 Michelle Nordkvist
Michelle Nordkvist Christer Björkman
Christer Björkman Maartje J. Klapwijk
Maartje J. Klapwijk- Department of Ecology, Swedish University of Agricultural Sciences, Uppsala, Sweden
Insect herbivore performance and arthropod communities can be affected by mammalian grazing and browsing via altered plant communities and vegetation structure. Far less is known about whether changes to plant architecture can cause similar effects. Browsing generated changes to within plant architecture could potentially have large consequences for arthropod communities, herbivore survival and eventually damage to plants. This study investigates plant-mediated effects of ungulate browsing on arthropod predator communities and on the survival of herbivorous insects. More specifically we studied how different levels of ungulate browsing (1) influenced the arthropod predator community on Scots pine (Pinus sylvestris) and (2) affected the survival of the European pine sawfly (Neodiprion sertifer). We related these response variables to browsing-inflicted changes in pine architecture. An observational study of generalist arthropod predators on pine trees revealed a trend toward a quadratic response of ants to browsing intensity—i.e., a higher abundance of ants on moderately browsed trees and lower abundance on intensively browsed trees. A field survey of sawfly larvae revealed a 19% lower larval survival on browsed compared to un-browsed pines, but no difference in survival comparing pines with moderate and high intensity of browsing. A structural equation model revealed that moose generated changes to pine architecture had only a small effect on sawfly larval survival, suggesting additional mediating pine traits affected by browsing. We conclude that insect survival can be altered by ungulate browsing, which could affect damage levels.
Introduction
Ungulates can have a strong effect on forest composition and structure (McInnes et al., 1992; Martin et al., 2010). Research has shown that intense grazing of forest understory vegetation will lead to decreased understory height as well as changes in understory composition in boreal forest (Speed et al., 2014; Lorentzen Kolstad et al., 2018). Other studies investigating the subsequent effects of browsing on arthropod communities observed a change in both diversity and composition (Suominen et al., 1999; Landsman and Bowman, 2017; Lilleeng et al., 2018). One consequence of an altered predator community is subsequent effects on predation rates of insect herbivores, and plant damage levels. The effect of grazing on understory vegetation and arthropod communities is rather well studied (Suominen et al., 2008; Landsman and Bowman, 2017; Lilleeng et al., 2018). Similarly, the structural complexity of individual plants (plant architecture) can influence both predator communities (Langellotto and Denno, 2004) and predation rates (Riihimäki et al., 2006; Šipoš and Kindlmann, 2012). Browsing has been demonstrated to affect the architecture of trees (Danell et al., 1994), and such changes could influence the community of arthropod predators (Danell and Huss-Danell, 1985) and/or their behavior. Browsing induced changes in top-down pressure could subsequently lead to altered survival of herbivorous insects. Browsing has previously been demonstrated to increase damage levels of insects on birch (Muiruri et al., 2015), but the indirect effect of browsing on herbivore survival mediated through architecture is largely unexplored.
Plant architecture can be defined as the structure and arrangement of plant parts and refers to characteristics such as size, growth form, and variety of above-ground parts (Lawton, 1983). Similar to structure of larger scale habitats, architecture of individual plants can impact animal communities, behaviors and interactions (Lawton, 1983; Langellotto and Denno, 2004; Reynolds and Cuddington, 2012a,b). Increased architectural complexity has been shown to primarily lead to higher natural enemy abundance and/or predation on plants (Halaj et al., 2000; Kaitaniemi et al., 2004; Langellotto and Denno, 2004; Reynolds and Cuddington, 2012a) and the opposite has been found for plants with decreased architectural complexity (Langellotto and Denno, 2004). Although in some cases high complexity has been demonstrated to decrease predation (Šipoš and Kindlmann, 2012), possibly depending on the specific species involved (Kaitaniemi et al., 2004; Langellotto and Denno, 2004; Šipoš and Kindlmann, 2012). Other aspects that are affected by plant architecture are predator movement (Cuddington and Yodzis, 2002) and behavior (Reynolds and Cuddington, 2012b), for example searching time and paths are found to be affected by plant morphology (Reynolds and Cuddington, 2012b). Structurally complex plants are also found to host a more diverse herbivore community (Lawton, 1983), which in turn can lead to higher diversity or abundance of predators (Forkner and Hunter, 2000). Changes to abundance, diversity, composition and behavior of natural enemies, via altered plant architecture, could thus influence the predation rate exerted on herbivorous insects and thereby their survival.
Although it is well established that ungulate browsing alters tree shape and growth (Wallgren et al., 2014), very little is known about the effect of such changes on tree-dwelling arthropods and subsequent effects on predation rates. Examples of browsing inflicted changes that could affect arthropods are diminished amount of biomass and retarded growth (Danell et al., 1994). Browsing is, in addition, often patchy within the tree (Danell et al., 1994) sometimes creating high variation of structures. Such changes in tree architecture can reduce the amount of suitable habitat available for arthropods but also increase the amount of habitat variation and complexity. Moreover, intensity of browsing is an important determinant of architectural features (Edenius et al., 1993; Danell et al., 1994), and may thus be important for arthropods. On birch, an increasing intensity of browsing (from slight to moderate) has been shown to increase abundance of ants (Danell and Huss-Danell, 1985), which are important generalist predators. Increasing intensity of browsing has also been demonstrated to increase damage level of herbivorous insects (Muiruri et al., 2015).
Mammalian browsing also influences plant quality (Nykänen and Koricheva, 2004), which could affect insect abundance (Den Herder et al., 2009), survival (Martinsen et al., 1998), and damage levels (Lind et al., 2012). A positive effect of browsing on nutritional quality could increase the abundance or quality of herbivorous insects and consequently increase the abundance of natural enemies (Forkner and Hunter, 2000). We believe that plant architecture and nutritional quality could jointly influence predation rates, and as a result herbivore survival (Kaitaniemi et al., 2004). Therefore, we set out to quantify the effect of browsing on tree architecture and nutritional quality and its subsequent effect on predator presence and herbivore survival.
Our study system consisted of Scots pine (Pinus sylvestris) browsed by moose (Alces alces) and the European pine sawfly (Neodiprion sertifer). We formulated the following research questions: (1) How does the occurrence and different intensities of moose browsing affect arthropod predator community variables, i.e., abundance, presence/absence, richness, and diversity? (2) How does pine architecture and needle C:N ratio differ depending on the occurrence and intensity of moose browsing? (3) How does the occurrence and intensity of browsing affect sawfly larvae survival, and (4) How does pine architecture and needle C:N ratio relate to sawfly survival rates?
Materials and Methods
Study Design
To achieve our aims, we conducted two studies. The first study (study 1), an observational study of the natural enemy community on browsed and un-browsed trees will answer question 1. The design of the second study (study 2) is based on the results from the first study and aimed to answer questions 2, 3, and 4. In study 2 we survey sawfly survival on browsed trees (in the field), and related it to browsing induced changes in pine architecture, as well as pine needle C:N ratio.
