 Weipan Lei
Weipan Lei Yang Wu
Yang Wu Fuxing Wu
Fuxing Wu Theunis Piersma
Theunis Piersma Zhengwang Zhang1*
Zhengwang Zhang1* José A. Masero
José A. Masero- 1Key Laboratory for Biodiversity Science and Ecological Engineering, College of Life Sciences, Beijing Normal University, Beijing, China
- 2National Demonstration Center for Experimental Life Sciences and Biotechnology Education, College of Life Sciences, Beijing Normal University, Beijing, China
- 3Fujian Provincial Key Laboratory of Marine Ecological Conservation and Restoration, Ministry of Natural Resources, Third Institute of Oceanography, Xiamen, China
- 4Rudi Drent Chair in Global Flyway Ecology, Groningen Institute for Evolutionary Life Sciences (GELIFES), University of Groningen, Groningen, Netherlands
- 5Department of Coastal Systems, NIOZ Royal Netherlands Institute for Sea Research, Texel, Netherlands
- 6Department of Anatomy, Cell Biology and Zoology, University of Extremadura, Badajoz, Spain
Artificial wetlands such as coastal saltpans have replaced a number of coastal natural habitats worldwide and may have accommodated specific waterbird populations in the East Asian–Australasian Flyway (EAAF). The role of saltpans in the EAAF as foraging grounds for shorebirds is widely recognized, although their role as breeding grounds for waterbirds is very limited and contradictory. The Nanpu saltpans in northern Bohai Bay, China, are one of the largest saltpan complexes in the world. In this study, we monitored the nesting success (852 nests) of pied avocets (Recurvirostra avosetta) during three breeding seasons (2015, 2016, and 2018) in the Nanpu saltpans. The nest daily survival rate (DSR) was 0.970; hence, nest survival over the 27 exposure days was 44%. The apparent nest success was 51%. Surprisingly, 55% of nests failed during the laying period. Flooding and nest abandonment were the main causes of nest failure during both the laying and incubation periods. We found a strong positive relationship between the DSR and nest age, with nests that approached hatching having a greater probability of survival than freshly started nests. We also found a strong negative relationship between the DSR and precipitation, with the highest DSR observed for nests that experienced no precipitation. The DSR decreased over the course of the 71-days nesting season and followed a linear trend. The DSR was also density dependent and decreased slightly when nests were denser. A literature review showed that nest survival in the Nanpu saltpans was average compared with that of other studies and that nest success in artificial wetlands was significantly higher than that in natural wetlands or both habitats. Nevertheless, nest success decreased with the study date, suggesting that that breeding conditions for the pied avocet are worsening with time. The loss of saltpans could negatively affect the population of avocets and other ground-nesting waterbird species; therefore, conservation actions and research efforts should be strengthened to understand and conserve these functional wetlands for waterbirds.
Introduction
Human-induced habitat transformation has deeply changed ecosystems and is considered a cause of global biodiversity losses (Vitousek et al., 1997; Worm et al., 2006; Burns et al., 2016). Coastal areas account for only 4% of Earth’s land area but host 40% of the world’s human population (UNEP, 2006), which has caused and continues to cause declines in coastal wetlands worldwide (Lotze et al., 2005; Cloern et al., 2016). This large scale coastal wetland loss has been identified as a key driver of the decline in waterbirds, one of the many faunal groups that depend on wetlands for survival (Kirby et al., 2008). Asia exemplifies this alarming situation with the loss of 65% tidal flats over the past five decades in the Yellow Sea, one of the most important stopover sites of the East Asia–Australia Flyway (EAAF) for waterbirds (Murray et al., 2014); this loss has caused the EAAF to be one of the most threatened flyways in the world (Kirby et al., 2008; Studds et al., 2017).
Anthropogenic wetlands such as coastal saltpans could provide alternative coastal habitats for waterbirds in the EAAF (Murray et al., 2014; Lei et al., 2018; Jackson et al., 2020). The importance of coastal saltpans for waterbirds as feeding and roosting habitats in the EAAF and other flyways is increasingly recognized (Takekawa et al., 2001; Sripanomyom et al., 2011; Green et al., 2015; Lei et al., 2018; Jackson et al., 2020); however, data on the role of coastal saltpans as breeding habitats for waterbirds in this flyway are very limited and contradictory (Que et al., 2015; Wu et al., 2020). Research on Kentish plover (Charadrius lexandrinus) breeding in saltpans located in the EAAF showed that its nest survival was among the lowest reported for this species, and human disturbance was the main cause of nest failure (Que et al., 2015). Surprisingly, gull-billed tern (Gelochelidon nilotica) breeding in these saltpans had higher nest survival than previously published estimates from other regions (Wu et al., 2020). The relative strength of the factors shaping breeding success for ground-nesting waterbirds will vary according to the geographical position of saltpans on the flyways. For example, similar to other geographic regions, large rainfall events that cause flooding during the breeding season could be a major source of nest failure in Asian saltpans. However, rainfall is not a factor for nest failure among waterbird breeding in Mediterranean saltpans of southern Europe (Rocha et al., 2016). The limited current knowledge about the potential value of coastal saltpans for waterbird breeding populations using the EAAF indicates the need for studies that aim to better understand this potential value.
In this study, we assessed the breeding success of the pied avocet (Recurvirostra avosetta), a common breeding shorebird in the Nanpu saltpans of Bohai Bay, China, one of the largest saltpan complexes in the world and a key site for migratory shorebirds and other waterbirds using the EAAF (Lei et al., 2018). Overall, pied avocets breed in flat open areas and typically along shallow saline lakes, lagoons, pools, saltpans, and estuaries, with sparse vegetation across Eurasia and Africa (Pierce, 2017). The selection of saltpans by breeding avocets has been reported from several parts of the breeding range in Europe (see review in Hötker and West, 2005). To place our findings in context, we reviewed the relevant studies on breeding success in artificial and natural habitats around the world. Specifically, we identified the leading factors influencing nest survival and addressed the issue of the value of coastal saltpans as alternative breeding habitats for pied avocets and similar shorebirds.
Materials and Methods
Study Sites
Fieldwork was carried out in the Nanpu saltpan complex (290 km2) in northern Bohai Bay, China (39°N, 118°E) (Figure 1), which is a key stopover site for migratory shorebirds along the EAAF (Yang et al., 2011; Lei et al., 2018). Saltpans are surrounded by natural intertidal mudflats in the south and west and adjacent to a nature reserve in the northeast, which is mainly composed of aquaculture ponds and rice fields (Figure 1). Similar to other industrial saltpans, the Nanpu saltpans consist of shallow, interconnected pans of varying sizes (range: 0.6–1,685 ha) separated by dikes without vegetation growth. There are three types of ponds: storage, evaporation, and crystallization ponds. Seawater is pumped into storage ponds, from which water is circulated through several evaporation ponds by gravity or pumps (Lei et al., 2018). As water flows, salinity gradually increases by solar evaporation to near the saturation point, and then the brine is pumped to the crystallization ponds. The study sites were located in two active evaporation ponds (I and II) and one temporarily abandoned evaporation pond (III). There is a plant that produces bromine between ponds I and II, using water from pond I and wastewater discharge into pond II; thus, almost no food was available for shorebirds in pond II (Weipan Lei, personal observation). Four different types of structures for nest sites were distinguished: (1) temporarily inactive (empty) evaporation pond (pond III), (2) dike isolated by water in evaporation pond (A and G dikes), (3) accessible dikes connected to road (B–F and H dikes), and (4) islet in evaporation pond (islet J) (Figure 1). The nesting substrate of dikes and abandoned ponds is mud, while the islets are mainly made up of shells. We assumed that differences in nesting substrate, food resources, and accessibility could cause nest survival variations. Research sites were monitored in 2015 (pond III), 2016 (pond II), and 2018 (ponds I, II, and III). Kentish plover, black-winged stilt (Himantopus himantopus), common tern (Sterna hirundo), gull-billed tern, and little tern (Sterna albifrons) were also bred in the study area (Yang, 2006; Que et al., 2015).
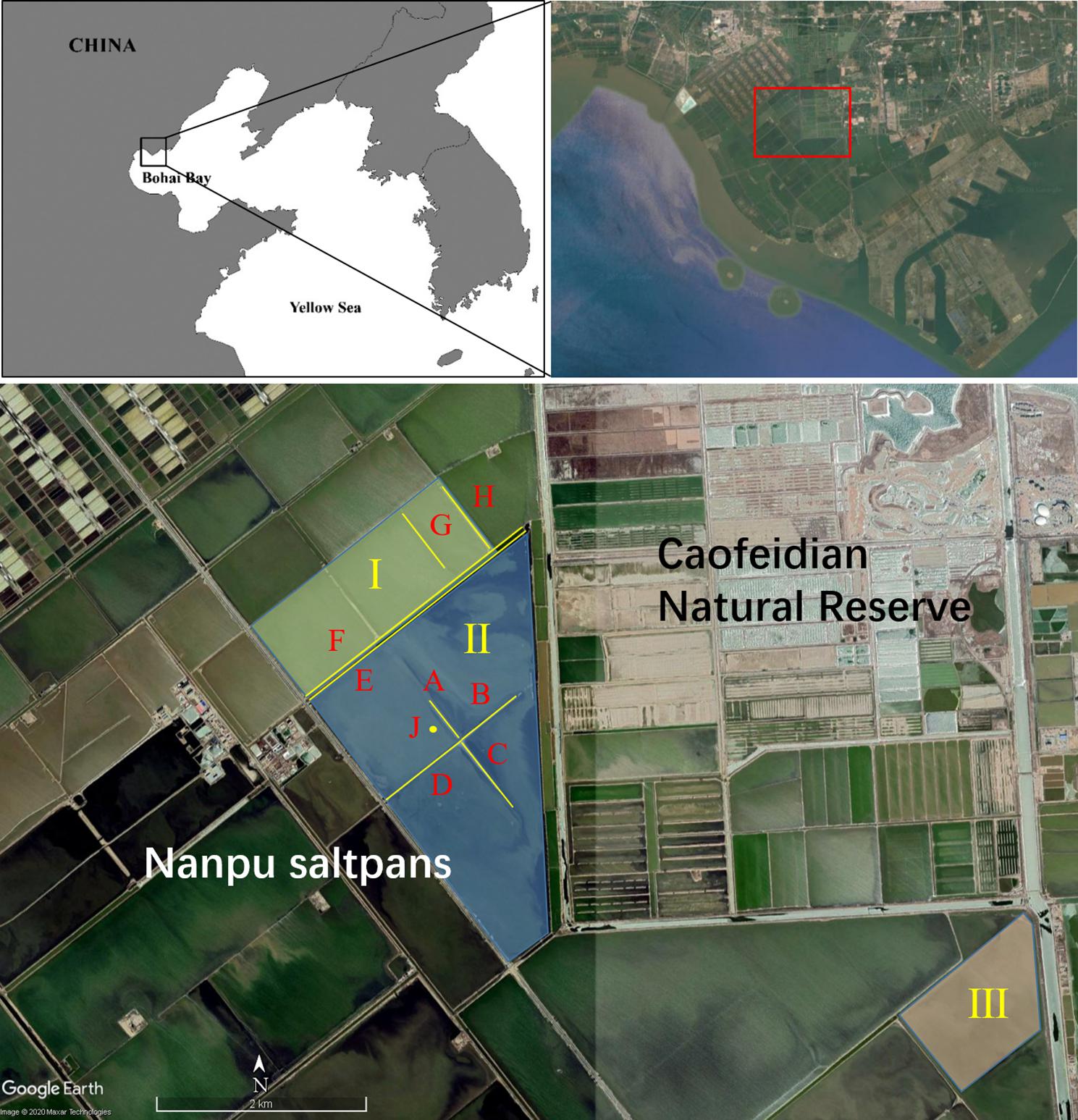
Figure 1. Study area in the Nanpu saltpans. Ponds I and II were active (flooded) ponds, but Pond III was a temporarily inactive (empty) evaporation pond. A–H, are dikes, and “J” is a small islet. See text for details.
