 Renato Crespo Pereira
Renato Crespo Pereira Ygor Jessé Ramos
Ygor Jessé Ramos Davyson de Lima Moreira3
Davyson de Lima Moreira3- 1Laboratory of Marine Chemical Ecology, Institute of Biology, Department of Marine Biology, Federal Fluminense University, Niterói, Rio de Janeiro, Brazil
- 2Earth Laboratory, Pharmacy College, Federal University of Bahia, Salvador, Brazil
- 3Laboratory of Natural Products and Biochemistry, Research Department, Rio de Janeiro Botanical Garden Research Institute, Rio de Janeiro, Brazil
Since time immemorial, a wide range of terrestrial, aquatic, and marine organisms have been used to treat illnesses and diseases, owing to the broad diversity of natural products they produce. Although widely recognized for their medicinal properties, natural products are fundamentally the result of adaptive chemistry - or phenotypic expression - of the organisms that produce them, shaped by evolutionary pressures from natural environmental enemies. This article examines natural products through ecological, evolutionary, and conservation lenses, highlighting how these perspectives add complexity to the already lengthy and costly process of bioprospecting. Due to their origin and evolutionary history, only a small fraction of the many screened natural products exhibits biological activity relevant to human health, as these compounds did not evolve to possess pharmacological properties. Our viewpoint emphasizes the true raison d’être of the vast diversity of natural products and argues that this perspective should be integrated into bioprospecting efforts. We propose focus on the ecological roles of natural products to offer a more rational, productive, and sustainable approach to drug discovery - one that aligns with biodiversity conservation and economic development.
1 Introduction
Medicinal plants have supported human health for millennia, as evidenced by ancient records across cultures from Mesopotamia to China (Arya et al., 2022). Ethnobotanical traditions and early pharmacological texts document extensive plant-based treatments, while aquatic and marine organisms have also contributed bioactive compounds to traditional medicine (Jia et al., 2004; Aasim et al., 2019). Over time, the isolation of plant secondary metabolites has led to major breakthroughs in treating cancer, infections, and neurological diseases, positioning natural products as vital resources in modern healthcare, pharmacology, and biotechnology (Papon et al., 2022; Labes, 2023; Rojo et al., 2023; Silveria and Boylan, 2023.
This article does not aim to provide a historical overview, but rather to reframe natural products not simply as drug leads, but as evolved adaptive traits shaped by ecological and evolutionary pressures. By highlighting their raison d’être, we argue for a shift toward a chemical ecology perspective - promoting sustainable, efficient, and biodiversity-informed bioprospecting.
2 Search strategy for scientific articles related to the raison d’être of natural products
To support our insights on the importance of considering the raison d’être of natural products in bioprospecting, a systematic search was conducted with the aim of answering the question: How has the integration of ecological and evolutionary approaches in marine, aquatic, and/or terrestrial bioprospecting contributed to the discovery of bioactive natural products with pharmaceutical potential? The methodological design followed the recommendations of the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA, Moher et al., 2009) to ensure rigor, transparency, and reproducibility through stages such as identification, screening, eligibility, and inclusion of studies.
Data collection took place in August 2024 and used the following indexed 4 databases: PubMed/MEDLINE, Scopus, Web of Science, and SciELO/LILACS. The search strategy was structured using a combination of controlled descriptors and keywords in English, Portuguese, and Spanish, articulated by Boolean operators (AND/OR). The main terms used were: ‘bioprospecting’, ‘natural products’, ‘metabolites’, ‘marine organisms’, ‘terrestrial plants’, ‘aquatic organisms’, ‘aquatic plants’, ‘chemical ecology’, ‘evolutionary approach’, ‘bioactive compounds’, ‘biotechnology’, and ‘pharmaceuticals’. A detailed description of the procedures used in the search (Supplementary Material) and workflow for selecting the relevant articles can be found in the Supplementary Material (Supplementary Table S1, Supplementary Material). Despite all the methodological rigor employed, it is not our intention to conduct a review, but rather to justify the innovative approach of the present perspective article.
The systematic search identified 115 records in the selected databases, PubMed/MEDLINE (n= 10), Scopus (n= 55), Web of Science (n= 49) and SciELO/LILACS (n= 1). After removing 29 duplicates, 86 records remained for initial evaluation. Of these, 14 studies were excluded for not meeting the minimum eligibility criteria. Thus, 72 records proceeded to the title and abstract screening phase, during which 24 were eliminated for not directly addressing bioprospecting or for not employing an ecological/evolutionary approach. In the full-text reading phase, 48 studies were assessed, with 24 excluded (4 due to being of an unsuitable study type and 20 for presenting a methodological approach outside the defined scope). In the end, 24 studies were included in the systematic review and comprised the analysis corpus (Supplementary Table S2, Supplementary Material). However, none of these 24 articles (Supplementary Table S2, Supplementary Material) explored the purpose of natural products or the importance of considering their ecological roles and evolution in the bioprospecting of natural products. This finding underscores the relevance of our insight, emphasizing the importance of incorporating the raison d’être of natural products into bioprospecting.
