 Sonia Meller
Sonia Meller Emmanuel Frossard
Emmanuel Frossard Marie Spohn
Marie Spohn Jörg Luster
Jörg Luster- 1Forest Soils and Biogeochemistry, Swiss Federal Institute for Forest, Snow and Landscape Research WSL, Birmensdorf, Switzerland
- 2Institute of Agricultural Sciences, Swiss Federal Institute of Technology ETH, Zurich, Switzerland
- 3Soil Ecology, University of Bayreuth, Bayreuth, Germany
Forests dominated by beech (Fagus sylvatica L.) cover large parts of Europe where they occupy a broad ecological niche in terms of soil fertility. This indicates a large potential to adapt to different soil conditions over long time periods. Recent changes in tree mineral nutrition across Europe raise the question to what degree beech can acclimate to changing soil conditions in the short term. In this study, we aimed at assessing the plasticity of root traits and rhizosphere properties of young beech trees from populations that are adapted to either high or low nutrient supply, when growing in soils differing in their fertility. We sampled beech saplings from two forest sites of contrasting nutrient supply, most distinctly in terms of phosphorus. We grew them for 2 years in rhizoboxes in mineral soil either from their own site or from the other site. We assessed the influence of the factors “plant origin” and “current soil” on root traits and rhizosphere properties. Fine root traits related to growth (biomass, length), architecture (branching), and morphology (diameter) responded strongly to the factor “current soil.” Provenance (factor “plant origin”) modified the response. The modifying effect was consistent with an influence of the plant status in those nutrients, which were not in sufficient supply in the soil. An additional genotypic difference in the sensitivity of the beech saplings to different soil nutrient supply could not be excluded. Fine root parameters normalized for length, mass, or volume (root tip density and frequency, specific root length and area, and root tissue density) did not differ among the treatments. Differences in percentage of mycorrhizal root tips and rhizosphere parameters related to phosphorus mobilization potential (pH, abundance of organic acid anions, and phosphatase activity) were small and mainly determined by the “current soil.” Provenance had only a minor modifying effect, possibly due to differences in the ability of the plants to transfer carbon compounds from the shoot to the root and the fungal partner. Our results indicate a high plasticity of young beech trees to adapt their root system to different soil nutrient supply, thereby also taking into account internal nutrient reserves.
Introduction
Forests dominated by European beech (Fagus sylvatica L.) cover large parts of Europe where climatic conditions are suitable (Durrant et al., 2016). Considering the distribution of important tree species in temperate forests, beech belongs to a group that is relatively insensitive to differences in soil nutrient supply (Walthert and Meier, 2017) and thus occurs at a wide range of soil chemical properties including strongly acid to alkaline pH, as well as low to high N and P availability (Leuschner et al., 2006). This indicates a large potential of beech to adapt to different soil nutrient supply over long periods of time. Considering the recent changes in tree mineral nutrition across Europe (Jonard et al., 2015), the question arises to what degree beech can acclimate to changing soil conditions in the short term. Of particular concern is phosphorus (P), since plant-available P occurs at only low concentrations in the soil solution, while most P is present in unavailable forms adsorbed to reactive surfaces of the soil solid phase or is bound in minerals or soil organic matter (Hinsinger, 2001).
Root–soil interactions have been shown to play a major role in adaptation to given soil conditions in relation to nutrient acquisition. Such interactions can include alterations of root growth, architecture and morphology, formation of mycorrhizae, and root exudation affecting nutrient availability in the rhizosphere (Richardson et al., 2009).
Root growth, architecture and morphology can be highly plastic in response to soil nutrient availability (Hodge et al., 2009). In particular, alterations in reaction to low soil availability of P and major nutrient cations (Mg, K, and Ca) that have been found across a large range of plant species include inhibition of primary root growth and promotion of lateral root growth (Gruber et al., 2013; Niu et al., 2013). Relations are not so clear in the case of N, which is demonstrated by maximum root length and branching of the model plant Arabidopsis at intermediate N limitation (Kiba and Krapp, 2016), and effects on branching depend in addition on the chemical form of N (nitrate or ammonium). While these mechanisms have been well established for crops, evidence for trees is scarce, and assessing effects of nutrient availability in the field is often made difficult by interaction with other soil properties such as texture (Weemstra et al., 2017), environmental factors such as the availability of water (Hertel et al., 2013) or light (Minotta and Pinzauti, 1996), or stand age (Finér et al., 2007). Preferential root proliferation in nutrient-enriched patches and layers has been observed frequently (Hodge, 2006; Chen et al., 2016). Particularly important for trees growing in nutrient-poor soils in temperate forests is preferential exploration of the topsoil, including the organic surface layer (Borken et al., 2007; Hauenstein et al., 2018).
Uptake via the mycorrhizal pathway is of major importance for N and P nutrition of trees in temperate zones (Plassard and Dell, 2010; Chalot and Plassard, 2011). While fertilization with P often decreases mycorrhization in inoculation experiments (Garbaye and Wilhelm, 1985; Kazantseva et al., 2009), under field conditions, relationships between soil P availability and measures of mycorrhization or mycorrhizal P uptake are less clear and might differ seasonally (Yang et al., 2016; Spohn et al., 2018). In most studies on the effect of N availability, mycorrhizal colonization increased with decreasing N availability (e.g., Brunner and Brodbeck, 2001; Sun et al., 2010); however, under natural concentration gradients, higher colonization was also found at lower C:N ratios (Hawkins et al., 2015). Under field conditions, the effects of both N and P supply on mycorrhization have to be considered as well (e.g., Bahr et al., 2013). Studies assessing host effects on ectomycorrhizal fungi provide a variable picture on the degree to which trees can actively shape the rhizosphere fungal community (Ishida et al., 2007; Lang et al., 2017; Spohn et al., 2018). There can also be competition between fungal partner and host plant, leading to limited nutrient transfer to the host (e.g., Simon et al., 2017).
On a small scale in the rhizosphere, root exudation can lead to an increase in the abundance of compounds that potentially increase the bioavailability of P, including protons, low-molecular-weight organic acid anions, and phosphatases (Hinsinger et al., 2011). However, under field conditions, it is often difficult to differentiate between the different sources of these compounds. Organic acid anions and phosphatases can be produced and released to the soil by roots, mycorrhizal hyphae, and free-living microorganisms (Gianfreda and Ruggiero, 2006; Oburger et al., 2011; Plassard et al., 2011). Therefore, also little is known to which degree plants are able to influence the P mobilization potential in their rhizosphere directly via root exudation and/or indirectly via stimulating microbial activity and growth or shaping the soil microbial community. For example, root exudates can stimulate P mineralization by heterotrophic bacteria in the rhizosphere (Spohn et al., 2013). Phosphatase activity (PA) in soil is often linked to soil P availability (Marklein and Houlton, 2012; Hofmann et al., 2016), but this relation can be masked, e.g., by the generally strong correlation with soil organic matter content (Nannipieri et al., 2011). The root exudation of organic acid anions may be induced by a low P nutritional status of the plant, as has been shown for crops (Hinsinger, 2001). However, it can also be a reaction to other conditions such as high Al concentrations in acid soils (Richardson et al., 2009) or be part of constitutive release of excess carbon (Heim et al., 2001; Eldhuset et al., 2007). Although proton exudation by roots can be induced by P deficiency (e.g., Shahbaz et al., 2006), alteration of rhizosphere pH often depends on the form of mineral nitrogen taken up by the plant (Riley and Barber, 1971; Hinsinger, 2001).
Signaling of the plant nutritional status has been shown to be involved in controlling root development, initiating mycorrhizal symbiosis, and producing and exuding mobilizing substances (Chalot and Plassard, 2011; George et al., 2011; Niu et al., 2013; Xuan et al., 2017). Thus, differences in this status could affect the root response of beech populations that are adapted to sites differing in resource availability, when growing in soils with different nutrient supply. However, genotypic differences related to adaptation to specific site conditions could also be involved. Beech populations across central Europe have been shown to be genetically closely related in terms of neutral markers, such as microsatellite loci, but to differ in genes related to adaptive traits (e.g., Buiteveld et al., 2007). Nevertheless, genotypic diversity has often been found to be larger within than among populations, also including also adaptive traits (e.g., Cuervo-Alarcon et al., 2018). In contrast to the lack of studies on acclimation to changes in nutrient supply, the ability of beech populations from sites with different climatic conditions to acclimate, in the short term, to increased drought frequency has recently received much attention (Meier and Leuschner, 2008; Cuervo-Alarcon et al., 2018). Specifically, Meier and Leuschner (2008) found that while root traits, such as relative fine root growth and turnover, of beech populations from sites differing in precipitation responded generally strongly to drought treatment, the effect of provenance was small. Aboveground adaptive traits related to resource acquisition such as photosynthetic activity have been considered in the so-called “resource economics” framework (Craine, 2009). This differentiates between “acquisitive” and “conservative resource strategies” exhibited by plants growing at resource-rich and resource-poor sites, respectively. However, Weemstra et al. (2016) concluded from their review that there is little evidence for root physiological and morphological traits being indicative of specific nutrient acquisition strategies. Specifically, fine root diameter had often been found to correlate with root longevity and therefore been considered a respective potential belowground indicator. Taking together the information on genotypic relations among beech populations, their acclimation to drought, and relation between belowground plant traits and nutrient acquisition strategies, we do not expect strong genotypic provenance effects on root traits during acclimation to a different soil nutrient supply.
In this study, we aimed at assessing the plasticity of root traits and rhizosphere properties of young beech trees from populations, which are adapted to either high or low nutrient supply, when growing in soils differing in their fertility. To this end, we sampled beech saplings from two forest sites differing most distinctly in the supply P. We grew the saplings in mineral soil either from their own site or from the other site. In all four experimental treatments, we assessed the influence of the factors “plant origin” and “current soil” on root growth, architecture and morphology, mycorrhization, and the occurrence of P mobilizing compounds in the rhizosphere. In this “cross-exchange” approach, the factor “current soil” was considered to reflect differences not only in physicochemical soil properties but also in microbial communities adapted to these properties. Potential confounding effects by differences in so-called “plant-soil feedback” (reviewed, e.g., by Bever et al., 2012), are expected to be small when comparing different populations of the same plant species (Wagner et al., 2011; Gundale et al., 2014).
We hypothesized, first, that the assessed root traits and rhizosphere parameters are determined mainly by the factor “current soil.” We hypothesized, second, that the factor “plant origin” modifies the effects of the soils and that the modifying effect can be attributed mainly to differences in the plant nutritional status.
Materials and Methods
Plant and Soil Materials
Plant and soil materials were collected at the core research sites of the priority programme 1685 “Ecosystem nutrition” of the German Science Foundation (DFG)1 in Unterlüss (Lower Saxony, Germany, LUE) and Bad Brückenau (northern Bavaria, Germany, BBR). The sites both sustain mature mono-specific beech stands, but differ in environmental conditions and soil properties (Lang et al., 2017), as summarized in Supplementary Table S1. The site LUE has a drier climate than the site BBR. The soil at BBR contains more N and P than the one at LUE in terms of total element stocks, as well as concentrations in organic surface layer and mineral soil. In particular, the P concentration in the mineral soil is much higher at BBR, whereas the organic surface layer is an important source of N and P at LUE. Both organic surface layer and mineral soil at LUE are more acidic and exhibit a lower base saturation than their BBR counterparts. Furthermore, the mineral soil at BBR has a loamy texture with a higher cation exchange capacity than the sandy mineral soil at LUE. Mature beeches are of similar age and height at both sites, however, their average diameter is much smaller at LUE.
Saplings of beech (F. sylvatica L.) of similar size were collected during their dormancy period in December 2014 and stored at 4°C with their roots embedded in soil until planting. Based on tree-ring counting, they were between 12 and 15 years old at the end of our experiment (Meller et al., 2019). Total N contents in various plant compartments were determined by combustion using an elemental analyzer (NC 2500, Carlo Erba Instruments), and total P, Mg, K, and Ca contents were determined by inductively coupled plasma optical emission spectrometry of digests (for details, see Meller et al., 2019).
Soil materials were taken from the Bh horizon in LUE and the uppermost part of the Bv horizon in BBR. This choice represented a compromise between root density within the soil profile—and thus potential importance for nutrient uptake—and organic matter content of the material being sufficiently low to not interfere with the assessment of rhizosphere properties. Soils were air-dried at 15°C, sieved to 4 mm, and homogenized. Plant residues were removed. Selected physical and chemical properties of the soils are summarized in Table 1 and were mostly determined as described by Meller et al. (2019). Sequential P extraction was performed according to Hedley et al. (1982) as modified by Tiessen and Moir (2006). In Table 1, resin exchangeable inorganic P (Presin), inorganic P (Pi) in various extracts (0.5 M NaHCO3, 0.1 M NaOH before and after sonication, 1 M HCl, concentrated HCl), and organic P (Porg) in the NaHCO3 and NaOH extracts are shown. The soil material from BBR exhibited a finer texture; a higher pH; a higher content in exchangeable nutrient cations; a higher organic carbon content; lower Corg/Ntot, Corg/Porg, and Ntot/Porg ratios; and much higher concentrations of all inorganic and organic P fractions than the material from LUE. On the other hand, the LUE soil exhibited a higher proportion of Porg than the BBR soil, and the base cation-to-Al ratio was similar in both soils.
