 Deborah Lawrence1*
Deborah Lawrence1* Wayne Walker
Wayne Walker Louis Verchot
Louis Verchot- 1Department of Environmental Sciences, University of Virginia, Charlottesville, VA, United States
- 2The Woodwell Climate Research Center, Falmouth, MA, United States
- 3The Alliance of Bioversity International and the International Center for Tropical Agriculture, Cali, Colombia
Climate policy has thus far focused solely on carbon stocks and sequestration to evaluate the potential of forests to mitigate global warming. These factors are used to assess the impacts of different drivers of deforestation and forest degradation as well as alternative forest management. However, when forest cover, structure and composition change, shifts in biophysical processes (the water and energy balances) may enhance or diminish the climate effects of carbon released from forest aboveground biomass. The net climate impact of carbon effects and biophysical effects determines outcomes for forest and agricultural species as well as the humans who depend on them. Evaluating the net impact is complicated by the disparate spatio-temporal scales at which they operate. Here we review the biophysical mechanisms by which forests influence climate and synthesize recent work on the biophysical climate forcing of forests across latitudes. We then combine published data on the biophysical effects of deforestation on climate by latitude with a new analysis of the climate impact of the CO2 in forest aboveground biomass by latitude to quantitatively assess how these processes combine to shape local and global climate. We find that tropical deforestation leads to strong net global warming as a result of both CO2 and biophysical effects. From the tropics to a point between 30°N and 40°N, biophysical cooling by standing forests is both local and global, adding to the global cooling effect of CO2 sequestered by forests. In the mid-latitudes up to 50°N, deforestation leads to modest net global warming as warming from released forest carbon outweighs a small opposing biophysical cooling. Beyond 50°N large scale deforestation leads to a net global cooling due to the dominance of biophysical processes (particularly increased albedo) over warming from CO2 released. Locally at all latitudes, forest biophysical impacts far outweigh CO2 effects, promoting local climate stability by reducing extreme temperatures in all seasons and times of day. The importance of forests for both global climate change mitigation and local adaptation by human and non-human species is not adequately captured by current carbon-centric metrics, particularly in the context of future climate warming.
Introduction
Failure to stabilize climate is in itself a large threat to biodiversity already at risk from deforestation. Protection, expansion, and improved management of the world’s forests represent some of the most promising natural solutions to the problem of keeping global warming below 1.5–2 degrees (Griscom et al., 2017; Roe et al., 2019). Forests sequester large quantities of carbon; of the 450–650 Pg of carbon stored in vegetation (IPCC, 2013), over 360 Pg is in forest vegetation (Pan et al., 2013). Adding the carbon in soils, forests contain over 800 PgC, almost as much as is currently stored in the atmosphere (Pan et al., 2013). In addition, forests are responsible for much of the carbon removal by terrestrial ecosystems which together remove 29% of annual CO2 emissions (∼11.5 PgC; Friedlingstein et al., 2019). Globally, forest loss not only releases a large amount of carbon to the atmosphere, but it also significantly diminishes a major pathway for carbon removal long into the future (Houghton and Nassikas, 2018). Tropical forests, which hold the greatest amount of aboveground biomass and have one of the fastest carbon sequestration rates per unit land area (Harris et al., 2021), face the greatest deforestation pressure (FAO, 2020). Given the long half-life and homogenous nature of atmospheric CO2, current forest management decisions will have an enduring impact on global climate through effects on CO2 alone. However, forests also impact climate directly through controls on three main biophysical mechanisms: albedo, evapotranspiration (ET) and canopy roughness.
The direct biophysical effects of forests moderate local climate conditions. As a result of relatively low albedo, forests absorb a larger fraction of incoming sunlight than brighter surfaces such as bare soil, agricultural fields, or snow. Changes in albedo can impact the radiation balance at the top of the atmosphere and thus global temperature. The local climate, however, is not only impacted by albedo changes but also by how forests partition incoming solar radiation between latent and sensible heat. Deep roots and high leaf area make forests very efficient at moving water from the land surface to the atmosphere via ET, producing latent heat. Thus, beneath the forest canopy, the sensible heat flux and associated surface temperature are relatively low, especially during the growing season when ET is high (Davin and de Noblet-Ducoudré, 2010; Mildrexler et al., 2011; Alkama and Cescatti, 2016). This cooling is enhanced by the relatively high roughness of the canopy, which strengthens vertical mixing and draws heat and water vapor away from the surface. Higher in the atmosphere, as water vapor condenses, the latent heat is converted to sensible heat. As a result, warming that began with sunlight striking the canopy is felt higher in the atmosphere rather than in the air near the land surface. These non-radiative processes stabilize local climate by reducing both the diurnal temperature range and seasonal temperature extremes (Lee et al., 2011; Zhang et al., 2014; Alkama and Cescatti, 2016; Findell et al., 2017; Forzieri et al., 2017; Hirsch et al., 2018; Lejeune et al., 2018). Their impact on global climate, however, is less clear.
Despite high spatial variability, forest biophysical impacts do follow predictable latitudinal patterns. In the tropics, higher incoming solar radiation and moisture availability provide more energy to drive ET and convection, which in combination with roughness overcome the warming effect of low albedo, and result in year round cooling by forests. At higher latitudes, where incoming solar radiation is highly seasonal, the impacts of ET and surface roughness are diminished (Anderson et al., 2011; Li et al., 2015) and albedo is the dominant biophysical determinant of the climate response. In boreal forests, relatively low albedo and low ET cause strong winter and spring warming. In the summer, higher incoming radiation and somewhat higher ET result in mild cooling by boreal forests (Alkama and Cescatti, 2016). In the mid-latitudes, forest cover results in mild biophysical evaporative cooling in the summer months and mild albedo warming in the winter months (Davin and de Noblet-Ducoudré, 2010; Li et al., 2015; Schultz et al., 2017). The latitude of zero net biophysical effect, the point at which the annual effect of the forest shifts from local cooling to local warming, ranges from 30 to 56°N in the literature (Figure 1). These generalized latitudinal trends can be modified by aridity, elevation, species composition, and other characteristics, which vary across a range of spatial scales (Anderson-Teixeira et al., 2012; Williams et al., 2021).
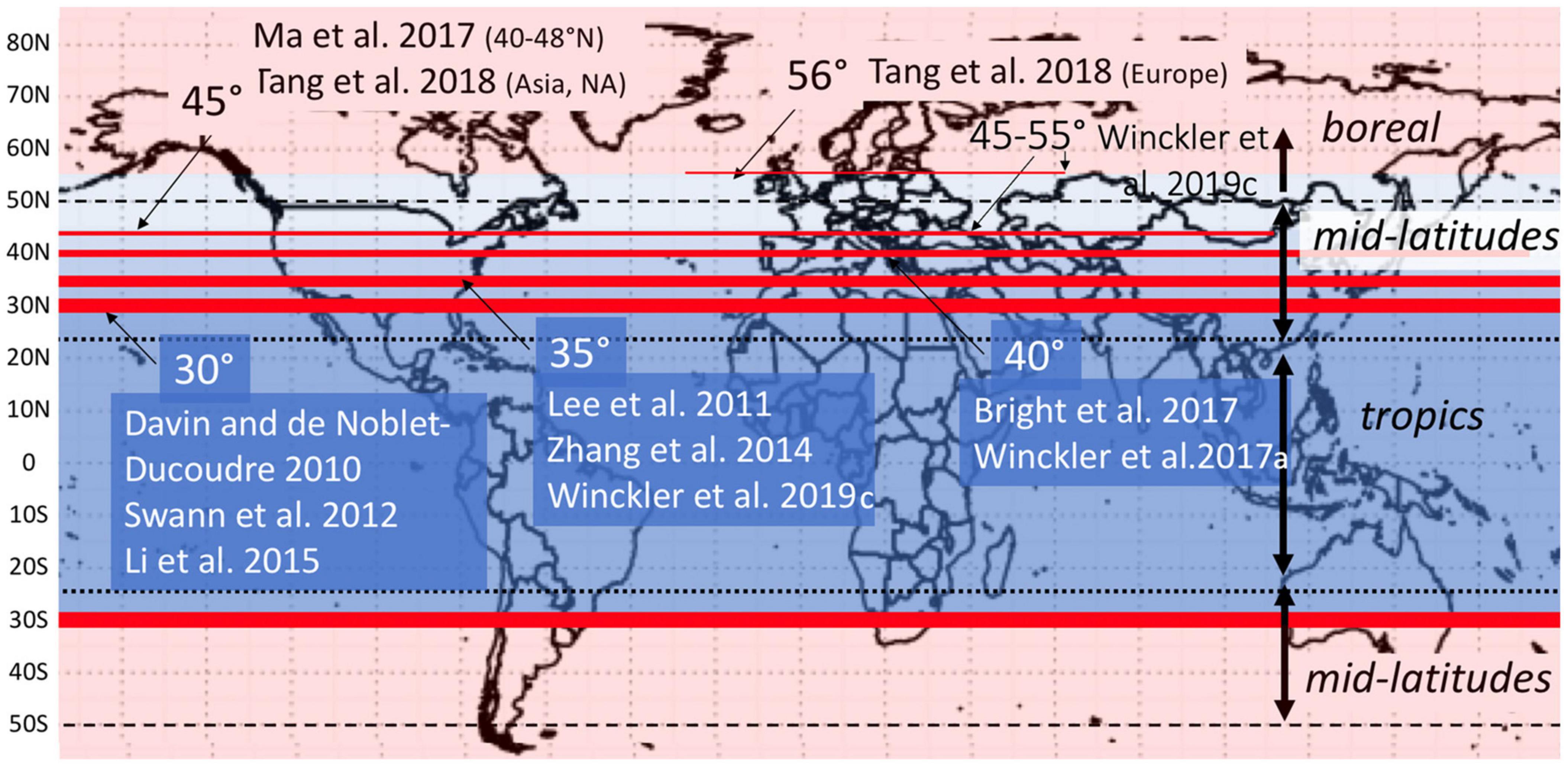
Figure 1. Latitude of net zero biophysical effect of forests on local temperature varies from 30 to 56°N. Above the line, forest cover causes local warming; below the line, forest cover causes local cooling. The thickness of the line indicates the number of studies that show forest cooling up to that threshold. Data sources as indicated.