Dikes were 10-m wide and 54-cm high, on average, from the water. Vegetation was not observed in these dikes and the islet. The mean water depth in pond II was 46 cm. We did not measure the water depth in pond I, but it was similar to that in pond II in 2018. The area of the islet is about 180 m2.
Field Methods
We systematically searched for nests on foot in the study area and recorded the location of nests with a handheld GPS. Every nest found was numbered and marked with inconspicuous shells, and eggs were marked individually and measured. We recorded the position of the nest in dikes or islets as top, slope, and bottom (nests in pond III without dikes/islets were all on the bottom) (Supplementary Figure 2). The avocet nesting period included egg laying and incubation periods. For the nests found during the laying period, we backward calculated the first laying date based on the avocet laying pattern: laying one egg each day and a 1-day break without laying between the third and fourth eggs (Hötker, 1998). Avocets generally started incubating after laying the third or fourth egg (Hötker, 1998); for estimating the hatching date, we assumed that they started to incubate after laying the last egg; thus, the hatching date was 24 days after laying the last egg. For the nests found with a full clutch, we floated the eggs to estimate the incubation days (i.e., the last egg laying date) (Alberico, 1995; Ackerman and Eagles-Smith, 2010), backward-calculated the first egg-laying date based on the number of eggs and estimated the hatching date. Nest age day 1 was defined as the first egg-laying date. We could not estimate the nest age of nests that failed before we found them. We checked each nest every 1–9 days (normally 3–5 days) from the day found until the final fate of the nest was determined. Each day that the nests were visited, they were coded as 1; otherwise, they were coded as 0 for subsequent nest survival analysis (Rotellam et al., 2000; Uher-Koch et al., 2015; Hu et al., 2020) (see “Data Analysis” section).
We considered the nest to be successful if (1) at least one chick was found in a nest or nearby, (2) we observed 1- to 4-mm shell fragments in or near the nest, or (3) the date of egg disappearance matched the expected hatching date with no sign of predation (Mabee, 1997; Rocha et al., 2016). We used the definition of incubation period from Drent (1975), i.e., the number of days between the last egg laying and the last chick hatching of each nest. Thus, hatching date was the day when the last chick hatched, the day after we observed a nest with all eggs pipping and fractures on the shell, 2 days after we observed multiple eggs with cracks but no pipping, or 3 days after we observed only one egg with fractures (Ackerman and Eagles-Smith, 2010). Avocet hatching usually lasts 2–3 days, i.e., from the first egg with fractures to the last chick hatched.
We considered the nest to fail if the following occurred: (1) flooding: eggs were flooded, covered with dirt, or blown out of the nest scrape; (2) predation: eggs were broken or yolk and albumen could be found around the nest, or eggs disappeared at least 5 days before the predicted hatching date; (3) poaching: previously observed eggs were collected by humans, or the nest was empty and human footprints were observed; (4) trampling: eggs were broken in the nest and footprints were obvious; (5) abandonment: eggs were unattended by adults for prolonged periods (eggs were considered abandoned if they were found cold and dirty for at least two visits); or (6) failing to hatch: nest was incubated more than 35 days but eggs did not hatch.
Nest fates were defined as unknown if the above criteria could not be determined (Walker et al., 2005; Colwell et al., 2011). Nests with unknown fates and those that did not finish at the end of the fieldwork seasons were not used in the analyses. Precipitation data were obtained from the Nanpu saltpan weather station.
Data Analysis
We standardized the season dates among years by using the earliest date in either year as the first day of the season and the latest hatching or failure date in either year as the last day of the season (Moynahan et al., 2007). We thus defined a 71-days nesting season beginning on April 20 and ending on June 29. Incubation days were calculated from the successful nests found during the laying period. The complete clutch size excluded nests that did not start to incubate with one or two eggs. Because various authors have used different terminology, we defined the following terms for this paper: (1) “nest daily survival rate” (DSR) is the probability that a nest will survive a single day, and (2) “nest success” is the probability that a nest will be successful, including both apparent nest success and cumulative nest success based on DSR.
We used ArcGIS 10.2 (ESRI, Redlands, CA, United States) to calculate spatial covariates: nearest distance to other avocet nests and number of avocet nests within a 25-m radius (nest density) (25 m was used because it is an indicator of colony nests; Hötker, 2000). For the nests on the islet, the distance between nests was beyond the GPS accuracy, and the radius of the islet was smaller than 25 m; therefore, we used 0.5 m as the average distance between nests, and density was calculated considering all nests (including the nests of common tern).
The nest survival analysis procedure in the program MARK 9.0 (White and Burnham, 1999; Cooch and White, 2006) via the R package “RMark” version 2.2.7 (Laake, 2013; Laake et al., 2019) was used to estimate the DSR and assess how environmental and temporal factors affected the DSR. Only the known nest fate and nest age were used in the analysis, and the nests that failed with unknown nest age were not used in this analysis, although they were still used to calculate the apparent nest success.
The nest survival analysis in MARK uses a generalized linear model with a logit link function and binominal error to estimate the relationship of the DSR with various covariates (Rotella et al., 2004; Hu et al., 2020). The parameters in the encounter history of the nests included FirstFound (date when the nest was first found), LastPresent (date when nests were last found present), LastChecked (date when the nest was last visited; for success nest, LastPresent is the same as LastChecked), Fate (fate of the nest; 0 = success and 1 = fail), Freq (frequency of the nests with the same encounter history), AgeFound (age of the nest when it was found), and AgeDay1 (age of the nest on the first day of the nesting season, i.e., April 20 in this study).
The covariates considered to explain the DSR in the model included year, nesting season, precipitation (1-day lag effect), nest age, nest position (top, slope, or bottom), pond, habitat, nest density, distance to nearest nest, and researchers’ nest visit (Table 1). A 1-day lag of precipitation was used because the initial analysis showed that it performed better than precipitation (Moynahan et al., 2007). Thus, precipitation from April 19 to June 27 was used in the model corresponding to 1–70 intervals within 71 nesting season days (the following precipitation just means a 1-day lag of precipitation for conciseness). We used the pairs function in R to check the colinearity among covariates of year, Ageday1, position, pond, habitat, density, and distance to nearest nest. The pairplot indicated colinearity between habitat (structure) and pond (Supplementary Figure 2). Then, we used the “corvif” function in the HighstatLibV4.R file (Zuur et al., 2013) to check the variance inflation factor values of pond and habitat with other covariates separately (an error occurred when pond and habitat were checked together), which showed that the habitat had the largest GVIF value (4.22); therefore, we dropped it then the GVIF values of the remaining covariates were smaller than 3.
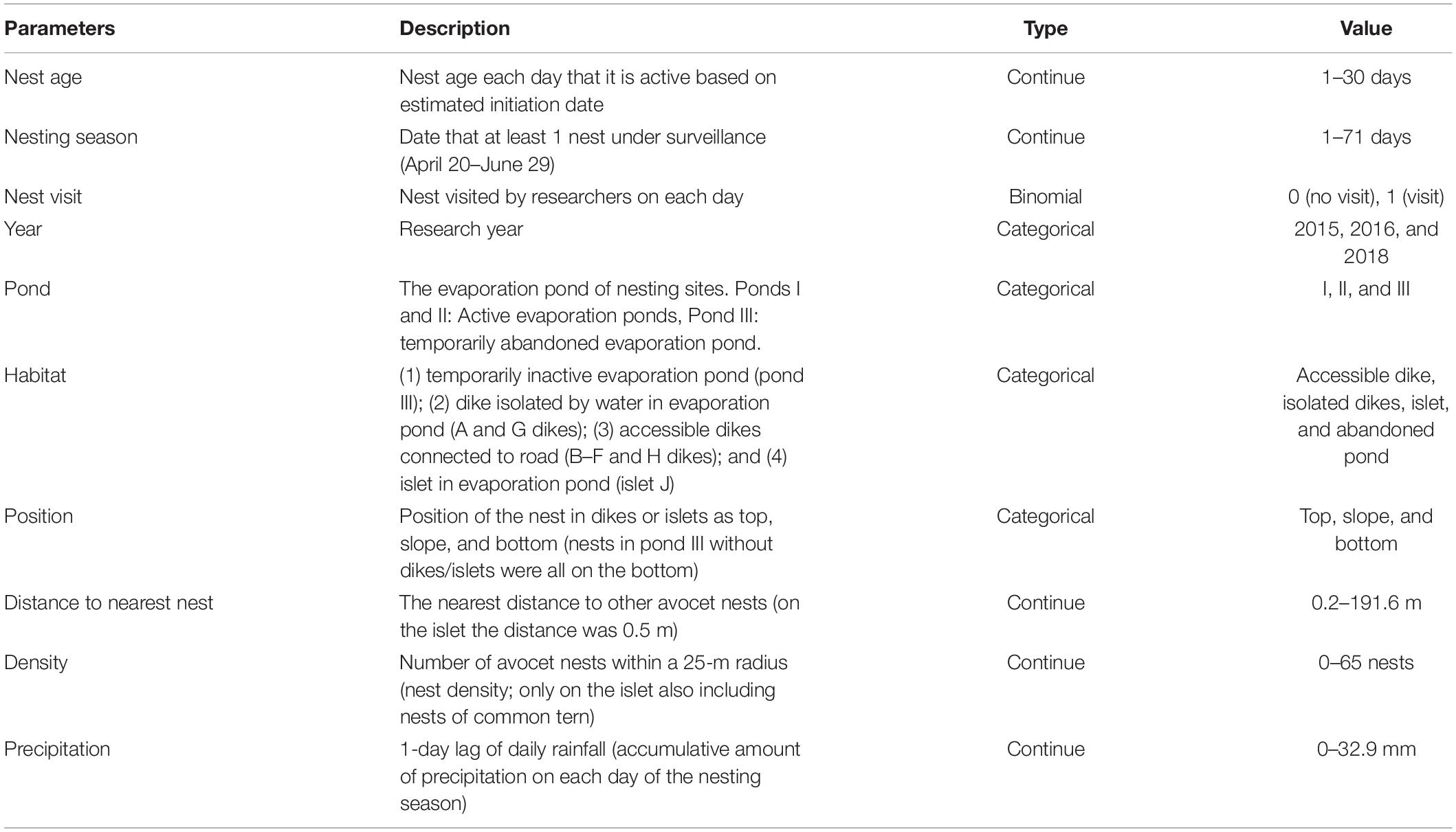
Table 1. Parameters used in models predict daily nest survival rate of pied avocet in Nanpu saltpans.