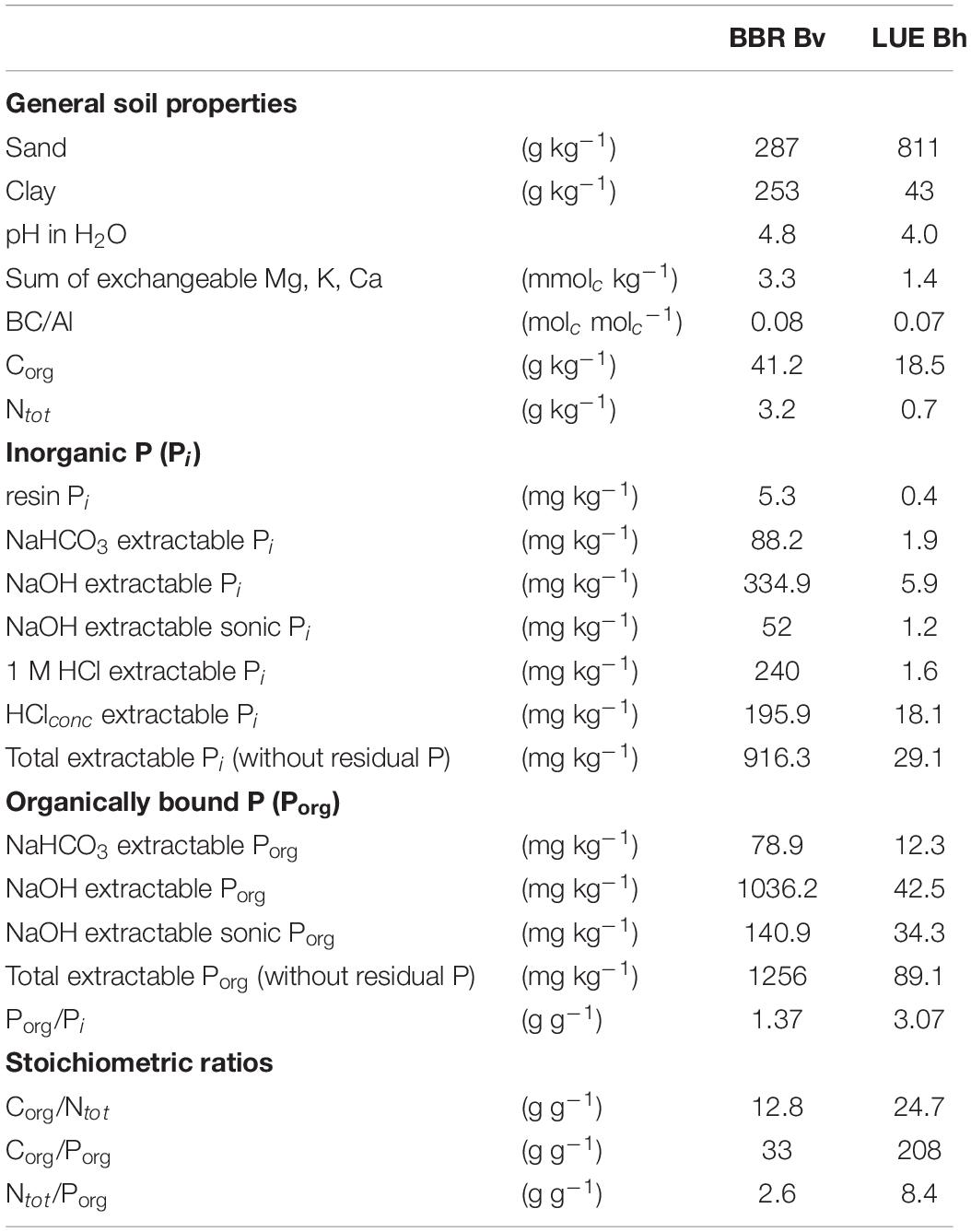
Table 1. Properties of the soil materials from the forest sites Bad Brückenau (BBR, Bv horizon) and Unterlüss (LUE, Bh horizon); BC/Al refers to the ratio between the sum of exchangeable base cations (Mg, K, Ca) and exchangeable Al; inorganic P (Pi) and organic P (Porg) used in element ratios refer to the respective total extractable fractions; data are from Meller et al., 2019 except for the concentration of P fractions.
Experimental Setup
In April 2015, rhizoboxes were set up with beech saplings planted either in the soil from their site of origin or in the contrasting soil from the other site. In a completely randomized design, each treatment was replicated six times. The rhizoboxes had inner dimensions of 60 cm × 25 cm × 1.5 cm. They consisted of PVC walls and a removable transparent front plate made of polymethyl methacrylate. The soil was filled into the boxes at a bulk density of 1.2 kg/dm3. After 1 week of soil conditioning under irrigation as described below, the saplings were planted. At this time point, saplings possessed up to 10-cm-long tap roots of 0.5–1.5 cm diameter but almost no fine roots, which presumably had died off during storage. The roots were washed with tap water to remove sticking soil, and approximately 2 cm of tap root was cut to stimulate new root formation. For each tree, the front plate of one rhizobox was opened, the roots pressed into the soil, and the front plate closed again. Rhizoboxes were placed in a greenhouse with temperature control (day, 22 ± 2°C; night, 18 ± 2°C), natural light, and shading from the direct sun. Since shading with movable blinds was the only means for active cooling, at some days in summer, temperatures higher than 22°C occurred for short periods. The soil was kept dark by covering the rhizoboxes with black plastic foil, and to stimulate the formation of a quasi-planar root system along the front plate, the rhizoboxes were inclined at an angle of about 30°. Soil water potential in the rhizoboxes was kept at approximately -8 kPa by using irrigation tubes (“Rhizon irrigators,” Rhizosphere research products, Wageningen, The Netherlands), providing P-free artificial rain solution based on the composition of natural precipitation [2.1 μM K2SO4, 3.7 μM Na2SO4, 3.0 μM CaCl2, 4.4 μM CaSO4, 1.9 μM MgCl2, 26.4 μM NH4NO3, and 2.0 μM Ca(NO3)2; Holzmann et al., 2016]. During summer, additional periodic irrigation from the top was needed to compensate for high evapotranspiration. At the end of the first growing season (end of September 2015), the rhizoboxes were placed outside of the greenhouse, but protected by a roof, to induce dormancy. In November 2015, they were moved to a dark cold room at 4°C and periodically irrigated with artificial rain from the top. End of March 2016, after the last frost, the rhizoboxes were moved first to the protected area outside of the greenhouse, and in May, after appearance of the first leaves, back into the greenhouse with temperature control set to the same conditions as in the year before.
Measurement of Rhizosphere Parameters
In August 2015 and 2016, non-destructive and minimally invasive membrane-based methods were applied to the surface-exposed roots after carefully removing the front plate. For each rhizobox, all measurements, as described in detail in the following paragraphs, were performed on the same day in the following order: pH (8–9 a.m.), exchangeable anions (10 a.m. to 1 p.m.), and potential PA (2–4 p.m.). For this, the rhizoboxes were laying horizontally on their back side. Two or three rhizoboxes were assessed per day within 2 weeks, and the order of replicates among the four treatments was selected randomly. Five replicates per plant/soil combination were selected. Since some of the LUE saplings growing in LUE soil died during the first months after planting, replication was only 3 in this case.
pH Distribution
The pH in the rhizosphere was mapped using prototypes of planar optodes with an optimal measurement range between pH 3.5 and 5.0 and signal detection by a VisiSens camera (PreSens GmbH, Regensburg, Germany). The optodes consisted of a 10-μm-thick foil composed of a proton-permeable polymer matrix, with a pH-sensitive and a pH-insensitive dye fixed in this matrix. Upon contact with soil or buffer solution, pH is measured as the ratio between the fluorescence of the pH-sensitive indicator dye and the fluorescence of the pH-insensitive reference dye (for more details of the principle, refer to Blossfeld and Gansert, 2007). For this, photos taken with the VisiSens camera were separated into a red and green channel corresponding to the fluorescence of the sensitive and non-sensitive fluorophores, respectively. For calibration, small pieces of optodes were equilibrated in a buffer of pH 4 overnight and then placed for 30 min in buffered standard solutions with pH values ranging from 3 to 5. Five to ten measurements of each membrane were averaged. The calibration curve was sigmoidal with a quasi-linear range and thus optimum sensitivity, between pH 3.5 and 4.5. For soil measurements, optodes of about 1 × 2 to 2 × 2 cm2 were equilibrated in pH 4 buffer overnight, applied to the terminal part of at least three newly grown roots per rhizobox including the surrounding soil, and left to equilibrate for 15 min, protected with a small piece of clear acrylic glass. In initial tests, an equilibration time of 15 min was found to be optimum to reach stable conditions and to avoid artifacts caused by drying of the membrane at longer application times. Then, photos were taken with the VisiSens camera using a cylindric aluminum spacer of 6 cm length between camera and acrylic glass. In order to cover its whole area, several overlapping partial areas of each optode were measured, and photos were subsequently merged into one image using Adobe Photoshop (Supplementary Figure S1). After use, optodes were rinsed with deionized water and stored in a buffer of pH 4 in a dark plastic bag at 4°C, because they are damaged by drying and re-wetting and exposure to light. After overlaying the pH maps with a mask for the location of the roots, using Adobe Photoshop, two zones were defined: root surface (values in the middle of the root) and bulk soil (>2 mm from root edge). Averaged values of bulk soil and root surface per rhizobox were used in the statistical analysis.
Nutrient and Organic Acid Anions
Nutrient and organic acid anions (nitrate, phosphate, sulfate, oxalate, and citrate) were collected from the rhizosphere using anion exchange membranes (AEMs; Shi et al., 2011). Strips of AEMs (2 cm × 0.5 cm; No. 55164 2S, BDH Laboratory Supplies, United Kingdom) were soaked in deionized water for 24 h and then converted into HCO3– form by equilibration with 2.2 ml of various agents per cm2 as follows: (i) for 10 min with 0.5 M HCl, (ii) twice for 1 h with 0.5 M NaCl, and (iii) three times for 30 min with 0.5 M NaHCO3. In between and at the end, the AEMs were rinsed with deionized water and stored in deionized water at 4°C until use. The membranes were applied to at least three newly grown roots including tip, elongation zone, side roots, and the respective rhizosphere per rhizobox for 3 h, covered with a plastic sheet to keep them moist during this time. After collection, the AEMs were rinsed with deionized water to remove sticking soil and extracted for 3 h with 0.3 ml of 1.75 M HCl in 2 ml Eppendorf tubes (opened periodically to release an excess of CO2 produced) using an end-over-end shaker at room temperature. The extracts were measured with ion chromatography (Thermo Scientific DIONEX ICS-3000 with InGuard Ag and Na Column 9∗24 mm, an Ultratrace Anion Concentrator Column and a conductivity detector). Data from all membranes per rhizobox were averaged.
Potential PA
Spatial distribution of potential PA in the rhizosphere and bulk soil was mapped using zymography as developed by Spohn and Kuzyakov (2013) with slight modifications. Polyamide membranes (pore size 0.45 μm, Sartorius Stedim Biotech GmbH, Goettingen, Germany) were coated with 4-methylumbelliferyl phosphate (MUF-P, Sigma-Aldrich) by soaking in a 12 mM solution of this substrate in 10–4 M HCl (unbuffered solution with pH similar to soil pH) directly before application. Membranes of approximately 150 cm2 and varying shapes—to match the roots—were applied to newly grown long roots including the surrounding soil with a 1 mm protective layer of agarose gel (in 10–4 M HCl) between membrane and soil for 20 min. After incubation, the membrane was exposed to UV light (366 nm) in a dark chamber (UV cabinet camag, Muttenz, Switzerland) to visualize the fluorescence of the reaction product (4-Methylumbelliferone, MUF). Images (RGB) of the membrane were taken from a fixed distance of 28 cm using a Nikon D3200 Camera with an AF-S Nikkor 18–55 mm lens. Of all zymograms, several images were taken with different exposures. For quantification, an image without overexposure was selected and compared with the image of a series of MUF standards taken with the same exposure. Standards were prepared by soaking small pieces of the membrane in solutions of MUF in 10–4 M HCl with concentrations of 0, 35, 70, 130, and 200 μM MUF, yielding a linear calibration curve. The amount of MUF per unit area was calculated based on the amount of solution taken up and the surface of the membrane. The images of the standard curve and zymograms required further processing for which the intensities of the green channel of the RGB images were used without conversion (Image J version 1.18; Schneider et al., 2012). First, a correction was made for variations in light reaching the camera sensor. In our setting, this variation had a circular shape with a maximum near the center of the image and decreasing linearly toward the peripheries. Maximum and gradient of the corresponding function were defined based on spatial variations in light reflection from the homogeneous table background, which was part of each image. For this, we wrote a custom R [version 3.1.2 (2014-10-31)] code. Then, the intensities of individual pixels of zymograms, membranes with unreacted substrate, and standards were scaled based on this function. Second, the zymograms were corrected for background fluorescence of unreacted substrate on a piece of membrane that was not placed in contact with the soil but photographed in the same image. Third, we overlaid the corrected zymograms with masks representing the root distribution. For this, visible roots in a photograph of the open rhizobox were traced manually using Adobe Photoshop. This drawing, together with markings for the edges of the rhizobox was laid over a photograph of the rhizobox with substrate-soaked membranes applied. From this, masks were created showing root distribution and markings for the edges of the membranes. Finally, the mirrored masks were laid over the zymograms and the membrane markings in the masks aligned with the membranes in the zymogram. Intensities assigned to pixels on the root surface and in the bulk soil (areas > 2 mm from the edge of a root mask) were ranked and calibrated using a standard curve obtained as described above. For each rhizobox value, median values of PA for root and bulk soil of all individual membranes applied to this box were averaged.
Root Morphology
The whole root systems of the saplings were excavated at the end of the experiment (August 2016), rinsed with tap water, and then scanned and analyzed for morphological characteristics using the WinRHIZO software employing 0.1 mm steps for root diameter. Specific root length (m g–1) was calculated as length of fine roots (diameter ≤ 2 mm) divided by their dry mass (M), specific root area (m2 kg–1) as surface of fine roots divided by M, and root tissue density (RTD) (kg m–3) as M divided by volume of fine roots. Root tip density and frequency are expressed as number of root tips per unit length and per unit dry mass of fine roots, respectively. Branching was calculated as the number of forks divided by the total length of fine roots. In order to determine the mycorrhization of root tips for a given sapling, the whole fine root system was cut into small pieces, which were mixed in a bowl filled with water. Three replicate subsamples of 30 ml of root suspension were transferred to a petri dish each. The number of mycorrhized and non-mycorrhized root tips was counted for each replicate using a binocular, and the percentages of mycorrhized root tips were averaged.
Carbon, Nitrogen, and Phosphorus in Soil Microbial Biomass
At the end of the experiment, bulk soil and rhizosphere soil—defined as soil sticking to the roots after gentle shaking—were collected from the rhizoboxes. Microbial biomass C and N (Cmic, Nmic) were determined using chloroform fumigation–extraction (Brookes et al., 1985), following the detailed instructions by Voroney et al. (2006). Specifically, soils were fumigated for 24 h and extracted with 0.5 M K2SO4 at a soil-to-extractant ratio of 1:5. Organic C and total N in the extracts were measured using a TOC/TN analyzer (Shimadzu TOC-V). The measured values for Cmic and Nmic were used without factors accounting for soil-specific recovery. Microbial biomass P (Pmic) was determined using the hexanol fumigation method introduced by Kouno et al. (1995) as described by Bünemann et al. (2004). Briefly, slurries of 0.5 g soil in 30 ml of deionized water with and without addition of 1 ml of hexanol were incubated together with an AEM in bicarbonate form (for preparation see above) for 16 h. Then, the AEMs were removed and extracted with 0.5 M HCl. Phosphate in the extracts was analyzed colorimetrically using malachite green (Ohno and Zibilske, 1991) and a phosphate standard curve in 0.5 M HCl. The re-sorption of P released by hexanol fumigation was accounted for by employing a third replicate incubation with addition of a suitable P spike as described by Bünemann et al. (2004).
Statistical Analysis
All analyses were performed in R [R version 3.1.2 (2014-10-31)]. Differences among individual treatments were assessed by using analysis of variance (ANOVA) followed by a Tukey post hoc test. In addition, results were tested for an influence of the factors “plant origin” and “current soil” as well as their interactions using two-way ANOVA. The following variables were log transformed to meet the requirements of the ANOVA: concentrations of resin-extractable phosphate in the first and second season, exchangeable nitrate in the first season, foliar N/Ca ratios in the first and second season, foliar K concentrations and N/P ratios in the second season. Because of unequal group sizes, we used the ANOVA model from the R package “car” that employs a “type II” test of the factors (for details of the definition of “type II” testing in “car,” refer to the respective reference manual)2.