Various mechanisms can amplify or dampen a forest’s direct effects on the energy and water balance, with climate impacts in the immediate vicinity, in remote locations, or both (Bonan, 2008). Indirect biophysical effects are particularly important in the boreal region where snow-forest albedo interactions are prevalent. Low albedo forests typically mask high albedo snow, resulting in local radiative warming (Jiao et al., 2017). At the larger scale this forest-induced warming is transferred to the oceans and further amplified by interactions with sea ice (Brovkin et al., 2004; Bala et al., 2007; Davin and de Noblet-Ducoudré, 2010; Laguë and Swann, 2016). In fact, indirect biophysical feedbacks appear to dominate the global temperature response to deforestation in the boreal region (Devaraju et al., 2018). Future climate warming may alter the strength of such feedbacks, depending on the rate at which forests expand northward and the extent and persistence of spring snow cover in a warmer world.
In the tropics, where ET and roughness are the dominant biophysical drivers, forests cool the lower atmosphere, but also provide the water vapor to support cloud formation (Teuling et al., 2017). Clouds whiten the atmosphere over forests and thus increase albedo, at least partially offsetting the inherently low albedo of the forest below (Heald and Spracklen, 2015; Fisher et al., 2017). However, the water vapor in clouds also absorbs and re-radiates heat, counteracting some of the cloud albedo-induced cooling (Swann et al., 2012). In the Amazon basin, evidence suggests that deep clouds may occur more frequently over forested areas as a result of greater humidity and consequently greater convective available potential energy (Wang et al., 2009). The impact of tropical deforestation on cloud formation is modified by biomass burning aerosols (Liu et al., 2020) and the net impact on global climate is unclear. Quantifying these indirect biophysical feedback effects is an ongoing challenge for the modeling community particularly in the context of constraining future climate scenarios.
Forest production of biogenic volatile organic compounds (BVOC), which affect both biogeochemical and biophysical processes, further complicate quantification of the net climate impact of forests. BVOC and their oxidation products regulate secondary organic aerosols (SOA), which are highly reflective and result in biophysical cooling. SOA also act as cloud condensation nuclei, enhancing droplet concentrations and thereby increasing cloud albedo, which leads to additional biophysical cooling (Topping et al., 2013). On the other hand, SOA can also cause latent heat release in deep convective cloud systems resulting in strong radiative warming of the atmosphere (Fan et al., 2012, 2013). Furthermore, through impacts on the oxidative capacity of the atmosphere, BVOC increase the lifetime of methane and lead to the formation of tropospheric ozone in the presence of nitrogen oxides (Arneth et al., 2011; McFiggans et al., 2019). The persistence of ozone and methane (both greenhouse gases) results in a biogeochemical warming effect. The net effect of forest BVOC at both local and global scales remains uncertain. Current evidence, from modeling forest loss since 1850, suggests that BVOC result in a small net cooling, if indirect cloud effects are included (Scott et al., 2018). The strongest effect is in the tropics, where BVOC production is highest (Messina et al., 2016).
An improved understanding of the combined effects of forest carbon and biophysical controls on both local and global climate is necessary to guide policy decisions that support global climate mitigation, local adaptation and biodiversity conservation. The relative importance of forest carbon storage and biophysical effects on climate depend in large part on the spatial and temporal scale of interest. Local surface or air temperature may not be sensitive to the incremental impact of atmospheric CO2 removed by forests growing in a particular landscape or watershed. In contrast, local temperature is sensitive to biophysical changes in albedo, ET and roughness. At regional and global scales, where the cumulative effects of forests on atmospheric CO2 become apparent in the temperature response, we can usefully compare these impacts. Estimates of the relative impact of biophysical and biogeochemical (e.g., carbon cycle) processes on global or zonal climate have been provided primarily by model simulations of large-scale deforestation or afforestation (Table 1). These studies generally show that CO2 effects on global temperature are many times greater than the biophysical effects of forest cover or forest loss. In models depicting global or zonal deforestation outside the tropics, however, global warming from CO2 release offsets only 10–90% of the global biophysical cooling. The global CO2 effects of total deforestation in the tropics greatly outweigh the global biophysical effects (Table 1). With the exception of Davin and de Noblet-Ducoudré (2010), these studies have estimated the net contribution of biophysical processes, without isolating the individual biophysical components. Here, we provide a new analysis of CO2-induced warming from deforestation by 10° latitudinal increments (Supplementary Information 1). We then compare the CO2 effect with the only published determination of biophysical effects by latitude (Davin and de Noblet-Ducoudré, 2010) to clarify the potential net impact of forest loss in a particular region on local and global climate.
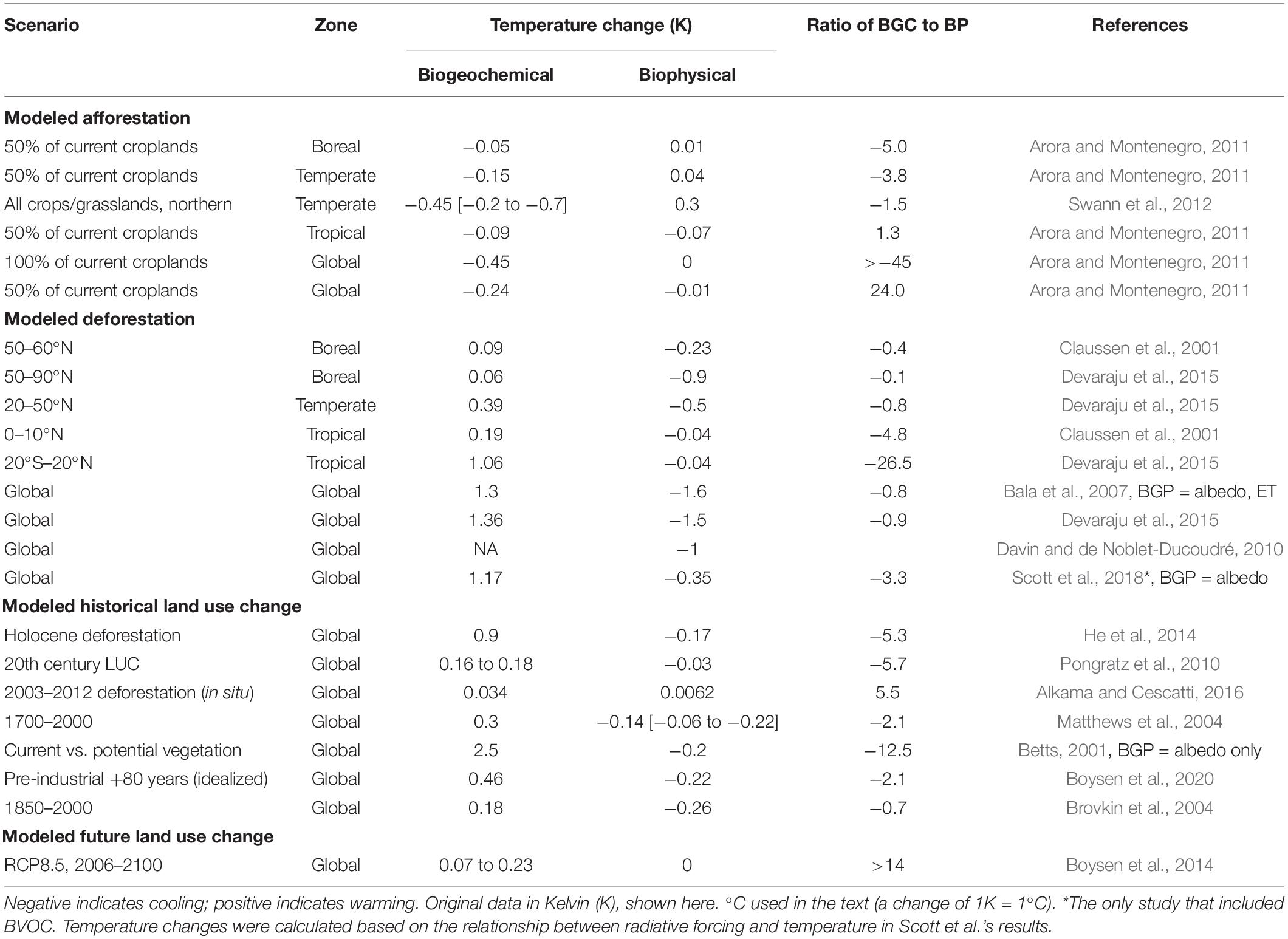
Table 1. Forest effects on global temperature in modeling experiments from biogeochemical (CO2) versus biophysical impacts (albedo, evapotranspiration and roughness as well as changes in atmospheric and ocean circulation, snow and ice, and clouds).
Materials and Methods
In the scientific literature, biophysical impacts have been quantified using a number of different methods. In situ observational data, including weather station and eddy flux measurements, have shaped our understanding of the direct biophysical impacts of forests on the surface energy balance. With the advantage of high temporal resolution, they allow for process level investigation of forest biophysical impacts and attribution of temperature changes to particular biophysical forcings, both radiative (albedo) and non-radiative (ET and roughness) (Lee et al., 2011; Luyssaert et al., 2014; Vanden Broucke et al., 2015; Bright et al., 2017; Liao et al., 2018). Remote sensing techniques have recently been used to extrapolate to larger scales, providing a global map of forest cover effects on local climate (Li et al., 2015; Alkama and Cescatti, 2016; Bright et al., 2017; Duveiller et al., 2018; Prevedello et al., 2019). However, in contrast to in situ approaches which generally measure near surface air temperature (generally but not always at 2 m), remote sensing studies have investigated the response of land surface temperature (i.e., skin temperature) which is 0.5–3 times more sensitive to forest cover change (Alkama and Cescatti, 2016; Novick and Katul, 2020).
Generally, both in situ and remote sensing analyses have adopted a space-for-time approach where differences in surface climate of neighboring forest and non-forest sites are used as proxies for the climate signal from deforestation/afforestation over time. This approach assumes that neighboring sites share a common background climate and that any temperature differences between them can be attributed solely to differences in forest cover. Consequently, large-scale biophysical feedback effects are ignored. New observation-based methodologies have been devised to investigate impacts from ongoing land use change rather than estimating climate sensitivities to idealized forest change (Alkama and Cescatti, 2016; Bright et al., 2017; Prevedello et al., 2019), however, they too measure only local biophysical impacts.