Study 1–Arthropod Predator Community
Experimental Set Up
The study was conducted between 20th of May and 10th of June in2017, in 14 sites in Uppland, Sweden (Supplementary Table 1). Within those sites, a total of 119 trees were randomly chosen, half of them browsed and half of them un-browsed. We specifically selected browsed trees that had been browsed the previous winter. There were generally more un-browsed than browsed trees within the sites, but the average level of browsing on stand scale was rather high, thus there was no shortage of browsed trees. Each site was a pine dominated forest stand, with pines of similar height (range of surveyed pines 130–170 cm). The intensity of browsing was recorded for the browsed trees and estimated to 25, 50, 75, and 100% of last year’s lateral shoots browsed. All trees were surveyed for arthropod natural enemies (e.g., spiders, ants, and lady beetles), using the beating tray method (Leather 2008). All predatory arthropods found on the beating tray were recorded and predators belonging to the orders Hymenoptera (exclusively family Formicate), Araneae and Coleoptera were found. Since the samples were dominated by ants and spiders (98% of the samples) and since these two groups are important predators for sawfly larvae (Olofsson, 1992; Lindstedt et al., 2006; Kaitaniemi et al., 2007), analyses were carried out only on these two groups. We surveyed branches at a height of 1–1.5 m. To be able to have comparable measures of abundance we surveyed the same amount of biomass on all trees approximated as 1.5 m length of green biomass, i.e., we surveyed branches adding up to a length of 1.5 m in total. The total number of arthropod natural enemies and the total number of morphospecies were counted directly, in the field. Similarly, number of individuals and number of species were counted within groups (ants, spiders etc.). This gave measures of abundance and community composition, i.e., presence/absence of different species groups as well as species richness and diversity.
Statistical Analysis
All analyses were performed in R version 3.6.2 (R Core Team, 2019). The lmer, glmer and glmer.nb functions in the lme4 package (Bates et al., 2015) were used to fit mixed effects models. Model results were calculated using the car package (Fox and Weisberg, 2019). In all models site was entered as a random effect and intensity of browsing as a fixed effect (five levels; un-browsed, 25, 50, 75, and 100% of the exterior shoots browsed). Abundance (total and within species groups) was analyzed with a generalized linear mixed model with negative binomial distribution, since the data was overdispersed. Richness (number of species) was analyzed with a generalized linear mixed model with Poisson distribution. Presence/absence data of species groups were analyzed using generalized linear mixed models with binomial distribution. Species diversity (Simpson’s index) was analyzed using a linear mixed model. We removed seven trees from the analysis of ant abundance due to them being outliers (extremely high values of ants, possibly due to close proximity to ant hills). The result from the initial analysis with all trees are presented in the supplementary material (Supplementary Table 7).
Study 2–Herbivore Survival
Study System and Current Knowledge
The European pine sawfly (Neodiprion sertifer) is a pine specialist herbivore. It is an outbreak species, and can be a severe pest in pine forest stands. Larvae feed gregariously on all pine needles except current year’s foliage in early summer, and group size is between 20 and 60 individuals (Gur’yanova, 2006). Pine sawfly larvae feed gregariously on the branch on which they hatched and normally only move to another nearby branch if there is a shortage of food. Therefore, the larvae are not expected to be influenced by pine architecture, but their predators could be, hence potentially altering sawfly survival. Sawflies are subjected to predation in their larval stage by various generalists, mainly ants (Olofsson, 1992; Lindstedt et al., 2006; Kaitaniemi et al., 2007) and spiders (Juutinen, 1967; Kaitaniemi et al., 2007). Sawfly performance is commonly related to pine needle nitrogen (Björkman et al., 1991, 1997) and diterpene content (Larsson et al., 1986; Björkman et al., 1997), but moose browsing has previously been shown not to influence sawfly bottom-up survival (Nordkvist et al., 2019). Predation of sawfly larvae is commonly related to pine needle diterpene content, due to sequestering and related anti-predator behavior by sawfly larvae (Lindstedt et al., 2006; Kollberg et al., 2015), but moose browsing has previously been shown not to affect diterpene content of pine needles (Nordkvist et al., 2019).
Experimental Set Up
Based on the results from the observational study we explored herbivore survival, using pine sawfly larvae, on browsed pines. The pines were either un-browsed, moderately browsed (50% of exterior lateral shoots browsed) or intensively browsed (100% of exterior lateral shoots browsed). The survey was conducted in 2018 in two field sites in Uppsala, Sweden (site 1: 59 52 01.7N, 18 11 06.4E, site 2: 59 58 00.9N, 18 13 37.0E). The dominant tree species in both sites is Scots pine. On each site, pines were selected in groups of three, one un-browsed, one moderately browsed and one intensively browsed. The selected pines had been browsed the previous winter. Each group consisted of pines growing in similar conditions to minimize effects of micro-climate and soil properties (these groups are henceforth referred to as blocks). Ten blocks in site 1 and six blocks in site 2 were set up, resulting in 48 trees in total. The rationale for using naturally browsed trees rather than clipping trees was that (i) natural browsing incorporates moose specific factors (such as saliva) which can induce responses that are not induced on clipped trees (Ohse et al., 2017) and (ii) the aim was to assess how browsing affects pine architecture rather than manipulating architecture. Needles were sampled from each tree for C:N ratio analysis. Six architecture variables were recorded and quantified on all trees (Table 1). Correlation between the individual architecture variables as well as C:N ratio are presented in Supplementary Table 2 (cor.test function; R package stats; R Core Team, 2019). The variables were chosen based on their potential effect on predator numbers (Halaj et al., 2000; Langellotto and Denno, 2004), predator behavior (Cuddington and Yodzis, 2002; Reynolds and Cuddington, 2012b) and herbivore numbers (Lawton, 1983) in combination with them likely being affected by browsing (Danell et al., 1994).

Table 1. Architectural variables measured on the experimental pines.
The sawfly larvae used for the survey were collected in May 2018 from an outbreak area around Ramnäs, Sweden (59 46 39.98N, 16 05 49.18E). Larvae were stored briefly in 5°C dark room, and then randomly assorted into groups to avoid maternal effects, related to performance like body size but also vertical transmission of Nuclear Polyhedrosis Virus (Olofsson, 1989). Thirty 2nd instar sawfly larvae were placed on each tree on the 24th of May 2018 and groups were thereafter monitored every second day to ensure that the larvae were feeding and to keep track of their growth. One group of larvae were placed on each tree. Groups were always placed on browsed branches and on the same whorl on all trees (third from the top). After 14 days the remaining number of larvae were counted. Larvae had not reached final instar at the end of the 2 weeks (all larvae had reached 3rd instar), no larvae had pupated and there was no apparent difference in growth rate between the groups. Survival was assessed at group level as the proportion of larvae remaining at the end of the survey and number of larvae at the start of the survey. Survival affected only by bottom-up processes was not assessed separately but has been previously shown not to differ between browsed and un-browsed trees (Nordkvist et al., 2019). On five trees larval groups failed to establish properly and these trees were removed from the analysis, resulting in a final sample of 43 trees.
C:N Ratio Analysis
To analyze nitrogen and carbon content of needles, needle samples were dried at 70°C and thereafter ground. An elemental analyzer vario EL CNS (Elementar Analysensysteme GmbH, Elementar−Strasse 1, D−63505, Langenselbold, Germany) was used to analyze total nitrogen and carbon (% dry weight).
Statistical Analysis
Analysis of architectural data
To determine whether the intensity of browsing affected overall pine architecture we performed a principal component analysis (prcomp() function; R package stats; R Core Team, 2019). Principal component analysis has been used in a similar manner to analyze the effect of habitat diversity on leaf traits (Muiruri et al., 2019). To assess if the measured tree variables (architecture variables and C:N ratio) contributed to the effect of browsing on tree architecture, we used multivariate analysis of variance (manova() function; R package stats; R Core Team, 2019); all architecture variables (Table 1) as well as C:N ratio were used as response variables, and site, block and browsing were entered as explanatory variables.