Results
Nutritional Status of the Beech Saplings
During the experiment, the nutritional status of the beech saplings underwent a drastic change from mainly reflecting their site of origin in the first growing season to a major influence of the experimental soil in the second growing season. Important exceptions were K with a strong additional influence of the factor “current soil” already in the first season and P with a still strong additional effect of the factor “plant origin” in the second season. The respective data are provided in Table 2 and described in more detail in the following.
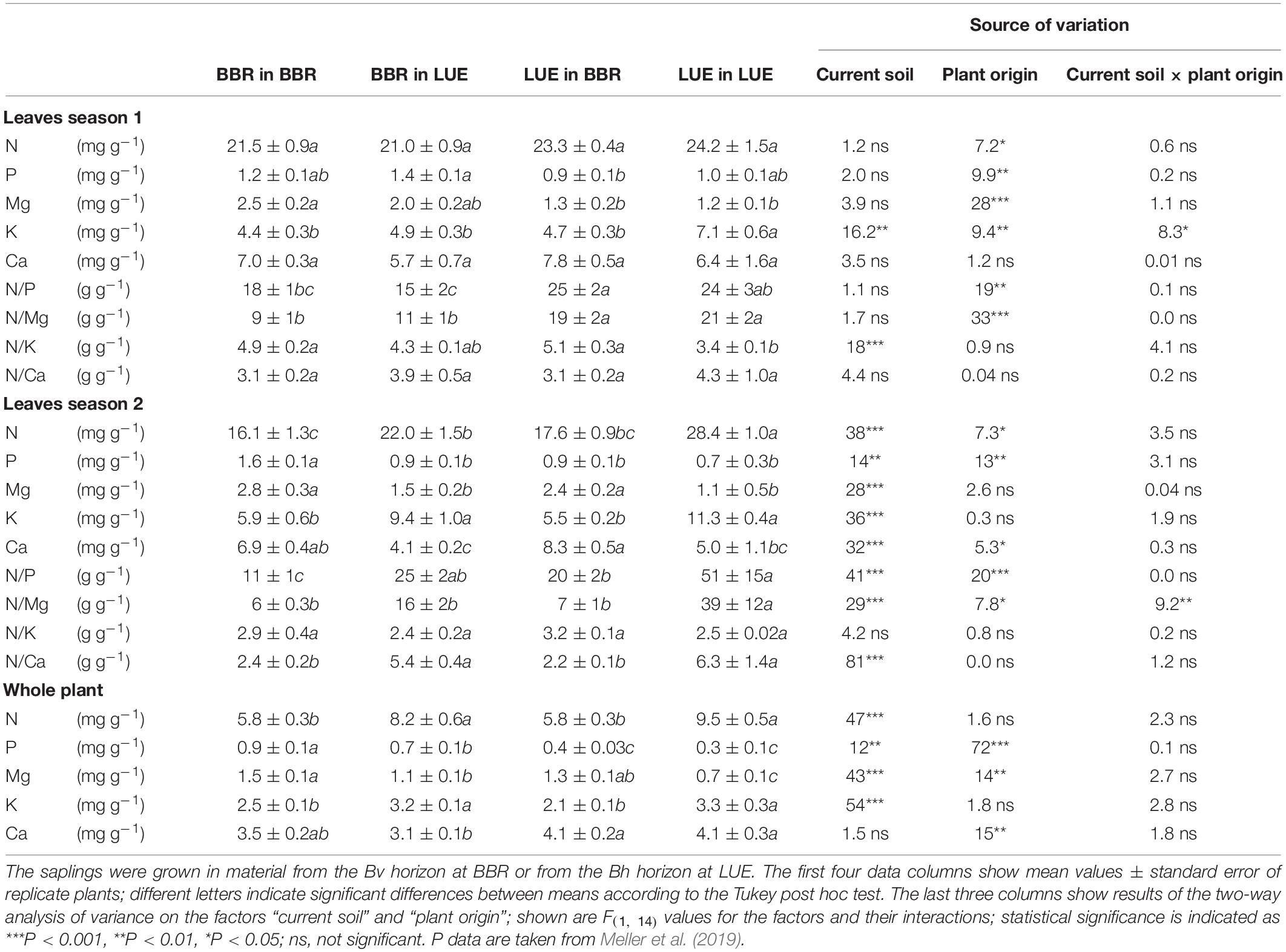
Table 2. Nutrient concentrations and ratios in full season leaves, as well as average nutrient concentrations in the whole plant for beech (Fagus sylvatica L.) saplings originating from the sites Bad Brückenau (BBR) and Unterlüss (LUE), respectively.
Foliar concentrations and concentration ratios in the first growing season revealed, irrespective of the soil, a higher supply of beech saplings from LUE with N and a lower supply with P and Mg than saplings from BBR. Compared to leaves of BBR plants, leaves of LUE plants exhibited about 10% higher N, 30% lower P, and 50% lower Mg concentrations, as well as about 50% higher N/P and twice as high N/Mg ratios. These differences were not all significant, but the corresponding effect of the factor “plant origin” was. The additional influence of the factor “current soil” on foliar K in the first growing season was stronger for saplings from LUE than from BBR. As a consequence, foliar K concentrations were about 40% higher for LUE plants growing in LUE soil than in all other treatments. These differences were significant, as were the effects of both experimental factors and their interaction. Similar differences were observed for the N/K ratios (35 and 15% lower for LUE and BBR saplings, respectively, when comparing growth in LUE with growth in BBR soil), but only the factor “current soil” was significant.
In the second growing season and irrespective of their site of origin, the leaves of saplings growing in the BBR soil exhibited a lower supply with N and K but a higher supply with Mg and Ca than leaves of saplings growing in the LUE soil. The corresponding effects of the factor “current soil” on foliar concentrations and concentration ratios were all significant except for N/K. The same applied to the respective differences between plants from the same origin growing in the different soils. The smallest difference was about 25% for N concentrations of BBR saplings, and the largest difference was about 100% for Mg concentrations of saplings from both sites of origin. Additional small but significant effects of the factor “plant origin” indicated that LUE plants were better supplied with N, and less supplied with Mg and Ca than BBR plants when comparing growth in the same soil. The respective differences in Ca concentrations were small and not significant. By contrast, differences in N concentrations and N/Mg ratios were large (30 and 250%, respectively) and significant, but only for growth in the LUE soil. The strong combined effects of both factors, “current soil” and “plant origin,” on foliar P were expressed by 50% lower P concentrations and five times higher N/P ratios for LUE than BBR plants growing in the soil from their own site, and values in between for the other two treatments.
The average concentrations of N, Mg, and K in the whole plant at the end of the experiment in the second growing season reflected the situation in the leaves and were also determined mainly by the factor “current soil.” By contrast, average P concentrations in the whole plant were still mainly determined by the factor “plant origin” and were about 50% lower in saplings from LUE than from BBR, when comparing growth in the same soil. In addition, average Ca concentrations were determined significantly by “plant origin” only with about 25% higher values in saplings from LUE than BBR.
Growth, Architectural, and Morphological Traits of Fine Roots
Fine root traits of the beech saplings at the end of the experiment related to size, branching, and diameter were mainly determined by the “current soil,” but with an additional effect of “plant origin,” which was more pronounced for growth in the LUE soil. The respective data are presented in Table 3 and Figure 1, and the results of the ANOVA are shown in Table 4.
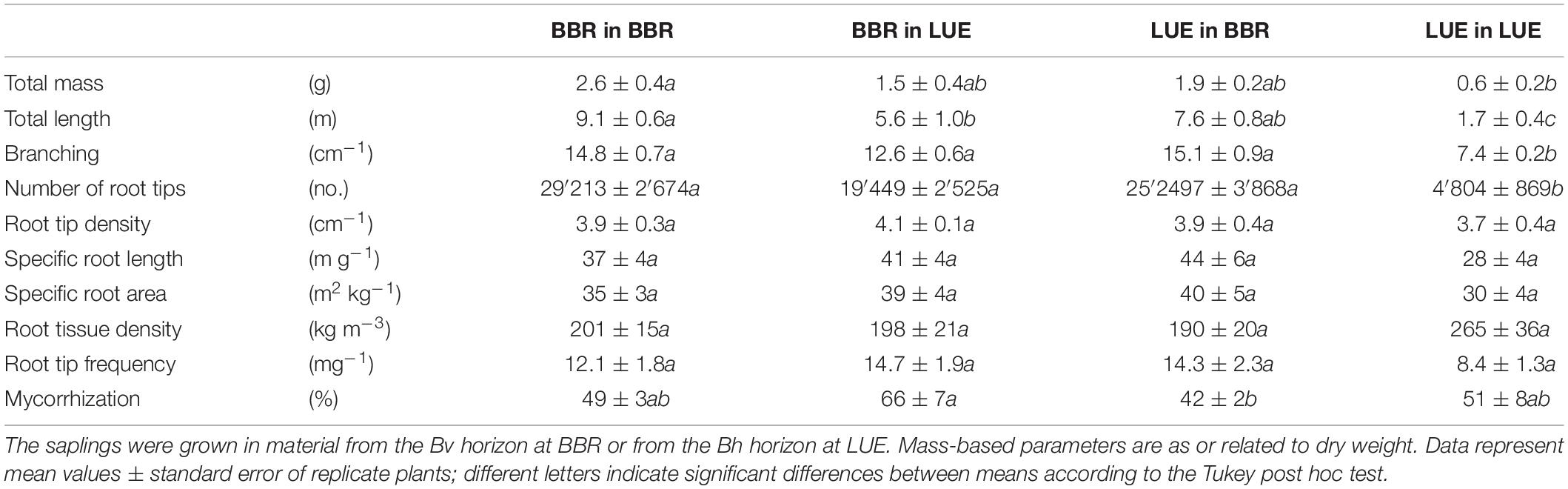
Table 3. Morphological traits of fine roots (<2 mm diameter), as measured during the second growing season in a rhizobox experiment with beech (Fagus sylvatica L.) saplings originating from the sites Bad Brückenau (BBR) and Unterlüss (LUE), respectively.
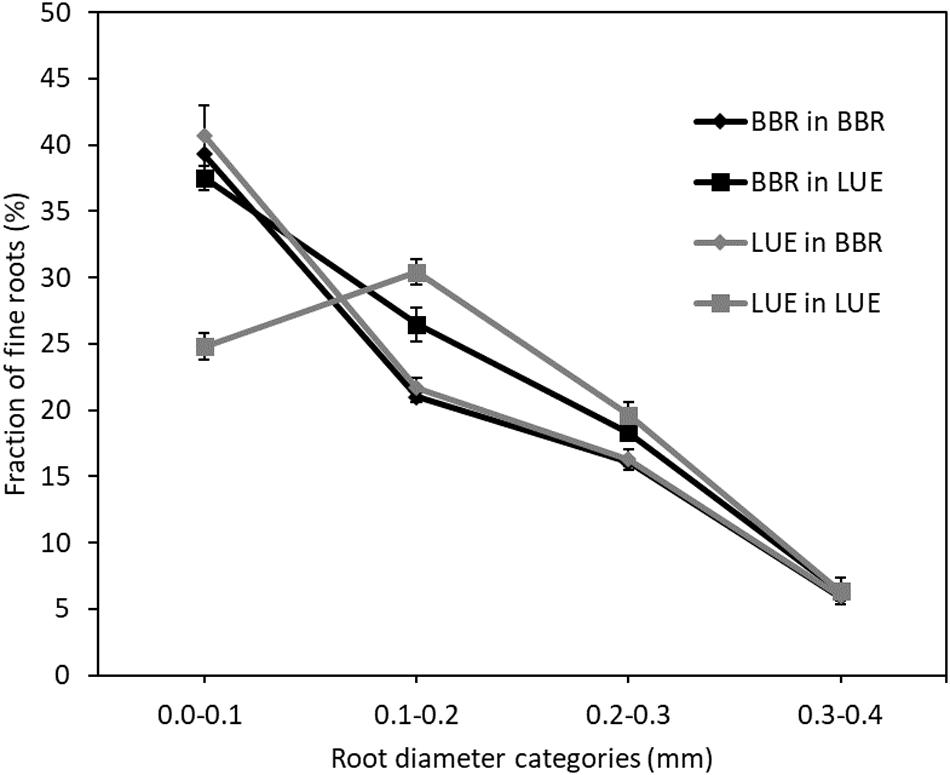
Figure 1. Fractions of fine roots in different diameter categories in (%) of total length of fine roots with diameter ≤ 2 mm, as measured in August of the second growing season in a rhizobox experiment with beech (Fagus sylvatica L.) saplings originating from the sites Bad Brückenau (BBR) and Unterlüss (LUE). The saplings were grown in material from the Bv horizon at BBR or from the Bh horizon at LUE. Data represent mean values ± SE for replicate saplings (n = 5, except n = 3 for LUE in LUE); lines serve as visual aid only.
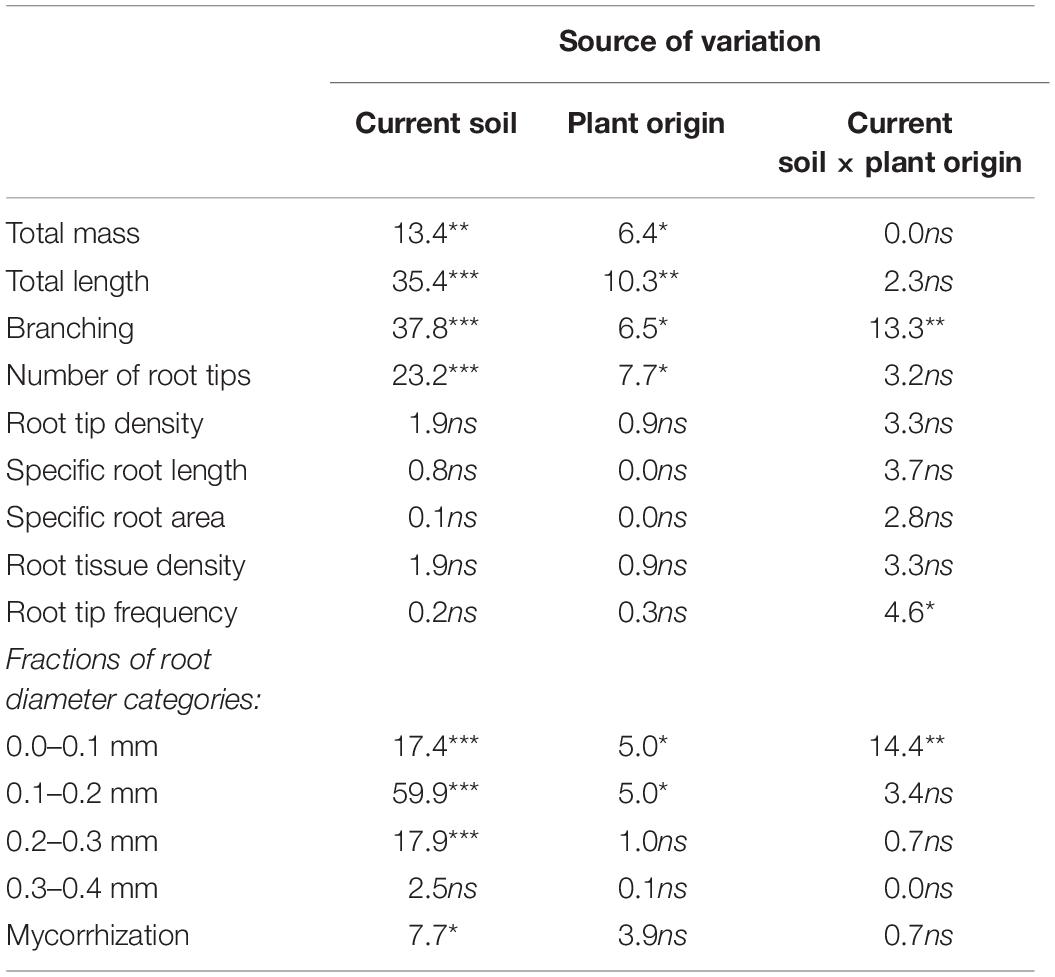
Table 4. Two-way analysis of variance for different fine root traits of beech (Fagus sylvatica L.) saplings, as measured in the second growing season of a rhizobox experiment; saplings originated from the sites Bad Brückenau (BBR) and Unterlüss (LUE) (factor “plant origin“) and were grown in material from the Bv horizon at BBR or from the Bh horizon at LUE (factor “current soil”); shown are F(1, 14) values for the factors and their interactions; statistical significance is indicated as ***P < 0.001, **P < 0.01, *P < 0.05; ns, not significant.