Numerical modeling of paired climate simulations with contrasting forest cover is necessary to investigate the net climate response to forest cover change, including both local and non-local impacts. Model simulations have focused on idealized scenarios of large-scale deforestation/afforestation which are more likely to trigger large-scale climate feedbacks than more realistic incremental forest cover change. Discrepancies between observed and modeled results may be due in part to the influence of indirect climate feedbacks that are not captured by observations (Winckler et al., 2017a,2019a; Chen and Dirmeyer, 2020). Unfortunately, model resolution is currently too coarse to guide local policy decisions. Modeling results are also plagued by a number of uncertainties associated with the partitioning of energy between latent or sensible heat (de Noblet-Ducoudré et al., 2012). The predicted impacts of similar land cover changes are model specific and can vary in sign, magnitude, and geographical distribution (Devaraju et al., 2015; Lawrence and Vandecar, 2015; Garcia et al., 2016; Laguë and Swann, 2016; Stark et al., 2016; Quesada et al., 2017; Boysen et al., 2020) and therefore must be viewed with caution. In this paper, we synthesize all types of observational data from the literature to illustrate the biophysical impacts of forests on local climate. However, given that local impacts have been extensively explored and summarized in the past (Anderson et al., 2011; Perugini et al., 2017), and because we wish to include indirect effects and feedbacks, we rely predominantly on modeling studies and our own calculations to elucidate the role of forests at different latitudes in shaping climate.
Effects on Global Temperature From Deforestation by 10° Latitude Band
We combined published data on biophysical effects of deforestation by latitude with our own analysis of CO2 effects from deforestation by latitude to compare the relative strength of biophysical factors and CO2 (the dominant biogeochemical factor) affecting global climate. Most modeling experiments available in the literature involve total deforestation at all latitudes, and the ocean feedbacks prove very strong (Davin and de Noblet-Ducoudré, 2010). Here, we consider land-only effects within a given 10° latitudinal band as this scale of impact is more indicative of the effects of regional or more incremental change on global temperature than the combined land/ocean effects. Finer scale, more realistic forest loss scenarios would not trigger massive cooling through albedo effects on the oceans. Area-scaled, land-only biophysical effects from deforestation provide the most realistic comparison with the effects of carbon stored by forests, and released through deforestation, at a given latitude. The biophysical response was derived from the results of Davin and de Noblet-Ducoudré (2010) who simulated total deforestation and decomposed the temperature response, by 10° latitude bands, into the fraction due to albedo, evapotranspiration, roughness and a non-linear response (see Supplementary Table 1).
The biogeochemical response was estimated by accounting for the CO2 effect of deforestation, using existing biomass data and known equilibrium temperature sensitivity to doubled CO2. The principal input to our analysis is a 2016 global extension of the 500-m resolution aboveground carbon density (ACD) change (2003–2016) product applied by Walker et al. (2020) to the Amazon basin. It is based on an approach to pantropical ACD change estimation developed by Baccini et al. (2017). The pantropical product combined field measurements with colocated NASA ICESat GLAS spaceborne light detection and ranging (LiDAR) data to calibrate a machine-learning algorithm that produced estimates of ACD using MODIS satellite imagery. This approach was modified for application to the extratropics, principally the temperate and boreal zones but also extratropical South America, Africa and Australia, using 47 allometric equations compiled from 27 unique literature sources for relating field-based measurements of aboveground biomass to airborne LiDAR metrics (Chapman et al., 2020). These equations were used to predict ACD within the footprints of GLAS LiDAR acquisitions in each region with the result being a pseudo-inventory of LiDAR-based estimates of ACD spanning the extratropics. This dataset was then combined with the pantropical dataset first generated by Baccini et al. (2012) to produce a global database of millions of spatially explicit ACD predictions. This database was used to calibrate six ecoregional MODIS-based models for the purposes of generating a global 500-m resolution map of ACD for the year 2016. Additional details on these methods can be found in Chapman et al. (2020).
The total aboveground carbon (GtC) was summed for each 10° latitude band and converted to CO2 (GtC*44/12 = GtCO2, Supplementary Information 1). The mass of CO2 was converted to ppm CO2 in the atmosphere (2.12 Gt/ppm). The derived CO2 concentration was reduced by 23% to account for ocean uptake (Global Carbon Project, 2019). We assumed that no uptake occurred on land, as the carbon stock in vegetation was completely removed in our experiment to match what occurred in Davin and de Noblet-Ducoudré (2010). Next, we calculated the global temperature response to the increase in atmospheric CO2 due to the CO2 released by completely deforesting each 10° latitudinal band using the equilibrium temperature sensitivity derived from general circulation models. Given the accepted value of 3°C (±1.5°C) for a doubling of atmospheric CO2 (an increase of 280 ppm) (IPCC, 2013), we determined that temperature sensitivity is equivalent to 0.107°C (±0.054°C) for every 10 ppm increase in atmospheric CO2 content.
To determine the global temperature response to deforestation of a given band, we calculated the area-weighted values for each biophysical response within each latitude band. The area encompassed by 10° of latitude increases toward the equator. Thus, to determine the contribution of a given band to a global temperature response, scaling by the surface area within the band was essential. We used average temperature responses over the land only to avoid the strong bias associated with ocean feedbacks from global scale implementation of deforestation.
For the global analysis, we also determined the contribution of BVOC to global temperature change for deforestation of each 10° of latitude. Scott et al. (2018) described the warming from deforestation due to BVOC in relation to the amount of cooling due to changes in albedo. For the tropics, the BVOC effect on global temperature was 17% of the albedo effect. For the temperate zone, it was 18% and for the boreal, it was 2% of the albedo effect. We applied these scalars (with an opposite sign) to the albedo figures for each 10° latitude band.
Effects on Regional (Local) Temperature From Deforestation by 10° Latitude Band
To analyze the effect of deforesting 10° of latitude on the temperature within that latitude zone (‘local’ effect), we did not scale by area within the band. Rather we assessed the average temperature change across the band, locally felt, as reported in the original study. The CO2 effect was calculated as above and then scaled to reflect the sensitivity of a given latitudinal band to a global forcing. Only the CO2 emitted by the latitudinal band itself was considered when determining the locally felt effects of CO2 in a given band. Our experimental design involved global deforestation and all emitted CO2 would have had an effect in a given band, but the point of the analysis was to isolate the temperature change caused by forests in a given latitude. We determined the latitudinal sensitivity to warming in response to added CO2 from a re-analysis of global 2 m temperature data (CERA-20C) obtained from the European Centre for Medium-Range Weather https://www.ecmwf.int/en/forecasts/datasets/reanalysis-datasets/cera-20c. We compared average temperatures from 1901 to 1910 and 2001–2010, by latitude on land only (inadequate land only data for 50–60S and 80–90N; for those, we do not report a locally felt CO2 effect). Then we divided the temperature change for each latitude band by the change in global temperature over the same period. We scaled the effect of CO2 emitted by a given 10° latitude band by this sensitivity to represent the influence of non-linear responses such as polar amplification (see Supplementary Information 1 and Supplementary Table 2).
Results
Biophysical Effects of Deforestation on Local Climate: A Broader Context
Our analysis is the first to compare regional scale biophysical and CO2 impacts from regional scale deforestation but the literature is replete with data on local biophysical impacts. The results for local biophysical effects (100s of m to 100s of km) agree with our results at the regional scale (below). Figures 2, 3 synthesize local biophysically-driven temperature responses to deforestation, as indicated by forest/no-forest comparisons or forest change over time, from the scientific literature. Satellite and flux tower data indicate that surface temperatures in tropical forests are significantly lower than in cleared areas nearby. On an annual basis, local surface cooling of 0.2–2.4°C has been observed (mean 0.96°C, Figure 2 and Supplementary Information 2). In the temperate zone, satellite studies of land surface temperature (which is more sensitive than the temperature of the air at 2 m) have shown biophysical cooling from forest cover, or biophysical warming from deforestation (0.02–1.0°C, mean of 0.4°C; see Figure 2 and Supplementary Information 2). Both in situ and satellite data generally indicate an average annual cooling of under 1°C from boreal deforestation (Figure 2). Across latitudinal zones, warming from deforestation is generally greater during the day, and during the dry (hot) season (Figure 3).
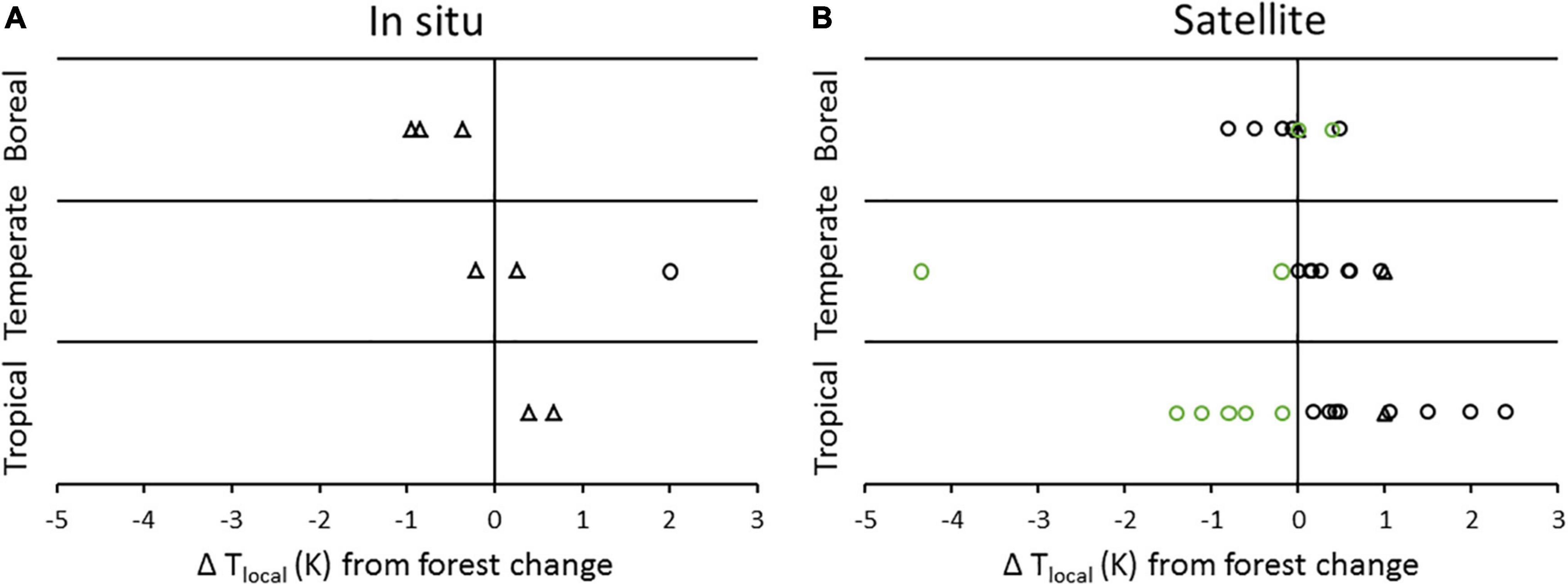
Figure 2. Local average annual temperature change in response to deforestation (black symbols) or afforestation (green symbols) as determined by comparing neighboring forested and open land (space for time approach) or measuring forest change over time in the tropics, temperate and boreal zones, by (A) in situ or (B) satellite based land surface temperature measurements (0 m, triangles) or air temperature measurements (2 m, circles). See Supplementary Information 2 for data sources.