Analysis of sawfly survival and relation to architecture
To test whether survival differed between the different intensities of browsing we used a linear mixed model with binomial distribution. The browsing intensities and site was used as fixed factors and block as random factor. To tease apart which pine variables (architecture and C:N ratio) differed between the intensities of browsing and affected survival we used piecewise structural equation modeling (psem; piecewiseSEM package; Lefcheck, 2016). Piecewise structural equation modeling allows for simultaneous assessment of multiple potential relationships within complex networks of factors and effects (Lefcheck, 2016). The structural equation model included sawfly survival as a response to browsing, the architecture variables and C:N ratio. Sawfly survival was measured as proportion, but in the piecewiseSEM package the use of a binomial error structure for proportion data has not yet been implemented, therefore we applied arc-sine square-root transformation of survival prior to analysis. First, the simplest structural equation model was modeled (Supplementary Figure 1). Based on the test of the directed separation, significant direct relationships were added to the model either as direct relationships or covariance structure. We added browsing as a direct relationship with survival, and the others as covariance structure.
Results
Study 1–Arthropod Predator Community
The predator community was dominated by ants and spiders (98% of the total sample). On average five predators were found per 1.5 m sampled biomass per tree. There was a significant difference in abundance of ants between the different intensities of browsing (p = 0.05; Figure 1A and Table 2A). The highest abundance of ants was found on 50% browsed trees and lowest on 100% browsed trees (Figure 1). There was no difference in the number of spiders (Figure 1B and Table 2B). There was no difference in presence/absence (ants: Df = 2, χ2 = 3.7, p = 0.16, spiders: Df = 2, χ2 = 4.4, p = 0.11), species richness (Df = 2, χ2 = 2.5, p = 0.29) or diversity (Df = 2, χ2 = 2.4, p = 0.30) between the intensities of browsing.
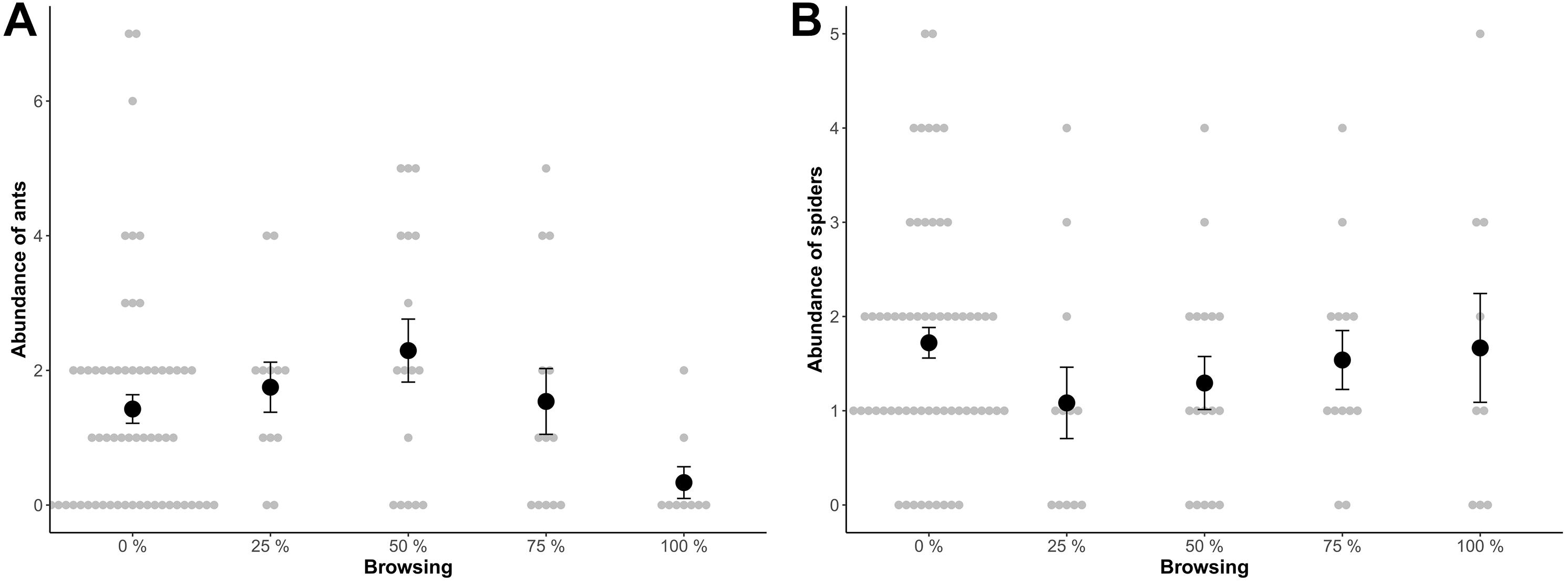
Figure 1. (A) Abundance of ants and (B) abundance of spiders in relation to browsing intensity. Mean abundances were: (A) 1.43, 1.75, 2.29, 1.54, and 0.33 ants per 1.5 m sampled biomass per tree and (B) 1.76, 1.08, 1.22, 1.5, 1.7, spiders per 1.5 m samples biomass on un-browsed (control), 25, 50, 75, and 100% browsed trees, respectively. Error bars show standard error.
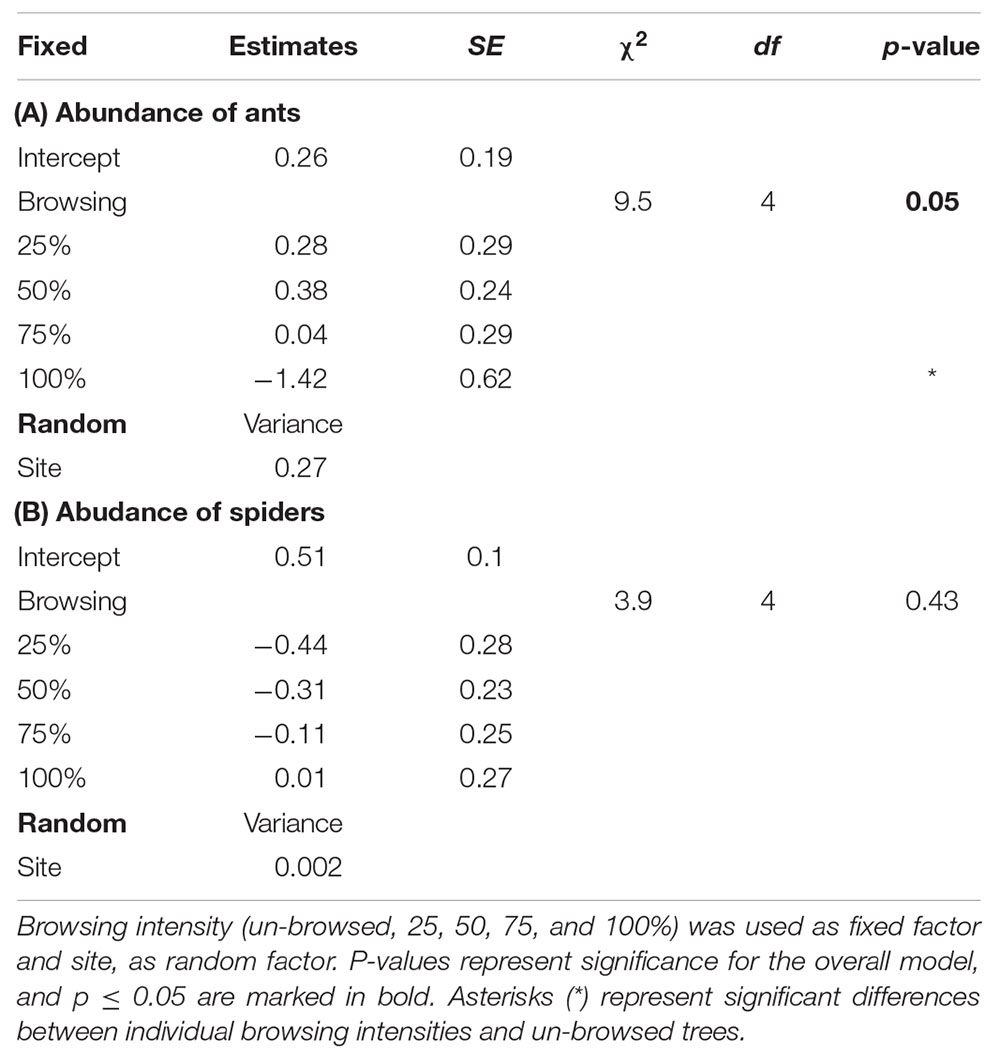
Table 2. ANOVA (type II test) and summary table for generalized linear mixed effects models testing the difference in abundance of (a) ants and (b) spiders, in relation to browsing intensity.
