 Philippe Cadieux1
Philippe Cadieux1 Pierre Drapeau1*
Pierre Drapeau1* Ugo Ouellet-Lapointe1Alain Leduc1
Ugo Ouellet-Lapointe1Alain Leduc1 Louis Imbeau2Réjean Deschênes1Antoine Nappi3
Louis Imbeau2Réjean Deschênes1Antoine Nappi3- 1Centre for Forest Research, NSERC-UQAT-UQAM Industrial Chair in Sustainable Forest Management, Université du Québec à Montréal, Montréal, QC, Canada
- 2Centre for Forest Research, NSERC-UQAT-UQAM Industrial Chair in Sustainable Forest Management, Université du Québec en Abitibi-Témiscamingue, Rouyn-Noranda, QC, Canada
- 3Direction de l’expertise sur la Faune Terrestre, l’herpétofaune et l’avifaune, Ministère des Forêts, de la Faune et des Parcs, Québec, QC, Canada
Structural complexity generated by forest development processes and tree species compositional changes provide key habitat features for vertebrate communities that rely upon tree size and decay processes for foraging, denning or nesting. Complexity of forest structure in old stands could not only be key for harboring increased taxonomic species diversity but also greater functional diversity through more complexity in networks of tree cavity dependent species. Using a nest web approach that hierarchically links cavity-bearing trees with cavity formation agents (natural decay processes and avian excavators) and cavity users (non-excavator species), we compared network characteristics of nest webs along a time since fire gradient in a naturally disturbed boreal mixedwood forest landscape in eastern North America. Since 2003, twelve 24 to 40 ha plots ranging from 61 to more than 245 years after fire were surveyed at the Lake Duparquet Research and Teaching Forest in Abitibi, Quebec, Canada to detect active nesting, and denning cavities. We found that network complexity both in terms of number of vertebrate species and number of interactions among species, increased along the age gradient and was significantly higher in the older stands than predicted by chance. Whereas cavity-nesting communities in old forests used a higher diversity of tree species over a wide range of decay stages, trembling aspen remained a key cavity-bearing tree throughout the age gradient. Woodpeckers were the main cavity formation agents whereas less than 1% of cavities originated from natural decay. The structural development of older forests is thus a driver for functional diversity in cavity-using vertebrate communities through higher interaction richness in nest webs, among cavity-bearing trees, excavators and non-excavating users. The pivotal contribution of the entire gradient of old forest cover types to the overall complexity of nest webs in the boreal mixedwood zone is also a key for the resilience of the cavity-using vertebrate community to natural disturbances. We discuss how such resilience may be compromised by even-aged industrial timber harvesting with short rotations that shifts the age structure of boreal landscapes toward regenerating and young pole forests whereas old forest cover types become below their historical range of variability.
Introduction
In the last decades, increased knowledge in the understanding of compositional and structural development of forest ecosystems (Bergeron and Charron, 1994; Gauthier et al., 1996; Bergeron, 2000; Franklin et al., 2002; Kneeshaw and Gauthier, 2003) has emphasized the critical role of vertical and horizontal complexity of forest structure and composition on species taxonomic diversity when forest is aging and old large trees are decaying and dying (Harmon et al., 1986; Grove, 2002; Saint-Germain et al., 2007; Stokland et al., 2012). Aging of forest stands has indeed been linked to complexity of forests, notably through increasing diversity of tree decay stages (Franklin et al., 2002; Harper et al., 2003) that in turn harbor high taxonomic species diversity (Grove, 2002; Stokland et al., 2012) and are accompanied by the development of tree cavities and other dendromicrohabitats (Remm and Lõhmus, 2011; Larrieu et al., 2017). However, in depth knowledge of how interspecific interactions are mediated by these keystone structural features (sensu Tews et al., 2004) and how they drive functional diversity is less known in natural stand development conditions.
Such ecological knowledge can be pivotal in the context where organisms dependent on these structural features are likely to be affected in managed landscapes under industrial timber harvesting where their age structure has shifted toward a greater proportion of young forest stands at the expense of older forests (Spies et al., 1994; Esseen et al., 1997; Drapeau et al., 2000; Cyr et al., 2009), reducing the provision of large old, decaying and dead trees. Accordingly, ecologists have proposed forest managers silvicultural approaches that incorporate principles from natural stand development, notably the maintenance of biological legacies (more large old trees, standing dead and down wood) to enhance species diversity in managed forest landscapes (Bergeron et al., 1999; Franklin et al., 2000, 2002; Lindenmayer et al., 2000; Harvey et al., 2002; Gauthier et al., 2009).
Studying species interactions with biological legacies in forest ecosystems can lead to the identification of suites of species with functional redundancies (same roles shared by multiple species) or species that have disproportionate roles (keystone species). Such knowledge allows in turn to make predictions about the stability and resilience of these interaction networks (McCann, 2000; Messier and Puettmann, 2011) and perhaps provide better monitoring tools for assessing how managed forests are sustainable (Drapeau et al., 2009a,2016; Simard et al., 2013) from an functional ecology viewpoint.
For vertebrates that use tree cavities for denning or nesting, the quality and quantity of cavity-bearing trees can vary considerably between forest cover types as forest ages, thus changing cavity availability through tree species dynamics. This, in turn, can limit population density of cavity users, particularly for species that cannot excavate holes [Newton, 1994; Holt and Martin, 1997; but see Wesolowski and Martin (2018)]. In addition, changes in the availability of cavity-bearing trees play a critical role in the interspecific network of interactions that hierarchically link trees, cavity formation agents (natural decay and avian excavators) and non-excavating users into what Martin and Eadie (1999) have described as nest webs.
In most North American forest ecosystems where nest web studies were conducted, it was noted that avian excavators, mainly woodpeckers are the main cavity formation agents in these networks (Martin et al., 2004; Aitken and Martin, 2007; Blanc and Walters, 2008; Cockle et al., 2011; Cooke and Hannon, 2011). Accordingly, the excavation process in the structuring of nest webs is considered a keystone process (Bednarz et al., 2004) whereas some excavators can have a disproportionate role (keystone species) by providing a rare resource to numerous cavity users that cannot excavate a cavity (Martin et al., 2004; Edworthy and Martin, 2014).
Assessing changes in the nest web structure with forest structural development along succession requires prior knowledge of natural disturbances and the resulting age structure and composition of forest landscapes under investigation. Studies of natural disturbance history reconstruction of forest landscapes with dendrochronological or paleological approaches (Bergeron et al., 2004; Cyr et al., 2009) may provide such information but they are, however, rarely available for areas close to one another where forest successional pathways can be identified. The southern fringe of the boreal mixedwood forest in northwestern Québec offers such unique conditions, where detailed fire reconstruction studies (Bergeron, 1991; Dansereau and Bergeron, 1993) and natural forest succession pathways along this time since fire gradient have been documented (Bergeron and Dubuc, 1989; Bergeron, 1991, 2000; Bergeron and Charron, 1994; Bergeron and Harvey, 1997) in an area lightly affected by timber harvesting.
In this paper, we analyze how changes in forest composition and structure along this time since fire gradient influence the availability of cavity-bearing trees and how this has cascading effects on interspecific interactions within the cavity-using community in a naturally disturbed boreal mixedwood forest landscape in eastern North America. We hypothesize that nest web complexity of tree-cavity vertebrate communities is driven by structural and compositional development as the forest is aging. We predict that increases in tree species richness and structural diversity (stem degradation and diameter) with time since fire will increase the functional diversity of nest webs through increases in the number of interactions between species at all levels of their hierarchical structure from cavity-bearing trees to cavity formation agents to non-excavator users.
Materials and methods
Study design
Our study was conducted in the boreal mixedwood forest of northern Quebec in the balsam fir-white birch bioclimatic domain, located in the northern Clay Belt of Quebec and Ontario. This region is characterized by clay soils and low rocky hills (Robitaille and Saucier, 1998). The average annual temperature is 0.6oC and the average total annual precipitation is 822.7 mm (Environment Canada, 2022). The study site is located within the conservation area of the Lake Duparquet Research and Teaching Forest (hereafter LDRTF) at the southern fringe of the boreal forest in Québec (48°30’ N, 79°22’ W). The area spans ∼2,000 ha and contains a complex natural forest mosaic formed by different forest cover types originating from a variety of natural disturbances (Bergeron, 1991; Harvey, 1999). Over the last century, the balsam fir-dominated old-growth stands were affected by three spruce budworm outbreaks (1919–1929, 1930–1950, 1970–1987), (Morin et al., 1993). These outbreaks reduced the abundance of large balsam firs, killing as much as 75% of firs of more than 15 cm in diameter at breast height (DBH) in the study area in the most recent outbreak (Bergeron et al., 1995). Morever, Bergeron (2000) found that the cyclic spruce budworm outbreaks of the 20th century are important drivers in old forest stand dynamics that drive the high mortality and recruitment of balsam fir, which in turn allows trembling aspen individual or small patches to occur in conifer-dominated old forests.
Fire history reconstruction at the LDRTF is well-documented (Bergeron, 1991; Dansereau and Bergeron, 1993) and offered a unique opportunity to investigate within a relatively limited study area (35 km2) how structural complexity related to forest aging and succession affects the relationships of cavity-using communities. It includes eight major fires (1717, 1760, 1797, 1816, 1823, 1870, 1923, 1944) that occurred over the last three centuries, which within our study area comprises the chronosequence of stands covering the entire forest succession of the eastern boreal mixedwood forest. Tree species compositional changes in the study area have been well-studied (Bergeron and Charron, 1994; Bergeron and Harvey, 1997; Harvey et al., 2002). They are characterized by the transition from young and mature forests dominated by early successional tree species, such as trembling aspen (Populus tremuloides) and paper birch (Betula papyrifera) whereas older forests are characterized by mixed stands where balsam fir (Abies balsamea), black spruce (Picea mariana), and white spruce (Picea glauca) share the forest canopy with the deciduous species. Old-growth stands are dominated by coniferous species and are characterized by a higher abundance of white cedar (Thuja occidentalis) (Bergeron and Charron, 1994; Bergeron, 2000).
In 2003, we established 12 study plots (24–40 ha) distributed in four age classes; 61–89, 90–149, 149–244, and over 245 years after fire, with year 2005 as a reference point (Figure 1). These age classes were chosen [see Nappi et al. (2015) for more details] to cover the gradient of structural and compositional changes that occur in the boreal mixedwood forest from early to late successional stages following wildfire disturbances that historically occurred in the study area (Bergeron, 1991, 2000; Dansereau and Bergeron, 1993; Bergeron and Charron, 1994).