Corrected Akaike’s information criterion (AICc) was used to rank candidate models for small sample sizes to assess model likelihood and parameter parsimony given the data (Burnham and Anderson, 2004). First, we built nine models with each single covariate and a constant model. The model with nest age was much better than the others (Supplementary Table 2), and then, we built models with two covariates (nest age and other single covariates). Only the models better than the previous top model were advanced to the next stage of model building. A new top model as a foundation for model construction and other single covariates (from models advanced to this stage) were added into the top models each time. Keep this process until the AICc was not smaller. The models with △AICc < 2 were considered the best candidate models. Furthermore, we averaged all models using the “model. avg” function using the package “MuMIn” (Barton and Barton, 2015). The relative importance of the Akaike weights of response variables (by the sum of Akaike weights of models where explanatory variable was present) (Burnham and Anderson, 2004) and 95% confidence intervals (CIs) were used to evaluate the contribution of the variables to the models. We considered covariates with 95% CIs that did not overlap 0 as biologically informative.
We visually assessed the sensitivity of biologically informative covariates in the selected model by plotting back-transformed estimates of the DSR using a mean or range of values representative of those recorded during the nesting season and confidence intervals derived using the delta method (Seber, 1982; Sexson and Farley, 2012). We calculated estimates of cumulative nest success by multiplying the DSR by the 27th exponent, which is consistent with a 27-days laying and incubation period (see result), and apparent nest success = succeeded nests/(succeeded nests + failed nests).
We also searched the literature for nest survival values for pied avocet to model nest success based on different predictor variables, including habitat type (see below). To identify similar studies that estimated nest success in pied avocet, we queried the “Web of Science” and “Google Scholar” databases using the terms “pied avocet,” “Recurvirostra avosetta,” “breed,” and “nest survival”. Studies not included in those databases but listed in Hötker and Segebade (2000); MacDonald and Bolton (2008), and Chokri and Selmi (2011) were also included in the literature review. We classified habitats as natural (lake, islet, salt marsh, and delta), artificial (saltpans, fish ponds, grasslands, summer polder, and dikes), or both. Studies including habitats we could not classify were excluded from the analysis. If only the DSR value was available, we calculated the cumulative nest success by multiplying the DSR by the 27th exponent. If the results of several years were presented, we pooled the nests to calculate the apparent nest success or used the average DSR to calculate the cumulative nest success. Beta regression (with a logit link function; betareg 3.1-4 in R) (Zeileis et al., 2016) was used to detect whether the habitat, study date (year, or last year, used in the study to estimate nest parameters), number of nests, and method for estimating nest success (apparent nest success or cumulative nest success) affected the nest success. All GVIF values were <2, indicating no colinearity among the predictive variables.
All analyses were carried out in R studio (Version 1.1.456, RStudio, Inc.). The significance levels were set at P < 0.05. The mean values were given as means ± SE.
Results
A total of 852 nests were found, and among the 804 known nest fates, 408 nests succeeded and 396 nests failed, which represented an apparent nest success of 50.8% (Table 2). The clutch size was 3.88 ± 0.02 eggs (n = 566), with four eggs as the most common clutch size that had the highest apparent nest success (Figure 2). The incubation period was 23.2 ± 0.16 days (19–31 days, n = 135). The nesting period, including incubation and laying, was 27.1 ± 0.16 days (23–35 days, n = 135). The overall DSR, assuming a constant survival rate without the influence of covariates, was 0.970; hence, the cumulative nest success during 27 nesting exposure days was 43.8%.
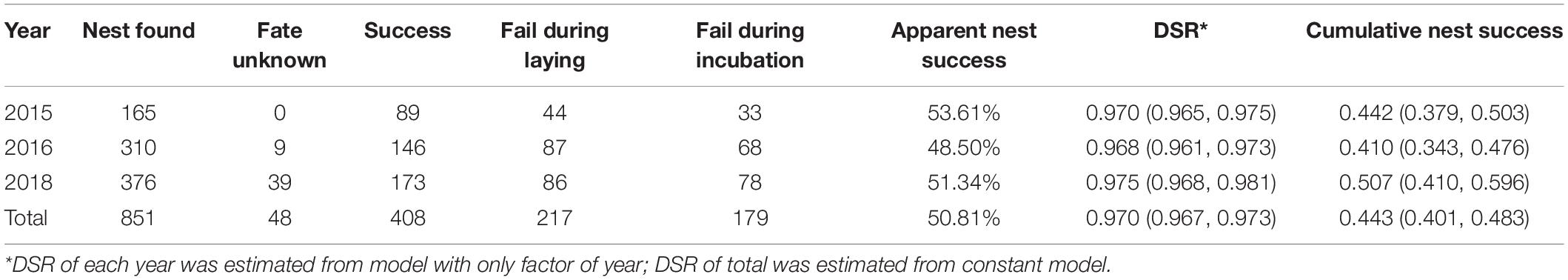
Table 2. Total number of pied avocet nests monitored and used to predict daily survival rate (DSR) in nest survival models in program MARK.
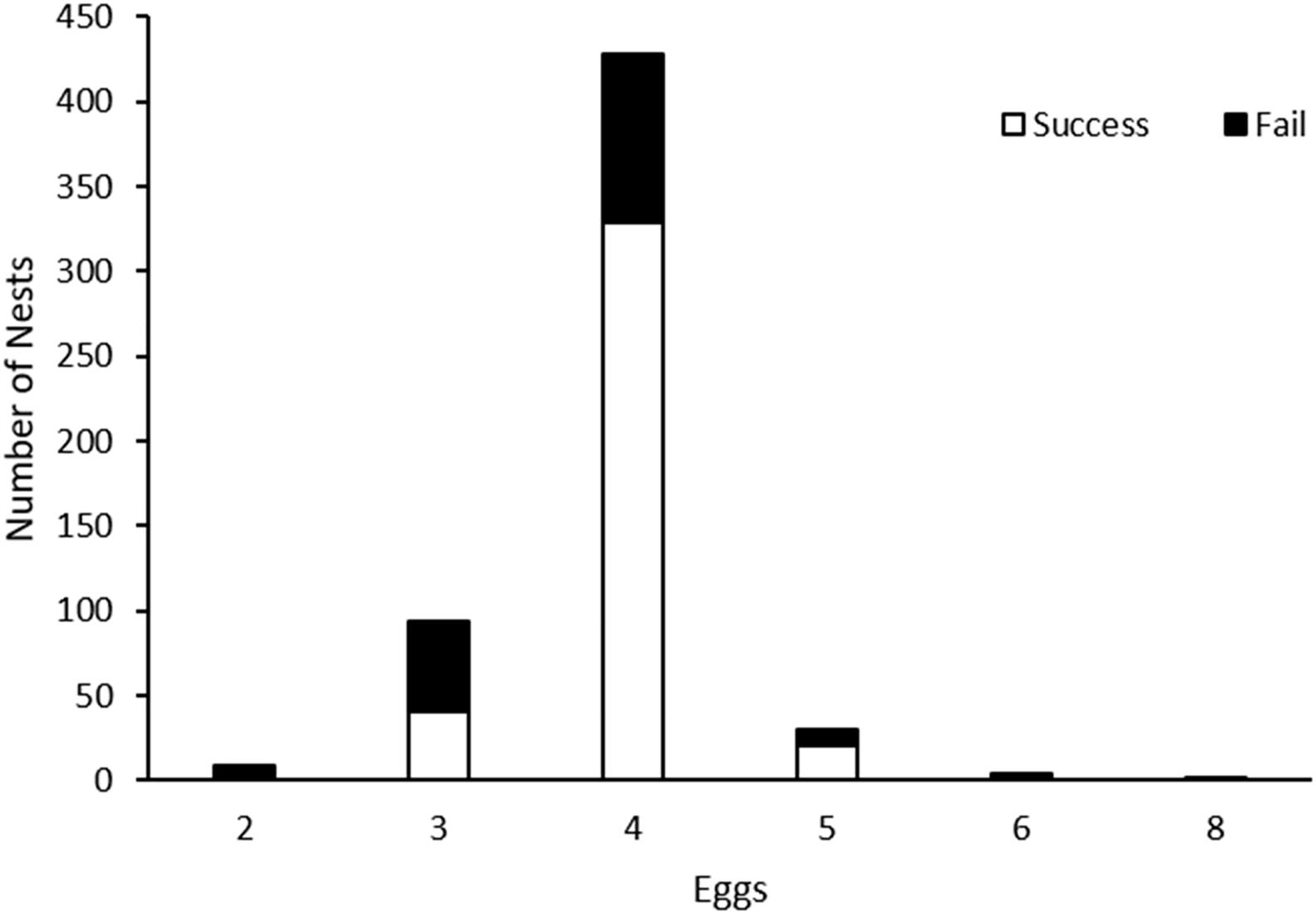
Figure 2. Clutch size of pied avocet found in the Nanpu saltpans and their fate. Nests with uncompleted clutches were excluded from the analysis.
Among the failed nests, 217 nests (54.8%) failed during the laying period (170 nests with one egg, 46 nests with two eggs, and one nest with the number of eggs unknown), and 179 nests failed during the incubation period. Nest abandonment was the main cause leading to nest failure during both the laying (44.7%, n = 97 out of 217 nests) and incubation periods (27.3%, n = 49 out of 179 nests), followed by flooding (laying: 38.7%, n = 84; incubation: 44.1%, n = 79), predation (15.7%, n = 34; 16.2%, n = 29), and poaching (0.9%, n = 2; 9.5%, n = 17). Other reasons included trampling (n = 1) and unviable eggs (n = 4), which only occurred during the incubation period (Figure 3). Most nests were located at the bottom of the pond, dikes, or islet (n = 556). Although fewer nests were found on the top (n = 126) and slope (n = 122) of dikes or islet, they had higher apparent nest success than those on the bottom (Figure 4). Nest abandonment and flooding were the main reasons for nest failure at all nest positions. Nests placed on the top of dikes and islets had a high predation rate (Figure 4).