Study 2-Herbivore Survival
There was a difference in overall architecture between the three levels of browsing (Manova results: p < 0.001, Df = 2, F = 12.9; Figure 2 and Supplementary Tables 3, 4). For the principal component analysis, the first two axes together explain 68% of the variance, and there was a clear separation between the trees from the different browsing intensities (Figure 2). The importance of each individual architecture variable to principal component 1 and 2 are presented in Supplementary Table 3. According to the result from the Manova analysis five out of six individual architectural variables contributed to the overall differences among browsing levels (height, damaged whorls, branches, shoot units and growth abnormalities) while number of branches without needles did not, nor did needle C:N ratio (Supplementary Table 4). Browsed trees generally had fewer branches and fewer shoots compared to un-browsed trees (Figure 2 and Supplementary Table 4) and the difference between browsed and un-browsed trees was bigger when the intensity of browsing was higher (Figure 2). Intensively browsed trees had a higher amount of growth abnormalities, fewer shoots and branches, and the browsing damage was more dispersed compared to moderately browsed trees (Figure 2 and Supplementary Table 4).
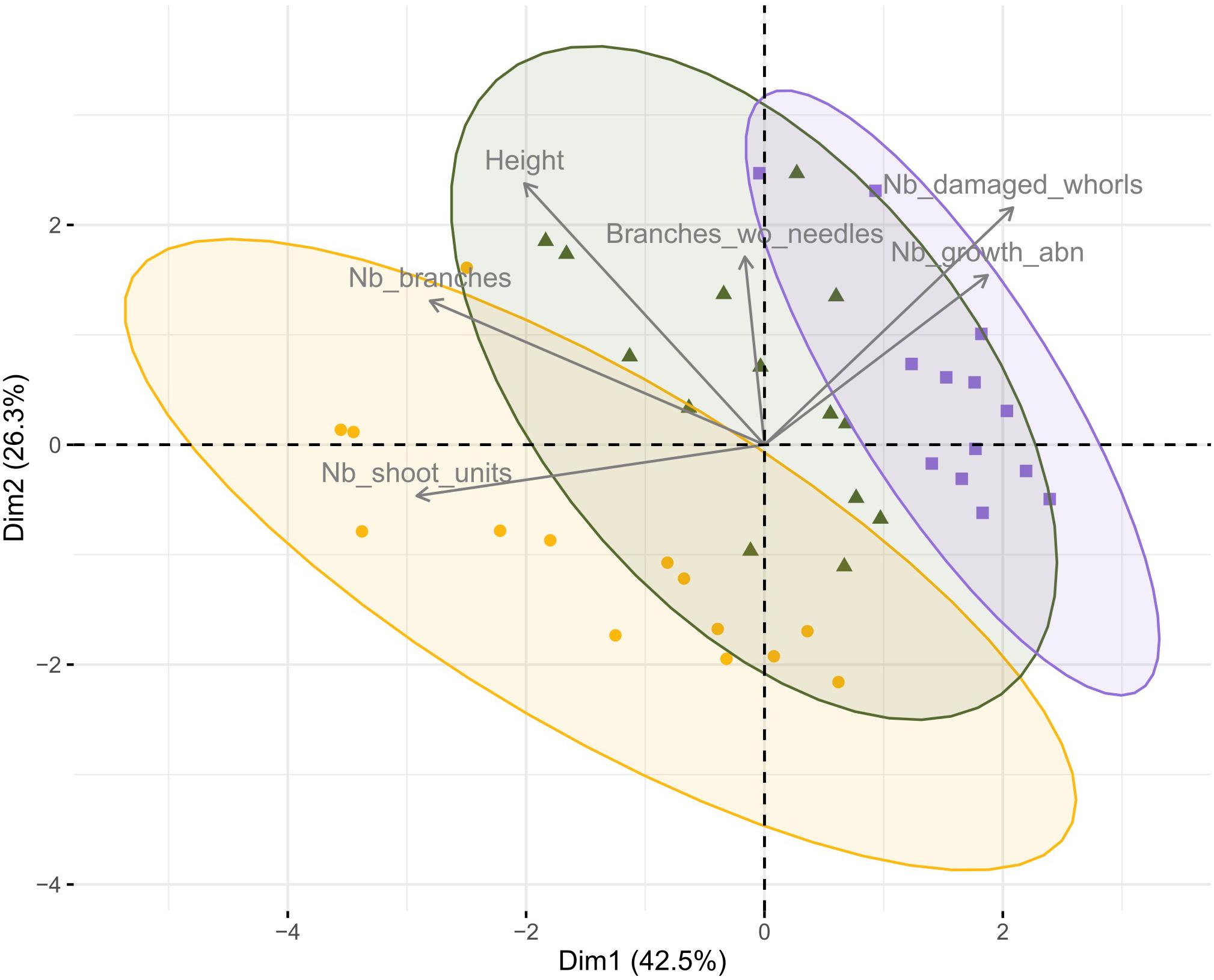
Figure 2. Principal component analysis of pine architecture variables (Table 1). The plot shows the first and second axis, which together explained 68% of the variance. Arrows (and corresponding names) represent the architectural traits (Nb_shoot_units = number of shoot units, Nb_branches = number of branches, Height = tree height, Branches_wo_needles = branches without needles, Nb_damaged_whorls = number of damaged whorls and Nb_growth_abn = number of growth abnormalities). Dots correspond to the individual trees. Shape and color represent the different levels of browsing (orange circles = control, green triangles = moderately browsed, blue squares = intensively browsed). Ellipses are drawn at the 95% confidence level according to browsing intensity.
Sawfly survival was lower on browsed trees compared to un-browsed trees but did not differ significantly between the two levels of browsing (Df = 2, χ2 = 15, p < 0.001; Figure 3 and Table 3). On average 50% of the larvae survived on un-browsed trees whereas on average 43% and 35% of the larvae survived on 50 and 100% browsed trees, respectively. The complete results of the piecewise structural equation model are presented in Figure 4 and Supplementary Material (summary statistics: Supplementary Table 5, initial model Supplementary Figure 1 and R2-values Supplementary Table 6). The path model showed that browsing affected five out of six architecture variables (all except height and branches without needles; Figure 4 and Supplementary Table 5). Survival was affected by four architecture variables; number of shoot units, damaged whorls, branches without needles and tree height. Tree height was the single most important measured pine variable determining survival (Figure 4), and survival decreased with tree height. C:N ratio was not affected by browsing nor did it affect survival. Browsing had a small but significant indirect effect on sawfly survival that can be attributed to tree architecture, through alteration of the number of shoot units and damaged whorls (Figure 4). Neither number of growth abnormalities nor number of branches affected survival. There was also a direct negative effect of browsing on survival (i.e., not explained by any of the measured pine variables; Figure 4).