Irrespective of their origin, beech saplings exhibited a larger root system—in terms of mass, length, and number of root tips—in the BBR than the LUE soil. However, the respective trait differences were larger for saplings from LUE (three to five times higher values in BBR than LUE soil) than for saplings from BBR (about 50–75% higher values in BBR than LUE soil). The differences were not all significant, but the effects of both experimental factors were in all cases. Furthermore, beech saplings from LUE growing in soil from LUE had developed a fine root system with half the branching of plants in all other treatments. This led to a significant interaction between the two experimental factors.
Fine root diameters were identical for saplings from both sites of origin when growing in soil from BBR, with 40% of all fine roots being thinner or equal to 0.1 mm. By contrast, the distribution of root diameter categories differed between the plants from the two sites of origin for growth in the LUE soil. When compared to the growth in the BBR soil, LUE plants growing in LUE soil exhibited a lower proportion of the thinnest roots but higher proportions of roots with diameters between 0.1 and 0.3 mm. On the other hand, BBR plants growing in LUE soil exhibited the same proportion of thinnest roots, a higher proportion of roots between 0.1 and 0.3 mm, and a lower proportion of fine roots larger than 0.4 mm, when compared to growth in BBR soil.
In contrast to the aforementioned root traits, fine root parameters normalized for length, mass, or volume (root tip density and frequency, specific root length and area, and RTD) were equal for all four plant-soil combinations (Table 3) and the experimental factors did not exert any significant effect (Table 4).
Mycorrhizal Colonization of Fine Roots and Soil Microbial Biomass
After two growing seasons, mycorrhizal colonization of root tips did not differ strongly among the four treatments. Fine roots of saplings from the same site of origin tended to be more highly colonized when growing in the LUE than the BBR soil (Table 3). The corresponding effect of the factor “current soil” was significant (Table 4). On the other hand, in the same soil, BBR plants tended to have more highly colonized roots than LUE plants. The corresponding effect of the factor “plant origin” was weak [F(1, 14) = 3.9; P = 0.06].
Microbial biomass in bulk soil and rhizosphere at the end of the experiment was much higher in the BBR than the LUE soil with almost no effect of the origin of the beech saplings growing in the soil. Microbial biomass C (Cmic) in BBR soil was about 4.5 times, that in Nmic was about 6 times, and that in Pmic was about 3 times higher than in LUE soil (Table 5). The differences in Cmic and Nmic were significant, while those in Pmic were not. The corresponding effects of the factor “current soil” were highly significant for Cmic and Nmic, but only weakly significant for Pmic (Table 6). The Cmic:Nmic ratios in LUE soil tended to be a bit higher, the Cmic:Pmic ratios were up to three times lower, and the Nmic:Pmic ratios were two to three times lower than in the BBR soil. Considering both bulk soil and rhizosphere data for all plant-soil combinations, there was a weakly significant rhizosphere effect for Cmic (P = 0.02) and Nmic (P = 0.008) with slightly higher concentrations in the rhizosphere.
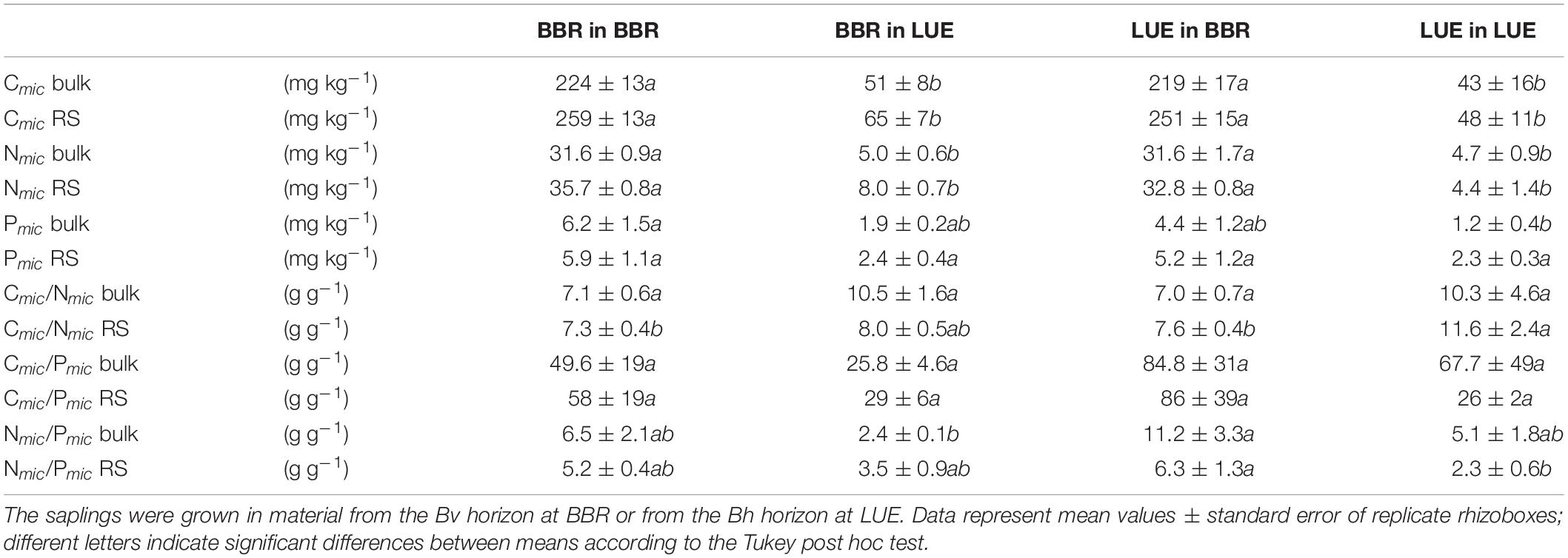
Table 5. Soil microbial biomass C, N, and P concentrations (per dry weight) in the bulk soil (bulk) and the rhizosphere (RS), as well as ratios in the rhizosphere, as measured during the second growing season in a rhizobox experiment with beech (Fagus sylvatica L.) saplings originating from the sites Bad Brückenau (BBR) and Unterlüss (LUE).
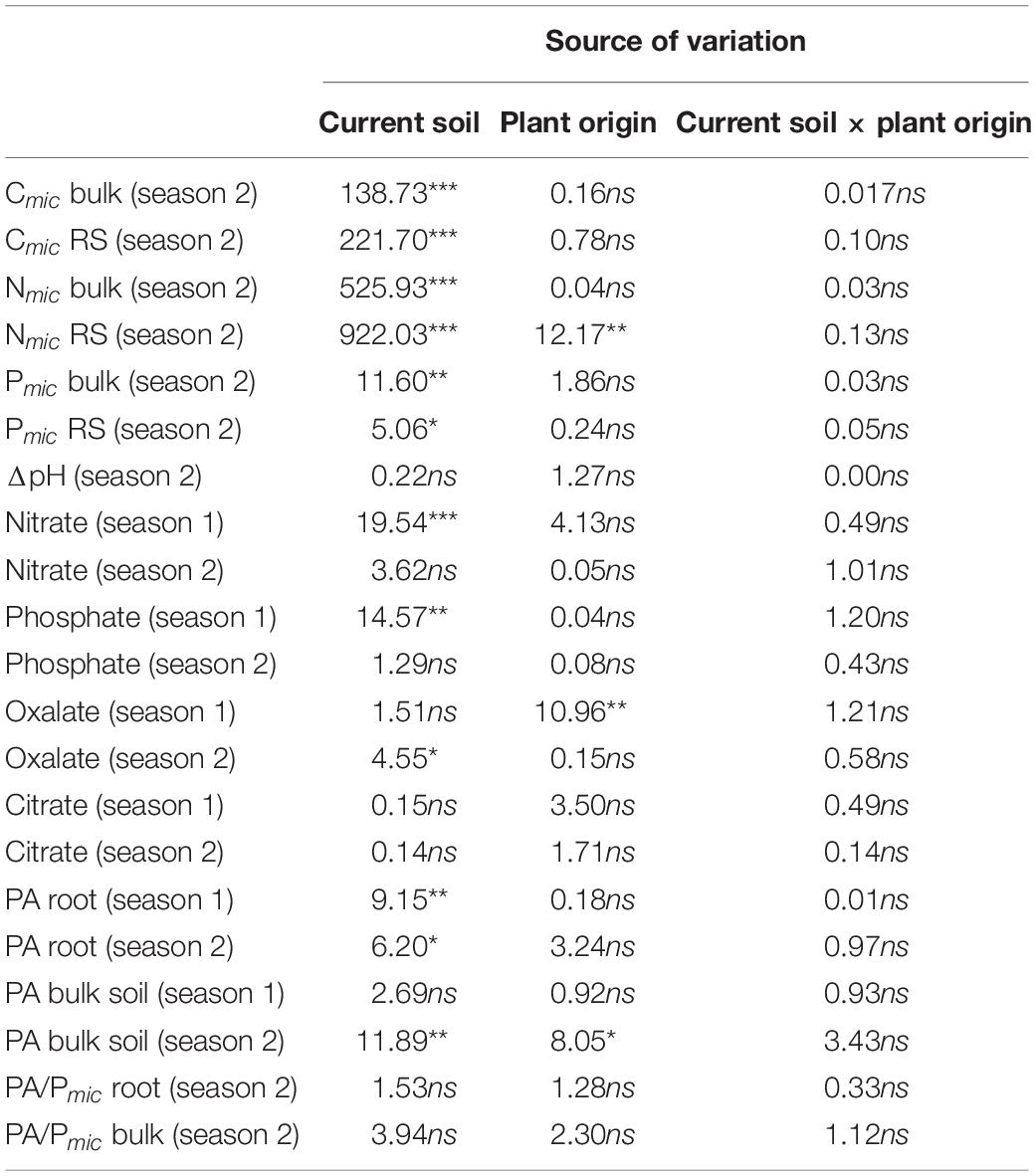
Table 6. Analysis of variance for microbial biomass C, N, and P (Cmic, Nmic, Pmic) in bulk soil (bulk) and rhizosphere (RS), the difference between pH on the root and in the bulk soil (ΔpH), resin extractable anions (nitrate, phosphate, oxalate, and citrate) in the rhizosphere, and phosphatase activity (PA) on the root and in the bulk soil, as measured in the first and/or second growing season of a rhizobox experiment with beech (Fagus sylvatica L.) saplings; saplings originated from the sites Bad Brückenau (BBR) and Unterlüss (LUE) (factor “plant origin”), and were grown in material from the Bv horizon at BBR or from the Bh horizon at LUE (factor “current soil”); shown are F(1, 14) values for the factors and their interactions; statistical significance is indicated as ***P < 0.001, **P < 0.01, *P < 0.05; ns, not significant.
Rhizosphere Properties
Although the variability of all measured rhizosphere properties was large and differences among treatments were mostly not significant, ANOVA revealed significant effects of the experimental factors. Most of the parameters differed between the first and the second growing season in terms of magnitude, effects of the two experimental factors, or both. The influence of the factor “current soil” was higher in most cases in both years except for a larger effect of “plant origin” on the abundance of low-molecular-weight organic acids in the first growing season. The respective data are shown in Figures 2–4, and the results of the ANOVA are shown in Table 6.
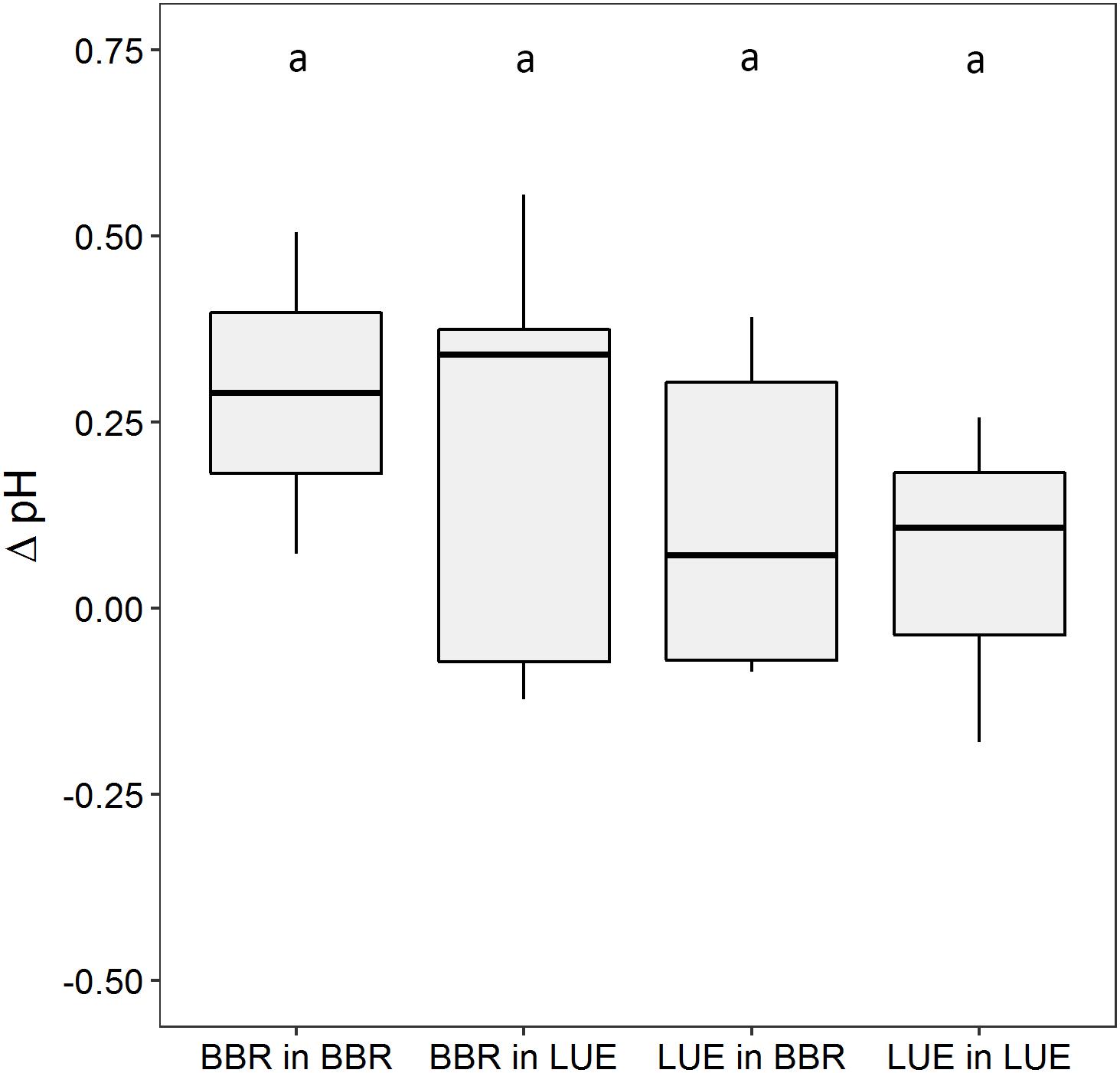
Figure 2. Difference between pH measured with optodes on the surface of roots and in the bulk soil (ΔpH) in August of the second growing season in a rhizobox experiment with beech (Fagus sylvatica L.) saplings originating from the sites Bad Brückenau (BBR) and Unterlüss (LUE). The saplings were grown in material from the Bv horizon at BBR or from the Bh horizon at LUE. Shown are box plots based on values for replicate rhizoboxes in the second growing season (n = 5, except n = 3 for LUE in LUE and n = 2 for BBR in BBR); different letters indicate significant differences between means according to the Tukey post hoc test.
