 Fusong Han
Fusong Han Chengqun Yu
Chengqun Yu Gang Fu
Gang Fu- 1Lhasa Plateau Ecosystem Research Station, Key Laboratory of Ecosystem Network Observation and Modeling, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing, China
- 2School of Resource and Environment and Safety Engineering, Hunan University of Science and Technology, Xiangtan, Hunan, China
It is well known that asymmetric warming among elevations (i.e., warming magnitude increases with increasing elevation) will weaken the difference of air temperature among elevations. However, it remains controversial on whether asymmetric warming among elevations can homogenize plant α-diversity and above-ground net primary production (ANPP) in alpine regions. In the present study, we conducted an experiment of asymmetric warming among elevations in alpine grasslands, Northern Tibet since 2010. There were four experiment treatments, including a treatment under natural conditions at elevation 4,313 m (C4313), a treatment under natural conditions at elevation 4,513 m (C4513), a treatment under warming conditions at elevation 4,513 m (W4513) and a treatment under warming conditions at elevation 4,693 m (W4693). We investigated ANPP, taxonomic α-diversity (i.e., species richness, Shannon, Simpson and Pielou) and phylogenetic α-diversity (mean nearest taxon distance, MNTD; phylogenetic diversity, PD) in 2011–2019. There were no significant differences of mean air temperature between C4313 and W4513, or between C4513 and W4693 in 2011–2019, indicating the differences of air temperature were eliminated among elevations. Then we found that the differences of plant α-diversity and ANPP were also eliminated among elevations: (1) there were no significant differences of ANPP, Pielou and MNTD between C4313 and W4513, or between C4513 and W4693 in 2011–2019. (2) There were also no significant differences of mean species richness, Shannon and Simpson between C4513 and W4693 in 2011–2019. (3) There were also no significant differences of ANPP, species richness, Shannon, Simpson, Pielou, PD and MNTD between C4313 and W4513, or C4513 and W4693 in 2019. Therefore, asymmetric warming among elevations may homogenize plant α-diversity and aboveground net primary production in alpine grasslands, at least in Northern Tibet.
1. Introduction
Climate warming is an accepted fact, and the warming magnitudes vary by region (Tian and Fu, 2022). The asymmetric warming among elevations, as a case of spatial asymmetric warming, can generally refer to warming magnitude increases with increasing elevation for some special regions, especially in some highland areas (Yao et al., 2000; Rangwala and Miller, 2012). However, there is still argument on how such asymmetric warming among elevations can influence terrestrial ecosystems, which can be mainly due to the following facts. First, previous studies have focused on the phenomenon itself of asymmetric warming among elevations and possible driving mechanisms (Rangwala and Miller, 2012) rather than the effects of asymmetric warming among elevations on ecosystem structure and function. Second, asymmetric warming generally includes daily asymmetric warming (Zhong and Fu, 2022), seasonal asymmetric warming (Fu et al., 2019; Fu and Shen, 2022; Han et al., 2023) and spatial asymmetric warming (Wang et al., 2021b, 2022; Fu et al., 2022). Previous studies have primarily explored the impacts of daily and seasonal asymmetric warming on terrestrial ecosystems (Bokhorst et al., 2011; Tozzi et al., 2014; Ji et al., 2017; Yang et al., 2017; Fu et al., 2019; Fu and Shen, 2022; Zhong and Fu, 2022), but only a small number of studies have tried to probe the responses of terrestrial ecosystems to asymmetric warming among elevations (Wang et al., 2021b). Symmetric warming among elevations may overestimate the impact of asymmetric warming among elevations on plant species and phylogenetic composition (Wang et al., 2021a,b; Fu and Sun, 2022), which implies that some related conclusions from previous studies may exaggerate the impacts of climate warming on ecosystem structure and function. There is still a great deal of uncertainty about the effects of climate warming on various ecosystems, because asymmetric warming among elevations has not received enough attention. Third, warming magnitude is one of the core problems for spatial and temporal asymmetric warming, and thus many previous studies have tried to investigate how warming magnitude can alter the impacts of climate warming on terrestrial ecosystems and found that temperature sensitivities of ecosystem structure and function can generally change with warming magnitude (Fu et al., 2015b, 2018, 2022; Yu et al., 2019b; Fu and Sun, 2022). Such studies may indirectly support that the effects of asymmetric warming among elevations on various ecosystems are different from those of symmetric warming among elevations, but can hardly obtain the impacts of asymmetric warming among elevations on terrestrial ecosystems directly. Therefore, further investigation on the impacts of asymmetric warming among elevations on terrestrial ecosystems will be necessary.
Grasslands, as an important vegetation type in global ecosystems, are key resources for high-quality development of livestock industry and an important place for wildlife protection (Fu et al., 2022; Zha et al., 2022; Han et al., 2022a; Wang and Fu, 2023). Alpine grasslands are important sections of global grasslands, and may be the most sensitive and fragile grassland types to global change (Zhang et al., 2015, 2022; Fu and Sun, 2022). Alpine meadows and alpine steppes on the Qinghai-Tibet Plateau are not only two of the most typical alpine grasslands on the world, but also the grassland types closest to the sky on the earth (Zhang et al., 2015; Zong and Fu, 2021; Han et al., 2022b). Aboveground net primary production (ANPP) determines the largest carrying capacity of grazing livestock and wild animals in grasslands (Wang et al., 2022). A certain size of plant α-diversity and relatively rational plant community composition are the basis for sustaining large ANPP in grasslands (Wang et al., 2021a; Fu and Sun, 2022). The ANPP and plant diversity of alpine grasslands are survival basis for maintaining yaks and other specific plateau grazing livestock and wild animals, which can ensure the livelihood of Tibetan people’s on the Qinghai-Tibet Plateau (Fu et al., 2022; Wang et al., 2022). Alpine grasslands of the Qinghai-Tibet Plateau are not only the typical regions with asymmetric warming among elevations (Qin et al., 2009; Du et al., 2019), but also the perfect areas to trace alterations in ANPP and biodiversity under external disturbance conditions (Trisos et al., 2020; Tian and Fu, 2022; Zhang et al., 2022). However, as we refer to the best knowledge, only one study has tried to explore the effects of asymmetric warming among elevations on plant species and phylogenetic composition in alpine grasslands on the Tibetan Plateau (Wang et al., 2021b) and no reports have investigated the impacts of asymmetric warming among elevations on ANPP and plant α-diversity in alpine grasslands of the Tibetan Plateau. This limits our ability to accurately predict plant productivity and diversity in alpine grassland ecosystems under future scenes of asymmetric warming among elevations, which in turn may limit our abilities to fully use grassland forages for scientific and high-quality livestock development, as well as biodiversity conservation. Therefore, investigating the impacts of asymmetric warming among elevations on plant α-diversity and ANPP in alpine grasslands of the Tibetan Plateau will undisputablly broaden our mind.