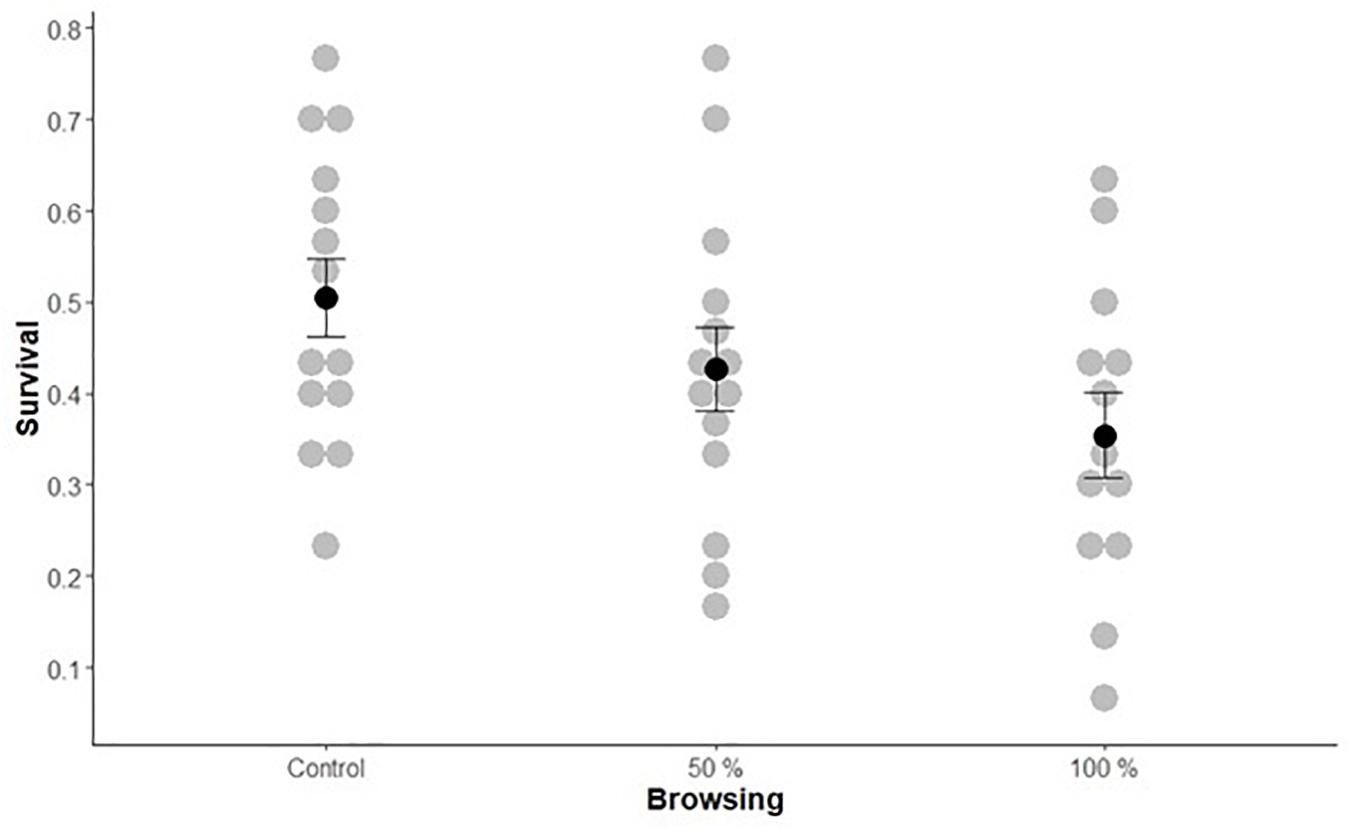
Figure 3. Survival of larvae on un-browsed, moderately and intensively browsed trees. Mean survival was 51, 43, and 35%. Gray dots represent survival on individual trees, and the black dots represent mean survival for each browsing intensity. Error bars represent standard error.
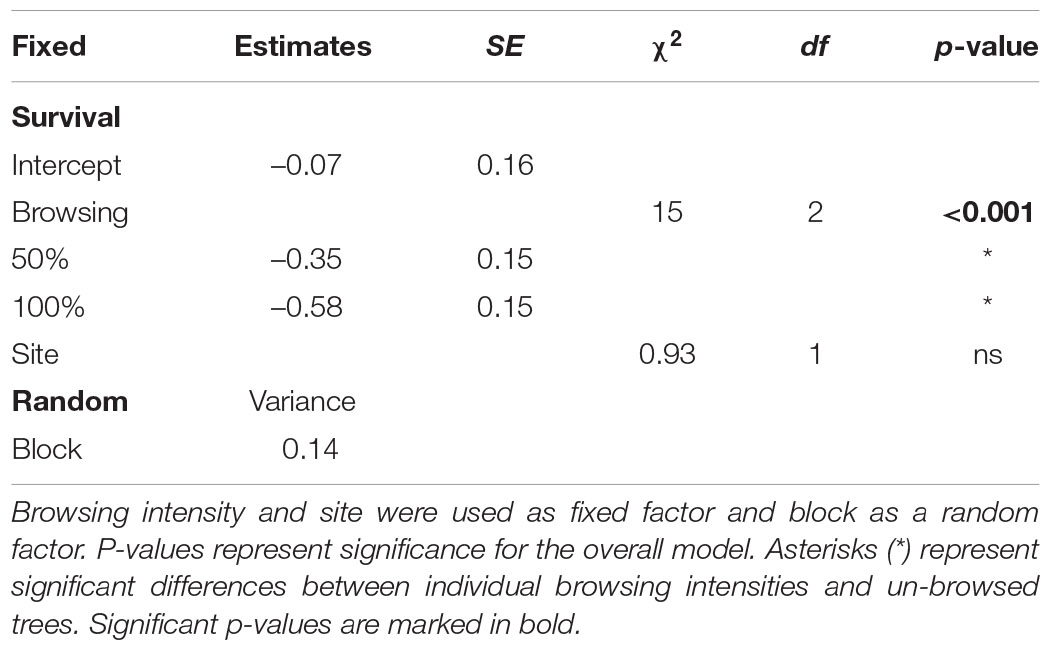
Table 3. ANOVA (type II test) and summary table for generalized linear mixed effects models testing the difference in sawfly survival in relation to browsing intensity (un-browsed, 50 and 100%).