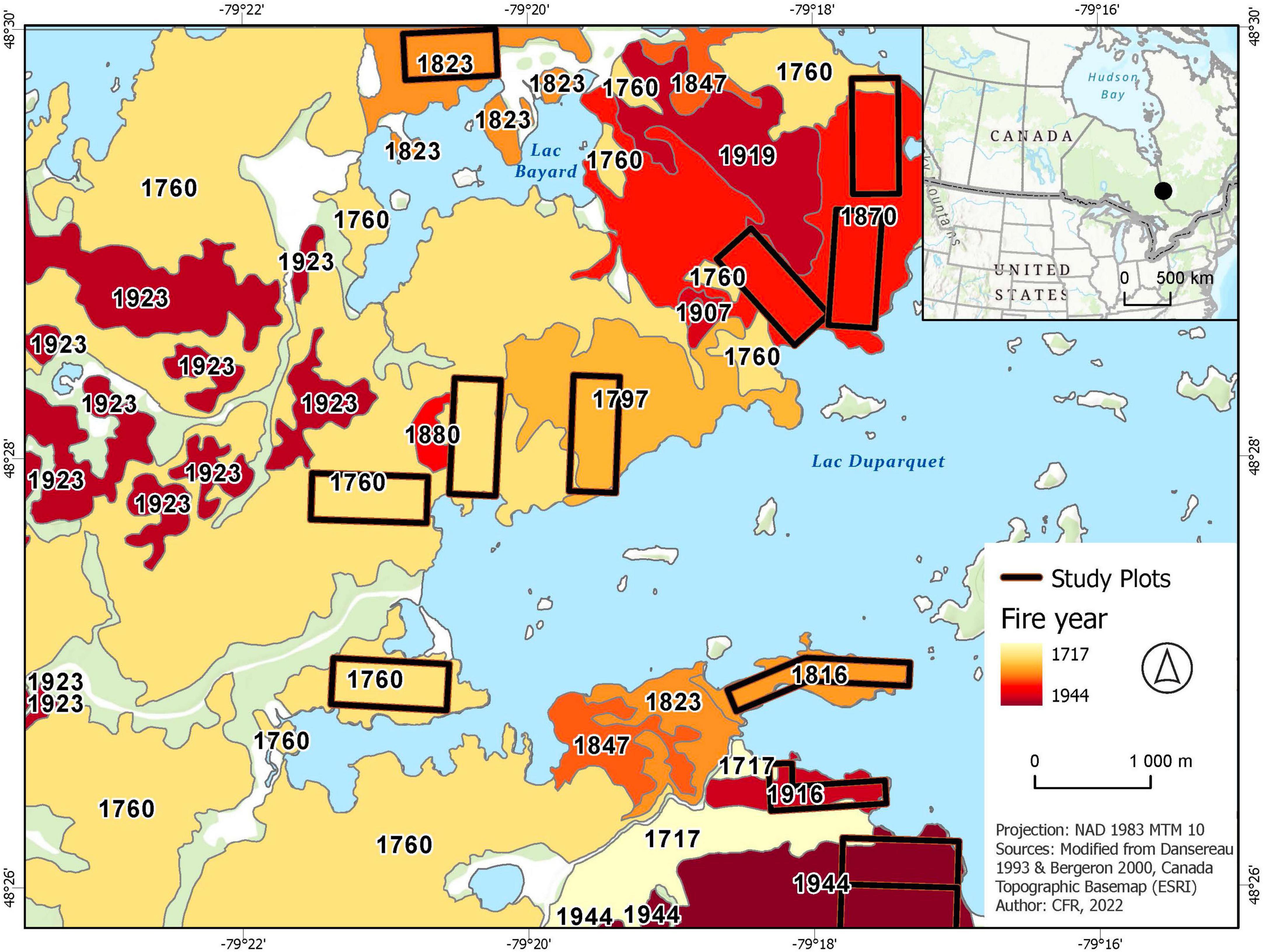
Figure 1. Localization of 12 forest plots where nest searches and cavity monitoring were conducted, from 2003 to 2012. Plots were distributed in four stand age categories (61–89, 90–149, 150–244, and over 245 years after fire) in the conservation area of the Lake Duparquet Research and Teaching Forest. Time since the last fire was calculated using 2005 as year of reference (based on detailed studies of fire history reconstruction: Bergeron, 1991; Dansereau and Bergeron, 1993, and Bergeron, 2000).
Vegetation sampling
Changes in forest composition and structure with time since disturbance of the forest cover are likely to influence the availability of adequate nesting trees for cavity users, which may in turn, change the structure and complexity of nest webs. Hence, vegetation sampling was conducted in the study area to compare changes in stand characteristics along the age gradient. In each study plot, nine to ten rectangular shaped vegetation-sampling stations measuring 600 m2 (10 m × 60 m) were distributed 200 m apart in each plot for a total 112 vegetation stations. Three variables were used to characterize the available trees for cavity-using communities: tree species, decay stage and DBH. Vegetation was sampled in 2005 and all trees having a DBH greater than 10 cm were characterized. Decay of standing trees was classified based on visual appearance in 5 categories (Nappi, 2009; Nappi et al., 2015): Decay 1 (alive, > 20% foliage); Decay 2 (declining, < 20% foliage); Decay 3 (recently dead, hard wood, firm bark cover); Decay 4 (moderate degradation, soft wood, no dead foliage present, no small twigs, some branches remaining, usually intact top); Decay 5 (high degradation, soft decomposed wood, very few branches, often broken top).
An additional vegetation survey was completed in 2013 to further characterize aspen decay along the age gradient. This survey consisted of 180 vegetation-sampling stations measuring 400 m2 (15 sampling stations per plot). As trembling aspens are often agregated in boreal mixedwood forest landscapes, the sampling stations were randomly distributed in aspen patches previously identified using areal photos of our study area (Ministère des Ressources naturelles et de la Faune du Québec, 2006). Trembling aspen stems > 20 cm in DBH were characterized by measuring the DBH, the decay class and the presence or absence of wood-decaying heart rot conks (Phellinus tremulae).
Nest search and cavity monitoring
This study is based on data compiled between 2003 and 2012. Data was collected using nest search surveys and long-term cavity monitoring. Nest search surveys were carried out almost every year between 2003 and 2012. However, they were conducted in all age classes in years 2003, 2004, 2005, and 2009. From 2003 to 2004, data on cavity nesters were collected as part of a study on the foraging ecology of woodpeckers (Nappi, 2009), which consisted of surveying 2 transect lines per plot with 100 m buffers on each side covering a total of 104–120 ha in each age category (1 km long and 400 m wide, ∼40 ha plots). Unlike later years, nest search effort was mainly directed toward finding woodpeckers (excavators) nests. In these 2 years, 51 occupied cavities were found. In 2008, systematic cavity density surveys were done on areas of 6.4 ha in each sampling plot [see Ouellet-Lapointe et al. (2012)] and nest searches were done in 8 of the 12 plots (2 plots in each age category; 80 ha in each age category). This search effort allowed the detection of 41 occupied cavities. In 2009, all 12 plots were sampled and the size of the sampling plots was reduced to 30 ha, covering 90 ha in each age class. Observers followed 8 line transects per plots having a buffer zone of 50 m on each side to increase the detection rate of active nests. This search effort allowed the detection of 132 occupied cavities. Occupied cavities were found during nest search surveys for other years (2008; n = 56 occupied cavities, 2010; n = 74, 2011; n = 42, 2012: n = 32), but given the unequal nest search efforts that were made between age classes, these cavities were not used in network comparisons. No nest search surveys were conducted in 2006–2007.
Nest searches were realized between the beginning of May and the middle of July. Observers walked along transect lines and used visual and auditory cues that would indicate the presence of an occupied cavity: breeding-bird behavior, begging nestlings or wood chips on the ground. When possible, the excavator of the cavity was visually identified. If no excavator was present at the site, we identified the excavator with the size and characteristics of the cavity. If there was a doubt, the excavator remained unidentified. All trees with occupied cavities were marked with an aluminum tag and geo-referenced. These cavities were then inspected with a camera mounted on a telescopic pole [TreeTop Peeper(TM), Sandpiper Technologies, Manteca, CA, USA] to determine the state of the cavity: non-completed excavation, suitable cavity or occupied cavity. A cavity was considered “occupied” if it contained at least one egg or nestling (Martin et al., 2004), or if a mammal was present inside the cavity. To collect additional information on the use of cavity by small mammals, particularly the Sciuridae family, we recorded the presence or absence of nesting material in each cavity (branches, straw, leaves, or bark). For nests that were not accessible with the telescopic pole and camera (higher than 15.24 m) and for nests that were in cavities with an opening too small for the camera [ex: some of the Red-breasted Nuthatch (Sitta canadensis) nests], we used nesting clues (behavior of adult cavity nesters) to confirm cavity activity. Nests found opportunistically in a 50 m radius around the plots were also georeferenced and characterized.
To assess the interactions of cavity producing agents with non-excavating users, which can be more elusive (Ouellet-Lapointe et al., 2012), we inspected existing cavities once a year from 2008 to 2012. Since it would have been logistically impossible to inspect every cavity found, we selected a subset of cavities to be monitored in order to get reasonable sample sizes on various cavity types (excavated or non-excavated, excavator species) in the stand ages in which they were located. Accordingly, the last 2 years of cavity monitoring (2011 and 2012) were more focused on increasing the sample size for rarer cavity types in our study area such as large cavities excavated by the Pileated Woodpecker (Dryocopus pileatus) and Northern Flicker (Colaptes auratus) and small cavities excavated by the Red-breasted Nuthatch.
Statistical analyses
Forest structural development along the time since fire gradient
We used linear and polynomial mixed effects models to compare stand characteristics of our response variables: tree species composition, tree degradation (the five categories of tree decay), tree size (DBH), and tree structural diversity (calculated with a Shannon-Weaver diversity index that combined degradation and DBH classes) along the time since fire gradient (explanatory variable) of our study sites. We used generalized mixed models with a binomial distribution to compare the probability of trembling aspen having a conk of Phellinus tremulae along the time since fire gradient. Plot location was treated as a random variable in all models to account for the hierarchical structure of the sampling design (Bates et al., 2012). We used a bootstrap procedure with a 1,000 permutations to estimate parameters and 95% confidence intervals (Bates et al., 2012). Statistical analyses were conducted using the R version 3.2.3 (R Core Team, 2015).
Nest web production
The complete boreal mixedwood nest web was created as a reference for illustrating the complexity and diversity of the cavity-nesting community at the landscape scale across the time since fire gradient of forest cover types. It is composed of all the nest data that were collected in the study plots and their surroundings from the entire 2003 to 2012 period for the whole study area (35 km2) in the continuous boreal mixedwood forest of Lake Duparquet. To assess changes in the structure and functional diversity of nest webs along the time since fire gradient, we constructed nest webs within the four age classes of our 12 study plots (61–89, 90–149, 150–244, and > 245 years after fire). Each age class had three study plots and only nest surveys conducted during 2003, 2004, 2005, and 2009 characterized by a similar nest search effort between age classes were used to build nest webs and compare their network characteristics.
For each nest web, strength of interactions was measured for each link between species (trees–excavating agent and excavating agent–non-excavating users) and was calculated separately between the tree and the excavator level and the excavator and non-excavator level. The linkage strength between a tree species and a cavity formation agent (either natural decay or an excavator) was defined as the proportion of cavities produced by the cavity formation agent in a given tree species. The linkage strength for cavity reuse by cavity excavators (woodpeckers), facultative cavity excavators (nuthatches and chickadees) or non-excavator users, was calculated as the proportion of total nests located in cavities not excavated by the focal species. In our nest webs, three categories of linkage strength were used: weak (< 10%), medium (11–50%), and strong links (> 50%).
Network characteristics of nest webs
Nest webs were characterized with four different network characteristics including interaction richness, species richness, number of links and mean link strength. Interaction richness was defined as the number of different interspecific interactions found in nest webs. Every used cavity was categorized on the basis of its interspecific interactions stemming from the three different levels: the cavity-bearing tree level, the cavity formation level and the non-excavator user level (Dyer et al., 2010). Species richness was calculated by summing all species identified in each nest web i.e., cavity-bearing trees, excavators and non-excavators. The number of links was calculated by summing all connections between cavity-bearing trees and cavity formation agents (natural decay and excavator species) and between excavators and non-excavator users. Mean link strength was calculated by summing all link strengths divided by the total number of species in the nest web.
To verify if network characteristics changed along the age gradient, observed nest webs were compared with simulated nest webs created by the random sampling of the complete boreal mixedwood nest web. To do so a bootstrap procedure with replacement of 10 000 permutations was used to generate nest webs and calculate confidence intervals for each network characteristic. For each nest web, we used the observed abundances of used cavities (80 occupied cavities in stands between 61 and 89 years after fire, 106 occupied cavities in stands of 90 to 149 years after fire, 70 occupied cavities in stands of 150 to 244 years after fire, and 40 occupied cavities in stands of > 245 years after fire) as the sample size of nests to construct our randomly sampled nest webs. We tested the null hypothesis that interspecific interactions in nest webs were randomly distributed between age-class categories thus expecting to find the different network metrics of our nest webs to fall between the 95% confidence interval for each age-class category of simulated nest webs.
Interactions between cavity producers and users
We assessed the probability of cavity occupancy by cavity users (a cavity used by a bird or mammal species other than the one that created it) and identified key cavity producing agents in nest webs along the stand age gradient of our study plots. This analysis was performed using cavity monitoring data collected between 2008 and 2012. A cavity was considered occupied if it contained eggs or fledglings for birds, if one or more individuals for mammals were found during cavity inspection or if mammals’ nest material (loose branches and leaves) filled the cavity without an individual. Based on our empirical observations these filled cavities were occupied by red squirrels (Tamiasciurus hudsonicus) and northern flying squirrels (Glaucomys sabrinus) denning sites.