Here, since May 2010, we performed an experiment of asymmetric warming among elevations in alpine grasslands of the Tibet, and measured ANPP, plant phylogenetic and species α-diversity in 2011–2019. The main goal of the present study was to explore whether asymmetric warming among elevations may homogenize ANPP, plant phylogenetic and species α-diversity. Previous studies found that asymmetric warming among elevations may homogenize plant phylogenetic and species composition in alpine grasslands of the Qinghai-Tibet Plateau (Wang et al., 2021b). Previous studies also found that the temperature sensitivity of plant production were mainly related to temperature itself (Fu and Sun, 2022) and asymmetric warming among elevations can result in homogeneous temperature among elevations in alpine grasslands (Wang et al., 2021b). Therefore, we hypothesized that asymmetric warming among elevations may homogenize ANPP, plant phylogenetic and species α-diversity in alpine grasslands.
2. Materials and methods
2.1. Study area and experimental design
The detailed information about the study area and experiment design have been published in our previous studies (Zhang et al., 2021; Wang et al., 2021b; Fu and Sun, 2022). Since May 2010, we chose three alpine grassland sites with elevation about 4,313, 4,513, and 4,693 m in the Northern Tibet Plateau, respectively. Four open top chambers (top with 1.00 m in diameter, bottom with 1.45 m in diameter, and height with 0.40 m) were established at elevation 4,513 m (W4513) and 4,693 m (W4693), and four control plots (no open top chambers) were established at elevation 4,313 m (C4313) and 4,513 m (C4513), respectively.
2.2. Microclimate observations, soil sampling, and analyses
The detailed information about the monitoring and observations of soil temperature (Ts, 0.05 m), air temperature (Ta, 0.15 m), soil moisture (SM, 0.10 m), and relative humidity (RH, 0.15 m), and a derived microclimate variable (i.e., vapor pressure deficit, VPD), could be discovered by our prior reports (Zhang et al., 2021; Wang et al., 2021b). The HOBO microclimate observation stations were used to monitor Ts, Ta, SM and RH, and the original sampling frequency of these climate data was 1 min (Zhang et al., 2021; Fu and Sun, 2022). The VPD was calculated from Ta and RH (Yu et al., 2019b; Fu and Shen, 2022). The Ts, Ta, SM, and VPD during the period from June to September in 2011–2019 were used in the present study. The topsoil (0–0.10 m) samples were gathered in August, 2013 and 2015–2019, and then used to measure soil nitrate nitrogen (NO3−-N), ammonium nitrogen (NH4+-N), pH and available phosphorus (AP; Yu et al., 2019a). The comparisons of Ts, Ta, SM, VPD, NO3−-N, NH4+-N, AP, and pH between C4313 and W4513, C4313 and W4693, or C4513 and W4693 were published by our a previous study (Wang et al., 2021b).
2.3. Community investigation and ANPP simulations
The detailed information about plant community investigation could be discovered by our prior reports (Wang et al., 2021a,b). We used measured species height and coverage to obtain the important value for each species. Important values were used to obtain species α-diversity, including species richness, Shannon, Simpson and Pielou (Sun et al., 2021). We obtained plant phylogenetic tree from taxonomic information for each species (Sun et al., 2021). Then we calculated phylogenetic α-diversity (Faith’s phylogenetic diversity, PD; mean nearest taxon distance, MNTD) using important values matrix of all species and phylogenetic tree (Sun et al., 2021). All the detected species in our experimental plots were categorized into sedge, graminoid and forb function groups. Aboveground net primary production of sedge (ANPPsedge), graminoid (ANPPgraminoid), and forb (ANPPforb) were estimated from measured sedge, graminoid and forb coverage and height, respectively (Wang et al., 2021a). Community aboveground net primary production (ANPP) was obtained from the following equation.
2.4. Statistical analyses
The temperature difference among elevations can be decreased and even approached zero under the asymmetric warming among elevations conditions (Wang et al., 2021b). If there were no significant differences of plant productivity and α-diversity among elevations, asymmetric warming among elevations were supposed to homogenize ANPP and plant α-diversity. We calculated geographic distance among the three elevations. For each year, we used t-test to compare the differences of ANPP, species richness, Shannon, Simpson, Pielou, PD and MNTD between C4313 and W4513, between C4513 and W4693, or between C4313 and W4693. Then we used repeated measure analysis of variance to test the differences of ANPP, species richness, Shannon, Simpson, Pielou, PD, and MNTD between C4313 and W4513, between C4513 and W4693, or between C4313 and W4693 based on all the nine-year measured data. We used environment variables to partition the variations of plant α-diversity and ANPP into four parts. All the tests of significance referred to p < 0.05 and performed in R 4.1.2. The package used in the present study included the SPAA, vegan, Plantlist and Picante.
3. Results
3.1. Comparisons of plant α-diversity and aboveground plant production between the C4313 versus W4513, C4313 versus W4693, and C4513 versus W4693
Significant dissimilarities for the multi-year average ANPP of the C4313 versus W4513, C4313 versus W4693, and C4513 versus W4693 in 2011–2019 were not detected (Table 1). Significant dissimilarities for multi-year average ANPPsedge and ANPPgraminoid of the C4313 versus W4513, and significant dissimilarity for multi-year average ANPPforb of the C4313 versus W4693 in 2011–2019 were not detected (Table 1). The multi-year average ANPPsedge of the W4693 was larger than that of the C4313 and C4513, while the multi-year average ANPPgraminoid of the W4693 was lower than that of the C4313 and C4513 in 2011–2019 (Table 1). A significant dissimilarity of multi-year average ANPPforb was also detected between the C4313 versus W4513, and the C4513 versus W4693 in 2011–2019 (Table 1). Two of the six variables of plant α-diversity (i.e., Pielou and MNTD) between the C4313 versus W4513 were not significant dissimilar in 2011–2019 (Table 1). None of the six variables of plant α-diversity between the C4313 versus W4693 was significant dissimilar in 2011–2019 (Table 1). Only one of the six variables of plant α-diversity (i.e., PD) between the C4513 versus W4693 was significant dissimilar in 2011–2019 (Table 1). The dissimilarities of the ANPP, ANPPsedge, ANPPgraminoid, ANPPforb, and the six variables of plant α-diversity between the C4313 versus W4513, the C4313 versus W4693, and the C4513 versus W4693 varied with year (Figures 1, 2).