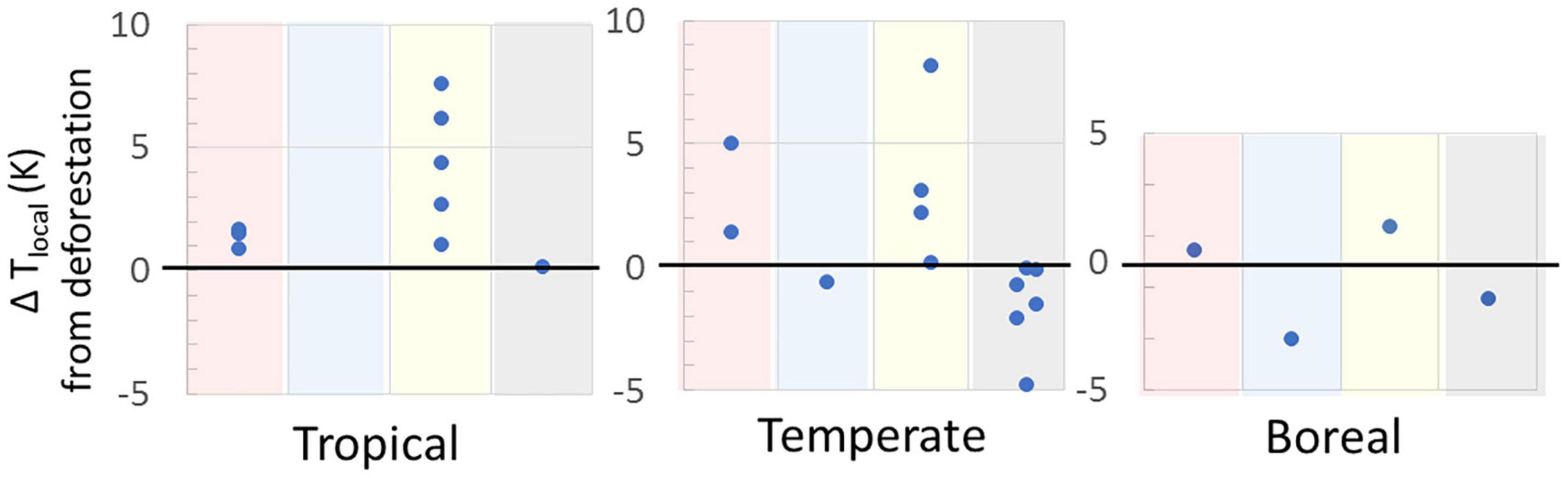
Figure 3. Local temperature change in response to deforestation by season and time of day in the various climate zones as determined by comparing neighboring forested and open land (space for time approach) or measuring forest change over time. Warm/dry season response, averaged over the entire diurnal cycle, in red shading and cold/wet season response in blue shading. Daytime response, averaged over the entire annual cycle, in yellow shading and nighttime response in gray shading. See Supplementary Information 3 for data sources.
CO2-Induced Warming Versus Biophysical Effects on Regional (Local) Temperature From Deforestation by 10° Latitude Band
As expected, the regionally felt effect of regionally (10° band) produced CO2 is very small compared to any individual biophysical effect or the sum of all non-CO2 effects (Figure 4). These results indicate that the net impact of all non-CO2 effects is negligible between 20 and 30N. Beyond 30N the local biophysical response to deforestation is cooling. In the broader literature, this latitude of net zero biophysical effect on local temperature is generally between 30 and 40N (Figure 1).
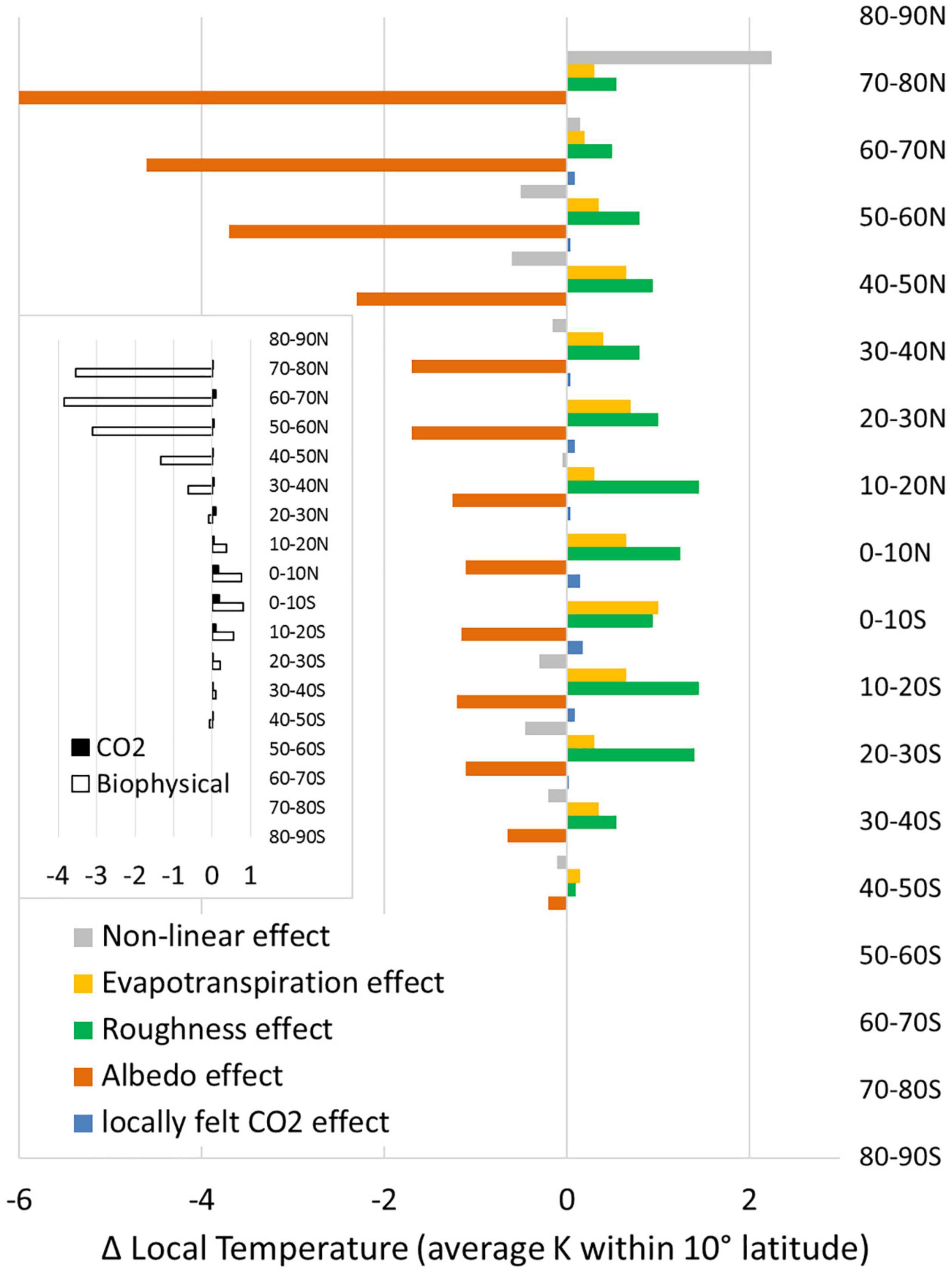
Figure 4. Effect of complete deforestation on local annual temperature by climate factor, averaged across the land surface within a 10° latitudinal band. Complete deforestation was implemented globally and analyzed by 10° latitudinal bands (Davin and de Noblet-Ducoudré, 2010). The CO2 effect was determined from total aboveground biomass in each 10° band after Walker et al. (2020) and scaled by CERA-derived sensitivity by latitude. Inset distinguishes the sum of all local biophysical effects from local CO2 effects.
Biophysical Effects on Global Temperature From Deforestation by 10° Latitude Band
For most latitudinal bands, the strongest biophysical effect of deforestation is cooling from albedo changes. In the tropics, however, the warming effect of lost roughness is comparable to or greater than the albedo effect (Figure 5A). Adding the warming from lost evapotranspiration, the net biophysical effect from tropical deforestation is global warming, as much as 0.1°C contributed each by latitudes 0°–10°S and 0°–10°N. The net biophysical effect of intact tropical forest, therefore, is global cooling; slightly more cooling if BVOCs are also considered (see Figure 5B). Roughness effects are generally greater than evapotranspiration effects across latitudes providing a strong counterbalance to albedo effects (Davin and de Noblet-Ducoudré, 2010; Burakowski et al., 2018; Winckler et al., 2019b; Figure 5A). Albedo almost balances the combined effect of roughness, evapotranspiration, BVOC and non-linear effects between 20 and 30°N resulting in close to zero net biophysical effect on global temperature (Figure 5B). From 30–40°N and northward, albedo dominates, and the net biophysical effect of deforestation is cooling.