For each cavity we determined its probability of occupancy by cavity users as a function of the excavator’s identity and its position along the stand age gradient. We used generalized mixed models with a binomial error distribution of the response variable (use or non-use of a cavity) and a logit link (Bates et al., 2012). Since some trees had more than one cavity and some cavities were inspected multiple times (repeated measures), the variable TreeID was used as random effects to ensure that the hierarchical structure of the sampling design was considered. We used a theoretical-information approach to compare the strength of support of a list of 7 candidate models constructed using combinations of the variables Excavator (5 categories), Forest Age (time since fire; 7 categories, ordinal, but treated as a numeric continuous variable) and Year (Burnham and Anderson, 2002; Anderson, 2008). We conducted separate analyses for cavities where non-cavity users were found than those filled with mammals’ nest material.
Results
Forest structural development along the time since fire gradient
Aspen, paper birch, balsam fir, white cedar and white spruce were co-dominant tree species across forest cover types. As expected, forest vegetation was found to follow a transition in tree species composition along the time since fire gradient from a shade-intolerant deciduous to a mixed and a dominant coniferous forest cover for canopy trees (Figure 2). This is particularly the case for trembling aspen, a tree species highly selected by cavity users because of its vulnerability to heart-rot fungi (Phellinus tremulae). However, Figure 3 shows an increase in the probability for stems of trembling aspens to harbor heart rot disease as stand age increases, despite its net reduction in basal area. This increased probability for aspens to be infected by heart rot disease is also linked with the size of trembling aspens stems where mean DBH increased significantly from mature stands (32.4 cm) to old forests (43.6 cm; Table 1). When combining all tree species, mean diameter, amount of dying and dead trees and trees structural diversity defined by tree decay increased significantly along the age gradient, particularly when forest become older than 150 years old (Table 1).
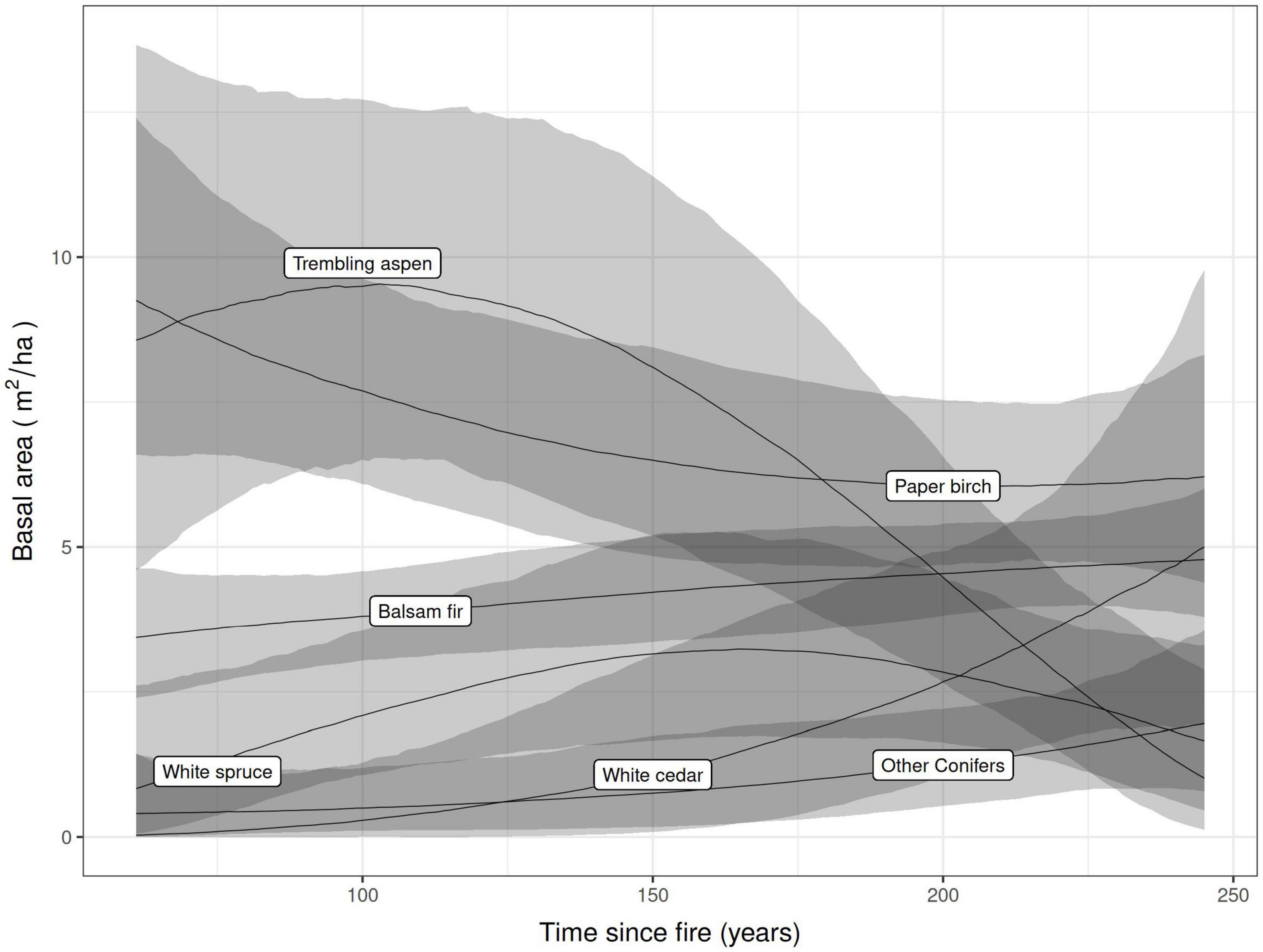
Figure 2. Polynomial mixed effects models of tree species basal area in relation with time since fire in the 12 forest plots where cavity-using communities were monitored. The gray shaded ribbons represent 95% confidence intervals.
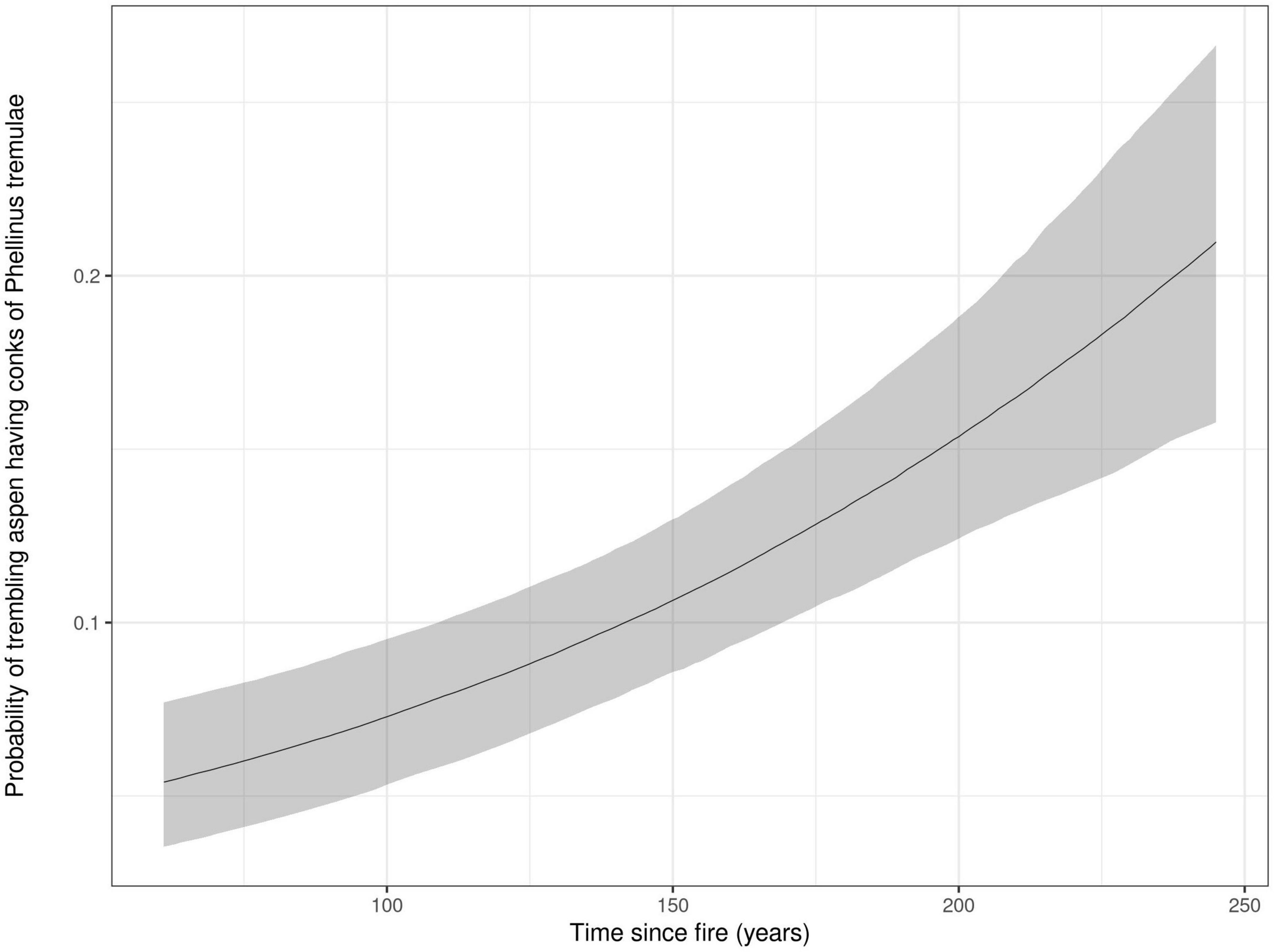
Figure 3. Probability of trembling aspen bearing Phellinus tremulae fungal conks in relation to forest stand age. The gray shaded ribbons represent the 95% confidence intervals.
