 João Pedro Saldanha
João Pedro Saldanha Alisson Klayton Martins
Alisson Klayton Martins Gabriel Ladeira Osés
Gabriel Ladeira Osés Flavia Callefo
Flavia Callefo- 1Programa de Pós-graduação em Geociências, Departamento de Paleontologia e Estratigrafia, Instituto de Geociências, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil
- 2Laboratório de Arqueometria e Ciências Aplicadas ao Patrimônio Cultural, Departamento de Física Nuclear, Instituto de Física, Universidade de São Paulo, São Paulo, Brazil
- 3Laboratório Nacional de Luz Síncrotron, Linha Carnaúba, Centro Nacional de Pesquisa em Energia e Materiais, Campinas, Brazil
Biogenicity assessments in paleobiology encompass a broad range of criteria, arguments, and methodological approaches for evaluating the origin of fossil objects and purported biological materials. Amid this uncertainty, paleometry emerges as a critical tool for deepening and refining our understanding of biogenicity. This study aims to systematize procedures observed across recurring patterns in the literature, to critically examine the type of data acquired, and to expand the application of high-resolution analytical techniques. Rather than debating optimal criteria, we propose a direct and structured framework for biogenicity assessment, outlining standardized steps, choosing the most powerful techniques for solving specific issues, result interpretation, and scientific discussion. Biogenicity-related studies are categorized into three major groups: (1) dubiofossils, including microfossil-, ichnofossil-, and biomineral-like forms; (2) fossilization processes involving microbial mediation; and (3) potential biominerals associated with skeletal metazoans. Despite their differences, these categories share a fundamental reliance on reconstructing diagenetic history, a prerequisite for any meaningful biogenicity inference. In this context, paleometry plays a central role across all stages of investigation. Our analysis highlights the widespread use of high-resolution, non-destructive techniques such as optical microscopy (OM), scanning electron microscopy with energy-dispersive spectroscopy (SEM-EDS), micro-Raman spectroscopy (µ-RS), and Fourier-transform infrared spectroscopy (FT-IR), largely applied for their minimal sample preparation and versatile micrometer-scale capabilities. We also propose the utilization of other techniques, still underexplored in biogenicity studies. Furthermore, the temporal distribution of case studies reveals a concentration of research on geologic intervals associated with key evolutionary and environmental transitions, which we argue the distribution of biogenicity studies in such periods. The protocols proposed herein offer a flexible and reproducible methodological pathway. By guiding researchers from analytical technique selection to data interpretation, this framework aims to facilitate future investigations and foster more coherent discussions around the biogenicity of paleobiological materials.
1 Introduction
The term biogenicity carries diverse connotations and applications across various scientific domains (Malaterre et al., 2023). Within paleontology, it denotes the recognition of an object, substance, and/or pattern whose origin specifically involves a biological agent (Des Marais et al., 2008). This encompasses processes exclusively tied to biological activity and the assessment of the preservation of biomaterials in the face of post-depositional transformations. Consequently, the detection of life in deep time and/or distant space is exceedingly complex and necessitates caution (McMahon et al., 2021; McMahon and Cosmidis, 2022; Malaterre et al., 2023). In both astrobiology and paleontology (mainly in Precambrian research), discussions on biogenicity revolve around specific sets of arguments formulated and tested in particular cases, designated as biogenicity criteria, which are in constant debate (Buick, 1990; García-Ruiz et al., 2002; Brasier et al., 2002; Schopf et al., 2002; Cady et al., 2003; Brasier et al., 2004; Sugitani et al., 2007; Westall, 2008; Noffke, 2009, 2021; Slater, 2009; Wacey, 2010; Brasier and Wacey, 2012; Schopf and Kudryavtsev, 2012; Bernard and Papineau, 2014; McLoughlin and Grosch, 2015; Callefo et al., 2019a; Gomes et al., 2019; Neveu et al., 2018; McMahon et al., 2021; Rouillard et al., 2021). Within this framework, paleometry – a transdisciplinary effort that aims to develop and combine more effective and efficient techniques for recovering information from the fossil record (Riquelme et al., 2009; Delgado et al., 2014) – has played a pivotal role from the very beginning. Whether through the implementation of novel technologies, the refinement of data acquisition processes, or the integration of results, paleometry stands as a potential key to distinguishing life across time and space (Riquelme et al., 2009; Delgado et al., 2014; Callefo et al., 2019a; Gomes et al., 2019; Prado et al., 2021).
Despite advancements in formulating and discussing biosignatures in recent decades, there is a lack of protocols and biogenicity tests that could formalize and propel scientific progress further (see Nevue et al., 2018 and Rouillard et al., 2021). This is particularly evident given the diversity of objects and objectives in astrobiological and paleontological sciences (Callefo et al., 2019a). Despite the need to be protocolizing procedures to minimize misinterpretations, these parameters must be constantly reevaluated, given the frequent advances in understanding the different biosignatures, and the advancement of new technologies (especially at subcellular scales and high resolution), which enable the development of knowledge about the structure and composition of materials. Furthermore, the very understanding of what is life, and its evolution are also concepts in constant progress. And obviously, this impacts the very understanding and interpretation of biosignatures and their biogenicity parameters.
The challenges surrounding biogenicity issues can be categorized into four main categories: 1) dubiofossils, forms of uncertain origin; 2) fossilization processes via microbial activity; 3) minerals from candidate biomineralizing metazoans; and 4) chemical record or compositional influence in abiotic processes. Each category exhibits unique characteristics, distinct scientific interests, and varied study methodologies. Once it focusses on different objects and objectives, research within these categories tends to be concentrated on specific geological periods, resulting in a distinct temporal range.
Given the diverse research areas in paleontology, as well as the uniqueness of the fossil record, this contribution aims to attempt to formalize specific protocols for biogenicity assessment based on the current state of knowledge. It focuses on systematizing procedures compiled from research articles, in a critical way, aiming to document how biogenicity is assessed in different areas, possible causes for that, and consequently debating future research avenues. The compiled data is compared against the geological timescale, which offers interesting insights on the distribution of biogenicity studies over time. Rather than debating the best biogenicity criteria, this work provides a direct and assertive approach by outlining standardized steps for data acquisition, result interpretation, and discussion.
2 Methods and definitions
To achieve biogenicity protocols, case study articles in paleontology, geobiology, geosciences and related areas from various biogenicity themes were compiled, detailing the equipment used and the primary data acquired. Analyzing the results and interpretations allowed the recognition of patterns through recurring discussion styles, which culminated in the proposal of standardized protocols for understanding biogenicity in the respective research areas.
The bibliographic survey for this review was conducted in two complementary stages. The first stage involved identifying key references cited in two categories of publications: (1) highly cited and widely recognized articles considered classic in the discussion of biogenicity criteria, and (2) review articles specifically addressing biogenicity criteria. These initial sources were identified through searches using the keywords “biogenicity” and “criteria”, which served as the starting point for mapping the field. From this initial mapping, two main groups of articles were established: (a) Review articles on biogenicity criteria focused on specific material types (e.g., microfossils, ichnofossils, dubiofossils), resulting in more study case articles; (b) Case study articles applying biogenicity criteria to distinct material types.
In the second stage, this survey was expanded through systematic searches in the electronic databases Scopus, Web of Science, and Google Scholar, conducted between July 2022 and March 2025. Boolean operators (AND, OR) were applied to combine both general and specific keywords. The general search terms included: “biogenicity”, “biogenicity criteria”, “biosignatures”, “biotic”, “abiotic”, “bioindicator”, “fossil diagenesis”, and “paleometry”. These were combined with specific terms for each thematic focus: “dubiofossils”, “pseudofossils”, “ichnofossil”, “microfossils”, “artifact”, “biomineral”, “abiotic mineralization”, “induced mineralization”, “influenced mineralization”, “controlled mineralization”, “phosphatization”, “pyritization”, “silicification”, “concretion”, “biofilm”, “microbial mat”, “biomineralizing”, “biometazoans”, and “preservation”. Experimental taphonomy articles were selectively included in the fossilization processes category when they employed high-resolution analytical techniques or explicitly discussed fossil biogenicity criteria. Articles exclusively addressing chemical fossils or chemical fossil bioindicators were excluded, as these fell outside the scope of this review.
This contribution does not include chemical bioindicators. Due to their distinct nature as chemical signals, they are different from fossil objects and artifacts that are investigated in other biogenicity challenges categories and allow for a more direct comparison. In addition, there is a vast and growing literature on fossil bioindicators that requires greater attention in a future article.
Here we present a list of definitions with explanations utilized to categorize and quantify the articles reviewed.
Techniques: Several articles discuss the application and limitations of high-resolution or non-destructive techniques in paleontology (Carlson et al., 2003; Delgado et al., 2014; Marshall and Marshall, 2015; Cusack, 2016; Borgwardt and Wells, 2017; Callefo et al., 2019a; Gomes et al., 2019; Pan et al., 2019; Prado et al., 2021). Abbreviations are explained in Supplementary Material 1. The techniques were grouped to facilitate categorization based on similar responses or shared applications (Supplementary Materials 2, 3). This grouping does not imply identical data acquisition methods nor underlying physical principles.
Biogenicity categories: biogenicity in paleontology is a subject of research in several objects that we categorize here into four main groups:
1: Dubiofossils: fossil-like objects formerly related to life with an ambiguous origin (Hofmann, 1972). Biogenicity criteria are proposed that allow access to their abiotic or biotic origin. Through testing, many times through paleometry, the biological nature of a dubiofossil can be established, leading to its classification as a genuine fossil; alternatively, if its origin is determined to be the result of abiotic processes, it is categorized as a pseudofossil (Hofmann, 1972; Monroe and Dietrich, 1990; McMahon et al., 2021). After the survey, we categorized the dubiofossils into three types of material classes:
1a: microfossil-like dubiofossils: generally elongated/filamentous or globular/spherical objects ranging from nanometric to microscopic scales that morphologically resemble a microfossil.
1b: ichnofossil-like dubiofossils: structures that resemble bioturbations, microbially induced sedimentary structures (MISS), or stromatolites and can be associated with ichnofossils.
1c: biomineral-like dubiofossils: branched, rounded, unusual, and complex mineral shapes that could be comparable to those produced by controlled, induced or influenced biomineralizations processes (see Dupraz et al., 2009). These materials are unusual mineral forms, which due to their shape and complexity may suggest biotic origin.
2: Fossilization processes: biogenicity of fossilization process means that via microbial activity was responsible for fossil preservation. Fossils with materials non-original from organisms that may have been precipitated, recrystallized or modified with microbiological activity. Classification by fossilization modes:
2a: silicification: silica precipitation.
2b: preservation by carbonates: calcite, aragonite, dolomite or other type of carbonate minerals that help in fossilization.
2c: concretions: hard, compact mass of mineral, often spherical or oval in shape around a fossil or fossil fragment within sedimentary host rock.
2d: phosphatization: phosphate minerals precipitated or recrystallized during fossilization.
2e: preservation by clay minerals: clay minerals preserving fossils.
2f: pyritization: pyrite precipitation.
2g: biofilms: biofilm evidence preserved along with fossils.
2h: Iron (oxyhydr)oxides: iron-based oxyhydroxides precipitation.
3: Biomineralizing metazoans: animals able to form skeletons with a mineral component, the so-called biominerals. Thus, the terms biomineral and biomineralization differ from that of class 1c.
4: Bioindicators: diagnostic compositions that could only have occurred in the presence of life, or that at least have a greater chance of occurrence due to biotic origin. They can be isolated elements, isotopic fractionation values or molecules/minerals that are specific indicators of the presence of life in deep time and on other planets. Bioindicators are not integrated into the review and discussions.
Data: type of data that can be obtained from each technique. Classified into Morphological, Texture or Composition.
Morphological: morphological data divided into superficial or internal data;
S: superficial morphological data;
I: internal morphological data;
Texture: spatial data associated with a group of crystals, like distribution, orientation, packing and inclusion;
T: textural spatial data, topological or organizational distribution regarding the host rock;
M: spatial data composition, mapping and distribution of the composition relative to the matrix;
Composition: compositional data;
C: inorganic compositional data, including elemental or molecular/mineralogical data;
O: organic compositional data, including elemental or molecular data; Note: these data refer to the detection of organic molecules, such as hydrocarbon chains and carbon detection, and have no connotation of the origin, whether biotic or abiotic, of the material.
i: detailed composition, trace elements or isotopic data;
s: detailed organic compositional data, including protein and DNA characterization or genetic sequencing.
Result: characteristics extracted from the studied material obtained through some high-resolution technique. Here it is categorized within a data type.
Interpretation: product of the results, an interpretation of the identified attributes that can serve as criteria for evaluating biogenicity.
Discussion: pattern of argumentation and integration of information that culminates in the characterization of biogenicity. Here it is summarized from the repetition of discussion styles found in the bibliographic survey.
Biosignature: signatures exclusively generated and/or transformed by past or present organisms that unambiguously diagnose life. These signatures can be created from the growth or decay of (once) living organisms and cannot be produced by purely abiotic processes (Slater, 2009; McLoughlin, 2011).
Biogenicity criteria: arguments proposed to defend or refute the biotic origin of a given object, substance or process (see Buick, 1990; Gargaud et al., 2015).
Biotic: substances, materials, or processes that are derived from or involve living entities.
Abiotic: substances, materials, or processes that are derived from non-living entities (for example, physical, and chemical processes without biological participation).
Pseudofossil: objects that appear, at least at first glance, to be fossils, but after more refined analysis are considered to be of abiotic origin, that is, they are not fossils (Monroe and Dietrich, 1990).
Indigeneity: refers to the origin of the material, with an emphasis on eliminating the possibility of recent or procedural contamination through cross-cutting relationships (Gargaud et al., 2015; Rouillard et al., 2021).
Syngenicity: material’s temporal relationship with its surrounding matrix, confirming synchrony with the host medium (Buick, 1990; Wacey, 2009; McLoughlin and Grosch, 2015; Gargaud et al., 2015; Rouillard et al., 2021).
Extracellular polymeric substance (EPS): polymeric substance biosynthesized by microorganisms which contain mainly polysaccharides, genetic material (i.e., DNA) and proteins, offering several benefits to microbes, including protection against external agents, and the enhancement of metabolic changes inside biofilms (Flemming and Wingender, 2010).
Controlled biominerals: minerals that are directly produced and regulated by living organisms that exercise a high level of control over their formation and composition (Dupraz et al., 2009).
Induced biominerals: minerals indirectly formed by living organisms; these play an active role in triggering or influencing their formation, producing certain organic compounds or creating specific environmental conditions, often as an indirect result of the metabolic action (Dupraz et al., 2009).
Influenced biominerals: minerals in which the presence of living or dead organisms plays a passive role in their formation or modification (see Dupraz et al., 2009, for a broader review).
Abiotic minerals: by exclusion, abiotic minerals are the result of physicochemical reactions without any biological interference (see Dupraz et al., 2009, for a broader review).
Supplementary materials: The supplementary material presents crucial details of the review and quantification. Supplementary Material 1 – presents the techniques found in the review, organized by abbreviations used in the rest of the text, date, results and which classes have the greatest application. Supplementary Material 2 – presents all case study articles compiled in the review. Supplementary Material 3 – presents tables of quantification of data by date and technique by class from Supplementary Material 2. Supplementary Material 4 – reference list of all articles cited in the text and or compiled in the review. Supplemental Material 2 and 3 were used to base the figures and tables in the main text.
3 Biogenicity for paleobiology
A total of 85 case study articles were collected and are presented in Supplementary Material 2, classified by age, main contribution, techniques used, and main data obtained. In general, the techniques of OM, SEM-EDS/FIB-SEM, and RS/FT-IR are widely used in all categories of biogenicity categories, and are more relevant for categories 1, 2, and 3, respectively, as can be seen in Table 1. A summary of the quantification is presented in Supplementary Material 3. It is important to note that more than 3/4 of the articles apply more than one high-resolution technique with the aim of complementing the data obtained. Other well-used techniques are TEM, Cryo-TEM, FIB-TEM; XRF, WD-XRF, SR-XRF; XRD, XPS, EBSD; elemental analysis, isotopic analysis, MS, LA-ICP-MS, ICP-MS, GC, GC-MS, NanoSIMS, (Tabel 1).
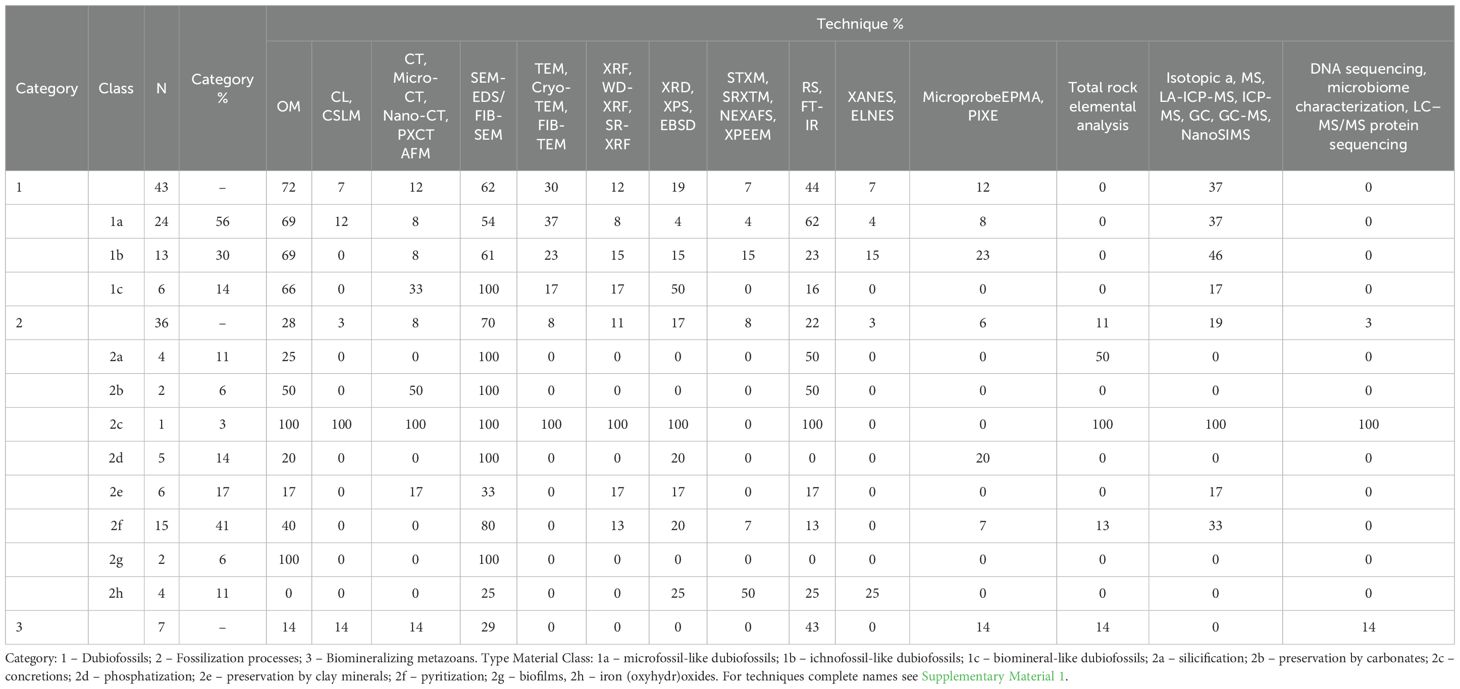
Table 1. Percentage of articles that used each type of technic by category and class of material type (percentages refer to the quantity within the class and not to the total number of articles).