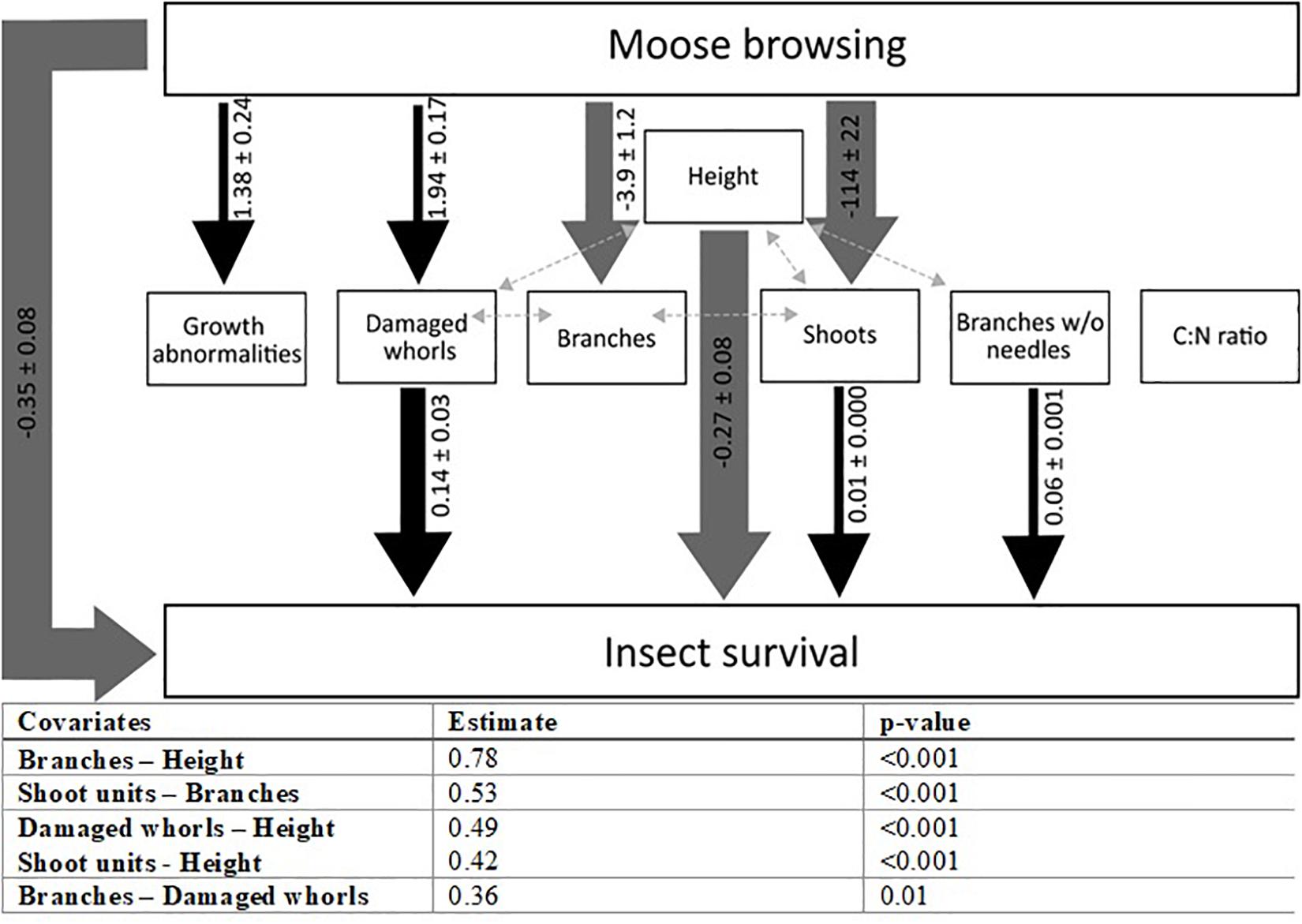
Figure 4. Illustration of the final structural equation model showing significant direct and indirect effects of moose browsing on insect survival as well as co-variances. Significant positive and negative relationships are shown with black and gray arrows, respectively. Path coefficients and standard error are indicated on or beside the arrows, and the thickness of the arrow correspond to the magnitude of those coefficients. Light gray dashed double-headed arrows denote significant co-variances between variables. The level of co-variance per combination is provided in the box below. Summary statistics for the initial and final model is provided in Supplementary Table 5 and marginal and conditional R2-values are provided in Supplementary Table 6.
Discussion
We demonstrate that ungulate browsing can alter the survival of herbivorous insects through plant-mediated effects, on tree level. We observe reduced survival on browsed trees vs. non-browsed trees but the effect can only partly be attributed to changes in tree architecture caused by browsing.
Study 1–Arthropod Predator Community
Our results demonstrate a correlation between browsing intensity and ant abundance. Ant abundance increase with intermediate levels of browsing but decreased with high levels of browsing and on non-browsed trees (Figure 1A and Table 2A), which is in line with a previous study conducted on birch (Danell and Huss-Danell, 1985). Ants are important generalist predators, and thus browsing could affect predation rate of several different herbivores, including sawflies (Lindstedt et al., 2006). Browsing on birch by reindeer has been shown to increase aphid abundance (Den Herder et al., 2009), which could be one of the mechanisms causing increased abundance of ants. Conversely, the abundance of spiders showed a trend toward decreasing with intermediate levels of browsing and increasing at high levels of browsing and on non-browsed trees (Figure 1). The opposite response of spiders and ants to browsing intensity might be a result of the positive effect of browsing on ants in combination with negative interference between ants and spiders (Halaj et al., 1997; Kaitaniemi et al., 2007). Ants also recruit conspecifics to patches of high quality (such as trees with high abundance of prey), which could increase the proportional differences due to e.g., interference. A change like this in the composition of predator community as a result of browsing could in turn change the herbivore community. Ants and spiders often have different predation strategies and behavior thus a shift from ants to spiders and vice versa could result in a shift of prey targeted. By only considering ants and spiders as broad groups (i.e., not determining species identities of the samples ants and spiders), we may have missed important differences in spider and ant community composition as a result of browsing. Knowledge of species identities could have provided a deeper insight to how and why browsing alters predator communities and which implications that might have for predation rates. Another caveat is that by using naturally browsed trees, rather than clipping trees to simulate browsing, we cannot be certain that the different intensities of browsing and the differences in ant abundance is not a result of another unmeasured variable. Still, the effect on herbivorous prey like the sawflies remain irrespective of the causes behind the patterns. The results presented here provides an interesting starting point to continue to study how browsing can influence predator communities at tree level.
Study 2–Herbivore Survival
Overall, survival of sawfly larvae was lower on browsed compared to un-browsed trees, but no significant difference between moderately and intensively browsed trees was observed (Figure 3 and Table 3). The measured architecture variables only partly explained the effect on survival of sawflies (Figure 4). The most important measured pine variable was tree height (Figure 4), and survival of sawfly larvae decreased with tree height. Taller trees may simply harbor more predators (Campos et al., 2006), increasing predation rate on the sawflies. We hypothesized that browsing would decrease needle C:N ratio (Nykänen and Koricheva, 2004) and that such changes would influence predation, and thus survival, through increased abundance or quality of alternative prey (Forkner and Hunter, 2000). However, we found that C:N ratio did not affect survival, nor was it affected by browsing (Figure 4).
The main effect of browsing on sawfly survival was negative and not mediated by any of the measured plant variables (Figure 4). Therefore, we draw the conclusion that survival is affected by plant traits affected by browsing other than the ones measured in our study. This could either be plant quality traits (other than C:N ratio) or additional architecture traits. Sawfly larvae sequester di-terpenoids which they use as defense against predators, and therefore survival of larvae can be higher on trees with high needle di-terpene content (Björkman et al., 1991). Pines with low needle di-terpene content could therefore cause lower survival due to a decreased ability in anti-predator behavior of sawfly larvae. Browsing could potentially alter di-terpene content of pine needles (Nykänen and Koricheva, 2004), which could thus affect sawfly survival. However, browsing has been shown to not affect pine needle di-terpenes (Nordkvist et al., 2019). Another possibility is that moose selectively forage on pines with low di-terpene content, and therefore we observe lower survival on browsed trees. Another candidate plant quality trait is tannins, which reduce available nitrogen (Feeny, 1968), and has been shown to be affected by browsing (du Toit et al., 1990; Hrabar and du Toit, 2014). Tannins could potentially alter the amount of alternative prey and consequently predation. Browsing could potentially alter other architectural traits than the ones measured in this study, such as the proportion of leaf vs. branch morphologies or the amount of edges, which can serve as attachment points for predators (Reynolds and Cuddington, 2012a,b), and subsequently affect predation rates. Further studies targeting these and other architectural traits, such as branch length (which could be importance in terms of ant movement), are needed if we want to understand how browsing affects arthropod predators and predation. Moreover, studies simulating browsing induced changes to architecture could be beneficial in elucidating the mechanisms, as it is then possible to simulate certain architectural responses in isolation. To us it does not seem plausible that the difference in survival is caused only by browsing induced bottom-up plant quality effects on the sawfly larvae. Our assertion is based on a previous study using the same study system in which we did not find an effect of browsing on bottom-up survival (Nordkvist et al., 2019).