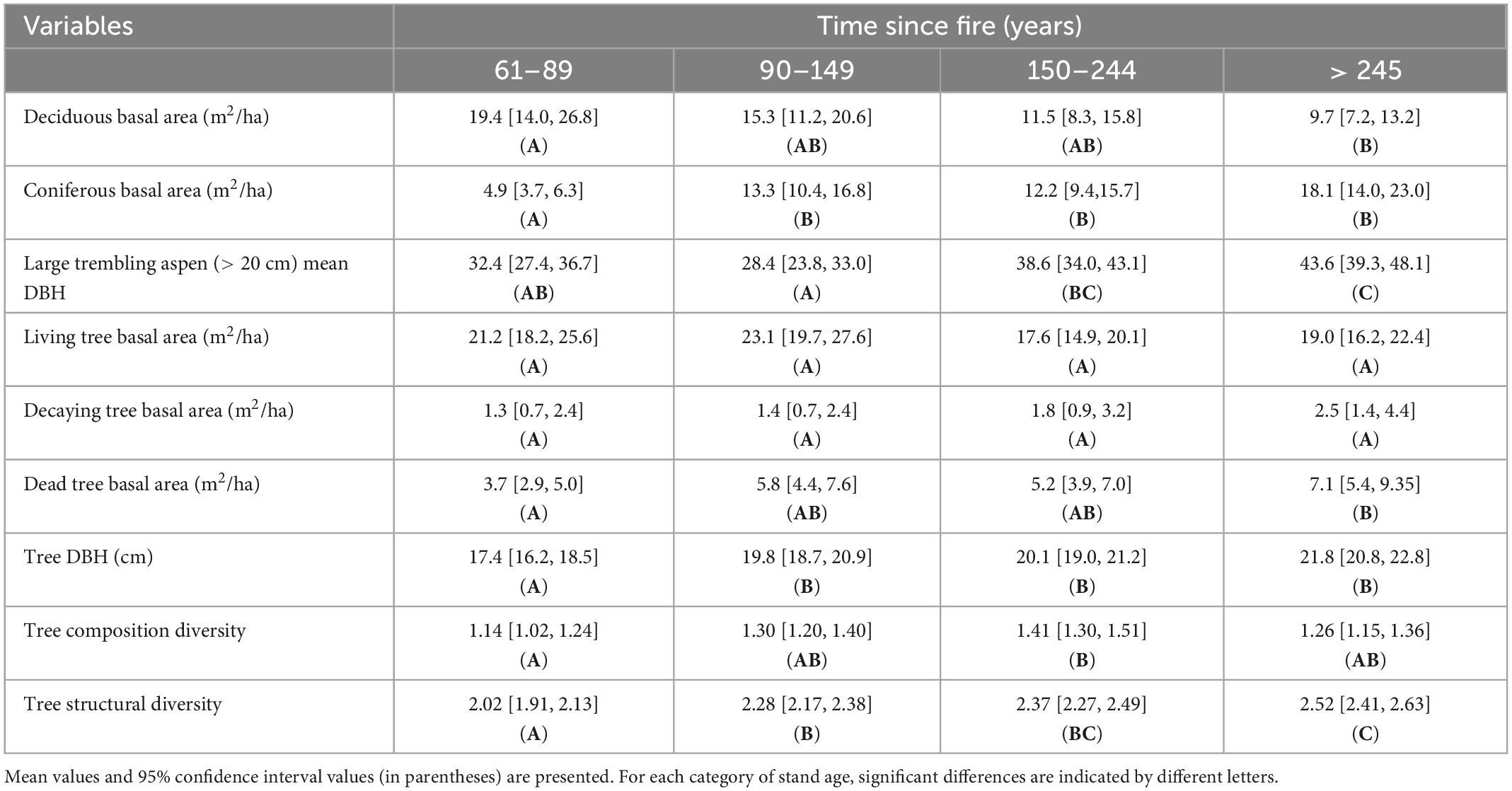
Table 1. Stand characteristics of the twelve forest stands of our study design for assessing nest webs along the age gradient of forest cover types in the Lake Duparquet Research and Teaching Forest in the Abitibi region, Québec, Canada.
The cavity-using vertebrate community
During 2003–2012 we collected data on 423 occupied cavities of 14 species of cavity users, which were located in 4 tree species (the complete boreal mixedwood nest web in Figure 4). All excavators had strong links with the trembling aspen which harbored 95% of nests. Apart from two occupied nests found in non-excavated natural cavities, the non-excavators exclusively reused cavities excavated by excavators. Occupied cavities were created by 7 excavators; 2 species of weak cavity excavators (nuthatches and chickadees) and 5 strong cavity excavators (woodpeckers). Overall, 7 species of non-excavating users were found.
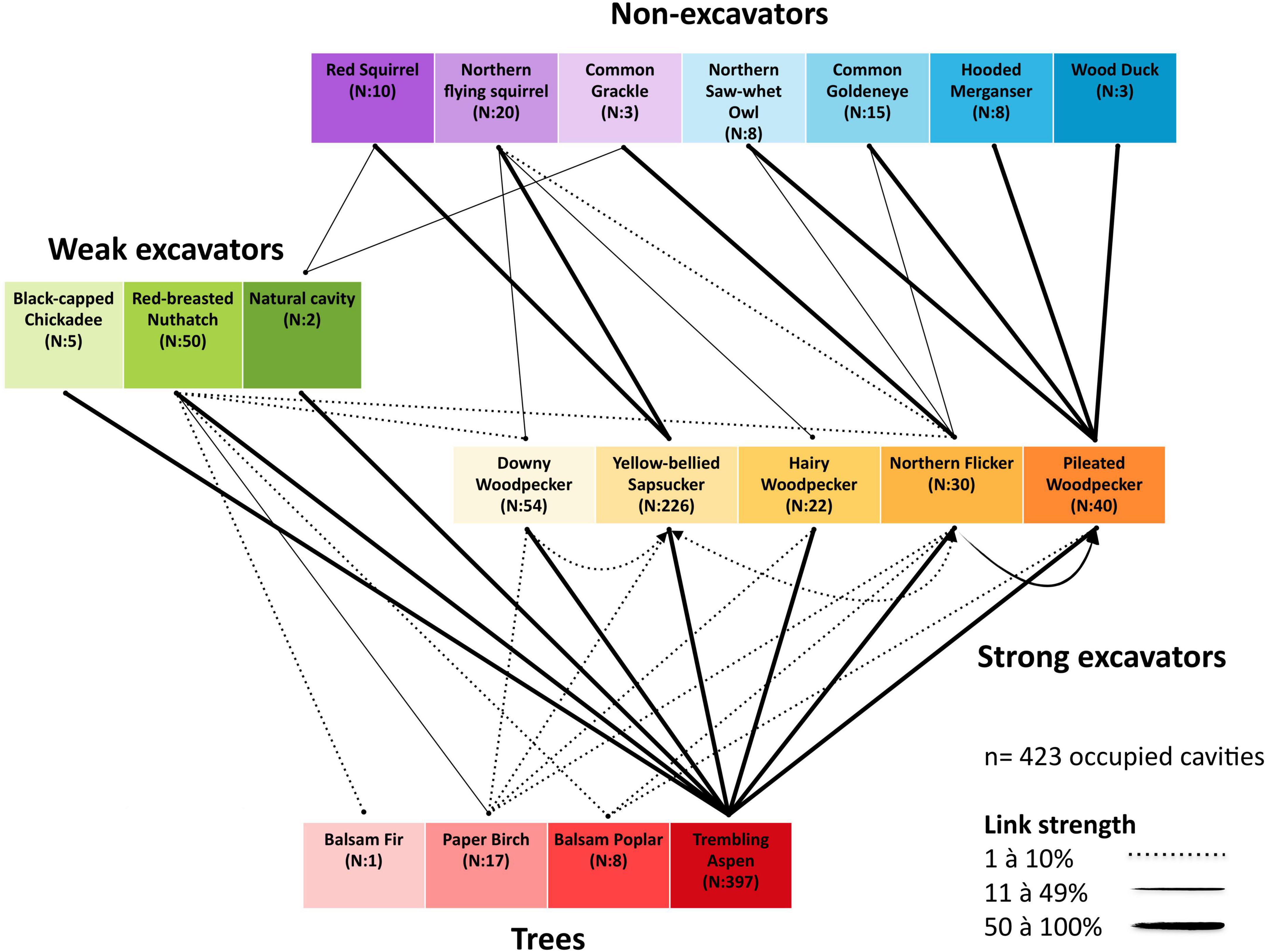
Figure 4. Complete boreal mixedwood nest web in Lake Duparquet, Abitibi, Québec. Nest data were collected from 2003 to 2012. Lines between species represent the use of a cavity resource. N is the number of nests found. Links between the excavator level and the cavity-bearing tree level represent the proportion of nests found in a tree species and links between non-excavator users and excavators or natural cavities represent the proportion of nests found in reused cavities.
Small to medium-sized cavities were produced by three woodpecker species, the Downy Woodpecker (Picoides pubescens), the Yellow-bellied Woodpecker (Sphyrapicus varius) and the Hairy Woodpecker (Picoides villosus). The Yellow-bellied Sapsucker was the dominant excavator with the highest relative nest abundance and was the supplier of numerous small cavities used by small-bodied non-excavators. Two small-bodied non-excavators, the red squirrel and the northern flying squirrel, had strong links with sapsucker cavities. However, the latter also had medium links with Downy Woodpecker cavities. The majority of large-bodied non-excavators reused Pileated Woodpecker cavities. However, the Northern Saw-Whet Owl (Aegolius acadicus) and the Common Goldeneye (Bucephala clangula) had medium links with Northern Flicker cavities. Excavators reuse of cavities excavated by other species was observed for almost every species, but not for the Hairy Woodpecker and the Pileated Woodpecker. Notably, the Northern Flicker had intermediate links with the Pileated Woodpecker cavities (Figure 4).
Nest webs along the age gradient
Important changes in nest webs’ structure occurred at different stages of forest succession. Compared to older forest nest webs, the mature deciduous forest nest web (61–89 years after fire) was simplified (Figure 5). This network consisted in a very limited number of cavity-nesting species that was substantially reduced in complexity. Moreover, the links between Pileated Woodpecker, Northern Flicker cavities and large-bodied non-excavating users were restricted to the 89 years old stand plot as shown with species with an asterisk (*) in Figure 5. The other two 61 years old plots harbored no large cavity dwellers and users (Figure 5). Trembling aspen was the only cavity-bearing tree in this nest web. In the three other stand age classes whereas all excavators maintained a strong link with trembling aspen the use of other cavity-bearing tree species increased from 0 to 3% to 17 to 10% along the age gradient. The small-bodied non-excavating users had strong links with the Yellow-bellied Sapsucker cavities, except in older forest nest webs where they seemed to use a wider range cavity resources. All large-bodied non-excavating users had strong links with the Pileated Woodpecker cavities (Figure 5).
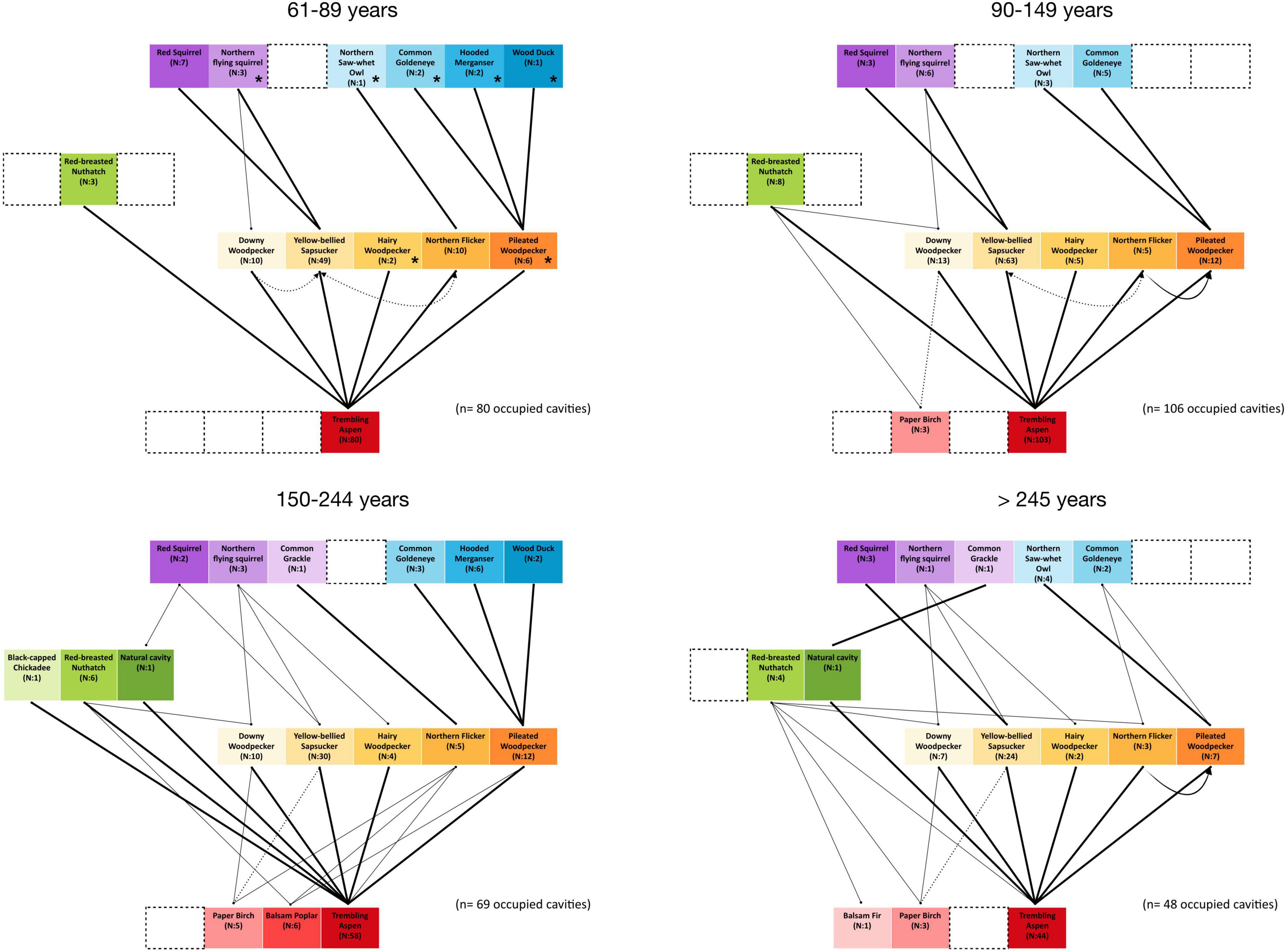
Figure 5. Nest webs along the forest stands age gradient for 61–89, 90–149, 150–244, and over 245 years after fire in Lake Duparquet, Abitibi, Québec. Nest data were collected from 2003 to 2005 and in 2009. Lines between species represent the use of a cavity resource. N is the number of nests found. Links between the excavator level and the cavity-bearing tree level represent the proportion of nests found in a tree species and links between non-excavator users and excavators or natural cavities represent the proportion of nests found in reused cavities. In the 61–89 years old nest web, species with an asterisk (*) were only detected in the 89 years old stand.