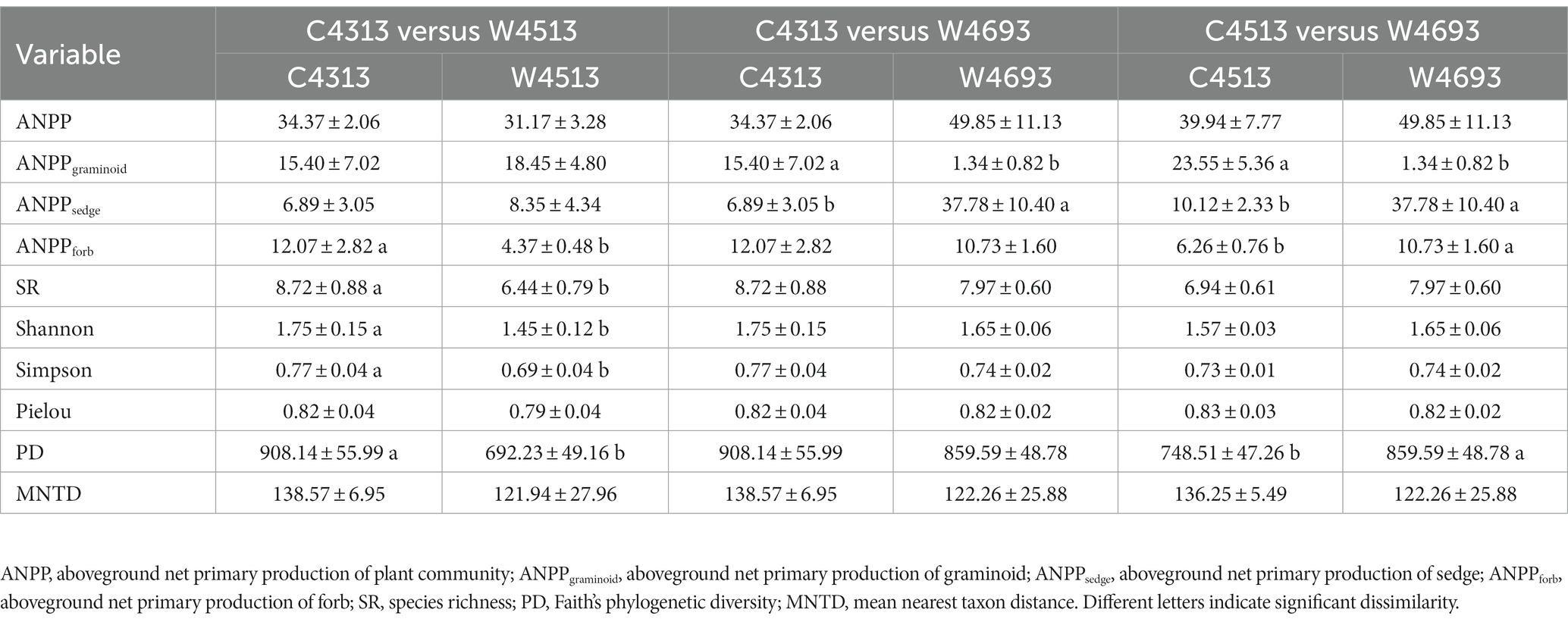
Table 1. Comparison of average ANPP, ANPPgraminoid, ANPPsedge, ANPPforb, SR, Shannon, Simpson, Pielou, PD, and MNTD in 2011–2019 of the C4313 versus W4513, C4313 versus W4693, and C4513 versus W4693 based on a repeated ANOVA.
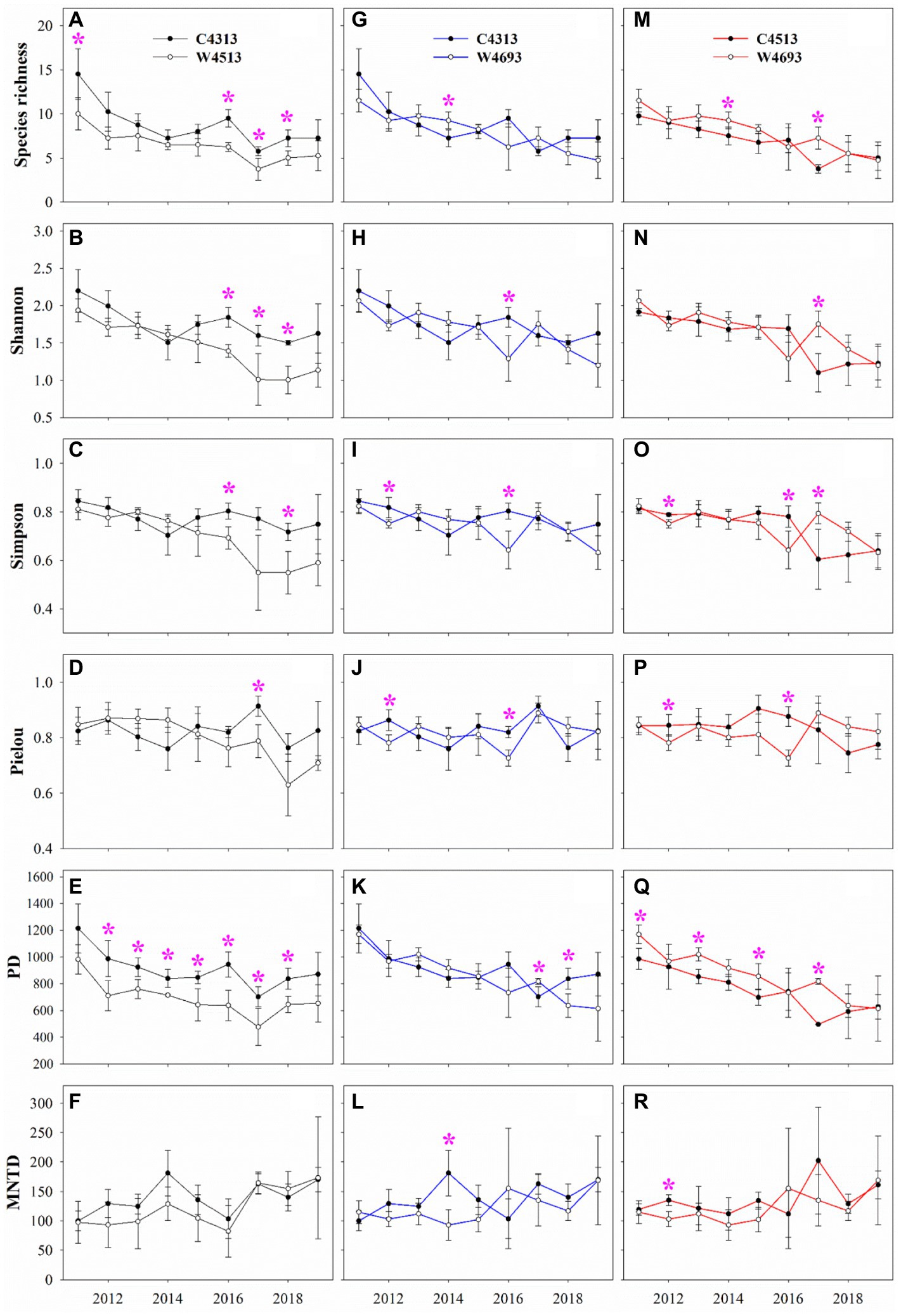
Figure 1. Comparison of species richness (SR), Shannon, Simpson, Pielou, Faith’s phylogenetic diversity (PD) and mean nearest taxon distance (MNTD) of plant community (A–F) between the non-warming plots at elevation 4,313 m (C4313) and warming plots at elevation 4,513 m (W4513), (G–L) between the C4313 and warming plots at elevation 4,693 m (W4693), and (M–R) between the non-warming plots at elevation 4,513 m (C4513) and the W4693.
