 Andrew J. Plumptre1,2*†
Andrew J. Plumptre1,2*† Daniele Baisero1†
Daniele Baisero1† R. Travis Belote3
R. Travis Belote3 Ella Vázquez-Domínguez4†
Ella Vázquez-Domínguez4† Soren Faurby5†
Soren Faurby5† Włodzimierz Jȩdrzejewski6†
Włodzimierz Jȩdrzejewski6† Henry Kiara7Hjalmar Kühl8,9Ana Benítez-López10†Carlos Luna-Aranguré4†
Henry Kiara7Hjalmar Kühl8,9Ana Benítez-López10†Carlos Luna-Aranguré4† Maria Voigt11
Maria Voigt11 Serge Wich12,13William Wint14
Serge Wich12,13William Wint14 Juan Gallego-Zamorano15†
Juan Gallego-Zamorano15† Charlotte Boyd16†
Charlotte Boyd16†- 1Key Biodiversity Areas Secretariat, BirdLife International, Cambridge, United Kingdom
- 2Conservation Science Group, Department of Zoology, University of Cambridge, Cambridge, United Kingdom
- 3The Wilderness Society, Bozeman, MT, United States
- 4Instituto de Ecología, Universidad Nacional Autónoma de México, Ciudad Universitaria, Mexico City, Mexico
- 5Department of Biological and Environmental Sciences, University of Gothenburg, Gothenburg, Sweden
- 6Centro de Ecología, Instituto Venezolano de Investigaciones Científicas (IVIC), Caracas, Venezuela
- 7International Livestock Research Institute, Nairobi, Kenya
- 8Max Planck Institute for Evolutionary Anthropology, Department of Primatology, Leipzig, Germany
- 9German Centre for Integrative Biodiversity Research (iDiv), Leipzig, Germany
- 10Integrative Ecology Group, Estación Biológica de Doñana (EBD-CSIC), Seville, Spain
- 11Durrell Institute of Conservation and Ecology (DICE), School of Anthropology and Conservation, University of Kent, Canterbury, United Kingdom
- 12School of Natural Sciences and Psychology, Liverpool John Moores University, Liverpool, United Kingdom
- 13Institute for Biodiversity and Ecosystem Dynamics, University of Amsterdam, Amsterdam, Netherlands
- 14Environmental Research Group Oxford Ltd., Department of Zoology, Oxford, United Kingdom
- 15Department of Environmental Science, Radboud University Nijmegen, Nijmegen, Netherlands
- 16International Union for Conservation of Nature (IUCN) North America Office, Washington DC, WA, United States
Conservation efforts should target the few remaining areas of the world that represent outstanding examples of ecological integrity and aim to restore ecological integrity to a much broader area of the world with intact habitat and minimal species loss while this is still possible. There have been many assessments of “intactness” in recent years but most of these use measures of anthropogenic impact at a site, rather than faunal intactness or ecological integrity. This paper makes the first assessment of faunal intactness for the global terrestrial land surface and assesses how many ecoregions have sites that could qualify as Key Biodiversity Areas (KBAs – sites contributing significantly to the global persistence of biodiversity) based on their outstanding ecological integrity (under KBA Criterion C). Three datasets are combined on species loss at sites to create a new spatially explicit map of numbers of species extirpated. Based on this map it is estimated that no more than 2.9% of the land surface can be considered to be faunally intact. Additionally, using habitat/density distribution data for 15 large mammals we also make an initial assessment of areas where mammal densities are reduced, showing a further decrease in surface area to 2.8% of the land surface that could be considered functionally intact. Only 11% of the functionally intact areas that were identified are included within existing protected areas, and only 4% within existing KBAs triggered by other criteria. Our findings show that the number of ecoregions that could qualify as Criterion C KBAs could potentially increase land area up to 20% if their faunal composition was restored with the reintroduction of 1–5 species. Hence, if all necessary requirements are met in order to reintroduce species and regain faunal integrity, this will increase ecological integrity across much of the area where human impacts are low (human footprint ≤4). Focusing restoration efforts in these areas could significantly increase the area of the planet with full ecological integrity.
Introduction
Ecologically intact ecosystems have long been recognized as an important conservation objective for protection. Wilderness areas were identified as important priorities over 30 years ago (McCloskey and Spalding, 1989; Mittermeier et al., 1998), but more recently there have been attempts to be more explicit about what is being measured when referring to wilderness, with a focus on quantifying intact habitat (Potapov et al., 2008, 2017; Watson et al., 2016; Kennedy et al., 2019; Grantham et al., 2020) and the values of intact habitat (Watson et al., 2018). Many assessments have focused on mapping anthropogenic influence (Human Footprint: Sanderson et al., 2002a; Venter et al., 2016b; Anthromes: Goldewijk et al., 2017; Global Human Modification: Kennedy et al., 2018; Low Impact Areas: Jacobson et al., 2019; Intact Forest: Potapov et al., 2008, 2017) and have generated several maps of anthropogenic impact that independently estimate that 20–40% of the earth’s terrestrial surface is under low human influence (Riggio et al., 2020). Recently, the integrity of natural ecosystems has been recognized as an important goal in the post 2020 global biodiversity framework (CBD, 2020), and the value of intact forest also been recognized both for biodiversity conservation and for ecosystem services (Watson et al., 2018). Systematic identification of wilderness areas with outstanding ecological integrity requires a clear definition of ecological integrity and indicators of intactness that can be used to assess areas with potential ecological integrity at a global scale.
There is no clear definition of what is meant by intactness and the term is used loosely in the scientific literature. However, the KBA Standard, published in 2016, following an extensive global consultation process, provides a global standard for identifying “sites contributing significantly to the global persistence of biodiversity” (IUCN, 2016, p. 9). Five over-arching criteria are used to identify KBAs, with Criterion C identifying sites of globally outstanding ecological integrity. The KBA Standard defines Criterion C KBAs in terms of ecological intactness as: A site is one of ≤ 2 per ecoregion characterized by wholly intact ecological communities, comprising the composition and abundance of native species and their interactions. The KBA Standard further defines “intact ecological community” as: “An ecological community having the complete complement of species known or expected to occur in a particular site or ecosystem, relative to a regionally appropriate historical benchmark, which will often correspond to pre-industrial times.” The KBA Standard provides some guidance on how ecological integrity should be measured and states that it “should be observed or inferred from both direct measures of species composition and abundance/biomass across taxonomic groups (particularly for species indicative of long-term structural stability and functionality or those known to be highly sensitive to human impact) and absence (or very low levels) of direct industrial human impact (as quantified by appropriate indices at the scale of interest and verified on the ground or in the water).” There is also a recommendation that Criterion C KBAs “should ideally be delineated to be at least 10,000 km2 in size, within the confines of manageability” and “based on historical bounds of variation using a regionally appropriate benchmark (e.g., the past 500 years).” Ecological integrity needs to be assessed against some regionally appropriate historical benchmark. Species distributions have been changing for millennia because of human activity [e.g., the loss of large mammals across continents (Sandom et al., 2014)] and we chose to focus on changes that have occurred since the year 1500 AD, because this is the baseline date for assessing species extinctions within the IUCN Red List of Threatened species (IUCN, 2020).
Building on these definitions, we distinguish three types of intactness which together make up ecological integrity. Habitat intactness occurs where there is no anthropogenic habitat conversion/transformation. Habitat intactness can be estimated and monitored using remote sensing techniques and measures of the impact of the human footprint on ecosystem structure, although it is easier to estimate for forests than some other habitat types. However, habitat intactness does not necessarily equate to faunal intactness (Benítez-López et al., 2019; Plumptre et al., 2019). Several studies have shown that habitats that appear intact are often missing large, wide-ranging species that play important ecological roles in the ecosystem (Morrison et al., 2007; Plumptre et al., 2019). Medium-size and even small species also play key ecological and functional roles in ecosystems (e.g., small carnivores, bats), significantly contributing to integrity and intactness (Camargo-Sanabria et al., 2015; Brodie et al., 2018). Faunal intactness is therefore differentiated here and occurs where there is no loss of faunal species. We also include the term functional intactness to refer to where there is no reduction in faunal densities below ecologically functional densities to differentiate this intactness at sites from those where species are just present but at diminished density. Identifying sites with outstanding ecological integrity requires an assessment of habitat, faunal and functional intactness.
Efforts to measure intactness to date have mostly mapped human influence that is visible from remotely sensed data (e.g., roads, human settlement, night lights, forest canopy intactness, etc.) or where global-scale data are available from other sources of human activity (e.g., human population density; livestock density, etc.), and thus provide measures of the human footprint on habitat/land cover change. Habitat loss and resulting ecosystem degradation is one of the key drivers of species loss (Chase et al., 2020) but it is not the only driver. While measurements of the Human Footprint (HF) on habitat are useful, they do not fully capture the effects of human activities on biodiversity. Overexploitation, effects of invasive species, and disease, for example, are more difficult to assess using HF-type datasets. In particular, hunting is one threat that is much more difficult to assess because hunters can penetrate large areas of intact habitat, spending days by moving camps, and this activity is impossible to detect remotely (Redford, 1992; Peres et al., 2006). Benítez-López et al. (2019) provided the first attempt to map the impact of hunting across the tropical forests of the world, developing a model of hunting impacts using data from many local hunting studies from South America, Africa, and Asia, and predicting mammal defaunation (both extirpation and reduction in density) across the tropical forested region. This analysis showed that at least 50% of the tropical forests likely have partial defaunation of their mammal biota, with 52% of Intact Forests and 62% of Wilderness Areas (where the Human Footprint ≤ 2) being partially depleted of large mammals. Clearly habitat loss and hunting have compounded impacts on mammal populations (Romero-Muñoz et al., 2020); reducing the area of occupied habitat by more than half, with 29% of the total area attributed to hunting activities (Gallego-Zamorano et al., 2020).
