functional richness (FRic) and functional dispersion (FDis) with the “dbFD” function of the R-package “FD” (Laliberté and Legendre, 2010; Laliberté et al., 2014), which uses Euclidean distance matrix based on the species trait values. We both estimated single-trait diversity measures (using continuous traits), as well as multi-trait diversity (including both continuous traits and variables derived from functional groups). To test whether there are habitat level differences in the estimated functional diversity indices, we fitted log-linked Gamma GLMMs (as described above).
We estimated the mean species pairwise dissimilarity (MPD) of specialists per sampling plots based on de Bello et al. (2016), using the “taxondive” function of the R-package “vegan” (Oksanen et al., 2019). MPD expresses taxonomic diversity and distinctness as mean taxonomic distances among species. The advantage of MPD is that it is not influenced by species richness (de Bello et al., 2016). For the calculations, we used the mega-phylogeny of plants updated by Qian and Jin (2016). To see whether there are habitat-type differences in MPD scores, we used a Gaussian mixed-effect linear regression model (LMM), with MPD as a response variable and habitat type as a fixed predictor.
Notably, in Gamma GLMMs we incremented all values of the response variables by one, because this model type is not able to handle zeros in the data. In all mixed-effect models, we used sampling site ID as random factor to control for the non-independence in the sampling data. Furthermore, we used the “emmeans” function (“emmeans” package) for the acquisition of marginal mean estimates and sequential post-hoc comparisons among factor levels (Russell, 2019); Tukey HSD method was used to correct the P-values after performing multiple comparisons.
Results
We recorded a total of 55 specialist species in the study sites: 41 on kurgans, 27 in verges, and 33 in extensive grasslands (Supplementary Table 1). Plot-level species richness and cover of specialists were equally high on kurgans and in extensive grasslands, whilst specialist species were underrepresented in verges (Figure 1, Supplementary Table 2). We observed a similar pattern regarding the cover of insect- and wind-pollinated specialists (Figure 2, Supplementary Table 2). However, autonomous specialists were typical of kurgans. Possessing persistent seed bank was not typical in any of the studied habitat types. Kurgans and extensive grasslands were characterized by a high cover of perennial species. Rosette species were characteristic to extensive grasslands.
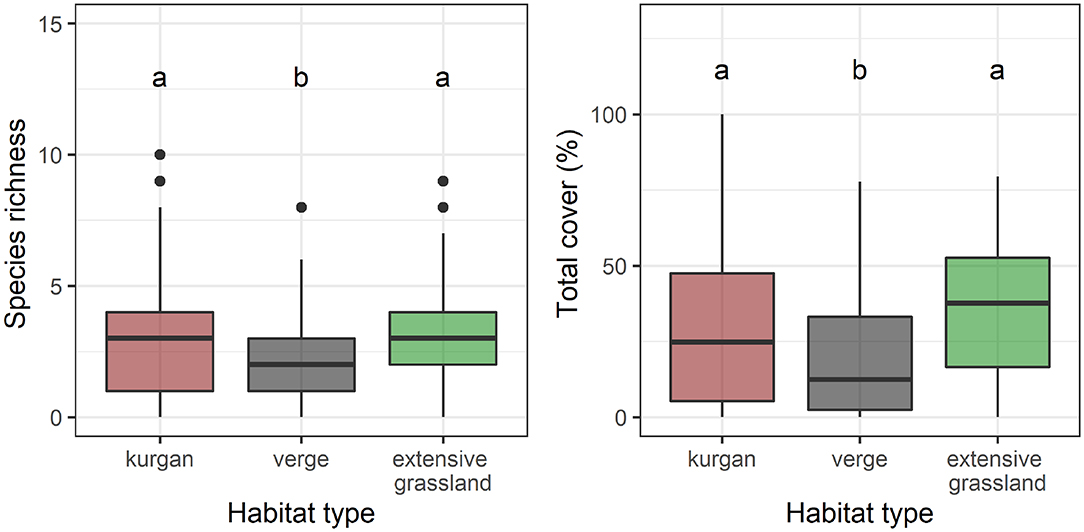