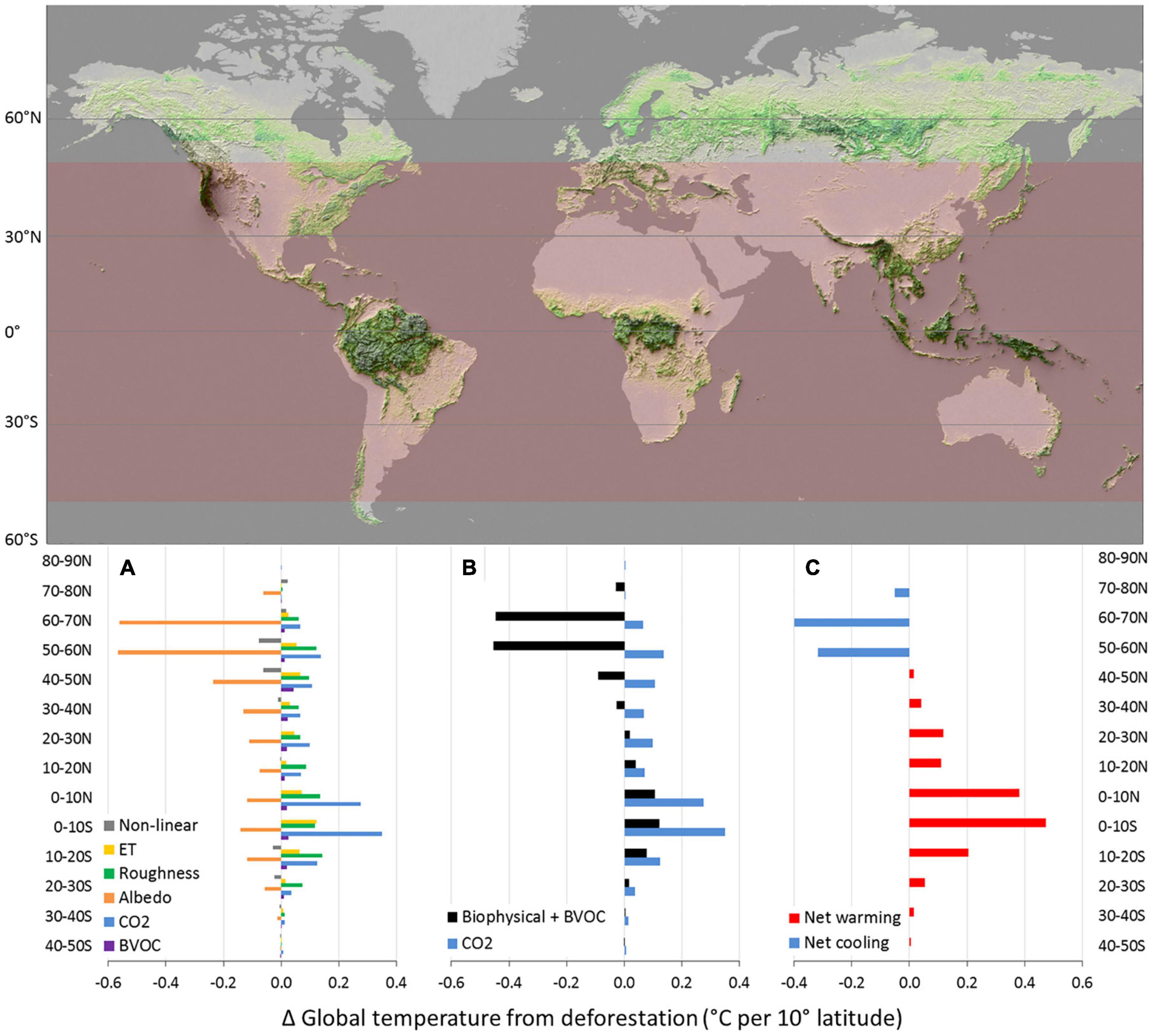
Figure 5. Effect of complete deforestation on global temperature by 10° band of latitude. (A) Contribution to global temperature change by climate forcing factor. Biophysical factors are from Davin and de Noblet-Ducoudré, 2010, area-weighted. BVOC effects are estimated relative to albedo effects based on Scott et al., 2018. CO2 effect is based on aboveground live biomass for each 10° latitudinal band following Baccini et al., 2017 and Walker et al., 2020. (B) Net biophysical and BVOC effect versus CO2 effect. (C) Cooling or warming effects of deforestation by 10° latitudinal band (BVOC included). “Forests as mountains” map of aboveground biomass carbon in woody vegetation ca. 2016 courtesy of Woodwell Climate Research Center and shaded to indicate where deforestation results in net global warming. See Supplementary Information 1 for details.
CO2-Induced Warming Versus Biophysical Effects on Global Temperature From Deforestation by 10° Latitude Band
From 30°S to 30°N, the biophysical effect of deforesting a given 10° latitudinal band is about half as great and in the same direction as the CO2 effect: global warming. Biophysical warming is around 60% as great as warming from released CO2 in the outer tropics (20°S–10°S and 10°N–20°N) and about 35% as great in the heart of the tropics (10°S–10°N). Biophysical cooling due to deforestation from 30°N to 40°N offsets about 40% of the warming associated with carbon loss from deforestation; from 40°N to 50°N biophysical effects offset 85% of CO2 effects (Figure 5B). Above 50°N, biophysical global cooling is 3–6 times as great as CO2 induced global warming. The net impact of deforestation (effects of CO2, biophysical processes and BVOC combined) is warming at all latitudes up to 50N (Figure 5C). Thus, from 50S to 50N, an area that encompasses approximately 65% of global forests (FAO, 2020), deforestation results in global warming (Figure 5C).
Discussion
All Forests Provide Local Climate Benefits Through Biophysical Effects
Ignoring biophysical effects on local climate means casting aside a powerful inducement to promote global climate goals and advance forest conservation: local self-interest. The biogeochemical effect of forests tends to dominate the biophysical effect at the global scale because physical effects in one region can cancel out effects in another. Nevertheless, biophysical effects are very important, and can be very large, at the local scale (e.g., Anderson-Teixeira et al., 2012; Bright et al., 2015; Jiao et al., 2017; Figures 2–4). The role of forests in maintaining critical habitat for biodiversity is well known, but new research on extinction confirms the role of forests in maintaining critical climates to support biodiversity. Changes in maximum temperature are driving extinction, not changes in average temperature (Román-Palacios and Wiens, 2020). Deforestation is associated with an increase in the maximum daily temperature throughout the year in the tropics and during the summer in higher latitudes (Lee et al., 2011; Zhang et al., 2014). Of course deforestation also increases average daytime temperatures in boreal, mid-latitude and tropical forests (Figure 3). The biophysical effects of forests also moderate local and regional temperature extremes such that extremely hot days are significantly more common following deforestation even in the mid- and high latitudes (Vogel et al., 2017; Stoy, 2018). Historical deforestation explains ∼1/3 of the present day increase in the intensity of the hottest days of the year at a given location (Lejeune et al., 2018). It has also increased the frequency and intensity of hot dry summers two to four fold (Findell et al., 2017). Local increases in extreme temperatures due to forest loss are of comparable magnitude to changes caused by 0.5°C of global warming (Seneviratne et al., 2018). Forests provide local cooling during the hottest times of the year anywhere on the planet, improving the resilience of cities, agriculture, and conservation areas. Forests are critical for adapting to a warmer world.
Forests also minimize risks due to drought associated with heat extremes. Deep roots, high water use efficiency, and high surface roughness allow trees to continue transpiring during drought conditions and thus to dissipate heat and convey moisture to the atmosphere. In addition to this direct cooling, forest ET can influence cloud formation (Stoy, 2018), enhancing albedo and potentially promoting rainfall. The production of BVOCs and organic aerosols by forests accelerates with increasing temperatures, enhancing direct or indirect (cloud formation) albedo effects. This negative feedback on temperature has been observed to counter anomalous heat events in the mid-latitudes (Paasonen et al., 2013).
Some Forests Provide Global Climate Benefits Through Biophysical Effects
Disregarding the biophysical effects of specific forests on global climate means under-selling some forest actions and over-selling others. The response to local forest change is not equivalent for similar sized areas in different latitudes. According to Arora and Montenegro (2011) warming reductions per unit reforested area are three times greater in the tropics than in the boreal and northern temperate zone due to a faster carbon sequestration rate magnified by year-round biophysical cooling. Thus, considering biophysical effects significantly enhances both the local and global climate benefits of land-based mitigation projects in the tropics (see Figures 4, 5).
Constraints on Forest Climate Benefits in the Future
Climate change is likely to alter the biophysical effect of forests in a variety of ways. Deforestation in a future (warmer) climate could warm the tropical surface 25% more than deforestation in a present-day climate due to stronger decreases in turbulent heat fluxes (Winckler et al., 2017b). In a warmer climate, reduced snow cover in the temperate and boreal regions will lead to a smaller albedo effect and thus less biophysical cooling with high latitude deforestation. In addition to snow cover change, future rainfall regimes will affect the response of climate to changes in forest cover (Pitman et al., 2011) as rainfall limits the supply of moisture available for evaporative cooling. Increases in water use efficiency due to increasing atmospheric CO2 may reduce evapotranspiration (Keenan et al., 2013), potentially reducing the local cooling effect of forests and altering atmospheric moisture content and dynamics at local to global scales. Future BVOC production may increase due to warming and simultaneously decline due to CO2 suppression (Lathière et al., 2010; Unger, 2014; Hantson et al., 2017). The physiological and ecological responses of forests to warming, rising atmospheric CO2 and changing precipitation contribute to uncertainty in the biophysical effect of future forests on climate.
Forest persistence is essential for maintaining the global benefits of carbon removals from the atmosphere and the local and global benefits of the physical processes described above. Changing disturbance regimes may limit forest growth and regrowth in many parts of the world. Dynamic global vegetation models currently show an increasing terrestrial carbon sink in the future. This sink is thought to be due to the effects of fertilization by rising atmospheric CO2 and N deposition on plant growth as well as the effects of climate change lengthening the growing season in northern temperate and boreal areas (Le Quéré et al., 2018). Free-air carbon dioxide enrichment (FACE) experiments often show increases in biomass accumulation under high CO2 but results are highly variable due to nutrient limitations and climatic factors (Feng et al., 2015; Paschalis et al., 2017; Terrer et al., 2018). Climate change effects on the frequency and intensity of pest outbreaks are poorly studied, but are likely to be significant, particularly at the margins of host ranges. Warmer springs and winters are already increasing insect-related tree mortality in boreal forests through increased stress on the tree hosts and direct effects on insect populations (Volney and Fleming, 2000; Price et al., 2013).
Climate also affects fire regimes. In the tropics, fire regimes often follow El Niño cycles (van der Werf et al., 2017). As temperatures increase, however, fire and rainfall are decoupled as the flammability of forests increases even in normal rainfall years (Fernandes et al., 2017; Brando et al., 2019). Fire frequency is also increasing in some temperate and boreal forests, with a discernable climate change signal (Abatzoglou and Williams, 2016). Modeling exercises indicate that this trend is expected to continue with increasing damage to forests as temperatures rise and fire intensity increases (De Groot et al., 2013).