Where species have fallen below a “functional density” at a site, it would not be considered functionally intact because important biotic interactions are likely no longer fulfilling their ecological role (Soulé et al., 2003). Overexploitation, invasive species and disease may reduce species below functional densities leading to loss of functional intactness even if species are not completely extirpated. Functional densities are particularly important for species that play key ecological roles in an ecosystem, such as seed dispersal, nutrient cycling or top-down regulation (Estes et al., 2011; Camargo-Sanabria et al., 2013; Peres et al., 2016). Where mesopredators or competitive species have been introduced by humans, these can have an impact on both species composition and on ecosystem function (Gordon and Letnic, 2016). Assessment of functional intactness should therefore assess both loss of species at a site (faunal intactness) as well as decline in the functional role of individual species as a result of human influence.
To support identification of sites with outstanding ecological integrity, we provide an initial scoping analysis and explore how measuring habitat, faunal and functional intactness narrow down the areas where sites might be identified as having outstanding ecological integrity under KBA Criterion C. More detailed analyses specific to single ecoregions would be needed for a complete assessment of Criterion C sites. We assess how the percentage of land that would qualify for Criterion C KBA would change if we consider just habitat intactness or if we include faunal and functional intactness, which is arguably more difficult to measure. We compile data across multiple taxonomic groups, but the focus is primarily on mammals, particularly large- and medium-sized mammals, because these are often the first species to be affected by hunting (Redford, 1992; Atwood et al., 2020; Gallego-Zamorano et al., 2020). We then assess what proportion of the terrestrial surface of the world could be considered faunally and functionally intact at a minimum. We compiled three sets of data layers, representing areas that have low human footprint, are faunally intact, and functionally intact. We applied these layers cumulatively to identify areas that are ecologically intact (i.e., low human footprint and faunally intact and functionally intact). We then calculated the percentage of the earth’s land surface and the number/percent of ecoregion with at least one polygon remaining in the different grades of intactness, and identified areas with potential for restoration of ecological integrity through reintroduction of extirpated species.
Materials and Methods
To undertake a global scoping of ecological integrity we used three main data layers and an analytical process to: (1) Identify areas where human impact is low; (2) Assess where species have been historically extirpated; and (3) Assess where species densities are low where this is likely to have been caused by human influence.
Areas of Low Human Impact
We used the Human Footprint (HF) map (Venter et al., 2016a) as our index of human impact. The HF is a composite index of eight global pressures and ranges between 0 and 50. HF scores of four or more are considered modified habitat; scores below this are considered as low impact (Venter et al., 2016b; Mokany et al., 2020; Williams et al., 2020), although, in some recent papers, a value of below three is considered low impact (Di Marco et al., 2018; O’Bryan et al., 2020). The HF mainly maps human infrastructure, population or agriculture, but does attempt to predict impacts up to 15 km on either side of linear features (rivers and roads). For our scoping we selected a threshold value for the HF of four or lower, to ensure that sites with scores between three and four that might still meet faunal intactness were considered. Following the methodology in Plumptre et al. (2019), we smoothed the global HF scores before identifying where the HF score is equal to or lower than four. This was done to eliminate isolated micro-patches of high or low HF scores that would add to analysis time while providing no substantial benefit to global findings. We then converted the result to a vector file. This process developed our baseline map of estimated habitat intactness.
We calculated the percentage loss of area for each ecoregion (Dinerstein et al., 2017) and the average percentage loss across all ecoregions where there was habitat with a HF score ≤ 4. Criterion C sites are suggested to be large (10,000 km2 or larger). However, our initial results showed that few ecoregions can meet this size with complete functional intactness. We therefore also assessed how the number of ecoregions and the area of land surface would change when selecting sites with minimum areas of 1,000, 5,000, and 10,000 km2. We excluded Antarctica as it is not designated as an ecoregion but included within a category of rock and ice (Dinerstein et al., 2017).
Areas of Where Fauna Have Not Been Extirpated
We used three data layers that provided measures of species extirpation at a site: two global, and one focused on tropical forested areas. The first layer was an estimate of the number of historical mammal extinctions across the world (Mammal extirpations). The data for this were taken from Faurby et al. (2018), with the original data mainly coming from Faurby and Svenning (2015) and IUCN 2016-3. We note that these data are not explicitly linked to any specific time point but rather compare current ranges with estimated non-human modified ones. However, we omitted any global or continental extinctions happening before 1500 AD. Whilst a few species have had prolonged range contractions within the continents they still occupy, the data are generally an adequate approximation of the mammalian community if 1500 AD is used as a baseline and have previously been used as such (Belote et al., 2020). The resolution of this map was at ∼ 100 × 100 km and we downscaled it to 10 × 10 km for this study using a bilinear interpolation in ArcGIS 10.5.
The second layer was a map of extinct and possibly extinct range areas compiled from the IUCN Red List of Threatened Species for all taxonomic groups assessed by June 2020 (IUCN 2020 – henceforth IUCN Red List). The IUCN Red List assesses where species have been extirpated (extinct) since the year 1500 AD and this is recorded in the “Presence” field of the range polygons (Plumptre et al., 2019). We updated the map of all extinct ranges provided by Plumptre et al. (2019) for the IUCN Red List 2020 v2. Range mapping on the IUCN Red List is not consistent and has changed over time, so that not all species have their extinct range mapped, but it provides a measure across many taxonomic groups. For this analysis we excluded the extinct ranges of mammal species as these were included already in the first layer (Mammal extirpations). We refer to this layer as non-mammal extirpations (Non-mammal extirpations). Because IUCN Red List Assessors do not consistently map extirpated range for species when making assessments, we preferred to use the first layer for mammals as the mammal layer uses the same approach for nearly all mammal species.
To account for the localized effect of hunting, we used a third layer, a binary raster map of defaunation (i.e., defaunated or not) within tropical forest areas which represented where tropical mammals are predicted to be locally extirpated due to hunting pressure (Gallego-Zamorano et al., 2020). This dataset contains tropical mammal species with at least 95% of their range within the tropical biome. In its preparation, each species was modeled separately at 1 km resolution using presence information in hunted and unhunted areas. A binomial mixed-effects model that quantified the species-specific probability of persistence under hunting pressure was used to identify local extirpations per species (Gallego-Zamorano et al., 2020). We calculated this layer by aggregating all species to create a map of predicted numbers of large and medium mammals extirpated within the areas identified as “defaunated” in the binary map (Tropical forest hunting extirpations).
From these three extirpation maps we created two outputs: (1) A binary map where at least one species has been extirpated by combining all three maps and selecting all cells where extirpated species exceed zero; and (2) A raster of numbers of species extirpated at a scale of 10 × 10 km which was derived by summing the numbers of mammal extirpations (Belote et al., 2020) and the non-mammal extirpations generated for this analysis, scaled to the same 10 × 10 km resolution.
Assess Areas Where Species Are Reduced Below Functional Density
We compiled maps for widespread species for which predicted density or probability of suitable habitat has been modeled across their global range. We targeted wide-ranging large mammals (including large ungulates, great apes, and carnivores), and landscape species that are often the first species to be lost from an ecological community because of anthropogenic impacts (Sanderson et al., 2002b). While this is not a comprehensive mapping of potential reduction in densities of species across the world, these species may be considered indicators of ecological integrity because they are sensitive to human impacts. We assembled density data for the following species or subspecies: African forest elephant (Loxodonta africana cyclotis) (Maisels et al., 2013), western lowland gorilla (Gorilla gorilla gorilla) and central chimpanzee (Pan troglodytes troglodytes) (Strindberg et al., 2018), western chimpanzee (P.t.verus) (Heinicke et al., 2019), Grauer’s gorilla (Gorilla beringei graueri) (Plumptre et al., 2016), Bornean Orangutan (Pongo pygmaeus) (Voigt et al., 2018), Sumatran Orangutan (P. abelii) (Wich et al., 2016), Tapanuli Orangutan (P. tapanuliensis) (Nater et al., 2017), jaguar (Panthera onca) (Jȩdrzejewski et al., 2018), lion (Panthera leo) (Riggio et al., 2013), and African buffalo (Syncerus caffer) (Wint and Kiara, 2017). In addition, species distribution models showing probability of suitable habitat were compiled for eastern chimpanzee (P. t. schweinfurthii) (Plumptre et al., 2010), and four bear species: polar bear (Ursus maritimus), brown bear (Ursus arctos), American black bear (Ursus americanus) and Asian black bear (Ursus thibetanus) (Luna-Aranguré et al., 2020). Threshold densities or probabilities were identified for each species and for this study were assumed to predict where a species is at low abundance and will not be playing its functional role in the ecosystem (Supplementary Table 1). We compiled these into an overall map showing where densities are likely to be low (Selected large mammal reduced density).
As a second input layer to measure reduced density, Benítez-López et al. (2019) mapped areas of predicted defaunation (defaunation map) resulting from hunting across tropical forests. Their maps show the areas where mammal species are predicted to have declined in abundance, but do not identify areas of local extinction. However, the same data were used to predict where species have been extirpated by hunting (Gallego-Zamorano et al., 2020) – our Tropical Forest Hunting Extirpation map above – and we used this as a mask to remove areas from the defaunation map which have lost species, thereby leaving sites where species have been reduced in absolute abundance, but not lost. We used a threshold of <0.9 (i.e., more than 10% average decline in abundance across mammal species) as recommended by Benítez-López et al. (2019) on their defaunation score to represent areas where species were below functional density (Tropical forest hunting reduced density).