Figure 1. Species richness and total cover of specialists in the three studied habitat types (kurgans, verges, and extensive grasslands). Boxes, vertical whiskers, bold horizontal lines, and individual dots represent interquartile range (IQR: value range between 25 and 75% quartiles), minimum and maximum value range (excluding outliers), median and outliers (values being at least 1.5 times the IQR lower than the lower quartile, or 1.5 times IQR higher than the upper quartile), respectively. Different letters denote significant differences between groups (GLMM; p ≤ 0.05 after Tukey's test).
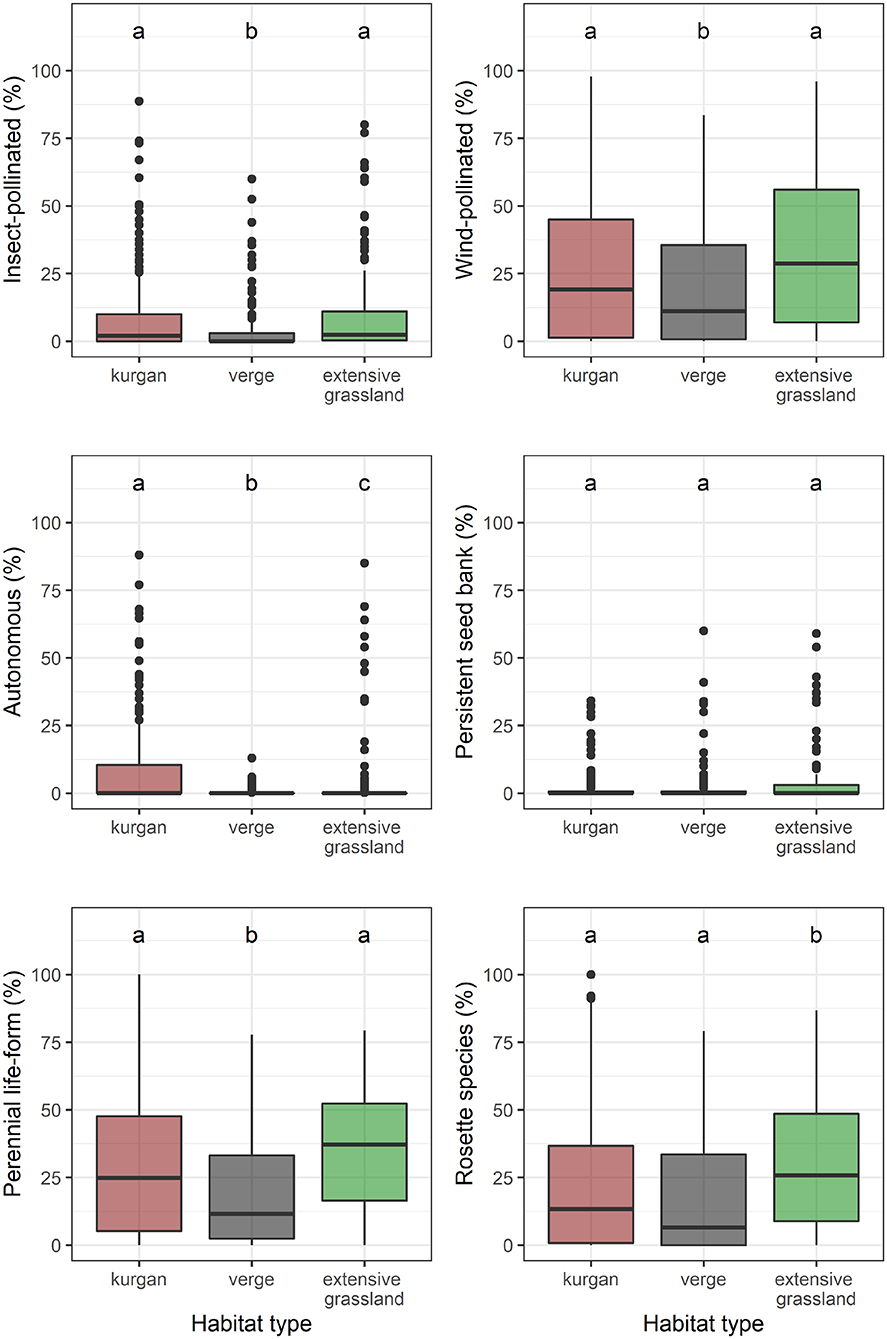
Figure 2. Percentage cover values of specialists belonging to different functional groups observed in the three studied habitat types (kurgans, verges, and extensive grasslands). Boxes, vertical whiskers, bold horizontal lines, and individual dots represent interquartile range (IQR: value range between 25 and 75% quartiles), minimum and maximum value range (excluding outliers), median, and outliers (values being at least 1.5 times the IQR lower than the lower quartile, or 1.5 times IQR higher than the upper quartile), respectively. Different letters denote significant differences between groups (GLMM; p ≤ 0.05 after Tukey's test).
There were no differences in the CWMs of the start and the length of flowering period across the three habitat types (Figure 3). Specialist species occurring on kurgans tended to produce heavier seeds shown by the larger CWM of seed mass compared to the other habitat types. Clonality was the most important in verges indicated by the highest CWMs of the clonal index in this habitat type. Plants on kurgans tended to be taller than in the other habitat types, whereas shorter plants were the most abundant in extensive grasslands. Specialist plants were characterized by smaller specific leaf area on kurgans than in the other two habitat types.
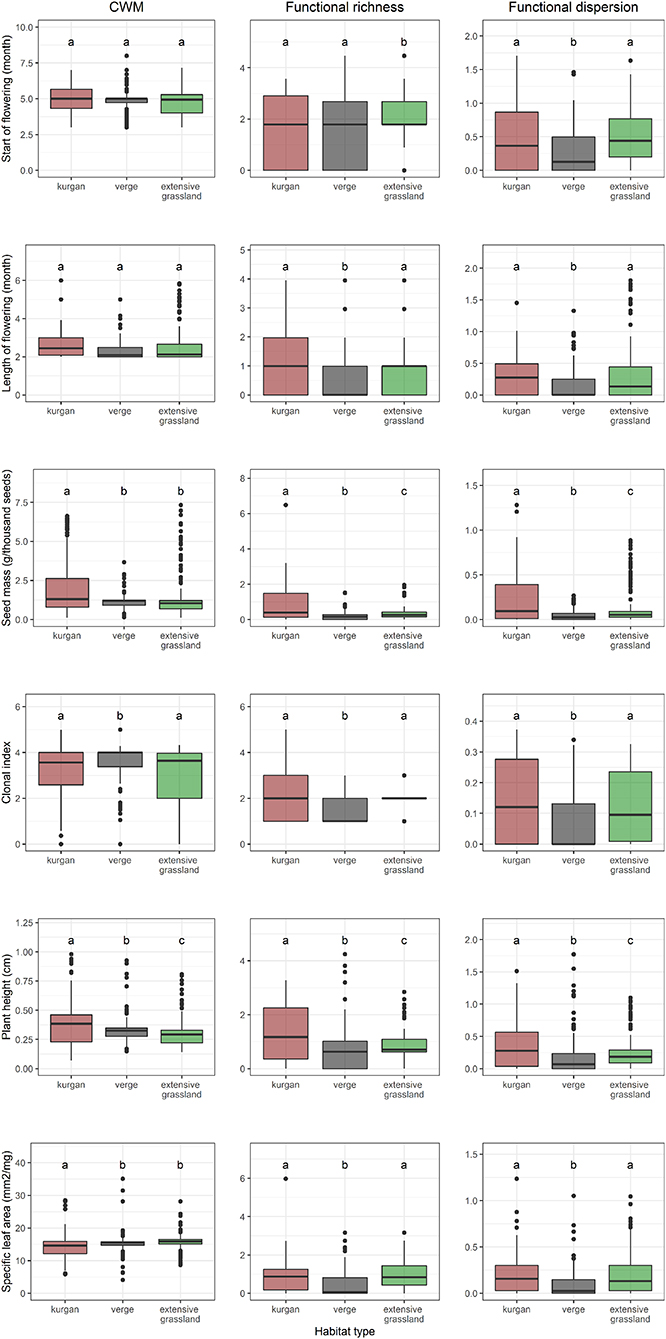
Figure 3. Distributions of community weighted mean (CWM), functional richness (FRic), and dispersion (FDis) of single traits of specialists in the three studied habitat types (kurgans, verges, and extensive grasslands). Boxes, vertical whiskers, bold horizontal lines, and individual dots represent interquartile range (IQR: value range between 25 and 75% quartiles), minimum and maximum value range (excluding outliers), median, and outliers (values being at least 1.5 times the IQR lower than the lower quartile, or 1.5 times IQR higher than the upper quartile), respectively. Different letters denote significant differences between groups (GLMM; p ≤ 0.05 after Tukey's test).
In all cases, functional diversity indices (FDis, FRic) calculated for single traits were lower in verges, while kurgans and grasslands were characterized by grassland specialist species with a higher variation in trait distributions. FDis calculated from multiple traits of specialist species was equally high on kurgans and in extensive grasslands (Figure 4, Supplementary Table 2). Phylogenetic diversity expressed by the MPD values was similar in the three habitat types (Figure 4).