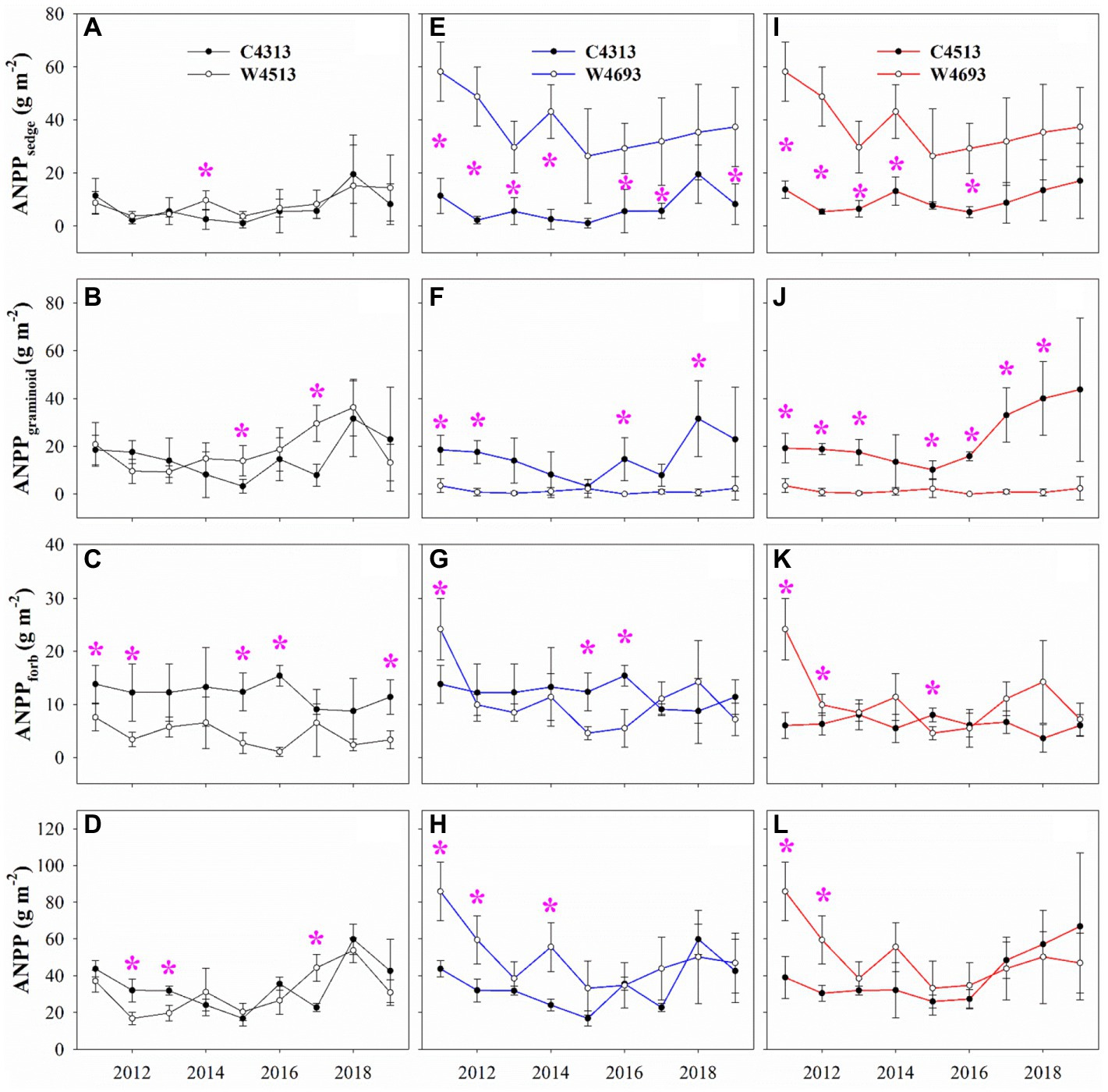
Figure 2. Comparison of aboveground net primary production of plant community (ANPP), aboveground net primary production of sedge (ANPPsedge), aboveground net primary production of graminoid (ANPPgraminoid), and aboveground net primary production of forb (ANPPforb; A–D) between the non-warming plots at elevation 4,313 m (C4313) and warming plots at elevation 4,513 m (W4513), (E–H) between the C4313 and warming plots at elevation 4,693 m (W4693), and (I–L) between the non-warming plots at elevation 4,513 m (C4513) and the W4693.
3.2. Correlations between ANPP, ANPPgraminoid, ANPPsedge, ANPPforb, and α-diversity versus environmental variables, and ANPP versus α-diversity
The ANPP dissimilarities of C4313 versus W4513, C4313 versus W4693, and C4513 versus W4693 decreased with warming duration, but that of Shannon, Simpson and Pielou increased with warming duration (Supplementary Table S1). The ANPP dissimilarity increased with phylogenetic composition dissimilarity of C4313 versus W4513, C4313 versus W4693, and C4513 versus W4693 (Supplementary Table S1). The ANPPsedge dissimilarity increased with the dissimilarity of Ts and pH of C4313 versus W4513, C4313 versus W4693, and C4513 versus W4693 (Supplementary Table S1). The Pielou dissimilarity decreased with the NO3−-N dissimilarity of C4313 versus W4513, C4313 versus W4693, and C4513 versus W4693 (Supplementary Table S1).
All the four explained parts had excluded impacts on Shannon, Simpson, PD, ANPPsedge and ANPPforb (Figure 3; Supplementary Figure S1). Only one of the four explained parts (i.e., soil pH) had a no excluded impact on species richness (Figure 3). Two of the four explained parts (i.e., “GeoDist.Dur.” and “Soil nutrition”) had excluded impacts on Pielou, but they did not have excluded impacts on MNTD (Figure 3). Two of the four explained parts (i.e., “Temp.Water” and “Soil nutrition”) had excluded impacts on ANPP (Supplementary Figure S1). Only one of the four explained parts (i.e., Temp.Water) had a no excluded impact on ANPPgraminoid (Supplementary Figure S1). Only one of the four explained parts (i.e., species β-diversity) had a no excluded impact on ANPP (Figure 4). Only the “Env.” had an excluded impact on ANPP (Figure 5). Besides the impact of the “Env.”, the β-diversity had interactive impacts with “GeoDist.Dur.” and α-diversity on ANPP (Figure 5).