Analyses
We assessed the cumulative effects of these layers for potential Criterion C sites by sequentially applying the following filters to vector files that we had derived from the various maps described above (Figure 1):
1. Habitat intactness: Select all areas where Human Footprint ≤ 4;
2. Faunally intact: Subtract areas from 1 (Habitat intactness), where one or more species have been extirpated;
3. Functionally intact: Subtract areas from 2 (Faunally intact) where selected large mammal density is low.
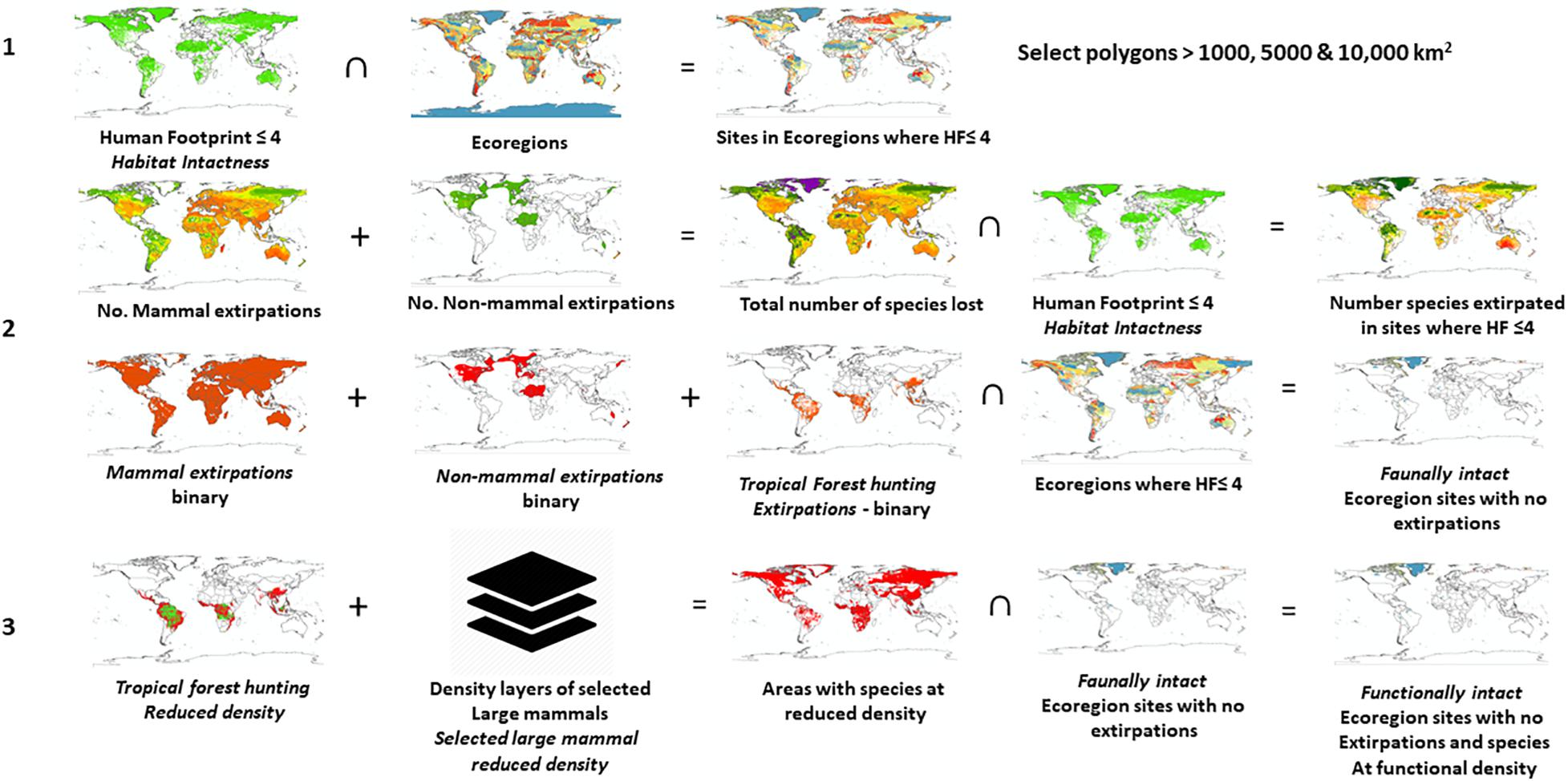
Figure 1. Diagram of the analysis process to calculate habitat intactness, faunal intactness, and functional intactness. (∩ = intersect with).
We assessed the effect of limiting sites to a minimum polygon area of 1,000, 5,000, and 10,000 km2 for each ecoregion. We calculated changes in numbers and percentage area of ecoregions and calculated median polygon area for each ecoregion that had at least one polygon for each minimum area class. We then calculated the total percentage of land that would meet a definition of faunal and functional intactness for each minimum area class.
We assessed which biomes of the world would contain areas of faunal intactness that are at least 10,000 km2 by intersecting the polygons generated above with the map of terrestrial biomes generated by Dinerstein et al. (2017) as attributes in their ecoregions shapefile. For the ecoregions that were identified as having functionally intact areas, we also assessed what percentage of these areas was covered by protected areas using the World Database of Protected Areas (UNEP-WCMC and IUCN, 2020) and percentage covered by existing KBAs using the World Database of KBAs (BirdLife International, 2020). We measured this coverage using the same threshold areas above to evaluate changes with minimum polygon area.
Finally, having mapped the number of species extirpated in each 10 km × 10 km cell, we also mapped the extent of land where Human Footprint ≤ 4 and where it might be possible to restore ecological integrity by assessing where few species (1–5) have been extirpated and might be reintroduced if they have not gone extinct globally. The number of species extirpated did not correlate well with the Human Footprint score (Supplementary Figure 1) but the requirement of Criterion C sites to be in areas with low human impact meant we restricted our analyses to the area of low human impact (Human footprint ≤ 4). We first removed the number that were globally extinct on the IUCN Red List from the map of number of species extirpated, so that we could assess areas where restoration might be possible in land where HF ≤ 4. Applying the three minimum area classes, the total area of a site was computed for sites incorporating 0–5 species extirpated (for example a 10,000 km2 site assessed for 2 species extirpated could contain areas within the 10,000 km2 where 0, 1 or 2 species were extirpated).
Results
Areas Under the Types of Intactness
Habitat intactness (as measured by HF ≤ 4), covered 43.4% of the terrestrial surface of the planet. When selecting areas of land exclusively based on the HF ≤ 4 score, only 214 of the 847 ecoregions (25.3%) qualify as having polygons of 10,000 km2 or larger. This rises to 464 ecoregions (54.8%) if areas as small as 1,000 km2 are considered. Removing each of the individual faunal layers from this baseline habitat intactness map for a minimum area of 10,000 km2 showed that most of the reduction in percentage of ecoregions or terrestrial land surface occurs as a result of mammal and non-mammal extirpations and the reduced densities of selected species (Figure 2).
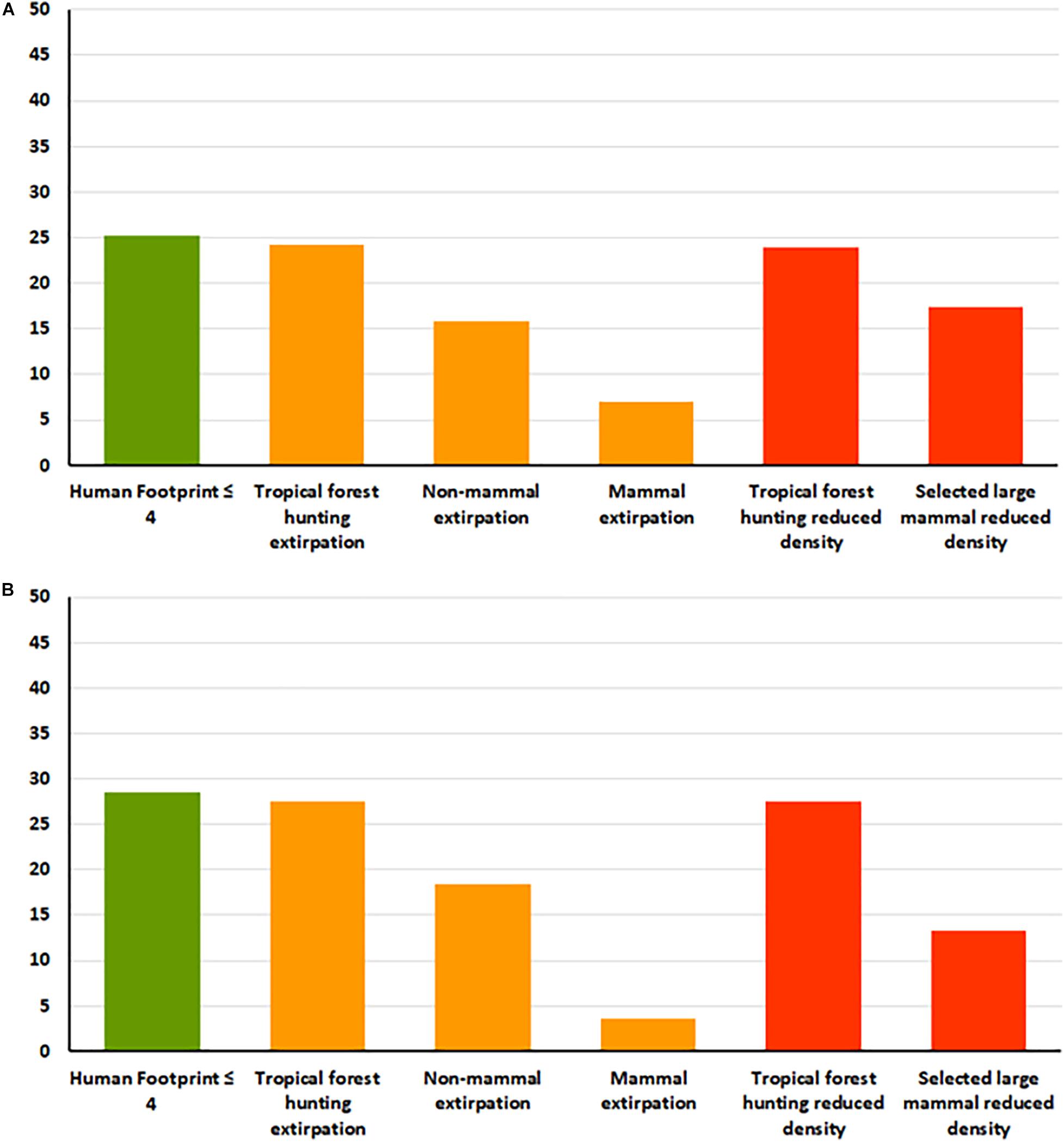
Figure 2. The effects of selecting a HF ≤ 4 (green) on the percentage of (A) ecoregions and (B) area of the land surface of the earth with the addition of three measures of faunal intactness (Orange – Intact fauna in tropical forest; non-mammal extirpations from the IUCN Red List; Mammal extirpations) and two measures of functional intactness (Red – Minimal impacts of hunting in tropical forest; High densities of selected large mammals). The percentages/areas are calculated as the amount remaining as each layer is selected within the HF ≤ 4 layer for polygon areas that are 10,000 km2 or larger.