Based on the results from study 1 (quadratic response of ants to browsing intensity Figure 1 and Table 2), our expectation was that sawfly survival would follow the same trend—i.e., be lower on moderately browsed trees but higher on intensively browsed trees. However, we found reduced survival at both browsing intensities. This inconsistency could be caused by diverging predator behavior on intensively browsed trees. Predation rate has been reported to differ between different architectural variations; Grevstad et al. (1992) showed changes in predator encounter rate, kill rate and forage rate on plants with varying stem and leaf morphologies as well as total leaf area. It could also be a result of stronger negative bottom-up effects on larval survival on intensively browsed trees, as level of induced bottom-up effects may increase with damage intensity (Underwood, 2000). In a previous study we did not find difference in sawfly larval survival between browsed and un-browsed trees (Nordkvist et al., 2019), in this study only moderately (50%) browsed pines were used. Since plant quality traits change not just with the presence but also with intensity of browsing (Guillet and Bergström, 2006; Kohi et al., 2010), there is a possibility that a different bottom-up effect emerges in intensively browsed trees. On the other hand, the diverging responses on intensively browsed trees in study 1 and 2 could be a result of altered predator community (at species level) on moderately and intensively browsed trees. In study 1 we did not assess species identity, hence we might have missed important differences in species composition such as that there could be fewer but more efficient predators on intensively browsed trees. Future studies would benefit from surveying the predator community and studying the survival of herbivores on the same pines.
Conclusion
We conclude that ungulate browsing can influence arthropod predator communities, survival rates of herbivorous insects and thereby potentially damage levels. In addition, the intensity of browsing is of importance for arthropod predators, and that browsing may induce changes in predator community composition. The underlying mechanisms are, however, still elusive, and further studies are needed.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
MN, CB, and MK designed the experiments. MN set up the experiments, collected, and analyzed the data and wrote the first draft of the manuscript. All authors contributed to revisions of the manuscript.
Funding
This study was financed by FORMAS, Grant no: 2015-55.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful to Karin Eklund and Lars Edenius for valuable comments on the experimental design. We also thank Kevin Cestrières, Daniel Leppert, Erik Karlsson, and Daniel Tooke for help in the field.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2021.666069/full#supplementary-material
References
Bates, D., Maechler, M., Bolker, B., and Walker, S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48. doi: 10.18637/jss.v067.i01
Björkman, C., Larsson, S., and Bommarco, R. (1997). Oviposition preferences in pine sawflies: a trade-off between larval growth and defence against natural enemies. Oikos 79:45. doi: 10.2307/3546088
Björkman, C., Larsson, S., and Gref, R. (1991). Effects of nitrogen fertilization on pine needle chemistry and sawfly performance. Oecologia 86, 202–209. doi: 10.1007/BF00317532
Campos, R. I., Vasconcelos, H. L., Ribeiro, S. P., Neves, F. S., and Soares, J. P. (2006). Relationship between tree size and insect assemblages associated with Anadenanthera macrocarpa. Ecography 29, 442–450. doi: 10.1111/j.2006.0906-7590.04520.x
Cuddington, K., and Yodzis, P. (2002). Predator-prey dynamics and movement in fractal environments. Am. Nat. 160, 119–134.
Danell, K., and Huss-Danell, K. (1985). Feeding by insects and hares on birches earlier affected by moose browsing. Oikos 44:75. doi: 10.2307/3544046
Danell, K., Bergström, R., and Edenius, L. (1994). Effects of large mammalian browsers on architecture, biomass, and nutrients of woody plants. J. Mammal. 75, 833–844. doi: 10.2307/1382465
Den Herder, M., Bergström, R., Niemelä, P., Danell, K., and Lindgren, M. (2009). Effects of natural winter browsing and simulated summer browsing by moose on growth and shoot biomass of birch and its associated invertebrate fauna. Ann. Zool. Fennici 46, 63–74. doi: 10.5735/086.046.0107
du Toit, J. T., Bryant, J. P., and Frisby, K. (1990). Regrowth and palatability of acacia shoots following pruning by African savanna browsers. Ecology 71, 149–154. doi: 10.2307/1940255
Edenius, L., Danell, K., and Bergström, R. (1993). Impact of herbivory and competition on compensatory in growth woody winter plants: on Scots pine browsing by moose. Oikos 66, 286–292. doi: 10.2307/3544816
Feeny, P. P. (1968). Effect of oak leaf tannins on larval growth of the winter moth Operophtera brumata. J. Insect Physiol. 14, 805–817. doi: 10.1016/0022-1910(68)90191-1
Forkner, R. E., and Hunter, M. D. (2000). What goes up must come down? Nutrient addition and predation pressure on oak herbivores. Ecology 81, 1588–1600.
Fox, J., and Weisberg, S. (2019). An {R} Companion to Applied Regression, 3rd Edn. Thousand Oaks, CA: Sage.
Grevstad, F. S., Klepetka, B. W., and Url, S. (1992). The influence of plant architecture on the foraging efficiencies of a suite of ladybird beetles feeding on aphids. Oecologia 92, 399–404. doi: 10.1007/BF00317466
Guillet, C., and Bergström, R. (2006). Compensatory growth of fast-growing willow (Salix) coppice in response to simulated large herbivore browsing. Oikos 113, 33–42. doi: 10.1111/j.0030-1299.2006.13545.x
Gur’yanova, T. M. (2006). Fecundity of the European pine sawfly Neodiprion sertifer (Hymenoptera, Diprionidae) related to cyclic outbreaks: invariance effects. Entomol. Rev. 86, 910–921. doi: 10.1134/S0013873806080069
Halaj, J., Ross, D. W., and Moldenke, A. R. (1997). Negative effects of ant foraging on spiders in Douglas-fir canopies. Oecologia 109, 313–322. doi: 10.1007/s004420050089
Halaj, J., Ross, D. W., and Moldenke, A. R. (2000). Importance of habitat structure to the arthropod food-web in Douglas-fir canopies. Oikos 90, 139–152. doi: 10.1034/j.1600-0706.2000.900114.x
Kohi, E. M., De Boer, W. F., Slot, M., Van Wieren, S. E., Ferwerda, J. G., Grant, R. C., et al. (2010). Effects of simulated browsing on growth and leaf chemical properties in Colophospermum mopane saplings. Afr. J. Ecol. 48, 190–196. doi: 10.1111/j.1365-2028.2009.01099.x
Hrabar, H., and du Toit, J. T. (2014). Interactions between megaherbivores and microherbivores: elephant browsing reduces host plant quality for caterpillars. Ecosphere 5:7. doi: 10.1890/ES13-00173.1
Juutinen, P. (1967). “Zur Bionomie und zum Vorkommen der Roten Kiefernbuschhornblattwespe (Neodiprion sertifer Geoffr.) in Finnland in den Jahren 1959–65,” in Mets ntutkimuslaitoksen julkaisuja 63. Available online at: http://jukuri.luke.fi/handle/10024/523102
Kaitaniemi, P., Riihimäki, J., Koricheva, J., and Vehviläinen, H. (2007). Experimental evidence for associational resistance against the European pine sawfly in mixed tree stands. Silva Fenn. 41, 259–268. doi: 10.14214/sf.295
