 Beatriz Modenutti
Beatriz Modenutti Evelyn Vega
Evelyn Vega Marcela Bastidas Navarro
Marcela Bastidas Navarro Nicolás Martyniuk
Nicolás Martyniuk Esteban Balseiro
Esteban Balseiro- Laboratorio de Limnología, Instituto de Investigaciones en Biodiversidad y Medioambiente (INIBIOMA), Universidad Nacional del Comahue-Consejo Nacional de Investigaciones Científicas y técnicas (CONICET), Bariloche, Argentina
The cryosphere is facing unprecedented changes due to global change, and one of its main consequences is on the connectivity of ice-snow and downstream habitats in the glacial and periglacial landscape. Here, we analyze the eukaryotic community composition and the ecological processes structuring eukaryotic communities in the glacial environment of Mount Tronador (North-Patagonia, Argentina). The study was carried out in Castaño Overo and Alerce glaciers, including three different compartments: glacial ice, snow, and ponds in the glacier forefield. All compartments showed low dissolved and particulate nutrient concentrations. Genomic DNA analyses revealed a high eukaryotic microorganism’s diversity, and a high proportion of shared taxa (>35%), resulting in low presence-absence ß-diversity. However, abundance-based ß-diversity is higher among compartments, underscoring that changes in abundances are significant regardless of the large number of shared ASVs. Ice and snow showed a predominance of Fungi, while Chlorophyta increased its importance in ponds. Ciliophora and Dinoflagellates were almost exclusively present in ice and ponds, respectively. The co-occurrence network analysis showed a higher number of modules and modularity in ice and snow relative to ponds. The presence of modules with algae and fungi (Pucciomycotina and Chytridiomycotina) would reflect a functional association due to the nutritional dependence of fungi on the algae. More connectors between modules were detected in ice and snow (5 and 6) relative to ponds (only 1). Most of these connectors were fungi that could also indicate their dependence on more than one species association (module). Overall, our results showed that as ice and snow melt due to global change, eukaryotes will undergo community reorganization that will affect downstream environments.
Introduction
Microeukaryotes in glacial and periglacial systems are a fascinating topic because these ecosystems, though extreme, host a variety of microbial life that has adapted to harsh conditions (Anesio et al., 2017). The eukaryotic organisms found in glaciers and ice-covered environments primarily include protists, fungi, and some algae, many of which are adapted to survive in low temperatures, nutrient-poor conditions, and high levels of ultraviolet (UV) radiation (Anesio and Laybourn-Parry, 2012).
In mountain regions, changes in near-surface temperature and in precipitation patterns lead to a decrease in snow accumulation, earlier spring melt, and prolonged summer ice melt (Fell et al., 2017). Consequently, these changes in the distribution, thickness, and permanence of ice sheets result in the retreat and thinning of many mountain glaciers (Zemp et al., 2015). For the year 2100, the estimated loss in glacier area is projected to range from 22 to 51%, depending on the climate scenario considered (Bosson et al., 2023). The retreat and imminent loss of glaciers have physical, chemical, and biological implications for the aquatic ecosystems receiving their meltwater (Vanderwall et al., 2024). In addition, the physical processes associated with glacial retreat and meltwater runoff erode the rock, modeling a new landscape that includes the formation of lakes and a still largely unexplored periglacial environment.
The periglacial environment is defined as that in which frost action and/or permafrost related processes dominate (French and Thorn, 2006). In addition, proglacial environments are located immediately in front of glaciers, ice caps, and ice sheets (Slaymaker, 2011). However, the criteria of ice-marginal location and intensive freeze-thaw and/or permafrost related processes are not exclusive of each other, and proglacial and periglacial environments commonly overlap (Slaymaker, 2011). Frozen water is an important erosive agent in these environments, as ice pressure can break the rock, allowing the formation of ponds (Martini et al., 2011). In particular, during the warm season, substantial melting of glaciers and snow contributes with water, nutrients, and microbial communities to downstream environments, where they play a crucial role in biogeochemical cycling (Anesio and Laybourn-Parry, 2012). These cold ecosystems are especially sensitive to climate change; thus, understanding the role of microbial life in these habitats is crucial in scenarios of glacier retreat and permafrost thawing (Margesin and Collins, 2019; Winkel et al., 2022).
Mountain peri/proglacial regions are changing rapidly due to loss of snow and ice, and this will produce important environmental changes, including physical parameters (temperature and light), chemical (nutrients), and biological (element stoichiometry, microorganisms’ diversity) (Ren et al., 2019; Elser et al., 2020; Vanderwall et al., 2024). In addition to ice melting, snow melt can also influence lake nutrient inputs (Vincent et al., 2011) and snow microbiome (Krug et al., 2020; Yakimovich et al., 2020) can remain in the active fraction of downstream aquatic environments (Comte et al., 2018). Thus, these cold environments represent an interesting spatial pattern of ice-snow-ponds of hydrological interaction, where microorganisms colonize the different water states.
Eukaryotic communities in glacial and peri/proglacial environments are dominated by heterotrophic microorganisms such as fungi and protozoa (flagellates and ciliates), and by autotrophs with algae such as Chlorophyceae, Zygnematophyceae, Dinophyceae, and Bacillariophyceae (Anesio and Laybourn-Parry, 2012). Microorganisms in glacial habitats exhibit metabolic adaptations such as the production of cold-active enzymes, antifreeze proteins, and exopolymeric substances that protect against low temperatures (Margesin and Miteva, 2011). These communities remain biogeochemically active, contributing to processes like carbon fixation, iron cycling, and methanogenesis, with implications at both local and global scales (Hood et al., 2009). Most studies on eukaryotic diversity in these cold environments have been carried out in the Arctic and Antarctic (Laybourn-Parry and Pearce, 2007; Cameron et al., 2015; Comte et al., 2018), in snow packs (Brown and Jumpponen, 2019; Vimercati et al., 2019), and in high altitude lakes (Triadó-Margarit and Casamayor, 2012; Kammerlander et al., 2015; Filker et al., 2016). Despite its ecological importance in nutrients and elements cycling (Anesio and Laybourn-Parry, 2012), our knowledge about eukaryotic communities from mountain glacial and periglacial environments is still scarce, and this is of particular importance since, as glaciers continue to recede, these communities will be gradually more vulnerable and lost (Vimercati et al., 2019). Studies of the connectivity and biogeography of the snow microbiome (Comte et al., 2018; Brown and Jumpponen, 2019) and biogeographic analyses of glacial lakes (Filker et al., 2016; Schiaffino et al., 2020) have indicated differences in community structure among regions, but little attention has been given to spatial patterns within connected hydrologic systems. Solid water environments such as glacial ice and snow packs can promote spatially structured microbial microhabitats and increase ecological complexity compared to the more homogeneous and diluted conditions typically found in meltwater ponds (Bachy et al., 2011; Anesio et al., 2017).
This study aims to analyze the eukaryotic communities of a glacial and periglacial environment of North Patagonian Andes. Glacial ice and snowpacks can sustain distinct microbial microhabitats (Bachy et al., 2011), whereas meltwater typically results in more diluted and mixed conditions. Based on these contrasting physicochemical settings, we hypothesize that the eukaryotic community varies among these hydrologically connected environments according to water state and physical heterogeneity. Specifically, we predict a more complex community structure in solid water (ice and snow) than in meltwater ponds.
To evaluate this, we analyzed eukaryotic community dissimilarities, diversity indices, diversity components, and performed a network analysis in the ice-snow-ponds hydrological system. The present study was carried out in Mount Tronador, the most important ice cap in the North Patagonian Andes, with different glaciers with radial flow (Ruiz et al., 2015). The retreat of Tronador glaciers has been well documented over recent decades (Ruiz et al., 2017). Samples were collected from ice and snow of the Alerce and Castaño Overo glaciers, and from ponds located in the glacier forefield.
Materials and methods
Study site and sampling
Mount Tronador is an inactive stratovolcano located in the North Patagonian Andes (41.15°S, 71.88°W), and has a height of 3478 m (Figure 1). The mountain straddles the border between Chile and Argentina. The upper slopes of Mount Tronador host the accumulation zones of 13 major glaciers, 5 of which are located in Argentina (Zorzut et al., 2020). In the last decades, the equilibrium line altitude changed due to increasing warming trends (Ruiz et al., 2017; Ghilardi Truffa and Ruiz, 2024). Thus, its glaciers have exhibited a continuous retreat trend (Ruiz et al., 2017; Masiokas et al., 2020). This retreat leaves new landscapes, modeling the existing ones and generating new aquatic environments, such as lakes and periglacial ponds (Ruiz et al., 2017; Modenutti et al., 2018, this study).