Regarding the type of data acquired and used in discussions of biogenicity in paleontology, it is clear that data on inorganic composition (C) and surface (S) and internal (I) morphology are the most relevent in all categories (Table 2; Figure 1). For dubiofossils, I and C are more pertinent than for biogenicity in fossilization processes, which require more S characterization, and for biomineralizers, being scrutinized by S, C and O. Despite this, the survey indicates that more than one type of data is always necessary for any inference and discussion of biogenicity. Surface (S) and internal morphological (I) and inorganic compositional data (C) are extremely relevant for all categories and, in some classes, data on texture, organic distribution and composition and chemical detailed characterization are valuable (Table 2). Compositional mapping (M) is less relevant for biomineralizing metazoans, detailed organic compositional data (s) appears to have little application in discussing the biogenicity of dubiofossils in general (Figure 1). From the data, results and interpretations are inferred and each category has a distinct pattern in discussing biogenicity, as described below.
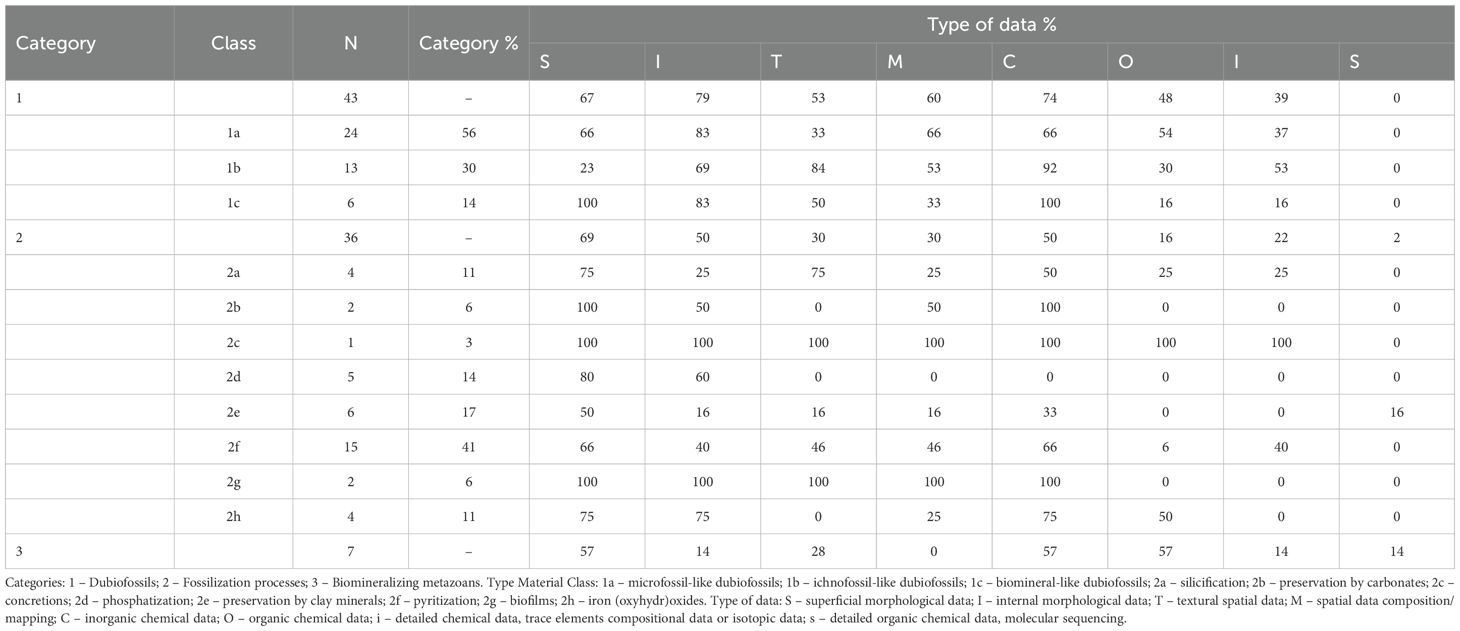
Table 2. Percentage of articles that obtained each type of data (results) by category and class of material type (percentages refer to the quantity within the class and not to the total number of articles).
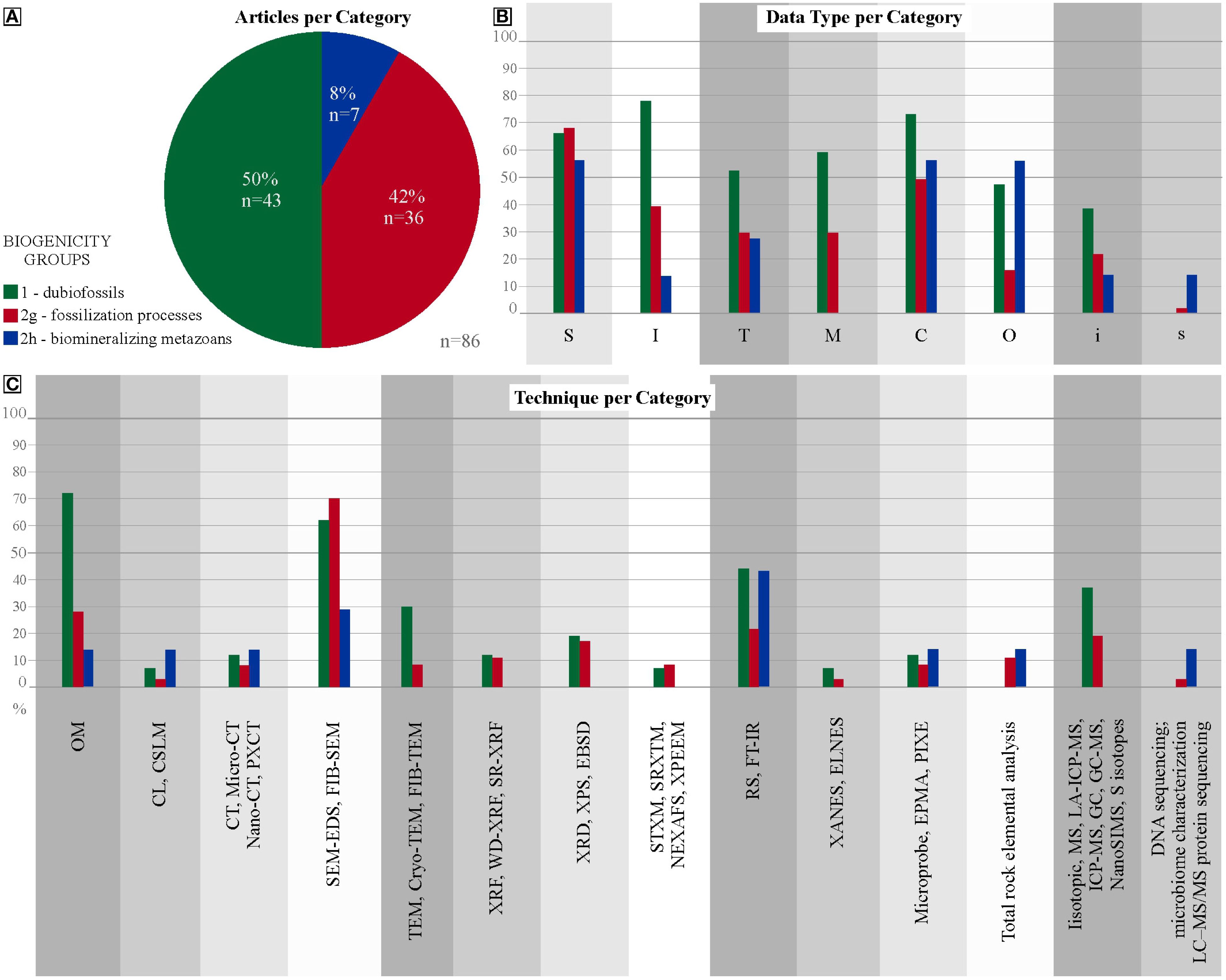
Figure 1. Distribution of all articles on biogenicity. (A) Comparative plot of the percentages of articles reviewed by each Category: 1 –Dubiofossils; 2 – Fossilization processes; 3 – Biomineralizing metazoans; (B) Percentages of each Category that obtain each type of data; (C) Percentages each Category that apply each type of technique.
3.1 Dubiofossils
Category 1 comprises dubiofossils, structures resembling fossils that were previously associated with life but possess an ambiguous origin (Hofmann, 1972), Supplementary Material 2 presents all articles compiled about dubiofossils in relation to the data and applied techniques, which were quantified in Supplementary Material 3. These dubiofossils are categorized into three classes (Figure 2A). The microfossil-like dubiofossils class (1a) has substantial biogenicity criteria, postulated and revised by Buick (1990); Cady et al. (2003); Benzerara and Menguy (2009); Schopf et al. (2010); Brasier and Wacey (2012); McMahon et al. (2021), and Rouillard et al. (2021). These criteria encompass internal morphology (I) – approximately 83% of the total 1a articles; external structure (S), inorganic composition (C) data and mapping (M) – both with roughly 66% of the total of 1a (Figure 2B). For ichnofossil-like dubiofossils (1b), the biogenicity criteria are outlined by Awramik and Grey (2005); Porada et al. (2008); Noffke (2009, 2010, 2021); Wacey (2010); Noffke and Awramik (2013); Davies et al. (2016), Davies et al. (2020) and Lucas and Lerner (2017), Sanchez et al. (2023), which reveal that it is important to characterize data on inorganic composition (C) – almost 92% of the total 1b articles; and texture (T) – approximately 84% of the 1b articles, supported by internal morphological characteristics (I) – roughly 69% of the 1b (Figure 2B). The biomineral-like dubiofossils (1c) lacks well established biogenicity criteria discussed by García-Ruiz et al. (2002); Braissant et al. (2003); Weiner and Dove (2003); Dupraz et al. (2009); Benzerara and Menguy (2009); Cailleau et al. (2009); Bindschedler et al. (2014); McMahon and Comidis (2022), and Saldanha et al. (2023). This highlight the need to obtain morphological data (S and I) – all 1c articles used S data and approximately 83% I data; and inorganic compositional data (C) – all 1c articles. In respect to texture (T) – half of the articles assessed this; and Compositional mapping (M) was performed by roughly 33% of the total 1c articles (Figure 2B).
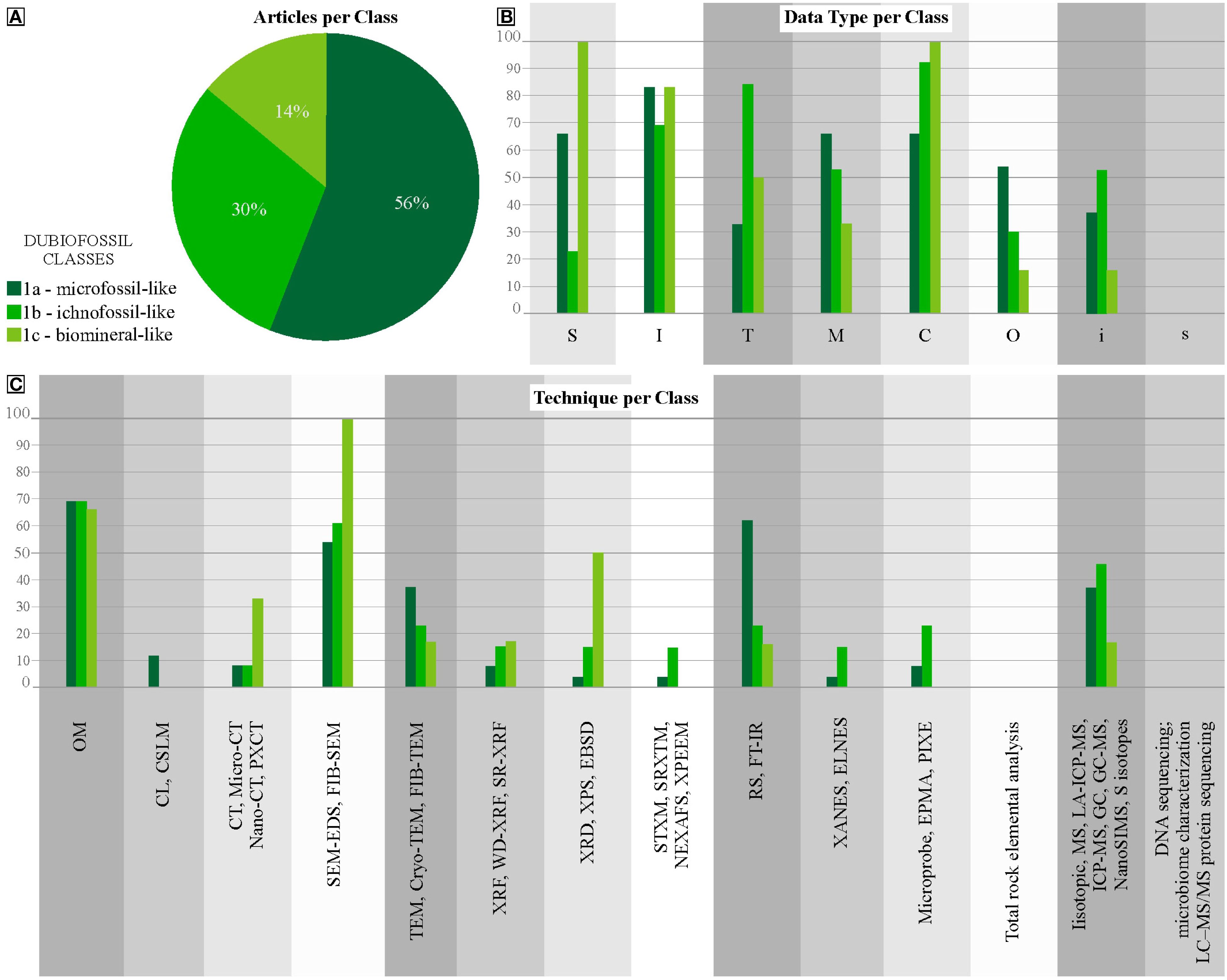
Figure 2. Distribution of Dubiofossils articles. (A) percentage of each class of the total articles in Category 1. (B) Percentages of dubiofossils classes relative to each type of data. (C) Percentages of dubiofossils classes that apply to each type of technique.
For 1a, morphology characterization is crucial, focusing on general shape, internal features, and population morphology range, best achieved through OM, SEM-EDS/FIB-SEM, nano or micro-CT (Buick, 1990; Javaux et al., 2003; Sugitani et al., 2007; McMahon et al., 2021). General size, population range, and morphometric consistency further support abiotic or biotic hypotheses (Figures 2C, 3). Details, ornamentation, and preservation textures can contribute secondarily, both obtained by nano- or micro-CT, PXCT or SEM (Hofmann et al., 2008; Dodd et al., 2017; Johannessen et al., 2020; Maldanis et al., 2020). Compositional analysis is less decisive due to potential confusion with abiotic objects (Brasier et al., 2002, 2004; Schiffbauer et al., 2007), as seen in the dispute over Apex Chert microfossils (Schopf et al., 2002; Brasier et al., 2004; Schopf and Kudryavtsev, 2012). Complementary analyses like RS, TEM, XANES, MS, LA-ICP-MS, GC-MS, are useful in such cases (Brasier et al., 2004; De Gregorio et al., 2009; Alleon et al., 2016; Dodd et al., 2017). Detecting kerogen associated with microbial morphologies may be relevant in specific contexts using RS, FT-IR or XANES (Schopf et al., 2002; 2005; Sugitani et al., 2007; Dodd et al., 2017; Gomes et al., 2019; Osterhout et al., 2021).
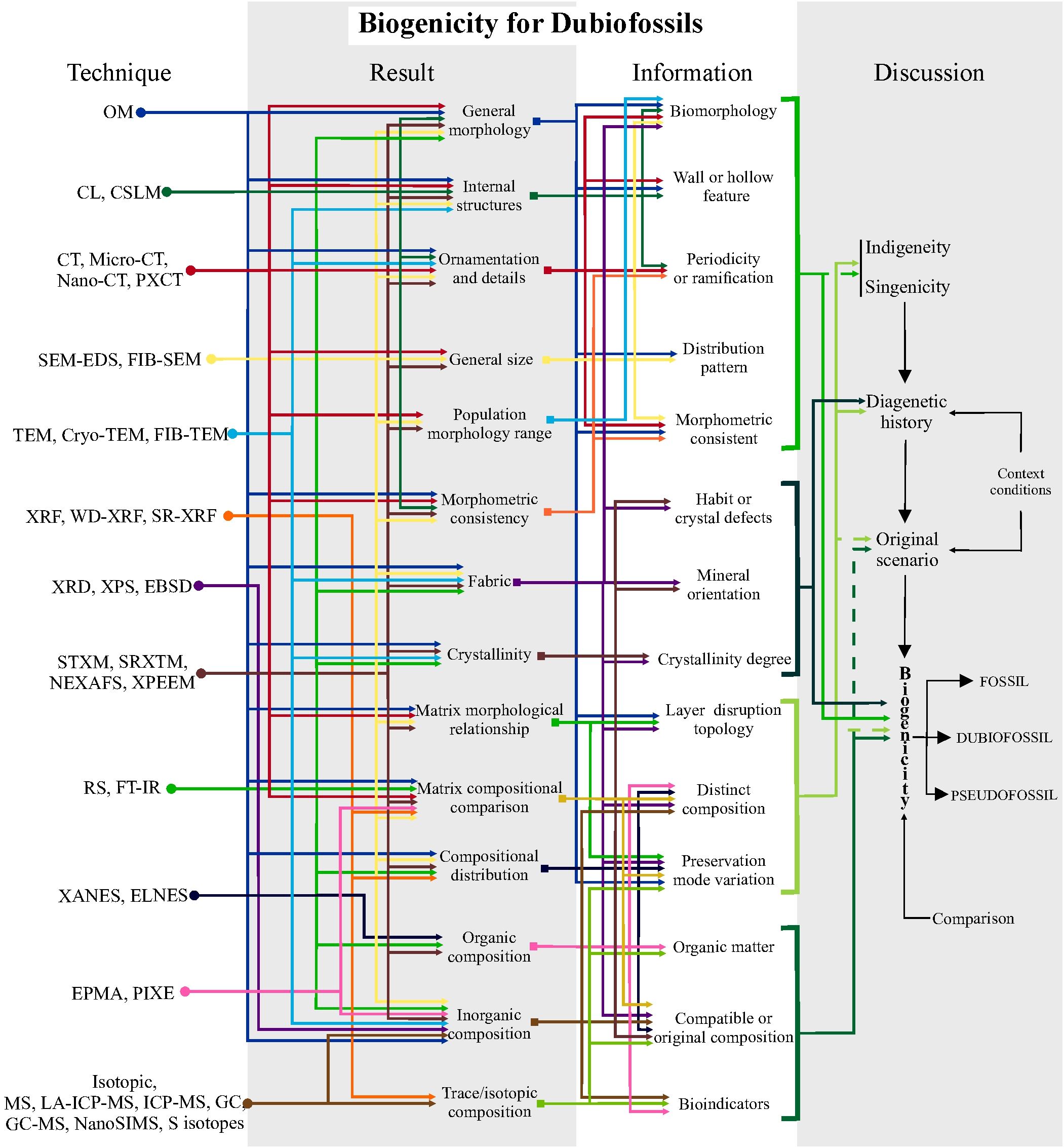
Figure 3. Schematic protocol for investigating biogenicity in Dubiofossil objects. Steps using high-resolution techniques to obtain results, information and how to discuss them. The first step involves deciphering indigeneity and syngenicity; the second, discovering the diagenetic history and assessing the original setting, involving any additional information about the geological context; and the final step is to argue and raise biogenicity criteria to propose the origin of the material, through comparison with other materials. This protocol is based on the table in Supplementary Material 2 with the high-resolution techniques used in the reviewed articles, together with their potential applications in results and information. Solid lines in the discussion of biogenicity indicate direct application of the information to the topic being discussed; dashed lines indicate indirect or lesser application to the discussion. See Supplementary Material 5 for a larger, high-resolution version.