Combining the three layers measuring extirpation (Tropical forest hunting extirpation, Non-mammal extirpation, and Mammal extirpation) into a single mask layer enabled us to assess faunal intactness. Removing regions where species have been lost from the HF ≤ 4 map has a huge impact, reducing the number of ecoregions to 42 at 10,000 km2 or 124 at 1,000 km2 (Figure 3A; Supplementary Figure 2). Only 2.9% of land surface (Figure 3B) could be considered faunally intact for a minimum area of 10,000 km2 (3.4% at 1,000 km2).
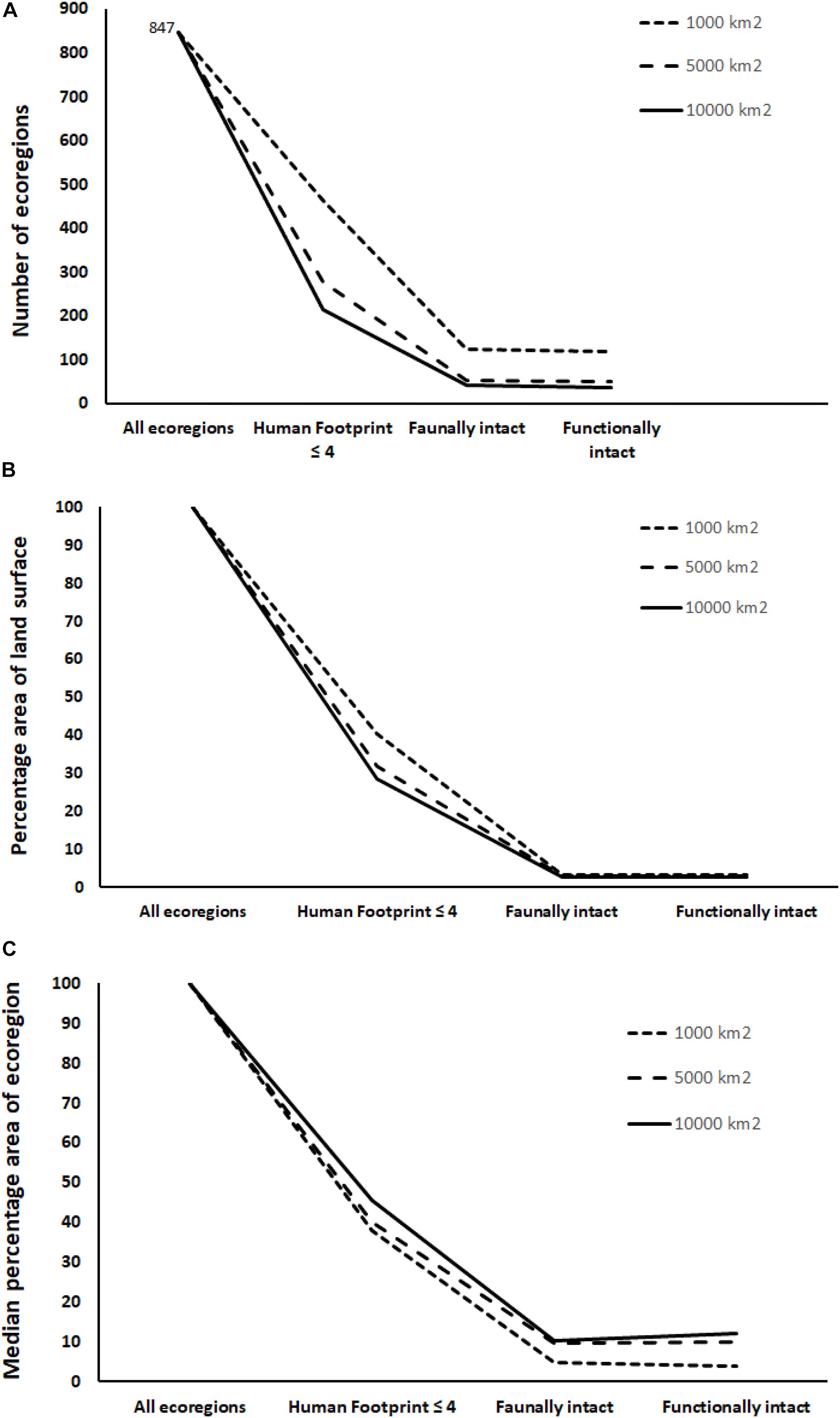
Figure 3. Changes in (A) number of ecoregions, (B) percentage area of terrestrial surface and (C) median percentage area of each ecoregion remaining as first HF ≤ 4, then faunally intact and finally functionally intact areas are selected, showing the cumulative loss with each layer. Results are shown for minimum areas of 1,000, 5,000, and 10,000 km2.
Finally, we combined the two layers measuring low density (tropical forest hunting reduced density and selected large mammal density) to assess functional intactness (i.e., faunally intact areas which also have functional densities of species). This reduced the number of ecoregions that would qualify further from 38 at 10,000 km2 to 119 at 1,000 km2 and only 2.8% of the land surface can be considered functionally intact for a minimum area of 10,000 km2 (3.3% at 1,000 km2).
Approximately 25% of the world’s terrestrial ecoregions have at least one habitat intact polygon >10,000 km2 remaining; whereas only 5.0% have at least one faunally intact polygon remaining; and 4.5% have at least one functionally intact polygon remaining. The median percentage area of ecoregions that remain in the analysis also drops as we sequentially filter for HF ≤ 4, faunal intactness, and functional intactness (Figure 3C). This shows that not only do the numbers of ecoregions decrease, but also the area remaining within the ecoregions that can be considered intact decreases greatly.
Biome Representation
A few biomes – tundra, deserts, tropical and subtropical grasslands – have an average of more than 40% of each ecoregion in faunally and ecologically intact areas of >10,000 km2. Other biomes have a much lower percentage (Figure 4 and Supplementary Figure 2).
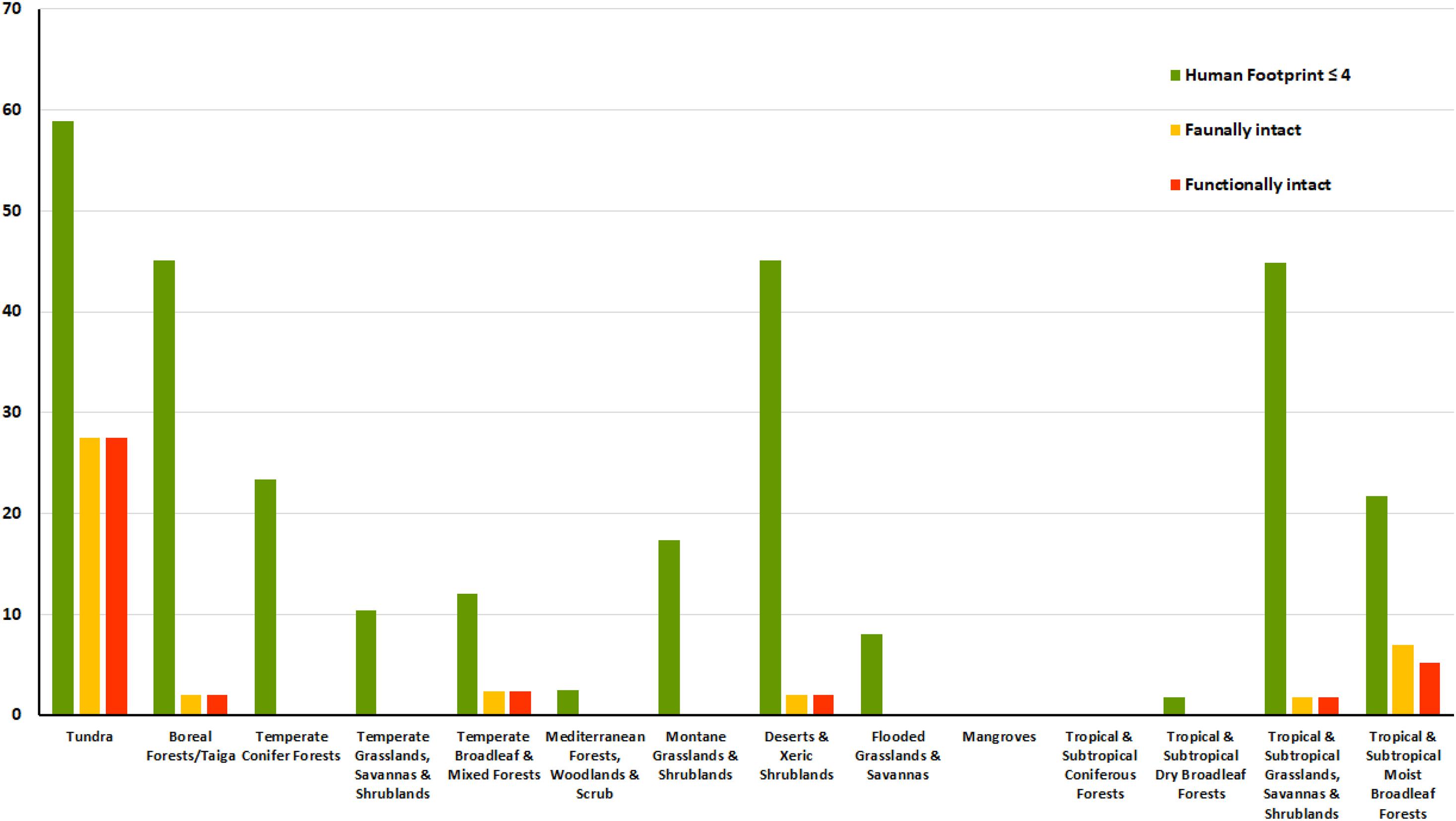
Figure 4. Changes in percentage of ecoregions in biomes selecting first areas of HF ≤ 4; then removing areas where species have been extirpated and finally removing areas where species are at low density. Results are presented for ecoregions with minimum polygon of 10,000 km2.