In addition to changes induced by warming, continued deforestation could severely stress remaining forests by warming and drying local and regional climates (Lawrence and Vandecar, 2015; Costa et al., 2019; Gatti et al., 2021). In the tropics, a tipping point may occur, potentially resulting in a shift to shorter, more savannah-like vegetation and altering the impact of vast, previously forested areas on global climate (Nobre et al., 2016; Brando et al., 2019). Some of these processes are included in climate models and some are not. The gaps leave considerable uncertainty. Nevertheless, a combination of observations, models, and theory gives us a solid understanding of the biophysical effects of forests on climate at local, regional and global scales. We can use that knowledge to plan forest-based climate mitigation and adaptation.
Mitigation Potential of Forests: Byond the Carbon/Biophysical Divide
If instead of focusing on the contrast between biophysical and biochemical impacts of forests and forest loss, we focus on the potential of forests to cool the planet through both pathways, another picture emerges. By our conservative estimate, through the combined effects on CO2, BVOC, roughness and evapotranspiration, forests up to 50°N provide a net global cooling that is enough to offset warming associated with their low albedo. Given the most realistic pathways of forest change in the future (not complete deforestation of a 10° latitudinal band, or an entire biome), global climate stabilization benefits likely extend beyond 50°N. For the 29% of the global land surface that lies beyond 50°N, forests may warm the planet, but only as inferred from assessing the effects of complete zonal deforestation with all the associated, and powerful, land-ocean feedbacks spawned by largescale forest change in the boreal zone. Forests above 50°N, like forests everywhere, provide essential local climate stabilization benefits by reducing surface temperatures during the warm season as well as periods of extreme heat or drought. Indeed, they also reduce extreme cold.
Creating a fair and effective global arena for market-based solutions to climate change requires attention to all the ways that forests affect climate, including the biophysical effects. Future metrics of forest climate impacts should consider the effects of deforestation beyond CO2. Only recently have modelers begun to include BVOC. Doing so means that the albedo of intact forests (or the atmosphere above them) is higher due to the creation of SOA and subsequent cloud formation. Modeled deforestation thus results in less of a change in albedo, reducing the biophysical cooling effect. Similarly, accounting for the ozone and methane effects of BVOC reduces the biogeochemical warming from deforestation (Scott et al., 2018). In addition, especially in the tropics, deforestation reduces the strength of the soil CH4 sink (Dutaur and Verchot, 2007). While a small change relative to the atmospheric pool of CH4 (the second most important greenhouse gas), the loss of this sink is equivalent to approximately 13% of the current rate of increase in atmospheric CH4 (Saunois et al., 2016). We already have the data (Figure 5) to begin conceptualizing measures to coarsely scale CO2 impacts of forest change by latitude. Finer resolution of latitude, background climate (current and future) and forest type would improve any such new, qualifying metric for the climate mitigation value of forests.
The role of forests in addressing climate change extends beyond the traditional concept of CO2 mitigation which neglects the local climate regulation services they provide. The biophysical effects of forest cover can contribute significantly to solving local adaptation challenges, such as extreme heat and flooding, at any latitude. The carbon benefits of forests at any latitude contribute meaningfully to global climate mitigation. In the tropics, however, where forest carbon stocks and sequestration rates are highest, the biophysical effects of forests amplify the carbon benefits, thus underscoring the critical importance of protecting, expanding, and improving the management of tropical forests. Perhaps it is time to think more broadly about what constitutes global climate mitigation. If climate mitigation means limiting global warming, then clearly the biophysical effects of deforestation must be considered in addition to its effects on atmospheric CO2. We may further consider whether mitigation is too narrow a scope for considering the climate benefits provided by forests. Climate policy often separates mitigation from adaptation, but the benefits of forests clearly extend into both realms.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author Contributions
DL conceived the presented idea. All authors helped perform the computations, discussed the results, and contributed to the final manuscript.
Funding
Financial support from the University of Virginia and the Climate and Land Use Alliance grant #G-1810-55876.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
Thanks to Frances Seymour, Michael Wolosin, Billie L. Turner, Ruth DeFries, and the reviewers for feedback on this manuscript and to the University of Virginia and the Climate and Land Use Alliance grant #G-1810-55876 for financial support.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2022.756115/full#supplementary-material
References
Abatzoglou, J. T., and Williams, A. P. (2016). Impact of anthropogenic climate change on wildfire across western US forests. Proc. Natl. Acad. Sci. U.S.A. 113, 11770–11775. doi: 10.1073/pnas.1607171113
Alkama, R., and Cescatti, A. (2016). Biophysical climate impacts of recent changes in global forest cover. Science 351, 600–604. doi: 10.1126/science.aac8083
Anderson, R. G., Canadell, J. G., Randerson, J. T., Jackson, R. B., Hungate, B. A., Baldocchi, D. D., et al. (2011). Biophysical considerations in forestry for climate protection. Front. Ecol. Environ. 9, 174–182. doi: 10.1890/090179
Anderson-Teixeira, K. J., Snyder, P. K., Twine, T. E., Cuadra, S. V., Costa, M. H., and DeLucia, E. H. (2012). Climate-regulation services of natural and agricultural ecoregions of the Americas. Nat. Clim. Change 2:177. doi: 10.1038/nclimate1346
Arneth, A., Schurgers, G., Lathiere, J., Duhl, T., Beerling, D. J., Hewitt, C. N., et al. (2011). Global terrestrial isoprene emission models: sensitivity to variability in climate and vegetation. Atmos. Chem. Phys. 11, 8037–8052. doi: 10.1002/2013JD021238
Arora, V. K., and Montenegro, A. (2011). Small temperature benefits provided by realistic afforestation efforts. Nat. Geosci. 4, 514–518. doi: 10.1038/ngeo1182
Baccini, A. G. S. J., Goetz, S. J., Walker, W. S., Laporte, N. T., Sun, M., Sulla-Menashe, D., et al. (2012). Estimated carbon dioxide emissions from tropical deforestation improved by carbon-density maps. Nat. Clim. Change 2, 182–185. doi: 10.1038/nclimate1354
Baccini, A., Walker, W., Carvalho, L., Farina, M., Sulla-Menashe, D., and Houghton, R. A. (2017). Tropical forests are a net carbon source based on aboveground measurements of gain and loss. Science 358, 230–234. doi: 10.1126/science.aam5962
Bala, G., Caldeira, K., Wickett, M., Phillips, T. J., Lobell, D. B., Delire, C., et al. (2007). Combined climate and carbon-cycle effects of large-scale deforestation. Proc. Natl. Acad. Sci. U.S.A. 104, 6550–6555. doi: 10.1073/pnas.0608998104
Betts, R. A. (2001). Biogeophysical impacts of land use on present-day climate: near-surface temperature change and radiative forcing. Atmos. Sci. Lett. 2, 39–51. doi: 10.1006/asle.2001.0023
Bonan, G. B. (2008). Forests and climate change: forcings, feedbacks, and the climate benefits of forests. Science 320, 1444–1449. doi: 10.1126/science.1155121
Boysen, L. R., Brovkin, V., Pongratz, J., Lawrence, D. M., Lawrence, P., Vuichard, N., et al. (2020). Global climate response to idealized deforestation in CMIP6 models. Biogeosciences 17, 5615–5638. doi: 10.5194/bg-17-5615-2020
Boysen, L., Brovkin, V., Arora, V. K., Cadule, P., de Noblet-Ducoudré, N., Kato, E., et al. (2014). Global and regional effects of land-use change on climate in 21st century simulations with interactive carbon cycle. Earth Syst. Dyn. 5, 309–319. doi: 10.5194/esd-5-309-2014
Brando, P. M., Paolucci, L., Ummenhofer, C. C., Ordway, E. M., Hartmann, H., Cattau, M. E., et al. (2019). Droughts, wildfires, and forest carbon cycling: a pantropical synthesis. Annu. Rev. Earth Planet. Sci. 47, 555–581. doi: 10.1146/annurev-earth-082517-010235
Bright, R. M., Davin, E., O’Halloran, T., Pongratz, J., Zhao, K., and Cescatti, A. (2017). Local temperature response to land cover and management change driven by non-radiative processes. Nat. Clim. Change 7:296. doi: 10.1038/nclimate3250
Bright, R. M., Zhao, K., Jackson, R. B., and Cherubini, F. (2015). Quantifying surface albedo and other direct biogeophysical climate forcings of forestry activities. Glob. Change Biol. 21, 3246–3266. doi: 10.1111/gcb.12951
Brovkin, V., Sitch, S., Von Bloh, W., Claussen, M., Bauer, E., and Cramer, W. (2004). Role of land cover changes for atmospheric CO2 increase and climate change during the last 150 years. Glob. Change Biol. 10, 1253–1266. doi: 10.1111/j.1365-2486.2004.00812.x
Burakowski, E., Tawfik, A., Ouimette, A., Lepine, L., Novick, K., Ollinger, S., et al. (2018). The role of surface roughness, albedo, and Bowen ratio on ecosystem energy balance in the Eastern United States. Agric. For. Meteorol. 249, 367–376. doi: 10.1016/j.agrformet.2017.11.030
Chapman, M., Walker, W. S., Cook-Patton, S. C., Ellis, P. W., Farina, M., Griscom, B. W., et al. (2020). Large climate mitigation potential from adding trees to agricultural lands. Glob. Change Biol. 26, 4357–4365. doi: 10.1111/gcb.15121
Chen, L., and Dirmeyer, P. A. (2020). Reconciling the disagreement between observed and simulated temperature responses to deforestation. Nat. Commun. 11:202. doi: 10.1038/s41467-019-14017-0
Claussen, M., Brovkin, V., and Ganopolski, A. (2001). Biogeophysical versus biogeochemical feedbacks of large-scale land cover change. Geophys. Res. Lett. 28, 1011–1014. doi: 10.1029/2000gl012471
Costa, M. H., Fleck, L. C., Cohn, A. S., Abrahão, G. M., Brando, P. M., Coe, M. T., et al. (2019). Climate risks to Amazon agriculture suggest a rationale to conserve local ecosystems. Front. Ecol. Environ. 17, 584–590. doi: 10.1002/fee.2124
Davin, E. L., and de Noblet-Ducoudré, N. (2010). Climatic impact of global-scale deforestation: radiative versus nonradiative processes. J. Clim. 23, 97–112. doi: 10.1175/2009jcli3102.1
De Groot, W. J., Flannigan, M. D., and Cantin, A. S. (2013). Climate change impacts on future boreal fire regimes. For. Ecol. Manage. 294, 35–44. doi: 10.1016/j.foreco.2012.09.027
de Noblet-Ducoudré, N., Boisier, J. P., Pitman, A., Bonan, G. B., Brovkin, V., Cruz, F., et al. (2012). Determining robust impacts of land-use-induced land cover changes on surface climate over North America and Eurasia: results from the first set of LUCID experiments. J. Clim. 25, 3261–3281. doi: 10.1175/jcli-d-11-00338.1
Devaraju, N., Bala, G., and Modak, A. (2015). Effects of large-scale deforestation on precipitation in the monsoon regions: remote versus local effects. Proc. Natl. Acad. Sci. U.S.A. 112, 3257–3262. doi: 10.1073/pnas.1423439112
Devaraju, N., de Noblet-Ducoudré, N., Quesada, B., and Bala, G. (2018). Quantifying the relative importance of direct and indirect biophysical effects of deforestation on surface temperature and teleconnections. J. Clim. 31, 3811–3829. doi: 10.1175/jcli-d-17-0563.1
Dutaur, L., and Verchot, L. V. (2007). A global inventory of the Soil CH4 Sink. Glob. Biogeochem. Cycles 21, GB4013.