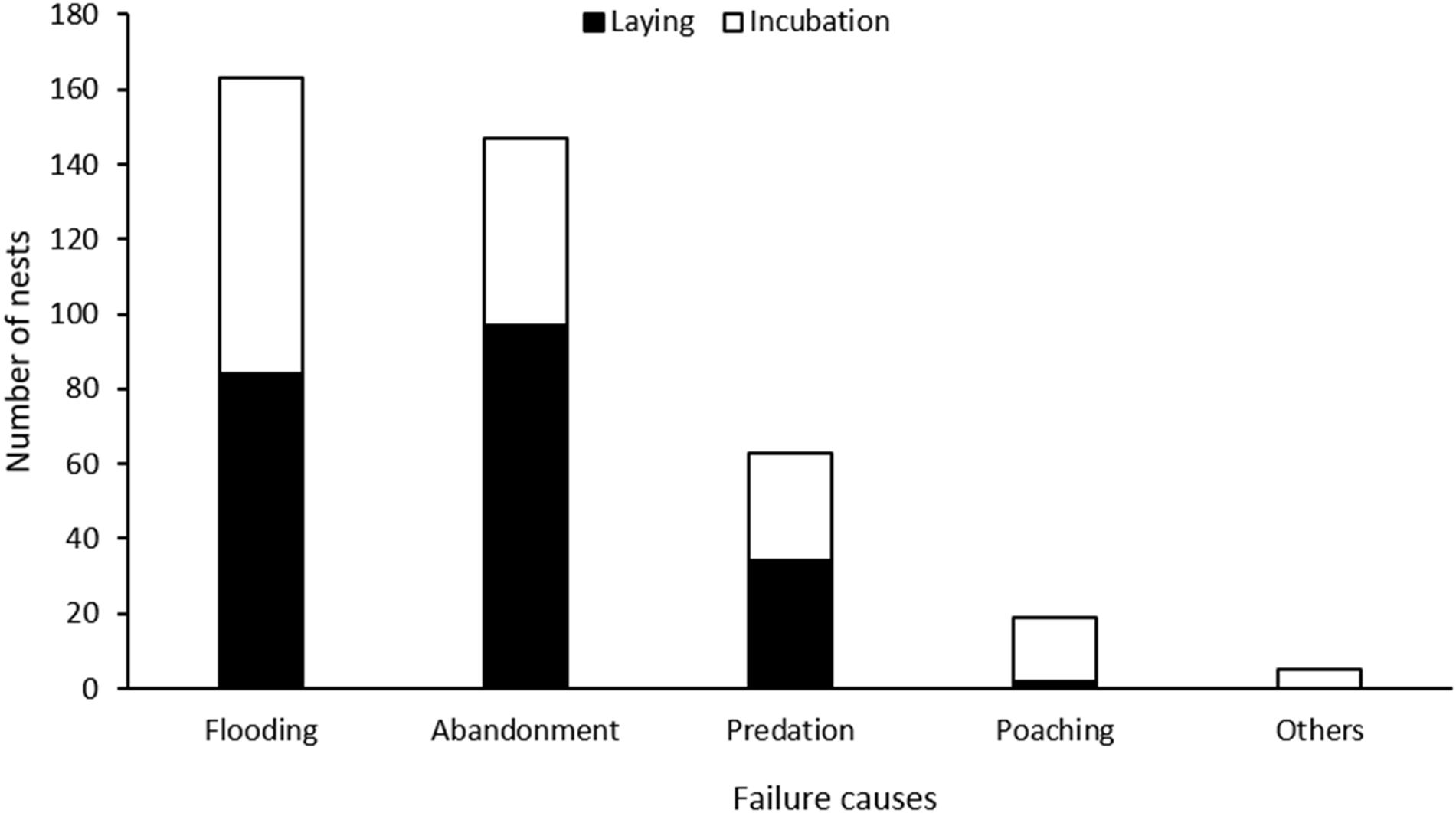
Figure 3. Causes of nest failure among pied avocets during the laying (black) and incubation (white) periods; other causes, including trampling (n = 1) and unviable eggs (n = 4), only occurred during the incubation period.
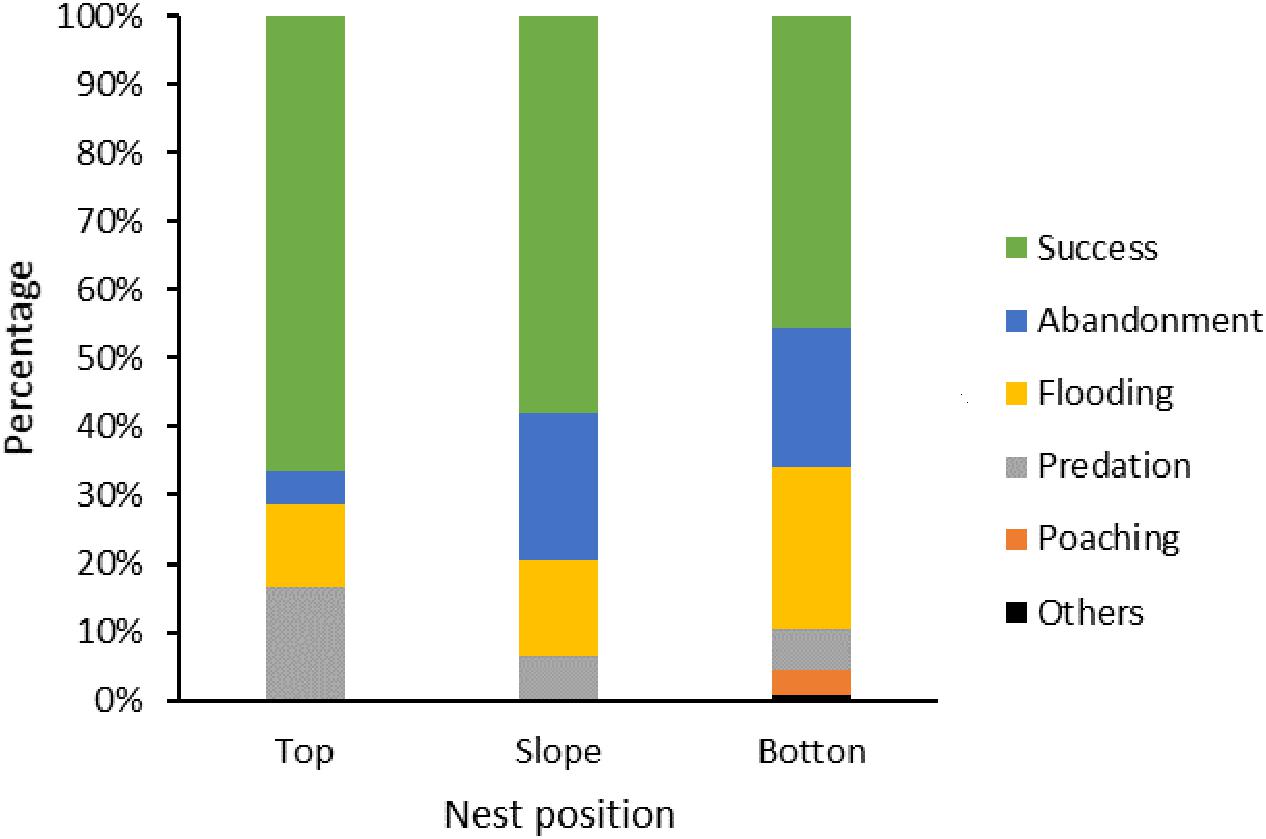
Figure 4. Proportion (%) of apparent nest success and failure causes according to nest position in dike or islet: top (n = 126), slope (n = 122), and bottom (n = 556).
We constructed 25 candidate models describing variations in the DSR (Supplementary Table 1). Only one model had △AICc < 2 (wi = 0.80), and it included nest age (β = 0.157 ± 0.011, 95% CI = 0.135, 0.178), nesting season (β = –0.035 ± 0.006, 95% CI = –0.047, –0.023), precipitation (β = –0.087 ± 0.017, 95% CI = –0.119, –0.054), nest density (β = –0.148 ± 0.004, 95% CI = –0.023, –0.007), and position (bottom β = 0.284 ± 0.215, 95% CI = –0.706, 0.138; slope β = 0.144 ± 0.270, 95% CI = –0.386, 0.673) (Table 3). The second and third models were nested in the top model (Table 3). After model averaging, the most biologically informative covariates (wi = 1) were nest age, nesting season, and precipitation, followed by nest density (wi = 0.99). Nest position (wi = 0.80), pond, year, nest visit, and distance to nearest nest (all wi < 0.01) were not biologically informative by evidence with a 95% CI that overlapped 0 (Table 4).
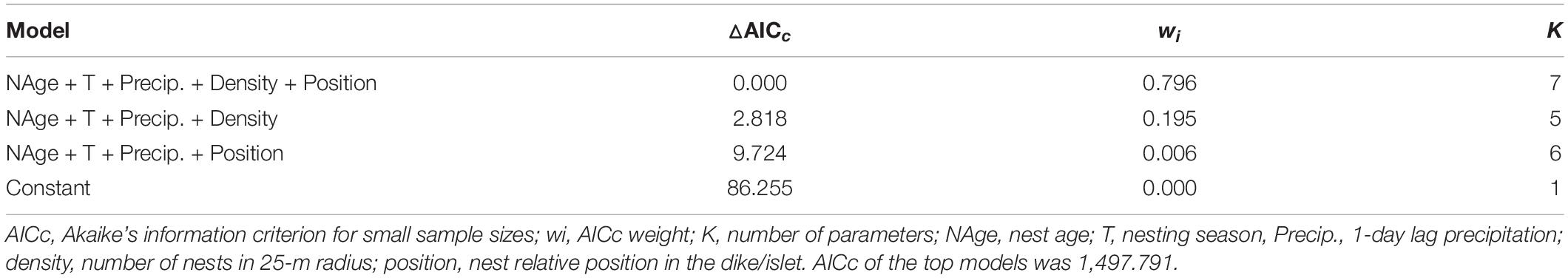
Table 3. Top three modes and constant model predicting pied avocet daily survival rate in Nanpu saltpans.