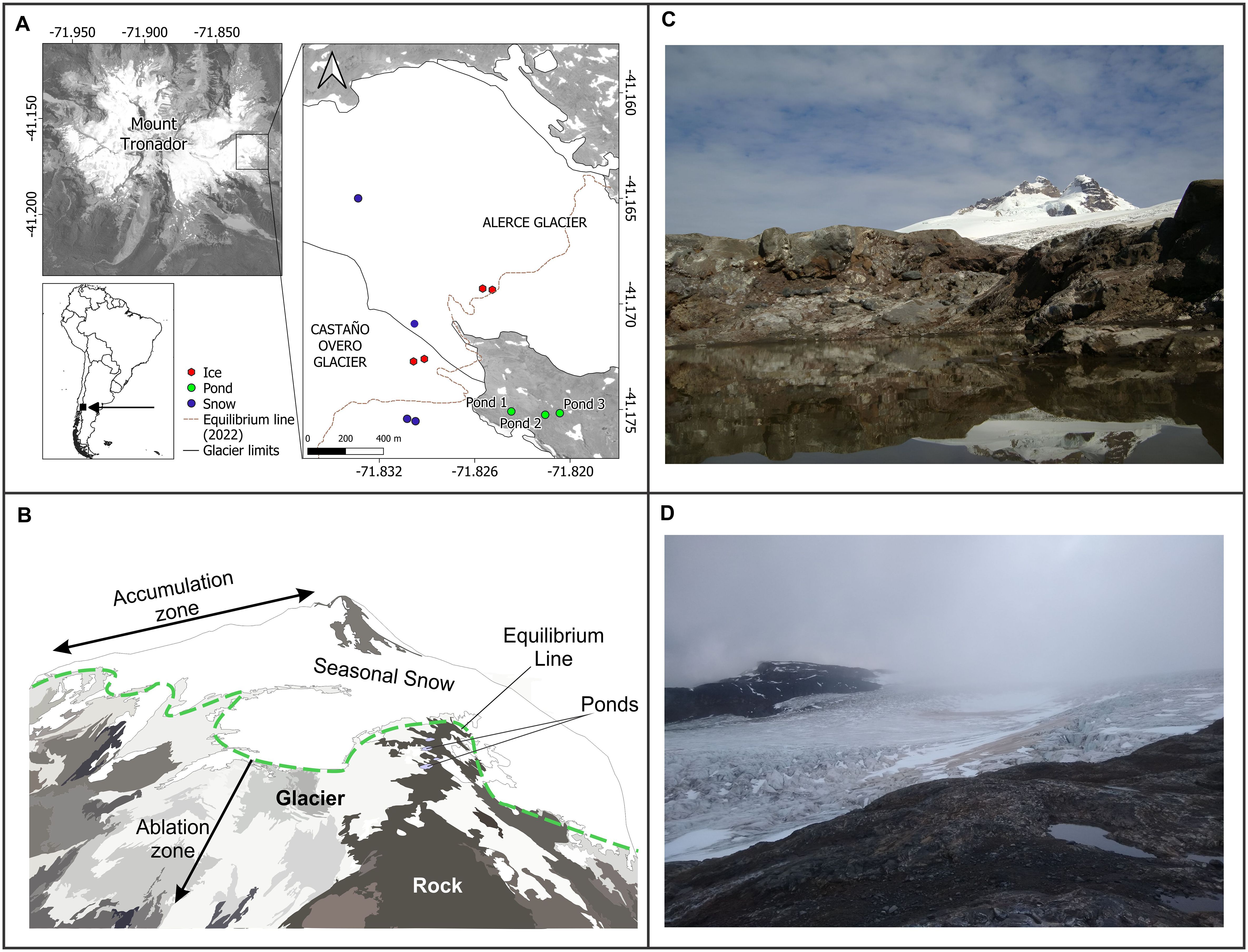
Figure 1. Map of the studied area. Sampling sites in the three glacial and periglacial environments (Glacial Ice, Snow and Ponds) are indicated with dots. (A) Map, (B) Scheme of the Tronador Mountain, (C) Photographs of Pond 2, and (D): Glacier Castaño Overa and Pond 1.
The study was carried out in Alerce and Castaño Overo glaciers (Argentina), where samples of glacial ice and snow were obtained. In addition, ponds in the glacier forefield at circa 1900 m a.s.l. were also sampled (Figure 1A). Sampling was carried out in summer 2021-2022. An ice drill (IDDO Hand Auger Kit) was used to obtain the ice core samples. This equipment is specially designed for boring and extraction in remote, high mountain locations. We sampled two ice sites, one in Alerce and the other in Castaño Overo glaciers (Figure 1A). At each sampling site, two cores (90 cm long) were obtained, and the upper 5 cm were discarded to avoid contamination from the equipment head. The remaining core sections were carried to the laboratory in sterile bags, divided into two portions. Considering the different sections and replicates, we analyzed 8 ice samples.
The snow samples were collected in four sampling sites (2 samples per site) located on both mentioned glaciers (Figure 1A). At each site, an area of 1 m2 of virgin snow cover was delimited, and the surface layer (10 cm) was collected in Nalgene® bottles with sterile gloves. Considering all sites, we analyzed 8 snow samples.
Finally, we obtained water samples from 3 ponds located in the glacier forefield (Figure 1A). These ponds resulted from proglacial processes that occurred around and within the margins of the former glacier and produced the formation of ponds scoured into bedrock (Figures 1B–D). Surface water samples (two samples per pond) were taken using sterile 1L Nalgene® containers. Thus, a total of 6 water samples were analyzed.
All samples were transported in thermally insulated containers until they arrived at the laboratory, where they were stored at -80°C until processing.
Laboratory procedures for chemical parameters
Sample processing consisted of filtering the water from the different sources (glacial ice, snow, and ponds). To determine the concentrations of total dissolved phosphorus (TDP), total dissolved nitrogen (TDN), and dissolved organic carbon (DOC) we filtered onto pre-combusted GF/F filters. The TDP samples were digested with potassium persulfate at 125 °C and 1.5 atm, and then the concentration was determined using the ascorbic acid reduction method with molybdenum (APHA, 2005). DOC was measured in 50 mL of sample using a high-combustion analyzer (Shimadzu TOC-VCSH) and TDN was determined using the TNM-1 unit on the Shimadzu TOC-VCSH.
In addition, unfiltered water samples (ice, snow, and ponds) were used for total phosphorus (TP) and total suspended solids (TSS) concentration determinations. TP was determined following APHA (2005) as explained for TDP, except for the filtering. TSS was quantified by filtration of 200 mL of unfiltered water onto GF/F filters previously combusted and weighed. Then, filters were dried for at least 48 h at 60°C and then reweighed. Particulate C and particulate N were measured on precombusted GF/F filters (200 mL) and processed in a High Temperature Combustion elemental analyzer (Thermo Finnigan EA1112).
DNA extraction, and amplicon sequencing
Water samples (250–300 mL) from different sources (ice, snow, and ponds) were filtered through 0.22 μm pore-size nucleopore filters (PALL®) for DNA extraction. The filters were frozen at -80 °C until DNA extraction.
Genomic DNA was extracted using Qiagen DNeasy® PowerSoil®Kit (Qiagen, Hilden, Germany), following the manufacturer’s protocols. The quantity and purity of DNA samples were determined with a Take3 Micro-Volume Plate (BioTek® Instruments, Inc., Winooski, VT, USA) in a Synergy™ HTX Multi-Mode Microplate Reader. The extracted DNA was stored at −80°C for further processing. The extracted DNA was sent to Novogene Bioinformatics Technology Co. (Beijing, China) to be processed using Illumina, the sequencing platform and strategy used was NovaSeq 6000 PE250.
To determine the eukaryotic community diversity, we used the tagged amplicons of the V4 region of 18S rRNA eukaryal gene obtained with the primers 528F (5′-GCGGTAATTCCAGCTCCAA-3′) and 706R (5′-AATCCRAGAATTTCACCTCT-3′) 350bp (Cheung et al., 2010). These primers amplify a specific hypervariable region of the eukaryotic 18S ribosomal RNA (rRNA) gene, which is widely used as a genetic marker for eukaryotic microorganisms. The 18S gene contains conserved regions (for primer binding) flanking variable regions that provide taxonomic resolution (Pawlowski et al., 2012). Amplification, library preparation, sequencing and demultiplexing were conducted by Novogene Bioinformatics Technology Co. The PCR conditions informed by Novogene Bioinformatics Technology Co. were: an initial denaturation of 3 min at 94°C, followed by 30 cycles of 30 s at 94°C, 30 s at 60°C, 1 min at 72°C, and a final cycle of 5 min at 72°C.
Processing of illumina sequencing data
Amplicon sequences provided by Novogene Bioinformatics Technology Co. were processed using DADA2 v1.22.0. The entire sequencing project was analyzed with the same filtering parameters following Callahan et al. (2016). We used the filterAndTrim function from DADA2 with the following quality values: maxEE = c(2,2) and truncLen = c(220,200). The chimera sequences were excluded using the function removeBimeraDenovo. Sequences were identified as unique amplicon sequence variants (ASV) based on single nucleotide differences. The taxonomic classification was performed using the PR2 database (version 2.0.2) as a reference. A second table was generated by discarding ASVs with fewer than 20 reads. Multicellular organisms were also eliminated to avoid bias due to multiple copies of the 18S rRNA gene, when they were amplified from the same organism. This last table was normalized to a sampling depth using the rarefy function of the vegan package (Oksanen et al., 2013). To have a lower taxonomic level, the taxonomic search for some ASVs was carried out on the National Center for Biotechnology Information (NCBI) website, introducing the sequence corresponding to each ASV in the tool “blasttn.” The sequencing data have been deposited in NCBI BioProject PRJNA1270387 with the Biosample accessions SAMN48824426-47.
Data analysis
Nutrients were compared among systems (Glacial ice, snow, and ponds) with a one-way ANOVA or Kruskal-Wallis test when normality or homoscedasticity were not fulfilled.
We constructed rarefaction curves to analyze the depth of sequencing and evaluate whether the richness of ASVs of each sample was adequate (Supplementary Figure S1). The obtained rarefaction matrix of ASVs was based on an iterative process to bring all samples to the smallest number of readings. These ASVs were used as taxonomic identities (see Processing of Illumina sequencing data) for the calculations of diversity indices, and network analysis.
To test our hypothesis, we calculated diversity indices, community dissimilarities, and performed network analysis as follows. We calculated alpha diversity indices (Shannon-Wiener index), ASV richness, and Pielou’s evenness index. Eukaryotic community dissimilarities were visualized using dendrograms and non-metric multidimensional scaling (NMDS) analyses based on Bray-Curtis distance with the total ASVs in 22 samples, followed by an analysis of similarities (ANOSIM) to determine if there were significant differences among the groups generated by the NMDS analysis. All the above statistical analyses and calculations were performed in R (v. 4.4.3) (R Core Team, 2025) with vegan v. 2.6.1. Heatmaps were constructed using pheatmap v.1.0.12, while Venn diagrams were created using the VennDiagram package v.1.7.3.