In the second growing season, we observed small pH increases from bulk soil to the roots for all plant-soil combinations (Figure 2). However, effects of neither “plant origin” nor “current soil” were significant. Unfortunately, measurements performed in the first growing season provided no reliable data.
Resin-extractable nitrate in the rhizosphere exhibited different patterns in the two growing seasons (Figure 3A). Comparing growth in the BBR with growth in the LUE soil in the first year, nitrate in the rhizosphere of roots of BBR and LUE plants was about three and five times higher, respectively. The respective influence of the factor “current soil” was significant. In the second growing season, nitrate concentrations remained similar to the first year in the BBR soil but increased strongly in the LUE soil and were larger than in the BBR soil. The differences were not significant, though, and the effect of the factor “current soil” was weak [F(1, 14) = 3.6; P = 0.08].
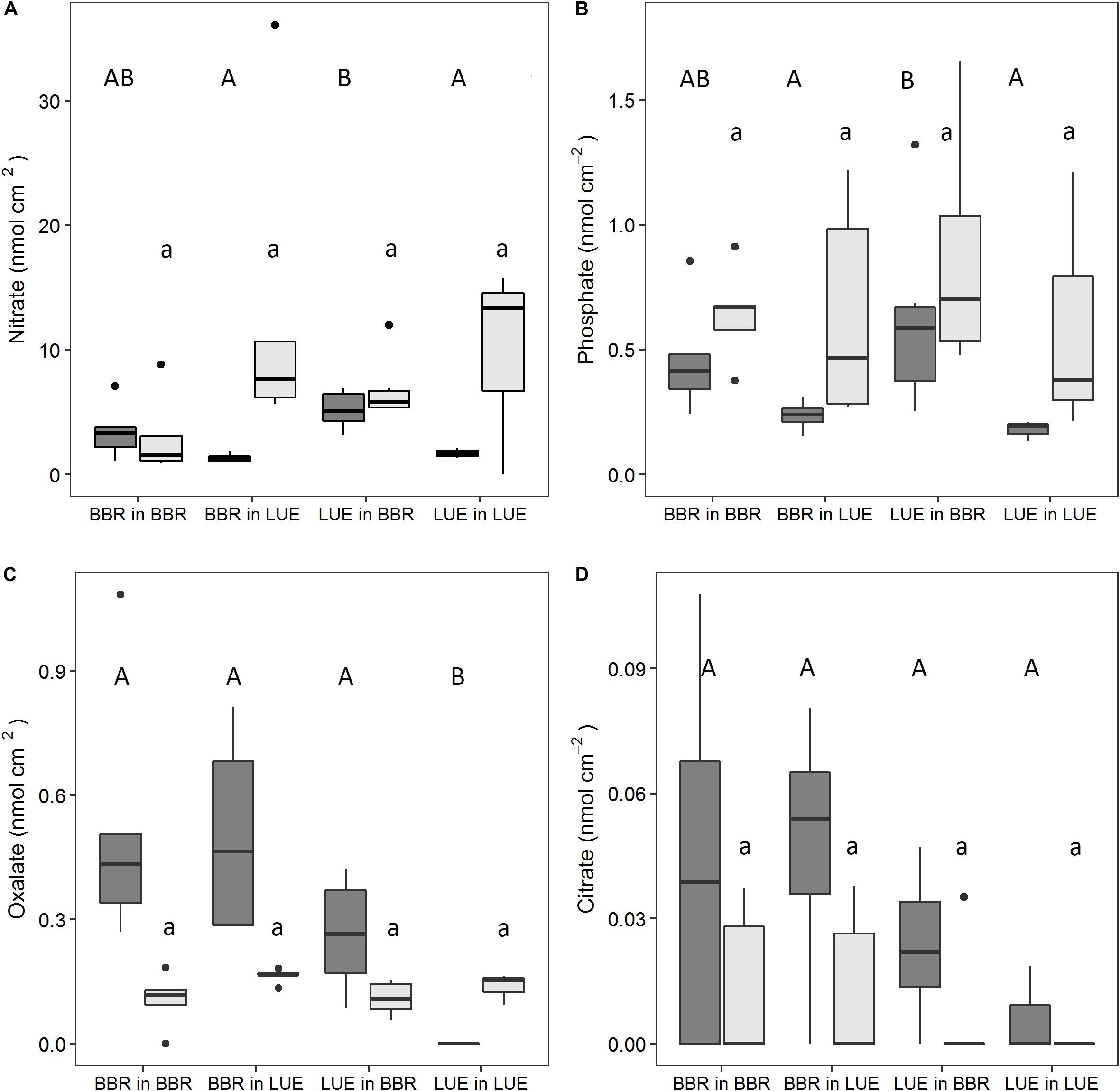
Figure 3. Nitrate (A), phosphate (B), oxalate (C), and citrate (D) collected with anion exchange membranes in the rhizosphere in a rhizobox experiment with beech (Fagus sylvatica L.) saplings originating from the sites Bad Brückenau (BBR) and Unterlüss (LUE). The saplings were grown in material from the Bv horizon at BBR or from the Bh horizon at LUE. Shown are box plots based on values for replicate rhizoboxes in August of the first (dark gray) and second (light gray) season (n = 5, except n = 3 for LUE in LUE); different letters indicate significant differences between means according to the Tukey post hoc test (uppercase letters, first growing season; lowercase letters, second growing season).
During the first growing season, resin-extractable phosphate was also significantly determined by the factor “current soil” with about two times higher concentrations in the BBR than the LUE soil (Figure 3B). In the second growing season, the pattern remained the same as a trend and concentrations were generally higher but also more variable than in the first growing season.
Oxalate and citrate were the only detectable resin-extractable low-molecular-weight organic acid anions, with oxalate occurring in about 10 times higher amounts than citrate (Figures 3C,D). In the first growing season, amounts of oxalate in the rhizosphere of the BBR plants were around 5 nmol cm–2, while amounts in the rhizosphere of LUE plants were about two times lower (BBR soil) or below the detection limit of 0.05 nmol cm–2 (LUE soil). This pattern led to a significant effect of “plant origin.” Citrate showed the same trends, but the effect of “plant origin” was not significant. In the second growing season, amounts of oxalate were generally lower than in the first year at around 0.15 nmol cm–2. There was no plant effect anymore, but a weak effect of “current soil” with a tendency to slightly higher amounts in the LUE soil. The few cases of detectable amounts of citrate suggest a similar, but not significant, effect of “plant origin” as in the first year.
In general, potential PA was highest on the main roots, intermediate on the side roots, and lowest in the bulk soil (see example in Supplementary Figure S2). During the first growing season, PA tended to be higher in the BBR than in the LUE soil, irrespective of the origin of the beech saplings (median values differed by about 15% in the bulk soil and by about 25% on the root; Figure 4A). This effect of “current soil” was significant for PA on roots but not for the bulk soil. In the second growing season, up to 30% lower PA were measured than in the first year, but the patterns among the treatments remained similar. The respective effect of “current soil” was significant for PA on roots and in the bulk soil. In contrast to the first year, the soil effect was overlaid by a weak effect of “plant origin“—albeit significant in the case of bulk soil—with a tendency to slightly higher values for saplings from BBR. In contrast to PA, PA per unit microbial P (PA/Pmic) exhibited a tendency toward higher values in the LUE soil, both on the root and in the bulk soil (Figure 4B).
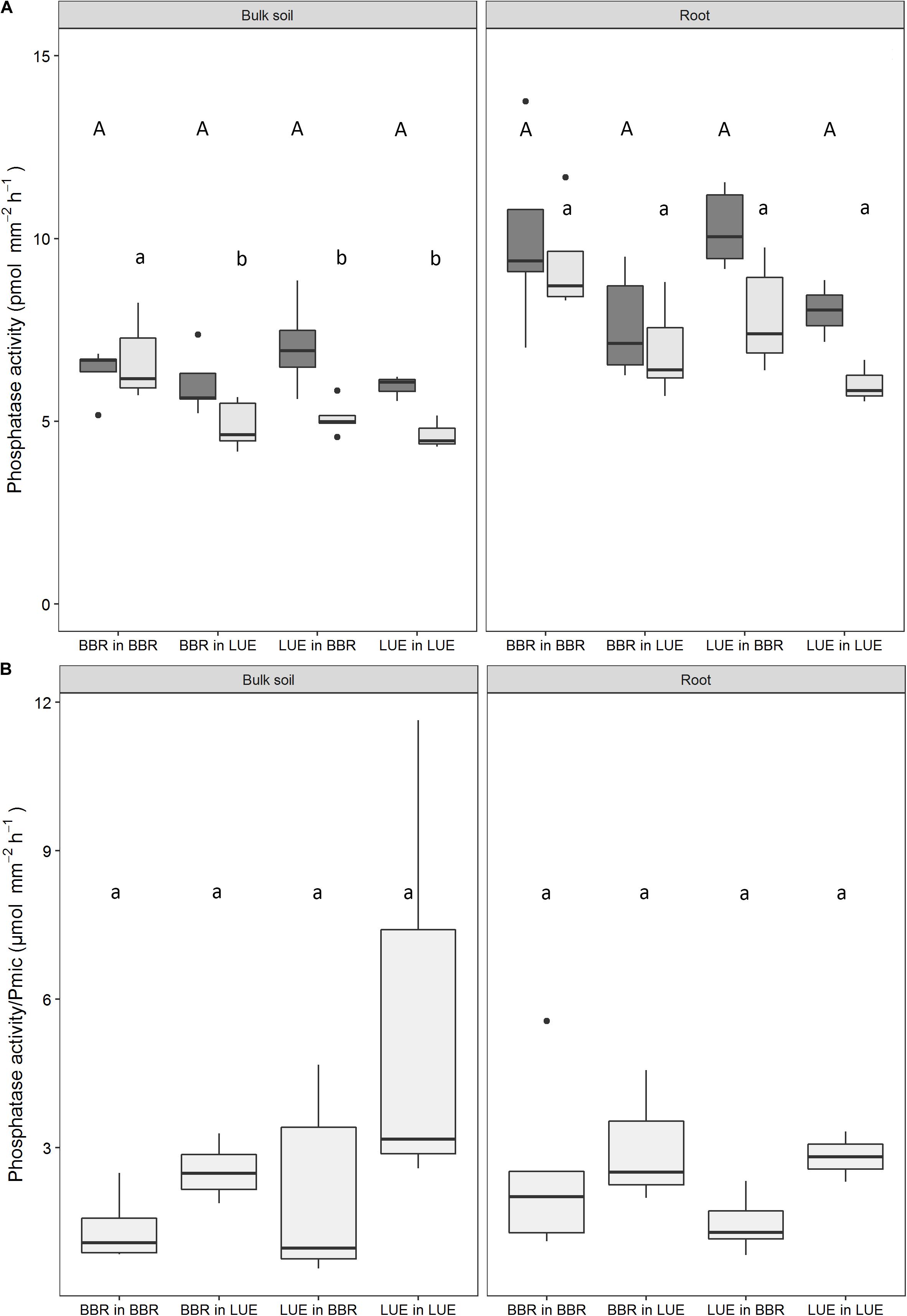
Figure 4. Potential phosphatase activity (PA) measured in the bulk soil and on the surface of the roots using zymography (A) and the ratio between potential PA and microbial P (Pmic) in bulk soil and in the rhizosphere (PA on the root divided by Pmic in the rhizosphere) (B) in a rhizobox experiment with beech (Fagus sylvatica L.) saplings originating from the sites Bad Brückenau (BBR) and Unterlüss (LUE). The saplings were grown in material from the Bv horizon at BBR or from the Bh horizon at LUE. Shown are box plots based on values for replicate rhizoboxes in August of the first (dark gray) and second (light gray) season (n = 5, except n = 3 for LUE in LUE); different letters indicate significant differences between means according to the Tukey post hoc test (uppercase letters, first growing season; lowercase letters, second growing season).
Discussion
Availability of Nutrients in the Soil and Plant Nutritional Status
Our results showed that after two growing seasons, the nutrient concentrations in different compartments of the beech saplings, irrespective of their provenance, had become largely determined by the experimental soil, with the notable exception of P (Table 2). In this section, we discuss these results by first reflecting on the soil conditions and plant characteristics at the sampling sites and then evaluating the plant nutritional status in our experiment as basis for the discussion of root and rhizosphere data in the following sections.
The comparison of the climatic conditions and soil properties at the two sites indicates more favorable conditions for plant growth at BBR than LUE in terms of both water and nutrients. An overall adaptation of the ecosystem at LUE to the low fertility of the mineral soil is the recycling of nutrients via a thick organic surface layer (Bünemann et al., 2016; Lang et al., 2017).