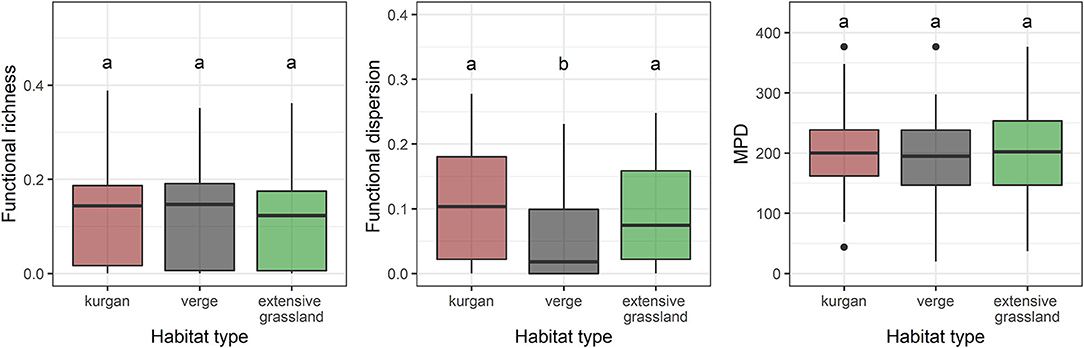
Figure 4. Multi-trait based functional richness (FRic), dispersion (FDis), and mean pairwise phylogenetic distance (MPD) of specialist plant assemblages in the three studied habitat types (kurgans, verges, and extensive grasslands). Boxes, vertical whiskers, bold horizontal lines, and individual dots represent interquartile range (IQR: value range between 25 and 75% quartiles), minimum and maximum value range (excluding outliers), median, and outliers (values being at least 1.5 times the IQR lower than the lower quartile, or 1.5 times IQR higher than the upper quartile), respectively. Different letters denote significant differences between groups (GLMM; p ≤ 0.05 after Tukey's test).
Discussion
Taxonomic and Functional Diversity Across the Habitat Types
Dry grassland specialists were equally diverse and abundant on kurgans as within extensively managed grasslands, suggesting that kurgans covered by dry grassland vegetation can have a high conservation importance in agricultural landscapes and may act as safe havens for a number of specialist species (see also in Deák et al., 2020b). Among the three studied habitats, verges representing highly disturbed linear grassland fragments held the lowest number and cover of specialists. Although the amount of functional space occupied by species assemblages (functional richness) was similar in the three studied habitat types, we found that functional dispersion was higher on kurgans and in extensive grasslands compared to verges. This result suggests that all studied habitat types were characterized by a wide range of plant trait values, and on the assemblage level the range of traits was not influenced by habitat specific abiotic and biotic factors (see also Mouillot et al., 2013). However, there were habitat specific factors that influenced the abundance-weighted distribution of the traits shown by the differences in functional dispersion. Kurgans and extensive grasslands were characterized by higher functional diversity (both at the level of single traits and multi-trait based functional dispersion) which is probably due to the higher level of environmental heterogeneity compared to the homogeneous environment in verges.
The high level of functional dispersion on kurgans might be attributed to the high environmental heterogeneity regarding soil moisture, microclimate, and soil nutrients (Deák et al., 2020c), which can support the co-existence of species with different trait combinations by niche partitioning (Zobel, 2015; Price et al., 2017). High functional dispersion on kurgans can have an important consequence for the long-term persistence of grasslands on these habitats, as high functional diversity may grant a high resilience for the community and by that, it supports an adaptive community-level response to changing environment (Keppel et al., 2018).
In extensive grasslands, the high functional dispersion values were likely maintained by the moderate level of cattle grazing that suppresses competitive species that otherwise would become dominant in homogenous environments, leading to competitive exclusion of disturbance-dependent species with a low competitive ability (Díaz et al., 2001). Furthermore, extensive grazing might contribute to the maintenance of a heterogeneous habitat structure due to a selective biomass removal and trampling, allowing the co-existence of specialists adapted to slightly different biotic and abiotic environments (Tälle et al., 2016; Deák et al., 2017). Although the disturbance of verges likely did not reach a level that would lead to the extinction of species with extreme trait values, the low functional dispersion in this habitat suggests that disturbance might act as a habitat filter, sorting out specialists with high abundances and extreme trait values (Zobel, 2015). In contrast to extensive grazing, frequent mowing usually provides a uniform biomass removal by which it can homogenize the habitat (Tälle et al., 2016).
Plant Traits of Specialist Species Across the Habitat Types
The high cover of autonomous specialists on kurgans suggests that these habitats can act as terrestrial habitat islands as previous studies found autonomous reproduction was characteristic to islands (Grossenbacher et al., 2017; Ottaviani et al., 2020). The high proportion of autonomous species on oceanic islands was first proposed by Baker (known as Baker's law; Baker, 1955), who argued that on islands species with uniparental reproduction have a considerable colonization advantage. As colonization is often achieved by only a few individuals, finding mates can be challenging in the early phase of establishment due to the small population size. In such a situation establishment success is higher for autonomous species (Ottaviani et al., 2020). In terrestrial islands, autonomous species are less dependent on incoming pollen sources, which makes them less dependent on the composition of the adjacent landscapes. As a result, they can sustain small populations for a long period (Heinken and Weber, 2013). A consequence of this strategy is that the start and length of the flowering period are largely independent of the availability of pollinator agents.
The cover of insect-pollinated specialists was similarly high on the kurgan and in the extensive grassland habitats, suggesting that both habitats can provide more stable resources for pollinators compared to verges. This pattern suggests that these extensive grasslands can maintain stable and abundant populations of various pollinator species that support the long-term survival of many plants relying on insect pollination. Although the cover of insect-pollinated specialists was also high on kurgans, in this habitat many of the specialists can be optionally pollinated either by insects or in an autonomous way. This possibility might be especially advantageous in transformed landscapes where food resources for pollinators are limited both in quality and in quantity resulting in reduced species richness and abundance of pollinators at a landscape-level, and a reduced period when they are available (Geppert et al., 2020).
Although isolated habitats are expected to hold species with a short flowering period due to the lack of nearby pollen sources and the absence of pollinator vectors (Janečková et al., 2017), we did not observe this pattern. It is possible that the high environmental heterogeneity typical to kurgans can counterbalance the effect of isolation. As was found by Olliff-Yang and Ackerly (2020), complementarity between microhabitats with different aspects can extend the flowering times by intra-specific variations and species turnover even at small spatial scales. On the studied kurgans environmental heterogeneity (Deák et al., 2020c) might override the effect of isolation regarding flowering phenology. As suggested by the high functional richness and dispersion values of flowering length, high environmental heterogeneity might result in a species turnover of specialists with a short and long flowering length among the kurgan microhabitats.
