 William J. Harvey1*
William J. Harvey1* Sandra Nogué2
Sandra Nogué2 Nathan Stansell3
Nathan Stansell3 Gillian Petrokofsky1
Gillian Petrokofsky1 Byron Steinman4
Byron Steinman4 Katherine J. Willis1
Katherine J. Willis1- 1Long–Term Ecology Laboratory, Department of Zoology, University of Oxford, Oxford, United Kingdom
- 2School of Geography and Environmental Science, University of Southampton, Southampton, United Kingdom
- 3Department of Geology and Environmental Geosciences, Northern Illinois University, DeKalb, IL, United States
- 4Large Lakes Observatory and Department of Earth and Environmental Sciences, University of Minnesota Duluth, Duluth, MN, United States
A Corrigendum on
The Legacy of Pre–Columbian Fire on the Pine–Oak Forests of Upland Guatemala
by Harvey, W. J., Nogué, S., Stansell, N., Petrokofsky, G., Steinman, B., and Willis, K. J. (2019). Front. For. Glob. Change 2:34. doi: 10.3389/ffgc.2019.00034
In the original article, there was an error. After discussions with the wider scientific community, the pollen taxa identified as Corylus is more likely to be Myrica; and the pollen taxa identified as Cercocarpus is more likely to be Terminalia.
A correction has therefore been made to the Results, subsection Palaeoecological Trends, paragraph two and three:
“Results from the CCA show that microscopic and macroscopic charcoal are significant environmental variables most associated with Zones 2 and 3, while Sporormiella is most associated with Zone 1 and is not statistically significant (Figure 4A). The PCA displays a distinct gradient and several associations between taxa and samples (Figure 4B). The first axis represents 20.2% of the variation, while axis two represents 10.5% of the variation. The arch between samples suggests that there is only one clear gradient. When the independently calculated palynological Zones are superimposed upon these quadrants, the top, and bottom right quadrants are most associated with Zone 1, the top left quadrant is most associated with Zone 2 and the bottom left quadrant is most associated with Zone 3 (Figure 4B). The first taxonomic association comprises canopy taxa Quercus, understory taxa Leguminosae, Myrica, Ericaceae, and the herbaceous and agrarian taxa Capsicum, which is most associated with Zone 1. The second taxonomic association comprises of understory taxa Juniperus and Terminalia, herbaceous taxa Polygalaceae, Apiaceae, Compositae Aphelandra, and agrarian taxa Capsicum most associated with Zone 2. The third taxonomic association comprises canopy taxa Pinus, Alnus, and Liquidambar together with understory taxa Morella cerifera, and herbaceous taxa Poaceae, and Campanulaceae, which is most associated with Zone 3 (Figure 4B).”
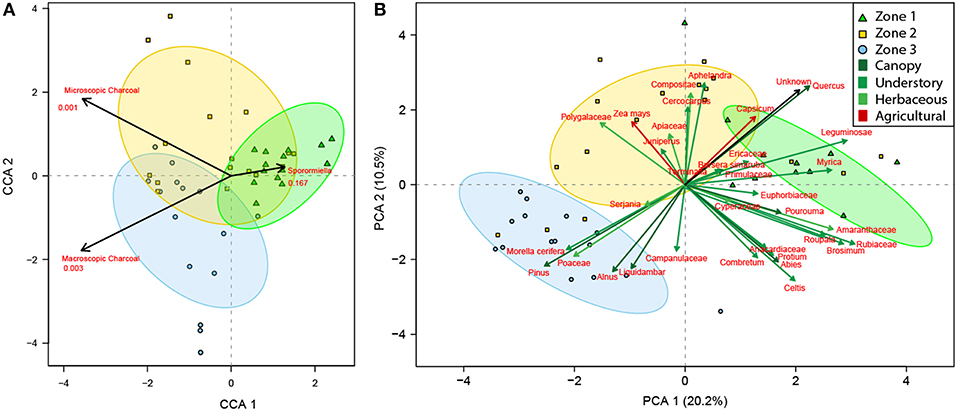
Figure 4. Canonical correspondence analysis of the palaeopalynological data set ordinated against independent environmental indicators of local fire (macroscopic charcoal), regional fire (microscopic charcoal), and herbivory (Sporormiella) (A). Principal component analysis of the palaeopalynological data set (B). Zones are derived from the broken stick model and are represented by ellipses at a confidence of 95%. Zone 1 = green triangles; Zone 2 = yellow squares; Zone 3 = blue circles.