For 1b (Figures 2C, 3), microscopic features like filamentous morphologies and putative EPS, and ornamentations, diagnosed with OM and SEM-EDS/FIB-SEM (sometimes complemented by TEM or FIB-TEM, STXM or SRXTM), may be pertinent, although macroscopic forms are of secondary relevance, and size is less crucial (Noffke et al., 2001, 2003, 2006; Porada et al., 2008; De Gregorio et al., 2009; Inglez et al., 2021). Verifying composition is important, especially for organic matter detection (RS, XANES) and bioindicator elements (EDS, XRD, XRF or SR-XRF, RS, MS) (De Gregorio et al., 2009; Wacey, 2010; Allwood et al., 2018; Callefo et al., 2019b; Alleon et al., 2021; Klug et al., 2021). Caution is advised due to the ongoing debate on reliability of biomarkers (Brasier et al., 2002; Marshall et al., 2012; Schopf and Kudryavstev, 2012; Bower et al., 2015).
For 1c (Figures 2C, 3), it is necessary to identify general morphology, internal structures, mineral habit, and crystallinity, described mainly through OM, accompanied by SEM-EDS and Micro-CT for determining general shape and texture (Cailleau et al., 2009; Bindschedler et al., 2014; Tisato et al., 2015; Green, 2022; Saldanha et al., 2023). Secondarily, mineral orientation, crystalline defects, twinning, and habits are considered (Braissant et al., 2003; Cailleau et al., 2009; Saldanha et al., 2023), with good results coming out from OM, SEM-EDS, and CT if the object differs compositionally from the matrix. The composition is key for identifying minerals and assessing diagenetic modifications, obtained through EDS, XRF, XRD, RS, MS (Braissant et al., 2003; Cailleau et al., 2009; Tisato et al., 2015).
The biogenicity arguments for 1a are based on the interpretation of biological morphologies, such as walls and hollow features, ornamentation, periodicity, and branching (Buick, 1990). This includes assessing consistent morphologies, normal distribution of population sizes, and taphonomic variations to identify globular, filamentous, or even colonial forms (Buick, 1990; Javaux et al., 2003; Sugitani et al., 2007; Maldanis et al., 2020; McMahon et al., 2021). Evidencing the organic/kerogen composition is desirable, but requires great caution in its interpretation, avoiding erroneous conceptions of abiotic origin or diagenetic modifications (Schopf et al., 2002, 2005; Brasier et al., 2002, 2004; Schiffbauer et al., 2007; Sugitani et al., 2007; Dodd et al., 2017; Gomes et al., 2019). For 1b, the biogenicity criteria are interpreted from textural relationships, diagnostics of filamentous features, and their distribution in the framework (Noffke et al., 2001). Mineral orientation may also be relevant (e.g., Noffke et al., 2001, 2003; Inglez et al., 2021). Verifying composition is important, especially for detecting organic matter and bioindicator elements (Brasier et al., 2002; Marshall et al., 2012; Schopf and Kudryavtsev, 2012; Bower et al., 2015). For 1c, the detection of crystal defects, variations in mineral habit, degree of crystallinity, and distinct matrix composition aids in discussing the biogenicity of biomineral dubiofossils (Braissant et al., 2003; Cailleau et al., 2009; Bindschedler et al., 2014; Tisato et al., 2015; Saldanha et al., 2023). Composition is key for identifying minerals and assessing diagenetic modifications (Braissant et al., 2003; Tisato et al., 2015). However, composition alone cannot serve as the main biogenicity argument, as no mineral species or characteristics are uniquely tied to life (Weiner and Dove, 2003; Benzerara and Menguy, 2009; Benzerara et al., 2019). Figure 3 summarizes a protocol of how to proceed using techniques to obtain specific data (result and information) aiming to test hypotheses (discussion).
The discussion of the biogenicity of any dubiofossil is typically conducted in four steps (compiled from Buick, 1990; García-Ruiz et al., 2002; Brasier et al., 2002; Schopf et al., 2002; Brasier et al., 2004; Westall, 2008; Noffke, 2009, 2021; Slater, 2009; Wacey, 2010; Brasier and Wacey, 2012; Schopf and Kudryavtsev, 2012; McLoughlin and Grosch, 2015; Callefo et al., 2019a; Gomes et al., 2019; Neveu et al., 2018; McMahon et al., 2021; Rouillard et al., 2021; Saldanha et al., 2023), presented in Figure 3, from the technique used to obtain the data and results relate to more specific information until reaching the steps to discuss and decipher the origin of the dubiofossil: 1) Deciphering syngenicity and indigeneity: evaluating the relationships of cutting, inclusion, and subsequent modifications to establish the timing of the processes and ensure there is no contamination (Buick, 1990; Wacey, 2009; McLoughlin and Grosch, 2015; Rouillard et al., 2021). 2) Understanding diagenetic history: detailing all the processes that may have transformed the material, throughout the entire trajectory from immediately after its deposition/formation until collection. 3) Reconstructing the original scenario: based on additional information about the environment, age, and context, this step involves hypothesizing the possible environmental processes that formed the material. 4) Integrating information: this final step involves evaluating the most plausible hypotheses about the origin of the object, discarding less likely processes. This evaluation is based mainly on the interpretations of the results and comparisons with similar modern and/or fossil objects and processes (Figure 3). It is recommended to use biotic versus abiotic hypotheses with equal weights (Hofmann, 1972; Buick, 1990; Benzerara and Menguy, 2009; McMahon et al., 2021; Rouillard et al., 2021). After the discussion, the object can be included as a fossil or a pseudofossil if it has sufficient evidence or alternatively remain as a dubiofossil (Hofmann, 1972).
3.2 Fossilization processes
It is well-established that authigenic mineralization resulting in fossilization depends on microorganisms, which decay organic matter, inducing mineral precipitation (e.g., Briggs, 2003). However, evidence for such direct mediation is rarely presented and is guided mainly by actualistic taphonomic experiments. Due to the number of articles on the various modes of fossil preservation and to reduce the overlap among experimental articles, some main and review articles were selected to compose Category 2 (Supplementary Material 2; Tables 1, 2). We consider that the very first step is to compile strategies, relying on analytical approach, to assess these challenges, thus aiming to guide future studies.
Considering Class 2a (silicification), it is noted that most studies rely on modern models of silica precipitation in association with microorganisms. Imaging and geochemical techniques are used to characterize, respectively, external structure (S), crystal morphology and texture (T) – both with approximately 75% of the total 2a articles; inorganic composition (C) – half of the 2a articles and less relevant biofilms (I) and organic (O), and isotopic (i) compositions (Table 2, Figure 4). Silica precipitation in microorganisms has a biotic component, with formation of polysaccharide as polymers evidenced by FT-IR, facilitating silica formation, and an inorganic component that is spherical nanosilica precipitation showed by SEM (Benning et al., 2004). This agrees with this habit occurring in biofilms and can be used to identify fossilized biofilms (Benning et al., 2004). Moore et al. (2021) indicated that silicification is facilitated by (1) bacterial EPS (FT-IR and EDS showed that sulphate-rich EPS facilitates silica precipitation; Si quantified by nanoSIMS); by (2) photosynthesis-based pH induces silica formation (SEM/EDS and FT-IR showed silica increase); (3) by Mg2+; and (4) by silica in solution. This study indicates that it is worth searching for life in silica deposits with Mg (both terrestrial and in other planets), and that combination of silica and Mg in ancient microbialite candidates can be evidence for their biogenicity. Melim et al. (2023) interpreted silicified biofilm coating trilobite sclerites as biologically mediated silicification using SEM to describe crystal morphology and identify biofilms.
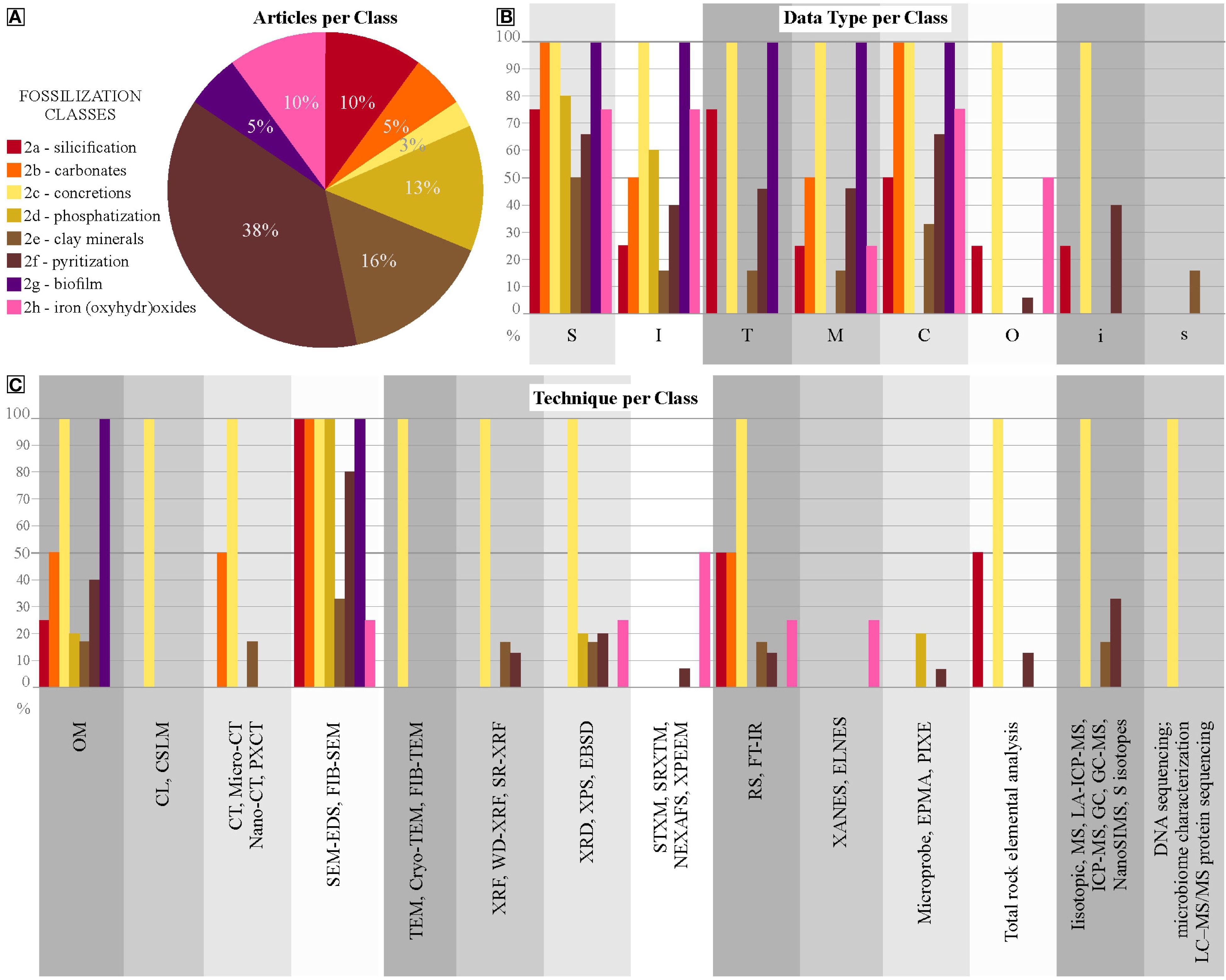
Figure 4. Distribution of Fossilization Processes articles. (A) percentage of each class of the total articles in Category 2. (B) Percentages of fossilization processes classes regarding each type of data. (C) Percentages of fossilization processes classes that apply each type of technique.
Class 2b (preservation by carbonate) comprises mainly calcitization, which is fossilization by calcite and less frequently the formation of dolomite, aragonite or other carbonate minerals. All the reviewed articles from 2b used data on external morphology (S) and inorganic composition (C), and half assessed internal morphology (I) and compositional distribution (M), these being the main data to discuss the biogenicity of fossilization by carbonates (Table 2; Figure 4). The investigation of modern environments and taphonomic experiments indicate that the bacterial surface has an affinity to Ca2+ and to Mg2+, creating a coating (observed by SEM and optical microscopy) and thus preservation of bacteria by calcite (Van Lith et al., 2003). The authors suggested that this could be a criterion for bacterial mediation in calcitization. Mähler et al. (2020) performed a taphonomic experiment of muscle calcitization with associated bacteria. SEM and micro-CT showed microfabric details, while the former technique imaged soft tissues and bacteria, and Raman yielded composition.
In an extensive review, Dhami et al. (2023) compiled the biological (microbial) mediation in the formation of calcium carbonate, iron carbonate and phosphatic concretions, which comprise our Class 2c (concretions). Several imaging, geochemical, microbiology and omic techniques were reviewed to explore their potential as indicators for microbial processes mediating concretion formation (Table 2; Figure 4).
The evaluation of Class 2d (phosphatization) provides one of the most explored fossilization modes. The works comprise several geological periods and experimental taphonomy also play an important role in understanding and testing the biogenicity of phosphatization. SEM was used to characterize preserved soft tissues (S and I, respectively 80 and 60% of the articles in class 2d; Table 2; Figure 4) and associated microfabrics (Wilby and Briggs, 1997), phosphate-rich microbial mats mediating phosphatization (Wilby et al., 1996). In this latter study, total rock elemental analysis showed phosphate enrichment in microbial levels. SEM showed calcium phosphate microspheres preserving polychaete muscles and pits left after possible bacteria degradation (Wilson et al., 2016). Gäb et al. (2024) employed SEM and TEM to show that apatite formed by phosphatase enzymes resemble that found in fossils. Crosby and Bailey (2012) showed that sulfide-oxidizing bacteria form modern phosphorite deposits, though a protocol for their identification in the geological record has not been established yet.
It is long considered that biological processes play a role in the preservation by clay minerals (Class 2e). Data of external structure (S) and composition (C) are more relevant for discussing compositions (Table 2, Figure 4). Butterfield (1995) suggested that enzymatic activity during decay is hampered by clay minerals, which would have facilitated the aluminosilicification of Cambrian fossils. Taphonomic experiments are consistent in showing that certain clay minerals are more efficient in soft-tissue preservation, specifically kaolinite (Wilson and Butterfield, 2014), as it slows decay in preventing bacterial proliferation (Briggs and McMahon, 2016, and references therein). This observation explains Burgess Shale-type preservation preferentially in originally kaolinite facies (Anderson et al., 2018, 2021). Corthésy et al. (2024) further used microbial community characterization/genetic sequence associated with decaying shrimps in different clay substrates, to show that kaolinite has the lowest bacterial diversity, particularly of bacteria that efficiently decay polysaccharides of the skeletons. Becker-Kerber et al. (2021) applied a suit of techniques to Ediacaran macrofossils, suggesting microbial mediation of clay mineral precipitation after volcanic material alteration.
Class 2f (pyritization) is also a well-explored taphonomic process, much focused on the understanding of microbial mediation in mineralization. To this end, more than half of the articles use data on external morphology (S) and inorganic composition (C), aided to a lesser extent by texture (T) and mapping data (M) (Table 2; Figure 4). Taphonomic experiments have been focused on understanding geochemical conditions controlling distinct pyrite habits and textures formed during plant decay with the mediation of sulphate-reducing bacteria in anoxic conditions (Grimes et al., 2001; Brock et al., 2006). Experiments showed the formation of FeS black films coating shrimp carcasses under microbial consortium of sulphate-reducing, sulphide-oxidizing, and fermentative bacteria (Sagemann et al., 1999). The controlled decay of carcasses simulating the preservation of Ediacaran soft-bodied macrofossils enlightened the role of biofilms in mediating the precipitation of authigenic combination of Fe/S (iron sulphides)? and Al/K/Mg (aluminosilicates)? (Darroch et al., 2012), as shown in the fossil record (Laflamme et al., 2011). In this study, SEM was applied to determine crystal morphology (S and T) and EDS and XPS to characterize chemical composition (C).
Associated to class 2f, the high-resolution characterization of mineral textures in association to biofilms, Class 2g, is still poorly-explored, with the exception of a few studies like that of MacLean et al. (2008), that employed SEM/EDS and FIB-SEM to describe the textural relationship between biofilms and pyrite nanocrystals forming framboids and biofilms (Class 2g), plus STXM, NEXAFS and XPEEM to yield the organic composition (O) of biofilms and sulphur isotopic (i) composition of pyrite, altogether confirming pyrite biogenicity. Several studies have used sulphur isotopes to test the biogenicity of pyritization, and to describe the sequence of pyritization (Briggs et al., 1991; Farrell et al., 2009; Schiffbauer et al., 2014; Georgieva et al., 2022). In some of these studies, SIMS has been proved to be a powerful tool in enabling in situ nano-/micro-measurements. Xiao et al. (2010) showed that rapid pyrite precipitation dropped pH yielding silicification, then forming chert nodules encasing fossil embryos. SEM work in fossil insects from the Jehol Biota (China) and from the Crato Formation (Brazil) has demonstrated preservation by framboidal pyrite in association with biofilms (Wang et al., 2012; Osés et al., 2016; Dias and Carvalho, 2022).