Total eukaryotic ß-diversity was calculated using the Sørensen dissimilarity index (ßSOR) and total Bray–Curtis dissimilarity index (βBC), applied to presence–absence and abundance data, respectively. Then, we applied the Baselga ß diversity partitioning (Baselga and Orme, 2012; Baselga and Chao, 2017) by calculating the contributions of turnover (ßSIM; Simpson dissimilarity index) and nestedness (ßNES) to the presence/absence dataset. In addition, we analyzed the contributions of balanced-variation (ßBC.BAL) and abundance-gradient (ßBC.GRA) components to the abundance dataset. The dissimilarity indices of eukaryotic communities were calculated in R, using the function ‘beta.pair’ and ‘beta.pair.abund’ in the R package ‘betapart’ (v1.6). Total eukaryotic ß-diversity and its partitioning were analyzed by comparing the systems: a) glacial ice-snow, b) snow-ponds, and c) glacial ice-ponds.
We generated null models of eukaryotic communities in the periglacial environments for both presence-absence and abundance data. In the former, we applied the functions ‘nullmodel’ and ‘simulate’ of the package ‘vegan’, with a non-sequential algorithm for binary matrices that preserves the site (row) frequencies (option ‘r1’). In the abundance null models, we used the function ‘taxo.null’ of the package ‘NST’ (v3.1.10), with the abundance option ‘region’. In both cases, we generated 1000 null models, we estimated the distance matrices (ß-diversity) and then we calculated the average distance matrix for each kind of index. Then, we compared the ß diversity of the observed datasets (glacial ice, snow, and ponds) with those of the corresponding null model with a t-test.
Network analysis was conducted for each environment (glacial ice, snow, and ponds) to determine the co-occurrence patterns among the eukaryotic taxa and to identify key ASVs or significant module members in eukaryotic communities (Guimera and Amaral, 2005). A module is a group of nodes (ASVs) with high interactions among them and low interactions with other nodes outside the module; while modularity measures how well a network separates into different modules (Deng et al., 2012). Modularity may indicate habitat heterogeneity and an increase in modularity seemed to reflect more intensive functional associations (Zhou et al., 2021). Analysis was performed using the Igraph package (v2.0.3) and MetaNet (v0.1.2) in R (v.4.4.3). A similarity matrix was constructed based on Pearson correlation coefficient between pairwise ASVs. The edge list was constructed based on the similarity matrix considering only absolute values > 0.8. The different roles of each node in the network were identified using the values that measure within-module connectivity (Zi) and the values that measure among-module connectivity (Pi). Then, we develop Zi–Pi diagrams`, which can be used to study key species in a community (Zhou et al., 2011; Deng et al., 2012). Co-occurrence graphs were constructed with Cytoscape v3.10.3 (Shannon et al., 2003).
Results
Chemical features of the glacial and periglacial environments
Dissolved nutrient concentrations (TDP, TDN and DOC) were significantly different among the glacial environments (Table 1). Glacial ice showed the highest DOC concentration and TDN: TDP ratio, while snow exhibited the highest TDN and TDP concentrations but the lowest TDN: TDP ratio. The ponds showed a very low DOC concentration (Table 1).
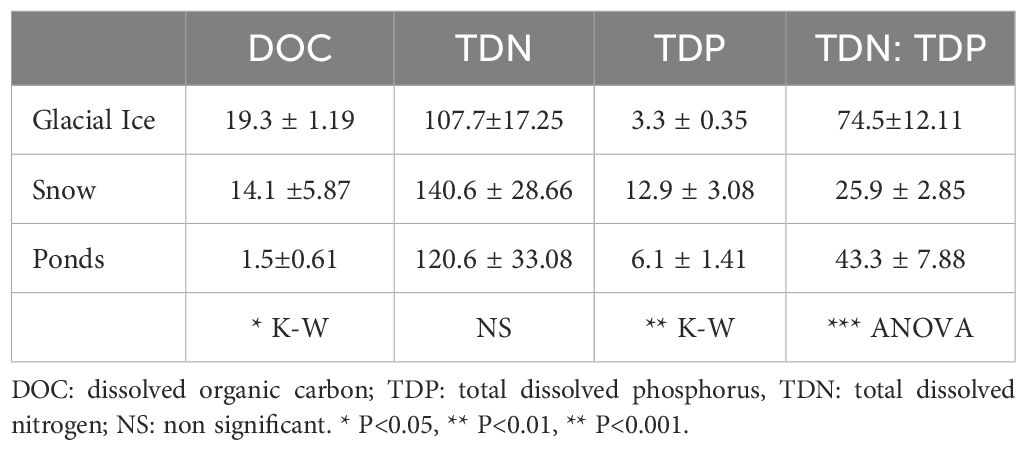
Table 1. Dissolved nutrients in the three glacial and periglacial environments (glacial ice, snow and ponds) and results of ANOVA and Kruskall Wallis post-hoc analysis.
The particulate fraction showed a different pattern. C concentration was highest in snow and lowest in glacial ice, similar to particulate N and P, but in the latter, the lowest values were observed in the ponds (Table 2). Elemental ratios showed only significant differences in N:P, with the lowest values in glacial ice (Table 2).
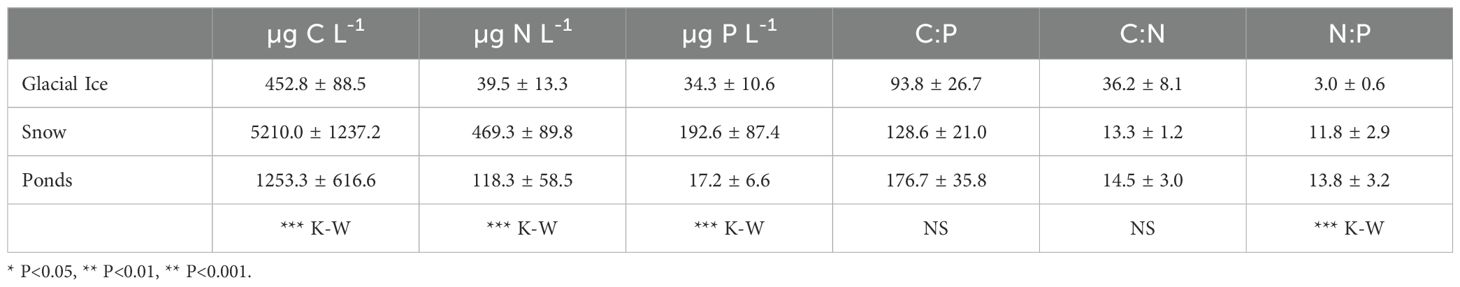
Table 2. Particulate carbon (C), phosphorus (P) and nitrogen (N) and elemental ratios in the three glacial and periglacial environments (glacial ice, snow and ponds) and results of Kruskall Wallis (K-W) post-hoc analysis.
Taxonomic composition of the eukaryotic community in glacial and periglacial environments
After the quality control and rarefaction, we obtained a total of 759550 reads in 1363 ASVs. The eukaryotic communities are dominated by Chlorophyta, fungi, Dinoflagellata, and Ciliophora with different relative abundances in the three systems (Figures 2A, B). Ice and snow showed a predominance of fungi, while in the ponds, Chlorophyta increased its importance. In addition, Ciliophora and Dinoflagellates were present almost exclusively in the ice and the ponds, respectively.
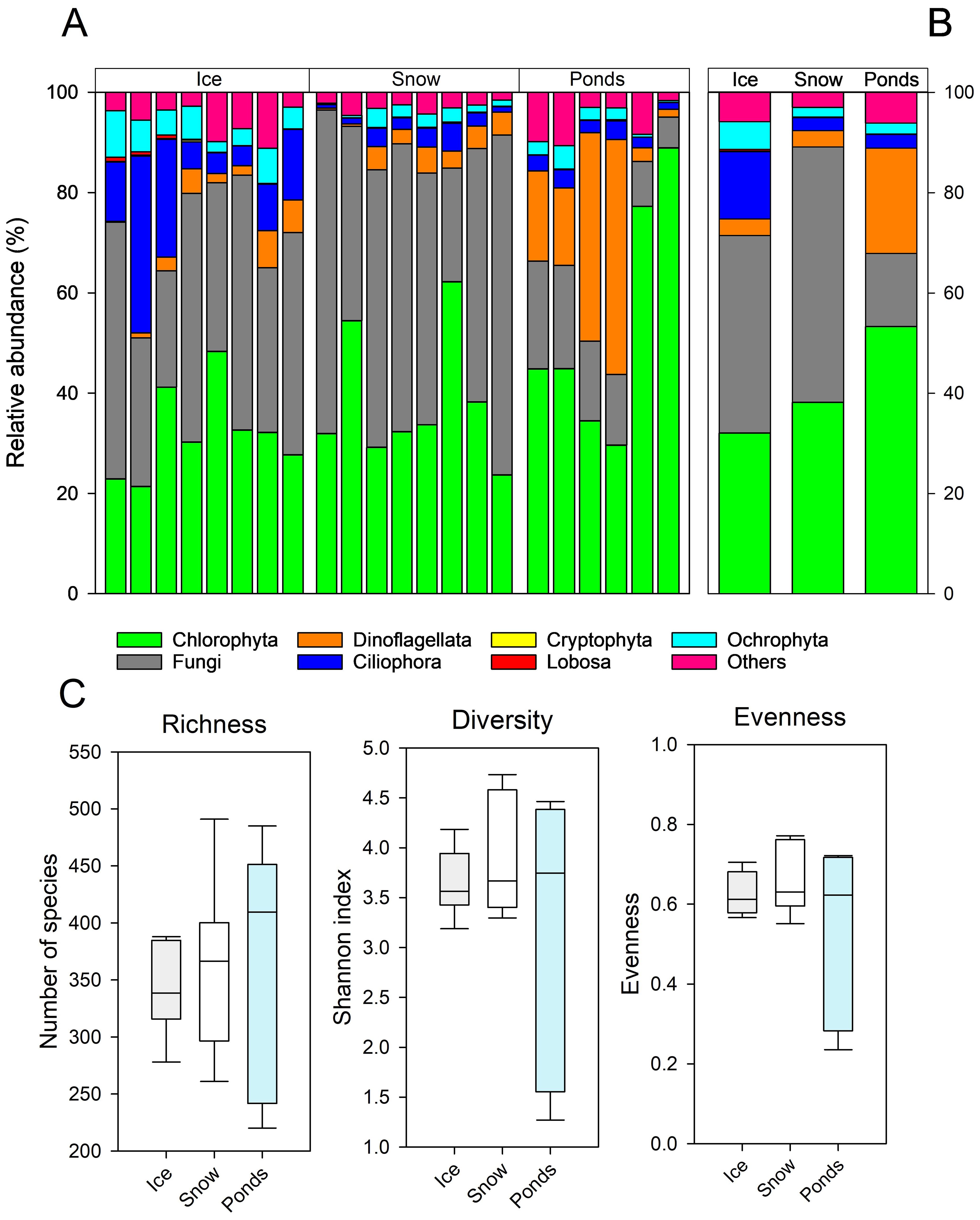
Figure 2. (A) Relative abundance of phylla in each sample. (B) Average of the relative abundance of phylla in each environment. (C) Boxplots of ASV Richness, Shannon diversity, and evenness of eukaryotic communities in each system of the glacial and periglacial environments.