3 General aspects on the development of pharmaceuticals
In the early stages of drug discovery, the process typically consists of two main steps: the collection of biological samples and the evaluation of its extracts for bioactivities. However, identifying promising biological activity is only the beginning of a long, expensive and complex process (Firn, 2003). Drug development is notoriously costly – estimate at approximately $ 900 million – and can take around 14 years from discovery to market approval. Moreover, only a small number of potential candidates, referred to as “hits”, exhibit sufficient bioactivity to warrant further investigation as viable drug candidates (DiMasi et al., 2003). From these hits, a “lead” compound is selected for preclinical studies (Hunt and Vicent, 2006).
Screening of bioactive chemicals is generally conducted through one or more of the following approaches: (1) Random screening, (2) Ethnopharmacological knowledge or (3) The examination of chemical diversity within a particular taxon in comparison to another previously studied bioactive compounds. However, all these approaches, bioprospecting can lead to significative negative ecological consequences. For example, large-scale bioprospecting can lead to populations decline, loss of genetic diversity, genetic drift, and even local extinction of source species (DiMasi, 2001). These concerns raise important questions about the sustainability of a widespread biological sampling. Therefore, these bioprospecting strategies – whether random, ethnopharmacological, or taxonomically guided – may not always be environmentally sustainable.
In recent years, the advanced analytical tools such as Gas Chromatography-Mass Spectrometry (CG-MS) and/or Liquid Chromatography-Mass Spectrometry in tandem (LC-MS/MS), often combined with bioinformatics database resources like the Global Natural Products Social Molecular Networking (GNPS) have helped to mitigate these impacts (Cachet et al., 2015; Raina et al., 2022). These methods require only a few milligrams of material and allow for high throughput, environmentally conscious screening. Additionally, artificial intelligence and advanced in vitro technologies are being employed to streamline the drug development pipeline and reduce associated costs and timelines – though many challenges remain (Singh et al., 2023).
Once a promising compound is identified, however, the process of isolation and characterization represent a major bottleneck. Bioactive compounds are typically produced in very small quantities by their source organisms. For instance, several marine-derived secondary metabolites - including ecteinascidin 743 from the ascidian Ecteinascidia turbinata, bryostatins from the bryozoan Bugula neritina, and halichondrins from the sponge Lissodendoryx, can only be isolated in trace quantities (Pereira and Costa-Lotufo, 2012). As a specific example, therapeutic use of halichondrins in cancer treatment would require 1–5 kg annually, which would necessitate harvesting between 3,000 –16,000 metric tons of sponge biomass per year (Proksch et al., 2003).
In summary, although bioprospecting has led to important discoveries, its current models remain conceptually fragmented and environmentally unsustainable. The predominant reliance on random screening or ethnopharmacological knowledge often fails to incorporate ecological principles, leading to costly and low-yield outcomes. Moreover, these approaches typically overlook the adaptive nature of natural products, treating them as pharmacological candidates rather than evolved traits. This disconnect highlights the need for transdisciplinary frameworks that not only integrate chemical, biological, and ecological knowledge, but also reframe bioprospecting as a process informed by eco-evolutionary logic (Rotter et al., 2020). However, it is necessary and urgent that bioprospecting and its associated costs be minimized through the recognition of the true origin and evolution of natural products in the organisms that produce them.
Despite more than a century of research, many questions remain about the functions and ecological roles of natural products in the organisms that produce them (Dixon et al., 2024). But while the origin of secondary metabolites is not fully understood, it is widely accepted that they evolved to facilitate chemical defense, inter-organism communication, and competition within ecological communities. Thus, regardless of the procedure used in the search for bioactive substances, the origin and evolution of natural products cannot be disregarded, as this is the main foundation of our current insight.
4 Ecological and evolutionary views on the bioprospecting area
Among the major challenges in natural product bioprospecting is the low success rate in discovering new drug leads (Liu et al., 2019). For over two decades, bioprospecting has been viewed as economically discouraging due to high costs and the low probability that a given compound will display meaningful biological activity (Firn, 2003; Liu et al., 2019). However, in our view, a more fundamental limitation lies in the fact that natural products have evolved – through natural selection – to function primarily as chemical mediators in ecological contexts in the natural environments (terrestrial, marine, and aquatic), rather than as therapeutic agents for human use. For this simple reason, many molecules are tested, but only a small fraction exhibit biological activity. This outcome is not surprising if one considers that current screening strategies are largely blind to the ecological context in which natural products evolved. The assumption that any given metabolite might display therapeutic activity disregards its original adaptive function. This methodological oversight represents a critical limitation in traditional bioprospecting pipelines, which often prioritize pharmacological potential over ecological relevance. A more productive strategy would reverse this logic, beginning with ecological function as a filter before pharmacological testing, thereby increasing both efficiency and sustainability.