Preservation by Category 2h (Fe (III) (oxyhydr(oxides)) has demonstrably been attributed to biological mediation, for this, three quarters of the articles used morphology (S and I) and composition (C) data. Li et al. (2013) demonstrated that Fe (II)-oxidizing bacteria can precipitate iron (III) (oxyhydr)oxides, either mineralizing EPS or the bacteria themselves. Experiments using TEM and geochemical techniques showed that the formation and excretion of Fe, in association with organic polymers, prevents cell encrustation of iron (II)-oxidizing bacteria (Chan et al., 2011). These biosignatures can be applied to iron (II)-oxidizing bacteria candidates in the geological record, which also has implications for Astrobiology. Miot et al. (2011) found Fe-binding/oxidizing sites at the outermost region of cell membranes. SEM was used to show biofilms in plants fossilized by iron (III) oxyhydr(oxides) (Martins et al., 2022).
Evidence of biogenicity in fossilization processes (Figure 5), in general, is inferred from coating/biofilm features associated with bacterial degradation in fossil tissues (Wilson et al., 2016). Specific morphologies such as microspheres or filaments and specific habits such as framboidal pyrite (see Grimes et al., 2001; Brock et al., 2006), mineral excretions, mineral growth patterns and microfabrics can help in defining induced or influenced mineralization and possible biofilms (Wilby et al., 1996; Wilby and Briggs, 1997; Benning et al., 2004; Van Lith et al., 2003; Chan et al., 2011; Mähler et al., 2020; Melim et al., 2023). The discussion can still be complemented by organic composition or isotopic fractionation to characterize biosynthesis and metabolic processes of fossilizing microorganisms (Briggs et al., 1991; Maclean et al., 2008; Farrell et al., 2009; Schiffbauer et al., 2014; Georgieva et al., 2022). Figure 5 summarizes a protocol of how to proceed to assess biogenicity of several fossilization modes.
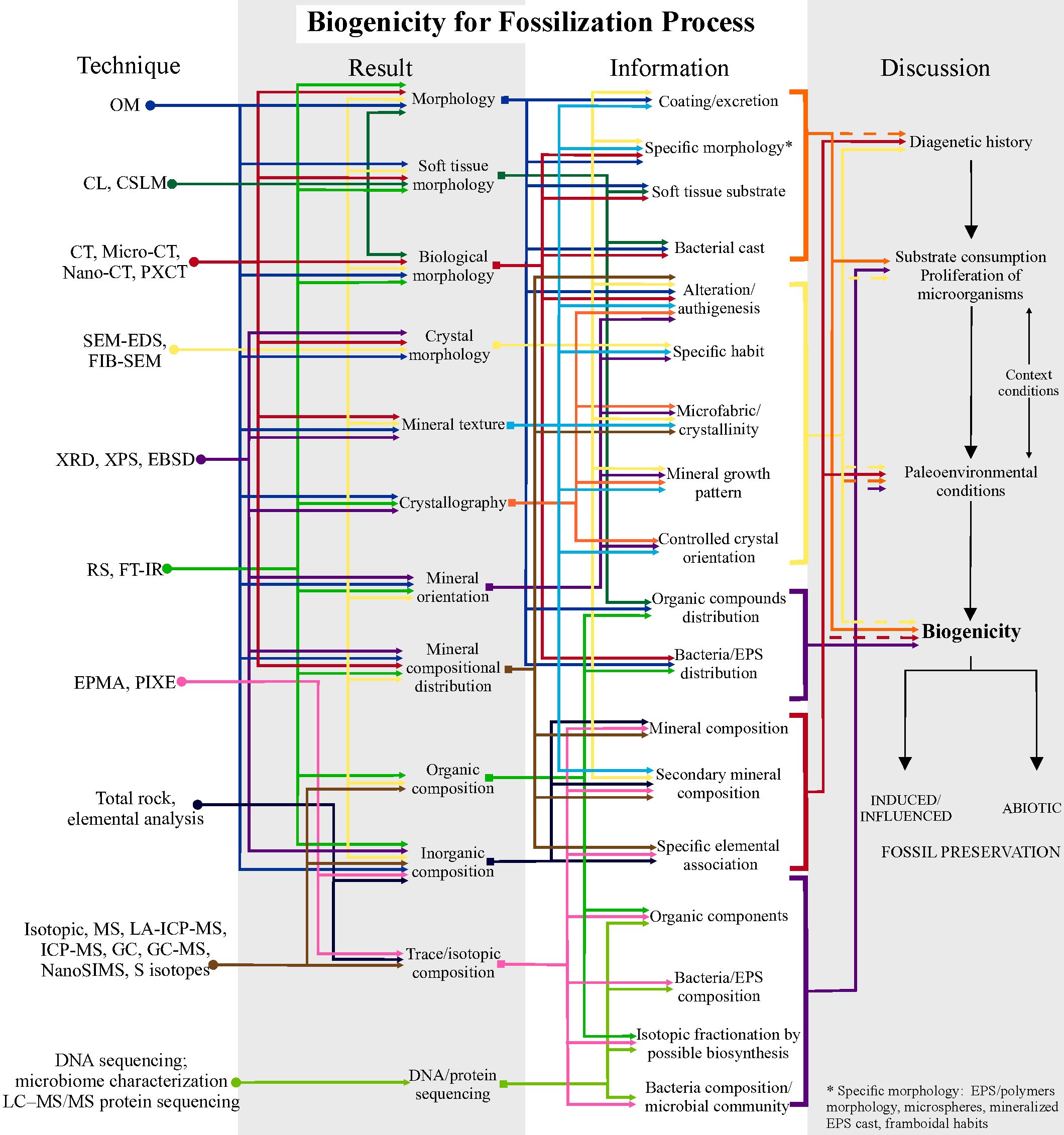
Figure 5. Schematic protocol for investigating biogenicity in Fossilization Processes. Steps using high-resolution techniques to obtain results, information and how to discuss them. The first step involves deciphering the diagenetic history; the second, attesting the substrate consumption and or proliferation of microorganisms; third, recovering the original biogeochemical conditions, involving any additional information about the geological context; and the final step is to argue and raise biogenicity criteria to propose the abiotic or induced/influenced fossil preservation. This protocol is based on the table in Supplementary Material 2 with the high-resolution techniques used in the reviewed articles, together with their potential applications in results and information. Solid lines in the discussion of biogenicity indicate direct application of the information to the topic being discussed; dashed lines indicate indirect or subordinate application to the discussion. See Supplementary Material 6 for a larger, high-resolution version.
To test the participation of life in fossilization processes (Figure 5), the strategy could be as follows: 1) track the diagenetic history, defining which processes have led to fossilization and which occurred later; 2) evaluate whether the substrate was consumed and allowed the proliferation of biomineralizing microorganisms by means of the description of preserved biofilms, mineral textures and geochemistry; and 3) retrieve data on the original environmental conditions using an integrated stratigraphic, sedimentological and geochemistry approach, which can shed light on biogeochemical cycles conducive to fossilization. If there is sufficient evidence, the fossilization process should be defined as biotically induced/influenced.
3.3 Biomineralizing metazoans
Category 3 includes a wealth of published data that builds a toolkit to evaluate biogenicity of minerals found in fossilized skeletal metazoans. For this purpose, data are mainly required on surface morphology (S) and inorganic and organic composition (C and O) – approximately 57% of total Category 3 articles requires this type of data; complemented by texture (T), 28% of Category 3 (Table 2; Figure 6). RS and FT-IR are well-used techniques, as well as SEM-EDS and EBSD (Trueman, 2013; Cusack, 2016; Gilbert et al., 2019; Osés et al., 2022). FT-IR can be used to assess the crystallinity of biominerals to evaluate diagenesis and, consequently, the use of biominerals to biological and paleoenvironmental studies (Trueman, 2013). EBSD has been largely used to modern and extinct taxa to characterize the mineralogy and the crystallographic orientation of crystals, giving information on the biological control of biomineralization (Cusack, 2016). Osés et al. (2022) employed a multitechnique characterization of Ediacaran fossils (with OM, CL, micro-CT, SEM, RS, EBSD and EPMA), tracking diagenesis to show a primary aragonite skeletal composition, precipitated under biological control, by using EBSD and EPMA. Pérez-Huerta et al. (2018) reviewed the application of CL and of EPMA to the evaluation of diagenesis in biomineralization studies. The organic components of biomineralized skeletons have also been analyzed to test their biogenicity. Stern (1996) applied GC/GC-MS spectrometry to identify lipids found in shells of fossil mollusks, and Osés et al. (2022) used micro-Raman spectra to show amorphous carbon associates to skeleton calcite. (plus see a compilation of articles reporting organic matter preservation and isotope research of organics in fossils by Pérez-Huerta et al., 2018).
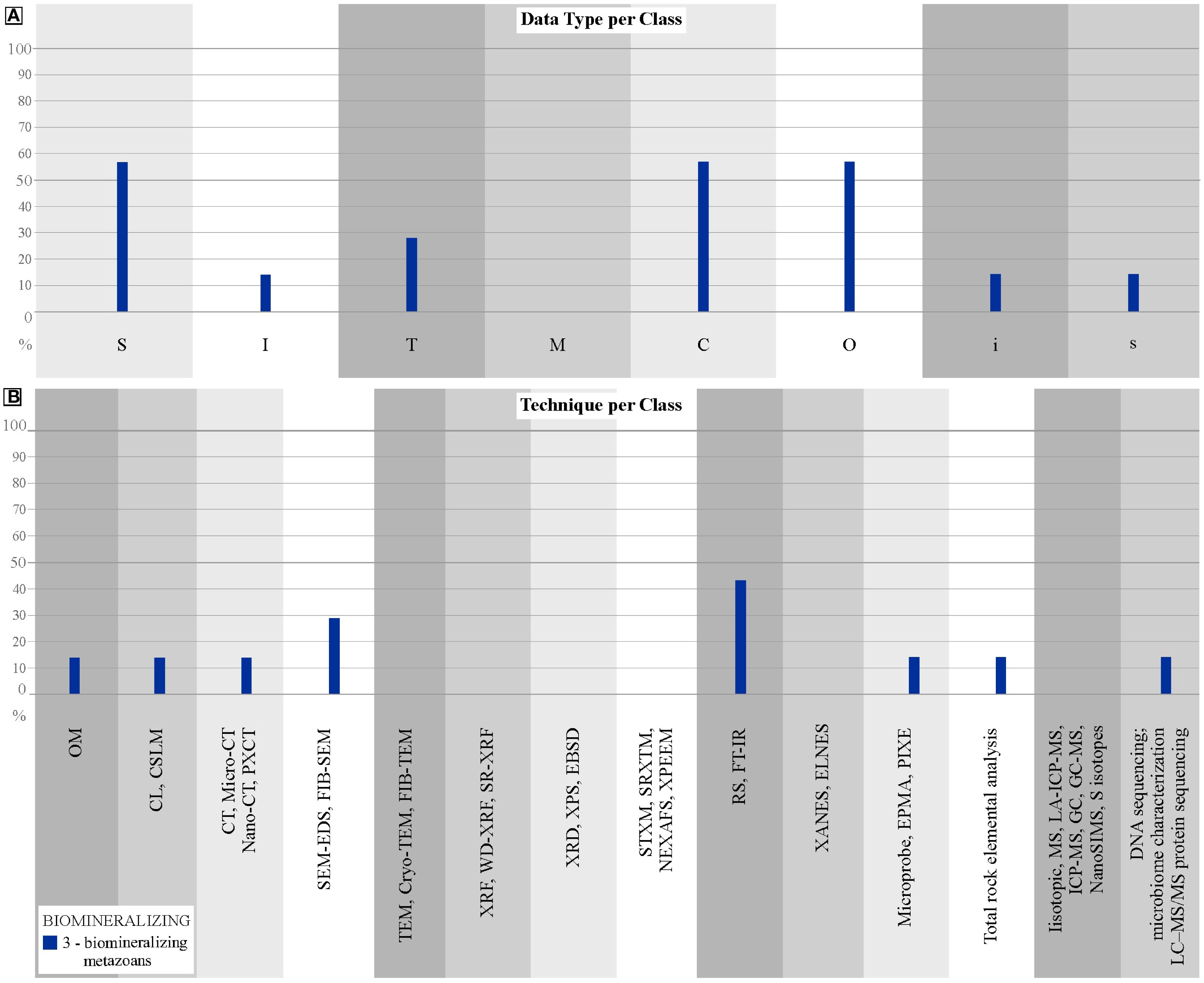
Figure 6. Distribution of biomineralizing metazoans articles. (A) Percentages of biomineralization articles that obtain each type of data. (B) Percentages of biomineralization articles that apply each type of technique.
When aiming to test biomineralizing metazoan candidates, it is generally necessary to gather evidence to assess two main aspects of biomineralization in the fossil record, that are primary composition of mineralized skeletons and biological control of biomineralization (Figure 7). The first goal deals with characterizing mineral habit, size, texture, and elemental and mineralogical composition. This allows for tracking diagenesis, to rule out secondary, alteration composition and give clues of primary mineralogy or, at least, aid in targeting less altered regions to be further investigated (e.g., Zhuravlev et al., 2012; Cusack, 2016; Wood et al., 2017; Pérez-Huerta et al., 2018; Pruss et al., 2018; Osés et al., 2022). The second goal aims to differ biomineralization control from biomineralization through passive action of the organism (e.g., Pérez-Huerta et al., 2018; Osés et al., 2022). This involves interpreting to what extent texture and microstructure are linked, yielding consistent patterns in allometric studies. The second goal also involves establishing the spatial relationship between organic matter and crystals that would suggest biomineral growth at an organic template, the identification of biomolecules involved in biomineralization, the type of skeletal microstructure, nanostructure of crystals and finding evidence for a preferential orientation of crystals (Pérez-Huerta et al., 2018; Osés et al., 2022). This approach could be complemented by the identification of macromolecules, organic compounds, and their distribution within the object of study (Stern, 1996; Cusack, 2016; Osés et al., 2022), in order to understand a genetic relationship between the organic and mineral components of a fossil (Figure 7).
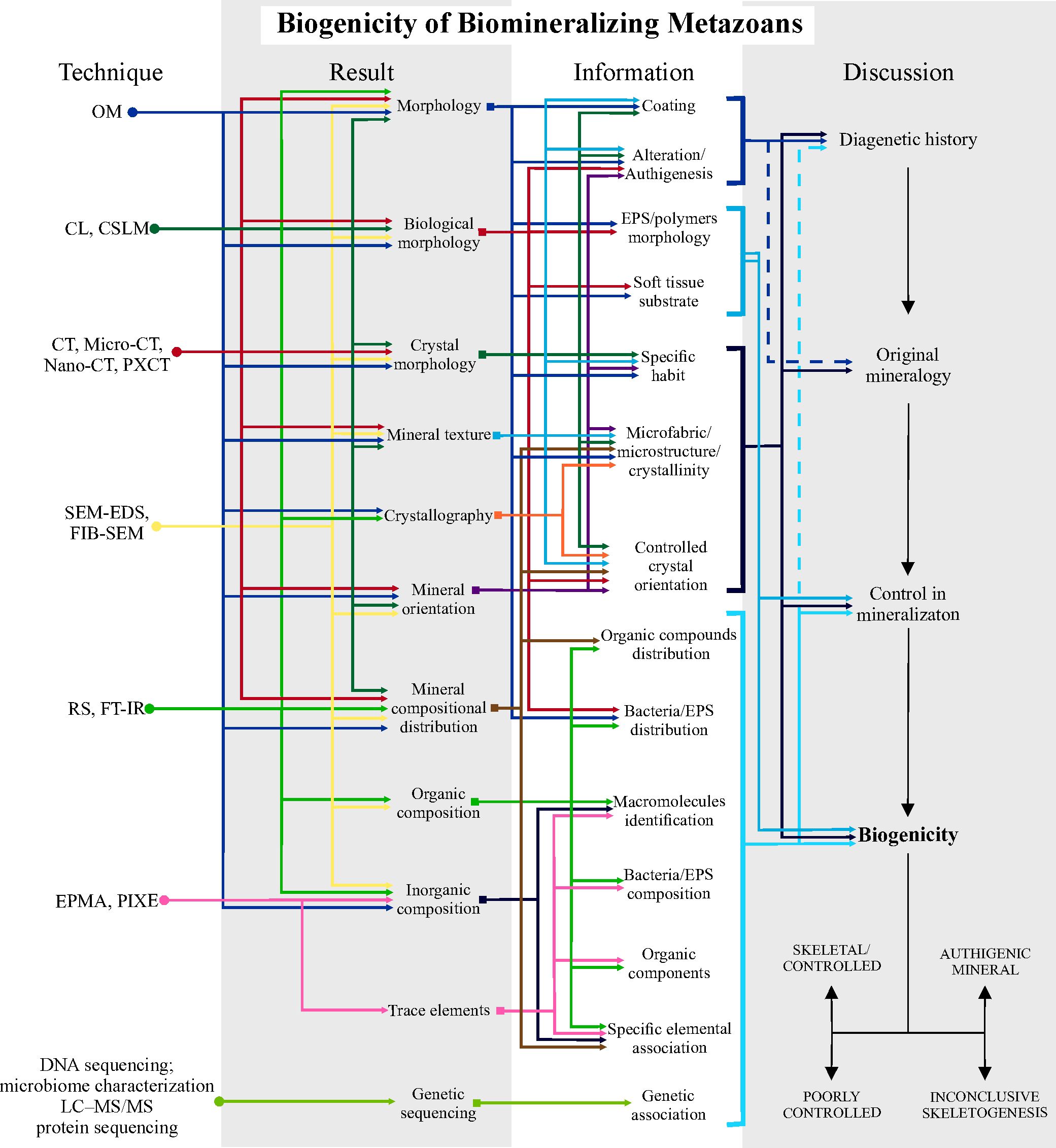
Figure 7. Schematic protocol for investigating biogenicity in Biomineralizing Metazoans. Steps using high-resolution techniques to obtain results, information and how to discuss them. The first step involves discovering diagenetic history; the second, recovering the original mineralogy, eliminating secondary alterations; the third step, investigate the biological control in the original mineralogy and the final step is to argue and raise biogenicity criteria to propose the origin by controlled biomineralization, or abiotic authigenesis, or inconclusive skeletogenesis due to lack of information. This protocol is based on the table in Supplementary Material 2 with the high-resolution techniques used in the reviewed articles, together with their potential applications in results and information. Solid lines in the discussion of biogenicity indicate direct application of the information to the topic being discussed; dashed lines indicate indirect or subordinate application to the discussion. See Supplementary Material 7 for a larger, high-resolution version.
Therefore, this approach can be followed in three steps, as summarized in Figure 7: 1) Understand the diagenetic history: identify the processes that have modified mineral composition, morphologies, and textures, or even formed new minerals. 2) Identifying the original mineralogy. 3) Discussing the biological control of mineralization by assessing textural evidence and other lines of evidence. If there is sufficient supporting information, the fossil can be interpreted as having been formed by biominerals either formed by biologically controlled growth, or as a poorly controlled mineralization (as soft tissue-mediated mineral agglutination) or with an inconclusive biomineralizing skeletogenesis. Otherwise, preserving skeletal minerals can be proved to be totally diagenetic in origin.