Comparing the three environments, the eukaryotic richness, Shannon diversity, and evenness did not show significant differences (richness: one-way ANOVA, F2,19 = 0.233, P = 0.794; Shannon diversity: one- way ANOVA F2,19 = 1.234, P = 0.313; Evenness: one-way ANOVA, F2,19 = 1.761, P = 0.199). However, in the three parameters, the greatest variation was observed in the ponds (Figure 2C). Glacial ice and snow presented 946 ASVs and 966 ASVs, respectively, while ponds presented a lower value (775 ASVs). A total of 436 ASVs were shared by the three environments, representing 32% of the total ASVs. A similar percentage of ASVs (34%) were unique for each one of the three environments (Figure 3A).
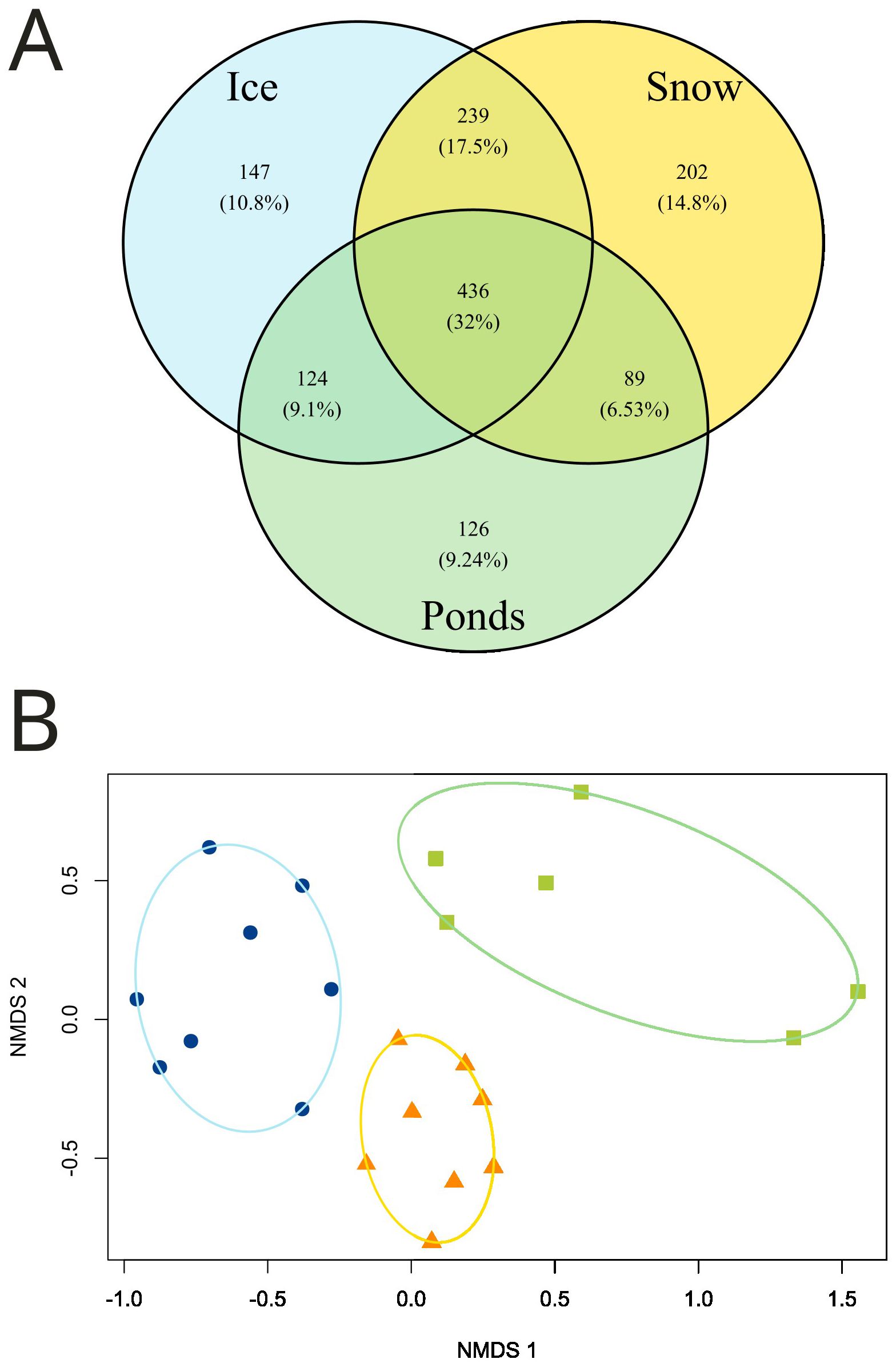
Figure 3. (A) Venn diagram of the ASVs present in the glacial and periglacial environments. (B) Results of the Nonmetric multidimensional analysis (NMDS) based on Bray–Curtis dissimilarity considering all ASVs present in all samples.
However, when we analyze communities based on abundance of ASVs (Bray Curtis index), the NMDS ordination analysis showed that the three environments constitute separate groups (Figure 3B). This was confirmed by the ANOSIM analysis, which showed significant differences among the sampled environments (ANOSIM, global R = 0.70, p < 0.001).
A more deeply analysis of the sequences (heatmap and the cluster analysis in Figure 4) confirmed the separation of the eukaryotic communities in the three environments (ice, snow, and ponds). These partitions resulted from differences in the abundance of the dominant phylotypes: Chlorophyta, fungi, Ciliophora, and Dinoflagellates. The two most abundant ASVs correspond to algae: Chlamydomonadales and Prasiolales, and contributed to the segregation of the ponds and ice, while the next two correspond to Pucciniomyocotina (Fungi) that are mostly exclusive from snow samples (Figure 4). In addition, the following ASVs were also abundant in different environments: ASV5 Chloromonas in snow, ASV6 Suessiaceae (Dinoflagellates) in ponds, and ASV7 Oxytrichidae (Ciliophora) in ice.
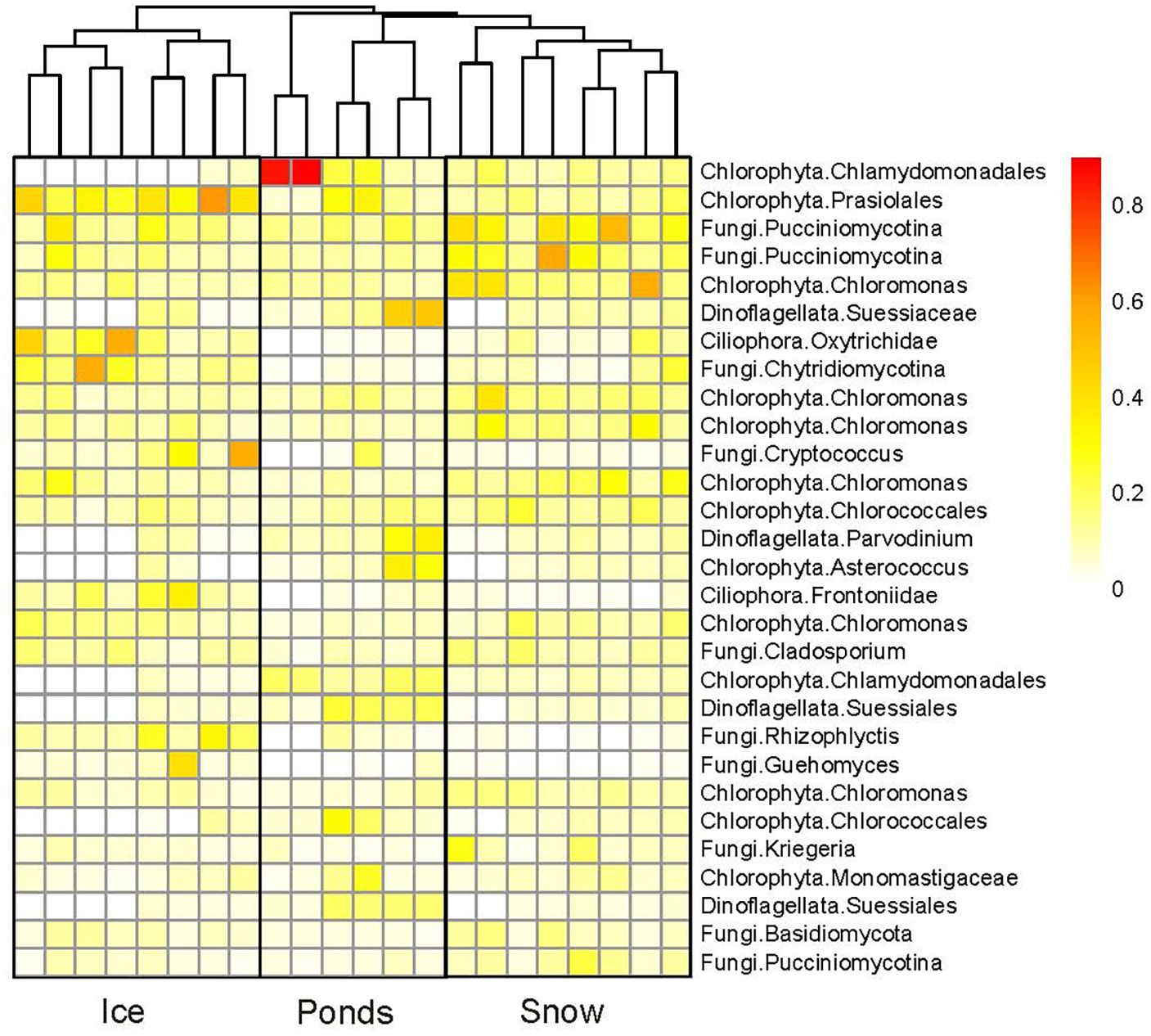
Figure 4. Heat map of the ASV abundances in the samples of the three glacial and periglacial environments. Cluster in the top is based on Bray-Curtis distance.