Although seeds preserving germination ability for an extended period may act as an effective buffer against extinction in fragmented habitats (Marini et al., 2012; Auffret et al., 2015), cover of specialists with a persistent seed bank was similarly rare in the studied habitat types. The reason for this might be that dry grasslands in the temperate forest-steppe zone can be considered as long-term stable habitats, where specialists do not invest resources in producing long-lived seeds (Bossuyt and Honnay, 2008; Kiss et al., 2016). The lack of persistent seed bank can be especially problematic on kurgans because they are generally isolated from other grassland stands. In these habitats, following a large scale stochastic event (such as tilling or the use of chemicals within agricultural landscapes) re-establishment of specialists is hindered both by the sparse persistent seed bank and the reduced amount of incoming seed rain from the neighboring areas (Deák et al., 2018).
Previous studies suggested that producing light seeds might buffer the effects of isolation and support the maintenance of metapopulation connections in fragmented landscapes as light seeds can disperse to larger distances than heavy ones (Lindborg et al., 2012; Helsen et al., 2013; Janečková et al., 2017). On the contrary, we observed that kurgans were characterized by larger-seeded specialists than verges or extensive grasslands. This result is in line with the findings of Rossetto and Kooyman (2005), Kooyman et al. (2011), and Auffret et al. (2015) who suggested that a large reduction in functional connectivity among fragmented populations might lead to a suppression of traits related to long-distance dispersal. In highly fragmented landscapes, propagules with good dispersal ability likely leave the original habitat patch and have a high chance to land in an inadequate habitat unsuitable for long-term establishment (Auffret et al., 2015). Thus, good dispersal ability supported by light seeds contributes neither to the maintenance of the local populations nor the establishment of metapopulation connections (Deák et al., 2018). Heavy seeds with a low potential for long-distance dispersal, generally remain on-spot supporting the long-term persistence of the local population that might be a beneficial strategy in isolated grasslands (Helsen et al., 2013).
High proportion of heavy seeded species on kurgans might also be a consequence of abandonment resulting in litter accumulation (Lindborg et al., 2014). Accumulated thick litter layer can hinder establishment processes by lighter seeds acting as a physical barrier, preventing them to land on the soil surface, while heavier seeds have a higher chance to penetrate through the thick litter layer (Janečková et al., 2017). Furthermore, larger nutrient storage of heavy seeds supports seedlings to outcompete seedlings of lighter-seeded species (Bossuyt and Honnay, 2008; Janečková et al., 2017). As was pointed out by Keppel et al. (2018) for island-like refugia, and by Rossetto and Kooyman (2005) and Kooyman et al. (2011) in Australian rainforest refugia, producing heavy seeds might also be an outcome of long-term stability, which in our study was provided by the lack of management on kurgans. Deák et al. (2020c) also found that there is a climatic buffering on kurgans due to the co-occurrence of several microhabitats (such as slopes with different aspects on kurgans) with different climatic parameters. Although our sampling setup does not allow testing the assumption, but it is possible that the co-existence of harsh and mild microhabitats could lead to large values of functional richness and dispersion for seed mass on kurgans: large-seeded specialists are probably more abundant in the harsh microhabitats because of the larger storing effect linked to larger seeds, and the small-seeded specialists are typical in the mild microhabitats (Leishman and Westoby, 1994).
In contrast to kurgans, producing light seeds seemed to be a good strategy in extensive grasslands and verges characterized by larger spatial dimensions and also a high number of dynamically forming establishment gaps (e.g., open surfaces created by grazing or mowing) (Tälle et al., 2016). In these habitats good dispersal ability promoted by light seeds supports specialists to overarch moderate spatial distances within the grassland patch, and to occupy newly formed gaps appropriate for seedling emergence of light-seeded specialists (Kahmen and Poschlod, 2008; Kiss et al., 2020). Heterogeneity of the habitat might be responsible for the high functional richness and dispersion of seed mass values in extensive grasslands, where ungrazed patches characterized by higher biomass and litter amount favor heavy-seeded specialists and highly grazed and trampled patches favor the establishment of light-seeded specialists (Kahmen and Poschlod, 2008).
We found that the cover of perennial specialists was high on kurgans likely contributing to the long-term maintenance of grassland vegetation despite the high proportion of those habitats in the adjacent landscape that are inadequate for the establishment for grassland specialist species. As long-lived species are less affected by short-term environmental changes and small-scale disturbances, they experience smaller fluctuations in their abundance compared to short-lived species, which might provide higher metapopulation stability (Marini et al., 2012; Heinken and Weber, 2013).
Although clonal growth providing an alternative life cycle loop (without pollination, seed production and seedling establishment) might be an advantageous strategy in long-term isolated habitats, we found that high CWM of clonal plants was typical only to less isolated verges. This result is in line with the findings of Lindborg et al. (2014), who found that in agricultural landscapes consisting of midfield islands, verges and large grassland stands, verges harbored the specialist species with the most clonal character. This pattern is likely due to the frequent mowing management and trampling typical to verges, both of which were proven to enhance clonal plants (Honnay and Bossuyt, 2005). Furthermore, due to the high level of disturbance, verges hold a higher cover of disturbance-tolerant generalist species (Deák et al., 2020b); thus, specialists that are able to spread or reproduce clonally have a higher chance to successfully compete with them. Despite the CWM of clonality was low on kurgans and in extensive grasslands, functional richness and dispersion of this trait were the highest in these habitats, likely due to the heterogeneous habitat structure where clonal plants are restricted only to certain microhabitats and suppressed from others.
We also found that kurgans hold the tallest specialist species. In abandoned and isolated areas, such as the surveyed kurgans, vertical growth supports species to cope with the accumulated thick litter layer that suppresses the growth and shade of shorter plant individuals (Janečková et al., 2017). Furthermore, an increased litter layer induces an increased competition for light, in which tall plants perform better than short ones (Marini et al., 2012). Enhanced growth in height can also be the result of the fertile soil conditions typical to kurgans providing high nutrient amounts for plant growth (Lisetskii et al., 2014). We found that CWM of plant height was smaller in extensive grasslands. The likely reason for this is that grazing benefits smaller species (Díaz et al., 2001) due to the preferential grazing of tall plants with larger biomass, and also due to the decreased competition for light by opening up the vegetation structure (Janečková et al., 2017). In extensive grasslands, grazing benefited specialists with rosette leaves that mitigate biomass removal by grazers (Janečková et al., 2017).