Network complexity increased along the age gradient even though there was a decrease in nest abundance. Network metrics such as interaction richness, species richness and mean number of links per species increased as the forest age increased and were significantly higher than predicted by random sampling of the complete boreal mixedwood nest web (Figure 6). The average link strength was significantly lower than predicted in older forests (150–244 years after fire and > 245 years after fire).
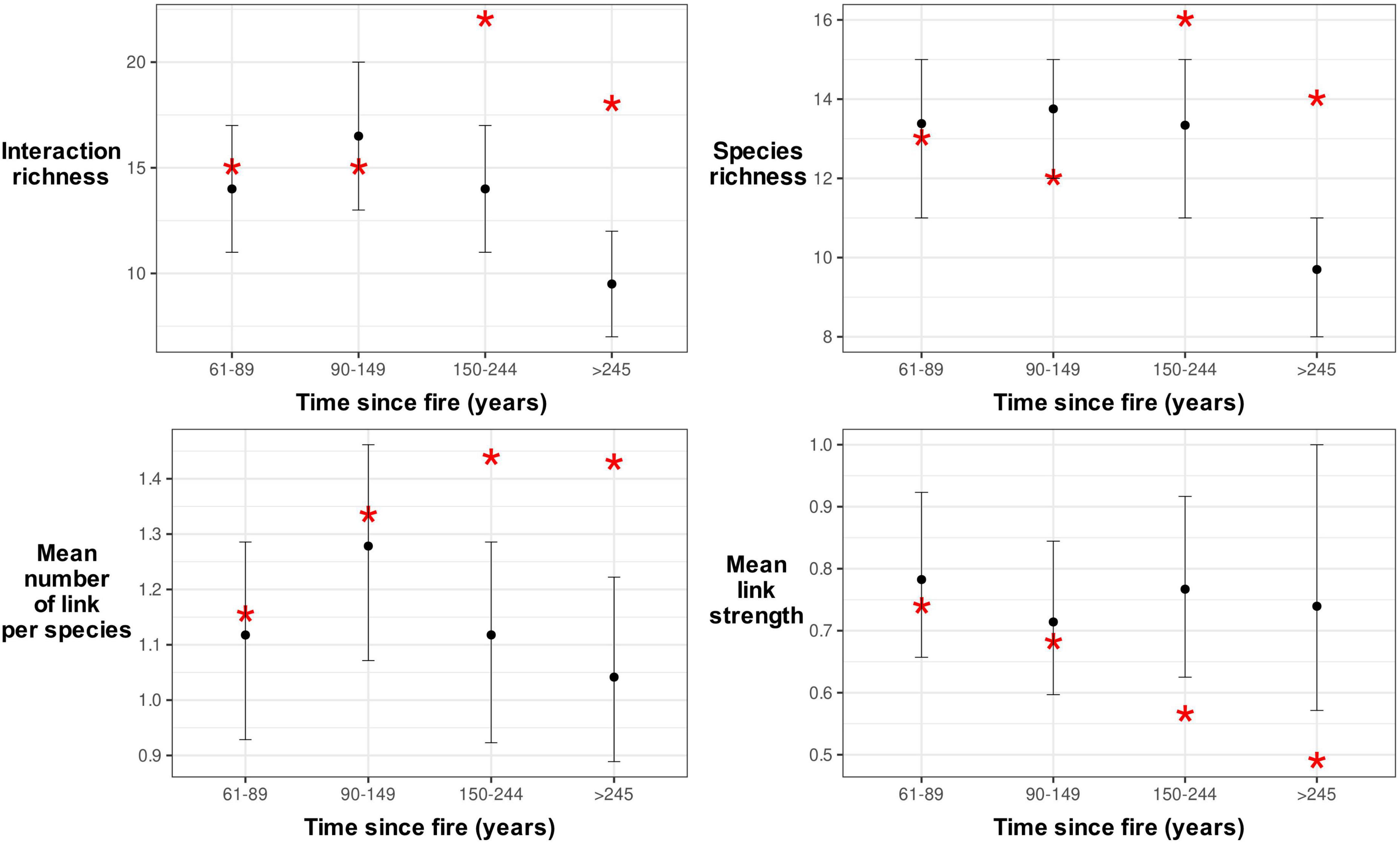
Figure 6. Network characteristics along the age gradient at the Lake Duparquet Research and Teaching Forest in Abitibi, Québec. Empirical network measures from our data (indicated by asterisks) were compared with those from expected simulated data. Random sampling with replacement of the complete boreal mixedwood nest web was used to build an expected distribution of interspecific interactions for each age class. Characteristics that are significantly different from those simulated are outside the 95% confidence interval depicted by a line for each age-class category.
Interactions between cavity producing agents and non-excavating users
We conducted 458 cavity inspections on 273 cavities located on 179 trees. These cavities were excavated by 6 different excavators: the Red-breasted nuthatch, the Downy Woodpecker, the Yellow-bellied Sapsucker, the Hairy Woodpecker, the Northern Flicker and the Pileated Woodpecker. However, cavities excavated by the Red-Breasted Nuthatch were excluded from this analysis because no other cavity user was found in this cavity type despite our inspection efforts (0 nests from non-excavators for 29 inspections in 25 nuthatch cavities).
Cavity users mainly occupied holes excavated by specific excavators whereas less than 1% of occupied cavities were in holes originating from natural decay. Models which included Excavators and Forest Age (Time since fire) had strong support (Wi = 0.82) for all excavators while the closest following models being Excavators only with a low weight (Wi = 0.13; Table 2). Using the predicted probabilities of multi-model inference, we plotted the probability of a cavity being occupied considering its excavator (Figure 7). Cavities excavated by the Pileated Woodpecker were significantly more likely to be occupied than cavities excavated by all other excavators with the exception of the Downy Woodpecker (Figure 7). Although confidence intervals were large in each case, cavities were more likely to be occupied by cavity users as forest was aging along our time since fire gradient (Figure 7).
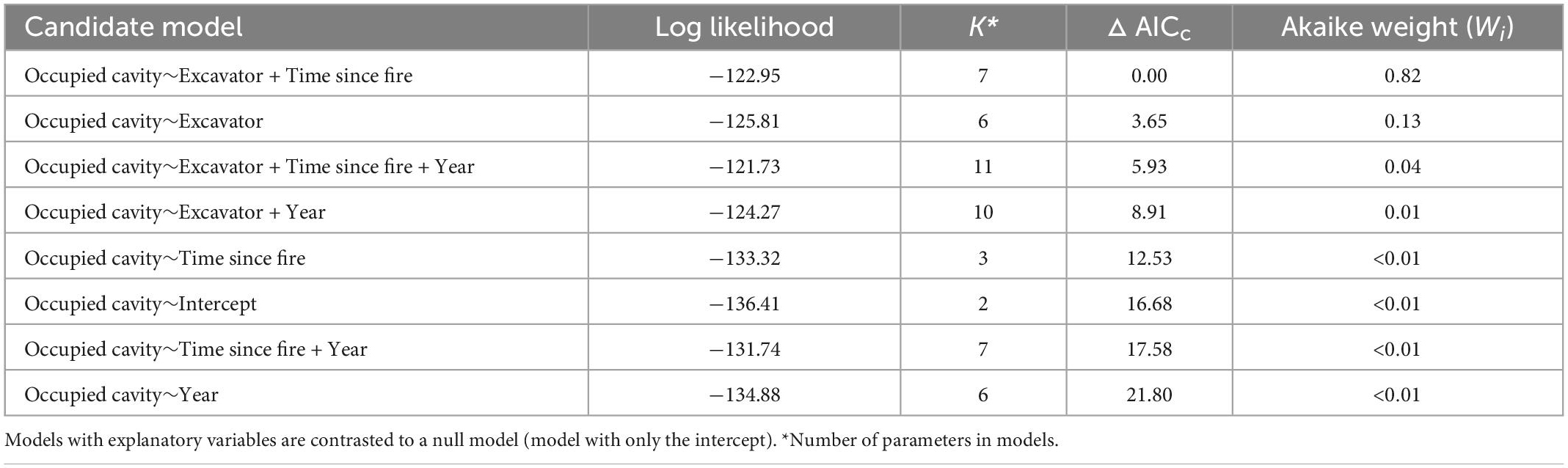
Table 2. Model selection based on Akaike’s Information Criterion corrected for small samples (AICc) for estimating the probability of cavity occupancy by cavity users in relation to excavator species, time since last fire, and sampled year.
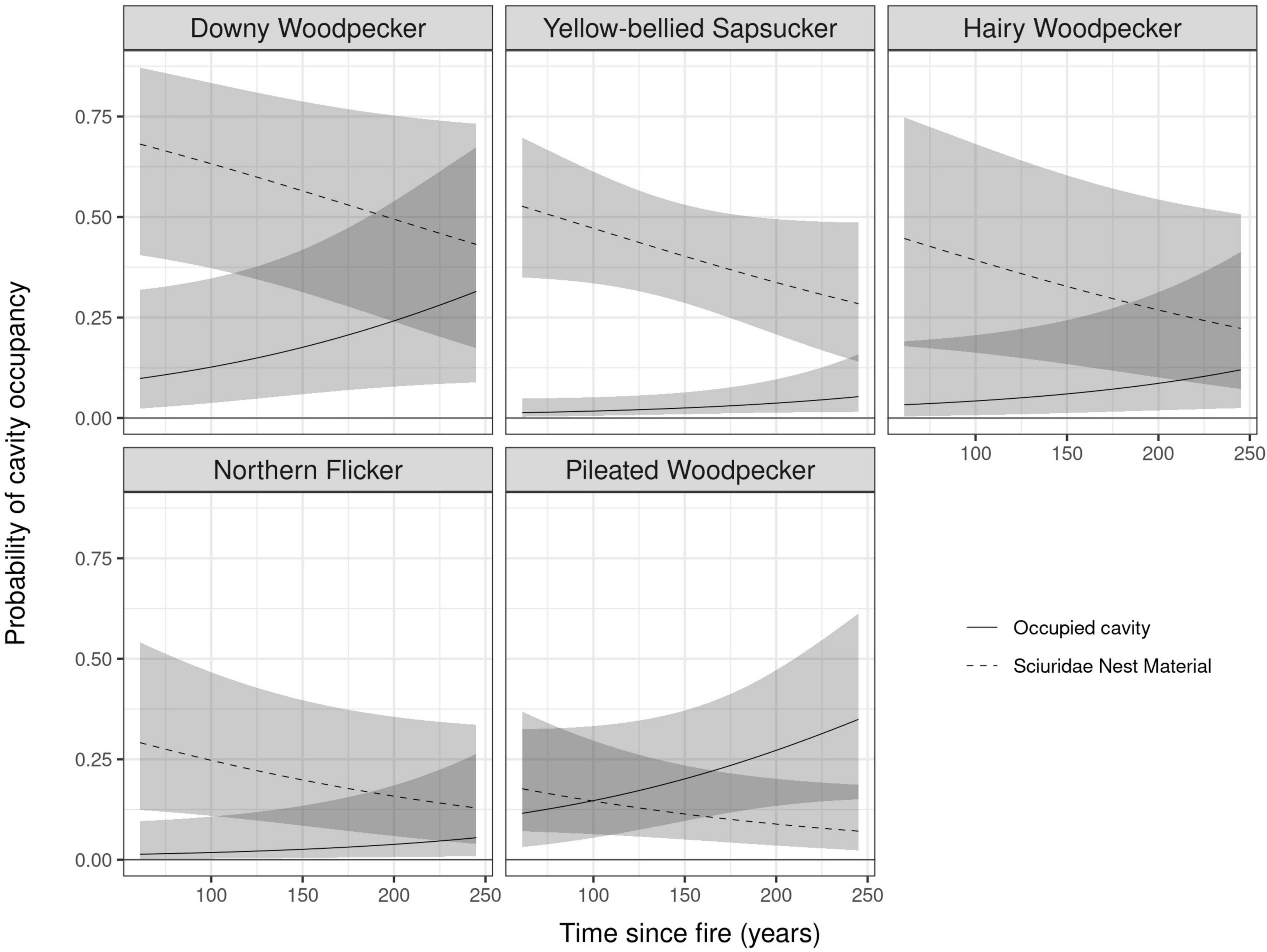
Figure 7. Probability that a cavity excavated by a woodpecker will be used by a cavity user (full line) or will be filled with nest material by sciuridae species that makes it unusable for other species (dotted line) along a time since fire gradient of forest structural development stages in a boreal mixedwood natural forest landscape in eastern Canada. The 95% confidence intervals are represented by the gray ribbons.