The analysis of ß-diversity indices enables comparison of communities across different environments, while the partitioning of these indices reveals whether changes in community composition are driven by species turnover or differences in relative abundances. The presence/absence dissimilarity analyses of the total ß-diversity (ßSOR) and turnover (ßSIM) showed low values among sectors (all βSOR and βSIM < 0.59) (Table 3, Supplementary Figure S2). Accordingly, nestedness (ßNES) values were very low (<0.08). On the contrary, values of ß-diversity based on ASVs abundance were higher (all βBC and βBC.BAL > 0.75). In all cases, maximum values were observed when comparing glacial ice and ponds (Table 3). When we compared the observed ß-diversity values of our presence/absence dataset with those generated by the null models, we observed that the differences between glacial ice and snow, and snow and ponds are significant, with an increase in the turnover component (Table 3). On the contrary, in the comparisons based on the abundance dataset, all pairwise comparisons gave significant results, indicating that although environments share ASVs, the changes in abundance differed more (Table 3, Supplementary Figure S2).
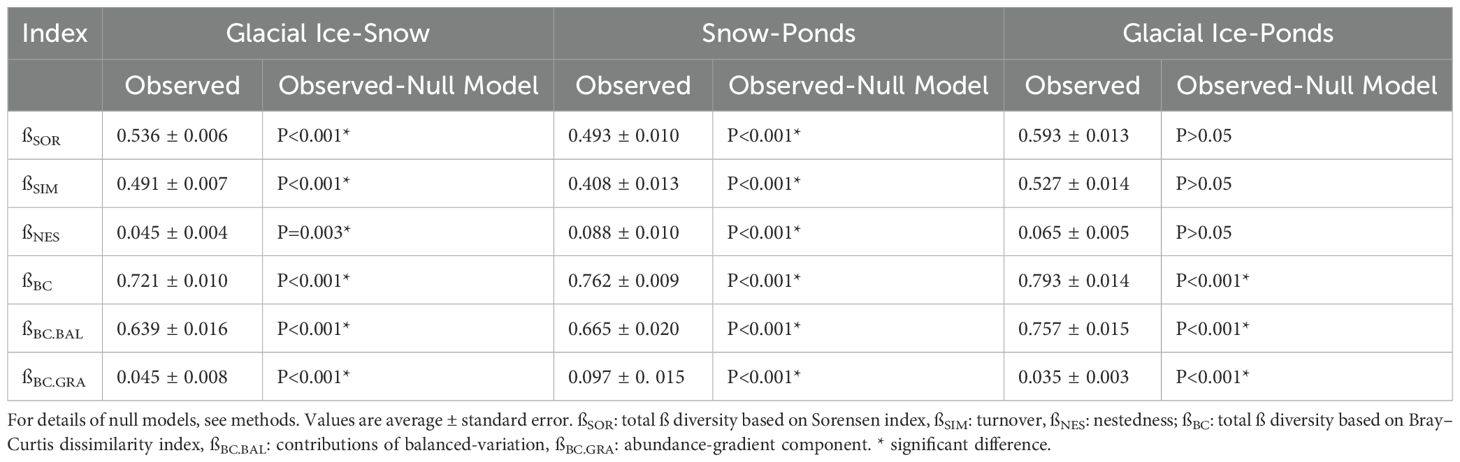
Table 3. Observed values of ß diversity and components among glacial and periglacial environments (glacial ice-snow and ponds) based on presence-absence data (Sorensen index) and abundance data (Bray Curtis index) and the statistical results of the comparison between observed and null models.
The network analysis was conducted to identify the eukaryotic organism co-occurrence patterns in the three glacial environments (Figure 5A). The glacial ice and snow exhibited higher modularity (0.74 in ice and 0.66 in snow) and module number (6 and 5, respectively). Modularity was lower in the ponds (0.52) and we also observed a low number (3) of modules. In the three systems, we identified modules with more than 100 ASVs. In the ponds, 45% of ASVs do not correspond to any module > 5 ASVs, and more than 25% of the total ASVs co-occurred in only one module. Algae and fungi co-occurred in most of the modules; however, there were modules composed mainly of algae. In particular, module 3 in ice and 2 in snow correspond to Chlamydomonadales, Chrysophyceae, Dinophyceae, among others, that were positively connected.
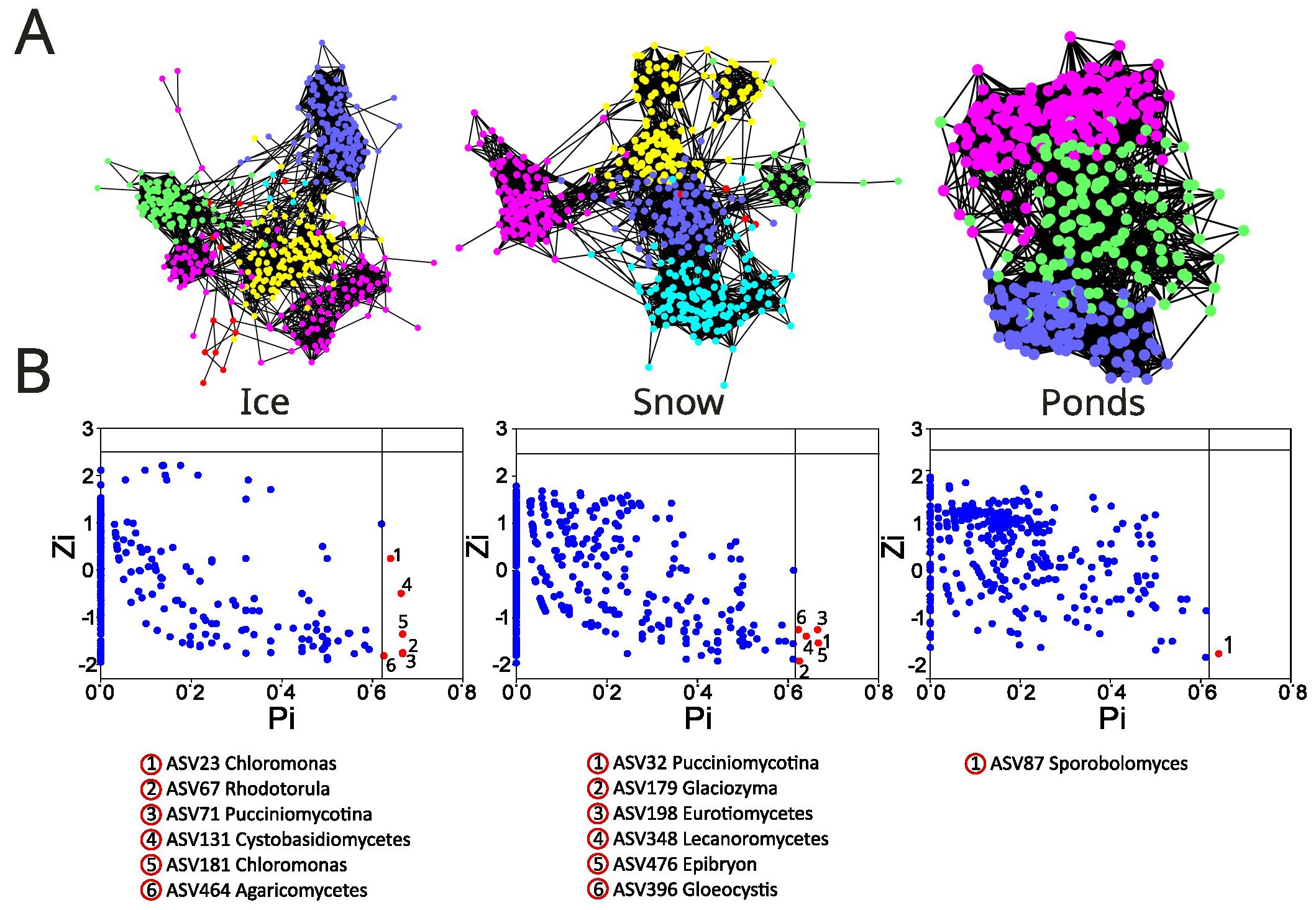
Figure 5. (A) Module structures of eukaryote co-occurrence networks in the three glacial and periglacial environments showing changes in modularity. (B) Topological roles in the network according to the within-module connectivity, Zi, and among-module connectivity, Pi. Blue dots: Peripherals (Zi ≤ 2.5 and Pi ≤ 0.62), and yellow dots: connectors (Zi ≤ 2.5 and Pi > 0.62). No nodes were categorized as modules and network hubs in any of the three glacial environments (Zi >2.5).