Kaitaniemi, P., Vehviläinen, H., and Ruohomäki, K. (2004). Movement and disappearance of mountain birch defoliators are influenced by the interactive effects of plant architecture and induced resistance. Ecol. Entomol. 29, 437–446. doi: 10.1111/j.0307-6946.2004.00617.x
Kollberg, I., Bylund, H., Jonsson, T., Schmidt, A., Gershenzon, J., and Björkman, C. (2015). Temperature affects insect outbreak risk through tritrophic interactions mediated by plant secondary compounds. Ecosphere 6, 1–17. doi: 10.1890/ES15-000021.1
Landsman, A. P., and Bowman, J. L. (2017). Discordant response of spider communities to forests disturbed by deer herbivory and changes in prey availability. Ecosphere 8:e01703. doi: 10.1002/ecs2.1703
Langellotto, G. A., and Denno, R. F. (2004). Responses of invertebrate natural enemies to complex-structured habitats: a meta-analytical synthesis. Oecologia 139, 1–10. doi: 10.1007/s00442-004-1497-3
Larsson, S., Björkman, C., and Gref, R. (1986). Responses of Neodiprion sertifer (Hym., Diprionidae) larvae to variation in needle resin acid concentration in Scots pine. Oecologia 70, 77–84. doi: 10.1007/BF00377113
Lawton, J. H. (1983). Plant architecture and the diversity of phytophagous insects. Annu. Rev. Entomol. 28, 23–39. doi: 10.1016/B978-0-12-374144-8.00000-0
Lefcheck, J. S. (2016). PiecewiseSEM: piecewise structural equation modelling in r for ecology, evolution, and systematics. Methods Ecol. Evol. 7, 573–579. doi: 10.1111/2041-210X.12512
Lilleeng, M. S., Rydgren, K., Halvorsen, R., Moe, S. R., and Hegland, S. J. (2018). Red deer structure the ground-dwelling beetle community in boreal forest. Biodivers. Conserv. 27, 2507–2525. doi: 10.1007/s10531-018-1550-x
Lind, E. M., Myron, E. P., Giaccai, J., and Parker, J. D. (2012). White-tailed deer alter specialist and generalist insect herbivory through plant traits. Environ. Entomol. 41, 1409–1416. doi: 10.1603/EN12094
Lindstedt, C., Mappes, J., Päivinen, J., and Varama, M. (2006). Effects of group size and pine defence chemicals on Diprionid sawfly survival against ant predation. Oecologia 150, 519–526. doi: 10.1007/s00442-006-0518-9
Lorentzen Kolstad, A., Austrheim, G., Solberg, E. J., De Vriendt, L., and Speed, J. D. M. (2018). Pervasive moose browsing in boreal forests alters successional trajectories by severely suppressing keystone species. Ecosphere 9:e02458. doi: 10.1002/ecs2.2458
Martin, J. L., Stockton, S. A., Allombert, S., and Gaston, A. J. (2010). Top-down and bottom-up consequences of unchecked ungulate browsing on plant and animal diversity in temperate forests: lessons from a deer introduction. Biol. Invasions 12, 353–371. doi: 10.1007/s10530-009-9628-8
Martinsen, G. D., Driebe, E. M., and Whitham, T. G. (1998). Indirect interactions mediated by changing plant chemistry: beaver browsing benefits beetles. Ecology 79, 192–200.
McInnes, P. F., Naiman, R. J., Pastor, J., and Cohen, Y. (1992). Effects of moose browsing on vegetation and litter of the boreal forest, Isle Royale, Michigan, USA. Ecology 73, 2059–2075. doi: 10.2307/1941455
Muiruri, E. W., Barantal, S., Iason, G. R., Salminen, J. P., Perez-Fernandez, E., and Koricheva, J. (2019). Forest diversity effects on insect herbivores: do leaf traits matter? New Phytol. 221, 2250–2260. doi: 10.1111/nph.15558
Muiruri, E. W., Milligan, H. T., Morath, S., and Koricheva, J. (2015). Moose browsing alters tree diversity effects on birch growth and insect herbivory. Funct. Ecol. 29, 724–735. doi: 10.1111/1365-2435.12407
Nordkvist, M., Klapwijk, M. J., Edenius, L., Gershenzon, J., Schmidt, A., and Björkman, C. (2019). Trait-mediated indirect interactions: moose browsing increases sawfly fecundity through plant-induced responses. Ecol. Evol. 9, 10615–10629. doi: 10.1002/ece3.5581
Nykänen, H., and Koricheva, J. (2004). Damage-induced changes in woody plants and their effects on insect herbivore performance: a meta-analysis. Oikos 104, 247–268. doi: 10.1111/j.0030-1299.2004.12768.x
Ohse, B., Hammerbacher, A., Seele, C., Meldau, S., Reichelt, M., Ortmann, S., et al. (2017). Salivary cues: simulated roe deer browsing induces systemic changes in and defence chemistry in wild−grown maple and beech saplings. Funct. Ecol. 31, 340–349. doi: 10.1111/1365-2435.12717
Olofsson, E. (1989). Transmission of the nuclear polyhedrosis virus of the European pine sawfly from adult to offspring. J. Invertebr. Pathol. 54, 322–330.
Olofsson, E. (1992). Predation by Formica polyctena Förster (Hym., Formicidae) on newly emerged larvae of Neodiprion sertifer (Geoffroy) (Hym., Diprionidae). J. Appl. Entomol. 114, 315–319. doi: 10.1111/j.1439-0418.1992.tb01132.x
R Core Team (2019). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Reynolds, P. G., and Cuddington, K. (2012a). Effect of plant gross morphology on predator consumption rates. Environ. Entomol. 41, 508–515. doi: 10.1603/EN11178
Reynolds, P. G., and Cuddington, K. (2012b). Effect of plant gross morphology on predator searching behaviour. Environ. Entomol. 41, 516–522. doi: 10.1603/EN11179
Riihimäki, J., Vehviläinen, H., Kaitaniemi, P., and Koricheva, J. (2006). Host tree architecture mediates the effect of predators on herbivore survival. Ecol. Entomol. 31, 227–235. doi: 10.1111/j.1365-2311.2006.00784.x
Šipoš, J., and Kindlmann, P. (2012). Effect of the canopy complexity of trees on the rate of predation of insects. J. Appl. Entomol. 137, 445–451. doi: 10.1111/jen.12015
Speed, J. D. M., Austrheim, G., Hester, A. J., Meisingset, E. L., Mysterud, A., Tremblay, J. P., et al. (2014). General and specific responses of understory vegetation to cervid herbivory across a range of boreal forests. Oikos 123, 1270–1280. doi: 10.1111/oik.01373
Suominen, O., Danell, K., and Bergstrom, R. (1999). Moose, trees, and ground-living invertebrates: indirect interactions in Swedish pine forests. Oikos 84, 215–226. doi: 10.2307/3546716
Suominen, O., Persson, I.-L., Danell, K., Bergström, R., and Pastor, J. (2008). Impact of simulated moose densities on abundance and richness of vegetation, herbivorous and predatory arthropods along a productivity gradient. Ecography 31, 636–645. doi: 10.1111/j.0906-7590.2008.05480.x
Underwood, N. (2000). Density dependence in induced plant resistance to herbivore damage: threshold, strength and genetic variation. Oikos 89, 295–300. doi: 10.1034/j.1600-0706.2000.890210.x
Keywords: trait-mediated effects, indirect interactions, neodiprion sertifer, plant architecture, alces alces, herbivory
Citation: Nordkvist M, Björkman C and Klapwijk MJ (2021) Plant Mediated Interactions: Lower Sawfly Survival on Pines Previously Browsed by Moose. Front. Ecol. Evol. 9:666069. doi: 10.3389/fevo.2021.666069
Received: 09 February 2021; Accepted: 06 April 2021;
Published: 26 April 2021.
Edited by:
Mingbo Yin, Fudan University, ChinaReviewed by:
Carita Lindstedt, University of Jyväskylä, FinlandElina Mäntylä, Academy of Sciences of the Czech Republic, Czechia
Copyright © 2021 Nordkvist, Björkman and Klapwijk. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Michelle Nordkvist, bWljaGVsbGUubm9yZGt2aXN0QHNsdS5zZQ==