For Sciuridae nest material analysis, the model that included Excavators, Forest Age (Time since fire) and Year had strong support (Wi = 0.83; Table 3). The model that included Excavators and Year was the closest following model (Wi = 0.12; Table 3). The Year effect in our best models highlights the fact that in the years 2011 and 2012 we deliberately under inspected Yellow-bellied Sapsucker cavities (a main cavity type filled with Sciuridae nest material) and prioritized inspections in cavities created by other excavators. Multi-model inference analyses of our competing models show that Sciuridae nesting material had a greater probability of being found in small cavities excavated by the Downy Woodpecker, the Yellow-bellied Sapsucker and the Hairy Woodpecker, than the Pileated Woodpecker and Northern Flicker, although Hairy Woodpecker had large confidence intervals similar to those of Northern Flicker (Figure 7). The probability of a cavity having Sciuridae nesting material decreased with stand age regardless of cavity origin (Figure 7) resulting in higher availability of cavities for other non-excavating species.
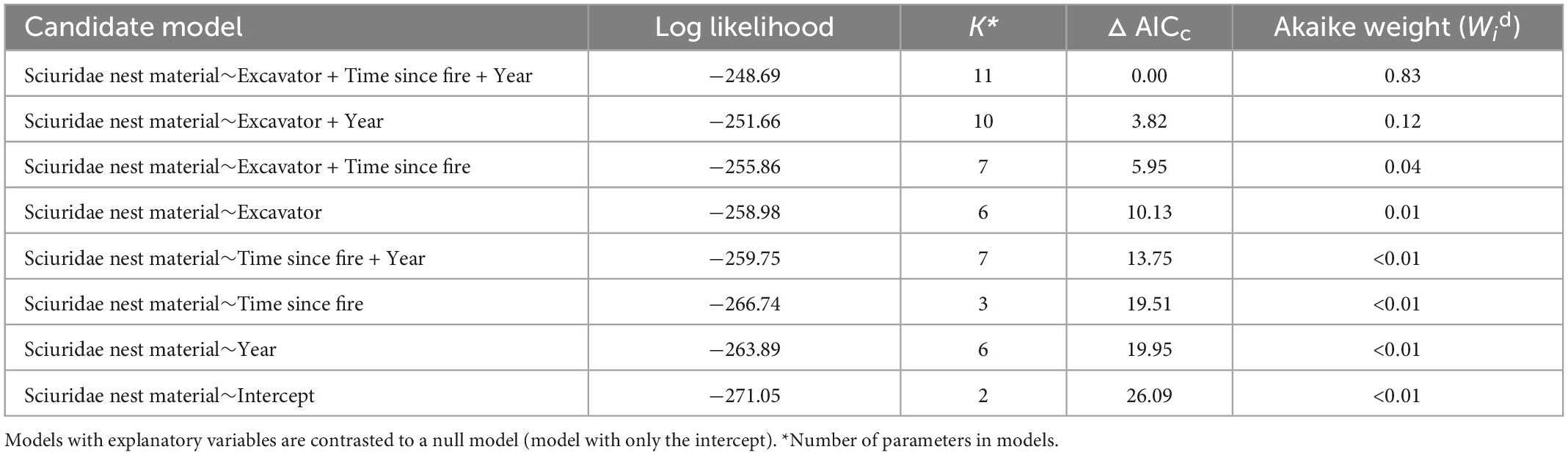
Table 3. Model selection based on Akaike’s Information Criterion corrected for small samples (AICc) for estimating the probability of cavity occupancy by Sciuridae nest material in relation to excavator species, time since last fire, and sampled year.
Discussion
Our main hypothesis, that network complexity increased along the age gradient of boreal mixedwood forest, was supported. Network complexity, quantified by species richness and interaction richness among species, were indeed higher than expected in older forest nest webs (> 150 years after fire). Moreover, as we predicted, this complexity was driven by a higher functional redundancy or equivalence of all hierarchical levels of the nest web. Diversification of tree species used by cavity excavators (links with more than one tree species), the main cavity formation agents in our nest webs, and the increased number of non-excavating cavity users, accounted for the greater complexity of nest webs in older forests.
Increase in network complexity of nest webs with forest structural development
Our study thus shows that forest structural development is a driver of complexity for ecological networks in the boreal mixedwood forest when using nest webs as a model. Along the forest age gradient, we found that the increased availability of structural elements such as large live and dead trees and a growing diversity of tree decay stages provided key structures for cavity formation by woodpeckers which in turn increased the complexity of interactions with non-excavating users.
Although we found a higher functional redundancy or equivalence between cavity substrates (tree species) and avian excavators in our oldest stands compared with our mature stands, the interactions between tree species and cavity formation agents were not as diversified as those found in tropical nest webs where a much larger number of tree species harbor cavities formed by either natural decay or woodpecker excavations (Cockle et al., 2012). Hence, whereas more complex in old boreal forests, our nest webs maintained throughout the age gradient an architecture being strongly structured around trembling aspen as the keystone cavity substrate.
Importance of trembling aspen for nest webs architecture throughout stand age gradient
The fact that a majority of nests were found in trembling aspen throughout our age gradient and even in our oldest conifer-dominated stands is noteworthy. Whereas several studies conducted in boreal and hemiboreal forests of North America have shown that cavity-nesting communities are strongly linked to trembling aspen in ecosystems where this tree species is naturally present (Li and Martin, 1991; Dobkin et al., 1995; Martin et al., 2004; Cooke and Hannon, 2012), our study is the first to show how this close association between trembling aspen and cavity-using communities is consistent across the entire range of forest age and composition in a natural successional gradient. The persistence of the shade-intolerant trembling aspen along forest succession including our oldest conifer-dominated forest stands is associated to its recruitment by root suckers when canopy gaps occur in old forests due to small to medium size disturbances of either single-tree gap replacement or spruce budworm outbreaks (Lavertu et al., 1994; Kneeshaw and Bergeron, 1998; Bergeron, 2000). When a disturbance occurs trembling aspen is one of the fastest boreal tree species to fill canopy gaps and reach a large diameter (Paré and Bergeron, 1995). For instance, large aspen individual trees and small patches that were indeed found in our 245 years old stands are likely associated with the recurrent outbreaks of spruce budworm that occurred throughout the 20th century (Morin et al., 1993) and that affected canopy cover of old balsam fir forests at variable severities (Bergeron, 2000). Hence, despite an important decrease in its availability along our age gradient, the quality of trembling aspen individual trees for cavity excavation persisted as its size and its propensity to be infected by heartrot fungi (Phellinus tremulae) were significantly higher in older forests (Figure 3, Table 1). This could explain why a majority of cavity nesters still strongly selected trembling aspen in older forests that can thus be considered a keystone structure (sensu Tews et al., 2004) in the eastern boreal mixedwood forests nest webs. Moreover, the strong association of cavity excavators with trembling aspen in the nest web architecture in our study area is likely to reflect evolutionary adaptation of cavity excavators to interactions between historical natural disturbance fire regimes (Drapeau et al., 2016) and spruce budworm cyclic outbreaks (Morin et al., 1993) of eastern boreal mixedwood forests that are characterized by high proportions of the forest cover with stands older than 100 years (Bergeron et al., 2004) where trembling aspen is persisting and available as a cavity-bearing tree along the entire stand age gradient (Bergeron and Charron, 1994; Bergeron, 2000).
Increases of interactions between cavity producers and users along stand age gradient
Complexity of nest webs along our age gradient also increased at the level of interactions between cavity producing agents (woodpeckers) and non-excavator users. Increases in tree structural diversity and corresponding availability of a wider range of tree decay stages also provided a wider range of cavity substrates for weak excavators such as Red-breasted nuthatches, Black-capped Chickadees (Poecele atricapillus) in older forest stands.
For numerous studies on cavity nesters in North America, an important pattern has emerged: only one or sometimes two excavator species provide the bulk of occupied cavities in the nest web (Martin et al., 2004; Blanc and Walters, 2008; Cooke and Hannon, 2012). In our study, the cavities of the Yellow-bellied Sapsucker and the Pileated Woodpecker represented 63% of all cavities found. The Pileated Woodpecker was a keystone excavator in our study since 57% of non-excavating species reused its cavities although they represented less than 10% of the available cavities in the nest web (Figure 3). Moreover, the Pileated Woodpecker maintained its status across the entire range of forest age and composition of our natural successional gradient (Figure 4). Although a number of studies have found this species to be a keystone excavator (Bonar, 2000; Aubry and Raley, 2002; Martin et al., 2004; Cooke and Hannon, 2011), to our knowledge, it is the first time that the functional role of Pileated Woodpecker is confirmed across a successional age gradient ranging from mature to old forest stands.
Contrary to other well-studied nest webs in North America (Martin et al., 2004; Edworthy et al., 2018), the Northern Flicker played a less important role as a large-cavity provider in our continuous forest landscape mosaïc with few forest edges and open habitats. It became, however, an important provider in remnant habitats (riparian buffers and cutblock separators) in managed forest landscapes nearby (Ouellet-Lapointe et al., 2012) as this generalist species is mostly associated to open woodlands (Wiebe and Moore, 2020). Two of its cavities were occupied by large non-excavating users strongly associated with the Pileated Woodpecker cavities, thus providing some functional redundancy as a large cavity provider for cavity-using birds and mammals in boreal habitats where Pileated Woodpecker cavities are absent.
A point often overlooked in most studies on cavity users but underlined by Cooke and Hannon (2012), is the probable underestimation of cavity use by non-obligate cavity nesters, like Sciuridae. Our investigation of Sciuridae nest material (branches, straw, leaves, or bark) shows that almost half of small cavities excavated in our study plots had nest material importation showing that small cavities were not only heavily used by Sciuridae but that this behavior probably limited their availability to other non-excavating users (Figure 7). This result corroborates Cooke and Hannon (2012) observations of chew marks on 45% of Yellow-bellied sapsucker cavity entry holes. In our study, we found a clear preference by Sciuridae (red squirrel and northern flying squirrel) for all smaller cavities regardless of the excavator with cavities formed by Downy Woodpecker with the highest probability of being used. Large cavities such as those created by the Northern Flicker and the Pileated Woodpecker had, however, the lowest probability of being utilized by squirrels (Figure 7).