4 Discussion
4.1 Biogenicity discussions in paleobiology
Although the study objects and goals differ across the three biogenicity categories, they share a common framework for assessment (Figures 3, 5, 7), with deciphering diagenesis as the central element and the starting point. Interpreting diagenetic history – including indigenicity and syngenicity, especially for dubiofossils – is essential for evaluating biogenicity rocks (Gargaud et al., 2015; Rouillard et al., 2021). Diagenesis, understood as a biogeochemical continuum involving processes such as compaction, recrystallization, dissolution, and, in some cases, low-grade metamorphism (Milliken et al., 1978; Worden and Burley, 2009), acts as both a preservative and a transformative force, capable of incorporating abiotic and biotic signals into the rock record or erasing them entirely. Its complexity arises from the wide range of overlapping physicochemical processes (e.g., compaction, recrystallization, authigenesis) and the prolonged timescales involved, which often lead to overprinted or ambiguous signatures (Milliken et al., 1978; Briggs, 2003; Worden and Burley, 2009; Schiffbauer et al., 2007; Boggs, 2009; Knoll, 2013; Muscente et al., 2017; Saldanha et al., 2023). This complexity makes replicating diagenetic pathways in laboratory settings particularly difficult (Briggs and McMahon, 2016). The following discussion explores how these challenges manifest across the categories analyzed in this review.
4.1.1 Challenges in assessing the biogenicity of dubiofossils
Despite technological advances, evaluating the biogenicity of dubiofossils remains a complex task due to their ambiguous morphologies and the overlapping signatures produced by abiotic and biotic processes. Dubiofossils, enigmatic fossils, and pseudofossils are structures that often exhibit features suggestive of biological origin, leading to uncertainty or debate regarding their true nature (Hofmann, 1972; Monroe and Dietrich, 1990). Complex or organized morphologies are found in both abiotic and biotic settings, and distinct abiotic and biotic processes can produce similar compositional signatures, complicating this distinction (Monroe and Dietrich, 1990; Lowe, 1995; Seilacher, 2001; García-Ruiz et al., 2002; Pasteris and Wopenka, 2003; McCollom and Seewald, 2006; Botta et al., 2008; McLoughlin et al., 2008; Westall, 2008; Bower et al., 2015; Gargaud et al., 2015; Westall et al., 2015; McMahon, 2019). Diagenetic and metamorphic processes can further obscure original features or produce artifacts that mimic life (Schiffbauer et al., 2007; Neveu et al., 2018; Rouillard et al., 2021).
Recent frameworks highlight life’s active role in shaping geological environments, emphasizing that biological and geological processes are deeply intertwined (Knoll, 2013; Davies et al., 2016; Zhang et al., 2017). These interconnected biological and geological processes are integral to Earth’s natural cycles, often overlapping across various temporal and spatial scales (Milliken et al., 1978; Worden and Burley, 2009; Zhang et al., 2017). Moreover, the discovery of life in nearly all terrestrial environments, including extreme ones, has extended the known limits of habitability and deepened the relevance of astrobiological perspectives research (Merino et al., 2019; McMahon and Ivarsson, 2019, and references therein).
These complexities reinforce the importance of assessing indigenicity and syngenicity. Both abiotic and biotic contaminants or asynchronous materials can enter the system at any stage of the sedimentary cycle, altering or obscuring original signatures, meaning that while indigeneity and syngenicity are not criteria for biogenicity, both are crucial steps in understanding the object’s origin (Rouillard et al., 2021 and references therein). Therefore, establishing robust evidence for the object’s formation within a specific temporal context – whether pre-depositional, depositional, or diagenetic – is essential to ruling out contamination, understanding its origin and the effects of subsequent processes on the material (Schiffbauer et al., 2007; Neveu et al., 2018; Rouillard et al., 2021; Saldanha et al., 2023). This temporality should be complemented with all available information about the location, bedrock, and geological context, which aid in reconstructing the diagenetic history and the original setting (Rouillard et al., 2021).
Understanding the environmental context in which potential biosignatures are found is critical to evaluating their biogenicity accurately (Benzerara et al., 2019; Rouillard et al., 2021; McMahon and Cosmidis, 2022). While extreme environments were once used to challenge biogenic interpretations, the ubiquity of life cautions against dismissing biosignatures based on environmental context alone (Merino et al., 2019; McMahon and Ivarsson, 2019, and references therein). The full extent of biological activity throughout the sedimentary cycle remains incompletely understood, making it premature to dismiss biogenicity based solely on the context of information, even under extreme, mesodiagenetic or metamorphic conditions (Schiffbauer et al., 2007; Bengtson et al., 2017; McMahon and Ivarsson, 2019; Ivarsson et al., 2020, 2021). The original environmental scenario must be carefully analyzed to identify plausible formative processes and comparative analysis with similar modern or ancient products can provide a useful benchmark for evaluating these processes. Thus, any geological object, whether of abiotic or biotic origin, must therefore be understood not only in terms of its formation and initial conditions but also in light of the subsequent processes that have contributed to its preservation, alteration, or destruction through geological time, considering chemical interactions, biological influence, and superimposed processes (Rouillard et al., 2021; Saldanha et al., 2023).
There is growing consensus that no universal reliable, distinguishable, and generic biosignature can definitively diagnose life in deep time or beyond Earth (as highlighted by Rouillard et al., 2021). The various features discussed here demonstrate that no single characteristic can be universally applied to determine biogenicity, since ambiguous results are often encountered in the detection of life (McMahon et al., 2021). Therefore, a set of evidence should be required to assess the origin of dubiofossils. However, even multiple lines of evidence may be inconclusive if individually ambiguous, underscoring the need for at least one uncontested criterion for biogenicity (McMahon et al., 2021; McMahon and Cosmidis, 2022). Therefore, given the series of ambiguous proposed arguments, testing a set of criteria to determine whether dubiofossils fit an abiotic or biotic history remains valuable.
Therefore, testing hypotheses about the origin of dubiofossils requires the impartial consideration of abiotic and biotic processes (Buick, 1990). A major challenge lies in the asymmetry of knowledge in which abiotic processes are often less understood and harder to falsify (McMahon et al., 2021) and may inadvertently bias interpretations toward a biotic explanation (e.g., Brasier et al., 2004; Davies et al., 2016; Dodd et al., 2017). This approach is based on the premise that if an argument does not support biogenicity, it is assumed to be abiotic (Wacey, 2009). As Buick (1990) pointed out, failing to meet all biogenic criteria does not confirm an abiotic origin; it simply reflects a lack of definitive evidence. The inverse is also true: inability to eliminate all abiotic alternatives prevents proof of a biological origin. (e.g., Dodd et al., 2017; Baucon et al., 2020). Therefore, it is essential to evaluate hypotheses with equal consideration (Hofmann, 1972; Buick, 1990; Benzerara and Menguy, 2009; Wacey, 2009; McMahon et al., 2021; Rouillard et al., 2021; McMahon and Cosmidis, 2022) and to rigorously assess the possibilities considering current understanding of both abiotic and biotic phenomena (McMahon and Cosmidis, 2022).
Our quantitative analysis (Table 2) further shows that Class 1a microfossil-like structures receive the most attention, accounting for 56% of all Category 1 studies, due to their relevance to early life detection. In contrast, Classes 1b and 1c, with greater morphological variability, are studied less often, possibly due to the difficulty in applying uniform biogenicity criteria. In summary, assessing dubiofossils requires a multidisciplinary and context-aware approach. The absence of a universal biosignature, the need to test competing hypotheses impartially, and the overlapping effects of geological processes all demand a robust, integrated strategy grounded in both biotic and abiotic knowledge.
4.1.2 Challenges in assessing the biogenicity of fossilization
Although it is well-established that microorganisms mediate mineralization processes in fossilization (Briggs, 2003), recognizing this relationship in the fossil record remains difficult. The challenges include mainly dissecting the diagenetic processes that a carcass has undergone. It is common to find fossils with significant diagenetic imprints that obscure the very early stages of diagenetic mineralization, which is the main source of information of the mediation of microorganisms in decay and mineralization of organic matter. For example, oxidation of primary sulfides might make it difficult to identify the original crystal morphology (e.g., Osés et al., 2016), or the complex diagenesis of clay minerals might hamper the identification of primary preserving mineralogy (e.g., Voltani et al., 2023). It is also challenging to identify with confidence candidate biofilms and bacteria, which can be a direct source of evidence for the role of microorganisms in fossilization (e.g., Dias and Carvalho, 2022).
Experimental taphonomy offers promising solutions by directly testing microbial roles in different mineralization pathways, as demonstrated for phosphatization (Wilby et al., 1996; Wilby and Briggs, 1997). This would enlighten geomicrobiological processes and provide a textural and a compositional database that can be used to assess the fossil record. Other promising research topic is to test whether the carcass original composition drives textural and compositional diagnostic characteristics, which could be then identified in the fossil record. On the other way round, taphonomic experiments can gauge our interpretations and expectations of the fossil record (i.e., taphonomic models).
When we consider our quantitative data (Table 1), it is clear that techniques are mostly applied to solve morphological and geochemical issues concerning taphonomy (OM, SEM-EDS/SEM-FIB, RS/FT-IR). This pattern does reflect the main approaches to study the biogenicity of fossilization processes discussed above. These techniques also allow for in situ, microscopical characterization of minerals. In contrast, techniques such as synchrotron radiation (SR), TEM, cathodoluminescence (CL), CT imaging, and biomolecular methods (e.g., DNA sequencing, microbiome analysis) are underutilized—likely due to limited accessibility, complex sample preparation, or lower perceived utility. Therefore, the development of new, portable instrumentation of in situ analysis could aid in expanding the use of XRF and XRD for example. When we consider different fossilization modes, pyritization, clay mineral preservation and phosphatization are the most studied in respect of probing their biogenicity (Table 2 and Figure 4). Interestingly, these also usually employ multiple techniques, including less used ones (Table 1).
4.1.3 Challenges in assessing the biogenicity of skeletal minerals
Challenges to assess the biogenicity of biomineralizing candidates include mainly understanding the diagenesis of these fossils. Particularly carbonates are prone to diagenetic alteration, which represents a great challenge to biomineral study (Pérez-Huerta et al., 2018). Therefore, techniques that can retrieve high-resolution spatial information on texture, chemical/mineralogical/organic/isotopic composition and crystallography have been widely applied to biomineralization research.
Indeed, it is possible to conclude from our data that OM, SEM-EDS, RS/FT-IR, XRD/EBSD and elemental analysis are the most commonly used techniques in the field, as for fossilization studies. This observation has a twofold interpretation. First, it supports the high magnification, spatial resolution of combined images and chemical maps, low detection limit (even of light elements) and mineralogical identification/crystallographic information provided by the combination of these techniques. Second, this pattern might reflect the easy use of them, as they are widely available in research centers and have been long used in research, at least OM, SEM-EDS and XRD.
However, there is much room for the application of extremely useful but still underexplored techniques, like CL and EPMA for diagenesis tracking, CT-based techniques for morphological characterization and its relationship with mineralogy (hierarchical organization in biomineralizers sensu Pérez-Huerta et al., 2018). As for the fossilization studies, SR-based techniques and those relying on molecular taphonomy have been largely underexplored, thus being promising avenues of research.
4.2 Paleometry as a key to assessing biogenicity
This compilation highlights the pivotal role of paleometry in studies of biogenicity. Notably, only a small fraction of the reviewed articles relies solely on one category of data (Table 2). Among these, only the 2d class is characterized by the exclusive use of morphological data (see Table 2; Supplementary Material 2). Moreover, fewer than one-quarter of the studies employed one or no high-resolution analytical technique (Table 1), emphasizing the broader value of paleometry, of in employing more than one high-resolution technique and the need to integrate data to extract more precise information from the fossil record (Riquelme et al., 2009; Delgado et al., 2014).
As emphasized in Section 4.1, reconstructing the diagenetic history is a fundamental step in assessing the biogenicity of any paleobiological material. Traditionally, interpretations of sedimentary rock diagenesis are grounded in petrographic analysis using OM, often complemented by SEM (e.g., Ketzer et al., 2003; Worden and Burley, 2009; Worden and Morad, 2003; Cardoso et al., 2024). Reconstructing the original conditions (original scenario, original biogeochemical conditions, or original mineralogy) requires integrating stratigraphic, geochemical, and sedimentological context (e.g., variations in grain size, texture, and sedimentary structures) with results from high-resolution analytical techniques. Methods commonly used for diagenetic and geochemical characterization include OM, CL, SEM-EDS, XRD, and XRF. In the specific context of biogenicity, additional tools such as RS, FT-IR, and compositional analysis via EDS provide critical morphological, textural, and chemical data, both for the fossil and its surrounding host rock. These integrated datasets enhance our understanding of depositional environments, post-depositional weathering, and fossil-diagenetic conditions (e.g., Osés et al., 2017), thus supporting more robust and nuanced interpretations of biogenicity (Figures 3, 5, 7).
The assessment of biogenicity in paleobiological materials relies heavily on the integration of S and I and C, O, i, s data, associated secondarily with T and M providing important but secondary support (Table 2; Figure 1B). This emphasis helps explain the widespread application of high-resolution techniques such as OM, SEM-EDS, µ-RS, FT-IR, and, to a lesser extent, µ-CT. OM offers insights into I, T, M, and O; SEM-EDS covers S, I, M, and C; µ-RS and FT-IR provide C and O data linked to T and M; µ-CT enables 3D imaging of S, I, T and can infer C and M via grayscale. (see Supplementary Material 1). Most of these techniques operate at the microscale and are favored not only for their versatility in generating multiple data types, but also due to their widespread availability in academic and research settings. They typically involve minimal sample preparation, are non-destructive or minimally destructive, and offer relatively low cost and rapid processing and interpretation, features that have made them standard in paleontological research (Carlson et al., 2003; Delgado et al., 2014; Borgwardt and Wells, 2017; Pan et al., 2019).
The integration of these high-resolution methods is thus central to current biogenicity protocols. However, when microscale tools reach their resolution limits, nanoscale methods become essential (Callefo et al., 2019b; Maldanis et al., 2020). This is where synchrotron radiation (SR)-based techniques become indispensable. Techniques such as PXCT, SR-XRF, STXM, SRXTM, NEXAFS, XPEEM, SR-FT-IR, and XANES offer non-destructive, high-resolution, and low-detection-limit analyses that do not require complex sample preparation or vacuum conditions. For example, these methods can detect trace and rare earth elements in minute concentrations, aiding both in biogenicity assessments (e.g., Allwood et al., 2018) and in reconstructing paleoenvironmental and diagenetic histories (Gueriau et al., 2014; Prado et al., 2021). Beyond synchrotron-based tools, other advanced methods, such as electron microprobe, EPMA, PIXE, bulk, isotopic studies, mass spectrometry (MS, LA-ICP-MS, ICP-MS), gas chromatography (GC, GC-MS), NanoSIMS, sulfur isotope analysis, FIB, TEM, DNA sequencing, and protein sequencing (LC–MS/MS), are less frequently used but provide essential insights into biomolecular preservation and fossil diagenesis (e.g., McLean et al., 2008; Table 1). Though more specialized, these techniques significantly enhance our ability to detect and validate biogenic signals and warrant greater application in future paleobiological studies.
4.3 Biogenicity assessment through geologic time
As observed in Figure 8, most studies analyzing fossilization processes over time primarily on the Ediacaran onwards, with some works restricted to the Mesoproterozoic These fossils, often exceptionally preserved, enable the application of various techniques at both micro- and macro-scales to understand fossilization.
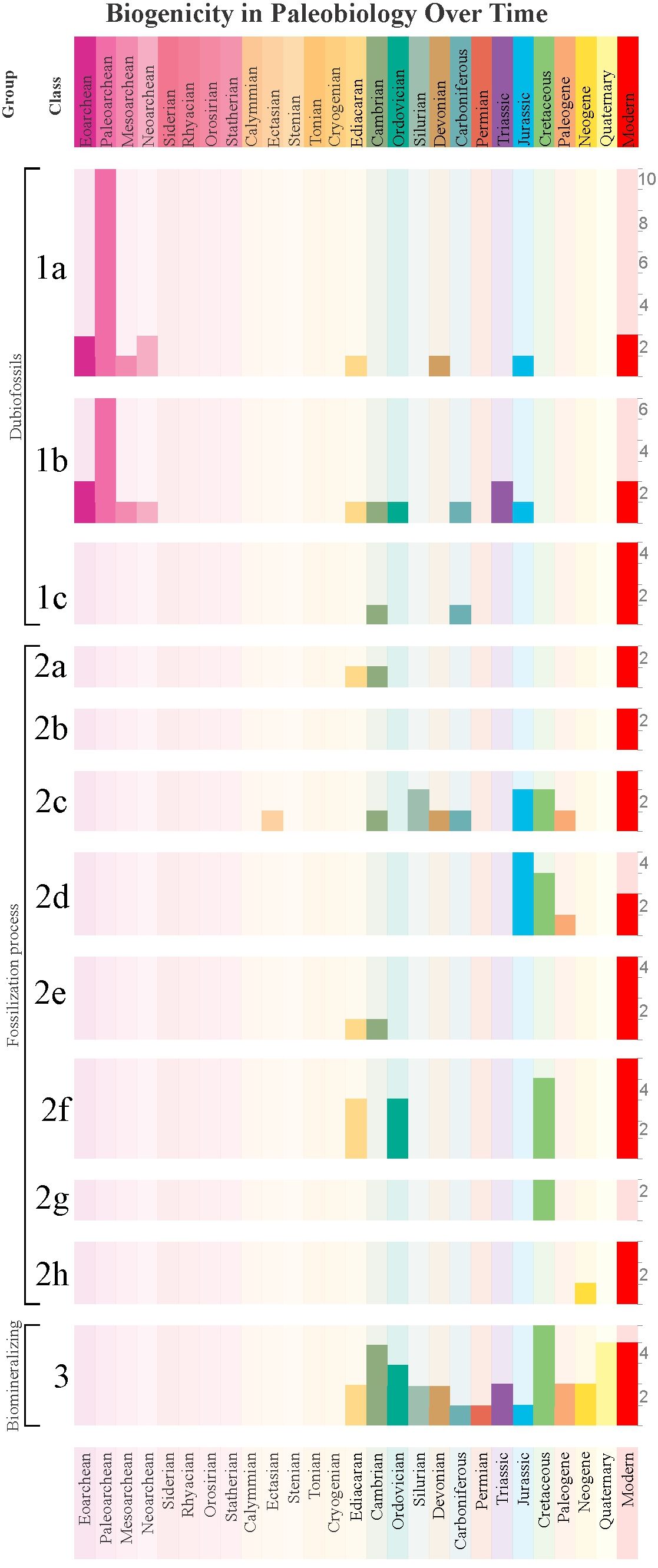
Figure 8. Distribution of the number of case study articles over geological time based on Supplementary Material 2, classified by Category and Class. The right sidebar indicates the absolute number of articles.