This idea is encapsulated in the known screening hypothesis, which suggest that potent biological activity is an uncommon property among natural products, given that most evolved to exhibit ecological roles rather than interact with human disease targets (Jones and Firn, 1991). This hypothesis was a pioneering attempt to conceptualize an evolutionary scenario in which significant constraints on the evolution of natural product diversity would have existed, given that most natural products did not possess a potent biological activity beneficial to the producer (Firn and Jones, 2003). As a result, bioprospecting screens tens of thousands of molecules, but only a small fraction shows promise as drug candidates. There is an evolutionary history behind the phenotypic expression of thousands of small molecules (natural products) that serve as chemical mediators in various ecological interactions among organisms and in their relationships with the environment. For many years, natural products have been repeatedly recognized as adaptive traits that have been shaped by natural selection during evolution (e.g. Wink, 2003; Erb and Kliebenstein, 2020). Assuming this is true, the organism that produce natural products have evolved distinct or unique types of these chemicals as selective response to numerous environmental factors or natural enemies. For example, the marine environments are significant variability in conditions such as pressure (1 atm for every 10 meters depth), salinity (salinities well below or above the average of 35), temperature (ranging from Antarctic waters and cold seeps to tropical regions and thermal vents), nutrient availability (from eutrophic to oligotrophic areas) and light, such as photic to aphotic zone (Rotter et al., 2021). It is therefore essential to consider the ecological rationale - or raison d’être - for the existence of these compounds when exploring their pharmacological potential. Unlike ecological functions, therapeutic activity in humans typically results from chance interactions, as most organisms producing natural products have not co-evolved with human pathogens or disease targets.
Contrary to ecological roles of evolved chemicals, biological activities may result from chance encounters, as most organisms have never had an evolutionary relationship with human diseases. Nevertheless, some biological activities help bridge the fields of bioprospecting and chemical ecology closer together. For this reason, studies on allelopathy have been suggested as promising avenues for the discovery and development of cytotoxic substances (Singh and Thakur, 2016), since this ecological role is often expressed through biological activities, such as: modification of membrane structure and transport receptors, changes in cell morphology, disruption of the cell cycle, altered phytohormone activity, disturbed energy metabolism, impaired water balance and stomata function, inhibited pigment synthesis, and blockage of key enzymes, especially regulatory ones. Chemicals from seaweeds (Alves et al., 2018) and marine sponges (Singh and Thakur, 2016) are known to influence cellular and biochemical processes, such as ATPase inhibition, microtubule stabilization, apoptosis, angiogenesis, migration and invasion. These biological activities have been observed in both in vitro and in vivo models, with some manifesting as allelopathic effects. In Table 1, we include some examples of allelopathic substances from aquatic marine organisms and terrestrial plants, along with the biological activities they exhibit and, therefore, their defensive role against competition. More recently, a review highlighted several findings from marine chemical ecology studies offer crucial insights for the sustainable discovery of new therapeutic pharmaceuticals (Tan, 2023). While these and other aspects are essential to advancing the field, they are not the only considerations. Bioprospecting must also incorporate evolutionary principles, as we discuss here. Objectively, it can argue that natural products were not primarily selected to function as drugs for treating human diseases, despite being commonly evaluated for this purpose in bioprospecting research. Since natural products represent evolutionary and ecological responses to various environmental selective pressures, it is understandable why bioprospecting remains so costly; for example, 50,000 - 100,000 compounds may be required to obtain a single hit (Kuhlmann, 1997).