Whereas the Yellow-bellied Sapsucker was the dominant small-cavity excavator with the highest nest abundance, cavities created by other small excavators like the Downy Woodpecker and the Hairy Woodpecker had very similar probability of being reused by cavity users than cavities of Yellow-bellied Sapsucker suggesting good functional redundancy of small cavity excavators for small cavity availability (Figure 7).
Another interesting result is the increase in the probability of occupancy of small cavities by cavity users along our stand age gradient which coincides with a decrease in the probability of Sciuridae nest material in small cavities for three woodpecker species of small excavators (Figure 7). This result corroborates Trudeau et al. (2011) study on northern flying squirrel tree-cavity use in our region which is indeed less in old conifer-dominated forests than in deciduous and mixed wood stands of earlier successional stages. In old coniferous stands, northern flying squirrels often nest in coniferous trees instead of tree cavities. Sciuridae are facultative cavity users often occupying multiple cavities and rendering many useless for other cavity dwellers by filling them with nest material (leaf, twigs, bark, and other vegetation). In addition to competing with small cavity nesters, Sciuridae, are also important nest predators (Wiebe et al., 2007) and could explain why few small cavity nesters reuse available cavities in boreal mixedwoods. In older forests small cavities are thus more available to a greater number of non-excavating species because of lesser use by the Sciuridae providing another insight why nest webs in these forests are more complex in terms of species richness, number of interactions and mean number of links between cavity producers and users (Figure 6).
Resilience of eastern boreal mixedwood nest webs
We clearly found that under natural disturbance regimes, boreal mixedwood old-growth stands harbor more complex nest webs that have higher interactions, weaker links and that are more robust to species lost (sensu Dunne et al., 2002) than those of younger forest stands.
Nest webs increased structural complexity in old forests also suggest that boreal mixedwood cavity-using vertebrate communities have evolved to be resilient to natural disturbances (ex: wildfires, insect outbreaks) and even to thrive on them as we see with mechanisms of continuous recruitment of the trembling aspen throughout forest succession along with the increasing diversity of decaying stages of other tree species, two drivers of the complexity of nest webs as forests are aging. Our study also clearly shows the pivotal contribution of old forests to the overall complexity of nest webs in the boreal mixedwood zone. This complexity of nest webs in old forests is thus a key for the resilience of the entire cavity-using vertebrate community to natural disturbance regimes. In managed landscapes, such resilience of the cavity-using community may, however, be compromised by even-aged industrial timber harvesting with short rotations that shifts the age structure of boreal landscapes toward regenerating and young pole forests and consequential loss of mature and old-growth forests (Bergeron et al., 2002, 2004). As the proportion of older forests becomes reduced, often falling outside its natural range of variability (Cyr et al., 2009), the complexity, stability (Barabási, 2009; Simard et al., 2013), and robustness to species’ loss (Dunne et al., 2002) of old forest nest webs, are likely to be altered. Therefore, even though landscapes may become dominated with young and mature aspen stands, the size, decay status and degree of fungi invasion of trees may be too low to provide quality substrates for excavation by woodpeckers (Drapeau et al., 2009a). This is exactly what we see with 61 years stands (Figure 5) that do not support large excavators and cavity users and that have simplified nest webs (Figure 5). The capacity of the cavity-using vertebrate community to bounce back in such human-disturbed regimes may thus be jeapardized by the strong reduction of the proportion old forests stands at the landscape level.
Conclusion
This study increases our understanding on the ecological processes that shape nest webs in the boreal mixed forest of eastern Canada and is among the first to document such patterns along an entire natural forest succession gradient. Old growth boreal forests take different structural profiles whereas continuous secondary disturbances that take place throughout stand dynamics (Martin et al., 2018, 2021). In our study system, disturbance dynamics of old forests (gaps and insect outbreaks) allows a pioneer fast growing short-lived species like the trembling aspen to persist in the forest canopy throughout succession, thus providing, along with increased decay stages of other tree species, continuous recruitment of suitable cavity substrates for excavators (mainly woodpeckers), the major cavity formation agents.
The critical importance of conserving old forest stands and old trees is obvious for cavity-using vertebrate communities that are highly dependent on processes of tree cavity formation by natural decay of old trees (Gibbons and Lindenmayer, 2002; Cockle et al., 2010, 2012; Politi et al., 2010). Our study shows that old forests are also keystone habitats for cavity-using communities that mainly rely on vertebrate excavators (less than 1% of used cavities in our nest webs are formed solely by tree decay). In such old stands, wood-decay processes of old trees (heart-rot fungi activity) become facilitator agents for woodpeckers, as most of the trees excavated by woodpeckers are affected by tree fungi.
Our results raise awareness on the importance of protecting the entire gradient old forest cover types at landscape and regional scales for maintaining the avifauna associated to old forests and wood-decaying trees (Imbeau et al., 2001; Drapeau et al., 2009b,2016). However, the conservation of old forest tracks alone is not sufficient in the context of extensive forestry presently occurring in the Canadian boreal forest. There is also a critical need to adjust forestry practices in managed forest landscapes to increase at the landscape level the amount of old forests by combining the use of longer rotations, partial harvesting under continuous forest cover (Bergeron et al., 2002; Gauthier et al., 2009) and in managed clear-cut landscapes, enhanced retention through a mesofilter approach (sensu Hunter, 2005) of keystone tree species for a range of tree decay stages (structural heterogeneity generated by tree senescence) for key excavators (Martin et al., 2004; Drapeau et al., 2009b; Drever and Martin, 2010; Edworthy and Martin, 2013) and other dendromicrohabitats that are critical for biodiversity (Larrieu et al., 2018; Martin et al., 2022).
Future research could explore how the network structure of nest webs and the diversity of species and interactions changes as we move north, from boreal mixedwood to coniferous-dominated boreal forests where trembling aspen stands becomes rare and scattered but deadwood associated birds are nevertheless strongly associated to this pivotal forest cover type (Cadieux and Drapeau, 2017).
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was reviewed and approved by Université du Québec à Montréal, Comité institutionnel de protection des animaux.
Author contributions
PC, PD, and AN conceived the ideas, study design, and methodology. RD, PD, PC, UO-L, and AN collected the data. PC performed the analyses and wrote the first draft of the manuscript. PD wrote the second draft. LI, UO-L, AL, and AN interpreted the results. All authors contributed critically to the drafts and gave final approval for publication.
Funding
The funding was provided by a Natural Sciences and Engineering Research Council of Canada (NSERC) graduate student scholarship to Cadieux, the Québec Fonds de la recherche nature et technologies (FRQNT) graduate student scholarship to Cadieux, a Centre for Forest Research scholarship for research communication, an NSERC Discovery grant to Drapeau, and logistic support from Lake Duparquet research and teaching forest station for lodging facilities throughout this project.
Acknowledgments
We thank for their assistance in the field the numerous undergraduate students that helped us gather data on cavity users during the years of this long-term study. We also thank Dr. Marc Mazerolle for statistical advice and Mélanie Desrochers from the Centre d’étude de la forêt (CEF)–Centre for forest research (CFR), for GIS support.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aitken, K., and Martin, K. (2007). The importance of excavators in hole-nesting communities: Availability and use of natural tree holes in old mixed forests of western Canada. J. Ornithol. 148, 425–434. doi: 10.1007/s10336-007-0166-9
Anderson, D. R. (2008). Model-based inference in the life sciences: A primer on evidence. New York, NY: Springer. doi: 10.1007/978-0-387-74075-1
Aubry, K. B., and Raley, C. M. (2002). “The Pileated Woodpecker as a keystone habitat modifier in the Pacific Northwest,” in Procedings of the symposium on the ecology and management of dead wood in western forests (Reno, NV, November 2–4, 1999), eds W. F. Laudenslayer, P. J. Shea, Jr. B. Valentine, C. P. Weatherspoon, and T. E. Et Lisle (Alabany, NY: USDA Forest Service), 257–274.
Barabási, A.-L. (2009). Scale-free networks: A decade and beyond. Science 325:412. doi: 10.1126/science.1173299
Bates, D., Maechler, M., and Bolker, B. (2012). lme4: Linear mixed-effects models using S4 classes On line. Available online at: http://CRAN.R-project.org/package=lme4 (accessed September 1, 2015).
Bednarz, J. C., Ripper, D., and Radley, P. M. (2004). Emerging concepts and research directions in the study of cavity-nesting birds: Keystone ecological processes. Condor 106, 1–4. doi: 10.1093/condor/106.1.1
Bergeron, Y. (1991). The influence of island and mainland lakeshore landscapes on boreal forest fire regimes. Ecology 72, 1980–1992. doi: 10.2307/1941553
Bergeron, Y. (2000). Species and stand dynamics in the mixed-woods of Quebec’s southern boreal forest. Ecology 81, 1500–1516. doi: 10.1890/0012-9658(2000)081[1500:SASDIT]2.0.CO;2
Bergeron, Y., and Charron, D. (1994). Postfire stand dynamics in the southern boreal forest (Quebec): A dendroecological approach. Ecoscience 1, 173–184. doi: 10.1080/11956860.1994.11682241
Bergeron, Y., and Dubuc, M. (1989). Succession in the southern part of the Canadian boreal forest. Vegetatio 79, 51–63. doi: 10.1007/s004420050289
Bergeron, Y., and Harvey, B. (1997). Basing silviculture on natural ecosystem dynamics: An approach applied to the southern boreal mixedwood forest of Quebec. For. Ecol. Manag. 92, 235–242. doi: 10.1016/S0378-1127(96)03924-2
Bergeron, Y., Gauthier, S., Flannigan, M., and Kafka, V. (2004). Fire regimes at the transition between mixedwood and coniferous boreal forest in northwestern Quebec. Ecology 85, 1916–1932. doi: 10.1890/02-0716
Bergeron, Y., Harvey, B., Leduc, A., and Gauthier, S. (1999). Forest management guidelines based on natural disturbance dynamics: Standand forest-level considerations. For. Chron. 75, 49–54. doi: 10.5558/tfc75049-1
Bergeron, Y., Leduc, A., Harvey, B. D., and Gauthier, S. (2002). Natural fire regime: A guide for sustainable management of the Canadian boreal forest. Silva Fenn. 36, 81–95. doi: 10.14214/sf.553
Bergeron, Y., Leduc, A., Morin, H., and Joyal, C. (1995). Balsam fir mortality following the last spruce budworm outbreak in northwestern Quebec. Can. J. For. Res. 25, 1375–1384. doi: 10.1139/x95-150
Blanc, L. A., and Walters, J. R. (2008). Cavity-nest webs in a longleaf pine ecosystem. Condor 110, 80–92. doi: 10.1525/cond.2008.110.1.80
Bonar, R. L. (2000). Availability of Pileated Woodpecker cavities and use by other species. J. Wildl. Manag. 64, 52–59. doi: 10.2307/3802974
Burnham, K. P., and Anderson, D. R. (2002). Model selection and multimodel inference: A practical information-theoretic approach. Berlin: Springer.