The topological roles of individual nodes (ASVs) in the network were classified according to the relationship within-module connectivity (Zi) and among-module connectivity (Pi) (Figure 5B). We did not observe the presence of network hubs or module hubs in any of the three glacial environments. The presence of connectors (Pi>0.625) was detected in the three environments, however, with different numbers. In the glacial ice and snow, the number of connectors was much higher (6) than in the ponds (1). In all cases, connectors are fungi (especially Pucciniomycotina) that include species that produce both yeast and hyphal states. In both the glacial ice and snow, there was one algal species (Chloromonas and Gloeocystis, respectively) acting as a connector. In the ponds, the only identified connector was Sporobolomyces (Pucciniomycotina, Microbotryomycetes).
Discussion
Dissolved nutrient concentrations are low, as is typical in many supraglacial environments (Ren et al., 2019). However, particulate elements are present at higher concentrations, suggesting that nutrients may exist in alternative forms such as decaying organic matter, particulate debris, and other materials (Antony et al., 2017). Additionally, the input of glacial clay affects both light penetration and nutrient availability in lakes and streams (Martyniuk et al., 2014; Rose et al., 2014). As glacial inputs decrease, an increase in phosphorus (P) limitation is expected (Elser et al., 2020; Martyniuk et al., 2022), or alternatively, nitrogen–phosphorus (N:P) co-limitation may occur (Ren et al., 2019). Ren et al. (2019) predicted a more rapid decline in nitrogen (N) availability compared to P as glacier retreat progresses, leading to a shift from P limitation to N–P co-limitation. In contrast, Elser et al. (2020) suggested that N availability would increase while P availability would decrease with diminishing glacial influence, resulting in a shift from N limitation to P limitation.
Studies have highlighted that certain aspects of phosphorus cycling in glacial environments have been largely overlooked, particularly the processes of P adsorption and re-adsorption onto mineral surfaces, which can lead to different outcomes in terms of elemental limitation (Stibal et al., 2008). In our study, the low particulate N:P ratios suggest nitrogen (N) limitation; however, the dissolved fraction presents a contrasting picture. Dissolved N:P ratios were well above 15, indicating potential phosphorus (P) limitation. Indeed, in a previous study, Modenutti et al. (2018) also found that total phosphorus (TP) concentrations were high and strongly correlated with total suspended solids, whereas total dissolved phosphorus (TDP) showed no such relationship. Thus, the apparent discrepancy in nutrient limitation may stem from P adsorption onto various mineral surfaces, such as glacial clays.Supraglacial primary production (by algae in glacial ice and snow) promotes the accumulation of autochthonous organic C (Williamson et al., 2019; Vanderwall et al., 2024). During summer, with warm temperatures and high solar radiation, there is a near constant melt. Thus, any organic matter that was fixed on the supraglacial surface (ice and snow) and not consumed by local heterotrophs (in our study, mainly fungi and ciliates, but also bacteria) is exported to downstream environments (ponds in our study). In hydrologically connected habitats, the metabolic activity of supraglacial organisms may affect the environmental availability of nutrients and shape trophic relationships in downstream habitats.
The analyses of the eukaryotic communities showed a clear dominance of fungi, algae, and ciliophora, though with different relative abundances in the three systems. We did not observe an important number of ASVs losses in the hydrologically connected systems but a community reorganization with different relative abundances. This was particularly evident when comparing beta-diversity based on presence-absence (Sorensen index) and relative abundances (Bray-Curtis index). This means that changes in relative abundances dominate over species turnover in the reorganization of the communities across environments. Moreover, the maximum differences were observed when comparing ice and ponds, with the balanced-variation (changes in relative abundances) having a higher contribution, while the abundance-gradient is low. In addition, the ß-diversity obtained from the null models differed from that of the observed ones. This implies that the relative abundances observed across environments were not due to random processes, but due to changes in the environmental conditions where the communities are established. As physical conditions of the systems changed (ice-snow-ponds), some species will be favored (environmental filtering) and this seemed to be the case of algae that increased their importance in the ponds. Several studies have discussed that non-motile algal cell stages rest beneath the snow during winter and rely on cellular motility to emerge, swimming upward during summer meltwater (Détain et al., 2025). Indeed, in the ponds that received meltwater from ice and snow, we observed an increase in the abundance of motile cells such as Dinoflagellates.
Microbial communities can be considered as a network of species connected by interspecific interactions (i.e. competition, predation, mutualism, and parasitism) (Barberan et al., 2012; Jia et al., 2021), that play substantial roles in promoting microbial diversity and ecosystem functioning (Ratzke et al., 2020). A module is a group of nodes (ASVs) with high interactions among them and low interactions with other nodes outside the module; while modularity measures how well a network separates into different modules (Deng et al., 2012). The co-occurrence network analysis showed a decrease in modularity in the ponds in which only three modules (with five or more nodes) were identified. Modularity may indicate habitat heterogeneity (Olesen et al., 2007) and an increase in modularity seemed to reflect more intensive functional associations (Zhou et al., 2021). Thus, the increased modularity observed in ice and snow suggests greater heterogeneity, likely driven by higher physical complexity (Bachy et al., 2011; Détain et al., 2025). At the same time, the presence of modules (6 in ice and 5 in snow) composed of fungi and algae would reflect the increase in functional associations between these components.
Algae generate bioavailable dissolved organic matter profited by heterotrophic communities (Anesio et al., 2010), and their presence in snow and ice has been reported on all the continents (Lutz et al., 2016). The most frequent algae belong to the Chlamydomonadales (Chlorophyta) group, that includes species responsible for snow algal blooms (Brown and Tucker, 2020; Hoham and Remias, 2020). The interplay between carbon-fixing algae and carbon-respiring heterotrophic microorganisms ultimately controls the amount and composition of organic matter in ice and snow (Rossel et al., 2025).
Among the fungi we identified Pucciniomycotina psychrophilic yeasts (ASV3 and ASV4, Figure 4) that inhabit snowfields and glacial ecosystems worldwide, including glaciers in Greenland and Svalbard, the Alps, and Antarctica (Margesin and Collins, 2019; Perini et al., 2021). Different Pucciniomycotina species have a key role in snow and ice ecosystems, since they are capable of degrading organic molecules during algal blooms, connecting the carbon cycle in cold environments (Marechal et al., 2025). We also found members of the Chytridiomycotina subdivision (ASV 8, Figure 4) that were previously cited in the studied area (Duo Saito et al., 2018). Chytridiomycota have been described as abundant in alpine and Arctic snow, snowpacks of Colorado and Switzerland (Lutz et al., 2017), and in glacier meltwater and sediments in a fjord in Chilean Patagonia (Gutiérrez et al., 2015). Chytridiomycetes are nutritionally dependent on the algae enrichment of snow patches (Brown et al., 2018), and the presence of this group associated with cryophilic algae in our ice and snow environments suggests a tied dependence. Moreover, the nutritional dependence of fungi on algae (Rossel et al., 2025) would be related to the observed increase in functional associations in the modules of ice and snow.
In the context of network analysis, the presence of a connector refers to a node (ASV) that links two modules within a network (Zhou et al., 2021). These nodes play a role in connecting or facilitating interactions across different modules of organisms within the community. In our case, the presence of more connectors in ice and snow can also be related to the interaction between fungi and algae (Brown et al., 2018; Rossel et al., 2025). Most of these connectors are fungi that would indicate their dependence on more than one species association (module). On the one hand, fungi depend on algae for nutrition, and on the other hand, fungal and algal communities may be influenced by similar environmental drivers (Brown et al., 2018). In our case study, in all three systems, the connectors are fungi, and two algal genera (Chloromonas and Gloeocystis, in ice and snow, respectively). The identification of fungi as the main connectors across all environments suggests their potential role as functional integrators within the eukaryotic community network, linking different modules and possibly mediating resource flows or ecological interactions between algae and other microbial groups (Brown et al., 2018). The few algae identified as connectors and the existence of modules composed mainly of algae would suggest that primary producers are more involved as module members.
The change in water state towards meltwater in the ponds produces a substantial reduction in the number of modules in the network. Indeed, in the ponds, we only observed three modules and one connector. The unique connector was Sporobolomyces (Pucciniomycotina), a common yeast previously found on glaciers in Patagonia (de Garcia et al., 2012). Studies have shown that these yeasts are cold-adapted and can produce extracellular enzymatic activities even at low temperatures (de Garcia et al., 2007). This capacity may allow this yeast to prevail as a connector in early aquatic ecosystems.
Currently, the ponds located at the glacier forefield of Castaño Overo and Alerce are still hydrologically connected to glaciers as the direct contact with glaciers has been lost. As glaciers retreat, direct contact is lost; however, some ponds continue to receive meltwater, maintaining the hydrological connectivity. Glaciers in the northern Patagonian Andes have shown a retreat in the last decades (Masiokas et al., 2020). Here, we showed that the organization of the eukaryotic community changes substantially as water melts, in particular in the relative abundance of taxa and in the community network organization. Based on the present data on glacial-periglacial environments, we did not find evidence of potential biodiversity loss. Although we cannot predict if these environments will definitely lose their glacial inputs, the reductions in ice masses in Patagonia will affect downstream aquatic environments, reducing the glacial influence in their biotic communities.