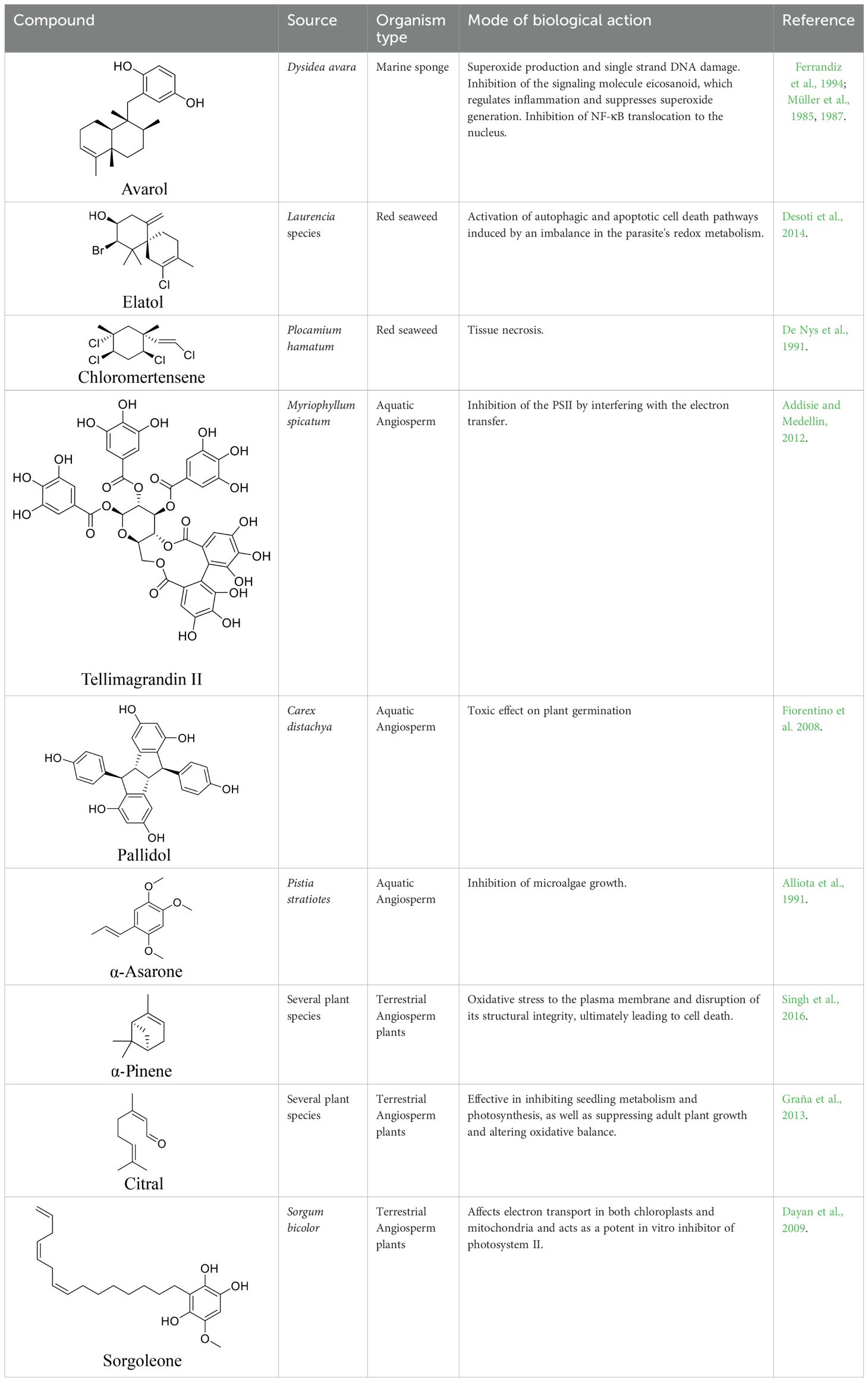
Table 1. Examples of ecological roles of allelochemicals derived from terrestrial and aquatic plants, as well as marine organisms, along with their modes of action in various biological activities.
To illustrate this point, consider the example antifouling bioprospecting, which is considerably more efficient than drug discovery. Marine organisms such as seaweeds face similar selection pressures as man-made submerged structures (e.g., piers and boats) and have evolved natural antifouling compounds do deter colonization by epibionts. These compounds protect against threats that include reduced growth (Meichssner et al., 2020), impaired reproduction (Saier and Chapman, 2004), increased drag during storms (Dixon et al., 1981), and higher susceptibility to consumers (Da Gama et al., 2008). Thus, antifouling compounds offer a clear case of evolved chemical function aligned with potential applied use, demonstrating the benefit of integrating ecological understanding into bioprospecting.
The concept of integrating bioprospecting with chemical ecology is not entirely new. For instance, Ledoux and Antunes (2018) proposed a functional workflow that prioritizes the ecological roles of marine natural products before chemical identification, in order to narrow the existing gap between the growing number of known these chemicals and the limited understanding of their ecological roles. However, what we emphasize here is the pressing need to take into account the origin and evolution of natural products in bioprospecting initiatives, given the various aspects highlighted in this discussion. Moreover, understanding the mechanisms and evolutionary pathways underlying the occurrence of natural products in nature will ultimately enable the efficient bioengineering of desirable metabolic traits in the organisms that produce them (Weng et al., 2021).
However, we are not aware of bioprospecting approaches that emphasize the fact that natural products are adaptive chemistry - phenotypic expressions of the species that produce them. Over the course of the evolution, genes encoding enzymes involved in primary metabolism have been duplicated, recruited, and diversified to perform new functions in response to the ever-changing selective pressures of the environment (Hartmann, 2007). Therefore, for these evolutionary reasons, a metabolite that exhibits an ecological role will not necessarily exhibit biological activity (such as anti-cancer or anti-viral effects); on the contrary.
5 Concluding remarks – future perspectives and limitations
Undoubtedly, bioprospecting efforts have expanded our understanding of the vast chemical diversity produced by terrestrial, marine, and aquatic organisms. However, only a small fraction of this diversity has had its ecological role elucidated. If, instead of focusing predominantly on drug discovery, bioprospecting efforts over the past decades had been directed toward understanding the ecological roles of these compounds, we might today possess deeper knowledge of how these chemicals operate at multiple levels - from cells to entire ecosystems. Such understanding could have provided critical scientific support for biodiversity conservation of across global ecosystems.
Bioprospecting has long been heralded as a dual solution for drug discovery and biodiversity conservation. However, this narrative has proven overly optimistic. In practice, the economic and ecological costs of current strategies often outweigh their benefits, and industry investment remains minimal. This disconnects stems largely from the failure to critically assess the ecological foundations of natural product diversity. Without a conceptual shift (one that recognizes natural products as evolved chemical traits rather than mere repositories of bioactivity) the full potential of biodiversity will remain underexploited and poorly protected (Firn, 2003).