While the beech population at BBR is putatively autochthonous, this is not certain for the one at LUE (Dietrich and Meesenburg, personal communications). Nevertheless, considering (i) the generally close relationship between beech populations from different sites (see section “Introduction”), (ii) the age of the stands, and (iii) the generally strong selection of trees at the juvenile stage, which allows for fast adaptation (e.g., Kremer et al., 2012), trait differences between beeches at the two sites are likely to a large degree due to adaptation to the specific site conditions. These adaptations are expressed in a slower growth of the mature beeches at LUE than at BBR, as is indicated by a smaller stem diameter. When grown under identical climatic conditions in undisturbed soil cores from their own site, saplings from natural rejuvenation at LUE produced smaller leaves with a lower photosynthetic activity and stomatal conductance than saplings from natural rejuvenation at BBR (Yang et al., 2016; Zavišić et al., 2018). In particular, the difference in photosynthetic activity can be considered an indication of a more conservative resource strategy of the beech saplings at LUE (Craine, 2009; Weemstra et al., 2016). Comparing foliar nutrient concentrations and nutrient ratios with threshold values by Mellert and Göttlein (2012); ratios) and Göttlein (2015); concentrations in mature trees), the mature beeches at BBR exhibited a balanced nutrition except for a latent deficiency in K and N/K ratios above the normal range, while the trees at LUE showed a latent deficiency in P and N/P, N/Mg, and N/Ca ratios above the normal range. A lower P nutritional status of juvenile beech trees from LUE than from BBR was documented by Zavišić et al. (2018). They found a respective difference in P concentration for all plant compartments.
The beech saplings from natural rejuvenation used in our experiment were of similar age to those investigated by Yang et al. (2016) and Zavišić et al. (2018). In contrast to the strong influence of “current soil” on the N, Mg, and K concentrations in the second year, P concentrations in different plant compartments were still similar to P concentrations of saplings from the two sites when grown in their natural soil (Zavišić et al., 2018). In the following, we assess the relative nutritional status of the saplings and its change during the experiment (i) by evaluating the foliar nutrient concentrations based on threshold values for juvenile beech trees by Göttlein (2015) and foliar N-to-nutrient ratios based on threshold values by Mellert and Göttlein (2012) and (ii) by comparing the average concentrations in the whole plant among the treatments. Figure 5 summarizes the results of this assessment for the second year. The figure also shows the results of an assessment of the effective nutrient availability in the soil. This assessment is based on the influence of the factor “current soil” on the nutrient concentrations in the plant as it should reflect what the plant effectively took up from the soil. According to this assessment, saplings from LUE growing in the soil from LUE exhibited in both years a low P, Mg, and Ca status and thus reflected well the nutritional status at the provenance site throughout the experiment. When saplings from LUE were growing in the soil from BBR, they also maintained a low P status throughout the experiment, but were low in K instead of Mg and Ca in the first year, and became low in N in the second year (although N/P was still above normal). In the first year, and irrespective of the “current soil” the saplings from BBR were low in K, as were the mature trees at this site. In the second year, the K status had improved, but N was low when growing in the soil from BBR, and Ca was low when growing in the soil from LUE. In the latter case, the P status was low in terms of foliar concentrations but still high when considering the concentrations in the whole plant. The reason for the low N status in the second year of saplings growing in the BBR soil becomes apparent neither from the chemical properties of our experimental soils, nor when inspecting the microbial C, N, and P concentrations and related nutrient ratios in the second year. A possible explanation is competition between beech saplings and mycorrhizal partner expressed as limited transfer of N to the plant (Simon et al., 2017). While ammonium has been shown to be the preferred N form taken up by mycorrhizal roots of beech (Gessler et al., 2005), nitrate appears to be much better transferred to the plant (Leberecht et al., 2016). This explanation is thus consistent with the tendency to lower resin-extractable nitrate in the BBR than the LUE soil in the second year. In summary, based on the assessment of the plant nutritional status at the end of the experiment, two of the experimental treatments represent the growth of beech saplings differing in P and Mg status in a soil that does not provide sufficient P, Mg, and Ca. In the other two treatments, beech saplings differing in P status grew in soil that did not provide sufficient N (Figure 5).
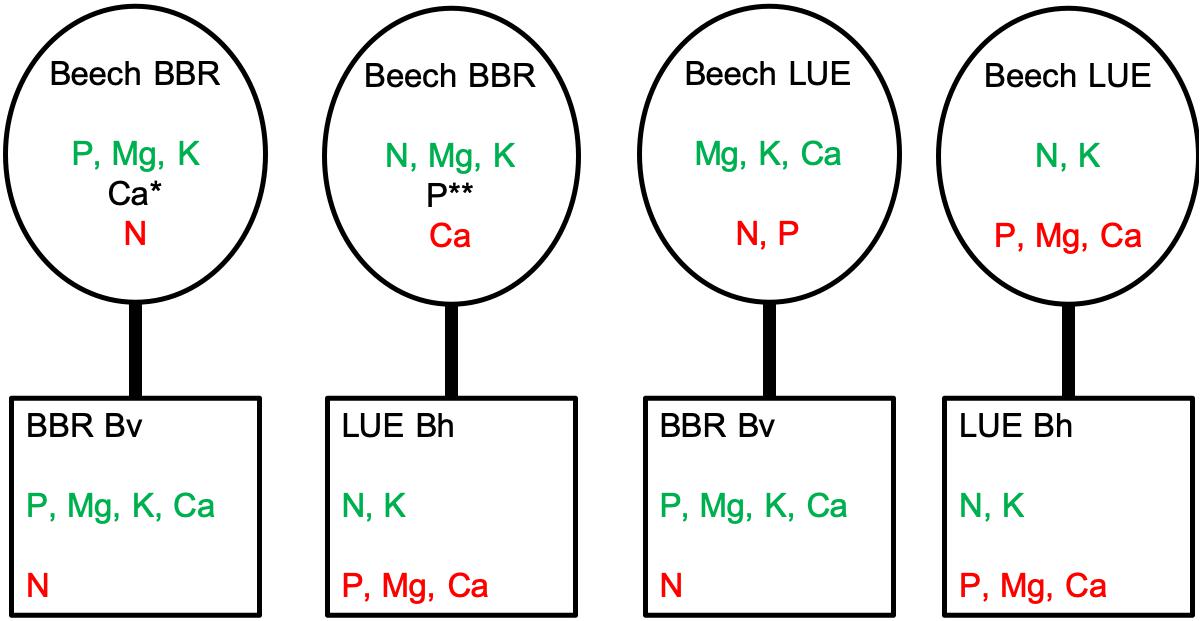
Figure 5. Evaluation of plant nutritional status and effective nutrient availability in the soil in a rhizobox experiment with beech (Fagus sylvatica L.) saplings originating from the sites Bad Brückenau (BBR) and Unterlüss (LUE). The saplings were grown in material from the Bv horizon at BBR or from the Bh horizon at LUE. For the plant nutritional status (i) the comparison of foliar nutrient concentrations and ratios (Table 2) with published threshold values (Mellert and Göttlein, 2012; Göttlein, 2015; green normal or surplus; red deficient or above/below normal) and (ii) the average nutrient concentrations in the whole plant (green low, red high) are considered; the assessments agree except for *: foliar Ca in deficient range, N/Ca in normal range; **: foliar P in deficient range, N/P above normal range, whole plant P high; effective nutrient availability in the soil (green sufficient, red not sufficient) is estimated from the influence of the factor “current soil” on the plant nutritional status in the second year of the experiment.
Root Growth and Morphology
The dominant effect of the experimental factor “current soil” on root growth, as well as on architectural and morphological root traits, demonstrates a high plasticity of the root system of juvenile beech trees when growing in soils differing in nutrient supply (Tables 3, 4 and Figure 1). The stronger reaction of the beech saplings from LUE than BBR to the differences in soil nutrient supply points to a modifying effect of the factor “plant origin” and thus of provenance. We cannot rule out that this may be due to a genotypic difference in sensitivity. However, taking into account the assessment of plant nutritional status and effective soil nutrient availability at the end of our experiment from the previous section, the provenance effect is also consistent with an influence of the plant nutritional status. In this respect, and for growth in a given soil, those nutrients are critical, which are not in sufficient supply in this soil. Specifically, when grown in the LUE soil, which did not provide sufficient P, Mg, and Ca, the beech saplings from the two sites differed in their P and Mg status and their fine roots strongly differed in growth, branching, and diameter. By contrast, when grown in the BBR soil, the saplings from the two sites exhibited a very similar root system, particularly in terms of branching and fine root diameter. In this case, the saplings differed mainly in their nutritional status in P, which was supplied sufficiently by the soil, but not in their status in N, which was not supplied sufficiently by the soil. Furthermore, when accepting the modifying role of the plant nutritional status, the results suggest that the observed differences in root traits can be mainly attributed to differences in the supply of P and/or Mg rather than N. To further examine this, we compare in the following our findings on specific root traits with the results from earlier studies on the effects of P, Mg, or N supply.
Our results on biomass and length of fine roots (Table 3) are consistent with either inhibition of root growth by low P and/or Mg supply or its stimulation by low N supply. They are thus in agreement with the general finding that P and Mg starvation inhibit primary root growth (Gruber et al., 2013; Niu et al., 2013). Effects of differences in N supply on root growth are less clear. This is emphasized, e.g., by results for the model plant Arabidopsis showing that mild N deficiency increases and strong N deficiency decreases root growth (Gruber et al., 2013). Independent on the considered nutrient, findings are more variable under natural than under controlled laboratory conditions. For beech under natural conditions, only small effects of soil fertility on root growth have been found (Leuschner et al., 2004). This can be attributed partly to interaction with other factors that can affect root growth, such as soil texture (Hertel et al., 2013; Weemstra et al., 2017), water availability (Leuschner et al., 2004; Hertel et al., 2013), stand age (Finér et al., 2007), or light (Minotta and Pinzauti, 1996; Yang et al., 2016). Soil texture cannot be excluded as a factor explaining differences in root growth between our two experimental soils. However, such an effect would be opposite to results of earlier studies that found higher fine root growth of beech in sandy than loamy or clayey soils (Hertel et al., 2013; Weemstra et al., 2017).
The lower root branching of the saplings from LUE growing in LUE soil than of the ones in all other treatments (Table 3) is mainly consistent with inhibition by a low combined P and/or Mg supply from soil and plant reserves. This is in contrast to the findings of many studies across different plant species, including trees, that P starvation promotes lateral root growth (Niu et al., 2013; Zhou et al., 2018). However, a lower branching at low P availability was observed earlier for pine (Theodorou and Bowen, 1993).
Our results on root diameter (Figure 1) are consistent with thicker fine roots at low P and/or Mg supply or with finer fine roots at low N supply. There are conflicting reports on how fine root diameter depends on soil P availability. Considering only trees, Yan et al. (2019) found thicker fine roots at low P supply for various tree species, but Razaq et al. (2017) found the opposite for maple. Reports on the effect of different N supply on root diameter are more coherent. Mostly thinner roots at low N supply have been reported for trees (e.g., Razaq et al., 2017; Yan et al., 2017). There are little indications that the differences in fine root diameter in our experiment are affected by genotypic differences in plant “resource strategy” (Weemstra et al., 2016; see also the section “Introduction”). On one hand, the relatively thick fine roots of saplings from LUE when growing in the soil from LUE could reflect a “conservative resource strategy” as expressed by these saplings at their site of origin. On the other hand, when the same saplings were grown in the BBR soil, they exhibited thin roots of the same diameter as those of saplings from BBR, which are rather indicative of an “acquisitive resource strategy.” Vice versa, the BBR saplings formed significantly thicker roots when growing in the soil from LUE.
In contrast to growth, branching, and diameter, fine root traits normalized for length, mass, or volume were insensitive to differences in nutrient supply (Tables 3, 4). Generally, these traits appear not to be affected strongly or uniformly by soil P availability. Again, considering only trees, specific root length (SRL) was found to be smaller at lower P supply, e.g., for spruce (Clemensson-Lindell and Asp, 1995), while the opposite was found, e.g., for pine (Zhang et al., 2013). The few reports on RTD show rather higher values at lower P supply (e.g., Zhang et al., 2013). There are more studies that investigated these root traits in dependence on N supply, but also for this nutrient, the results are variable. A higher SRL at higher N supply was observed, e.g., for larch (Liu et al., 2009) and the opposite was observed, e.g., for maple (Razaq et al., 2017). A higher specific root area at low N supply was reported, e.g., for spruce (Gong and Zhao, 2019). RTD was higher at low N availability, e.g., for spruce (Gong and Zhao, 2019), while the opposite was observed, e.g., for poplar (Yan et al., 2019).
In summary, the comparison with the literature shows that attributing the differences in root traits mainly to differences in the combined supply from soil and plant reserves in P and/or Mg, is consistent with earlier findings on root growth. On the other hand, N supply may have played an additional role determining fine root diameter.
In our “cross-exchange” setup, it cannot be completely ruled out that differences in “plant-soil feedback” may have affected the results on root growth and morphology. In particular, better growth of beech saplings from LUE in BBR than LUE soil would be consistent with negative plant-soil feedback. This kind of feedback is a commonly observed mechanism that has developed between plant species and soil microbial communities over longer time periods and helps to maintain the diversity of plant communities (Bever et al., 2012). It is based on specific root exudates stimulating host-specific pathogens, which in turn restrict root development. This can be enhanced by “auto-toxicity” effects of extracellular DNA leached from decomposed litter and accumulated in the soil (Mazzoleni et al., 2015; Nagler et al., 2018). The absence of such inhibitory effects can lead to better growth when plants are transferred to a “foreign” soil. However, in contrast to different plant species, different populations of the same plant species appear to exhibit similar feedback when grown in the same soil, as was shown for grasses (Wagner et al., 2011) and trees (Gundale et al., 2014). Considering the latter studies and the similar root growth of the beech saplings from BBR in both soils, we conclude that negative plant-soil feedback likely had a negligible effect on root growth and morphology in our experiment.
Mycorrhizal Colonization
Compared to root traits, mycorrhizal inoculation appeared to be relatively insensitive to the treatments. Nevertheless, as for root traits, the results indicate soil properties as the main and plant provenance as a modifying factor (Tables 3, 4). Taking into account our assessment of plant nutritional status and effective nutrient availability (section “Availability of Nutrients in the Soil and Plant Nutritional Status”), there are two potential explanations for higher mycorrhization of beech saplings from the same site when growing in the LUE than the BBR soil. This behavior could be related either to the lower P availability in the LUE soil or to a reaction of the plants to the limited transfer of N from the fungal partners in the BBR soil, as postulated above.
Although a weak effect, the somewhat higher mycorrhization of beech saplings from BBR than LUE, when considering growth in the same soil (Table 3), may point to a higher ability of the saplings with an overall higher nutritional status to provide the fungal partner with carbon compounds. A similar enhancing effect of high plant nutritional status was observed for stimulation of root growth of Helianthus in reaction to reduced N supply (Bowsher et al., 2016).
Phosphorus Mobilization Potential in the Rhizosphere
As for mycorrhizal inoculation, the results from the second year of our experiment indicate a less sensitive reaction of rhizosphere properties related to P mobilization to the treatments than root traits (Figures 2–4 and Table 6). Among these properties, the potential to mineralize organic P appeared to be most strongly affected with a main influence of the factor “current soil.”