Proportion of Intact Land Covered by Protected Areas or KBAs
Only about 11% of the area identified as functionally intact is included in existing protected areas (categories I-IV). The median percentage of functionally intact land covered by protected areas of all types is larger but still only 34% (Figure 5). The percentage of functionally intact land area covered by KBAs identified with other KBA criteria is only 4% at 10,000 km2 but rises to 7% at 1,000 km2.
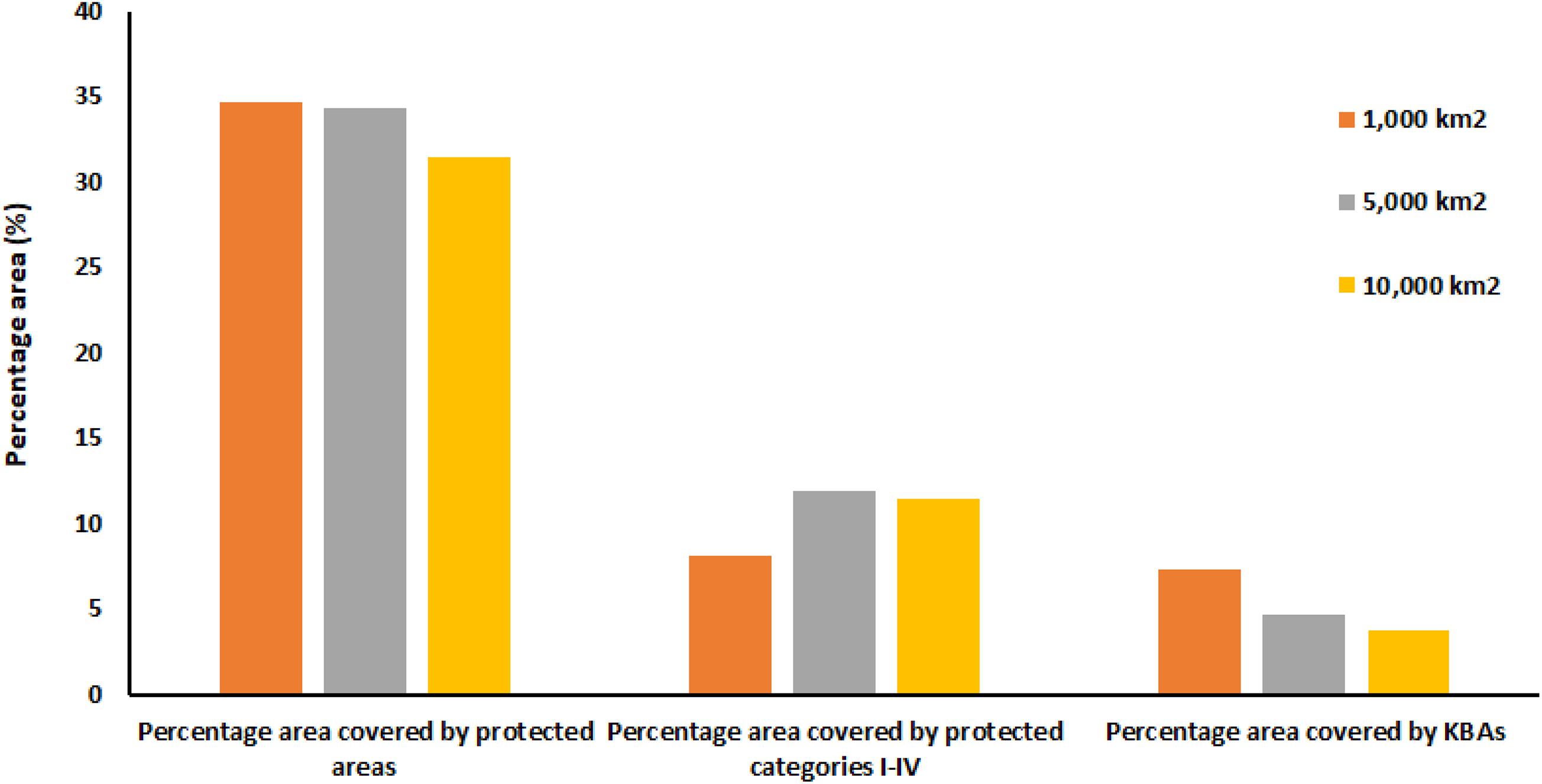
Figure 5. The percentage of ecologically intact sites (polygons) covered by protected areas of all types (left), protected areas in IUCN categories I-IV (center), and covered by KBAs (right).
Potential for Recovery of Functional Intactness
The very low percentage of land that might qualify as functionally intact led us to assess what remaining land exists where only five or fewer species have been extirpated (but are not globally extinct) within the area of habitat intactness (Figure 6). We calculated changes in land surface and number of ecoregions that could in future become faunally intact (and eventually functionally intact) if restoration occurred by sequentially adding 1, 2, up to 5 extant species where these have been extirpated (Table 1). Restoring 1–5 species would increase the percentage of the earth’s land surface that is faunally intact from approximately 3% at present up to 19% (10,000 km2 minimum area) to 24% (1,000 km2).
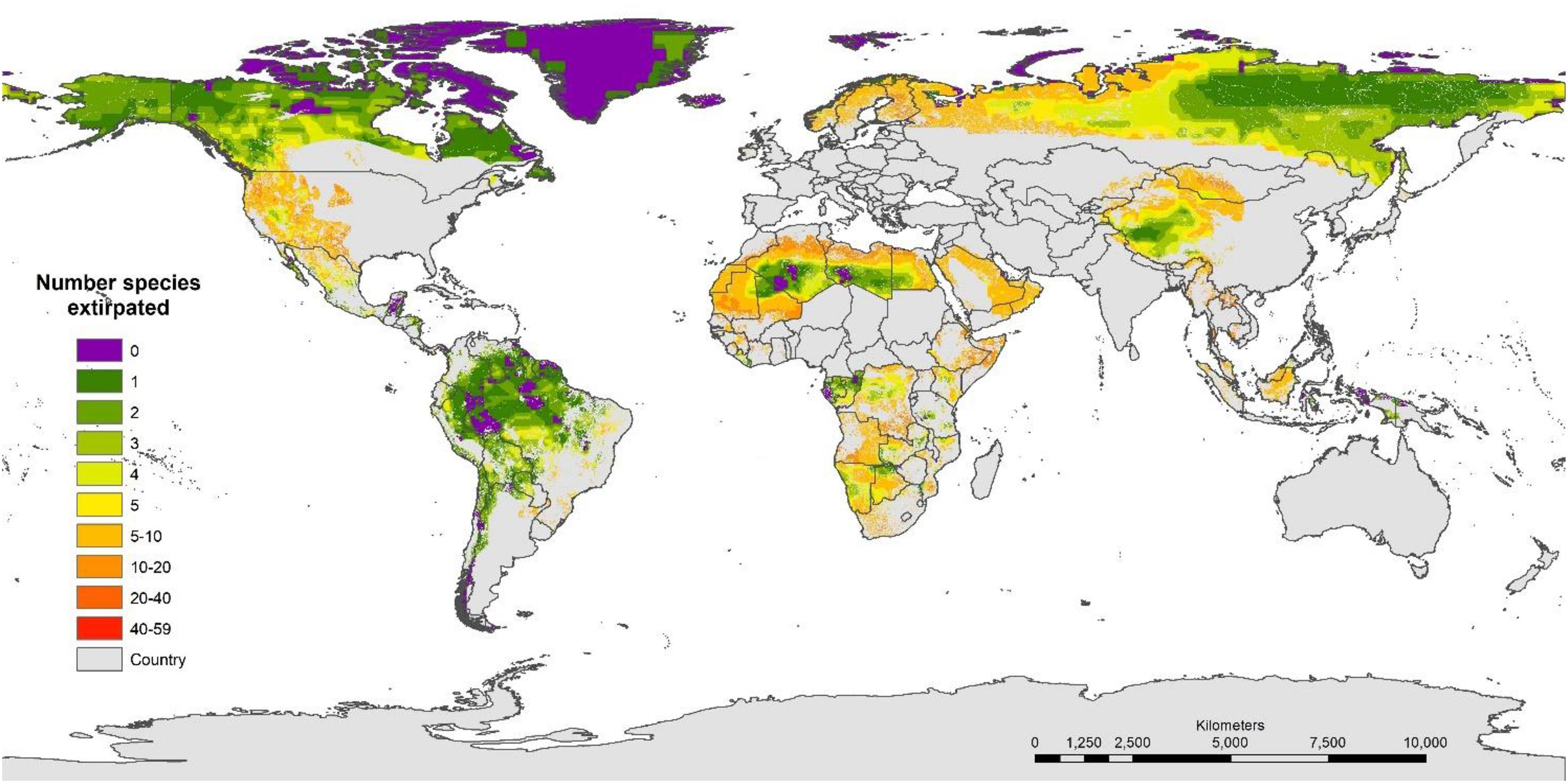
Figure 6. The number of extirpated species (mammals and all other taxa mapped on the IUCN Red List) mapped in areas of habitat intactness (Human Footprint ≤ 4) for the land surface of the earth (removing areas (gray) where species have become globally extinct since 1500 AD, HF >4, and Antarctica).