Considering the reasons that separate and connect chemical ecology and bioprospecting areas, we argue that there are strong reasons for integrating them. Some key reasons include: 1) Improved cost/benefit ratio, as investigating ecological roles may lead to more positive outcomes; 2) Bridging the gap between the number of known metabolites and the understanding of their ecological roles; 3) Lowering research costs, as a single metabolite could be tested both for biological activity and ecological roles, making one organism or collection serve dual purposes and increase the meaningful results through ecological “screening”; 4) Ecological Investigation can provide libraries of chemicals (e.g., Nocchi et al., 2017) that may later be explored for pharmacological and other biological properties (e.g., Desoti et al., 2014). Thus, a workflow that integrates ecological approaches - either prior to or alongside traditional bioprospecting methods (see Figure 1) - could be both scientifically productive and mutually beneficial for the fields of bioprospecting and chemical ecology.
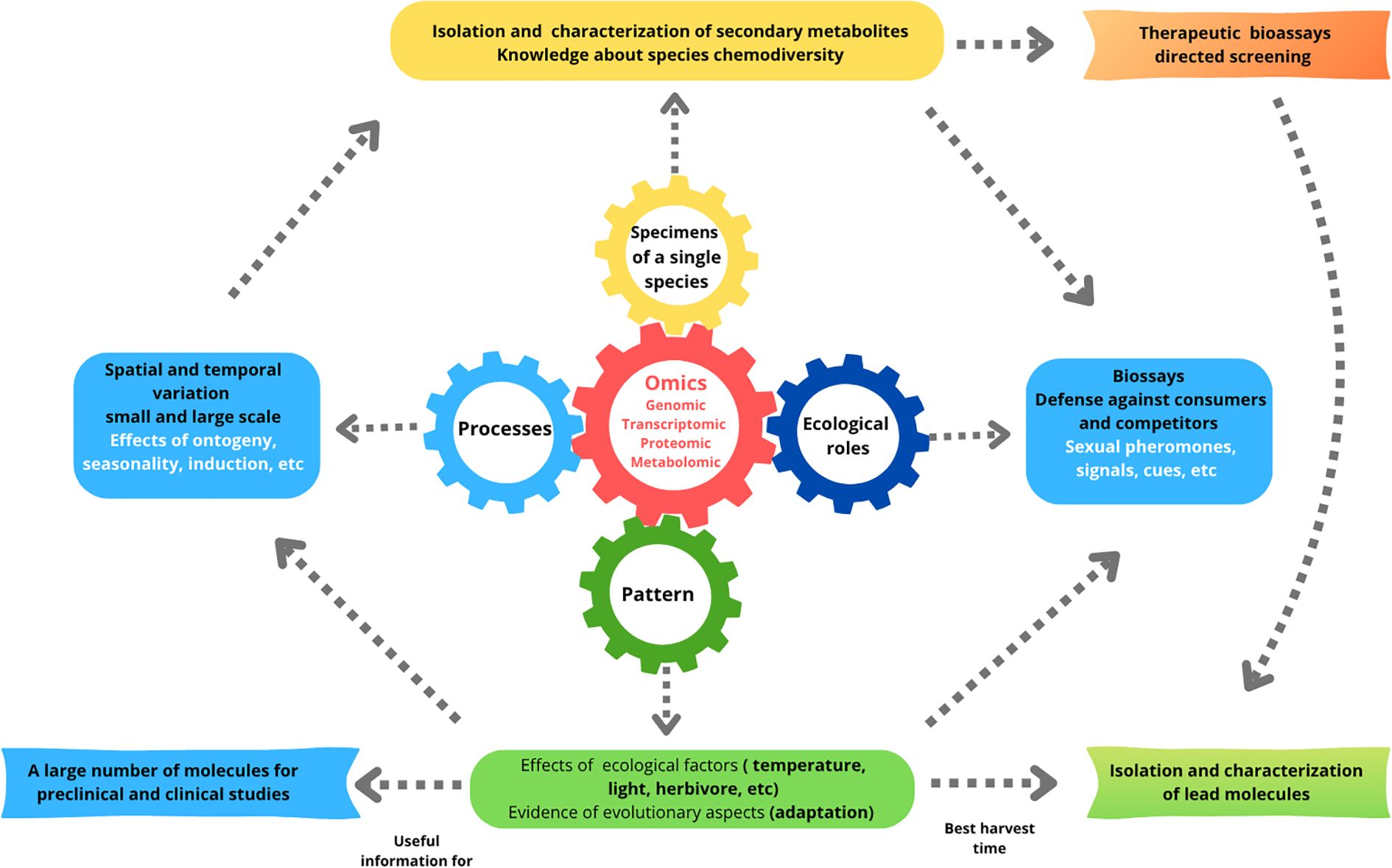
Figure 1. Ecological workflow depicting the integration of omics studies with ecological roles, processes, and patterns for the isolation and characterization of natural products. It includes sections on spatial and temporal variation, bioassays, ecological factors, and the development of therapeutic bioassays and lead molecules. Arrows connect these elements, highlighting the processes for preclinical and clinical studies. Adapted from Ledoux and Antunes, 2018.