The concentration of Category 1a and 1b dubiofossil studies in the Paleoarchean reflects efforts to detect early life (graphs 1a and 1b in Figure 8). However, two major obstacles hinder biogenicity recognition: (1) extensive geological overprinting (e.g., metamorphism, tectonic recycling), and (2) divergence between Archean and Phanerozoic life forms, which challenge comparisons and require more robust criteria (Buick, 1990; Wacey, 2009; McLoughlin and Grosch, 2015b; McMahon et al., 2021; Rouillard et al., 2021). These challenges fueled significant debate over early biosignatures (e.g., Brasier et al., 2002, 2004, 2015; Schopf et al., 2002, 2005, 2007, 2017; Schopf and Kudryavtsev, 2012).
Studies on silicification (graph 2a in Figure 8) concentrate in the Ediacaran–Cambrian, driven by interest in “Ediacara-style preservation.” Recent work suggests early diagenetic siliceous cements enabled preservation of soft-bodied fossils (Tarhan et al., 2025; Liu et al., 2019). Furthermore, the persistence of Ediacara-style fossilization linked to authigenic silica cementation in Cambrian strata was considered to support the view that the disappearance of the Ediacara Biota at the end of the Ediacaran was due to evolutionary rather than taphonomic phenomena (Tarhan et al., 2025). Biofilms can promote the three-dimensional preservation of macroscopic organisms in siliciclastic deposits by facilitating silica precipitation and contributing to mouldic preservation of the organisms (Slagter et al., 2022; Melim et al., 2023). Precambrian oceans were enriched in silica as biomineralizers did not promote silica uptake, becoming an important silica reservoir with the diversification of radiolarians and sponges in the Ordovician, and of diatoms in the Cretaceous (Maliva et al., 2005; Butts, 2014).
Studies on the investigation of biotic role in carbonate precipitation focus on experimental approach. However, the mediation of microorganisms in carbonate concretion formation has been subject of investigation (Dhami et al., 2023). This fossilization mode has a high potential for exceptional preservation, and therefore for the preservation of soft tissues (McCoy, 2014; Cotroneo et al., 2016). When this process is sufficiently rapid, it decreases permeability and consequently protects organic matter from significant degradation, allowing the preservation of soft tissues in a ‘closed chemical system’ (Allison and Pye, 1994; Farmer and Des Marais, 1999; McCoy et al., 2015a). This has led to the fossilization and burial of soft parts of various organisms in concretions across a vast spatiotemporal range (Marshall and Pirrie, 2013; McCoy, 2014; Trinajstic et al., 2022); As observed in graph 2c in Figure 8, concretions have a wide distribution in geological time. A factor contributing to the extensive study of fossil concretions and biogenic aspects of the process is the existence of Lagerstätten bearing concretions restricted to the Phanerozoic. These are known for representing a variety of well-preserved fossils in different paleoenvironments, ranging from hypersaline to marine, estuarine to lacustrine, and freshwater to terrestrial (Marshall and Pirrie, 2013; McCoy, 2014), although most appear to belong to transitional environments, i.e., those at the interface between marine and terrestrial realms (McCoy et al., 2015a). This may explain in part the lack of this fossilization mode in the Ediacaran, when at least metazoans were restricted to oceans, rarely encompassing shallow settings.
Phosphatization studies (graph 2d in Figure 8) are concentrated in the Jurassic–Paleogene (graph 2c in Figure 8). This process represents a critical taphonomic pathway responsible for exceptionally well-preserved fossils, including soft tissues and cellular structures (Briggs, 2003). According to Dornbos (2011), phosphatization is an extremely rare taphonomic window that generally depends on the formation of favorable microenvironments except during the two major pulses of global phosphatization during the Phanerozoic: Cambrian-Ordovician and Cretaceous-Eocene. The latter coincides with widespread euxinic seas and Tethyan transgressions (Ahmed and Kurzweil, 2002), favoring phosphate preservation and resulting in several Konservat-Lagerstätten (Dornbos, 2011) and explaining the highest occurrence of research related to phosphatization processes is during the Mesozoic.
The study of the biogenicity of clay mineral preservation (graph 2e in Figure 8) concentrates in the Ediacaran (Becker-Kerber et al., 2021) and mostly in the Cambrian Burgess Shale-type preservation (Anderson et al., 2018, 2021). In the former case, the authors argued for the role of volcanic material to clay precipitation, while in the latter, diagenetic, paleogeographic and climatic controls favoured this preservational mode (Muscente et al., 2017; Anderson et al., 2018).
As previously stated, exceptionally well-preserved fossils tend to be studied more due to the amount of information that can be retrieved from them. This is the same for the process of pyritization, a rapid process that account for soft-tissue preservation in several Konservat-Lagerstätten (Cai et al., 2012; Briggs et al., 1991; Farrell et al., 2013; Farrell, 2014; Liu, 2016; Liu et al., 2019; Wang et al., 2012; Osés et al., 2016, 2017). Pyritization (graph 2f in Figure 8) is well represented in the Ediacaran, Ordovician, and Cretaceous, often preserving soft tissues via early authigenic pyrite (e.g., Liu, 2016; Briggs et al., 1991). Examples include the “death-mask” model (Callow and Brasier, 2009), Beecher’s Trilobite Bed (Farrell et al., 2013), and the pyritized fossils of the Araripe and Jehol biotas (Osés, 2016; Wang et al., 2012; Osés et al., 2016, 2017; Dias and Carvalho, 2022). Ocean redox evolution may explain temporal trends, with modern pyritization increasingly limited to anoxic freshwater settings (Bowyer et al., 2017; Thompson and Kah, 2012). The progressive ventilation of oceans through the Phanerozoic, pyritization became more restricted to lacustrine settings with anoxic conditions (Osés et al., 2017; Wang et al., 2012).
The investigation of the role of biofilms in fossilization (graph 2g in Figure 8) are focused on Cretaceous deposits bearing pyritized fossils (Wang et al., 2012; Osés et al., 2016). Meanwhile, studies on oxy(hydr)oxide preservation (graph 2h in Figure 8) remain limited, likely due to difficulties in distinguishing primary from secondary Fe-minerals (Martins et al., 2022).
Finally, biomineralization in metazoans (Figure 8) has been explored mainly in the Ediacaran (Wood et al., 2018), Cambrian (Marshall, 2006), and during the Great Ordovician Biodiversification Event (GOBE; Servais et al., 2023). These biological radiations necessitate refined biogenicity criteria for skeletal remains. Expanding this refinement to other geological periods, specifically the determination of primary mineralogy and the testing of biological controls on biomineralization, will deepen knowledge and determine its controlling factors on a geological scale.
4.4 Future perspectives for biogenicity assessment
The protocols proposed here offer a flexible and evolving framework to guide future discussions on the origin and biogenicity of materials, with the aim of deepening and structuring the debate across multiple scientific contexts. They support the identification of high-resolution analytical tools with broad applicability and help reveal methodological gaps, encouraging the integration of more specialized techniques where current approaches are insufficient.
Crucially, these protocols are not limited to paleontology. They can be applied to: (a) ancient fossil records; (b) modern and ancient abiotic structures; (c) experimental taphonomy; (d) studies on life in extreme environments; and (e) astrobiology and the search for extraterrestrial life. Their adoption promotes a systematic and reproducible approach to biogenicity, reducing reliance on isolated case studies and addressing the knowledge gap in abiotic products, which often limits balanced evaluation of biotic vs. abiotic origins (McMahon et al., 2021).
More specifically, the application of biogenicity assessment frameworks in modern extreme environments, such as ophiolites, hydrothermal vents, and acid lakes (McMahon and Ivarsson, 2019; Merino et al., 2019), can provide critical insights into the origin of mineralization processes and related products. Additionally, astrobiology would benefit from structured protocols to evaluate potential biosignatures on other planetary bodies. Dubiofossil-like features, including mineral structures, ichnofossil-like traces, or microfossil analogs, may eventually be detected elsewhere (e.g., Baucon et al., 2020), demanding a systematic and reproducible biogenicity assessment framework comparable to those used for dubiofossils on Earth. Ultimately, integrating these protocols into future research will not only refine biogenicity criteria but also contribute to a more robust and interdisciplinary dialogue, connecting paleobiology, geochemistry, experimental taphonomy, and planetary science.
5 Conclusions
This review of case studies on biogenicity in paleobiology demonstrates that, despite the diversity of study objects—dubiofossils, fossilization processes, and biomineralizing metazoans—there are shared methodological patterns. Most assessments begin with the acquisition of morphological and compositional data, often complemented by texture analysis, followed by the integration of results and a stepwise discussion. A consistent and critical element across all categories is the interpretation of diagenesis, which underpins the reconstruction of original conditions and supports biogenicity evaluations. Each category applies this framework in a specific way:
● Dubiofossil studies assess syngenicity, indigenicity, and environmental context to classify materials as fossils, pseudofossils, or unresolved cases.
● Fossilization studies examine microbial mediation through diagenetic signatures and geochemical conditions to distinguish between biotic and abiotic mineralization.
● Biomineralizing metazoan research focuses on identifying diagenetic overprints and evaluating biological control over mineral formation.
Across these efforts, paleometry, the quantitative integration of morphological, textural, and compositional data, proves essential. Techniques such as OM, SEM-EDS, µ-RS, and FT-IR are widely used due to their accessibility and resolution. Moreover, synchrotron-based methods offer promising advances with their high spatial resolution and non-destructive precision, and their broader application is strongly encouraged. Ultimately, this study highlights the value of combining multiple high-resolution techniques in biogenicity research and underscores its inherently interdisciplinary nature, bridging paleontology, geochemistry, and planetary sciences to advance our understanding of life’s traces in deep time.
Author contributions
JS: Investigation, Conceptualization, Validation, Data curation, Writing – review & editing, Writing – original draft, Formal analysis, Visualization, Project administration. AM: Writing – original draft, Formal analysis, Visualization, Validation, Investigation, Writing – review & editing. GO: Visualization, Writing – review & editing, Writing – original draft, Investigation, Validation, Formal analysis, Data curation. FC: Writing – review & editing, Visualization.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This study was financed in part by the Coordenacão de Aperfeicoamento de Pessoal de Nıvel Superior (CAPES– Finance Code 001) and part by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq – grant 161149/2023-5) to JS and Programa de Pesquisador Colaborador do Instituto de Física da USP, São Paulo research Foundation (FAPESP) for the grants #2023/04501-6, #2022/06485-5, and #2023/14250-0 to GO.
Acknowledgments
The authors would like to thank Editor Prof. Mirían L.A.F. Pacheco for the invitation to contribute to this special volume and for all the previous discussions that resulted in this contribution. We would also like to thank Professors Joice Cagliari, Rodrigo S. Horodyski and Renata G. Netto for help and guidance. We appreciate and are thankful for the support of the Universidade do Vale do Rio do Sinos, Universidade Federal do Rio Grande do Sul and Universidade de São Paulo. We also thank the reviewers who kindly read and suggested modifications that improved our research.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2025.1646756/full#supplementary-material
References
Ahmed E. A. and Kurzweil J. (2002). “Sedimentological, mineralogical and geochemical characteristics of Upper Cretaceous Egyptian phosphorites with special reference to the microbial role in phosphogenesis,” In: Wagreich M. Aspects of Cretaceous Stratigraphy and Palaeobiogeography. Wien: Österr. Akad. Wiss., Schriftenr. Erdwiss. Komm, 11–34.
Alleon J., Bernard S., Le Guillou C., Marin-Carbonne J., Pont S., Beyssac O., et al. (2016). Molecular preservation of 1.88 Ga Gunflint organic microfossils as a function of temperature and mineralogy. Nat. Commun. 7, 11977. doi: 10.1038/ncomms11977
Alleon J., Bernard S., Olivier N., Thomazo C., and Marin-Carbonne J. (2021). Inherited geochemical diversity of 3.4 Ga organic films from the Buck Reef Chert, South Africa. Commun. Earth Environ. 2 (6), 1–7. doi: 10.1038/s43247-020-00066-7
Allison P. A. and Pye K. (1994). Early diagenetic mineralization and fossil preservation in modern carbonate concretions. PALAIOS 9, 561–575. doi: 10.2307/3515128
Allwood A. C., Rosing M. T., Flannery D. T., Hurowitz J. A., and Heirwegh C. M. (2018). Reassessing evidence of life in 3,700-million-year-old rocks of Greenland. Nature 563, 241–244. doi: 10.1038/s41586-018-0610-4
Anderson R. P., Tosca N. J., Gaines R. R., Koch N. M., and Briggs D. E. G. (2018). A mineralogical signature for Burgess Shale–type fossilization. Geology 46, 347–350. doi: 10.1130/G39941.1
Anderson R. P., Tosca N. J., Saupe E. E., Wade J., and Briggs D. E. G. (2021). Early formation and taphonomic significance of kaolinite associated with Burgess Shale fossils. Geology 49, 355–359. doi: 10.1130/G48067.1
Awramik S. M. and Grey K. (2005). Stromatolites: biogenicity, biosignatures, and bioconfusion. Astrobiology Planetary Missions 59060P. doi: 10.1117/12.625556
Baucon A., De Carvalho C. N., Felletti F., and Cabella R. (2020). Ichnofossils, cracks or crystals? A test for biogenicity of stick-like structures from Vera Rubin Ridge, Mars. Geosciences 39. doi: 10.3390/geosciences10020039
Becker-Kerber B., El Albani A., Konhauser K., Abd Elmola A., Fontaine C., Paim P. S. G., et al. (2021). The role of volcanic-derived clays in the preservation of Ediacaran biota from the Itajaí Basin (ca. 563 Ma, Brazil). Sci. Rep. 11, 1–10. doi: 10.1038/s41598-021-84433-0
Bengtson S., Rasmussen B., Ivarsson M., Muhling J., Broman C., Marone F., et al. (2017). Fungus-like mycelial fossils in 2.4-billion-year-old vesicular basalt. Nat. Ecol. Evol. 1, 1–6, 0141. doi: 10.1038/s41559-017-0141
Benning L. G., Phoenix V. R., Yee N., and Konhauser K. O. (2004). The dynamics of cyanobacterial silicification: An infrared micro-spectroscopic investigation. Geochimica Cosmochimica Acta 68, 743–757. doi: 10.1016/S0016-7037(03)00488-5
Benzerara K., Bernard S., and Miot J. (2019). “Mineralogical identification of traces of life in cavalazzi, B., westall, F editors. Biosignatures for astrobiology,” in Advances in astrobiology and biogeophysics (Springer, Cham). doi: 10.1007/978-3-319-96175-0_6
Benzerara K. and Menguy N. (2009). Looking for traces of life in minerals. Comptes Rendus Palevol 8, 617–628. doi: 10.1016/j.crpv.2009.03.006
Bernard S. and Papineau D. (2014). Graphitic carbons and biosignatures. Elements 10, 435–440. doi: 10.2113/GSELEMENTS.10.6.435
Bindschedler S., Cailleau G., Braissant O., Millière L., Job D., and Verrecchia E. P. (2014). Unravelling the enigmatic origin of calcitic nanofibres in soils and caves: Purely physicochemical or biogenic processes? Biogeosciences 11, 2809–2825. doi: 10.5194/bg-11-2809-2014
Boggs J. S. (2009). Petrology of sedimentary rocks (Cambridge: Cambridge University Press). doi: 10.1017/CBO9780511626487
Borgwardt T. C. and Wells D. P. (2017). What does non-destructive analysis mean? Cogent Chem. 3, 1405767. doi: 10.1080/23312009.2017.1405767
Botta O., Bada J. L., Gomez-Elvira J., Javaux E., Selsis F., and Summons R. (2008). Strategies of life detection (Boston: Springer US). doi: 10.1007/978-0-387-77516-6
Bower D. M., Hummer D. R., Steele A., and Kyono A. (2015). The co-evolution of fe-oxides, ti-oxides, and other microbially induced mineral precipitates in sandy sediments: understanding the role of cyanobacteria in weathering and early diagenesis. J. Sedimentary Res. 85, 1213–1227. doi: 10.2110/jsr.2015.76
Bowyer F., Wood R. A., and Poulton S. W. (2017). Controls on the evolution of Ediacaran metazoan ecosystems: a redox perspective. Geobiology 15, 516–551. doi: 10.1111/gbi.12232
Braissant O., Cailleau G., Dupraz C., and Verrecchia E. P. (2003). Bacterially induced mineralization of calcium carbonate in terrestrial environments: the role of exopolysaccharides and amino acids. J. Sedimentary Res. 73, 485–490. doi: 10.1306/111302730485
Brasier M. D., Green O. R., Jephcoat A. P., Kleppe A. K., Van Kranendonk M. J., Lindsay J. F., et al. (2002). Questioning the evidence for Earth's oldest fossils. Nature 416 (6876), 76–81. doi: 10.1038/416076a
Brasier M. D., Antcliffe J., Saunders M., and Wacey D. (2015). Changing the picture of Earth’s earliest fossils (3.5-1.9 Ga) with new approaches and new discoveries. Proc. Natl. Acad. Sci. U. S. A. 112, 4859–4864. doi: 10.1073/pnas.1405338111
Brasier M., Green O., Lindsay J., and Steele A. (2004). Earth’s oldest (~ 3.5 ga) fossils and the `Early eden hypothesis’: questioning the evidence. Origins Life Evol. Biosphere 34, 257–269. doi: 10.1023/B:ORIG.0000009845.62244.d3
Brasier M. D. and Wacey D. (2012). Fossils and astrobiology: new protocols for cell evolution in deep time. Int. J. Astrobiology 11, 217–228. doi: 10.1017/S1473550412000298
Briggs D. E. G. (2003). The role of decay and mineralization in the preservation of soft-bodied fossils. Annu. Rev. Earth Planetary Sci. 31, 275–301. doi: 10.1146/annurev.earth.31.100901.144746
Briggs D. E. G., Bottrell S. H., and Raiswell R. (1991). Pyritization of soft-bodied fossils: Beecher’s Trilobite Bed, Upper Ordovician, New York State. Geology. 19, 1221–1224. doi: 10.1130/0091-7613(1991)019<1221:POSBFB>2.3.CO;2
Briggs D. E. G. and McMahon S. (2016). The role of experiments in investigating the taphonomy of exceptional preservation. Palaeontology 59, 1–11. doi: 10.1111/pala.12219
Brock F., Parkes R. J., and Briggs D. E. G. (2006). Experimental pyrite formation associated with decay of plant material. PALAIOS, 21, 499–506. doi: 10.2110/palo.2005.P05-077R
Buick R. (1990). Microfossil recognition in Archean rocks: an appraisal of spheroids and filaments from a 3500 my old chert-barite unit at North Pole, Western Australia. Palaios 5, 441–459. doi: 10.2307/3514837
Butterfield N. J. (1995). Secular distribution of Burgess-Shale-type preservation. Lethaia 28, 1–13. doi: 10.1111/j.1502-3931.1995.tb01587.x
Butts S. H. (2014). Silicification. Paleontological Soc. Papers 20, 15–34. doi: 10.1017/S1089332600002783