Management targeting biomass removal such as grazing (Pakeman, 2013) and mowing (Lindborg et al., 2014—verges; Bátori et al., 2020b—river embankments) was proven to considerably increase SLA (i.e., species with soft and large leaves) of the community by promoting disturbance-tolerant fast-growing species. These species can occupy newly formed open gaps and able to grow new leaves by which they can optimize light capture for photosynthesis shortly after losing biomass. Our results derived from the extensive grasslands and verges confirm these patterns. We found low SLA values on kurgans, which might be due to abandonment that can considerably decrease SLA by the exclusion of fast-growing disturbance-dependent species (Auffret et al., 2015; Janečková et al., 2017). Low SLA can also be attributed to the dry environmental conditions on kurgans. Similarly to midfield islets (Lindborg et al., 2014), inselbergs (Ottaviani et al., 2016; De Smedt et al., 2018; de Paula et al., 2019; Yates et al., 2019), and karst hillocks (Filibeck et al., 2020), kurgans provide relatively dry habitat conditions due to their elevated position and intensive water run-off on the slopes (Deák et al., 2020c). Like in midfield islets and inselbergs, dry environmental conditions led to a low CWM of SLA values because smaller SLA decreases the chance for water to be lost by the plant (Pakeman, 2013; Conti et al., 2017). Like inselbergs, kurgans also hold contrasting harsh (dry and warm south-facing slopes and top; similar to “monocot mats” on inselbergs characterized by low SLA values) and mild (north-facing slopes similar to “shallow depressions” on inselbergs characterized by high SLA values) microhabitats (Deák et al., 2017, 2020c). Although in this study we did not aim to compare differences in plant traits among these microhabitats, high functional richness and dispersion values suggest that the high environmental variability on kurgans results in a high variability of SLA values, which are likely lower in harsh microhabitats and higher in mild microhabitats.
Interestingly, we did not find any differences in the phylogenetic diversity of the three studied habitat types. The lack of differences might be attributed to the applied phylogenetic diversity measure (MPD) that predominantly reflects large evolutionary differences (e.g., within families; Tucker et al., 2017). Abiotic and biotic filters typical to the study area and its habitats might influence species richness and functional diversity, but their magnitude is not large enough to affect the deeper levels of the phylogenetic tree. These results suggest that despite their relatively small size, habitat fragments embedded in agricultural landscapes are able to preserve a considerable amount of the evolutionary history typical to the landscape.
Conclusions
Our study revealed that kurgans covered by grassland vegetation may act as important safe havens for dry grassland specialist plants in transformed landscapes. The undisturbed status, elevated structure and high microhabitat diversity typical to kurgans support the existence of a high number of specialists. These species are characterized by specific traits characteristic of islands in general, such as large seed mass, tall growth and autonomous reproduction which are less typical to other grassland habitats (i.e., extensive grasslands and verges) in the landscape. Despite their small area, kurgans hold a high environmental heterogeneity, resulting in large variability in traits related to reproduction strategy, stress and disturbance avoidance and tolerance, dispersal ability, on-spot persistence, resource economy and space occupancy. Although these differences were not present on the level of phylogenetic diversity, we found that both kurgans and verges can preserve a considerable amount of the evolutionary history. Additional studies aiming to assess whether species turnover among habitat patches affects the taxonomic, functional and phylogenetic beta diversity would be highly desirable.
Our results suggest that besides acting as safe havens for dry grassland specialist plants, kurgans might also be considered as important microrefugia for many grassland species in the future. They provide high environmental heterogeneity with cooler/moister and warmer/drier microhabitats that might allow species to locally track shifting climates over time with minimal spatial movement. Rearrangement of species within kurgans is also supported by the high variability of traits supporting community-level adaptive responses for changing environmental conditions. Taking all these into consideration, we stress that besides the remaining extensive grasslands and verges, kurgans covered by grassland vegetation can be important sites for conservation in Eurasian agricultural landscapes.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
BD and OV contributed to the study conception and design. Data preparation and data collection was performed by BD, OV, AK, KL, and RK. Data analyses were performed by ZR. The first draft of the manuscript was written by BD and OV. All authors commented on previous versions of the manuscript and read and approved the final manuscript.
Funding
This study was supported by the NKFI KH 130338, NKFI FK 135329, NKFI FK 124404, and NKFI KH 126476 grants. OV, AK, and BD were supported by the Bolyai János Scholarship of the Hungarian Academy of Sciences. ZB was supported by the NKFI K 124796 grant. RK was supported by the NTP-NFTÖ-20-B-0093 grant. BD was supported by the Hungarian National Research, Development and Innovation Office (NKFIH KKP 133839).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2021.619812/full#supplementary-material
Footnote
1. ^Pladias – Database of the Czech Flora and Vegetation. Available online at: www.pladias.cz
References
Auffret, A. G., Plue, J., and Cousins, S. A. O. (2015). The spatial and temporal components of functional connectivity in fragmented landscapes. Ambio 44, 51–59. doi: 10.1007/s13280-014-0588-6
Baker, H. G. (1955). Self-compatibility and establishment after long-distance dispersal. Evolution 9, 347–349. doi: 10.1111/j.1558-5646.1955.tb01544.x
Batáry, P., Báldi, A., Ekroos, J., Gallé, R., Grass, I., and Tscharntke, T. (2020). Biol. Futura: landscape perspectives on farmland biodiversity conservation. Biol. Futura 71, 9–18. doi: 10.1007/s42977-020-00015-7
Bates, D., Mächler, M., Bolker, B., and Walker, S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Soft. 67, 1–48. doi: 10.18637/jss.v067.i01
Bátori, Z., Kiss, P. J., Tölgyesi, C., Deák, B., Valkó, O., Török, P., et al. (2020b). River embankments mitigate the loss of grassland biodiversity in agricultural landscapes. River Res. Appl. 36, 1160–1170. doi: 10.1002/rra.3643
Bátori, Z., Lőrinczi, G., Tölgyesi, C., Módra, G., Juhász, O., Aguilon, D. J., et al. (2020a). Karstic microrefugia host functionally specific ant assemblages. Front. Ecol. Evol. 8:613738. doi: 10.3389/fevo.2020.613738
Bátori, Z., Vojtkó, A., Farkas, T., Szabó, A., Havadtői, K., Vojtkó, E.-A., et al. (2017). Large- and small-scale environmental factors drive distributions of cool-adapted plants in karstic microrefugia. Ann. Bot.-London. 119, 301–309. doi: 10.1093/aob/mcw233
Bátori, Z., Vojtkó, A., Maák, I. E., Lörinczi, G., Farkas, T., Kántor, N., et al. (2019). Karst dolines provide diverse microhabitats for different functional groups in multiple phyla. Sci. Rep. 9:7176. doi: 10.1038/s41598-019-43603-x
Bhagwat, S. A., and Rutte, C. (2006). Sacred groves: potential for biodiversity management. Front. Ecol. Environ. 4, 519–524. doi: 10.1890/1540-9295(2006)4[519:SGPFBM]2.0.CO;2
Biró, M., Bölöni, J., and Molnár, Z. (2018). Use of long-term data to evaluate loss and endangerment status of Natura 2000 habitats and effects of protected areas. Conserv. Biol. 3, 660–671. doi: 10.1111/cobi.13038
Borhidi, A. (1995). Social behaviour types, the naturalness and relative ecological indicator values of the higher plants in the Hungarian Flora. Acta Bot. Hung. 39, 97–181.