Reflecting on the bioprospecting strategy proposed here, we suggest that screening based on the ecological roles of secondary metabolites represents a more rational and strategic alternative to conventional bioprospecting approaches. Moreover, this approach has the potential to foster sustainable development and biodiversity conservation while also supporting the economic development of biodiversity-rich countries.
While we argue that integrating eco-evolutionary reasoning into bioprospecting is a promising avenue, this approach is not without its challenges. One limitation lies in the current lack of standardized methodologies to systematically evaluate the ecological roles of natural products in situ or in complex community contexts. Furthermore, many organisms remain poorly studied from an ecological perspective, particularly in marine and microbial ecosystems, limiting our ability to formulate predictive frameworks. The development of functional ecological assays, metabolomic databases annotated with ecological metadata, and interdisciplinary collaborations will be critical to advance this field. Additionally, scaling this approach requires institutional support, funding mechanisms, and training in chemical ecology, areas that remain underrepresented in many bioprospecting programs. Nevertheless, the convergence of ecological theory, evolutionary biology, and chemical analytics offers a fertile ground for reimagining natural product discovery. Future research should focus on validating ecological screening models, developing bioinformatic tools for ecological annotation, and assessing the conservation implications of targeting ecologically functional compounds.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author/s.
Author contributions
RP: Writing – review & editing, Writing – original draft, Funding acquisition, Conceptualization. YR: Visualization, Conceptualization, Writing – review & editing. DM: Methodology, Conceptualization, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This study was partially supported by Funda̧cão de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), through the “Scientist of the State” Program for RCP and DLM, grants (Proc. E-26/201.141/2022 and E-26/201.211/2022), respectively. RCP and DLM also thank the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for their Research Productivity Fellowships (Proc. 306417/2025-1 and 306861/2023-2, respectively). YJR thanks the Federal University of Bahia for the Young Researcher Fellowship (PRPPG 010/2024).
Acknowledgments
The authors express their gratitude to their institutions (Federal Fluminense University, Federal University of Bahia, and Rio de Janeiro Botanical Garden Research Institute, Rio de Janeiro, Brazil) for providing the administrative and research support necessary to conduct this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2025.1658630/full#supplementary-material
References
Aasim M., Khawar K. M., Ahmed S. I., and Karatas M. (2019). “Multiple uses of some important aquatic and semiaquatic medicinal plants,” in Plant and Human Health, vol. 2 . Eds. Ozturk M. and Hakeem K., 541–577, Cham, Switzerland: Springer. doi: 10.1007/978-3-030-03344-6_23
Addisie Y. and Medellin A. C. (2012). Allelopathy in aquatic macrophytes: Effects on growth and physiology of phytoplanktons. Afr. J. Plant Sci. 6, 270–276. doi: 10.5897/AJPS12.008
Aliotta G., Monaco P., Pinto G., Pollio A., and Pevitera L. (1991). Potential allelochemicals from Pistia stratiotes L. J. Chem. Ecol. 17, 2223–2234. doi: 10.1007/BF00988003
Alves C., Silva J., Pinteus S., Gaspar H., Alpoim M. C., Botana L. M., et al. (2018). From marine origin to therapeutics: The antitumor potential of marine algae-derived compounds. Front. Pharmacol. 9. doi: 10.3389/fphar.2018.00777
Arya A. K., Durgapal M., Bachheti A., Deepti, Joshi K. K., Gonfa Y. H., et al. (2022). Ethnomedicinal use, phytochemistry, and other potential application of aquatic and semiaquatic medicinal plants. eCam 2022, 4931556. doi: 10.1155/2022/4931556
Cachet N., Genta-Jouve G., Ivanisevic J., Chevaldonne P., Sinniger F., Culioli G., et al. (2015). Metabolomic profiling reveals deep chemical divergence between two morphotypes of the zoanthid Parazoanthus axinellae. Sci. Rep. 5, 8282. doi: 10.1080/08927010802041253
Da Gama B. A. P., Santos R. P. A., and Pereira R. C. (2008). The effect of epibionts on the susceptibility of the red seaweed Cryptonemia seminervis to herbivory and fouling. Biofouling 24, 209–218. doi: 10.1080/08927010802041253