Higher rhizosphere concentrations of phosphate in the BBR than LUE soil, irrespective of plant origin, as observed in the first year (Figure 3B), reflect the better P availability in the former soil. The general increase in exchangeable phosphate concentrations between the first and the second year points to a respective difference in the balance between P mobilization and uptake by organisms. A similar reasoning may apply for the increase in nitrate from the first to the second year in the LUE soil (Figure 3A). The importance of nitrate for the N nutrition of our beech saplings was also indicated by the general increase of pH in the rhizosphere. Root exudation of OH– in exchange for NO3– is well known (e.g., Marschner et al., 1996). As a side effect, in acid soils, a pH increase in the rhizosphere, as observed in our experiment (Figure 2), could contribute to a better P solubility due to a decrease of the positive soil surface charge density and thus weaker sorption of phosphate (Hinsinger, 2001).
As reviewed in section “Introduction,” the source of organic acid anions in the rhizosphere can be root exudation, exudation by mycorrhizal hyphae, or release by free-living microorganisms, but it is difficult to distinguish between the different sources. Following a similar argument as for exchangeable phosphate and nitrate, the much larger oxalate and citrate concentrations in the first than the second year (Figures 3C,D), in particular in the BBR soil, point to a difference in the balance between exudation/release to and microbial degradation in the rhizosphere. The significance of microbial degradation for the effectiveness of low-molecular-weight organic acid anions in mobilization of sorbed or mineral bound inorganic P was emphasized earlier (e.g., Hinsinger, 2001). The higher concentration of organic acid anions in the rhizosphere of the BBR than the LUE saplings in the first year is likely related to a higher constitutive root exudation by the saplings from BBR, which increased the organic acid anion concentrations either directly or via stimulation of the production and release by free living microorganisms. Studies on trees have so far provided no evidence for organic acid anion exudation by roots being induced by specific soil conditions (Heim et al., 2001; Eldhuset et al., 2007). The weak effect of “current soil” in the second growing season with slightly higher abundance of oxalate in the rhizosphere of saplings growing in LUE than BBR soil was thus more likely due to lower microbial degradation than a reaction to lower soil P availability.
The higher PA in the BBR than the LUE soil in both years (Figure 4A) is in good agreement with results for mineral bulk soil from the same sites (Bünemann et al., 2016; Spohn et al., 2018), and with the often observed good correlation between PA and soil organic matter content (Feller et al., 1994; Nannipieri et al., 2011). However, PA per unit microbial P, representing the relative investment of the microorganisms in phosphatases, tended to be higher in the LUE than the BBR soil and suggests a reaction to the differences in P supply (Figure 4B). The lower PA in the second than the first growing season in all treatments, both on the roots and in the soil, may be explained as feedback to an increased P availability in the rhizosphere as indicated by the resin-extractable phosphate concentrations. Soil PA has been shown to react sensitively to increased P availability (Marklein and Houlton, 2012; Hofmann et al., 2016). The stronger reaction in the treatments with saplings from LUE is in good agreement with the results by Hofmann et al. (2016) who found a significant fertilization effect only in the rhizosphere of LUE saplings and not in the one of BBR saplings when growing in soil from their own site.
As organic acid anions, phosphatases can be produced and released by roots, mycorrhizal hyphae, and free-living microorganisms (see the section “Introduction”). The higher PA on the roots than in the bulk soil in our experiment can then be attributed either to production and release by the roots or to stimulation of microbial enzyme production in the rhizosphere by root exudation of easily degradable carbon compounds. Hofmann et al. (2016) argued that root exudation of such compounds could, in addition, alleviate the main C limitation of the microorganisms, as found for mineral soils from BBR and LUE by Heuck et al. (2015), and as a consequence induce P deficiency and further stimulate production of phosphatases. Similarly, the significant plant effect in the second growing season with slightly higher PA in the treatments with saplings from BBR could be explained by the higher constitutive root exudation by these saplings, as discussed above.
Conclusion
From our cross-exchange experiment with beech saplings and mineral soil material from two acid forest sites differing in nutrient supply, we draw the following conclusions.
Beech saplings exhibited a high plasticity in adapting their root system to soils, differing in their nutrient supply, in terms of root traits related to growth, architecture, and morphology. The results confirm our first hypothesis that the plastic reactions were determined mainly by the soil properties. Confirming our second hypothesis, plant provenance had a modifying effect that was consistent with an influence of the plant status in those nutrients, which were not in sufficient supply in the soil. However, we cannot completely rule out an additional genotypic difference between the beech saplings from LUE and BBR in their sensitivity to differences in soil nutrient supply.
Compared to root traits, differences among treatments in mycorrhizal inoculation and rhizosphere parameters related to P mobilization were small and mainly determined by the soil properties. Plant origin had only a minor modifying effect, possibly due to differences in the ability to transfer carbon compounds from the shoot to the root and the fungal partner.
Data Availability Statement
The datasets generated for this study are available on request to the corresponding author.
Author Contributions
JL, EF, and SM designed the experiment. SM and JL collected the plant and soil materials used in the study. SM set up and carried out the experiment, performed most of the chemical analyses, analyzed the data, and wrote the first version of the manuscript. MS provided the expertise in measuring phosphatase activity. All authors contributed significantly to the final version of the manuscript.
Funding
The project was carried out in the framework of the Priority Program SPP 1685 “Ecosystem Nutrition” of the German and Swiss National Science Foundations (DFG and SNF, respectively). This particular project was funded by the SNF project no. 149138 and by internal funds of the Swiss Federal Research Institute WSL.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The forest research stations “Bayerische Landesanstalt für Wald und Forstwirtschaft” (LWF) and “Nordwestdeutsche Forstliche Versuchsanstalt” (NW-FVA) provided access to the field sites. The following groups at the Swiss Federal Research Institute WSL provided technical assistance: the technical support at Birmensdorf and the workshop at Davos helped with construction of the rhizoboxes, the experimental garden operated and maintained the greenhouse, the central analytical laboratories carried out part of the chemical analyses, and the soil physical and chemical laboratories provided various technical support. We thank Jaane Krüger (Soil Ecology, University of Freiburg i.B.), Klaus Kaiser (Soil Science and Soil Protection, Martin-Luther University Halle-Wittenberg), and Jörg Prietzel (Soil Science, TU Munich) for providing data on soil properties at the sampling sites; Henning Meesenburg (NW-FVA) and Hans-Peter Dietrich (LWF) for providing data on foliar concentrations at the sampling sites and information on site history; and Christoph Sperisen (WSL) for advice on genetic relationships among beech forests.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2020.535117/full#supplementary-material
Footnotes
- ^ http://www.ecosystem-nutrition.uni-freiburg.de/
- ^ https://cran.r-project.org/web/packages/car/car.pdf
References
Bahr, A., Ellström, M., Akselsson, C., Ekblad, A., Mikusinska, A., and Wallander, H. (2013). Growth of ectomycorrhizal fungal mycelium along a Norway spruce forest nitrogen deposition gradient and its effect on nitrogen leakage. Soil Biol. Biochem. 59, 38–48. doi: 10.1016/j.soilbio.2013.01.004
Bever, J. D., Platt, T. G., and Morton, E. R. (2012). Microbial population and community dynamics on plant roots and their feedbacks on plant communities. Annu. Rev. Microbiol. 66, 265–283. doi: 10.1146/annurev-micro-092611-150107
Blossfeld, S., and Gansert, D. (2007). A novel non-invasive optical method for quantitative visualization of pH dynamics in the rhizosphere of plants. Plant Cell Environ. 30, 176–186. doi: 10.1111/j.1365-3040.2006.01616.x
Borken, W., Kossmann, G., and Matzner, E. (2007). Biomass, morphology and nutrient contents of fine roots in four Norway spruce stands. Plant Soil 292, 7–93. doi: 10.1007/s11104-007-9204-x
Bowsher, A. W., Miller, B. J., and Donovan, L. A. (2016). Evolutionary divergences in root system morphology, allocation, and nitrogen uptake in species from high- versus low-fertility soils. Funct. Plant Biol. 43, 129–140. doi: 10.1071/FP15162
Brookes, P. C., Landman, A., Pruden, G., and Jenkinson, D. S. (1985). Chloroform fumigation and the release of soil nitrogen: a rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 17, 837–842.
Brunner, I., and Brodbeck, S. (2001). Response of mycorrhizal Norway spruce seedlings to various nitrogen loads and sources. Environ. Poll. 114, 223–233.
Buiteveld, J., Vendramin, G. G., Leonardi, S., Kamer, K., and Geburek, T. (2007). Genetic diversity and differentiation in European beech (Fagus sylvatica L.) stands varying in management history. For. Ecol. Manag. 247, 98–106. doi: 10.1016/j.foreco.2007.04.018
Bünemann, E. K., Augstburger, S., and Frossard, E. (2016). Dominance of either physicochemical or biological phosphorus cycling processes in temperate forest soils of contrasting phosphate availability. Soil Biol. Biochem. 101, 85–95. doi: 10.1016/j.soilbio.2016.07.005
Bünemann, E. K., Bossio, D. A., Smithson, P. C., Frossard, E., and Oberson, A. (2004). Microbial community composition and substrte use in a highly weathered soil as affected by crop rotation and P fertilization. Soil Biol. Biochem. 36, 889–901. doi: 10.1016/j.soilbio.2004.02.002
Chalot, M., and Plassard, C. (2011). “Ectomycorrhiza and nitrogen provision to the host tree,” in Ecological Aspects of Nitrogen Metabolism in Plants, First Edition, eds J. C. Polacco and C. D. Todd (Hoboken, NJ: John Wiley & Sons Inc), 69–94.
Chen, W., Koide, R. T., Adams, T. S., DeForest, J. L., Cheng, L., and Eissenstat, D. M. (2016). Root morphology and mycorrhizal symbioses together shape nutrient foraging strategies of temperate trees. Proc. National Acad. Sci. U.S.A. 113, 8741–8746. doi: 10.1073/pnas.1601006113
Clemensson-Lindell, A., and Asp, H. (1995). Fine-root morphology and uptake of 32P and 35S in a Norway spruce (Picea abies (L.) Karst.) stand subjected to various nutrient and water supplies. Plant Soil 173, 147–155.
Cuervo-Alarcon, L., Arend, M., Muller, M., Sperisen, C., Finkeldey, R., and Krutovsky, K. V. (2018). Genetic variation and signatures of natural selection in populations of European beech (Fagus sylvatica L.) along precipitation gradients. Tree Genet. Genom. 14:84. doi: 10.1007/s11295-018-1297-2
Durrant, T. H., de Rigo, D., and Caudullo, G. (2016). Fagus sylvatica and other beeches in Europe: distrubution, habitat, usage and threats. Eur. Atlas For. Tree Spec. 2016, 130–131. doi: 10.2788/038466
Eldhuset, T. D., Swensen, B., Wickstrøm, T., and Wollebæk, G. (2007). Organic acids in root exudates from Picea abies seedlings influenced by mycorrhiza and aluminum. J. Plant Nutr. Soil Sci. 170, 645–648. doi: 10.1002/jpln.200700005
Feller, C., Frossard, E., and Brossard, M. (1994). Activité phosphatasique de quelques sols tropicaux à argile 1:1. Répartition dans les fractions granulométriques. Can. J. Soil Sci. 74, 121–129.
Finér, L., Helmisaari, H.-S., Lõhmus, K., Majdi, H., Brunner, I., Børja, I., et al. (2007). Variation in fine root biomass of three European tree species: beech (Fagus sylvatica L.), Norway spruce (Picea abies L. Karst.), and Scots pine (Pinus sylvestris L.). Plant Biosyst. 141, 394–405.
Garbaye, J., and Wilhelm, M. E. (1985). Facteurs limitants et aspects dynamiques de la mycorhization contrôlée de Fagus silvatica Lin. par Hebeloma crustuliniforme (Bull. ex Saint-Amans) Quél, sur tourbe fertilisée. Annal. Sci. For. 42, 53–68. doi: 10.1051/forest:19850104
George, T. S., Fransson, A.-M., Hammond, J. P., and White, P. J. (2011). “Phosphorus nutrition: rhizosphere processes, plant response and adaptations,” in Phosphorus in Action, Soil Biology, Vol. 26, eds E. K. Bünemann, A. Oberson, and E. Frossard (Berlin: Springer), 245–271.
Gessler, A., Jung, K., Gasche, R., Papen, H., Heidenfelder, A., Börner, E., et al. (2005). Climate and forest management influence nitrogen balance of European beech forests: microbial N transformations and inorganic N net uptake capacity of mycorrhizal roots. Eur. J. For. Res. 124, 95–111. doi: 10.1007/s10342-005-0055-9
Gianfreda, L., and Ruggiero, P. (2006). “Enzyme activities in soil,” in Nucleic Acids and Proteins in Soil, eds P. Nannipieri and K. Smalla (Berlin: Springer-Verlag), 257–311.
Gong, L., and Zhao, J. (2019). The response of fine root morphological and physiological traits to added nitrogen in Schrenk’s spruce (Picea schrenkiana) of the Tianshan mountains, China. PeerJ 7:e8194. doi: 10.7717/peerj.8194
Göttlein, A. (2015). Ranges of threshold values for the nutritional assessment of the main tree species spruce, pine, oak and beech. Allg. Forst Jagd-Ztg. 186, 110–116.
Gruber, B. D., Giehl, R. F. H., Friedel, S., and von Wirén, N. (2013). Plasticity of the Arabidopsis root system under nutrient deficiencies. Plant Phys. 163, 161–179. doi: 10.1104/pp.113.218453
Gundale, M. J., Kardol, P., Nilsson, M.-C., Nilsson, U., Lucas, R. W., and Wardle, D. A. (2014). Interactions with soil biota shift from negative to positive when a tree species is moved outside its native range. New Phytol. 202, 415–421. doi: 10.1111/nph.12699
Hauenstein, S., Neidhardt, H., Lang, F., Krüger, J., Hofmann, D., Pütz, T., et al. (2018). Organic layers favor phosphorus storage and uptake by young beech trees (Fagus sylvatica L.) at nutrient poor ecosystems. Plant Soil 432, 289–301. doi: 10.1007/s11104-018-3804-5
Hawkins, B. J., Jones, M. D., and Kranabetter, J. M. (2015). Ectomycorrhizae and tree seedling nitrogen nutrition in forest restoration. New For. 46, 747–771. doi: 10.1007/s11056-015-9488-2
Hedley, M. J., White, R. E., and Nye, P. H. (1982). Plant-induced changes in the rhizosphere of Rape (Brassica napus ver. Emerald) seedlings. III. Changes in L Value, Soil Phosphate Fractions and Phosphatase Activity. New Phytol. 91, 45–56.
Heim, A., Brunner, I., Frey, B., Frossard, E., and Luster, J. (2001). Root exudation, organic acids, and element distribution in roots of Norway spruce seedlings treated with aluminum in hydroponics. J. Plant Nutr. Soil Sci. 164, 519–526. doi: 10.1002/1522-2624(200110)164:5<519::AID-JPLN519<3.0.CO;2-Y
Hertel, D., Strecker, T., Müller-Haubold, H., and Leuschner, C. (2013). Fine root biomass and dynamics in beech forests across a precipitation gradient - Is optimal resource partitioning theory applicable to water-limited mature trees? J. Ecol. 101, 1183–1200. doi: 10.1111/1365-2745.12124
Heuck, C., Weig, A., and Spohn, M. (2015). Soil microbial biomass C:N:P stoichiometry and microbial use of organic phosphorus. Soil Biol. Biochem. 85, 119–129. doi: 10.1016/j.soilbio.2015.02.029
Hinsinger, P. (2001). Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes?: a review. Plant Soil 237, 173–195.