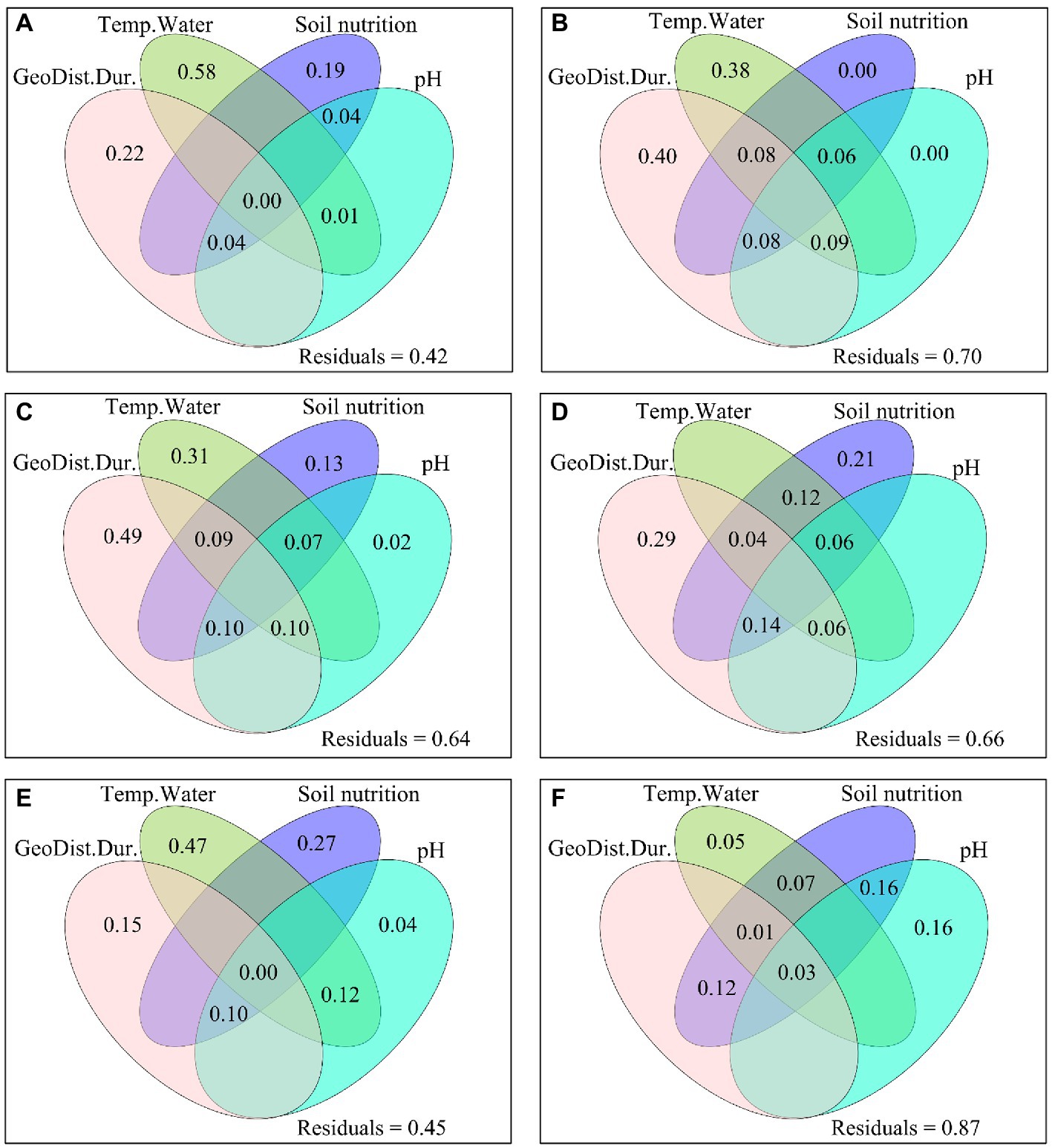
Figure 3. The excluded and interactive impacts of GeoDist.Dur., the dissimilarity of Temp.Water, soil nutrition and pH on the dissimilarity of (A) species richness, (B) Shannon, (C) Simpson, (D) Pielou, (E) PD, and (F) MNTD between C4313 versus W4513, C4313 versus W4693, and C4513 versus W4693. PD: phylogenetic diversity; MNTD: mean nearest taxon distance; GeoDist.Dur.: geographic distance and warming duration; Temp.Water: soil and air temperatures, vapor pressure deficit and/or soil moisture; soil nutrition: available phosphorus, nitrate and ammonium nitrogen. C4313: non-warming plots at elevation 4,313 m; C4513: non-warming plots at elevation 4,513 m; W4513: warming plots at elevation 4,513 m; W4693: warming plots at elevation 4,693 m.
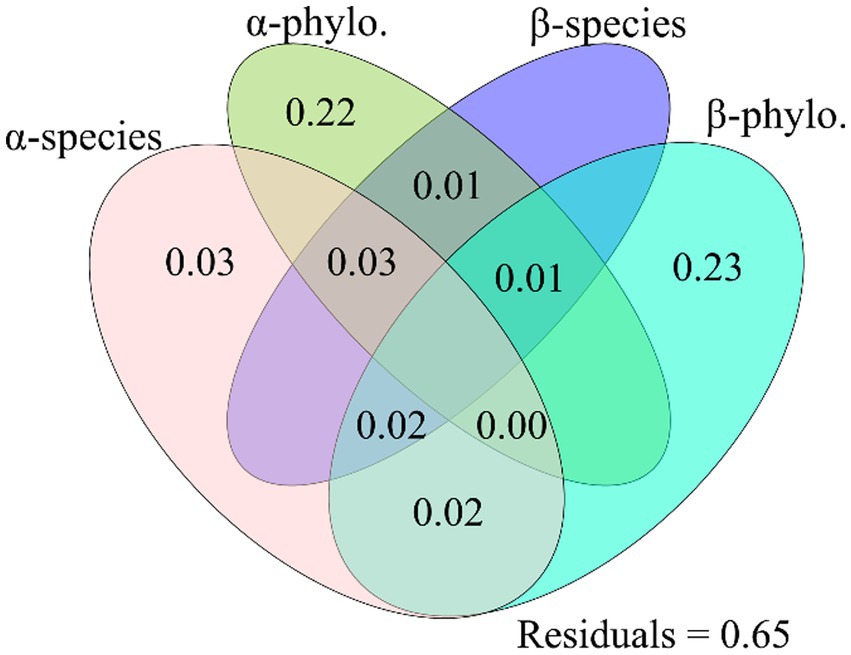
Figure 4. The excluded and interactive impacts of the dissimilarity of α-species and α-phylo., β-species and β-phylo. on the dissimilarity of plant community aboveground net primary production between C4313 versus W4513, C4313 versus W4693, and C4513 versus W4693. The α-phylo. and α-species indicated phylogenetic and species α-diversity, and β-phylo,. and β-species indicated phylogenetic and species β-diversity between C4313 versus W4513, C4313 versus W4693, and C4513 versus W4693, respectively. C4313: non-warming plots at elevation 4,313 m; C4513: non-warming plots at elevation 4,513 m; W4513: warming plots at elevation 4,513 m; W4693: warming plots at elevation 4,693 m. Species and phylogenetic β-diversity data were obtained from Wang et al. (2021a,b), Frontiers in Ecology and Evolution.