Bossuyt, B., and Honnay, O. (2008). Can the seed bank be used for ecological restoration? An overview of seed bank characteristics in European communities. J. Veg. Sci. 19, 875–884. doi: 10.3170/2008-8-18462
Brooks, M. E., Kristensen, K., van Benthem, K. J., Magnusson, A., Berg, C. W., Nielsen, A., et al. (2017). GlmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 9, 378–400. doi: 10.32614/RJ-2017-066
Chetcuti, J., Kunin, W. E., and Bullock, J. M. (2020). Habitat fragmentation increases overall richness, but not of habitat-dependent species. Front. Ecol. Evol. 8:607619. doi: 10.3389/fevo.2020.607619
Conti, L., de Bello, F., Lepš, J., Acosta, A. T. R., and Carboni, M. (2017). Environmental gradients and micro-heterogeneity shape fine-scale plant community assembly on coastal dunes. J. Veg. Sci. 28, 762–773. doi: 10.1111/jvs.12533
Council Directive 92/43/EEC (1992). Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off. J. L 206, 7–50.
de Bello, F., Carmona, C. P., Lepš, J., Szava-Kovats, R., and Pärtel, M. (2016). Functional diversity through the mean trait dissimilarity: resolving shortcomings with existing paradigms and algorithms. Oecologia 180, 933–940. doi: 10.1007/s00442-016-3546-0
de Paula, L. F. A., Kolb, R. M., Porembski, S., Silveira, F. A. O., and Rossatto, D. R. (2019). Rocks and leaves: can anatomical leaf traits reflect environmental heterogeneity in inselberg vegetation? Flora 250, 91–98. doi: 10.1016/j.flora.2018.11.020
De Smedt, P., Ottaviani, G., Wardell-Johnson, G., Sýkora, K. V., and Mucina, L. (2018). Habitat heterogeneity promotes intraspecific trait variability of shrub species in Australian granite inselbergs. Folia Geobot. 53, 133–145. doi: 10.1007/s12224-018-9311-x
Deák, B., Kovács, B., Rádai, Z., Apostolova, I., Kelemen, A., Kiss, R., et al. (2020c). Linking environmental heterogeneity and plant diversity: the ecological role of small natural features in homogeneous landscapes. Sci. Tot. Environ. 763:144199. doi: 10.1016/j.scitotenv.2020.144199
Deák, B., Rádai, Z., Lukács, K., Kelemen, A., Kiss, R., Bátori, Z., et al. (2020b). Fragmented dry grasslands preserve unique components of plant species and phylogenetic diversity in agricultural landscapes. Biodivers. Conserv. 29, 4091–4110. doi: 10.1007/s10531-020-02066-7
Deák, B., Tölgyesi, C., Kelemen, A., Bátori, Z., Gallé, R., Bragina, T. M., et al. (2017). Vegetation of steppic cultural heritage sites in Kazakhstan – effects of micro-habitats and grazing intensity. Plant. Ecol. Divers. 10, 509–520. doi: 10.1080/17550874.2018.1430871
Deák, B., Tóthmérész, B., Valkó, O., Sudnik-Wójcikowska, B., Moysienko, I., Bragina, T. M., et al. (2016). Cultural monuments and nature conservation: the role of kurgans in maintaining steppe vegetation. Biodivers. Conserv. 25, 2473–2490. doi: 10.1007/s10531-016-1081-2
Deák, B., Valkó, O., Nagy, D. D., Török, P., Torma, A., Gábor, L., et al. (2020a). Habitat islands outside nature reserves – threatened biodiversity hotspots of grassland specialist plant and arthropod species. Biol. Conserv. 241:108254. doi: 10.1016/j.biocon.2019.108254
Deák, B., Valkó, O., Török, P., Kelemen, A., Bede, Á., Csathó, A. I., et al. (2018). Landscape and habitat and filters jointly drive richness and abundance of grassland specialist plants in terrestrial habitat islands. Landscape Ecol. 33, 1117–1132. doi: 10.1007/s10980-018-0660-x
Dembicz, I., Moysiyenko, I., Kozub, Ł., Dengler, J., Zakharova, M., and Sudnik-Wójcikowska, B. (2020). Steppe islands in a sea of fields: where island biogeography meets the reality of a severely transformed landscape. J. Veg. Sci. e12930. doi: 10.1111/jvs.12930
Dengler, J., Janišová, M., Török, P., and Wellstein, C. (2014). Biodiversity of Palearctic grasslands: a synthesis. Agric. Ecosys. Environ. 182, 1–14. doi: 10.1016/j.agee.2013.12.015
Díaz, S., Noy-Meir, I., and Cabido, M. (2001). Can grazing response of herbaceous plants be predicted from simple vegetative traits? J. Appl. Ecol. 38, 497–508. doi: 10.1046/j.1365-2664.2001.00635.x
Fekete, R., Löki, V., Urgyán, R., Süveges, K., Lovas-Kiss, Á., Vincze, O., et al. (2019). Roadside verges and cemeteries: comparative analysis of anthropogenic orchid habitats in the Eastern Mediterranean. Ecol. Evol. 9, 6655–6664. doi: 10.1002/ece3.5245
Fick, S. E., and Hijmans, R. J. (2017). Worldclim 2: new 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315. doi: 10.1002/joc.5086
Filibeck, G., Sperandii, M. G., Bragazza, L., Bricca, A., Chelli, S., Maccherini, S., et al. (2020). Competitive dominance mediates the effects of topography on plant richness in a mountain grassland. Basic Appl. Ecol. 48, 112–123. doi: 10.1016/j.baae.2020.09.008
Geppert, C., Hass, A., Földesi, R., Donkó, B., Akter, A., Tscharntke, T., et al. (2020). Agri-environment schemes enhance pollinator richness and abundance but bumblebee reproduction depends on field size. J. Appl. Ecol. 57, 1818–1828. doi: 10.1111/1365-2664.13682
Götzenberger, L., de Bello, F., Bråthen, K. A., Davison, J., Dubuis, A., Guisan, A., et al. (2012). Ecological assembly rules in plant communities – approaches, patterns and prospects. Biol. Rev. 87, 111–127. doi: 10.1111/j.1469-185X.2011.00187.x
Grossenbacher, D. L., Brandvain, Y., Auld, J. R., Burd, M., Cheptou, P.-O., and Conner, J. K. (2017). Self-compatibility is overrepresented on islands. New. Phyt. 215, 469–478. doi: 10.1111/nph.14534
Heinken, T., and Weber, E. (2013). Consequences of habitat fragmentation for plant species: do we know enough? Perspect. Plant. Ecol. Evol. Syst. 15, 205–216. doi: 10.1016/j.ppees.2013.05.003