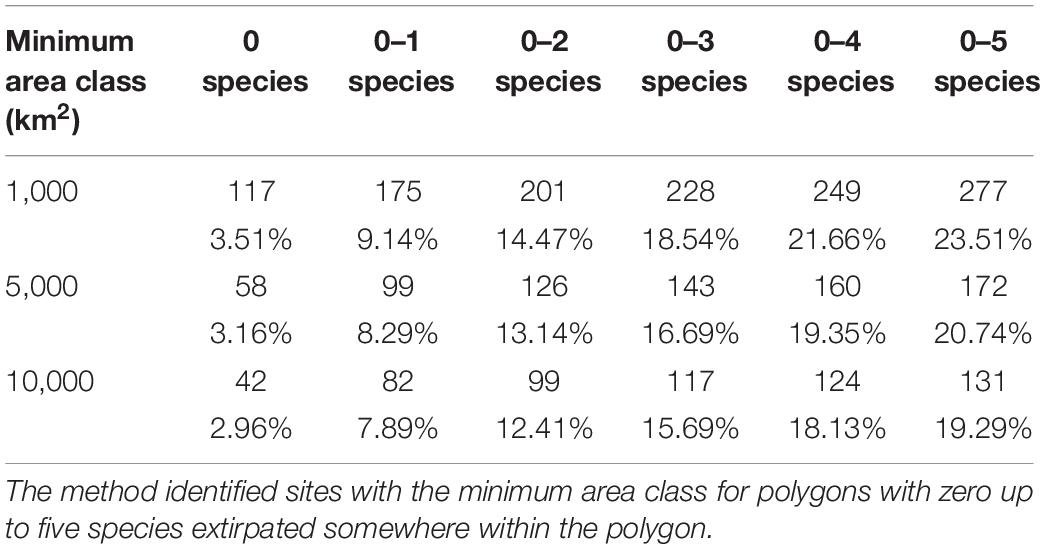
Table 1. The change in number of ecoregions (top row) and percentage land surface area (second row) that could potentially be conserved with the reintroduction of extant species that have been locally extirpated onto land where HF ≤ 4.
Discussion
While wilderness areas are increasingly recognized as important for biodiversity conservation, few areas of the world remain that can be characterized as having outstanding ecological integrity, (i.e., retaining intact species assemblages at ecologically functional densities). We found that only 2.8% of the terrestrial surface of the planet is represented in areas of 10,000 km2 or larger with low human footprint, no known species loss and no species known to be reduced below functional densities. This compares with estimates of 20–40% from mapped habitat intactness in the literature (Riggio et al., 2020).
The datasets we have applied in our analysis are the best we could compile at a global scale; we recognize that at an ecoregional scale it will be possible to make more detailed analyses of human footprint, identification of areas which have experienced species loss, and mapping of population densities of species likely impacted by humans to assess whether they are likely functional. In particular, we selected a subset of large mammals to assess functional density, choosing species that were relatively widespread and likely hunted, as surrogates for other species. Few species have their density mapped across their global range and we compiled what we could find that was relevant to the analysis. We strongly encourage more detailed analyses to be made to start scoping where ecologically functional KBAs might be identified and that sensitivity analyses are made using variation in the datasets. Detailed assessments will likely reduce the area that can be described as ecologically intact still further. However, detailed assessments may also reveal that smaller sites are sufficient to maintain populations and ecological processes in a few ecoregions, especially on islands or island-like ecosystems with few area-demanding species. For example, the East African montane moorlands ecoregion is a high altitude ecoregion where few large-bodied fauna occur naturally. The total area of this ecoregion is less than 5,000 km2 but it could potentially have areas that are considered functionally intact. We therefore explored the effect of relaxing the area requirement, but emphasize that sites that are smaller than 10,000 km2 would still need to maintain functionally intact populations of all species for that ecoregion to qualify as KBAs under Criterion C.
A surprisingly low percentage of the area identified as ecologically intact in our analysis is covered by protected areas (11% covered by PAs in categories I-IV), indicating that Other Effective Conservation Measures (OECMs) have an important role to play in maintaining ecological integrity. Many of the areas identified as ecologically intact coincide with territories managed by indigenous communities, who have played a vital role in maintaining the ecological integrity of these areas (Figure 6). We show that there is potential to restore ecological integrity over a much broader area. We found that up to 20% of the earth’s land surface could be restored to faunal intactness through the reintroduction of up to five species. Clearly restoring faunal intactness will involve meeting the requirements for successful species’ reintroduction and the ability to build up population numbers so that they can reach functional densities. Applying the IUCN/SSC reintroduction and translocation guidelines (IUCN/SSC, 2013) will be necessary to minimize disease risks and possible conflicts with people, but our findings show there is potential to increase faunal intactness, and ultimately functional intactness over time, if species can be reintroduced to sites with low human footprint. In this “decade of restoration” (Aronson et al., 2020), we believe that, as well as focusing on restoring habitat, a proactive approach should assess which areas would be amenable to restore faunal intactness with the aim of regaining full functional intactness, by tackling the threats that led to the loss in the first place. These areas will likely be located in eastern Russia, northern Canada and Alaska, the Amazon Basin, parts of the Sahara, and the Congo Basin. Planning for ecological restoration to include the reintroduction of extirpated but globally extant species, as has occurred in several conservation projects around the world (Hale and Koprowski, 2018), could significantly increase the number of ecoregions that could achieve sites with outstanding ecological integrity. Examples would include reintroducing forest elephants in areas of the Congo Basin where they have been extirpated, or reintroducing some of the large ungulates that have been lost from much of Africa’s woodlands and savannas because of overhunting (e.g., buffalo, giraffe, zebras etc.), as long as overhunting has ceased. We acknowledge that other key factors need be taken into account for successful species reintroduction projects, including but not limited to adequate source populations, pedigree and genetics, and disease assessments (Dinca et al., 2018; Hunter-Ayad et al., 2020). Restoration projects should also consider assessing which ecoregions currently cannot meet the requirement of faunal intactness and determine where they could best support restoration in these ecoregions to rebuild faunal intactness and, over time, functional intactness as densities rebuild.
Surprisingly, human footprint indices did not correlate well with faunal intactness (Supplementary Figure 1), because while threats such as habitat loss may correlate well, several major threats to faunal intactness (e.g., hunting, invasive species) are not properly accounted for by the human footprint maps (Benítez-López et al., 2019; Belote et al., 2020). There is considerable geographic variation in patterns of hunting pressure, but many tropical forest regions characterized by relatively low human densities and land cover change have also suffered from extensive and unsustainable hunting pressure (Redford, 1992; Tilker et al., 2019); invasive predators have encroached into wilderness areas far from infrastructure networks (Doherty et al., 2016); and diseases such as ebola can spread deep into remote forests (Rizkalla et al., 2007). None of these threats are picked up by remote sensing tools. Threats such as hunting primarily affect large- and medium-sized mammals (Benítez-López et al., 2019; Gallego-Zamorano et al., 2020), although regional variations exist because of hunters’ preferences, cultural beliefs, and taboos regarding the consumption of some species (Ngoufo et al., 2014; Bobo et al., 2015). For instance, in Uganda ungulates but not primates are targeted by hunters, unlike just over the border in the Democratic Republic of Congo where primates are widely hunted as well (A. Plumptre pers. obs.). Along the Gabon and Congo coastline, chimpanzees but not gorillas are considered taboo, and this is reversed in some areas of Cameroon; a factor “eat gorillas” or “eat chimpanzees” was included in the spatial model that predicted the density of both taxa in western Equatorial Africa (Strindberg et al., 2018). Predicting the impacts of hunting needs much more detailed analysis (Benítez-López et al., 2019; Bogoni et al., 2020; Romero-Muñoz et al., 2020) than making assumptions about impacts from access distance to road and river networks. Similarly, the impacts of introduced species cannot be assessed from maps such as the Human Footprint but have had a major impact on extirpated species numbers in places such as Australia (Woinarski et al., 2015).
Nevertheless, human footprint indices can play an essential role in identifying areas with potential for ecological restoration through reintroduction of extirpated species, as there will typically be fewer barriers to successful restoration in areas with low human footprint. As the world develops the next goals and targets for the Convention on Biological Diversity under the post 2020 global biodiversity framework, “intact habitat” has been recognized as an important target. We would argue here that “intact habitat” is clearly an important conservation target, and that we must also consider faunal and functional intactness, and direct resources to areas that still retain a fully functioning ecological community as this is clearly very rare. Retaining large areas will give these sites a better chance of retaining their functional intactness. Working to restore ecological integrity to those areas of the world that are close to ecological integrity should be a focus of the current decade of restoration also. Moreover, identifying Criterion C KBAs could help focus attention on these sites for conservation action and investment.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
AP and DB conceived the original idea, compiled layers, made analyses, and drafted manuscript. RB, EV-D, SF, WJ, HKi, HKü, AB-L, CL-A, MV, SW, WW, and JG-Z provided data layers and contributed to the manuscript. CB contributed ideas on manuscript structure and analyses. All authors contributed to the article and approved the submitted version.
Funding
AP was supported by the KBA Partnership. DB supported by Daniel K. Thorne Foundation and CCI Collaborative Fund. SF received funding from the Swedish Research Council (2017-03862). AB-L was supported by a Juan de la Cierva-Incorporación grant (IJCI-2017-31419) from the Spanish Ministry of Science, Innovation and Universities.
Disclaimer
The views implied or expressed here are those of the authors and do not necessarily represent the views of the institutions they work for or represent and are meant to inform, but not pre-empt, planned workshops to discuss the application of KBA criterion C.
Conflict of Interest
WW is employed by a consultancy, Environmental Research Group Oxford Ltd. which has no potential conflict of interest with these research topics and provided input free of charge.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was undertaken by AJP while working for the KBA Partnership as head of the KBA Secretariat and he wishes to thank BirdLife International and the KBA Partnership who support his position.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2021.626635/full#supplementary-material
Supplementary Figure 1 | Plot of number of species extirpated against human footprint score for 3,474 randomly assigned points across the land surface of the planet. While a positive linear correlation exists R2 is only 0.21.
Supplementary Figure 2 | Location of land with (A) HF ≤ 4 and (B) faunal intactness. Colors represent different ecoregions, gray is land that does not meet the criteria.
Supplementary Table 1 | The species name, threshold density and source of data for rangewide species distribution maps for the species that were used to assess impacts on functional intactness.