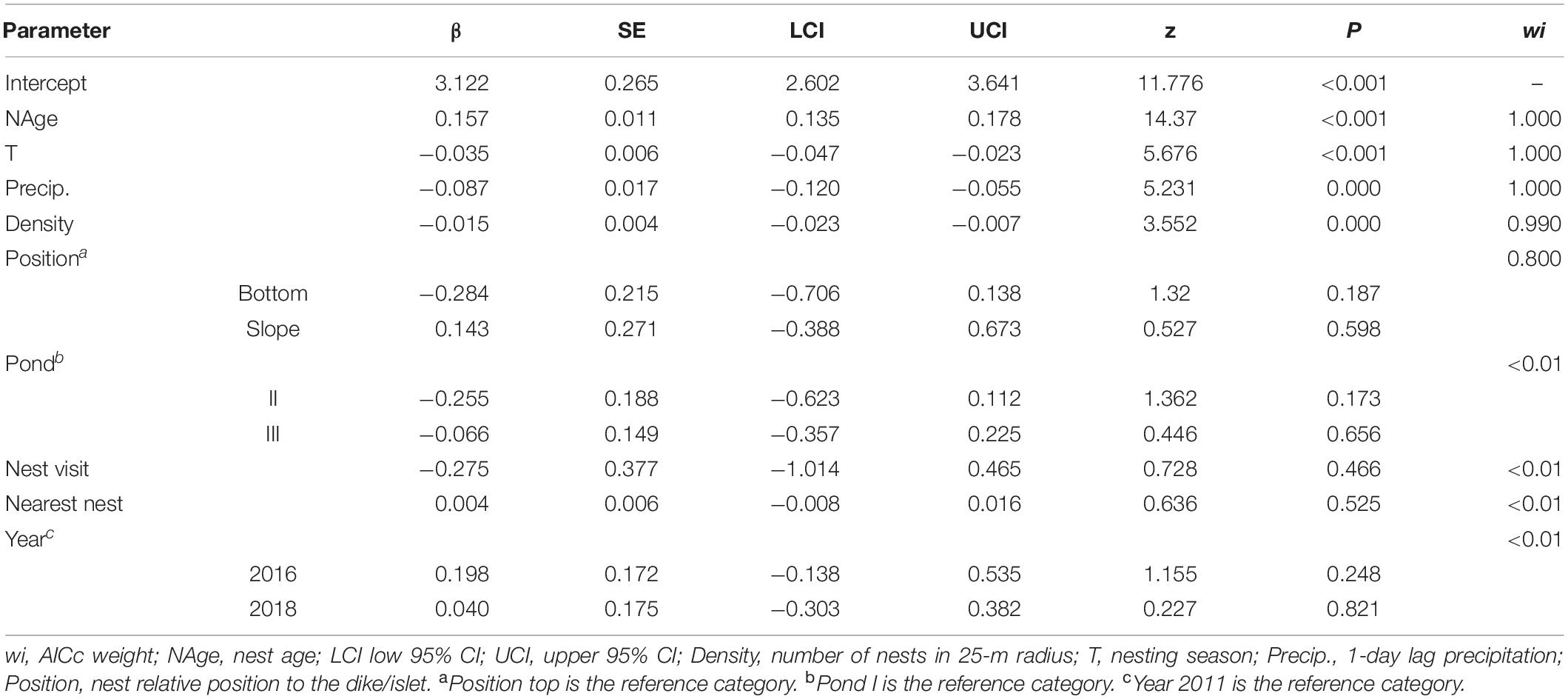
Table 4. Model-averaged parameter estimates and descriptive statistics.
Model averaging had a similar predictive ability to the single top model; therefore, we plotted back-transformed estimates of the DSR based on the top model. We found a strong positive relationship between the DSR and nest age, with nests near hatching showing a greater probability of survival (Figure 5A). We also found a strong negative relationship between the DSR and precipitation, with the greatest DSR values at nests that experienced no precipitation (Figure 5B). The DSR decreased over the course of the 71-days nesting season following a linear trend, so nests active in late season had a smaller probability of survival (Figure 5C). The DSR also decreased slightly with nest density (Figure 5D). The nests on the bottom had a lower DSR than those on the top, but the difference was not significant, with a 95% CI that overlapped 0.
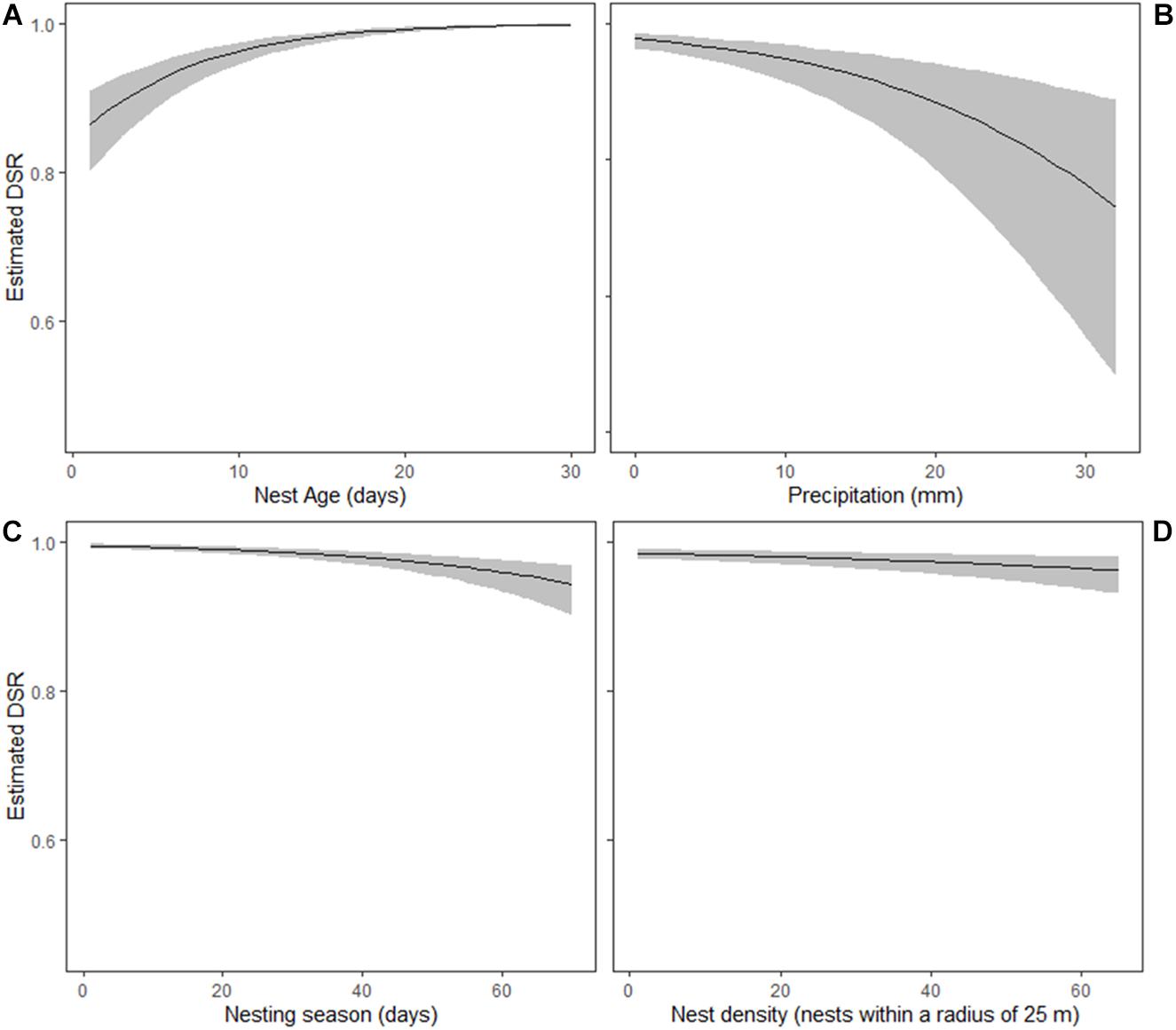
Figure 5. Predicted daily survival rates (95% CI) of pied avocet nests in relation to nest age (A), precipitation (B), nesting season (C), and nest density (D). Predicted based on the top nest survival model (Table 3). For illustrative purposes, we only plotted daily survival rate (DSR) values for the top position of nests.
The literature review showed that nest survival of pied avocet was highly variable: 58.3 ± 5.3% (n = 13 studies, apparent nest success) and 58.0 ± 11.1% (n = 5, cumulative nest success), ranging from 23 to 88%. Our average apparent nest success was 50.8 ± 1.5% (n = 3 years), and the cumulative nest success was 44.3% (40.1, 48.3% 95% CI); however, the nest-abandonment rate in the Nanpu saltpans was clearly superior to that of other studies (Supplementary Table 2). The beta regression showed that habitat and year significantly affected nest success (Table 5). Nest success in artificial habitats was significantly higher than that in natural or mixed habitats and decreased over time (Figure 6). The effects of the number of nests and nest success calculation method were not significant (Table 5).
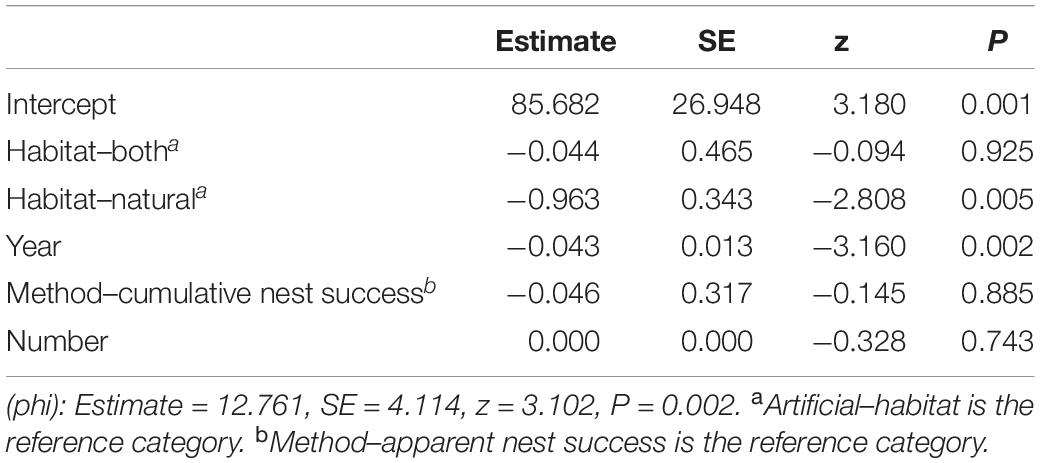
Table 5. Beta regression predicting the nest success of avocet in natural and artificial habitats.
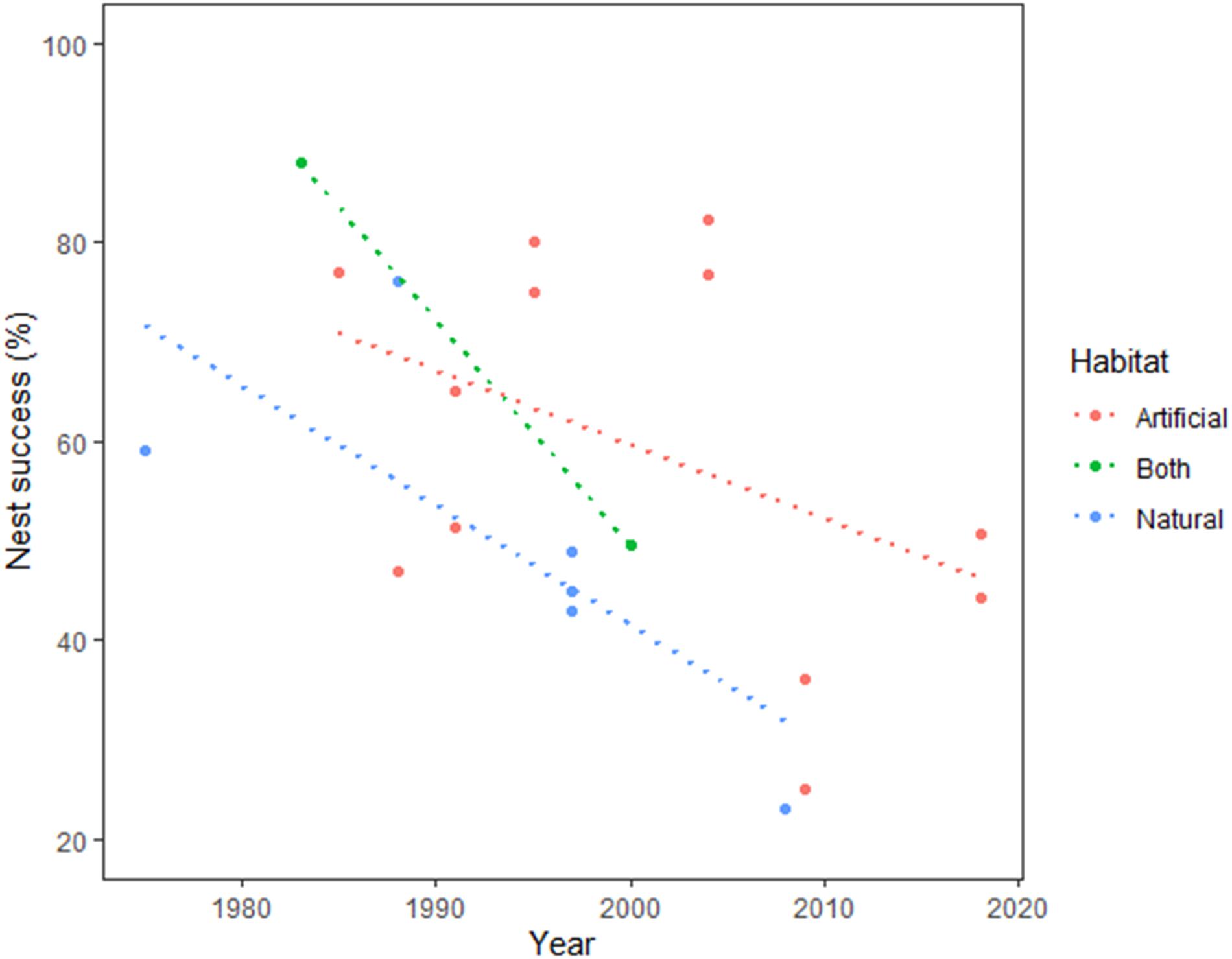
Figure 6. Relationships between nest success and study date and between nest success and nesting habitat. Data from this study and literature review.