Helsen, K., Hermy, M., and Honnay, O. (2013). Spatial isolation slows down directional plant functional group assembly in restored semi-natural grasslands. J. Appl. Ecol. 50, 404–413. doi: 10.1111/1365-2664.12037
Honnay, O., and Bossuyt, B. (2005). Prolonged clonal growth: escape route or route to extinction? Oikos 108, 427–432. doi: 10.1111/j.0030-1299.2005.13569.x
Horsák, M., Hájek, M., Spitale, D., Hájková, P., Dítě, D., and Nekola, J. C. (2012). The age of island-like habitats impacts habitat specialist species richness. Ecology 93, 1106–1114. doi: 10.1890/0012-9658-93.5.1106
Hunter, M. L., Acuña, V., Bauer, D. M., Bell, K. P., Calhoun, A. J. K., Felipe-Lucia, M. R., et al. (2017). Conserving small natural features with large ecological roles: a synthetic overview. Biol. Conserv. 211, 88–95. doi: 10.1016/j.biocon.2016.12.020
Janečková, P., Janeček, S., Klimešová, J., Götzenberger, L., Horník, J., Lepš, J., et al. (2017). The plant functional traits that explain species occurrence across fragmented grasslands differ according to patch management, isolation, and wetness. Landscape Ecol. 32, 791–805. doi: 10.1007/s10980-017-0486-y
Kahmen, S., and Poschlod, P. (2008). Effects of grassland management on plant functional trait composition. Agric. Ecosyst. Environ. 128, 137–145. doi: 10.1016/j.agee.2008.05.016
Keppel, G., Ottaviani, G., Harrison, S., Wardell-Johnson, G. W., Marcantonio, M., and Mucina, L. (2018). Towards an eco-evolutionary understanding of endemism hotspots and refugia. Ann. Bot. 122, 927–934. doi: 10.1093/aob/mcy173
Király, G. (2009). Új magyar Füvészkönyv. Magyarország Hajtásos Növényei (New Hungarian Herbal. The Vascular Plants of Hungary. Identification Key) [in Hungarian]. Jósvafő: Aggtelek National Park Directorate.
Kiss, R., Deák, B., Tóthmérész, B., Miglécz, T., Tóth, K., Török, P., et al. (2020). Establishment gaps in species-poor grasslands: artificial biodiversity hotspots to support the colonization of target species. Rest. Ecol. doi: 10.1111/rec.13135
Kiss, R., Valkó, O., Tóthmérész, B., and Török, P. (2016). “Seed bank research in Central-European grasslands - an overview,” in Seed Banks: Types, Roles and Research, ed J. Murphy (New York, NY: Nova Science Publishers), 1–34.
Kleyer, M., Bekker, R. M., Knevel, I. C., Bakker, J. P., Thompson, K., Sonnenschein, M., et al. (2008). The LEDA Traitbase: a database of life-history traits of Northwest European flora. J. Ecol. 96, 1266–1274. doi: 10.1111/j.1365-2745.2008.01430.x
Klimešová, J., Danihelka, J., Chrtek, J., de Bello, F., and Herben, T. (2017). CLO-PLA: a database of clonal and bud bank traits of Central European flora. Ecology 98:1179. doi: 10.1002/ecy.1745
Kooyman, R., Rossetto, M., Cornwell, W., and Westoby, M. (2011). Phylogenetic tests of community assembly across regional to continental scales in tropical and subtropical rain forests. Global Ecol. Biogeogr. 20, 707–716. doi: 10.1111/j.1466-8238.2010.00641.x
Kuli-Révész, K., Korányi, D., Lakatos, T., Szabó, Á. R., Batáry, P., and Gallé, R. (2021). Smaller and isolated grassland fragments are exposed to stronger seed and insect predation in habitat edges. Forests 12:54. doi: 10.3390/f12010054
Laliberté, E., and Legendre, P. (2010). A distance-based framework for measuring functional diversity from multiple traits. Ecology 91, 299–305. doi: 10.1890/08-2244.1
Laliberté, E., Legendre, P., Shipley, B., and Laliberté, M. E. (2014). Package ‘FD'. Measuring Functional Diversity From Multiple Traits, and Other Tools for Functional Ecology.
Leishman, M., and Westoby, M. (1994). The role of seed size in seedling establishment in dry soil conditions - experimental evidence from semi-arid species. J. Ecol. 82, 249–258. doi: 10.2307/2261293
Lindborg, R., Helm, A., Bommarco, R., Heikkinen, R. K., Kühn, I., Pykälä, J., et al. (2012). Fragmentation effects on plant trait distribution in European forests and grassland. Ecography 35, 356–363. doi: 10.1111/j.1600-0587.2011.07286.x
Lindborg, R., Plue, J., Andersson, K., and Cousins, S. A. O. (2014). Function of small habitat elements for enhancing plant diversity in different agricultural landscapes. Biol. Conserv. 169, 206–213. doi: 10.1016/j.biocon.2013.11.015
Lisetskii, F. N., Goleusov, V. P., Moysiyenko, I. I., and Sudnik-Wójcikowska, B. (2014). Microzonal distribution of soils and plants along the catenas of mound structures. Contemp. Probl. Ecol. 7, 282–293. doi: 10.1134/S1995425514030111
Löki, V., Molnár, V. A., Süveges, K., Heimer, H., Takács, A., Nagy, T., et al. (2019). Predictors of conservation value of Turkish cemeteries: a case study using orchids. Landscape Urban Plan. 186, 36–44. doi: 10.1016/j.landurbplan.2019.02.016
Marini, M., Bruun, H. H., Heikkinen, R. K., Helm, A., Honnay, O., Krauss, J., et al. (2012). Traits related to species persistence and dispersal explain changes in plant communities subjected to habitat loss. Divers. Distrib. 18, 898–908. doi: 10.1111/j.1472-4642.2012.00893.x
Mouillot, D., Graham, N. A. J., Villéger, S., Mason, N. W. H., and Bellwood, D. R. (2013). A functional approach reveals community responses to disturbances. Trends. Ecol. Evol. 28, 167–177. doi: 10.1016/j.tree.2012.10.004
Oksanen, J., Blanchet, F. G., Friendly, M., Kindt, R., Legendre, P., McGlinn, D., et al. (2019). Vegan: Community Ecology Package. R package version 2.5-6. Available online at: https://CRAN.R-project.org/package=vegan (accessed January 11, 2021).