Cady S. L., Farmer J. D., Grotzinger J. P., Schopf J. W., and Steele A. (2003). Morphological biosignatures and the search for life on Mars. Astrobiology 3, 351–368. doi: 10.1089/153110703769016442
Cai Y., Schiffbauer J. D., Hua H., and Xiao S. (2012). Preservational modes in the Ediacaran Gaojiashan Lagerstätte: Pyritization, aluminosilicification, and carbonaceous compression. Palaeogeography Palaeoclimatology Palaeoecol. 326–328, 109–117. doi: 10.1016/J.PALAEO.2012.02.009
Cailleau G., Verrecchia E. P., Braissant O., and Emmanuel L. (2009). The biogenic origin of needle fibre calcite. Sedimentology 56, 1858–1875. doi: 10.1111/j.1365-3091.2009.01060.x
Callefo F., Maldanis L., Teixeira V. C., Abans R. A., de O., Monfredini T., et al. (2019a). Evaluating biogenicity on the geological record with synchrotron-based techniques. Front. Microbiol. 10. doi: 10.3389/fmicb.2019.02358
Callefo F., Ricardi-Branco F., Hartmann G. A., Galante D., Rodrigues F., Maldanis L., et al. (2019b). Evaluating iron as a biomarker of rhythmites — An example from the last Paleozoic ice age of Gondwana. Sedimentary Geology 383, 1–15. doi: 10.1016/j.sedgeo.2019.02.002
Callow R. H. T. and Brasier M. D. (2009). Remarkable preservation microbial mats Neoproterozoic siliciclastic settings: Implications Ediacaran taphonomic models. Earth Sci. Rev. 96, 207–219. doi: 10.1016/j.earscirev.2009.07.002
Cardoso A. R., Basilici G., and Mesquita Á.F. (2024). The Cretaceous palaeodesert of the Sanfranciscana Basin (SE Brazil): A key record to track dissolved evaporites in the West Gondwana. Cretaceous Res. 155, 105788. doi: 10.1016/J.CRETRES.2023.105788
Carlson W. D., Rowe T., Ketcham R. A., and Colbert M. W. (2003). Applications of high-resolution X-ray computed tomography in petrology, meteoritics and palaeontology. Geological Soc. Special Publ. 215, 7–22. doi: 10.1144/GSL.SP.2003.215.01.02
Chan C. S., Fakra S. C., Emerson D., Fleming E. J., and Edwards K. J. (2011). Lithotrophic iron-oxidizing bacteria produce organic stalks to control mineral growth: Implications for biosignature formation. ISME J. 5, 717–727. doi: 10.1038/ismej.2010.173
Corthésy N., Saleh F., Thomas C., Antcliffe J. B., and Daley A. C. (2024). The effects of clays on bacterial community composition during arthropod decay. Swiss J. Palaeontology 143, 1–18. doi: 10.1186/s13358-024-00324-7
Cotroneo S., Schiffbauer J. D., McCoy V. E., Wortmann U. G., Darroch S. A. F., Peng Y., et al. (2016). A new model of the formation of Pennsylvanian iron carbonate concretions hosting exceptional soft-bodied fossils in Mazon Creek, Illinois. Geobiology 14, 543–555. doi: 10.1111/GBI.12197
Crosby C. H. and Bailey J. V. (2012). The role of microbes in the formation of modern and ancient phosphatic mineral deposits. Front. Microbiol. 3. doi: 10.3389/fmicb.2012.00241
Cusack M. (2016). Biomineral electron backscatter diffraction for palaeontology. Palaeontology 59, 171–179. doi: 10.1111/pala.12222
Darroch S. A. F., Laflamme M., Schiffbauer J. D., and Briggs D. E. G. (2012). Experimental formation of a microbial death mask. Palaios 27, 293–303. doi: 10.2110/palo.2011.p11-059r
Davies N. S., Liu A. G., Gibling M. R., and Miller R. F. (2016). Resolving MISS conceptions and misconceptions: A geological approach to sedimentary surface textures generated by microbial and abiotic processes. Earth-Science Rev. 154, 210–246. doi: 10.1016/j.earscirev.2016.01.005
Davies N. S., Shillito A. P., Slater B. J., Liu A. G., and McMahon W. J. (2020). Evolutionary synchrony of Earth’s biosphere and sedimentary-stratigraphic record. Earth-Science Rev. 201, 102979. doi: 10.1016/j.earscirev.2019.102979
De Gregorio B. T., Sharp T. G., Flynn G. J., Wirick S., and Hervig R. L. (2009). Biogenic origin for Earth’s oldest putative microfossils. Geology 37, 631–634. doi: 10.1130/G25683A.1
Delgado A., de O., Buck P. V., Osés G. L., Ghilardi R. P., Rangel E. C., et al. (2014). Paleometry: A brand new area in Brazilian science. Materials Res. 17, 1434–1441. doi: 10.1590/1516-1439.288514
Des Marais D. J., Nuth J. A., Allamandola L. J., Boss A. P., Farmer J. D., Hoehler T. M., et al. (2008). The NASA astrobiology roadmap. Astrobiology 8, 715–730. doi: 10.1089/ast.2008.0819
Dhami N. K., Greenwood P. F., Poropat S. F., Tripp M., Elson A., Vijay H., et al. (2023). Microbially mediated fossil concretions and their characterization by the latest methodologies: a review. Front. Microbiol. 14. doi: 10.3389/fmicb.2023.1225411
Dias J. J. and Carvalho I. de S. (2022). The role of microbial mats in the exquisite preservation of Aptian insect fossils from the Crato Lagerstätte, Brazil. Cretaceous Res. 130, 105068. doi: 10.1016/j.cretres.2021.105068
Dodd M. S., Papineau D., Grenne T., Slack J. F., Rittner M., Pirajno F., et al. (2017). Evidence for early life in Earth’s oldest hydrothermal vent precipitates. Nature 543, 60–64. doi: 10.1038/nature21377
Dornbos S. Q. (2011). Phosphatization through the Phanerozoic in Taphonomy: Process and bias throught time. Eds. Allison P. A. and Bottjer D. J. (Dordrecht: Springer), 435–456. doi: 10.1007/978-90-481-8643-3_12
Dupraz C., Reid R. P., Braissant O., Decho A. W., Norman R. S., and Visscher P. T. (2009). Processes of carbonate precipitation in modern microbial mats. Earth-Science Rev. 96, 141–162. doi: 10.1016/j.earscirev.2008.10.005
Farmer J. D. and Des Marais D. J. (1999). Exploring for a record of ancient Martian life. J. Geophysical Research: Planets 104, 26977–26995. doi: 10.1029/1998JE000540
Farrell Ú. C., Martin M. J., Hagadorn J. W., Whiteley T., and Briggs D. E. (2009). Beyond Beecher's Trilobite Bed: widespread pyritization of soft tissues in the Late Ordovician Taconic foreland basin. Geology 37 (10), 907–910. doi: 10.1130/G30177A
Farrell Ú.C. (2014). Pyritization of soft tissues in the fossil record: an overview. Paleontological Soc. Papers 20, 35–58. doi: 10.1017/S1089332600002795
Farrell U. C., Briggs D. E. G., Hammarlund E. U., Sperling E. A., and Gaines R. R. (2013). Paleoredox and pyritization of soft-bodied fossils in the ordovician Frankfort Shale of New York. Am. J. Sci. 313, 452–489. doi: 10.2475/05.2013.02
Flemming H. C. and Wingender J. (2010). The biofilm matrix. Nat. Rev. Microbiol. 8, 623–633. doi: 10.1038/nrmicro2415
Gäb F., Bierbaum G., Wirth R., Bultmann C., Palmer B., Janssen K., et al. (2024). Enzymatic phosphatization of fish scales—a pathway for fish fossilization. Sci. Rep. 14, 1–11. doi: 10.1038/s41598-024-59025-3
García Ruiz J. M., Carnerup A., Christy A. G., Welham N. J., and Hyde S. T. (2002). Morphology: an ambiguous indicator of biogenicity. Astrobiology 2, 353–369. doi: 10.1089/153110702762027925
Gargaud M., Irvine W. M., Amils R., Cleaves H. J., Pinti D. L., Quintanilla J. C., et al. (2015). Encyclopedia of astrobiology (Berlin: Springer Berlin Heidelberg). doi: 10.1007/978-3-662-44185-5
Georgieva M. N., Little C. T. S., Herrington R. J., Boyce A. J., Zerkle A. L., Maslennikov V. V., et al. (2022). Sulfur isotopes of hydrothermal vent fossils and insights into microbial sulfur cycling within a lower Paleozoic (Ordovician-early Silurian) vent community. Geobiology 20, 465–478. doi: 10.1111/gbi.12495
Gilbert P. U. P. A., Porter S. M., Sun C. Y., Xiao S., Gibson B. M., Shenkar N., et al. (2019). Biomineralization by particle attachment in early animals. Proc. Natl. Acad. Sci. United States America 116, 17659–17665. doi: 10.1073/pnas.1902273116
Gomes A. L. S., Becker-Kerber B., Osés G. L., Prado G., Becker Kerber P., de Barros G. E. B., et al. (2019). Paleometry as a key tool to deal with paleobiological and astrobiological issues: some contributions and reflections on the Brazilian fossil record. Int. J. Astrobiology 18, 575–589. doi: 10.1017/S1473550418000538
Green S. (2022). Polymerized tubular silicates in lower cambrian carbonates– biology or chemistry? [Dissertation thesis. Uppsala University, Uppsala. Available online at: https://urn.kb.se/resolve?urn=urn:nbn:se:uu:diva-476830.
Grimes S. T., Brock F., Rickard D., Davies K. L., Edwards D., Briggs D. E. G., et al. (2001). Understanding fossilization: Experimental pyritization of plants. Geology 29, 123–126. doi: 10.1130/0091-7613(2001)029<0123:UFEPOP>2.0.CO;2
Gueriau P., Mocuta C., Dutehil D. B., Cohen S. X., Thiaudière, Charbonnier S., et al. (2014). Trace elemental imaging of rare earth elements discriminates tissues at microscale in flat fossils. PLOS ONE 9 (1), 1–9. e86946. doi: 10.1371/journal.pone.0086946
Hofmann H. J. (1972) Precambrian remains in Canada: fossils, dubio fossils, and pseudofossils. In: International Geological Congress, 24th Session, Proceedings, Montreal. 1, p. 20–30.
Hofmann B. A., Farmer J. D., von Blanckenburg F., and Fallick A. E. (2008). Subsurface filamentous fabrics: an evaluation of origins based on morphological and geochemical criteria, with implications for exopaleontology. Astrobiology 8, 87–117. doi: 10.1089/ast.2007.0130
Inglez L., Warren L. V., Quaglio F., Netto R. G., Okubo J., Arrouy M. J., et al. (2021). Scratching the discs: evaluating alternative hypotheses for the origin of the Ediacaran discoidal structures from the Cerro Negro Formation, La Providencia Group, Argentina. Geological Magazine 159, 1192–1209. doi: 10.1017/S0016756821000327
Ivarsson M., Drake H., Neubeck A., Snoeyenbos-West O., Belivanova V., and Bengtson S. (2021). Introducing palaeolithobiology. GFF 143 (2–3), 305–319. doi: 10.1080/11035897.2021.1895302
Ivarsson M., Drake H., Neubeck A., Sallstedt T., Bengtson S., Roberts N. M. W., et al. (2020). The fossil record of igneous rock. Earth-Science Rev. 210, 103342. doi: 10.1016/j.earscirev.2020.103342
Javaux E. J., Knoll A. H., and Walter M. (2003). Recognizing and interpreting the fossils of early eukaryotes. Origins Life Evol. Biosphere 33, 75–94. doi: 10.1023/A:1023992712071
Johannessen K. C., McLoughlin N., Vullum P. E., and Thorseth I. H. (2020). On the biogenicity of Fe-oxyhydroxide filaments in silicified low-temperature hydrothermal deposits: Implications for the identification of Fe-oxidizing bacteria in the rock record. Geobiology 18, 31–53. doi: 10.1111/gbi.12363
Ketzer J. M., Holz M., Morad S., and Al-Aasm I. S. (2003). Sequence stratigraphic distribution of diagenetic alterations in coal-bearing, paralic sandstones: evidence from the Rio Bonito Formation (early Permian), southern Brazil. Sedimentology 50, 855–877. doi: 10.1046/J.1365-3091.2003.00586.X
Klug C., Silvestro G., Hoffmann R., Schweigert G., Fuchs D., Clements T., et al. (2021). Taphonomic patterns mimic biologic structures: Diagenetic Liesegang rings in Mesozoic coleoids and coprolites. PeerJ 9. doi: 10.7717/peerj.10703
Knoll A. H. (2013). Systems paleobiology. Geological Soc. America Bull. 125, 3–13. doi: 10.1130/B30685.1
Laflamme M., Schiffbauer J. D., Narbonne G. M., and Briggs D. E. G. (2011). Microbial biofilms and the preservation of the Ediacara biota. Lethaia 44, 203–213. doi: 10.1111/j.1502-3931.2010.00235.x
Li J., Benzerara K., Bernard S., and Beyssac O. (2013). The link between biomineralization and fossilization of bacteria: Insights from field and experimental studies. Chem. Geology 359, 49–69. doi: 10.1016/j.chemgeo.2013.09.013
Liu A. G. (2016). Framboidal pyrite shroud confirms the ‘death mask’ model for moldic preservation of Ediacaran soft-bodied organisms. Palaios 31, 259–274. doi: 10.2110/palo.2015.095
Liu A. G., Mcmahon S., Matthews J. J., Still J. W., and Brasier A. T. (2019). Petrological evidence supports the death mask model for the preservation of Ediacaran soft-bodied organisms in South Australia. Geology 47, 215–218. doi: 10.1130/G45918.1
Lowe D. R. (1995). Abiological origin of described stromatolites older than 3.2 Ga: reply. Geology 23, 191–192.
Lucas S. G. and Lerner A. J. (2017). Gallery of geology: the rare and unusual pseudofossil Astropolithon from the lower permian abo formation near Socorro, New Mexico. New Mexico Geology 39, 40–42. doi: 10.58799/NMG-v39n2.40
MacLean L. C. W., Tyliszczak T., Gilbert P. U. P. A., Zhou D., Pray T. J., Onstott T. C., et al. (2008). A high-resolution chemical and structural study of framboidal pyrite formed within a low-temperature bacterial biofilm. Geobiology 6, 471–480. doi: 10.1111/j.1472-4669.2008.00174.x
Mähler B., Janssen K., Menneken M., Tahoun M., Lagos M., Bierbaum G., et al. (2020). Calcite precipitation forms crystal clusters and muscle mineralization during the decomposition of cambarellus diminutus (Decapoda: Cambaridae) in freshwater. Palaeontologia Electronica 23, 1–17. doi: 10.26879/992
Malaterre C., ten Kate I. L., Baqué M., Debaille V., Grenfell J. L., Javaux E. J., et al. (2023). Is there such a thing as a biosignature? Astrobiology 23, 1213–1227. doi: 10.1089/ast.2023.0042
Maldanis L., Hickman-Lewis K., Verezhak M., Gueriau P., Guizar-Sicairos M., Jaqueto P., et al. (2020). Nanoscale 3D quantitative imaging of 1.88 Ga Gunflint microfossils reveals novel insights into taphonomic and biogenic characters. Sci. Rep. 10, 1–9. doi: 10.1038/s41598-020-65176-w
Maliva R. G., Knoll A. H., Simonson B. M., Maliva R. G., Knoll A. H., and Simonson B. M. (2005). Secular change in the Precambrian silica cycle: Insights from chert petrology Secular change in the Precambrian silica cycle: Insights from chert petrology. Geological Soc. America Bull. 117. doi: 10.1130/B25555.1
Marshall C. R. (2006). Explaining the Cambrian explosion of animals. Annu. Rev. Earth Planet. Sci. 34, 355–384. doi: 10.1146/annurev.earth.33.031504.103001
Marshall A. O., Emry J. R., and Marshall C. P. (2012). Multiple generations of carbon in the apex chert and implications for preservation of microfossils. Astrobiology 12, 160–166. doi: 10.1089/ast.2011.0729
Marshall A. O. and Marshall C. P. (2015). Vibrational spectroscopy of fossils. Palaeontology 58, 201–211. doi: 10.1111/pala.12144
Marshall J. D. and Pirrie D. (2013). Carbonate concretions-explained. Geology Today 29, 53–62. doi: 10.1111/GTO.12002
Martins A. K., Kerkhoff M. L. H., Dutra T. L., Horodyski R. S., Kochhann K. G. D., and Forancelli Pacheco M. L. A. (2022). Exceptional preservation of Triassic-Jurassic fossil plants: integrating biosignatures and fossil diagenesis to understand microbial-related iron dynamics. Lethaia 55, 1–16. doi: 10.18261/let.55.3.4
McCollom T. and Seewald J. (2006). Carbon isotope composition of organic compounds produced by abiotic synthesis under hydrothermal conditions. Earth Planetary Sci. Lett. 243, 74–84. doi: 10.1016/j.epsl.2006.01.027
McCoy V. E. (2014). Concretions as agents of soft-tissue preservation: A review. Paleontological Soc. Papers 20, 147–162. doi: 10.1017/S1089332600002849
Mccoy V. E., Young R. T., and Briggs D. E. G. (2015a). Factors controlling exceptional preservation in concretions. PALAIOS 30, 272–280. doi: 10.2110/PALO.2014.081
McLoughlin S. (2011). New records of leaf galls and arthropod oviposition scars in Permian–Triassic Gondwanan gymnosperms. Aust. J. Bot. 59, 156–169. doi: 10.1071/BT10297
McLoughlin N. and Grosch E. G. (2015b). A hierarchical system for evaluating the biogenicity of metavolcanic- and ultramafic-hosted microalteration textures in the search for extraterrestrial life. Astrobiology 15, 901–921. doi: 10.1089/ast.2014.1259
McLoughlin N., Wilson L. A., and Brasier M. D. (2008). Growth of synthetic stromatolites and wrinkle structures in the absence of microbes - Implications for the early fossil record. Geobiology 6, 95–105. doi: 10.1111/j.1472-4669.2007.00141.x
McMahon S. (2019). Earth’s earliest and deepest purported fossils may be iron-mineralized chemical gardens. Proc. R. Soc. B: Biol. Sci. 286, 20192410. doi: 10.1098/rspb.2019.2410
McMahon S. and Cosmidis J. (2022). False biosignatures on Mars: anticipating ambiguity. J. Geological Soc. 179. doi: 10.1144/jgs2021-050
McMahon S. and Ivarsson M. (2019). A new frontier for palaeobiology: earth’s vast deep biosphere. BioEssays 41, 1900052. doi: 10.1002/bies.201900052
McMahon S., Ivarsson M., Wacey D., Saunders M., Belivanova V., Muirhead D., et al. (2021). Dubiofossils from a Mars-analogue subsurface palaeoenvironment: The limits of biogenicity criteria. Geobiology 19, 473–488. doi: 10.1111/gbi.12445
Melim L., Terrile A. S., Hegna T., Bellott B. J., and Lerosey-Aubril R. (2023). New evidence for bacterial mediation of the silicification of carbonate fossils. GSAA 55, 394704. doi: 10.1130/ABS/2023AM-394704
Merino N., Aronson H. S., Bojanova D. P., Feyhl-Buska J., Wong M. L., Zhang S., et al. (2019). Living at the extremes: extremophiles and the limits of life in a planetary context. Front. Microbiol. 10. doi: 10.3389/fmicb.2019.00780
Milliken K. L., Diagenesis, In Middleton G. V., Church M. J., Coniglio M., Hardie L. A., et al. (1978). Encyclopedia of sediments and sedimentary rocks (Dordrecht: Springer Netherlands), 339–349. doi: 10.1007/978-1-4020-3609-5_66
Miot J., MacLellan K., Benzerara K., and Boisset N. (2011). Preservation of protein globules and peptidoglycan in the mineralized cell wall of nitrate-reducing, iron(II)-oxidizing bacteria: A cryo-electron microscopy study. Geobiology 9, 459–470. doi: 10.1111/j.1472-4669.2011.00298.x
Monroe J. S. and Dietrich R. V. (1990). Pseudofossils. Rocks Minerals 65, 150–158. doi: 10.1080/00357529.1990.11761667
Moore K. R., Gong J., Pajusalu M., Skoog E. J., Xu M., Feliz T., et al. (2021). A new model for silicification of cyanobacteria in Proterozoic tidal flats. Geobiology 19, 438–449. doi: 10.1111/gbi.12447
Muscente A. D., Schiffbauer J. D., Broce J., Laflamme M., O’Donnell K., Boag T. H., et al. (2017). Exceptionally preserved fossil assemblages through geologic time and space. Gondwana Res. 48, 164–188. doi: 10.1016/j.gr.2017.04.020
Neveu M., Hays L. E., Voytek M. A., New M. H., and Schulte M. D. (2018). The ladder of life detection. Astrobiology 18, 1375–1402. doi: 10.1089/ast.2017.1773
Noffke N. (2009). The criteria for the biogeneicity of microbially induced sedimentary structures (MISS) in Archean and younger, sandy deposits. Earth-Science Rev. 96, 173–180. doi: 10.1016/j.earscirev.2008.08.002
Noffke N. (2010). Geobiology (Berlin, Heidelberg: Springer Berlin Heidelberg). doi: 10.1007/978-3-642-12772-4
Noffke N. (2021). Microbially induced sedimentary structures in clastic deposits: implication for the prospection for fossil life on mars. Astrobiology 21, 866–892. doi: 10.1089/ast.2021.0011
Noffke N. and Awramik S. M. (2013). Stromatolites and MISS-differences between relatives. GSA Today 23, 4–9. doi: 10.1130/GSATG187A.1
Noffke N., Eriksson K. A., Hazen R. M., and Simpson E. L. (2006). A new window into Early Archean life: Microbial mats in Earth’s oldest siliciclastic tidal deposits (3.2 Ga Moodies Group, South Africa). Geology 34, 253–256. doi: 10.1130/G22246.1
Noffke N., Gerdes G., Klenke T., and Krumbein W. E. (2001). Microbially induced sedimentary structures–a new category within the classification of primary sedimentary structures–reply. J. Sedimentary Res. 72, 589–590. doi: 10.1306/010302720589
Noffke N., Hazen R., and Nhleko N. (2003). Earth’s earliest microbial mats in a siliciclastic marine environment (2.9 Ga Mozaan Group, South Africa). Geology 31, 673. doi: 10.1130/G19704.1
Osés G. L. (2016). Tafonomia de grupos fó́sseis do Membro Crato (Formaç̧ão Santana, Bacia do Araripe, Eocretácio, NE do Brasil): Implicações geobiológicas, paleoecoloógicas e paleoambientais [dissertation thesis]. [São Carlos, BRA]. Universidade de São Carlos.