“Zone 1 (545.75–421 cm, 14 samples, 4000–2200B.C.E.) spans c.1800 years concurrent with the last 2000 years of the Archaic Period (10,000–2000B.C.E.) (Figure 5). This Zone is predominantly defined by POF taxa Quercus (25.9%) and Pinus (11%) alongside herbaceous taxa Compositae (27.6%). Between 4000 and 3300 B.C.E. there is evidence for a decline in canopy taxa (from 61.3 to 37.3% of the total pollen sum), particularly Quercus (18–9.7%). In contrast, Pinus became abundant during this period, rising from 4000B.C.E. (2.3%) to 3300B.C.E. (16.7%). Myrica and Alnus are mostly present throughout this Zone, peaking at 3100B.C.E. (15.3%). Brosimum (0–4.3%), Anacardiaceae (0.3–4.3%), Leguminosae (0–6.3%), Morella cerifera, and Rubiaceae (0–9%) are present in low abundance and on average decrease from 4000 to 2200B.C.E. The ratio of coniferous to MHWF is on average 30:70. The coniferous to MHWF ratio changes from 25:75 to 66:34 between 3700 and 3300B.C.E. Compositae abundance increases between 4000 and 3200B.C.E. (17.3–35.3%) and peaks at 2900B.C.E. (53.3%) and 2700B.C.E. (47.3%). Amaranthaceae is present at 10.3% from 4000B.C.E. but subsequently declines leading up to 3700B.C.E. (1.3%). Capsicum appears and increases from 3850 to 3300 B.C.E. (0.3–5.3%), and Polygalaceae follows a similar trend, peaking at 3300B.C.E. (4.3%). Poaceae is stable and in low abundance throughout the record (2.3–5.6%). Pollen influx is low, decreasing from 4000 to 3300B.C.E. (965–344 grains x103 cm2 per year) and increasing after 2300B.C.E. (1,919 grains x104 cm2 per year). Sporomiella abundance is relatively high and stable (2.6–10.5%), falling below 4% abundance at 3100B.C.E. (2.6%), 2700B.C.E. (2.9%), and 2320B.C.E. (3.8%); there is a sustained high abundance of >10% between 2600 and 2500B.C.E. (10.2–13.3%). Macroscopic and microscopic charcoal are relatively low, decreasing between 4000 and 3000B.C.E. (macroscopic charcoal: 0.44–0.32 particles cm2 per year; and, microscopic charcoal: 552–86 particles cm2yr1) then increasing through to 2300B.C.E (macroscopic: 1.2 particles cm2 per year; and, microscopic: 770 particles cm2 per year).”
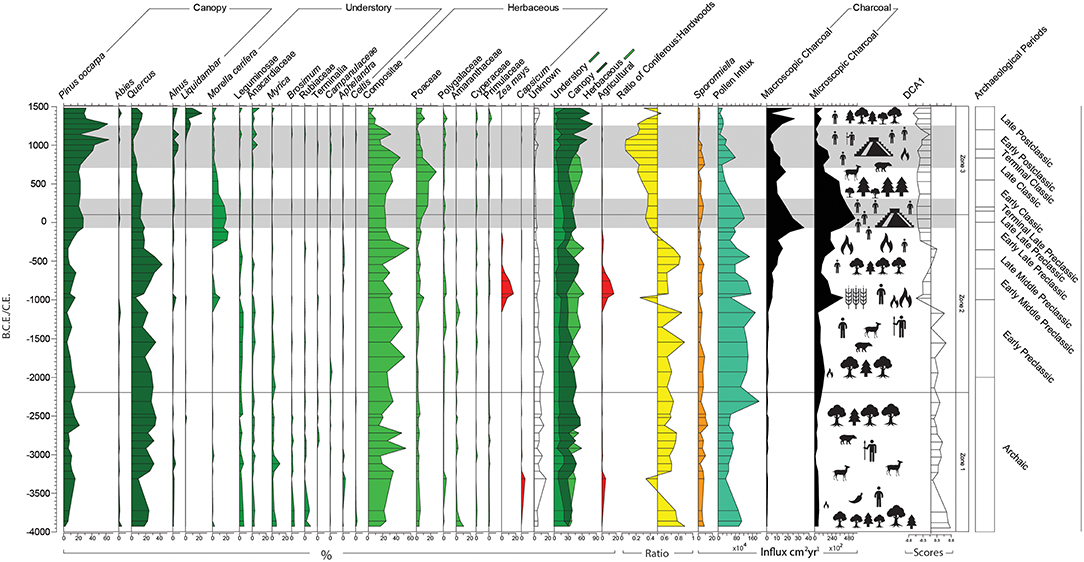
Figure 5. Palynological percentage diagram of taxa appearing in an abundance >2%; forest structure; coniferous to hardwood ratio; pollen influx; macroscopic and microscopic influx; DCA axis 1; and Occupation of Chinkultic (dark bands). The palynological data are expressed as a percentage of total land pollen.