Dayan F. E., Howell J. L., and Weidenhamer J. D. (2009). Dynamic root exudation of sorgoleone and its in planta mechanism of action. J. Exp. Bot. 60, 2107–2117. doi: 10.1093/jxb/erp082
De Nys R., Coll J. C., and Price I. R. (1991). Chemically mediated interactions between the red alga Plocamium hamatum (Rhodophyta) and the octocoral Sinularia cruciata (Alcyonacea). Mar. Biol. 108, 315–320. doi: 10.1007/bf01344346
Desoti V. C., Lazarin-Bidóia D., Sudatti D. B., Pereira R. C., Ueda-Nakamura T., Nakamura C. T., et al. (2014). Additional evidence of the trypanocidal action of (-)-elatol on amastigote forms through the involvement of reactive oxygen species. Mar. Drugs 12, 4973–4983. doi: 10.3390/md12094973
DiMasi J. A. (2001). New drug development in the United States from 1963 to 1999. Clin. Pharmacol. Ther. 69, 286–296. doi: 10.1067/mcp.2001.115132
DiMasi J. A., Hansen R. W., and Grabowski H. G. (2003). The price of innovation: New estimates of drug development costs. J. Health Econ. 22, 151–185. doi: 10.1016/S0167-6296(02)00126-1
Dixon J., Schroeter S. C., and Kastendiek J. (1981). Effects of the encrusting bryozoan, Membranipora membranacea, on the loss of blades and fronds by the giant kelp, Macrocystis pyrifera (Laminariales). J. Phycol. 17, 341–345. doi: 10.1111/j.1529-8817.1981.tb00860.x
Dixon R. A. and Dickinson A. J. (2024). A century of studying plant secondary metabolism—From “what?” to “where, how, and why?”. Plant Physiol. 195, 48–66. doi: 10.1093/plphys/kiad596
Erb M. and Kliebenstein. D. J. (2020). Plant secondary metabolites as defenses, regulators, and primary metabolites: The blurred functional trichotomy. Plant Physiol. 184, 39–52. doi: 10.1104/pp.20.00433
Ferrandiz M. L., Sanz M. J., Bustos G., Paya M., Alcaraz M. J., and De Rosa S. (1994). Avarol and avarone, two new anti-inflammatory agents of marine origin. Eur. J. Pharmacol. 253, 75e82. doi: 10.1016/0014-2999(94)90759-5
Fiorentino A., D’Abrosca B., Pacifico S., Izzo A., Letizia M., Esposito A., et al. (2008). Potential allelopatic effects of stilbenoids and flavonoids from leaves of Carex distachya Desf. Biochem. Syst. Ecol. 36, 691–698. doi: 10.1016/j.bse.2008.07.002
Firn R. D. (2003). Biopropsecting – Why is it so unrewarding? Biodivers. Conserv. 12, 207–216. doi: 10.1023/A:1021928209813
Firn R. D. and Jones C. G. (2003). Natural products – a simple model to explain Chemical diversity. Nat. Prod. Rep. 20, 382–391. doi: 10.1039/B208815K
Graña E., Sotelo T., Diíaz-Tielas C., Reigosa M. J., and Sanchez-Moreiras A. M. (2013). The phytotoxic potential of the terpenoid citral on seedlings and adult plants. Weed Sci. 61, 469e481. doi: 10.1016/j.phytochem.2007.09.017
Hartmann T. (2007). From waste products to ecochemicals: Fifty years research of plant secondary metabolism. Phytochemistry 68, 2831–2846. doi: 10.1016/j.phytochem.2007.09.017
Hunt B. and Vicent A. C. J. (2006). Scale and sustainability of marine bioprospecting for pharmaceuticals. Ambio 35, 57–64. doi: 10.1002/ptr.1506
Jia W., Gao W. E.-Y., Yan Y.-Q., Wang J., Xu Z.-H., Zheng W.-J., et al (2004). The rediscovery of ancient Chinese herbal formulas. Phytother. Res. 18, 681–686. doi: 10.1002/ptr.1506
Jones C. G. and Firn R. D. (1991). On the evolution of secondary plant chemical diversity. Philos. Trans. R. Soc Lond. B 333, 273–280. doi: 10.1098/rstb.1991.0077
Kuhlmann J. (1997). Drug research: From the idea to the product. Int. J. Clin. Pharmacol. Ther. 35, 541–552.
Labes A. (2023). Marine resources offer new compounds and strategies for the treatment of skin and soft tissue infections. Mar. Drugs 21, 387. doi: 10.1080/07388551.2017.1331335
Ledoux J.-P. and Antunes A. (2018). Beyond the beaten path: Improving natural products bioprospecting using an eco-evolutionary framework – The case of the octocorals. Chem. Rev. Biotechnol. 38, 184–198. doi: 10.1080/07388551.2017.1331335
Liu M., El-Hossary E. M., Oelschlaeger T. A., Donia M. S., Quinn R. J., and Abdelmohsen U. R. (2019). Potential of marine natural products against drug resistant bacterial infections. Lancet Infect. Dis. 7, E237–E245. doi: 10.1016/S1473-3099(18)30711-4
Müller W. E., Maidhof A., Zahn R. K., Schröeder H. C., Gasic M. J., Heidemann D., et al. (1985). Potent antileukemic activity of the novel cytostatic agent avarone and its analogues in vitro and in vivo. Cancer Res. 45, 4822e4826.
Müller W. E., Sladic D., Zahn R. K., Bässler K.-H., Dogovic N. H., Gerner M. J., et al. (1987). Avarol-induced DNA strand breakage in vitro and in Friend erythroleukemia cells. Cancer Res. 47, 6565–6571.