Osés G. L., Petri S., Becker-Kerber B., Romero G. R., Rizzutto M., de A., et al. (2016). Deciphering the preservation of fossil insects: A case study from the Crato Member, Early Cretaceous of Brazil. PeerJ 2016, e2756. doi: 10.7717/PEERJ.2756/SUPP-1
Osés G. L., Petri S., Voltani C. G., Prado G. M. E. M., Galante D., and Rizzutto M. A. (2017). Deciphering pyritization-kerogenization gradient for fish soft-tissue preservation. Sci. Rep. 7, 1–15. doi: 10.1038/s41598-017-01563-0
Osés G. L., Wood R., Romero G. R., Evangelista Martins Prado G. M., Bidola P., and Herzen J. (2022). Ediacaran Corumbella has a cataphract calcareous skeleton with controlled biomineralization. iScience 25. doi: 10.1016/j.isci.2022.105676
Osterhout J. T., Schopf J. W., Williford K. H., McKeegan K. D., Kudryavtsev A. B., and Liu M.-C. (2021). Carbon isotopes of Proterozoic filamentous microfossils: SIMS analyses of ancient cyanobacteria from two disparate shallow-marine cherts. Geomicrobiology J. 38, 719–731. doi: 10.1080/01490451.2021.1939813
Pan Y., Hu L., and Zhao T. (2019). Applications of chemical imaging techniques in paleontology. Natl. Sci. Rev. 6, 1040–1053. doi: 10.1093/NSR/NWY107
Pasteris J. D. and Wopenka B. (2003). Necessary, but not sufficient: raman identification of disordered carbon as a signature of ancient life. Astrobiology 3, 727–738. doi: 10.1089/153110703322736051
Pérez-Huerta A., Coronado I., and Hegna T. A. (2018). Understanding biomineralization in the fossil record. Earth-Science Rev. 179, 95–122. doi: 10.1016/j.earscirev.2018.02.015
Porada H., Ghergut J., and Bouougri E. H. (2008). Kinneyia-type wrinkle structures–critical review and model of formation. PALAIOS 23, 65–77. doi: 10.2110/palo.2006.p06-095r
Prado G., Arthuzzi J. C. L., Osés G. L., Callefo F., Maldanis L., Sucerquia P., et al. (2021). Synchrotron radiation in palaeontological investigations: Examples from Brazilian fossils and its potential to South American palaeontology. J. South Am. Earth Sci. 108, 102973. doi: 10.1016/j.jsames.2020.102973
Pruss S. B., Blättler C. L., Macdonald F. A., and Higgins J. A. (2018). Calcium isotope evidence that the earliest metazoan biomineralizers formed aragonite shells. Geology 46, 763–766. doi: 10.1130/G45275.1
Riquelme F., Ruvalcaba Sil J. L., and Alvarado Ortega J. (2009). Palaeometry: Non-destructive analysis of fossil materials. Boletín la Sociedad Geológica Mexicana 61, 177–183. doi: 10.18268/BSGM2009v61n2a4
Rouillard J., van Zuilen M., Pisapia C., and Garcia-Ruiz J.-M. (2021). An alternative approach for assessing biogenicity. Astrobiology 21, 151–164. doi: 10.1089/ast.2020.2282
Sagemann J., Bale S. J., Briggs D. E. G., and Parkes R. J. (1999). Controls on the formation of authigenic minerals in association with decaying organic matter: an experimental approach. Geochimica Cosmochimica Acta 63, 1083–1095. doi: 10.1016/S0016-7037(99)00087-3
Saldanha J. P., Cagliari J., Scalise Horodyski R., Del Mouro L., and Alves Forancelli Pacheco M. L. (2023). Deciphering the origin of dubiofossils from the Pennsylvanian of the Paraná Basin, Brazil. Biogeosciences 20, 3943–3979. doi: 10.5194/bg-20-3943-2023
Sanchez E. A. M., Romero G. R., Callefo F., Cardoso A. R., and Fairchild T. R. (2023). A guide for microscopic description of fossil stromatolites. Braz. J. Geology 53. doi: 10.1590/2317-4889202220220031
Schiffbauer J. D., Xiao S., Cai Y., Wallace A. F., Hua H., Hunter J., et al. (2014). A unifying model for Neoproterozoic-Palaeozoic exceptional fossil preservation through pyritization and carbonaceous compression. Nat. Commun. 5, 5754. doi: 10.1038/ncomms6754
Schiffbauer J. D., Yin L., Bodnar R. J., Kaufman A. J., Meng F., Hu J., et al. (2007). Ultrastructural and geochemical characterization of archean–paleoproterozoic graphite particles: implications for recognizing traces of life in highly metamorphosed rocks. Astrobiology 7, 684–704. doi: 10.1089/ast.2006.0098
Schopf J. W. and Kudryavtsev A. B. (2012). Biogenicity of Earth’s earliest fossils: A resolution of the controversy. Gondwana Res. 22, 761–771. doi: 10.1016/j.gr.2012.07.003
Schopf J. W., Kudryavtsev A. B., Agresti D. G., Czaja A. D., and Wdowiak T. J. (2005). Raman imagery: A new approach to assess the geochemical maturity and biogenicity of permineralized Precambrian fossils. Astrobiology 5, 333–371. doi: 10.1089/ast.2005.5.333
Schopf J. W., Kudryavtsev A. B., Czaja A. D., and Tripathi A. B. (2007). Evidence of Archean life: Stromatolites and microfossils. Precambrian Res. 158, 141–155. doi: 10.1016/j.precamres.2007.04.009
Schopf J. W., Kudryavtsev A. B., Osterhout J. T., Williford K. H., Kitajima K., Valley J. W., et al. (2017). An anaerobic ∼3400 Ma shallow-water microbial consortium: Presumptive evidence of Earth’s Paleoarchean anoxic atmosphere. Precambrian Res. 299, 309–318. doi: 10.1016/J.PRECAMRES.2017.07.021
Schopf J. W., Kudryavtsev A. B., Sugitani K., and Walter M. R. (2010). Precambrian microbe-like pseudofossils: A promising solution to the problem. Precambrian Res. 179, 191–205. doi: 10.1016/j.precamres.2010.03.003
Schopf J., Kudryavtsev A., Agresti D., Wdowiak T. J., and Czaja A. D. (2002). Laser–Raman imagery of Earth's earliest fossils. Nature 416, 73–76. doi: 10.1038/416073a
Seilacher A. (2001). Concretion morphologies reflecting diagenetic and epigenetic pathways. Sedimentary Geology 143, 41–57. doi: 10.1016/S0037-0738(01)00092-6
Servais T., Cascales-Miñana B., Harper D. A., Lefebvre B., Munnecke A., Wang W., et al. (2023). No (Cambrian) explosion and no (Ordovician) event: a single long-term radiation in the early Palaeozoic. Palaeogeogr. Palaeoclimatol. Palaeoecol. 623, 111592. doi: 10.1016/j.palaeo.2023.111592
Slagter S., Hao W., Planavsky N. J., Konhauser K. O., and Tarhan L. G. (2022). Biofilms as agents of Ediacara-style fossilization. Sci. Rep. 12, 8631. doi: 10.1038/s41598-022-12473-1
Slater G. F. (2009). Biosignatures: interpreting evidence of the origins and diversity of life. Geosci. Canada 36, 170–178.
Stern B. (1996). Biomineral lipids in living and fossil molluscs. [Doctoral dissertation]. Newcastle University, Newcastle.
Sugitani K., Grey K., Allwood A., Nagaoka T., Mimura K., Minami M., et al. (2007). Diverse microstructures from Archaean chert from the Mount Goldsworthy–Mount Grant area, Pilbara Craton, Western Australia: Microfossils, dubiofossils, or pseudofossils? Precambrian Res. 158, 228–262. doi: 10.1016/j.precamres.2007.03.006
Tarhan L. G., Hood A., v. S., and Droser M. L. (2025). Elevated marine dissolved silica levels explain a wide range of Ediacaran–Cambrian Ediacara-style fossil deposits. Geobiology 23, e70017. doi: 10.1111/GBI.70017
Thompson C. K. and Kah L. C. (2012). Sulfur isotope evidence for widespread euxinia and a fluctuating oxycline in Early to Middle Ordovician greenhouse oceans. Palaeogeogr. Palaeoclimatol. Palaeoecol. 313–314, 189–214. doi: 10.1016/j.palaeo.2011.10.020
Tisato N., Torriani S. F. F., Monteux S., Sauro F., De Waele J., Tavagna M. L., et al. (2015). Microbial mediation of complex subterranean mineral structures. Sci. Rep. 5, 15525. doi: 10.1038/srep15525
Trinajstic K., Long J. A., Sanchez S., Boisvert C. A., Snitting D., Tafforeau P., et al. (2022). Exceptional preservation of organs in Devonian placoderms from the Gogo Lagerstätte. Science 377, 1311–1314. doi: 10.1126/SCIENCE.ABF3289/SUPPL_FILE/SCIENCE.ABF3289_DATA_S1_TO_S4.ZIP
Trueman C. N. (2013). Chemical taphonomy of biomineralized tissues. Palaeontology 56, 475–486. doi: 10.1111/pala.12041
Van Lith Y., Warthmann R., Vasconcelos C., and McKenzie J. A. (2003). Microbial fossilization in carbonate sediments: A result of the bacterial surface involvement in dolomite precipitation. Sedimentology 50, 237–245. doi: 10.1046/j.1365-3091.2003.00550.x
Voltani C. G., Osés G. L., Freitas B. T., Prado G. M. E. M., Rohn R., Pacheco M. L. A. F., et al. (2023). Taphonomy of fish, invertebrates and plant remains in the first Tethyan-South Atlantic marine ingression along Cretaceous rift systems in NE-Brazil. Cretaceous Res. 147, 105508. doi: 10.1016/J.CRETRES.2023.105508
Wacey D. (2009). Early life on earth: a practical guide. Dordrecht: Springer Netherlands. 274 p. doi: 10.1007/978-1-4020-9389-0_4
Wacey D. (2010). Stromatolites in the ∼3400 ma strelley pool formation, western Australia: examining biogenicity from the macro- to the nano-scale. Astrobiology 10, 381–395. doi: 10.1089/ast.2009.0423
Wang Q., Wang X. L., Zhao Z. K., and Jiang Y. G. (2012). A new oofamily of dinosaur egg from the Upper Cretaceous of Tiantai Basin, Zhejiang Province, and its mechanism of eggshell formation. Chin. Sci. Bull. 57, 3740–3747. doi: 10.1007/S11434-012-5353-2/METRICS
Weiner S. and Dove P. (2003). An overview of biomineralization processes and the problem of the vital effect. Rev. Mineralogy Geochemistry 54, 1–29. doi: 10.2113/0540001
Westall F. (2008). Morphological biosignatures in early terrestrial and extraterrestrial materials. Space Sci. Rev. 135, 95–114. doi: 10.1007/s11214-008-9354-z
Westall F., Foucher F., Bost N., Bertrand M., Loizeau D., Vago J. L., et al. (2015). Biosignatures on mars: what, where, and how? Implications for the search for martian life. Astrobiology 15, 998–1029. doi: 10.1089/ast.2015.1374
Wilby P. R. and Briggs D. E. G. (1997). Taxonomic trends in the resolution of detail preserved in fossil phosphatized soft tissues. Geobios 30, 493–502. doi: 10.1016/S0016-6995(97)80056-3
Wilby P. R., Briggs D. E. G., Bernier P., and Gaillard C. (1996). Role of microbial mats in the fossilization of soft tissues. Geology 24, 787–790. doi: 10.1130/0091-7613(1996)024<0787:ROMMIT>2.3.CO;2
Wilson L. A. and Butterfield N. J. (2014). Sediment effects on the preservation of burgess shale-type compression fossils. Palaios 29, 145–154. doi: 10.2110/palo.2013.075
Wilson P., Parry L. A., Vinther J., and Edgecombe G. D. (2016). Unveiling biases in soft-tissue phosphatization: Extensive preservation of musculature in the Cretaceous (Cenomanian) polychaete Rollinschaeta myoplena (Annelida: Amphinomidae). Palaeontology 59, 463–479. doi: 10.1111/pala.12237
Wood R., Bowyer F., Penny A., and Poulton S. W. (2018). Did anoxia terminate Ediacaran benthic communities? Evidence from early diagenesis. Precambrian Research 313, 134–147. doi: 10.1016/j.precamres.2018.05.011
Wood J. R., Paul Scofield R., Hamel J., Lalas C., and Wilmshurst J. M. (2017). Short communication: Bone stable isotopes indicate a high trophic position for New Zealand’s extinct South Island adzebill (Aptornis defossor) (Gruiformes: Aptornithidae). New Z. J. Ecol. 41. doi: 10.20417/NZJECOL.41.24
Worden R. H. and Burley S. D. (2009). “Sandstone diagenesis: the evolution of sand to stone,” in Sandstone diagenesis (Blackwell Publishing Ltd., Oxford, UK), 1–44. doi: 10.1002/9781444304459.ch
Xiao S., Schiffbauer J. D., McFadden K. A., and Hunter J. (2010). Petrographic and SIMS pyrite sulfur isotope analyses of Ediacaran chert nodules: Implications for microbial processes in pyrite rim formation, silicification, and exceptional fossil preservation. Earth Planetary Sci. Lett. 297, 481–495. doi: 10.1016/j.epsl.2010.07.001
Zhang Y., Sun H., Stowell H. H., Zayernouri M., and Hansen S. E. (2017). A review of applications of fractional calculus in Earth system dynamics. Chaos Solitons Fractals 102, 29–46. doi: 10.1016/j.chaos.2017.03.051
Keywords: biosignatures, dubiofossil, biomineralization, biominerals, diagenesis, abiotic process, high-resolution analytical techniques
Citation: Saldanha JP, Martins AK, Osés GL and Callefo F (2025) Using paleometry to address biogenicity in paleobiology. Front. Ecol. Evol. 13:1646756. doi: 10.3389/fevo.2025.1646756
Received: 13 June 2025; Accepted: 03 September 2025;
Published: 29 September 2025.
Edited by:
Dina M. Bower, University of Maryland, College Park, United StatesReviewed by:
Seung Choi, Seoul National University, Republic of KoreaAhmed Abdelhady, Minia University, Egypt
Copyright © 2025 Saldanha, Martins, Osés and Callefo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: João Pedro Saldanha, c2FsZGFuaGFqcGVkcm9AZ21haWwuY29t