Figure 5. The excluded and interactive impacts of GeoDist.Dur., the dissimilarity of Env. and α-diversity, and β-diversity on the dissimilarity of plant community aboveground net primary production between C4313 versus W4513, C4313 versus W4693, and C4513 versus W4693. GeoDist.Dur.: geographic distance and warming duration; Env.: soil and air temperatures, soil moisture, vapor pressure deficit, available phosphorus, nitrate and ammonium nitrogen, and/or pH; C4313: non-warming plots at elevation 4,313 m; C4513: non-warming plots at elevation 4,513 m; W4513: warming plots at elevation 4,513 m; W4693: warming plots at elevation 4,693 m. Species and phylogenetic β-diversity data were obtained from Wang et al. (2021a,b), Frontiers in Ecology and Evolution.
4. Discussion
Concordant with the hypothesis, asymmetric warming among elevations might homogenize plant phylogenetic and species α-diversity. This phenomenon might be consequent to at least one of the subsequent mechanisms. First, each species has its special niche width of temperature, water, nitrogen and phosphorus availability (Thuiller et al., 2005; Brathen et al., 2018). Elevated temperature, water, nitrogen and phosphorus availability can alter plant α-diversity (Klein et al., 2004; Fu and Shen, 2016; Wang et al., 2021a). Asymmetric warming among elevations may homogenize temperature, water, nitrogen and phosphorus availability among elevations, which in turn may result in similar niche width of temperature, water, nitrogen and phosphorus availability among elevation. Second, wind may also impact plant diversity (Wang et al., 2021b), and warming may result in the declines in gale days and wind speed (Xiaomei et al., 2012; Yao and Li, 2019). Asymmetric warming among elevations may homogenize gale days and wind speed among elevations, which in turn may increase dispersal limitation of anemophilae plants among elevations. Third, radiation is an important limit variable of plant species α-diversity (Tian and Fu, 2022), and generally closely correlated with precipitation (Fu et al., 2022; Han et al., 2022a). Homogenization of wind among elevations may lead to homogenization of precipitation among elevation, which in turn may weaken the difference of radiation among elevations. Fourth, asymmetric warming among elevations may homogenize species composition in alpine grasslands (Wang et al., 2021a).
Concordant with the hypothesis, asymmetric warming among elevations might also homogenize ANPP. This phenomenon might be consequent to at least one of the subsequent mechanisms. First, our previous study (Wang et al., 2021a) and this study showed that asymmetric warming among elevations can homogenize plant α- and β-diversity. As is well known, both plant α- and β-diversity can be closely correlated with ANPP (Wu et al., 2014; Wang et al., 2021a). This phenomenon was strengthened by this study (Figures 2, 3). Second, climate change can significantly affect soil moisture (Wang and Fu, 2023) and asymmetric warming among elevations can homogenize temperature and water availability among elevations (Wang et al., 2021b). Ambient temperature and humidity conditions can influence plant phenology (Han et al., 2023), which are closely correlated with ANPP in alpine grasslands of the “Third Pole of the Earth” (Fu and Shen, 2022; Wang et al., 2022). Ambient temperature and humidity conditions can also impact plant net photosynthesis rate of the “Third Pole of the Earth” (Fu et al., 2015a). Temperature can also impact the allocation of photosynthetic products between plant aboveground and belowground parts (Friend and Woodward, 1990; Weih and Karlsson, 2001). Temperature sensitivities and precipitation sensitivities of plant production can be closely related to ranges of temperature and precipitation in alpine grasslands of the “Third Pole of the Earth” (Fu et al., 2018; Fu and Sun, 2022). Temperature sensitivities of plant production may decline with warming magnitude and may not be significantly correlated with elevation in alpine grasslands (Fu et al., 2018; Fu and Sun, 2022), which in turn may further imply that asymmetric warming among elevations may homogenize the temperature sensitivities of plant production in alpine grasslands. Third, asymmetric warming among elevations may homogenize soil nitrogen and phosphorus availability among elevations (Wang et al., 2021b), and nitrogen input can generally increase plant biomass of the “Third Pole of the Earth” (Fu and Shen, 2016; Fu et al., 2019).
This study and our previous study (Wang et al., 2021a) exhibited that symmetric warming among elevations may overestimate the impact of asymmetric warming among elevations on ANPP. Moreover, the reaction of plant α-diversity to asymmetric warming among elevations was different from that to symmetric warming among elevations (Wang et al., 2021a). This phenomenon might be consequent to at least one of the subsequent mechanisms. First, each plant species may have its own elevation range and optimum elevation, and climate change can result in the upward of the optimum elevation for a specific plant species (Lenoir et al., 2008). However, asymmetric warming and symmetric warming among elevations may cause different potentials of elevation upward. Second, asymmetric warming and symmetric warming among elevations may have different impacts on gale days and wind speed (Xiaomei et al., 2012; Yao and Li, 2019), and in turn the dispersal ability for wind-pollination plants (Wang et al., 2022). Third, asymmetric warming but not symmetric warming among elevations may homogenize environmental conditions among elevations (Wang et al., 2021a,b). Fourth, the reaction of plant β-diversity to asymmetric warming among elevations may underestimate that to symmetric warming in the identical alpine grasslands (Wang et al., 2021a,b).
Some previous studies found that the warming effects on plant production and α-diversity varied with years in alpine grasslands on the Tibetan Plateau (Wang et al., 2012; Zhang et al., 2021). Similar with these some previous studies, the effects of asymmetric warming among elevations on plant ANPP, species and phylogenetic α-diversity can vary with years. This phenomenon might be consequent to the fact that the effects of asymmetric warming among elevations on water availability, plant species and phylogenetic β-diversity, soil nitrogen and phosphorus availability, and soil pH varied with years (Wang et al., 2021b). Moreover, the effects of warming on plant production and α-diversity can be related to the background values of climatic conditions (Zhang et al., 2021; Fu and Sun, 2022), and climate conditions generally change with years (Wang et al., 2022; Han et al., 2022a; Wang and Fu, 2023). Third, the effects of warming on plant production and α-diversity can be also related to warming duration, and warming may have lagging effects on plant production and α-diversity (Fu et al., 2019; Wang et al., 2021a; Fu and Shen, 2022; Han et al., 2023). Fourthly, the effects of warming on plant production can be also related to its effects on plant precipitation use efficiency, which varied with years (Han et al., 2023). Fifthly, the effects of warming on plant production and α-diversity can be also related to its effects on soil microbial community structure, which also varied with years (Yu et al., 2019a; Zhang et al., 2021; Zhang and Fu, 2021).
Indeed, there may be some flaws in our study. For example, only two grassland types and only a relatively narrow elevation range (about 4,313–4,693 m) were included in our study (Wang et al., 2021b). These uncertainties implied that when the findings observed by this current study were extrapolated to the regions outside our study area, it needed to be careful. Therefore, we need to further carry out related studies in regions outside our study area to explore these uncertainties. However, our findings demonstrated that asymmetric warming among elevations can homogenize ANPP and plant α-diversity among elevations, at least in our study area.