Data availability statement
The data presented in this study are publicly available. This data can be found here: https://www.ncbi.nlm.nih.gov, BioProject accession number PRJNA1270387 and BioSample accession numbers SAMN48824426-SAMN48824447.
Author contributions
BM: Conceptualization, Funding acquisition, Investigation, Project administration, Supervision, Validation, Writing – original draft, Writing – review & editing. EV: Data curation, Formal analysis, Investigation, Writing – review & editing. MBN: Data curation, Investigation, Writing – review & editing. NM: Data curation, Investigation, Writing – review & editing. EB: Conceptualization, Formal analysis, Funding acquisition, Investigation, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was supported by grants FONCyT2018-1563 and FONCyT2020-0383.
Acknowledgments
We are grateful to IANIGLA for lending us the sampling ice device. We thank Lucas Ruiz (IANIGLA), Agustina Iglesias Louis and Luca Schenone for the sampling of snow and ice. We thank the Administración del Parque Nacional Nahuel Huapi for the permission for sampling Tronador Mountain.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2025.1670481/full#supplementary-material
Supplementary Figure 1 | Rarefaction curves.
Supplementary Figure 2 | ß-diversity matrices.
References
Anesio A. M. and Laybourn-Parry J. (2012). Glaciers and ice sheets as a biome. Trends Ecol. Evol. 27, 219–225. doi: 10.1016/j.tree.2011.09.012
Anesio A. M., Lutz S., Chrismas N. A. M., and Benning L. G. (2017). The microbiome of glaciers and ice sheets. NPJ Biofilms Microbiomes 3. doi: 10.1038/s41522-017-0019-0
Anesio A. M., Sattler B., Foreman C., Telling J., Hodson A., Tranter M., et al. (2010). Carbon fluxes through bacterial communities on glacier surfaces. Ann. Glaciol. 51, 32–40. doi: 10.3189/172756411795932092
Antony R., Willoughby A. S., Grannas A. M., et al. (2017). Molecular insights on dissolved organic matter transformation by supraglacial microbial communities. Environ. Sci. Technol. 51, 4328–4337. doi: 10.1021/acs.est.6b05780
APHA (2005). Standard methods for the examination of water and wastewater (Washington DC, USA: American Public Health Association).
Bachy C., Lopez-Garcia P., Vereshchaka A., and Moreira D. (2011). Diversity and vertical distribution of microbial eukaryotes in the snow, sea ice and seawater near the north pole at the end of the polar night. Front. Microbiol. 2. doi: 10.3389/fmicb.2011.00106
Barberan A., Bates S. T., Casamayor E. O., and Fierer N. (2012). Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 6, 343–351. doi: 10.1038/ismej.2011.119
Baselga A. and Chao A. (2017). Partitioning abundance-based multiple-site dissimilarity into components: balanced variation in abundance and abundance gradients. Methods Ecol. Evol. 8, 799–808. doi: 10.1111/2041-210x.12693
Baselga A. and Orme C. D. L. (2012). betapart: an R package for the study of beta diversity. Methods Ecol. Evol. 3, 808–812. doi: 10.1111/j.2041-210X.2012.00224.x
Bosson J.-B., Huss M., Cauvy-Fraunié S., Clément J.-C., Costes G., Fischer M., et al. (2023). Future emergence of new ecosystems caused by glacial retreat. Nature 620, 562–569. doi: 10.1038/s41586-023-06302-2
Brown S. P. and Jumpponen A. (2019). Microbial ecology of snow reveals taxa-specific biogeographical structure. Microb. Ecol. 77, 946–958. doi: 10.1007/s00248-019-01357-z
Brown S. P., Olson B. J. S. C., and Jumpponen A. (2018). Fungi and algae co-occur in snow: an issue of shared habitat or algal facilitation of heterotrophs? Arct. Antarct. Alp. Res. 47, 729–749. doi: 10.1657/aaar0014-071
Brown S. P. and Tucker A. E. (2020). Distribution and biogeography of Sanguina snow algae: Fine-scale sequence analyses reveal previously unknown population structure. Ecol. Evol. 10, 11352–11361. doi: 10.1002/ece3.6772
Callahan B. J., McMurdie P. J., Rosen M. J., Han A. W., Johnson A. J., and Holmes S. P. (2016). DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583. doi: 10.1038/nmeth.3869
Cameron K. A., Hagedorn B., Dieser M., Christner B. C., Choquette K., Sletten R., et al. (2015). Diversity and potential sources of microbiota associated with snow on western portions of the Greenland Ice Sheet. Environ. Microbiol. 17, 594–609. doi: 10.1111/1462-2920.12446
Cheung M. K., Au C. H., Chu K. H., Kwan H. S., and Wong C. K. (2010). Composition and genetic diversity of picoeukaryotes in subtropical coastal waters as revealed by 454 pyrosequencing. ISME J. 4, 1053–1059. doi: 10.1038/ismej.2010.26
Comte J., Culley A. I., Lovejoy C., and Vincent W. F. (2018). Microbial connectivity and sorting in a High Arctic watershed. ISME J. 12, 2988–3000. doi: 10.1038/s41396-018-0236-4
de Garcia V., Brizzio S., Libkind D., Buzzini P., and van Broock M. (2007). Biodiversity of cold-adapted yeasts from glacial meltwater rivers in Patagonia, Argentina. FEMS Microbiol. Ecol. 59, 331–341. doi: 10.1111/j.1574-6941.2006.00239.x
de Garcia V., Brizzio S., and van Broock M. R. (2012). Yeasts from glacial ice of Patagonian Andes, Argentina. FEMS Microbiol. Ecol. 82, 540–550. doi: 10.1111/j.1574-6941.2012.01470.x
Deng Y., Jiang Y.-H., Yang Y., He Z., Luo F., and Zhou J. (2012). Molecular ecological network analyses. BMC Bioinf. 13, 1–20. Available online at: http://www.biomedcentral.com/1471-2105/13/113.
Détain A., Suzuki H., Wijffels R. H., Leborgne-Castel N., and Hulatt C. J. (2025). Snow algae exhibit diverse motile behaviors and thermal responses. mBio, e02954–e02924. doi: 10.1128/mbio.02954-24
Duo Saito R. A., Connell L., Rodriguez R., Redman R., Libkind D., and de Garcia V. (2018). Metabarcoding analysis of the fungal biodiversity associated with Castaño Overa Glacier – Mount Tronador, Patagonia, Argentina. Fungal Ecol. 36, 8–16. doi: 10.1016/j.funeco.2018.07.006
Elser J. J., Wu C., Gonzalez A. L., Shain D. H., Smith H. J., Sommaruga R., et al. (2020). Key rules of life and the fading cryosphere: Impacts in alpine lakes and streams. Glob Chang Biol. 26, 6644–6656. doi: 10.1111/gcb.15362
Fell S. C., Carrivick J. L., and Brown L. E. (2017). The multitrophic effects of climate change and glacier retreat in mountain rivers. Bioscience 67, 897–911.
Filker S., Sommaruga R., Vila I., and Stoeck T. (2016). Microbial eukaryote plankton communities of high-mountain lakes from three continents exhibit strong biogeographic patterns. Mol. Ecol. 25, 2286–2301. doi: 10.1111/mec.13633
French H. and Thorn C. E. (2006). The changing nature of periglacial geomorphology. Géomorphologie: relief processus environnement 12, 3. doi: 10.4000/geomorphologie.119
Ghilardi Truffa J. C. and Ruiz L. (2024). “Debris-covered area increased in the Central Andes of Argentina glaciers over the past four decades,” in European Geosciences Union, General Assembly Conference Abstracts, (Vienna, Austria), 1171.