Duveiller, G., Hooker, J., and Cescatti, A. (2018). The mark of vegetation change on Earth’s surface energy balance. Nat. Commun. 9:679. doi: 10.1038/s41467-017-02810-8
Fan, J., Leung, L. R., Rosenfeld, D., Chen, Q., Li, Z., Zhang, J., et al. (2013). Microphysical effects determine macrophysical response for aerosol impacts on deep convective clouds. Proc. Natl. Acad. Sci. U.S.A. 110, E4581–E4590. doi: 10.1073/pnas.1316830110
Fan, J., Rosenfeld, D., Ding, Y., Leung, L. R., and Li, Z. (2012). Potential aerosol indirect effects on atmospheric circulation and radiative forcing through deep convection. Geophys. Res. Lett. 39:L09806.
Feng, Z., Rütting, T., Pleijel, H., Wallin, G., Reich, P. B., Kammann, C. I., et al. (2015). Constraints to nitrogen acquisition of terrestrial plants under elevated CO 2. Glob. Change Biol. 21, 3152–3168. doi: 10.1111/gcb.12938
Fernandes, K., Verchot, L., Baethgen, W., Gutierrez-Velez, V., Pinedo-Vasquez, M., and Martius, C. (2017). Heightened fire probability in Indonesia in non-drought conditions: the effect of increasing temperatures. Environ. Res. Lett. 12:054002. doi: 10.1088/1748-9326/aa6884
Findell, K. L., Berg, A., Gentine, P., Krasting, J. P., Lintner, B. R., Malyshev, S., et al. (2017). The impact of anthropogenic land use and land cover change on regional climate extremes. Nat. Commun. 8:989. doi: 10.1038/s41467-017-01038-w
Fisher, J. B., Melton, F., Middleton, E., Hain, C., Anderson, M., Allen, R., et al. (2017). The future of evapotranspiration: global requirements for ecosystem functioning, carbon and climate feedbacks, agricultural management, and water resources. Water Resour. Res. 53, 2618–2626. doi: 10.1002/2016wr020175
Forzieri, G., Alkama, R., Miralles, D. G., and Cescatti, A. (2017). Satellites reveal contrasting responses of regional climate to the widespread greening of Earth. Science 356, 1180–1184. doi: 10.1126/science.aal1727
Friedlingstein, P., Jones, M., O’Sullivan, M., Andrew, R., Hauck, J., Peters, G., et al. (2019). Global carbon budget 2019. Earth Syst.Sci. Data 11, 1783–1838.
Garcia, E. S., Swann, A. L., Villegas, J. C., Breshears, D. D., Law, D. J., Saleska, S. R., et al. (2016). Synergistic ecoclimate teleconnections from forest loss in different regions structure global ecological responses. PLoS One 11:e0165042. doi: 10.1371/journal.pone.0165042
Gatti, L. V., Basso, L. S., Miller, J. B., Gloor, M., Gatti Domingues, L., Cassol, H. L., et al. (2021). Amazonia as a carbon source linked to deforestation and climate change. Nature 595, 388–393. doi: 10.1038/s41586-021-03629-6
Global Carbon Project (2019). Supplemental Data of Global Carbon Budget 2019 (Version 1.0). Global Carbon Project. doi: 10.18160/gcp-2019
Griscom, B. W., Adams, J., Ellis, P. W., Houghton, R. A., Lomax, G., Miteva, D. A., et al. (2017). Natural climate solutions. Proc. Natl. Acad. Sci. U.S.A. 114, 11645–11650.
Hantson, S., Knorr, W., Schurgers, G., Pugh, T. A. M., and Arneth, A. (2017). Global isoprene and monoterpene emissions under changing climate, vegetation, CO2 and land use. Atmos. Environ. 155, 35–45. doi: 10.1016/j.atmosenv.2017.02.010
Harris, N. L., Gibbs, D. A., Baccini, A., Birdsey, R. A., De Bruin, S., Farina, M., et al. (2021). Global maps of twenty-first century forest carbon fluxes. Nat. Clim. Change 11, 234–240. doi: 10.1038/s41558-020-00976-6
He, F., Vavrus, S. J., Kutzbach, J. E., Ruddiman, W. F., Kaplan, J. O., and Krumhardt, K. M. (2014). Simulating global and local surface temperature changes due to Holocene anthropogenic land cover change. Geophys. Res. Lett. 41, 623–631. doi: 10.1002/2013gl058085
Heald, C. L., and Spracklen, D. V. (2015). Land use change impacts on air quality and climate. Chem. Rev. 115, 4476–4496. doi: 10.1021/cr500446g
Hirsch, A. L., Guillod, B. P., Seneviratne, S. I., Beyerle, U., Boysen, L. R., Brovkin, V., et al. (2018). Biogeophysical impacts of land-use change on climate extremes in low-emission scenarios: results from HAPPI-Land. Earths Future 6, 396–409. doi: 10.1002/2017EF000744
Houghton, R. A., and Nassikas, A. A. (2018). Negative emissions from stopping deforestation and forest degradation, globally. Glob. Change Biol. 24, 350–359. doi: 10.1111/gcb.13876
IPCC (2013). Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, eds T. F. Stocker, D. Qin, G.-K. Plattner, M. Tignor, S. K. Allen, J. Boschung, et al. (Cambridge: Cambridge University Press), 1535.
Jiao, T., Williams, C. A., Ghimire, B., Masek, J., Gao, F., and Schaaf, C. (2017). Global climate forcing from albedo change caused by large-scale deforestation and reforestation: quantification and attribution of geographic variation. Clim. Change 142, 463–476. doi: 10.1007/s10584-017-1962-8
Keenan, T. F., Hollinger, D. Y., Bohrer, G., Dragoni, D., Munger, J. W., Schmid, H. P., et al. (2013). Increase in forest water-use efficiency as atmospheric carbon dioxide concentrations rise. Nature 499:324. doi: 10.1038/nature12291
Laguë, M. M., and Swann, A. L. (2016). Progressive midlatitude afforestation: impacts on clouds, global energy transport, and precipitation. J. Clim. 29, 5561–5573. doi: 10.1175/jcli-d-15-0748.1
Lathière, J., Hewitt, C. N., and Beerling, D. J. (2010). Sensitivity of isoprene emissions from the terrestrial biosphere to 20th century changes in atmospheric CO2 concentration, climate, and land use. Glob. Biogeochem. Cycles 24:GB1004.
Lawrence, D., and Vandecar, K. (2015). Effects of tropical deforestation on climate and agriculture. Nat. Clim. Change 5:27. doi: 10.1038/nclimate2430
Le Quéré, C., Andrew, R. M., Friedlingstein, P., Sitch, S., Hauck, J., Pongratz, J., et al. (2018). Global carbon budget 2018. Earth Syst. Sci. Data 10, 2141–2194.
Lee, X., Goulden, M. L., Hollinger, D. Y., Barr, A., Black, T. A., Bohrer, G., et al. (2011). Observed increase in local cooling effect of deforestation at higher latitudes. Nature 479:384. doi: 10.1038/nature10588
Lejeune, Q., Davin, E. L., Gudmundsson, L., Winckler, J., and Seneviratne, S. I. (2018). Historical deforestation locally increased the intensity of hot days in northern mid-latitudes. Nat. Clim. Change 8:386. doi: 10.1038/s41558-018-0131-z
Li, Y., Zhao, M., Motesharrei, S., Mu, Q., Kalnay, E., and Li, S. (2015). Local cooling and warming effects of forests based on satellite observations. Nat. Commun. 6:6603. doi: 10.1038/ncomms7603
Liao, W., Rigden, A. J., and Li, D. (2018). Attribution of local temperature response to deforestation. J. Geophys. Res. Biogeosci. 123, 1572–1587. doi: 10.1029/2018jg004401
Liu, L., Cheng, Y., Wang, S., Wei, C., Pohlker, M. L., Pohlker, C., et al. (2020). Impact of biomass burning aerosols on radiation, clouds, and precipitation over the Amazon during the dry season: relative importance of aerosol–cloud and aerosol–radiation interactions. Atmos. Chem. Phys. 20, 13283–13301. doi: 10.5194/acp-20-13283-2020
Luyssaert, S., Jammet, M., Stoy, P. C., Estel, S., Pongratz, J., Ceschia, E., et al. (2014). Land management and land-cover change have impacts of similar magnitude on surface temperature. Nat. Clim. Change 4, 389–393. doi: 10.1038/nclimate2196
Matthews, H. D., Weaver, A. J., Meissner, K. J., Gillett, N. P., and Eby, M. (2004). Natural and anthropogenic climate change: incorporating historical land cover change, vegetation dynamics and the global carbon cycle. Clim. Dyn. 22, 461–479. doi: 10.1007/s00382-004-0392-2
McFiggans, G., Mentel, T. F., Wildt, J., Pullinen, I., Kang, S., Kleist, E., et al. (2019). Secondary organic aerosol reduced by mixture of atmospheric vapours. Nature 565:587. doi: 10.1038/s41586-018-0871-y
Messina, P., Lathière, J., Sindelarova, K., Vuichard, N., Granier, C., Ghattas, J., et al. (2016). Global biogenic volatile organic compound emissions in the ORCHIDEE and MEGAN models and sensitivity to key parameters. Atmos. Chem. Phys. 15, 14169–14202. doi: 10.5194/acp-16-14169-2016
Mildrexler, D. J., Zhao, M., and Running, S. W. (2011). A global comparison between station air temperatures and MODIS land surface temperatures reveals the cooling role of forests. J. Geophys. Res. Biogeosci. 116:G03025.