Discussion
We found that the nest survival, which is a proxy of breeding success, of pied avocet in the Nanpu saltpans was similar to the reported values in the literature, which included studies performed in natural and artificial wetlands. Our analysis including data about nest survival of pied avocets across different geographical regions suggested that breeding conditions seem more favorable in anthropogenic wetlands, such as our saltpans, where nest survival values were as high or higher than those reported for natural and artificial wetlands. This analysis also showed that nest survival decreased over time. Thus, coastal saltpans in the EAAF could provide alternative breeding sites for pied avocets, although regardless of the type of habitat, breeding conditions may have worsened over the last decades.
Field activities may lead to biased estimates of nest success (Rotellam et al., 2000) because researchers’ visits may provide cues for predators and/or brood parasites finding nests (Nichols et al., 1984; Westmoreland and Best, 1985; Major, 1990) or may prevent some predators from visiting nests (MaCivor et al., 1990; Ibáñez-Álamo and Soler, 2010). In this research, we failed to detect the visit effect, suggesting that the bias of the observing effect should be limited. This finding may be related to our attempt to minimize the disturbance during nest checks, as we inconspicuously marked the nests and avoided nest checks after heavy rainfall to avoid leaving the footprint. Additionally, the rate of predation and poaching was not high in this study. The low poaching rate, but not the low predator rate, may be related to our research activities because we have persuaded several egg collectors to stop egg collection; however, we never observed land predators at the nesting sites. A meta-analysis also showed that Charadriiformes did not show a significant increase in nest predation due to research perturbation (Ibáñez-Álamo et al., 2012).
Nest survival in the Nanpu saltpans was strongly affected by nest age and precipitation as well as by days from the beginning of the breeding season (nesting season) and nest density. Flooding and abandonment were the main causes of nest failure. In comparison with other waterbirds breeding in the Nanpu saltpans, this nest survival was much higher than that of Kentish plover (Que et al., 2015) but lower than that in gull-billed tern (Wu et al., 2020), both of which were also estimated in the Nanpu saltpans but at different sites. These differences may be due to the different human disturbance levels occurring within the Nanpu saltpans. The study sites for Kentish plovers were located in dikes formed from crystallization and shrimp ponds with high levels of anthropogenic disturbance (e.g., egg collection, irrigation, and trampling by pedestrians or vehicles) (Que et al., 2015), while the study sites for gull-billed terns were mainly isolated islets or abandoned ponds with low human disturbance (Wu et al., 2020). Both avocet and gull-billed tern nest survival were strongly and negatively affected by rainfall and anthropogenic activities (water pumping activity for salt production), which led to flooding events (Wu et al., 2020).
Our models showed that the DSR was negatively correlated with precipitation and decreased as the breeding season progressed. In the study area, large rainfall events occur annually after June, i.e., in the last stage of the breeding season, with up to 32.9 mm in a single day (Supplementary Figure 3). Precipitation leading to flooding may have an immediate and widespread effect on ground-nesting shorebirds that use flood-prone saltpans (Knopf, 1982; Winton et al., 2000; Sexson and Farley, 2012). We did not discriminate nest failure by flooding caused by precipitation or by flooding caused by saltworkers’ activity. Flooding is the main cause of nest failure in pied avocets in both natural (De Bie and Zijlstra, 1985; Hötker and Segebade, 2000; Thyen, 2005) and artificial habitats (Engelmoer and Blomert, 1985; Nogueira et al., 1996; Cuervo, 2005). In natural habitats, flooding is caused by rainfall events (e.g., Hötker and Segebade, 2000) or tides (Plaschke et al., 2019), and this negative effect on nest survival is habitat dependent. For example, Hötker and Segebade (2000) found that on natural saltmarshes, most avocet nest losses were due to flooding at storm tides. In artificial habitats, however, flooding is also caused by water manipulation (Nogueira et al., 1996; Chokri and Selmi, 2011). The threat of anthropogenic flooding can be more serious than other negative factors, such as predation, but this threat could be easily reduced or eliminated by controlling water levels during the breeding season or by building nesting structures to prevent flooding (Nogueira et al., 1996).
We did not find differences in nest survival among nest positions, although avocets seem to prefer nesting on the bottom. Nests in the Nanpu saltpans were threatened by both strong rainfall and wind events and anthropogenic water manipulation. Salt production requires massive influxes of water flow through different ponds during the breeding season. When the water level dramatically increases in a pond, water can drown and even wash away the nests on the bottom or slope of dikes and islets. If this water management by saltworkers coincides with heavy rainfall and strong wind, many nests will fail at a short time scale. The functionality of coastal saltpans for breeding would increase for waterbirds by avoiding or minimizing water management practices that negatively impact breeding conditions for ground-nesting waterbird species such as pied avocets.
Another main cause of nest failure was nest abandonment or desertion. Many reasons cause nest abandonment, such as predation risk, clutch loss, competition for nest sites, inclement weather, or human disturbance (see review in Roche et al., 2010). The abandonment rate in this study was much higher (especially during the laying period, which accounted for more than 50% of the cases) than in other avocet studies either in natural or artificial habitats (abandonment rate <20%) (Nogueira et al., 1996; Hötker and Segebade, 2000; Cuervo, 2005; Lengyel, 2006; Chokri and Selmi, 2011). Avocets face interspecific competition for nesting sites with black-winged stilts or common terns, all breeding at the same sites. In addition, thousands of gulls, such as the black-headed gull (Larus ridibundus) and relict gull (Larus relictus), use avocet nesting dikes (pond I) for roosting at night, and infrared cameras shows that the gulls preyed avocet eggs when they were unattended (Weipan Lei, pers. Observ.). Chokri and Selmi (2011) found that 54% of deserted nests were due to avocet competition with breeding slender-billed gull (Chroicocephalus genei). Another reason for nest abandonment may be the type of ground cover of the dikes, which would lead to eggs sticking to the ground when the muddy substrate is wet, and birds are unable to turn them (Arroyo and Hortas, 2005). This is unlikely to happen in natural habitats where the ground cover is composed of shells, sand, or grass.
Predation is the primary cause of nest failure in almost all avian species studied to date (Ricklefs, 1969; Martin, 1993; Smith and Wilson, 2010). In previous studies, the nest predation rate of pied avocet was high, mainly caused by mammalian or avian predators (Hötker and Segebade, 2000; Cuervo, 2004). Most predators, such as gulls (Nogueira et al., 1996), corvids (Goutner, 1985), foxes (Cadbury and Olney, 1978; Goutner, 1985; Hötker and Segebade, 2000), and Rattus spp. (Cuervo, 2004; Wu et al., 2020), hunt in both natural and artificial habitats, although certain predators, such as stray dogs (Chokri and Selmi, 2011) or poachers (egg collectors) (Que et al., 2015), are more likely to hunt in artificial habitats. However, the rate of nest predation in Nanpu saltpans was comparatively low for avocets.
The DSR also increased with nest age, such as in other biparental species, since parents defend older nests with greater intensity (Smith and Wilson, 2010). Avocets usually do not start to incubate until laying the third or fourth egg, so nests during the laying period are extremely vulnerable. When nest age increases, parents defend them more aggressively (reviewed in Montgomerie and Weatherhead, 1988; Forbes et al., 1994). In addition, the opportunity for renesting successfully declines because nests initiated in the late season are unlikely to succeed (Smith et al., 2010). Nesting season and nest age are often confounded. Nevertheless, our model showed that the DSR increased with nest age but decreased with nesting season, suggesting that older nests were defended by incubating adults with greater intensity.
In addition to avocet, many species use saltpans or other artificial habitats for breeding, such as American avocet (Recurvirostra americana) (Rintoul et al., 2003), black-necked stilt Himantopus mexicanus (Rintoul et al., 2003), black-winged stilt (Cuervo, 2005), Kentish plover (Que et al., 2015; Rocha et al., 2016), snowy plover (Charadrius alexandrinus nivosus) (Robinson, 2008), gull-billed tern (Wu et al., 2020), little tern (Catry et al., 2004), common tern (Škornik, 2019), Forster’s terns (Sterna forsteri) (Bluso-Demers et al., 2016), and slender-billed gull (Ramírez et al., 2012). All of these species prefer to breed in flat, open habitats with sparse vegetation for easy detection of predators, such as sand beaches, before land transformation. Catry et al. (2004) showed a slight shift in tern breeding from sand beaches to saltpans over 30 years but without a population decline. Species that prefer nesting in dense grass to hide from predators, such as redshank (Beintema and Muskens, 1987), seldom choose bare-land saltpans as breeding habitats. Thus, the different predator avoidance strategies of breeding birds may affect their use of artificial habitats.
In the Mediterranean basin, waterbirds tend to breed in artificial and semiartificial habitats, especially in saltpans, due to intensive human pressure on natural habitats (Britton and Johnson, 1987; Masero, 2003; Catry et al., 2004; Ali Chokri and Selmi, 2011). For example, saltpans and extensive fishponds support more than 75% of the avocet population breeding in Spain (Arroyo and Hortas, 2005). Saltpans provide stable and predictable feeding areas (Britton and Johnson, 1987; Masero and Pérez-Hurtado, 2001) as well as suitable nest sites and abundant food resources for chicks (e.g., Arroyo et al., 1997). The avocet population decline that occurred in some areas in Spain, such as the Bay of Cádiz, seems to be connected with the abandonment or transformation of saltpans into fishponds or other land uses (Arroyo and Hortas, 2005). From the Yellow Sea to southeast Asia, although saltpans are widely distributed and have a long history, knowledge of their value for breeding is still very limited, and large surfaces of saltpans are undergoing a similar transformation as in the Mediterranean region, without action to conserve them as suitable breeding habitats for waterbirds (Sripanomyom et al., 2011; Lei et al., 2018). Thus, action focused on improving the conservation of saltpans can help conserve avocet and other waterbirds in this flyway.