A correction has also been made to the Discussion, subsection Anthropogenic Impacts and Agrarian Practices, paragraph two:
Archaic populations from the Maya Area combined agriculture (e.g., pepper, beans, maize, squash, and chili) with hunting and gathering from as early as 5200B.C.E. (Pope et al., 2001). Evidence for agriculture from Centote Kail is first indicated by vegetation changes from 4000B.C.E. The observed changes include (i) a decline in canopy and understory taxa; (ii) increases in weedy disturbance taxa; (iii) a very small increase in local and regional burning; (iv) and the presence of cultivated taxa such as Capsicum (White, 1999), between c.3850 and 3300B.C.E. or Zea maysbetween c.970 and 550B.C.E. (Figure 5). A mixture of traditional hunter–gather practices combined with limited agriculture is exemplified by the abundance and variety of edible fruits and nuts (e.g., Brosimum, Myrica, and Anacardiaceae) and high abundance of fauna as indicated by the influx of Sporormiella.
Additionally, a correction has been made to Discussion, subsection Forest Structure, Composition and Successional Regeneration, paragraph one:
“Relatively equal abundances of canopy, understory, and herbaceous taxa from 4000 to 3700B.C.E. indicate a diverse and stratified forest structure comprising at least three vegetative levels. The high diversity of taxa, particularly in the understory (e.g., Leguminosae, Anacardiaceae, Myrica, and Rubiaceae), suggests a relatively low and open canopy allowing light to penetrate to the forest floor (Bush, 2000). This structure is typical of middle succession in mixed POF after a large clearance event (Peterson and Reich, 2001).”
Corrections have also been made in Figure 4 and Figure 5. The correct figures appear above.
Lastly, the Supplementary Material (including Supplementary Tables 1 and 2 and the raw data) has also been updated to reflect the correct taxa.
The authors apologize for this error and state that this does not change the scientific conclusions of the article in any way. The original article has been updated.
References
Bush, M. B. (2000). Deriving response matrices from Central American modern pollen rain. Quaternary Res. 54, 132–143. doi: 10.1006/qres.2000.2138
Peterson, D. W., and Reich, P. B. (2001). Prescribed fire in oak savanna: fire frequency effects on stand structure and dynamics. Ecol. Appl. 11, 914–927. doi: 10.1890/1051-0761(2001)011[0914:PFIOSF]2.0.CO;2
Pope, K. O., Pohl, M. E., Jones, J. G., Lentz, D. L., Von Nagy, C., Vega, F. J., et al. (2001). Origin and environmental setting of ancient agriculture in the lowlands of Mesoamerica. Science 292, 1370–1373.
Keywords: fire, pine-oak forest, Zea mais, early agriculture, Capsicum, Sporormiella, palaeoecocology, Maya
Citation: Harvey WJ, Nogué S, Stansell N, Petrokofsky G, Steinman B and Willis KJ (2019) Corrigendum: The Legacy of Pre–Columbian Fire on the Pine–Oak Forests of Upland Guatemala. Front. For. Glob. Change 2:44. doi: 10.3389/ffgc.2019.00044
Received: 19 July 2019; Accepted: 24 July 2019;
Published: 31 July 2019.
Edited and reviewed by: Tom Evans, Wildlife Conservation Society, United States
Copyright © 2019 Harvey, Nogué, Stansell, Petrokofsky, Steinman and Willis. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: William J. Harvey, d2lsbGlhbS5oYXJ2ZXlAc2VoLm94LmFjLnVr