5. Conclusion
Dependent on a decade experiment of asymmetric warming among elevations, this study discovered that asymmetric warming among elevations homogenized aboveground net primary production, plant phylogenetic and species α-diversity among elevations.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
GF and FH: conceptualization, formal analysis, writing – original draft preparation, and writing – review and editing. GF: methodology, validation, investigation, and data curation. CY: software, resources, project administration, and funding acquisition. FH: visualization and supervision. All authors contributed to the article and approved the submitted version.
Funding
This research was funded by the Youth Innovation Promotion Association of the Chinese Academy of Sciences (2020054), Pilot project of Chinese Academy of Sciences (XDA26050501), the National Natural Science Foundation of China (31600432), the Bingwei Outstanding Young Talents Program of the Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences (2018RC202), the Science and Technology Project of the Tibet Autonomous Region (XZ202101ZD0003N, XZ202101ZD0007G, XZ202201ZY0003N, XZ202202YD0009C, and XZ202301YD0012C), the construction of the fixed Observation and Experimental Station of the first Support System for Agricultural Green Development in Zhongba County, and the Study on the Path of Agricultural Green Development and Carbon Reduction and Sequestration in Typical Counties of Yarlung Zangbo River Basin.
Acknowledgments
We would like to thank the reviewers and editors for their valuable suggestions.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2023.1126651/full#supplementary-material
References
Bokhorst, S., Bjerke, J. W., Street, L. E., Callaghan, T. V., and Phoenix, G. K. (2011). Impacts of multiple extreme winter warming events on sub-Arctic heathland: phenology, reproduction, growth, and CO2 flux responses. Glob. Chang. Biol. 17, 2817–2830. doi: 10.1111/j.1365-2486.2011.02424.x
Brathen, K. A., Gonzalez, V. T., and Yoccoz, N. G. (2018). Gatekeepers to the effects of climate warming? Niche construction restricts plant community changes along a temperature gradient. Perspect. Plant Ecol. Evol. Syst. 30, 71–81. doi: 10.1016/j.ppees.2017.06.005
Du, M. Y., Liu, J., Li, Y., Zhang, F., Zhao, L., Niu, B., et al. (2019). Are high altitudinal regions warming faster than lower elevations on the Tibetan plateau? Int. J. Glob. Warm. 18, 363–384. doi: 10.1504/IJGW.2019.101094
Friend, A., and Woodward, F. (1990). Evolutionary and ecophysiological responses of mountain plants to the growing season environment. Adv. Ecol. Res. 20, 59–124. doi: 10.1016/S0065-2504(08)60053-7
Fu, G., and Shen, Z. X. (2016). Response of alpine plants to nitrogen addition on the Tibetan plateau: a meta-analysis. J. Plant Growth Regul. 35, 974–979. doi: 10.1007/s00344-016-9595-0
Fu, G., and Shen, Z. (2022). Asymmetrical warming of growing/non-growing season increases soil respiration during growing season in an alpine meadow. Sci. Total Environ. 812:152591. doi: 10.1016/j.scitotenv.2021.152591
Fu, G., Shen, Z. X., Sun, W., Zhong, Z. M., Zhang, X. Z., and Zhou, Y. T. (2015a). A meta-analysis of the effects of experimental warming on plant physiology and growth on the Tibetan plateau. J. Plant Growth Regul. 34, 57–65. doi: 10.1007/s00344-014-9442-0
Fu, G., Shen, Z. X., and Zhang, X. Z. (2018). Increased precipitation has stronger effects on plant production of an alpine meadow than does experimental warming in the Northern Tibetan plateau. Agric. For. Meteorol. 249, 11–21. doi: 10.1016/j.agrformet.2017.11.017
Fu, G., and Sun, W. (2022). Temperature sensitivities of vegetation indices and aboveground biomass are primarily linked with warming magnitude in high-cold grasslands. Sci. Total Environ. 843:157002. doi: 10.1016/j.scitotenv.2022.157002
Fu, G., Sun, W., Yu, C. Q., Zhang, X. Z., Shen, Z. X., Li, Y. L., et al. (2015b). Clipping alters the response of biomass production to experimental warming: a case study in an alpine meadow on the Tibetan plateau, China. J. Mt. Sci. 12, 935–942. doi: 10.1007/s11629-014-3035-z
Fu, G., Wang, J., and Li, S. (2022). Response of forage nutritional quality to climate change and human activities in alpine grasslands. Sci. Total Environ. 845:157552. doi: 10.1016/j.scitotenv.2022.157552
Fu, G., Zhang, H. R., and Sun, W. (2019). Response of plant production to growing/non-growing season asymmetric warming in an alpine meadow of the Northern Tibetan plateau. Sci. Total Environ. 650, 2666–2673. doi: 10.1016/j.scitotenv.2018.09.384
Han, F., Fu, G., Yu, C., and Wang, S. (2022a). Modeling nutrition quality and storage of forage using climate data and normalized-difference vegetation index in alpine grasslands. Remote Sens. 14. doi: 10.3390/rs14143410
Han, F. S., Yu, C. Q., and Fu, G. (2022b). Warming alters elevation distributions of soil bacterial and fungal communities in alpine grasslands. Glob. Ecol. Conserv. 39:e02306. doi: 10.1016/j.gecco.2022.e02306
Han, F., Yu, C., and Fu, G. (2023). Non-growing/growing season non-uniform-warming increases precipitation use efficiency but reduces its temporal stability in an alpine meadow. Front. Plant Sci. 14:1090204. doi: 10.3389/fpls.2023.1090204
Ji, S. N., Classen, A. T., Zhang, Z. H., and He, J. S. (2017). Asymmetric winter warming advanced plant phenology to a greater extent than symmetric warming in an alpine meadow. Funct. Ecol. 31, 2147–2156. doi: 10.1111/1365-2435.12909
Klein, J. A., Harte, J., and Zhao, X. Q. (2004). Experimental warming causes large and rapid species loss, dampened by simulated grazing, on the Tibetan plateau. Ecol. Lett. 7, 1170–1179. doi: 10.1111/j.1461-0248.2004.00677.x
Lenoir, J., Gegout, J. C., Marquet, P. A., de Ruffray, P., and Brisse, H. (2008). A significant upward shift in plant species optimum elevation during the 20th century. Science 320, 1768–1771. doi: 10.1126/science.1156831
Qin, J., Yang, K., Liang, S. L., and Guo, X. F. (2009). The altitudinal dependence of recent rapid warming over the Tibetan plateau. Clim. Chang. 97, 321–327. doi: 10.1007/s10584-009-9733-9