Meichssner A., Stegmann N., Cosin A. -S., Sachs D., Bressan M., Marx H., et al. (2020). Control of fouling in the aquaculture of Fucus vesiculosus and Fucus serratus by regular desiccation. J. Appl. Phycol. 32, 4145–4158. doi: 10.1007/s10811-020-02274-2
Moher D., Liberati A., Tetzlaff J., Altman D. G., and The PRISMA Group (2009). Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. Brit. Med. J. 339, 332–336. doi: 10.1371/journal.pmed.1000097
Nocchi N., Soares A. R., Souto M. L., Fernández J. J., Martin M. N., and Pereira R. C. (2017). Detection of a chemical cue from the host seaweed Laurencia dendroidea by the associated mollusk Aplysia brasiliana. PLoS One 12, e0187126. doi: 10.1016/j.bioteChadv.2021.107871
Papon N., Copp B. C., and Courdavault V. V. (2022). Marine drugs: Biology, pipelines, current and future prospects for production. Biotechnol. Adv. 54, 107871. doi: 10.1016/j.biotechadv.2021.107871
Pereira R. C. and Costa-Lotufo L. V. (2012). Bioprospecting for bioactives from seaweeds: potential, obstacles and alternatives. Rev. Bras. Farmacog. 22, 894–950. doi: 10.1590/S0102-695X2012005000077
Proksch P., Edrada-Ebel R., and Ebel R. (2003). Drugs from the sea - opportunities and obstacles. Mar. Drugs 1, 5–17. doi: 10.3390/md101005
Raina S.-B., Lambert B. S., Parks D. H., Rinke C., Siboni N., Bramucci A., et al. (2022). Chemotaxis shapes the microscale organization of the ocean’s microbiome. Nature 605, 132–138. doi: 10.1038/s41586-022-04614-3
Rojo F. P., Pillow J. J., and Kaur P. (2023). Bioprospecting microbes and enzymes for the production of pterocarpans and coumestans. Front. Bioeng. Biotechnol. 11. doi: 10.3389/fbioe.2023.1154779
Rotter A., Bacu A., Barbier M., Bertoni F., Bones A. M., Cancela M. L., et al. (2020). A new network for the advancement of marine biotechnology in Europe and beyond. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.00278
Rotter A., Barbier M., Bertoni F., Bones A. M., Cancela M. L., Carlsson J., et al. (2021). The essentials of marine biotechnology. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.629629
Saier B. and Chapman A. S. (2004). Crusts of the alien bryozoan Membranipora membranacea can negatively impact spore output from native kelps (Laminaria longicruris). Bot. Mar. 47, 265–271. doi: 10.1515/BOT.2004.031
Silveria D. and Boylan F. (2023). Medicinal plants: Advances in phytochemistry and ethnobotany. Plants 12, 1682. doi: 10.3390/plants12081682
Singh H. P., Batish D. R., Kaur S., Arora K., and Kohli R. K. (2006). Alpha-Pinene inhibits growth and induces oxidative stress in roots. Ann. Bot. 98, 1261e1269. doi: 10.1093/aob/mcl213
Singh A. and Thakur N. L. (2016). Significance of investigating allelopathic interactions of marine organisms in the discovery and development of cytotoxic compounds. Chem. Biol. Interact. 243, 135–147. doi: 10.1016/j.cbi.2015.09.009
Singh N., Vayer P., Tanwar S., Poyet J.-L., Tsaioun K., and Villoutreix B. O. (2023). Drug discovery and development: introduction to the general public and patient groups. Front. Drug Discov. 3, 1201419. doi: 10.3390/md21030174
Tan L. T. (2023). Impact of marine chemical ecology research on the discovery and development of new pharmaceuticals. Mar. Drugs 21, 174. doi: 10.3390/md21030174
Weng J.-K., Hynch I. J., Matos J. O., and Dudareva N. (2021). Adaptive mechanisms of plant specialized metabolism connecting chemistry to function. Nat. Chem. Biol. 17, 1037–1045. doi: 10.1038/s41589-021-00822-6
Keywords: secondary metabolites, bioprospection, chemical diversity, adaptive chemistry, biodiversity conservation
Citation: Pereira RC, Ramos YJ and Moreira DL (2025) The eco-evolutionary raison d’être of natural products as support for sustainable bioprospecting. Front. Ecol. Evol. 13:1658630. doi: 10.3389/fevo.2025.1658630
Received: 02 July 2025; Accepted: 29 September 2025;
Published: 07 November 2025.
Edited by:
Islam S. Sobhy, Cardiff University, United KingdomReviewed by:
Mohd Kamil Hussain, Govt. Raza Post Graduate College, Rampur, IndiaTao Liu, Guangdong Pharmaceutical University, China
Yehezkiel Steven Kurniawan, Universitas Gadjah Mada, Indonesia
Copyright © 2025 Pereira, Ramos and Moreira. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Renato Crespo Pereira, cmNyZXNwb0BpZC51ZmYuYnI=