References
Aronson, J., Goodwin, N., Orlando, L., Eisenberg, C., and Cross, A. T. (2020). A world of possibilities: six restoration strategies to support the United Nation’s Decade on Ecosystem Restoration. Restor. Ecol. 28, 730–736. doi: 10.1111/rec.13170
Atwood, T. B., Valentine, S. A., Hammill, E., McCauley, D. J., Madin, E. M., Beard, K. H., et al. (2020). Herbivores at the highest risk of extinction among mammals, birds, and reptiles. Sci. Adv. 6:eabb8458. doi: 10.1126/sciadv.abb8458
Belote, R. T., Faurby, S., Brennan, A., Carter, N. H., Dietz, M. D., Hahn, B., et al. (2020). Mammal species composition reveals new insights into earth’s remaining wilderness. Front. Ecol. Environ. 18, 376–383. doi: 10.1002/fee.2192
Benítez-López, A., Santini, L., Schipper, A. M., Busana, M., and Huijbregts, M. A. (2019). Intact but empty forests? Patterns of hunting-induced mammal defaunation in the tropics. PLoS Biol. 17:e3000247. doi: 10.1371/journal.pbio.3000247
BirdLife International (2020). World Database of Key Biodiversity Areas. Developed by the KBA Partnership: BirdLife International, International Union for the Conservation of Nature, Amphibian Survival Alliance, Conservation International, Critical Ecosystem Partnership Fund, Global Environment Facility, Global Wildlife Conservation, NatureServe, Rainforest Trust, Royal Society for the Protection of Birds, Wildlife Conservation Society and World Wildlife Fund. September 2020 version. Available online at: http://www.keybiodiversityareas.org/site/search (accessed September, 2020).
Bobo, K. S., Aghomo, F. F. M., and Ntumwel, B. C. (2015). Wildlife use and the role of taboos in the conservation of wildlife around the Nkwende Hills Forest Reserve; South-west Cameroon. J. Ethnobiol. Ethnomed. 11:2. doi: 10.1186/1746-4269-11-2
Bogoni, J. A., Peres, C. A., and Ferraz, K. M. (2020). Extent, intensity and drivers of mammal defaunation: a continental-scale analysis across the Neotropics. Sci. Rep. 10, 1–16. doi: 10.1038/s41598-020-72010-w
Brodie, J. F., Redford, K. H., and Doak, D. F. (2018). Ecological function analysis: incorporating species roles into conservation. Trends Ecol. Evol. 33, 840–850. doi: 10.1016/j.tree.2018.08.013
Camargo-Sanabria, A. A., Mendoza, E., Guevara, R., Martınez-Ramos, M., and Dirzo, R. (2015). Experimental defaunation of terrestrial mammalian herbivores alters tropical rainforest understorey diversity. Proc. R. Soc. B 282:20142580. doi: 10.1098/rspb.2014.2580
CBD (2020). Update of the Zero Draft of the Post2020 Global Biodiversity Framework. CBD/POST2020/PREP/2/1. Available online at: https://www.cbd.int/conferences/post2020/post2020-prep-01/documents (accessed September, 2020).
Chase, J. M., Blowes, S. A., Knight, T. M., Gerstner, K., and May, F. (2020). Ecosystem decay exacerbates biodiversity loss with habitat loss. Nature 584, 238–243. doi: 10.1038/s41586-020-2531-2
Di Marco, M., Venter, O., Possingham, H. P., and Watson, J. E. M. (2018). Changes in human footprint drive changes in species extinction risk. Nat. Commun. 9:4621. doi: 10.1038/s41467-018-07049-5
Dinca, V., Balint, Z., Voda, R., Dapporto, L., Hebert, P. D. N., and Vila, R. (2018). Use of genetic, climatic, and microbiological data to inform reintroduction of a regionally extinct butterfly. Conserv. Biol. 32, 828–837. doi: 10.1111/cobi.13111
Dinerstein, E., Olson, D., Joshi, A., Vynne, C., Burgess, N. D., Wikramanayake, E., et al. (2017). An ecoregion-based approach to protecting half the terrestrial realm. Bioscience 67, 534–545. doi: 10.1093/biosci/bix014
Doherty, T. S., Glen, A. S., Nimmo, D. G., Ritchie, E. G., and Dickman, C. R. (2016). Invasive predators and global biodiversity loss. Proc. Natl. Acad. Sci. U.S.A. 113, 11261–11265. doi: 10.1073/pnas.1602480113
Estes, J. A., Terborgh, J., Brashares, J. S., Power, M. E., Berger, J., Bond, W. J., et al. (2011). Trophic downgrading of planet Earth. Science 333, 301–306. doi: 10.1126/science.1205106
Faurby, S., Davis, M., Pedersen, R. Ø, Schowanek, S. D., Antonelli, A., and Svenning, J. -C. (2018). PHYLACINE 1.2: the phylogenetic atlas of mammal macroecology. Ecology 99:2626. doi: 10.1002/ecy.2443
Faurby, S., and Svenning, J. C. (2015). A species-level phylogeny of all extant and late Quaternary extinct mammals using a novel heuristic-hierarchical Bayesian approach. Mol. Phylogenet. Evol. 84, 14–26. doi: 10.1016/j.ympev.2014.11.001
Gallego-Zamorano, J., Benítez-López, A., Santini, L., Hilbers, J. P., Huijbregts, M. A. J., and Schipper, A. M. (2020). Combined effects of land use and hunting on distributions of tropical mammals. Conserv. Biol. 34, 1271–1280. doi: 10.1111/cobi.13459
Goldewijk, K. K., Beusen, A., Doelman, J., and Stehfast, E. (2017). Anthropogenic land use estimates for the Holocene – HYDE 3.2. Earth Syst. Sci. Data 9, 927–953. doi: 10.5194/essd-9-927-2017
Gordon, C. E., and Letnic, M. (2016). Functional extinction of a desert rodent: implications for seed fate and vegetation dynamics. Ecography 39, 815–824. doi: 10.1111/ecog.01648
Grantham, H. S., Duncan, A., Evans, T. D., Jones, K. R., Beyer, H. L., Schuster, R., et al. (2020) Anthropogenic modification of forests means only 40% of remaining forests have high ecosystem integrity. Nat. Commun. 11, 5978. doi: 10.1038/s41467-020-19493-3
Hale, S. L., and Koprowski, J. L. (2018). Ecosystem-level effects of keystone species reintroduction: a literature review. Restor. Ecol. 26, 439–445. doi: 10.1111/rec.12684
Heinicke, S., Mundry, R., Boesch, C., Amarasekaran, B., Barrie, A., Brncic, T., et al. (2019). Advancing conservation planning for western chimpanzees using IUCN SSC A.P.E.S.—the case of a taxon-specific database. Environ. Res. Lett. 14:064001. doi: 10.1088/1748-9326/ab1379
Hunter-Ayad, J., Ohlemüller, R., Recio, M. R., and Seddon, P. J. (2020). Reintroduction modelling: a guide to choosing and combining models for species reintroductions. J. Appl. Ecol. 57, 1233–1243. doi: 10.1111/1365-2664.13629
IUCN (2016). A Global Standard for the Identification of Key Biodiversity Areas, Version 1.0, 1st Edn. Gland: IUCN.
IUCN (2020). The IUCN Red List of Threatened Species. Version 2020-v2. Available online at: http://www.iucnredlist.org (accessed June, 2020).
IUCN/SSC (2013). Guidelines for Reintroductions and Other Conservation Translocations. The Reintroduction and Invasive Species Specialist Groups’ Task Force on Moving Plants and Animals for Conservation Purposes Version 1.0. Gland: IUCN Species Survival Commission.
Jacobson, A. P., Riggio, J., Tait, A. M., and Baillie, J. E. M. (2019). Global areas of low human impact (‘Low impact areas’) and fragmentation of the natural world. Sci. Rep. 9:14179. doi: 10.1038/s41598-019-50558-6
Jȩdrzejewski, W., Robinson, H. S., Abarca, M., Zeller, K. A., Velasquez, G., Paemelaere, E. A. D., et al. (2018). Estimating large carnivore populations at global scale based on spatial predictions of density and distribution - Application to the jaguar (Panthera onca). PLoS One 13:0194719. doi: 10.1371/journal.pone.0194719
Kennedy, C. M., Oakleaf, J. R., Theobald, D. M., Baruch-Mordo, S., and Kiesecker, J. (2019). Managing the middle: a shift in conservation priorities based on the global human modification gradient. Glob. Change Biol. 25, 811–826. doi: 10.1111/gcb.14549
Kennedy, C. M., Oaklefa, J. R., Theobald, D. M., Baruch-Mordo, S., and Kiesecker, J. (2018). Global Human Modification. Available online at: https://doi.org/10.6084/m9.figshare.7283087 (accessed September, 2020).
Luna-Aranguré, C., Soberón, J., and Vázquez-Domínguez, E. (2020). A tale of four bears: environmental signal on the phylogeographical patterns within the extant Ursus species. J. Biogeogr. 47, 472–486. doi: 10.1111/jbi.13752
Maisels, F., Strindberg, S., Blake, S., Wittemyer, G., Hart, J., Williamson, E. A., et al. (2013). Devastating decline of forest Elephants in Central Africa. PLoS One 8:e59469. doi: 10.1371/journal.pone.0059469
McCloskey, J. M., and Spalding, H. (1989). A reconnaissance level inventory of the amount of wilderness remaining in the world. Ambio 18, 221–227.
Mittermeier, R. A., Myers, N., Thomsen, J. B., Gustavo, A. B. D. F., and Olivieri, S. (1998). Biodiversity hotspots and major tropical wilderness areas: approaches to setting conservation priorities. Conserv. Biol. 12, 516–520.
Mokany, K., Ferrier, S., Harwood, T. D., Ware, C., and Di Marco, M. (2020). Reconciling global priorities for conserving biodiversity habitat. Proc. Natl. Acad. Sci. 117, 9906–9911.