In conclusion, our findings showed that coastal Nanpu saltpans in the EAAF can provide an alternative suitable habitat for pied avocets. Flooding and a lack of high-quality breeding sites probably result in a high nest-abandonment rate, especially during the laying period, which highlights the importance of water level management as well as the construction of dikes or islets to avoid flooding. Such management actions are likely to be beneficial for the entire waterbird community nesting in saltpans. As the saltpans are continually losing along the coast of Asia and other continents, we suggest that both conservation action and research effort should be strengthened to understand and conserve this habitat and waterbirds.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
Ethical review and approval was not required for the animal study because the research is about wild bird, so we apply the permit from local government, Luannan County Forestry Bureau, HeBei, China.
Author Contributions
ZZ and WL designed the experiments. WL, YW, and FW conducted fieldwork. WL and YW analysed the data. WL led the writing with contributions from ZZ, JM, and TP. All authors read and approved the final version of the manuscript.
Funding
This research was supported by the National Natural Science Foundation of China (Grant Numbers 31801985 and 31830089), The Fundamental Research Funds for the Central Universities, the Grant from WWF-China Programme, and SEE Foundation.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank lovely volunteers and colleagues, Katarzyna Kucharska, Tong Mu, Jian Zhao, Bingrun Zhu, Cheng Chen, Jingyi Liu, Hengjun Xiao, Siyang Liu, Jason Loghry, and Peter Crighton for their great help. We also thank Yang Liu and his father for local logistic support. We also thank the two reviewers who provided valuable and insightful comments which greatly improved the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2021.622756/full#supplementary-material
References
Ackerman, J. T., and Eagles-Smith, C. A. (2010). Accuracy of egg flotation throughout incubation to determine embryo age and incubation day in waterbird nests. Condor 112, 438–446. doi: 10.1525/cond.2010.090070
Alberico, J. A. (1995). Floating eggs to estimate incubation stage does not affect hatchability. Wildl. Soc. Bull. 23, 212–216.
Ali Chokri, M., and Selmi, S. (2011). Predation of Pied Avocet Recurvirostra avosetta nests in a salina habitat: evidence for an edge effect. Bird Study 58, 171–177. doi: 10.1080/00063657.2010.546390
Arroyo, G. M., and Hortas, F. (2005). Status and population dynamics of Avocets (Recurvirostra avosetta) in Spain. Bull. Wader Study Group 107, 98–102.
Arroyo, G. M., Ossorio, J. A. M., Hurtado, A. P., and Casas, M. C. (1997). “Uso de salinas industriales como hábitats de reproducción por la cigüeñuela (Himantopus himantopus) y la avoceta (Recurvirostra avosetta) en el Parque Natural de la Bahía de Cádiz (SW de España),” in Proceedings of the Actas de las XII Jornadas Ornitológicas Españolas: Almerimar (El Ejido-Almería), 15 a 19 de septiembre, 1994, (Almerimar: Instituto de Estudios Almerienses), 165–179.
Beintema, A. J., and Muskens, G. (1987). Nesting success of birds breeding in Dutch agricultural grasslands. J. Appl. Ecol. 24, 743–758. doi: 10.2307/2403978
Bluso-Demers, J. D., Ackerman, J. T., Takekawa, J. Y., and Peterson, S. H. (2016). Habitat selection by Forster’s Terns (Sterna forsteri) at multiple spatial scales in an urbanized estuary: the importance of salt ponds. Waterbirds 39, 375–387. doi: 10.1675/063.039.0407
Britton, R. H., and Johnson, A. R. (1987). An ecological account of a Mediterranean salina: the Salin de Giraud, Camargue (S. France). Biol. Conserv. 42, 185–230. doi: 10.1016/0006-3207(87)90133-9
Burnham, K. P., and Anderson, D. R. (2004). Multimodel Inference: understanding AIC and BIC in Model Selection. Soc. Methods Res. 33, 261–304. doi: 10.1177/0049124104268644
Burns, S. J., Godfrey, L. R., Faina, P., McGee, D., Hardt, B., Ranivoharimanana, L., et al. (2016). Rapid human-induced landscape transformation in Madagascar at the end of the first millennium of the Common Era. Quat. Sci. Rev. 134, 92–99. doi: 10.1016/j.quascirev.2016.01.007
Cadbury, C. J., and Olney, P. J. S. (1978). Avocet population dynamics in England. Br. Birds 71, 102–121.
Catry, T., Ramos, J. A., Catry, I., Allen-Revez, M., and Grade, N. (2004). Are salinas a suitable alternative breeding habitat for Little Terns Sterna albifrons? IBIS 146, 247–257. doi: 10.1046/j.1474-919x.2004.00254.x
Chokri, M. A., and Selmi, S. (2011). Nesting ecology of pied avocet Recurvirostra avosetta in Sfax salina, Tunisia. Ostrich 82, 11–16. doi: 10.2989/00306525.2011.556789
Cloern, J. E., Abreu, P. C., Carstensen, J., Chauvaud, L., Elmgren, R., Grall, J., et al. (2016). Human activities and climate variability drive fast-paced change across the world’s estuarine–coastal ecosystems. Glob. Change Biol. 22, 513–529. doi: 10.1111/gcb.13059
Colwell, M. A., Meyer, J. J., Hardy, M. A., Mcallister, S. E., Transou, A. N., Levalley, R. R., et al. (2011). Western Snowy Plovers Charadrius alexandrinus nivosus select nesting substrates that enhance egg crypsis and improve nest survival. IBIS 153, 303–311. doi: 10.1111/j.1474-919x.2011.01100.x
Cooch, E., and White, G. (2006). Program MARK: A Gentle Introduction. Available online at: http://www.phidot.org/software/mark/docs/book
Cuervo, J. J. (2004). Nest-site selection and characteristics in a mixed-species colony of Avocets Recurvirostra avosetta and Black-winged Stilts Himantopus himantopus. Bird Study 51, 20–24. doi: 10.1080/00063650409461328
Cuervo, J. J. (2005). Hatching success in Avocet Recurvirostra avosetta and Black-winged Stilt Himantopus himantopus: capsule daily survival rate of nests was influenced by laying date and nesting period (laying versus incubation) in both species and by colony size in Black-winged Stilts. Bird Study 52, 166–172. doi: 10.1080/00063650509461387
De Bie, S., and Zijlstra, M. (1985). Kluten (Recurvitostra avosetta) en waterpeil in de Oostvaardersplassen: broeden in een veilige omgeving? Limosa 58, 41–48.
Engelmoer, M., and Blomert, A.-M. (1985). Broedbiologie van de Kluut Langs de Friese Waddenkust Seizoen 1983. Lelystad: Rijksdienst voor de IJsselmeerpolders.
Forbes, M. R., Clark, R. G., Weatherhead, P. J., and Armstrong, T. (1994). Risk-taking by female ducks: intra-and interspecific tests of nest defense theory. Behav. Ecol. Sociobiol. 34, 79–85. doi: 10.1007/BF00164178
Goutner, V. (1985). Breeding ecology of the avocet (Recurvirostra avosetta) in the Evros delta (Greece). Bonn. Zool. Beitr. 36, 37–50.
Green, J. M. H., Sripanomyom, S., Giam, X., and Wilcove, D. S. (2015). The ecology and economics of shorebird conservation in a tropical human-modified landscape. J. Appl. Ecol. 52, 1483–1491. doi: 10.1111/1365-2664.12508
Hötker, H. (1998). Intraspecific variation in length of incubation period in avocets Recurvirostra avosetta. Ardea 86, 33–41.
Hötker, H. (2000). Intraspecific variation in size and density of avocet colonies: effects of nest-distances on hatching and breeding success. J. Avian Biol. 31, 387–398. doi: 10.2307/3677421
Hötker, H., and Segebade, A. (2000). Effects of predation and weather on the breeding success of Avocets Recurvirostra avosetta. Bird Study 47, 91–101. doi: 10.1080/00063650009461163
Hötker, H., and West, R. (2005). Population size, population development and habitat use by Avocets in Western Europe at the end of the 20th century. Bull. Wader Study Group 107, 57–65.
Hu, Q., Wen, Y., Yu, G., Yin, J., Guan, H., Lv, L., et al. (2020). Research activity does not affect nest predation rates of the Silver-throated Tit, a passerine bird building domed nests. Avian Res. 11:28. doi: 10.1186/s40657-020-00214-9
Ibáñez-Álamo, J. D., Sanllorente, O., and Soler, M. (2012). The impact of researcher disturbance on nest predation rates: a meta-analysis. IBIS 154, 5–14. doi: 10.1111/j.1474-919x.2011.01186.x
Ibáñez-Álamo, J. D., and Soler, M. (2010). Investigator activities reduce nest predation in blackbirds Turdus merula. J. Avian Biol. 41, 208–212. doi: 10.1111/j.1600-048x.2009.04805.x
Jackson, M. V., Choi, C.-Y., Amano, T., Estrella, S. M., Lei, W., Moores, N., et al. (2020). Navigating coasts of concrete: pervasive use of artificial habitats by shorebirds in the Asia-Pacific. Biol. Conserv. 247:108591. doi: 10.1016/j.biocon.2020.108591
Kirby, J. S., Stattersfield, A. J., Butchart, S. H., Evans, M. I., Grimmett, R. F., Jones, V. R., et al. (2008). Key conservation issues for migratory land-and waterbird species on the world’s major flyways. Bird Conserv. Int. 18, S49–S73. doi: 10.1017/S0959270908000439
Knopf, F. L. (1982). Habitat requirements and breeding success of Charadriiform birds nesting at Salt Plains National Wildlife Refuge, Oklahoma. J. Field Ornithol. 53, 139–148.
Laake, J. L. (2013). “Capture-recapture analysis with hidden Markov models”, AFSC Processed Rep. 2013-04, Alaska Fish. Sci. Cent., NOAA, Natl. Mar. Fish. Serv., Seattle WA, 34.
Lei, W., Masero, J. A., Piersma, T., Zhu, B., Yang, H.-Y., and Zhang, Z. (2018). Alternative habitat: the importance of the Nanpu Saltpans for migratory waterbirds in the Chinese Yellow Sea. Bird Conserv. Int. 28, 549–566. doi: 10.1017/S0959270917000508
Lengyel, S. (2006). Spatial differences in breeding success in the pied avocet Recurvirostra avosetta: effects of habitat on hatching success and chick survival. J. Avian Biol. 37, 381–395. doi: 10.1111/j.0908-8857.2006.03501.x
Lotze, H. K., Reise, K., Worm, B., van Beusekom, J., Busch, M., Ehlers, A., et al. (2005). Human transformations of the Wadden Sea ecosystem through time: a synthesis. Helgol. Mar. Res. 59, 84–95. doi: 10.1007/s10152-004-0209-z
Mabee, T. J. (1997). Using eggshell evidence to determine nest fate of shorebirds. Wilson Bull. 109, 307–313.