Nobre, C. A., Sampaio, G., Borma, L. S., Castilla-Rubio, J. C., Silva, J. S., and Cardoso, M. (2016). Land-use and climate change risks in the Amazon and the need of a novel sustainable development paradigm. Proc. Natl. Acad. Sci. U.S.A. 113, 10759–10768. doi: 10.1073/pnas.1605516113
Novick, K. A., and Katul, G. G. (2020). The duality of reforestation impacts on surface and air temperature. J. Geophys. Res. Biogeosci. 125:e2019JG005543.
Paasonen, P., Asmi, A., Petäjä, T., Kajos, M. K., Äijälä, M., Junninen, H., et al. (2013). Warming-induced increase in aerosol number concentration likely to moderate climate change. Nat. Geosci. 6, 438–442. doi: 10.1038/ngeo1800
Pan, Y., Birdsey, R. A., Phillips, O. L., and Jackson, R. B. (2013). The structure, distribution, and biomass of the world’s forests. Annu. Rev. Ecol. Evol. Syst. 44, 593–622.
Paschalis, A., Katul, G. G., Fatichi, S., Palmroth, S., and Way, D. (2017). On the variability of the ecosystem response to elevated atmospheric CO2 across spatial and temporal scales at the Duke Forest FACE experiment. Agric. For. Meteorol. 232, 367–383. doi: 10.1016/j.agrformet.2016.09.003
Perugini, L., Caporaso, L., Marconi, S., Cescatti, A., Quesada, B., de Noblet-Ducoudre, N., et al. (2017). Biophysical effects on temperature and precipitation due to land cover change. Environ. Res. Lett. 12:053002. doi: 10.1088/1748-9326/aa6b3f
Pitman, A. J., Avila, F. B., Abramowitz, G., Wang, Y. P., Phipps, S. J., and de Noblet-Ducoudré, N. (2011). Importance of background climate in determining impact of land-cover change on regional climate. Nat. Clim. Change 1:472. doi: 10.1038/nclimate1294
Pongratz, J., Reick, C. H., Raddatz, T., and Claussen, M. (2010). Biogeophysical versus biogeochemical climate response to historical anthropogenic land cover change. Geophys. Res. Lett. 37:L08702.
Prevedello, J. A., Winck, G. R., Weber, M. M., Nichols, E., and Sinervo, B. (2019). Impacts of forestation and deforestation on local temperature across the globe. PLoS One 14:e0213368. doi: 10.1371/journal.pone.0213368
Price, D. T., Alfaro, R. I., Brown, K. J., Flannigan, M. D., Fleming, R. A., Hogg, E. H., et al. (2013). Anticipating the consequences of climate change for Canada’s boreal forest ecosystems. Environ. Rev. 21, 322–365. doi: 10.1139/er-2013-0042
Quesada, B., Arneth, A., and de Noblet-Ducoudré, N. (2017). Atmospheric, radiative, and hydrologic effects of future land use and land cover changes: a global and multimodel climate picture. J. Geophys. Res. Atmos. 122, 5113–5131. doi: 10.1002/2016jd025448
Roe, S., Streck, C., Obersteiner, M., Frank, S., Griscom, B., Drouet, L., et al. (2019). Contribution of the land sector to a 1.5 C world. Nat. Clim. Change 9, 817–828.
Román-Palacios, C., and Wiens, J. J. (2020). Recent responses to climate change reveal the drivers of species extinction and survival. Proc. Natl. Acad. Sci. U.S.A. 117, 4211–4217. doi: 10.1073/pnas.1913007117
Saunois, M., Bousquet, P., Poulter, B., Peregon, A., Ciais, P., Canadell, J. G., et al. (2016). The global methane budget 2000–2012. Earth Syst. Sci. Data 8, 697–751. doi: 10.1016/j.scitotenv.2019.04.263
Schultz, N. M., Lawrence, P. J., and Lee, X. (2017). Global satellite data highlights the diurnal asymmetry of the surface temperature response to deforestation. J. Geophys. Res. Biogeosci. 122, 903–917. doi: 10.1002/2016jg003653
Scott, C. E., Monks, S. A., Spracklen, D. V., Arnold, S. R., Forster, P. M., Rap, A., et al. (2018). Impact on short-lived climate forcers increases projected warming due to deforestation. Nat. Commun. 9:157. doi: 10.1038/s41467-017-02412-4
Seneviratne, S. I., Wartenburger, R., Guillod, B. P., Hirsch, A. L., Vogel, M. M., Brovkin, V., et al. (2018). Climate extremes, land–climate feedbacks and land-use forcing at 1.5 C. Philos. Trans. A Math. Phys. Eng. Sci. 376:20160450. doi: 10.1098/rsta.2016.0450
Stark, S. C., Breshears, D. D., Garcia, E. S., Law, D. J., Minor, D. M., Saleska, S. R., et al. (2016). Toward accounting for ecoclimate teleconnections: intra-and inter-continental consequences of altered energy balance after vegetation change. Landsc. Ecol. 31, 181–194. doi: 10.1007/s10980-015-0282-5
Stoy, P. C. (2018). Deforestation intensifies hot days. Nat. Clim. Change 8, 366–368. doi: 10.1111/gcb.15279
Swann, A. L., Fung, I. Y., and Chiang, J. C. (2012). Mid-latitude afforestation shifts general circulation and tropical precipitation. Proc. Natl. Acad. Sci. U.S.A. 109, 712–716. doi: 10.1073/pnas.1116706108
Terrer, C., Vicca, S., Stocker, B. D., Hungate, B. A., Phillips, R. P., Reich, P. B., et al. (2018). Ecosystem responses to elevated CO 2 governed by plant–soil interactions and the cost of nitrogen acquisition. New Phytol. 217, 507–522. doi: 10.1111/nph.14872
Teuling, A. J., Taylor, C. M., Meirink, J. F., Melsen, L. A., Miralles, D. G., Van Heerwaarden, C. C., et al. (2017). Observational evidence for cloud cover enhancement over western European forests. Nat. Commun. 8:14065. doi: 10.1038/ncomms14065
Topping, D., Connolly, P., and McFiggans, G. (2013). Cloud droplet number enhanced by co-condensation of organic vapours. Nat. Geosci. 6:443. doi: 10.1038/ngeo1809
Unger, N. (2014). Human land-use-driven reduction of forest volatiles cools global climate. Nat. Clim. Change 4:907. doi: 10.1038/nclimate2347
van der Werf, G. R., Randerson, J. T., Giglio, L., Van Leeuwen, T. T., Chen, Y., Rogers, B. M., et al. (2017). Global fire emissions estimates during 1997-2016. Earth Syst. Sci. 9, 697–720. doi: 10.5194/essd-9-697-2017
Vanden Broucke, S., Luyssaert, S., Davin, E. L., Janssens, I., and Van Lipzig, N. (2015). New insights in the capability of climate models to simulate the impact of LUC based on temperature decomposition of paired site observations. J. Geophys. Res. Atmos. 120, 5417–5436. doi: 10.1002/2015jd023095
Vogel, M. M., Orth, R., Cheruy, F., Hagemann, S., Lorenz, R., van den Hurk, B. J., et al. (2017). Regional amplification of projected changes in extreme temperatures strongly controlled by soil moisture-temperature feedbacks. Geophys. Res. Lett. 44, 1511–1519. doi: 10.1002/2016gl071235
Volney, W. J. A., and Fleming, R. A. (2000). Climate change and impacts of boreal forest insects. Agric. Ecosyst. Environ. 82, 283–294. doi: 10.1016/s0167-8809(00)00232-2
Walker, W. S., Gorelik, S. R., Baccini, A., Aragon-Osejo, J. L., Josse, C., Meyer, C., et al. (2020). The role of forest conversion, degradation, and disturbance in the carbon dynamics of Amazon indigenous territories and protected areas. Proc. Natl. Acad. Sci. U.S.A. 117, 3015–3025. doi: 10.1073/pnas.1913321117
Wang, J., Chagnon, F. J., Williams, E. R., Betts, A. K., Renno, N. O., Machado, L. A., et al. (2009). Impact of deforestation in the Amazon basin on cloud climatology. Proc. Natl. Acad. Sci. U.S.A. 106, 3670–3674. doi: 10.1073/pnas.0810156106
Williams, C. A., Gu, H., and Jiao, T. (2021). Climate impacts of US forest loss span net warming to net cooling. Sci. Adv. 7:eaax8859. doi: 10.1126/sciadv.aax8859
Winckler, J., Lejeune, Q., Reick, C. H., and Pongratz, J. (2019a). Nonlocal effects dominate the global mean surface temperature response to the biogeophysical effects of deforestation. Geophys. Res. Lett. 46, 745–755. doi: 10.1029/2018gl080211
Winckler, J., Reick, C. H., Bright, R. M., and Pongratz, J. (2019b). Importance of surface roughness for the local biogeophysical effects of deforestation. J. Geophys. Res. Atmos. 124, 8605–8618. doi: 10.1029/2018jd030127
Winckler, J., Reick, C. H., and Pongratz, J. (2017a). Robust identification of local biogeophysical effects of land-cover change in a global climate model. J. Clim. 30, 1159–1176. doi: 10.1175/jcli-d-16-0067.1
Winckler, J., Reick, C. H., and Pongratz, J. (2017b). Why does the locally induced temperature response to land cover change differ across scenarios? Geophys. Res. Lett. 44, 3833–3840. doi: 10.1002/2017gl072519
Keywords: forest, biophysical effects, temperature, climate policy, deforestation/afforestation
Citation: Lawrence D, Coe M, Walker W, Verchot L and Vandecar K (2022) The Unseen Effects of Deforestation: Biophysical Effects on Climate. Front. For. Glob. Change 5:756115. doi: 10.3389/ffgc.2022.756115
Received: 10 August 2021; Accepted: 02 March 2022;
Published: 24 March 2022.
Edited by:
Christos Mammides, Frederick University, CyprusReviewed by:
Alvaro Montenegro, The Ohio State University, United StatesRita Von Randow, São Paulo State Technological College, Brazil
Copyright © 2022 Lawrence, Coe, Walker, Verchot and Vandecar. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Deborah Lawrence, ZGwzY0B2aXJnaW5pYS5lZHU=