Morrison, J. C., Sechrest, W., Dinerstein, E., Wilcove, D. S., and Lamoreux, J. F. (2007). Persistence of large mammal faunas as indicators of global human impacts. J. Mammal. 88, 1363–1380. doi: 10.1644/06-MAMM-A-124R2.1
Nater, A., Mattle-Greminger, M. P., Nurcahyo, A., Nowak, M. G., de Manuel, M., Desai, T., et al. (2017). Morphometric, behavioral, and genomic evidence for a new orangutan species. Curr. Biol. 27, 3487–3498. doi: 10.1016/j.cub.2017.09.047
Ngoufo, R., Yongyeh, N., Obioha, E., Bobo, K., Jimoh, S., and Waltert, M. (2014). Social norms and cultural services-community belief system and use of wildlife products in the Northern periphery of the Korup National Park, South-West Cameroon. Change Adapt. Socio-Ecol. Syst. 1, 26–34. doi: 10.2478/cass-2014-0003
O’Bryan, C. J., Allan, J. R., Holden, M., Sanderson, C., Venter, O., Di Marco, M., et al. (2020). Intense human pressure is widespread across terrestrial vertebrate ranges. Glob. Ecol. Conserv. 21:e00882. doi: 10.1016/j.gecco.2019.e00882
Peres, C. A., Barlow, J., and Laurance, W. F. (2006). Detecting anthropogenic disturbance in tropical forests. Trends Ecol. Evol. 21, 227–229. doi: 10.1016/j.tree.2006.03.007
Peres, C. A., Emilio, T., Schietti, J., Desmoulière, S. J., and Levi, T. (2016). Dispersal limitation induces long-term biomass collapse in overhunted Amazonian forests. Proc. Natl. Acad. Sci. U.S.A. 113, 892–897. doi: 10.1073/pnas.1516525113
Plumptre, A. J., Baisero, D., Jêdrzejewski, W., Kühl, H., Maisels, F., Ray, J. C., et al. (2019). Are we capturing faunal intactness? A comparison of intact forest landscapes and the “last of the wild in each ecoregion. Front. For. Glob. Change 2:24. doi: 10.3389/ffgc.2019.00024
Plumptre, A. J., Nixon, S., Kujirakwinja, D. K., Vieilledent, G., Critchlow, R., Williamson, E. A., et al. (2016). Catastrophic decline of World’s largest primate: 80% loss of grauer’s Gorilla (Gorilla beringei graueri) population justifies critically endangered status. PLoS One 11:e0162697. doi: 10.1371/journal.pone.0162697
Plumptre, A. J., Rose, R., Nangendo, G., Williamson, E. A., Didier, K., Hart, J., et al. (2010). Status Survey and Conservation Action Plan for the Eastern Chimpanzee (Pan troglodytes schweinfurthii). Gland: IUCN.
Potapov, P., Hansen, M. C., Laestadius, L., Turubanova, S., Yaroshenko, A., Thies, C., et al. (2017). The last frontiers of wilderness: tracking loss of intact forest landscapes from 2000 to 2013. Sci. Adv. 3:e1600821. doi: 10.1126/sciadv.1600821
Potapov, P., Yaroshenko, A., Turubanova, S., Dubinin, M., Laestadius, L., Thies, C., et al. (2008). Mapping the world’s intact forest landscapes by remote sensing. Ecol. Soc. 13:51.
Riggio, J., Baillie, J. E. M., Brumby, S., Ellis, E., Kennedy, C. M., Oakleaf, J. R., et al. (2020). Global human influence maps reveal clear opportunities in conserving Earth’s remaining intact terrestrial ecosystems. Glob. Change Biol. 26, 4344–4356. doi: 10.1111/gcb.15109
Riggio, J., Jacobson, A., Dollar, L., Bauer, H., Becker, M., Dickman, A., et al. (2013). The size of savannah Africa: a lion’s (Panthera leo) view. Biodivers. Conserv. 22, 17–35. doi: 10.1007/s10531-012-0381-4
Rizkalla, C., Blanco-Silva, F., and Gruver, S. (2007). Modeling the impact of ebola and bushmeat hunting on Western Lowland Gorillas. EcoHealth 4, 151–155. doi: 10.1007/s10393-007-0096-2
Romero-Muñoz, A., Benítez-López, A., Zurell, D., Baumann, M., Camino, M., Decarre, J., et al. (2020). Increasing synergistic effects of habitat destruction and hunting on mammals over three decades in the Gran Chaco. Ecography 43, 1–13. doi: 10.1111/ecog.05053
Sanderson, E. W., Jaiteh, M., Levy, M. A., Redford, K. H., Wannebo, A. V., and Woolmer, G. (2002a). The human footprint and the last of the wild. Bioscience 52, 891–904.
Sanderson, E. W., Redford, K. H., Vedder, A., Coppolillo, P. B., and Ward, S. E. (2002b). A conceptual model for conservation planning based on landscape species requirements. Landscape Urb. Plann. 58, 41–56. doi: 10.1016/S0169-2046(01)00231-6
Sandom, C., Faurby, S., Sandel, B., and Svenning, J.-C. (2014). Global late Quaternary megafauna extinctions linked to humans, not climate change. Proc. Biol. Sci. 281, 1–9. doi: 10.1098/rspb.2013.3254
Soulé, M. E., Estes, J. A., Berger, J., and Del Rio, C. M. (2003). ‘Ecological effectiveness: conservation goals for interactive species’. Conserv. Biol. 17, 1238–1250. doi: 10.1046/j.1523-1739.2003.01599.x
Strindberg, S., Maisels, F., Williamson, E. A., Blake, S., Stokes, E. J., and Aba’a, R. (2018). Guns, germs, and trees determine density and distribution of gorillas and chimpanzees in Western Equatorial Africa. Sci. Adv. 4:eaar2964. doi: 10.1126/sciadv.aar2964
Tilker, A., Abrams, J. F., Mohamed, A., Nguyen, A., Wong, S. T., Sollmann, R., et al. (2019). Habitat degradation and indiscriminate hunting differentially impact faunal communities in the Southeast Asian tropical biodiversity hotspot. Commun. Biol. 2:396. doi: 10.1038/s42003-019-0640-y
UNEP-WCMC, and IUCN (2020). Protected Planet: The World Database on Protected Areas (WDPA). Cambridge: UNEP-WCMC and IUCN.
Venter, O., Sanderson, E. W., Magrach, A., Allan, J. R., Beher, J., Jones, K. R., et al. (2016b). Sixteen years of change in the global terrestrial human footprint and implications for biodiversity conservation. Nat. Commun. 7:12558. doi: 10.1038/ncomms12558
Venter, O., Sanderson, E. W., Magrach, A., Allan, J. R., Beher, J., Jones, K. R., et al. (2016a). Data from: global terrestrial Human Footprint maps for 1993 and 2009. Dryad Digital Repository doi: 10.5061/dryad.052q5.2
Voigt, M., Wich, S. A., Ancrenaz, M., Meijaard, E., Abram, N., Banes, G. L., et al. (2018). Global demand for natural resources eliminated more than 100,000 bornean orangutans. Curr. Biol. 28, 761–769. doi: 10.1016/j.cub.2018.01.053
Watson, J. E. M., Evans, T., Venter, O., Williams, B., Tulloch, A., Stewart, C., et al. (2018). The exceptional value of intact forest ecosystems. Nat. Ecol. Evol. 2, 599–610. doi: 10.1038/s41559-018-0490-x
Watson, J. E. M., Shanahan, D. F., Di Marco, M., Allan, J., Laurance, W. F., Sanderson, E. W., et al. (2016). Catastrophic declines in wilderness areas undermine global environment targets. Curr. Biol. 26, 2929–2934. doi: 10.1016/j.cub.2016.08.049
Wich, S., Singleton, I., Nowak, M. G., Utami Atmoko, S. S., Nisam, G., Arif, S. M., et al. (2016). Land-cover changes predict steep declines for the Sumatran orangutan (Pongo abelii). Sci. Adv. 2:e1500789. doi: 10.1126/sciadv.1500789
Williams, B. A., Venter, O., Allan, J. R., Atkinson, S. C., Rehbein, J. A., Ward, M., et al. (2020). Change in terrestrial human footprint drives continued loss of intact ecosystems. One Earth 3, 371–382. doi: 10.1016/j.oneear.2020.08.009
Wint, W., and Kiara, H. (2017). Where Should the Muguga Cocktail be Used? The Distributions of Theileria Parva, its Hosts and Vectors in Africa. Unpublished Report of International Livestock Research Institute. Available online at: https://cgspace.cgiar.org/handle/10568/103218 (accessed October, 2020).
Keywords: ecological integrity, KBA, human impact, species restoration, faunal intactness, functional intactness
Citation: Plumptre AJ, Baisero D, Belote RT, Vázquez-Domínguez E, Faurby S, Jȩdrzejewski W, Kiara H, Kühl H, Benítez-López A, Luna-Aranguré C, Voigt M, Wich S, Wint W, Gallego-Zamorano J and Boyd C (2021) Where Might We Find Ecologically Intact Communities? Front. For. Glob. Change 4:626635. doi: 10.3389/ffgc.2021.626635
Received: 09 November 2020; Accepted: 17 March 2021;
Published: 15 April 2021.
Edited by:
Manuel R. Guariguata, Center for International Forestry Research (CIFOR), IndonesiaReviewed by:
Paola Isaacs, Alexander von Humboldt Biological Resources Research Institute, ColombiaRenato Crouzeilles, International Institute for Sustainability, Australia
Marina Schmoeller, Federal University of Rio de Janeiro, Brazil
Copyright © 2021 Plumptre, Baisero, Belote, Vázquez-Domínguez, Faurby, Jȩdrzejewski, Kiara, Kühl, Benítez-López, Luna-Aranguré, Voigt, Wich, Wint, Gallego-Zamorano and Boyd. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andrew J. Plumptre, YXBsdW1wdHJlQGtleWJpb2RpdmVyc2l0eWFyZWFzLm9yZw==
†ORCID: Andrew J. Plumptre, orcid.org/0000-0002-9333-4047; Daniele Baisero, orcid.org/0000-0002-1266-7174; Ella Vázquez-Domínguez, orcid.org/0000-0001-6131-2014; Soren Faurby, orcid.org/0000-0002-2974-2628; Włodzimierz Jȩdrzejewski, orcid.org/0000-0002-9934-0683; Ana Benítez-López, orcid.org/0000-0002-6432-1837; Carlos Luna-Aranguré, orcid.org/0000-0002-7493-2092; Juan Gallego-Zamorano, orcid.org/0000-0003-2317-5148; Charlotte Boyd, orcid.org/0000-0003-4095-9390