Guimera R. and Amaral L. A. (2005). Cartography of complex networks: modules and universal roles. J. Stat. Mech. 2005, nihpa35573. doi: 10.1088/1742-5468/2005/02/P02001
Gutiérrez M. H., Galand P. E., Moffat C., and Pantoja S. (2015). Melting glacier impacts community structure of B acteria, A rchaea and F ungi in a C hilean P atagonia fjord. Environ. Microbiol. 17, 3882–3897. doi: 10.1111/1462-2920.12872
Hoham R. W. and Remias D. (2020). Snow and glacial algae: A review. J. Phycol. 56, 264–282. doi: 10.1111/jpy.12952
Hood E., Fellman J., Spencer R. G., Hernes P. J., Edwards R., D'Amore D., et al. (2009). Glaciers as a source of ancient and labile organic matter to the marine environment. Nature 462, 1044–1047. doi: 10.1038/nature08580
Jia M., Gao Z., Gu H., Zhao C., Liu M., Liu F., et al. (2021). Effects of precipitation change and nitrogen addition on the composition, diversity, and molecular ecological network of soil bacterial communities in a desert steppe. PloS One 16, e0248194. doi: 10.1371/journal.pone.0248194
Kammerlander B., Breiner H. W., Filker S., Sommaruga R., Sonntag B., and Stoeck T. (2015). High diversity of protistan plankton communities in remote high mountain lakes in the European Alps and the Himalayan mountains. FEMS Microbiol. Ecol. 91. doi: 10.1093/femsec/fiv010
Krug L., Erlacher A., Markut K., Berg G., and Cernava T. (2020). The microbiome of alpine snow algae shows a specific inter-kingdom connectivity and algae-bacteria interactions with supportive capacities. ISME J. 14, 2197–2210. doi: 10.1038/s41396-020-0677-4
Laybourn-Parry J. and Pearce D. A. (2007). The biodiversity and ecology of Antarctic lakes: Models for evolution. Philos. Trans. R. Soc B 362, 2273–2289. doi: 10.1098/rstb.2006.1945
Lutz S., Anesio A. M., Edwards A., and Benning L. G. (2017). Linking microbial diversity and functionality of arctic glacial surface habitats. Environ. Microbiol. 19, 551–565. doi: 10.1111/1462-2920.13494
Lutz S., Anesio A. M., Raiswell R., Edwards A., Newton R. J., Gill F., et al. (2016). The biogeography of red snow microbiomes and their role in melting arctic glaciers. Nat. Commun. 7, 11968. doi: 10.1038/ncomms11968
Marechal E., Ezzedine J., Guenzi-Tiberi P., et al. (2025). Snow-and ice-ecosystem cleaning capability of the pucciniomycotinous yeast Phenoliferia psychrophenolica. Commun. Biol. 8, 1084. doi: 10.21203/rs.3.rs-5795686/v1
Margesin R. and Collins T. (2019). Microbial ecology of the cryosphere (glacial and permafrost habitats): current knowledge. Appl. Microbiol. Biotechnol. 103, 2537–2549. doi: 10.1007/s00253-019-09631-3
Margesin R. and Miteva V. (2011). Diversity and ecology of psychrophilic microorganisms. Res. Microbiol. 162, 346–361. doi: 10.1016/j.resmic.2010.12.004
Martini I. P., French H. M., and Alberti A. P. (2011). Ice-marginal and periglacial processes and sediments: an introduction. Geological Society London Special Publications 354, 1–13. doi: 10.1144/sp354.1
Martyniuk N., Modenutti B., and Balseiro E. (2014). Can increased glacial melting resulting from global change provide attached algae with transient protection against high irradiance? Freshwat. Biol. 59, 2290–2302. doi: 10.1111/fwb.12431
Martyniuk N., Souza M. S., Bastidas Navarro M., Balseiro E., and Modenutti B. (2022). Nutrient limitation affects biofilm enzymatic activities in a glacier-fed river. Hydrobiologia. 849, 2877–2894. doi: 10.1007/s10750-022-04857-1
Masiokas M. H., Rabatel A., Rivera A., Ruiz L., Pitte P., Ceballos J. L., et al. (2020). A review of the current state and recent changes of the andean cryosphere. Front. Earth Sci. 8. doi: 10.3389/feart.2020.00099
Modenutti B., Bastidas Navarro M., Martyniuk N., and Balseiro E. (2018). Melting of clean and debris-rich ice differentially affect nutrients, dissolved organic matter and bacteria respiration in the early ontogeny of the newly formed proglacial Ventisquero Negro Lake (Patagonia Argentina). Freshwat. Biol. 63, 1341–1351. doi: 10.1111/fwb.13161
Oksanen J., Blanchet F. G., Kindt R., Legendre P., Minchin P. R., O’hara R., et al. (2013). Package ‘vegan’. Community ecology package, version, Vol. 2. 1–295. Available online at: https://github.com/vegandevs/vegan.
Olesen J. M., Bascompte J., Dupont Y. L., and Jordano P. (2007). The modularity of pollination networks. PNAS 104, 19891–19896. doi: 10.1073/pnas.0706375104
Pawlowski J., Audic S., Adl S., Bass D., Belbahri L., Berney C., et al. (2012). CBOL Protist Working Group: Barcoding eukaryotic richness beyond the animal, plant, and fungal kingdoms. PloS Biol. 10, e1001419. doi: 10.1371/journal.pbio.1001419
Perini L., Andrejasic K., Gostincar C., Gunde-Cimerman N., and Zalar P. (2021). Greenland and Svalbard glaciers host unknown basidiomycetes: the yeast Camptobasidium arcticum sp. nov. and the dimorphic Psychromyces glacialis gen. and sp. nov. Int. J. Syst. Evol. Microbiol. 71, 004655. doi: 10.1099/ijsem.0.004655
Ratzke C., Barrere J., and Gore J. (2020). Strength of species interactions determines biodiversity and stability in microbial communities. Nat. Ecol. Evol. 4, 376–383. doi: 10.1038/s41559-020-1099-4
R Core Team (2025). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing).
Ren Z., Martyniuk N., Oleksy I. A., Swain A., and Hotaling S. (2019). Ecological stoichiometry of the mountain cryosphere. Front. Ecol. Evol. 7. doi: 10.3389/fevo.2019.00360
Rose K. C., Hamilton D. P., Williamson C. E., McBride C. G., Fischer J. M., Olson M. H., et al. (2014). Light attenuation characteristics of glacially-fed lakes. J. Geophys. Res. 119, 1446–1457. doi: 10.1002/2014jg002674
Rossel P. E., Antony R., Mourot R., Dittmar T., Anesio A. M., Tranter M., et al. (2025). Dynamics of organic matter in algal blooms on the Greenland ice sheet. Sci. Rep. 15, 8288. doi: 10.1038/s41598-025-92182-7
Ruiz L., Berthier E., Masiokas H. M., Pitte P., and Villalba R. (2015). First surface velocity maps for glaciers of Monte Tronador, North Patagonian Andes, derived from sequential Pléiades satellite images. J. Glaciol 61, 908–922. doi: 10.3189/2015JoG14J134
Ruiz L., Berthier E., Viale M., Pitte P., and Masiokas M. H. (2017). Recent geodetic mass balance of Monte Tronador glaciers, northern Patagonian Andes. Cryosphere 11, 619–634. doi: 10.5194/tc-11-619-2017
Schiaffino M. R., Huber P., Sagua M., Sabio Y. G. C. A., and Reissig M. (2020). Covariation patterns of phytoplankton and bacterioplankton in hypertrophic shallow lakes. FEMS Microbiol. Ecol. 96, fiaa161. doi: 10.1093/femsec/fiaa161
Shannon P., Markiel A., Ozier O., Baliga N. S., Wang J. T., Ramage D., et al. (2003). Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504. doi: 10.1101/gr.1239303
Slaymaker O. (2011). Criteria to distinguish between periglacial, proglacial and paraglacial environments. Quageo 30, 85–94. doi: 10.2478/v10117-011-0008-y
Stibal M., Tranter M., Telling J., and Benning L. G. (2008). Speciation, phase association and potential bioavailability of phosphorus on a Svalbard glacier. Biogeochemistry 90, 1–13. doi: 10.1007/s10533-008-9226-3
Triadó-Margarit X. and Casamayor E. O. (2012). Genetic diversity of planktonic eukaryotes in high mountain lakes (Central Pyrenees, Spain). Environ. Microbiol. 14, 2445–2456. doi: 10.1111/j.1462-2920.2012.02797.x
Vanderwall J. W., Muhlfeld C. C., Tappenbeck T. H., Giersch J., Ren Z., and Elser J. J. (2024). Mountain glaciers influence biogeochemical and ecological characteristics of high-elevation lakes across the northern Rocky Mountains, USA. Limnol. Oceanogr 69, 37–52. doi: 10.1002/lno.12434
Vimercati L., Darcy J. L., and Schmidt S. K. (2019). The disappearing periglacial ecosystem atop Mt. Kilimanjaro supports both cosmopolitan and endemic microbial communities. Sci. Rep. 9, 10676. doi: 10.1038/s41598-019-46521-0
Vincent W. F., Callaghan T. V., Dahl-Jensen D., Johansson M., Kovacs K. M., Michel C., et al. (2011). Ecological implications of changes in the arctic cryosphere. Ambio 40, 87–99. doi: 10.1007/s13280-011-0218-5
Williamson C. E., Neale P. J., Hylander S., Rose K. C., Figueroa F. L., Robinson S. A., et al. (2019). The interactive effects of stratospheric ozone depletion, UV radiation, and climate change on aquatic ecosystems. Photochem. Photobiol. Sci. 18, 717–746. doi: 10.1039/c8pp90062k
Winkel M., Trivedi C. B., Mourot R., Bradley J. A., Vieth-Hillebrand A., and Benning L. G. (2022). Seasonality of glacial snow and ice microbial communities. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.876848
Yakimovich K. M., Engstrom C. B., and Quarmby L. M. (2020). Alpine snow algae microbiome diversity in the coast range of british columbia. Front. Microbiol. 11. doi: 10.3389/fmicb.2020.01721
Zemp M., Frey H., Gärtner-Roer I., Nussbaumer S. U., Hoelzle M., Paul F., et al. (2015). Historically unprecedented global glacier decline in the early 21st century. J. Glaciol. 61, 745–762. doi: 10.3189/2015JoG15J017
Zhou J., Deng Y., Luo F., He Z., and Yang Y. (2011). Phylogenetic molecular ecological network of soil microbial communities in response to elevated CO2. MBio 2. doi: 10.1128/mBio.00122-11
Zhou L., Zhou Y., Tang X., Zhang Y., Zhu G., Székely A. J., et al. (2021). Eutrophication alters bacterial co-occurrence networks and increases the importance of chromophoric dissolved organic matter composition. Limnol. Oceanogr. 66, 2319–2332. doi: 10.1002/lno.11756
Keywords: eukaryotic communities, glacier, snow, glacial forefield, eukaryotic diversity, network analysis, North-Patagonian Andes
Citation: Modenutti B, Vega E, Bastidas Navarro M, Martyniuk N and Balseiro E (2025) Change in relative abundances, more so than species turnover, modulates ß-diversity and network complexity in communities of eukaryotic microorganisms from glacial to periglacial environments. Front. Ecol. Evol. 13:1670481. doi: 10.3389/fevo.2025.1670481
Received: 21 July 2025; Accepted: 09 October 2025;
Published: 30 October 2025.
Edited by:
Paulina Fermani, CONICET Instituto de Biología de Organismos Marinos (IBIOMAR), ArgentinaReviewed by:
Paula Huber, Federal University of São Carlos, BrazilManuel Castro Berman, Rensselaer Polytechnic Institute, United States
Copyright © 2025 Modenutti, Vega, Bastidas Navarro, Martyniuk and Balseiro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Beatriz Modenutti, Ym1vZGVudXR0aUBjb21haHVlLWNvbmljZXQuZ29iLmFy