MacDonald, M. A., and Bolton, M. (2008). Predation on wader nests in Europe. IBIS 150, 54–73. doi: 10.1111/j.1474-919X.2008.00869.x
MaCivor, L. H., Melvin, S. M., and Griffin, C. R. (1990). Effects of research activity on piping plover nest predation. J. Wildl. Manag. 54, 443–447. doi: 10.2307/3809656
Major, R. E. (1990). The effect of human observers on the intensity of nest predation. IBIS 132, 608–612. doi: 10.1111/j.1474-919x.1990.tb00285.x
Masero, J. A. (2003). Assessing alternative anthropogenic habitats for conserving waterbirds: salinas as buffer areas against the impact of natural habitat loss for shorebirds. Biodivers. Conserv. 12, 1157–1173. doi: 10.1023/A:1023021320448
Masero, J. A., and Pérez-Hurtado, A. (2001). Importance of the supratidal habitats for maintaining overwintering shorebird populations: how redshanks use tidal mudflats and adjacent saltworks in southern Europe. Condor 103, 21–30. doi: 10.1093/condor/103.1.21
Montgomerie, R. D., and Weatherhead, P. J. (1988). Risks and rewards of nest defence by parent birds. Q. Rev. Biol. 63, 167–187. doi: 10.1086/415838
Moynahan, B. J., Lindberg, M. S., Rotella, J. J., and Thomas, J. W. (2007). Factors affecting nest survival of greater sage-grouse in Northcentral Montana. J. Wildl. Manag. 71, 1773–1783. doi: 10.2193/2005-386
Murray, N. J., Clemens, R. S., Phinn, S. R., Possingham, H. P., and Fuller, R. A. (2014). Tracking the rapid loss of tidal wetlands in the Yellow Sea. Front. Ecol. Environ. 12:267–272. doi: 10.1890/130260
Nichols, J. D., Percival, H. F., Coon, R. A., Conroy, M. J., Hensler, G. L., and Hines, J. E. (1984). Observer visitation frequency and success of mourning dove nests: a field experiment. Auk 101, 398–402. doi: 10.1093/auk/101.2.384
Nogueira, H. C., Carrique, E. L., Romero, J. G., Ariza, J. N., and Aguilera, P. A. (1996). Management of Avocet breeding islands. Bull. Wader Study Group 81, 46–49.
Pierce, R. J. (2017). “Pied Avocet (Recurvirostra avosetta),” in Handbook of the Birds of the World Alive, eds J. del Hoyo, A. Elliott, J. Sargatal, D. A. Christie, and E. de Juana (Barcelona: Lynx Edicions).
Plaschke, S., Bulla, M., Cruz-López, M., Gómez del Ángel, S., and Küpper, C. (2019). Nest initiation and flooding in response to season and semi-lunar spring tides in a ground-nesting shorebird. Front. Zool. 16:15. doi: 10.1186/s12983-019-0313-1
Que, P. J., Chang, Y. J., Eberhart-Phillips, L., Liu, Y., Székely, T., and Zhang, Z. W. (2015). Low nest survival of a breeding shorebird in Bohai Bay. China. J. Ornithol. 156, 297–307. doi: 10.1007/s10336-014-1126-9
Ramírez, F., Navarro, J., Afán, I., Hobson, K. A., Delgado, A., and Forero, M. G. (2012). Adapting to a changing world: unraveling the role of man-made habitats as alternative feeding areas for slender-billed gull (Chroicocephalus genei). PLoS One 7:e47551. doi: 10.1371/journal.pone.0047551
Ricklefs, R. E. (1969). An analysis of nesting mortality in birds. Smithson. Contrib. Zool. 9, 1–48. doi: 10.5479/si.00810282.9
Rintoul, C., Warnock, N., Page, G. W., and Hanson, J. T. (2003). Breeding status and habitat use of black-necked stilts and American avocets in South San Francisco Bay. West. Birds 34, 2–14.
Robinson, C. W. (2008). Western Snowy Plover Use of Managed Salt Ponds at Eden Landing, Hayward, CA. Master’s theses, San Jose, CA: San Jose State University.
Rocha, A., Fonseca, D., Masero, J. A., and Ramos, J. A. (2016). Coastal saltpans are a good alternative breeding habitat for Kentish plover Charadrius alexandrinus when umbrella species are present. J. Avian Biol. 47, 824–833. doi: 10.1111/jav.00883
Roche, E. A., Arnold, T. W., and Cuthbert, F. J. (2010). Apparent nest abandonment as evidence of breeding-season mortality in great lakes piping plovers (Charadrius melodus)abandono aparente de nidos como evidencia de mortalidad durante la estacion reproductiva en charadrius melodus. Auk 127, 402–410. doi: 10.1525/auk.2009.09034
Rotella, J. J., Dinsmore, S. J., and Shaffer, T. L. (2004). Modeling nest–survival data: a comparison of recently developed methods that can be implemented in MARK and SAS. Anim. Biodivers. Conserv. 27, 187–205.
Rotellam, J. J., Taper, M. L., and Hansen, A. J. (2000). Correcting nesting-success estimates for observer effects: maximum-likelihood estimates of daily survival rates with reduced bias. Auk 117, 92–109. doi: 10.1093/auk/117.1.92
Seber, G. A. F. (1982). The Estimation of Animal Abundance and Related Parameters. New York, NY: MacMillan Press.
Sexson, M. G., and Farley, G. H. (2012). Snowy plover nest survival in Kansas and effective management to counter negative effects of precipitation. J. Wildl. Manag 76, 1587–1596. doi: 10.1002/jwmg.405
Škornik, I. (2019). 37 years of common tern sterna hirundo breeding at Sečovlje Salina. Acrocephalus 40, 105–112. doi: 10.1515/acro-2019-0008
Smith, P. A., Gilchrist, H. G., Forbes, M. R., Martin, J.-L., and Allard, K. (2010). Inter-annual variation in the breeding chronology of arctic shorebirds: effects of weather, snow melt and predators. J. Avian Biol. 41, 292–304. doi: 10.1111/j.1600-048X.2009.04815.x
Smith, P. A., and Wilson, S. (2010). Intraseasonal patterns in shorebird nest survival are related to nest age and defence behaviour. Oecologia 163, 613–624. doi: 10.1007/s00442-010-1644-y
Sripanomyom, S., Round, P. D., Savini, T., Trisurat, Y., and Gale, G. A. (2011). Traditional salt-pans hold major concentrations of overwintering shorebirds in Southeast Asia. Biol. Conserv. 144, 526–537.
Studds, C. E., Kendall, B. E., Murray, N. J., Wilson, H. B., Rogers, D. I., Clemens, R. S., et al. (2017). Rapid population decline in migratory shorebirds relying on Yellow Sea tidal mudflats as stopover sites. Nat. Commun. 8:14895. doi: 10.1038/ncomms14895
Takekawa, J. Y., Lu, C. T., and Pratt, R. T. (2001). Avian communities in baylands and artificial salt evaporation ponds of the San Francisco Bay estuary. Hydrobiologia 466, 317–328.
Thyen, S. (2005). Reproduction of Coastal Birds Breeding in the Wadden Sea: Variation, Influencing Factors and Monitoring (PhD Thesis). Oldenburg: Universität Oldenburg.
Uher-Koch, B. D., Schmutz, J. A., and Wright, K. G. (2015). Nest visits and capture events affect breeding success of Yellow-billed and Pacific loons. Condor 117, 121–129.
UNEP (2006). Marine and Coastal Ecosystems and Human Well-Being: A Synthesis Report Based on the Findings of the Millennium Ecosystem Assessment. Nairobi: UNEP.
Vitousek, P. M., Mooney, H. A., Lubchenco, J., and Melillo, J. M. (1997). Human domination of Earth’s ecosystems. Science 277, 494–499. doi: 10.1126/science.277.5325.494
Walker, J., Lindberg, M. S., MacCluskie, M. C., Petrula, M. J., and Sedinger, J. S. (2005). Nest survival of scaup and other ducks in the boreal forest of Alaska. J. Wildl. Manag. 69, 582–591.
Westmoreland, D., and Best, L. B. (1985). The effect of disturbance on mourning dove nesting success. Auk 102, 774–780. doi: 10.1093/auk/102.4.774
White, G. C., and Burnham, K. P. (1999). Program MARK: survival estimation from populations of marked animals. Bird Study 46, 120–138.
Winton, B. R., Leslie, D. M., and Rupert, J. R. (2000). Breeding ecology and management of snowy plovers in north-central oklahoma. J. Field Ornithol. 71, 573–584. doi: 10.1648/0273-8570-71.4.573
Worm, B., Barbier, E. B., Beaumont, N., Duffy, J. E., Folke, C., Halpern, B. S., et al. (2006). Impacts of biodiversity loss on ocean ecosystem services. Science 314, 787–790. doi: 10.1126/science.1132294
Wu, F., Lei, W., Lloyd, H., and Zhang, Z. (2020). Predictors of Gull-billed tern (Gelochelidon nilotica) nest survival in artificial coastal saltpans, Bohai Bay, China. PeerJ 8:e10054. doi: 10.7717/peerj.10054
Yang, H.-Y. (2006). Studies on Ecology of Shorebirds in the Coastal Area of Northern Bohai Bay. Beijing: Beijing Normal University.
Yang, H. Y., Chen, B., Barter, M., Piersma, T., Zhou, C. F., Li, F. S., et al. (2011). Impacts of tidal land reclamation in Bohai Bay, China: ongoing losses of critical Yellow Sea waterbird staging and wintering sites. Bird Conserv. Int. 21, 241–259. doi: 10.1017/S0959270911000086
Zeileis, A., Cribari-Neto, F., Gruen, B., Kosmidis, I., Simas, A. B., Rocha, A. V., et al. (2016). Package ‘betareg.’. R Package 3, 2.
Keywords: Bohai Bay, breeding, nest success, waterbirds, Yellow Sea
Citation: Lei W, Wu Y, Wu F, Piersma T, Zhang Z and Masero JA (2021) Artificial Wetlands as Breeding Habitats for Shorebirds: A Case Study on Pied Avocets in China’s Largest Saltpan Complex. Front. Ecol. Evol. 9:622756. doi: 10.3389/fevo.2021.622756
Received: 29 October 2020; Accepted: 22 March 2021;
Published: 25 May 2021.
Edited by:
Binbin Li, Duke Kunshan University, ChinaReviewed by:
Attila D. Sándor, University of Agricultural Sciences and Veterinary Medicine of Cluj-Napoca, RomaniaXingfeng Si, East China Normal University, China
Copyright © 2021 Lei, Wu, Wu, Piersma, Zhang and Masero. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zhengwang Zhang, enp3QGJudS5lZHUuY24=