Rangwala, I., and Miller, J. R. (2012). Climate change in mountains: a review of elevation-dependent warming and its possible causes. Clim. Chang. 114, 527–547. doi: 10.1007/s10584-012-0419-3
Sun, W., Li, S., Wang, J., and Fu, G. (2021). Effects of grazing on plant species and phylogenetic diversity in alpine grasslands, Northern Tibet. Ecol. Eng. 170:106331. doi: 10.1016/j.ecoleng.2021.106331
Thuiller, W., Lavorel, S., and Araujo, M. B. (2005). Niche properties and geographical extent as predictors of species sensitivity to climate change. Glob. Ecol. Biogeogr. 14, 347–357. doi: 10.1111/j.1466-822X.2005.00162.x
Tian, Y., and Fu, G. (2022). Quantifying plant species α-diversity using normalized difference vegetation index and climate data in alpine grasslands. Remote Sens. 14. doi: 10.3390/rs14195007
Tozzi, E., Lyons, E. M., and Van Acker, R. C. (2014). The effect of simulated winter warming spells on Canada fleabane [Conyza canadensis (L.) Cronq. var.canadensis] seeds and plants. Can. J. Plant Sci. 94, 963–969. doi: 10.4141/cjps2013-302
Trisos, C. H., Merow, C., and Pigot, A. L. (2020). The projected timing of abrupt ecological disruption from climate change. Nature 580, 496–501. doi: 10.1038/s41586-020-2189-9
Wang, S. P., Duan, J., Xu, G., Wang, Y., Zhang, Z., Rui, Y., et al. (2012). Effects of warming and grazing on soil N availability, species composition, and ANPP in an alpine meadow. Ecology 93, 2365–2376. doi: 10.1890/11-1408.1
Wang, S., and Fu, G. (2023). Modelling soil moisture using climate data and normalized difference vegetation index based on nine algorithms in alpine grasslands. Front. Environ. Sci. 11:1130448. doi: 10.3389/fenvs.2023.1130448
Wang, J., Li, M., Yu, C., and Fu, G. (2022). The change in environmental variables linked to climate change has a stronger effect on aboveground net primary productivity than does phenological change in alpine grasslands. Front. Plant Sci. 12:798633. doi: 10.3389/fpls.2021.798633
Wang, J., Yu, C., and Fu, G. (2021a). Warming reconstructs the elevation distributions of aboveground net primary production, plant species and phylogenetic diversity in alpine grasslands. Ecol. Indic. 133:108355. doi: 10.1016/j.ecolind.2021.108355
Wang, J. W., Yu, C. Q., and Fu, G. (2021b). Asymmetrical warming between elevations may result in similar plant community composition between elevations in alpine grasslands. Front. Ecol. Evol. 9:757943. doi: 10.3389/fevo.2021.757943
Weih, M., and Karlsson, P. S. (2001). Growth response of mountain birch to air and soil temperature: is increasing leaf-nitrogen content an acclimation to lower air temperature? New Phytol. 150, 147–155. doi: 10.1046/j.1469-8137.2001.00078.x
Wu, J. S., Shen, Z. X., and Zhang, X. Z. (2014). Precipitation and species composition primarily determine the diversity-productivity relationship of alpine grasslands on the northern Tibetan plateau. Alp. Bot. 124, 13–25. doi: 10.1007/s00035-014-0125-z
Xiaomei, Y., Zongxing, L., Qi, F., Yuanqing, H., Wenlin, A., Wei, Z., et al. (2012). The decreasing wind speed in southwestern China during 1969-2009, and possible causes. Quat. Int. 263, 71–84. doi: 10.1016/j.quaint.2012.02.020
Yang, Z. L., Zhang, Q., Su, F., Zhang, C., Pu, Z., Xia, J., et al. (2017). Daytime warming lowers community temporal stability by reducing the abundance of dominant, stable species. Glob. Chang. Biol. 23, 154–163. doi: 10.1111/gcb.13391
Yao, H. R., and Li, D. L. (2019). The gale concentration period and degree over the Tibetan plateau and related atmospheric circulation during the windy period. J. Desert Res. 39, 122–133.
Yao, T. D., Liu, X. D., Wang, N. L., and Shi, Y. F. (2000). Amplitude of climatic changes in Qinghai-Tibetan plateau. Chin. Sci. Bull. 45, 1236–1243. doi: 10.1007/BF02886087
Yu, C. Q., Han, F. S., and Fu, G. (2019a). Effects of 7 years experimental warming on soil bacterial and fungal community structure in the northern Tibet alpine meadow at three elevations. Sci. Total Environ. 655, 814–822. doi: 10.1016/j.scitotenv.2018.11.309
Yu, C. Q., Wang, J. W., Shen, Z. X., and Fu, G. (2019b). Effects of experimental warming and increased precipitation on soil respiration in an alpine meadow in the Northern Tibetan plateau. Sci. Total Environ. 647, 1490–1497. doi: 10.1016/j.scitotenv.2018.08.111
Zha, X. J., Tian, Y., Ouzhu,, and Fu, G. (2022). Response of forage nutrient storages to grazing in alpine grasslands. Front. Plant Sci. 13:991287. doi: 10.3389/fpls.2022.991287
Zhang, H., and Fu, G. (2021). Responses of plant, soil bacterial and fungal communities to grazing vary with pasture seasons and grassland types, northern Tibet. Land Degrad. Dev. 32, 1821–1832. doi: 10.1002/ldr.3835
Zhang, X. Z., Shen, Z. X., and Fu, G. (2015). A meta-analysis of the effects of experimental warming on soil carbon and nitrogen dynamics on the Tibetan plateau. Appl. Soil Ecol. 87, 32–38. doi: 10.1016/j.apsoil.2014.11.012
Zhang, G., Shen, Z., and Fu, G. (2021). Function diversity of soil fungal community has little exclusive effects on the response of aboveground plant production to experimental warming in alpine grasslands. Appl. Soil Ecol. 168:104153. doi: 10.1016/j.apsoil.2021.104153
Zhang, G., Shen, Z., and Fu, G. (2022). Geo-distribution patterns of soil fungal community of Pennisetum flaccidum in Tibet. J. Fungi 8. doi: 10.3390/jof8111230
Zhong, Z., and Fu, G. (2022). Response of soil fungal species, phylogenetic and functional diversity to diurnal asymmetric warming in an alpine agricultural ecosystem. Agric. Ecosyst. Environ. 335:107993. doi: 10.1016/j.agee.2022.107993
Keywords: biodiversity, symmetrical warming, phylogenetic diversity, Tibetan Plateau, asymmetric warming
Citation: Han F, Yu C and Fu G (2023) Asymmetric warming among elevations may homogenize plant α-diversity and aboveground net primary production of alpine grasslands. Front. Ecol. Evol. 11:1126651. doi: 10.3389/fevo.2023.1126651
Edited by:
Zhongqing Yan, Chinese Academy of Forestry, ChinaReviewed by:
Linfeng Li, University of Chinese Academy of Sciences, ChinaChitra Bahadur Baniya, Tribhuvan University, Nepal
Copyright © 2023 Han, Yu and Fu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gang Fu, ZnVnYW5nQGlnc25yci5hYy5jbg==; ZnVnYW5nMDlAMTI2LmNvbQ==
†These authors have contributed equally to this work