Olliff-Yang, R. L., and Ackerly, D. D. (2020). Topographic heterogeneity lengthens the duration of pollinator resources. Ecol. Evol. 10, 9301–9312. doi: 10.1002/ece3.6617
Ottaviani, G., Keppel, G., Götzenberger, L., Harrison, S., Opedal, Ø. H., Conti, L., et al. (2020). Linking plant functional ecology to island biogeography. Trends. Plant. Sci. 25, 329–339. doi: 10.1016/j.tplants.2019.12.022
Ottaviani, G., Marcantonio, M., and Mucina, L. (2016). Soil depth shapes plant functional diversity in granite outcrops vegetation of Southwestern Australia. Plant Ecol. Divers. 9, 263–276. doi: 10.1080/17550874.2016.1211192
Pakeman, R. J. (2013). Intra-specific leaf trait variation: management and fertility matter more than the climate at continental scales. Folia Geobot. 48, 355–371. doi: 10.1007/s12224-013-9168-y
Price, J., Tamme, R., Gazol, A., de Bello, F., Takkis, K., Uria-Diez, J., et al. (2017). Within-community environmental variability drives trait variability in species-rich grasslands. J. Veg. Sci. 28, 303–312. doi: 10.1111/jvs.12487
Qian, H., and Jin, Y. (2016). An updated megaphylogeny of plants, a tool for generating plant phylogenies and an analysis of phylogenetic community structure. J. Plant. Ecol. 9, 233–239. doi: 10.1093/jpe/rtv047
R Core Team (2020). R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna.
Rath, S., and Ormsby, A. (2020). Conservation through traditional knowledge: a review of research on the sacred groves of Odisha, India. Hum. Ecol. 48, 455–463. doi: 10.1007/s10745-020-00173-1
Ricotta, C., and Moretti, M. (2011). CWM and Rao's quadratic diversity: a unified framework for functional ecology. Oecologia 167, 181–188. doi: 10.1007/s00442-011-1965-5
Rossetto, M., and Kooyman, R. M. (2005). The tension between dispersal and persistence regulates the current distribution of rare palaeo-endemic rain forest flora: a case study. J. Ecol. 93, 906–917. doi: 10.1111/j.1365-2745.2005.01046.x
Russell, L. (2019). Emmeans: Estimated Marginal Means, Aka LEAST-Squares Means. R Package Version 1.4.2.
Sudnik-Wójcikowska, B., Moysiyenko, I., and Zachwatowicz, M. (2011). The value and need for protection of kurgan flora in the anthropogenic landscape of steppe zone in Ukraine. Plant Biosyst. 145, 638–653. doi: 10.1080/11263504.2011.601335
Szabó, S., Novák, T., and Elek, Z. (2012). Distance models in ecological network management: a case study of patch connectivity in a grassland network. J. Nat. Cons. 20, 293–300. doi: 10.1016/j.jnc.2012.06.002
Tälle, M., Deák, B., Poschlod, P., Valkó, O., Westerberg, L., and Milberg, P. (2016). Grazing vs. mowing: a meta-analysis of biodiversity benefits for grassland management. Agr. Ecosyst. Environ. 222, 200–212. doi: 10.1016/j.agee.2016.02.008
Török, P., Miglécz, T., Valkó, O., Tóth, K., Kelemen, A., Albert, Á.-J., et al. (2013). New thousand-seed weight records of the Pannonian flora and their application in analysing social behaviour types. Acta Bot. Hung. 55, 429–472. doi: 10.1556/ABot.55.2013.3-4.17
Török, P., Tóth, E., Tóth, K., Valkó, O., Deák, B., Kelbert, B., et al. (2016). New measurements of thousand-seed weights of species in the Pannonian Flora. Acta. Bot. Hung. 58, 187–198. doi: 10.1556/034.58.2016.1-2.10
Tóth, C. A., Deák, B., Nyilas, I., Bertalan, L., Valkó, O., and Novák, T. (2019). Iron age burial mounds as refugia for steppe specialist plants and invertebrates – case study from the Zsolca mounds (NE Hungary). Hacquetia 18, 195–206. doi: 10.2478/hacq-2019-0009
Tucker, C. M., Cadotte, M. W., Carvalho, S. B., Davies, T. J., Ferrier, S., Fritz, S. A., et al. (2017). A guide to phylogenetic metrics for conservation, community ecology and macroecology. Biol. Rev. 92, 698–715. doi: 10.1111/brv.12252
Vojtkó, E.-A., Balogh, N., Deák, B., Kelemen, A., Kis, S., Kiss, R., et al. (2020). Leaf trait records of vascular plant species in the Pannonian flora with special focus on endemics and rarities. Folia Geobot. 55, 73–79. doi: 10.1007/s12224-020-09363-7
Yates, C. J., Robinson, T., Wardell-Johnson, G. W., Keppel, G., Hopper, S. D., Schut, A. G. T., et al. (2019). High species diversity and turnover in granite inselberg floras highlight the need for a conservation strategy protecting many outcrops. Ecol. Evol. 9, 7660–7675. doi: 10.1002/ece3.5318
Keywords: habitat island, refuge site, verge, functional traits, phylogenetics, conservation, environmental heterogeneity, steppe
Citation: Deák B, Rádai Z, Bátori Z, Kelemen A, Lukács K, Kiss R, Maák IE and Valkó O (2021) Ancient Burial Mounds Provide Safe Havens for Grassland Specialist Plants in Transformed Landscapes—A Trait-Based Analysis. Front. Ecol. Evol. 9:619812. doi: 10.3389/fevo.2021.619812
Received: 21 October 2020; Accepted: 14 January 2021;
Published: 11 February 2021.
Edited by:
Gianluigi Ottaviani, Institute of Botany (ASCR), CzechiaReviewed by:
Lars Götzenberger, The Czech Academy of Sciences, CzechiaØystein Opedal, Lund University, Sweden
Copyright © 2021 Deák, Rádai, Bátori, Kelemen, Lukács, Kiss, Maák and Valkó. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Balázs Deák, ZGViYWxhenNAZ21haWwuY29t