Hinsinger, P., Brauman, A., Devau, N., Gérard, F., Jourdan, C., Laclau, J.-P., et al. (2011). Acquisition of phosphorus and other poorly mobile nutrients by roots. Where do plant nutrition models fail? Plant Soil 348, 29–61. doi: 10.1007/s11104-011-0903-y
Hodge, A. (2006). Plastic plants and patchy soils. J. Exp. Bot. 57, 401–411. doi: 10.1093/jxb/eri280
Hodge, A., Berta, G., Doussan, C., Merchan, F., and Crespi, M. (2009). Plant root growth, architecture and function. Plant Soil 321, 153–187. doi: 10.1007/s11104-009-9929-9
Hofmann, K., Heuck, C., and Spohn, M. (2016). Phosphorus resorption by young beech trees and soil phosphatase activity as dependent on phosphorus availability. Oecologia 181, 369–379. doi: 10.1007/s00442-016-3581-x
Holzmann, S., Missong, A., Puhlmann, H., Siemens, J., Bol, R., Klumpp, E., et al. (2016). Impact of anthropogenic induced nitrogen input and liming on phosphorus leaching in forest soils. J. Plant Nutr. Soil Sci. 179, 443–453. doi: 10.1002/jpln.201500552
Ishida, T. A., Nara, K., and Hogetsu, T. (2007). Host effects on ectomycorrhizal fungal communities insight from eight host species in mixed conifer–broadleaf forests. New Phytol. 174, 430–440.
Jonard, M., Fürst, A., Verstraeten, A., Thimonier, A., Timmermann, V., Potoèiæ, N., et al. (2015). Tree mineral nutrition is deteriorating in Europe. Glob. Chang. Biol. 21, 418–430. doi: 10.1111/gcb.12657
Kazantseva, O., Bingham, M., Simard, S. W., and Berch, S. M. (2009). Effects of growth medium, nutrients, water, and aeration on mycorrhization and biomass allocation of greenhouse-grown interior Douglas-fir seedlings. Mycorrhiza 20, 51–66. doi: 10.1007/s00572-009-0263-0
Kiba, T., and Krapp, A. (2016). Plant nitrogen acquisition under low availability: regulation of uptake and root architecture. Plant Cell Phys. 54, 707–714. doi: 10.1093/pcp/pcw052
Kouno, K., Tuchiya, Y., and Ando, T. (1995). Measurement of soil microbial phosphorus by anion exchange membrane method. Soil Biol. Biochem. 27, 1353–1357.
Kremer, A., Ronce, O., Robledo-Arnuncio, J. J., Guillaume, F., Bohrer, G., Nathan, R., et al. (2012). Long-distance gene flow and adaptation of forest trees to rapid climate change. Ecol. Lett. 15, 378–392.
Lang, F., Krüger, J., Amelung, W., Willbold, S., Frossard, E., Bünemann, E. K., et al. (2017). Soil phosphorus supply controls P nutrition strategies of beech forest ecosystems in Central Europe. Biogeochemistry 136, 5–29. doi: 10.1007/s10533-017-0375-0
Leberecht, M., Dannenmann, M., Tejedor, J., Simon, J., Rennenberg, H., and Polle, A. (2016). Segregation of nitrogen use between ammonium and nitrate of ectomycorrhizas and beech trees. Plant Cell Environ. 39, 2691–2700. doi: 10.1111/pce.12820
Leuschner, C., Hertel, D., Schmid, I., Koch, O., Muhs, A., and Hölscher, D. (2004). Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility. Plant Soil 258, 43–56.
Leuschner, C., Meier, I. C., and Hertel, D. (2006). On the niche breadth of Fagus sylvatica: soil nutrient status in 50 Central European beech stands on a broad range of bedrock types. Ann. For. Sci. 63, 355–368. doi: 10.1051/forest:2006016
Liu, J., Mei, L., Gu, J., Quan, X., and Wang, Z. (2009). Effects of nitrogen fertilization on fine root biomass and morphology of Fraxinus mandshurica and Larix gmelinii: a study with in-growth core approach. Shengtaixue Zazhi 28, 1–6.
Marklein, A. R., and Houlton, B. Z. (2012). Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems. New Phytol. 193, 696–704. doi: 10.1111/j.1469-8137.2011.03967.x
Marschner, H., Kirkby, E. A., and Cakmak, I. (1996). Effect of mineral nutritional status on shoot-root partitioning of photoassimilates and cycling of mineral nutrients. J. Exp. Bot. 47, 1255–1263. doi: 10.1093/jxb/47.Special_Issue.1255
Mazzoleni, S., Bonanomi, G., Incerti, G., Chiusano, M. L., and Termolino, P. (2015). Inhibitory and toxic effects of extracellular self-DNA in litter: a mechanism for negative plant-soil feedbacks? New Phytol. 205, 1195–1210. doi: 10.1111/nph.13121
Meier, I. C., and Leuschner, C. (2008). Genotypic variation and phenotypic plasticity in the drought response of fine roots of European beech. Tree Phys. 28, 297–309.
Meller, S., Frossard, E., and Luster, J. (2019). Phosphorus allocation to leaves of beech saplings reacts to soil phosphorus availability. Front. Plant Sci. 10:744. doi: 10.3389/fpls.2019.00744
Mellert, K. H., and Göttlein, A. (2012). Comparison of new foliar nutrient thresholds derived from van den Burg’s literature compilation with established central European references. Eur. J. For. Res. 131, 1461–1472. doi: 10.1007/s10342-012-0615-8
Minotta, G., and Pinzauti, S. (1996). Effects of light and soil fertility on growth, leaf chlorophyll content and nutrient use efficiency of beech (Fagus sylvatica L.) seedlings. For. Ecol. Manag. 86, 61–71.
Nagler, M., Insam, H., Pietramellara, G., and Ascher-Jenull, J. (2018). Extracellular DNA in natural environments: features, relevance and applications. Appl. Microbiol. Biotechnol. 102, 6343–6356. doi: 10.1007/s00253-018-9120-4
Nannipieri, P., Giagnoni, L., Landi, L., and Renella, G. (2011). “Role of phosphatase enzymes in soil,” in Phosphorus in Action, Soil Biology, Vol. 26, eds E. K. Bünemann, A. Oberson, and E. Frossard (Berlin: Springer), 215–243.
Niu, Y. F., Chai, R. S., Jin, G. L., Wang, H., Tang, C. X., and Zhang, Y. S. (2013). Responses of root architecture development to low phosphorus availability: a review. Ann. Bot. 112, 391–408. doi: 10.1093/aob/mcs285
Oburger, E., Jones, D. L., and Wenzel, W. W. (2011). Phosphorus saturation and pH differentially regulate the efficiency of organic acid anion-mediated P solubilization mechanisms in soil. Plant Soil 341, 363–382. doi: 10.1007/s11104-010-0650-5
Ohno, T., and Zibilske, L. M. (1991). Determination of low concentrations of phosphorus in soil extracts using malachite green. Soil Sci. Soc. Am. J. 55, 892–895.
Plassard, C., and Dell, B. (2010). Phosphorus nutrition of mycorrhizal trees. Tree Phys. 30, 1129–1139. doi: 10.1093/treephys/tpq063
Plassard, C., Louche, J., Ali, M. A., Duchemin, M., Legname, E., and Cloutier-Hurteau, B. (2011). Diversity in phosphorus mobilisation and uptake in Ectomycorrhizal fungi. Annals For. Sci. 68, 33–43. doi: 10.1007/s13595-010-0005-7
Razaq, M., Zhang, P., Shen, H., and Salahuddin, J. (2017). Influence of nitrogen and phosphorous on the growth and root morphology of Acer mono. PLoS One 12:e0171321. doi: 10.1371/journal.pone.0171321
Richardson, A. E., Hocking, P. J., Simpson, R. J., and George, T. S. (2009). Plant mechanisms to optimise access to soil phosphorus. Crop Pasture Sci. 60, 124–143. doi: 10.1071/CP07125
Riley, D., and Barber, S. A. (1971). Effect of ammonium and nitrate fertilization on phosphorus uptake as related to root-induced pH changes at the root-soil interface. Soil Sci. Soc. Am. J. 35, 300–306. doi: 10.2136/sssaj1971.03615995003500020035x
Schneider, C. A., Rasband, W. S., and Eliceiri, K. W. (2012). NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675.
Shahbaz, A. M., Oki, Y., Adachi, T., Murata, Y., and Khan, M. H. R. (2006). Phosphorus starvation induced root-mediated pH changes in solubilization and acquisition of sparingly soluble P sources and organic acids exudation by Brassica cultivars. Soil Sci. Plant Nutr. 52, 623–633. doi: 10.1111/j.1747-0765.2006.00082.x
Shi, S., Condron, L., Larsen, S., Richardson, A. E., Jones, E., Jiao, J., et al. (2011). In situ sampling of low molecular weight organic anions from rhizosphere of radiata pine (Pinus radiata) grown in a rhizotron system. Env. Exp. Bot. 70, 131–142. doi: 10.1016/j.envexpbot.2010.08.010
Simon, J., Dannenmann, M., Pena, R., Gessler, A., and Rennenberg, H. (2017). Nitrogen nutrition of beech forests in a changing climate: importance of plant-soil-microbe water, carbon, and nitrogen interactions. Plant Soil 418, 89–114.
Spohn, M., Ermak, A., and Kuzyakov, Y. (2013). Microbial gross organic phosphorus mineralization can be stimulated by root exudates–a 33P isotopic dilution study. Soil Biol. Biochem. 65, 254–263.
Spohn, M., and Kuzyakov, Y. (2013). Distribution of microbial-and root-derived phosphatase activities in the rhizosphere depending on P availability and C allocation–Coupling soil zymography with 14C imaging. Soil Biol. Biochem. 67, 106–113.
Spohn, M., Zavišić, A., Nassal, P., Bergkemper, F., Schulz, S., and Marhan, S. (2018). Temporal variations of phosphorus uptake by soil microbial biomass and young beech trees in two forest soils with contrasting phosphorus stocks. Soil Biol. Biochem. 117, 191–202. doi: 10.1016/j.soilbio.2017.10.019
Sun, Y., Gu, J.-C., Zhuang, H.-F., and Wang, Z.-Q. (2010). Effects of ectomycorrhizal colonization and nitrogen fertilization on morphology of root tips in a Larix gmelinii plantation in northeastern China. Ecol. Res. 25, 295–302. doi: 10.1007/s11284-009-0654-x
Theodorou, C., and Bowen, G. D. (1993). Root morphology, growth and uptake of phosphorus and nitrogen of Pinus radiata families in different soils. For. Ecol. Manag. 56, 43–56.
Tiessen, H., and Moir, J. O. (2006). “Chapter 25: Characterization of available P by sequential extraction,” in Soil Sampling and Methods of Analysis Canadian Society of Soil Science, 2nd Edn, eds M. Carter and E. Gregorich (Milton Park: Taylor and Francis).
Voroney, R. P., Brookes, P. C., and Beyaert, R. P. (2006). “Chapter 49: Soil microbial biomass C, N, P, and S,” in Soil Sampling and Methods of Analysis, Canadian Society of Soil Science, 2nd Edn, eds M. Carter and E. Gregorich (Milton Park: Taylor and Francis).
Wagner, V., Antunes, P. M., Ristow, M., Lechner, U., and Hensen, I. (2011). Prevailing negative soil biota effect and no evidence for local adaptation in a widespread Eurasian grass. PLoS One 6:e17580. doi: 10.1371/journal.pone.0017580
Walthert, L., and Meier, E. S. (2017). Tree species distribution in temperate forests is more influenced by soil than climate. Ecol. Evol. 7, 9473–9484. doi: 10.1002/ece3.3436
Weemstra, M., Mommer, L., Visser, E. J. W., van Ruijven, J., Kuyper, T. W., Mohren, G. M. J., et al. (2016). Towards a multidimensional root trait framework: a tree root review. New Phytol. 211, 1159–1169. doi: 10.1111/nph.14003
Weemstra, M., Sterck, F. J., Visser, E. J. W., Kuyper, T. W., Goudzwaard, L., and Mommer, L. (2017). Fine-root trait plasticity of beech (Fagus sylvatica) and spruce (Picea abies) forests on two contrasting soils. Plant Soil 415, 175–188. doi: 10.1007/s11104-016-3148-y
Xuan, W., Beeckman, T., and Xu, G. (2017). Plant nitrogen nutrition: sensing and signaling. Curr. Opin. Plant Biol. 39, 57–65. doi: 10.1016/j.pbi.2017.05.010
Yan, G., Chen, F., Zhang, X., Wang, J., Han, S., Xing, Y., et al. (2017). Spatial and temporal effects of nitrogen addition on root morphology and growth in a boreal forest. Geoderma 303, 178–187.
Yan, X.-L., Wang, C., Ma, X., and Wu, P. (2019). Root morphology and seedling growth of three tree species in southern China in response to homogeneous and heterogeneous phosphorus supplies. Trees 33, 1283–1297. doi: 10.1007/s00468-019-01858-x
Yang, N., Zavišić, A., Pena, R., and Polle, A. (2016). Phenology, photosynthesis, and phosphorus in European beech (Fagus sylvatica L.) in two forest soils with contrasting P contents. J. Plant Nutr. Soil Sci. 179, 151–158. doi: 10.1002/jpln.201500539
Zavišić, A., Yang, N., Marhan, S., Kandeler, E., and Polle, A. (2018). Forest soil phosphorus resources and fertilization affect ectomycorrhizal community composition, beech P uptake, and photosynthesis. Front. Plant Sci. 9:463. doi: 10.3389/fpls.2018.00463
Zhang, Y., Zhou, Z., and Yang, Q. (2013). Genetic variations in root morphology and phosphorus efficiency of Pinus massoniana under heterogeneous and homogeneous low phosphorus conditions. Plant Soil 364, 93–104. doi: 10.1007/s11104-012-1352-y
Keywords: root growth, root architecture, root morphology, mycorrhizal colonization, rhizosphere, potential phosphatase activity, organic acid anions, acid forest soil
Citation: Meller S, Frossard E, Spohn M and Luster J (2020) Plant Nutritional Status Explains the Modifying Effect of Provenance on the Response of Beech Sapling Root Traits to Differences in Soil Nutrient Supply. Front. For. Glob. Change 3:535117. doi: 10.3389/ffgc.2020.535117
Received: 14 February 2020; Accepted: 31 August 2020;
Published: 25 September 2020.
Edited by:
Sebastian Loeppmann, Christian-Albrechts-Universität zu Kiel, GermanyReviewed by:
Stefano Mazzoleni, University of Naples Federico II, ItalyMaria Teresa Ceccherini, University of Florence, Italy
Copyright © 2020 Meller, Frossard, Spohn and Luster. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jörg Luster, am9lcmcubHVzdGVyQHdzbC5jaA==