Cadieux, P., and Drapeau, P. (2017). Are old boreal forests a safe bet for the conservation of the avifauna associated with decayed wood in eastern Canada? For. Ecol. Manag. 385, 127–139. doi: 10.1016/j.foreco.2016.11.024
Cockle, K. L., Martin, K., and Drever, M. C. (2010). Supply of tree-holes limits nest density of cavity-nesting birds in primaryand logged subtropical Atlantic forest. Biol. Conserv. 143, 2851–2857. doi: 10.1016/j.biocon.2010.08.002
Cockle, K. L., Martin, K., and Robledo, G. (2012). Linking fungi, trees, and hole-using birds in a Neotropical tree-cavity network: Pathways of cavity production and implications for conservation. For. Ecol. Manag. 264, 210–219. doi: 10.1016/j.foreco.2011.10.015
Cockle, K. L., Martin, K., and Wesolowski, T. (2011). Woodpeckers, decay, and the future of cavity-nesting vertebrate communities worldwide. Front. Ecol. Environ. 9:377–382. doi: 10.1890/110013
Cooke, H. A., and Hannon, S. J. (2011). Do aggregated harvests with structural retention conserve the cavity web of old upland forest in the boreal plains? For. Ecol. Manag. 261, 662–674. doi: 10.1016/j.foreco.2010.11.023
Cooke, H. A., and Hannon, S. J. (2012). Nest-site selection by old boreal forest cavity excavators as a basis for structural retention guidelines in spatially-aggregated harvests. For. Ecol. Manag. 269, 37–51. doi: 10.1016/j.foreco.2011.12.042
Cyr, D., Gauthier, S., Bergeron, Y., and Carcaillet, C. (2009). Forest management is driving the eastern North American boreal forest outside its natural range of variability. Front. Ecol. Environ. 7:519–524. doi: 10.1890/080088
Dansereau, P., and Bergeron, Y. (1993). Fire history in the southern boreal forest of northwestern Quebec. Can. J. For. Res. 23, 25–32. doi: 10.1139/x93-005
Dobkin, D. S., Rich, A. C., Pretare, J. A., and Pyle, W. H. (1995). Nest-site relationships among cavity-nesting birds of riparian and snowpocket aspen woodlands in the northwestern Great Basin. Condor 97, 694–704. doi: 10.2307/1369178
Drapeau, P., Leduc, A., and Bergeron, Y. (2009a). “Bridging ecosystem and multiple species approaches for setting conservation targets in managed boreal landscapes,” in Setting conservation targets in managed forest landscapes, Chap. 7, eds M.-A. Villard and B. G. Jonsson (New York, NY: Cambrige University Press), 129–160. doi: 10.1017/CBO9781139175388.008
Drapeau, P., Nappi, A., Imbeau, L., and Saint-Germain, M. (2009b). Standing deadwood for keystone bird species in the eastern boreal forest: Managing for snag dynamics. For. Chron. 85, 227–234. doi: 10.5558/tfc85227-2
Drapeau, P., Leduc, A., Giroux, J.-F., Savard, J.-P. L., Bergeron, Y., and Vickery, W. K. (2000). Landscape-scale disturbances and changes in bird communities of boreal mixed-wood forests. Ecol. Monogr. 70, 423–444. doi: 10.1890/0012-9615(2000)070[0423:LSDACI]2.0.CO;2
Drapeau, P., Villard, M.-A., Leduc, A., and Hannon, S. J. (2016). Natural disturbance regimes as templates for the response of bird species assemblages to contemporary forest management. Divers. Distrib. 22, 385–399. doi: 10.1111/ddi.12407
Drever, M., and Martin, K. (2010). Response of woodpeckers to changes in forest health and harvest: Implications for conservation of avian biodiversity. For. Ecol. Manag. 259, 958–966. doi: 10.1016/j.foreco.2009.11.038
Dunne, J. A., Williams, R. J., and Martinez, N. D. (2002). Network structure and biodiversity loss in food webs: Robustness increases with connectance. Ecol. Lett. 5, 558–567. doi: 10.1046/j.1461-0248.2002.00354.x
Dyer, L. A., Walla, T. R., Greeney, H. F., Stireman, J. O. III, and Hazen, R. F. (2010). Diversity of interactions: A metric for studies of biodiversity. Biotropica 42, 281–289. doi: 10.1111/j.1744-7429.2009.00624.x
Edworthy, A. B., and Martin, K. (2013). Persistence of tree cavities used by cavity nesting verterbrates declines in harvested forests. J. Wild. Manag. 77, 770–776. doi: 10.1002/jwmg.526
Edworthy, A. B., and Martin, K. (2014). Long-term dynamics of the characteristics of tree cavities used for nesting by vertebrates. For. Ecol. Manag. 334, 122–128. doi: 10.1016/j.foreco.2014.09.001
Edworthy, A. B., Trzcinski, M. K., Cockle, K. L., Weibe, K. L., and Martin, K. (2018). Tree cavity occupancy by nesting vertebrates across cavity age. J. Wild. Manag. 82, 639–648.
Esseen, P.-A., Ehnström, B., Ericson, L., and Sjöberg, K. (1997). Boreal forests. Ecol. Bull. 46, 16–47.
Franklin, J. F., Lindenmayer, D., MacMahon, J. A., McKee, A., Magnuson, J., Perry, D. A., et al. (2000). Threads of continuity. Conserv. Pract. 1, 9–16. doi: 10.1016/S0378-1127(01)00575-8
Franklin, J. F., Spies, T. A., Van Pelta, R., Carey, A. B., Thornburghd, D. A., Berge, D. R., et al. (2002). Disturbances and structural development of natural forest ecosystems with silvicultural implications, using Douglas-fir forests as an example. For. Ecol. Manag. 155, 399–423.
Gauthier, S., Leduc, A., and Bergeron, Y. (1996). Forest dynamics modelling under a natural fi re cycle: A tool to defi ne natural mosaic diversity in forest management. Environ. Monit. Assess. 39, 417–434. doi: 10.1007/BF00396159
Gauthier, S., Vaillancourt, M.-A., Leduc, A., De Grandpré, L., Kneeshaw, D. D., Morin, H., et al. (2009). Ecosystem management in the boreal forest. Québec, QC: Presses de l’Université du Québec.
Gibbons, P., and Lindenmayer, D. (2002). Tree hollows and wildlife conservation in Australia. Collingwood, ON: CSIRO Publishing. doi: 10.1071/9780643090033
Grove, S. J. (2002). Saproxylic insect ecology and the sustainable management of forests. Annu. Rev. Ecol. Syst. 33, 1–23. doi: 10.1146/annurev.ecolsys.33.010802.150507
Harmon, M. E., Franklin, J. F., Swanson, F. J., Sollins, P., Gregory, S. V., Lattin, J. D., et al. (1986). Ecology of coarse woody debris in temperate ecosystems. Adv. Ecol. Res. 15, 133–302. doi: 10.1016/S0065-2504(08)60121-X
Harper, K., Boudreault, C., De Grandpré, L., Drapeau, P., Gauthier, S., and Bergeron, Y. (2003). Structure, composition, and diversity of old-growth black spruce boreal forest of the Clay Belt region in Quebec and Ontario. Environ. Rev. 11(Suppl. 1) S79–S98. doi: 10.1139/a03-013
Harvey, B. D. (1999). The lake duparquet research and teaching forest: Building a foundation for ecosystem management. For. Chron. 75, 389–393. doi: 10.5558/tfc75389-3
Harvey, B. D., Leduc, A., Gauthier, S., and Bergeron, Y. (2002). Stand-landscape integration in natural disturbance-based management of the southern boreal forest. For. Ecol. Manag. 155, 369–385. doi: 10.1016/S0378-1127(01)00573-4
Holt, R. F., and Martin, K. (1997). Landscape modification and patch selection: The demography of two secondary cavity nesters colonizing clearcuts. Auk 114, 443–455. doi: 10.2307/4089245
Hunter, M. L. Jr. (2005). A mesofilter conservation strategy to complement fine and coarse filters. Conserv. Biol. 19, 1025–1029. doi: 10.1111/j.1523-1739.2005.00172.x
Imbeau, L., Mönkkönen, M., and Desrochers, A. (2001). Long-term effects of forestry on birds of the eastern Canadian boreal forest: A comparison with Fennoscandia. Conserv. Biol. 15, 1151–1162. doi: 10.1046/j.1523-1739.2001.0150041151.x
Kneeshaw, D., and Bergeron, Y. (1998). Canopy gap characteristics and tree replacement in the southeastern boreal forest. Ecology 79, 783–794. doi: 10.1890/0012-9658(1998)079[0783:CGCATR]2.0.CO;2
Kneeshaw, D., and Gauthier, S. (2003). Old-growth in the boreal: A dynamic perspective. Environ. Rev. 11(Suppl. 1) S99–S114. doi: 10.1139/a03-010
Larrieu, L., Cabanettes, A., Gouix, N., Burnel, L., Bouget, C., and Deconchat, M. (2017). Development over time of the tree-related microhabitat profile: The case of lowland beech–oak coppice-with-standards set-aside stands in France. Eur. J. For. Res. 136, 37–49. doi: 10.1007/s10342-016-1006-3
Larrieu, L., Paillet, Y., Winter, S., Bütler, R., Kraus, D., Krumm, F., et al. (2018). Tree related microhabitats in temperate and Mediterranean European forests: A hierarchical typology for inventory standardization. Ecol. Indic. 84, 194–207. doi: 10.1016/j.ecolind.2017.08.051
Lavertu, D., Mauffette, Y., and Bergeron, Y. (1994). Suckering success of aspen (Populus tremuloides Michx.) in relation to stand age and soil disturbance. J. Veg. Sci. 5, 561–568. doi: 10.2307/3235983
Li, P., and Martin, T. E. (1991). Nest-site selection and nesting success of cavity-nesting birds in high elevation forest drainages. Auk 108, 405–418.
Lindenmayer, D. B., Cunningham, R. B., Donnelly, C. F., and Franklin, J. F. (2000). Structural features of old-growth Australian montane ash forests. For. Ecol. Manag. 134, 189–204. doi: 10.1016/S0378-1127(99)00257-1
Martin, K., Aitken, K. E. H., and Wiebe, K. L. (2004). Nest sites and nest webs for cavity-nesting communities in interior British Colombia, Canada: Nest characteristics and niche partitioning. Condor. 106, 5–19. doi: 10.1093/condor/106.1.5
Martin, K., and Eadie, J. M. (1999). Nest webs: A community-wide approach to the management and conservation of cavity-nesting forest birds. For. Ecol. Manag. 115, 243–257. doi: 10.1016/S0378-1127(98)00403-4
Martin, M., Fenton, N., and Morin, H. (2018). Structural diversity and dynamics of boreal old-growth forests case study in Eastern Canada. For. Ecol. Manag. 422, 125–136. doi: 10.1016/j.foreco.2018.04.007
Martin, M., Grondin, P., Lambert, M. C., Bergeron, Y., and Morin, H. (2021). Compared to wildfire, management practices reduced old-growth forest diversity and functionality in primary boreal landscapes of eastern Canada. Front. For. Glob. Change 4:639397. doi: 10.3389/ffgc.2021.639397
Martin, M., Paillet, Y., Larrieu, L., Kern, C., Raymond, P., Drapeau, P., et al. (2022). Tree-related microhabitats are promising yet underused tools for biodiversity and nature conservation: A systematic review for international perspectives. Front. For. Glob. Change 5:818474. doi: 10.3389/ffgc.2022.818474
Messier, C., and Puettmann, K. J. (2011). Forests as complex adaptive systems: Implications for forest management and modelling. Ital. J. For. Environ. 66, 249–258. doi: 10.1097/HMR.0b013e3182860937
Ministère des Ressources naturelles et de la Faune du Québec (2006). mos_06_32d11, mos_06_32d06 [orthophotos numérisées]. Échelle 1 : 15 000, noir & blanc, 8 bits, format MrSID, 5.43 GB & 5.45 GB. Québec, QC: Gouvernement du Québec.
Morin, H., Laprise, D., and Bergeron, Y. (1993). Chronology of spruce budworm outbreaks near Lake Duparquet, Abitibi region, Quebec. Can. J. For. Res. 23, 1497–1506. doi: 10.1139/x93-189
Nappi, A. (2009). Utilisation du bois mort et des forêts brûlées par le Pic à dos noir en forêt boréale. Ph.D. thesis. Montréal, QC: Université du Québec à Montréal.
Nappi, A., Drapeau, P., and Leduc, A. (2015). How important is dead wood for woodpeckers foraging in eastern North American boreal forests? For. Ecol. Manag. 346, 10–21. doi: 10.1016/j.foreco.2015.02.028
Newton, I. (1994). The role of nest sites in limiting the numbers of hole-nesting birds: A review. Biol. Conserv. 70, 265–276. doi: 10.1016/0006-3207(94)90172-4
Ouellet-Lapointe, U., Drapeau, P., Cadieux, P., and Imbeau, L. (2012). Woodpecker excavations suitability for and occupancy by cavity users in the boreal mixedwood forest of eastern Canada. Ecoscience 19, 391–397. doi: 10.2980/19-4-3582
Paré, D., and Bergeron, Y. (1995). Above-ground biomass accumulation along a 230-year chronosequence in the southern portion of the Canadian boreal forest. J. Ecol. 83, 1001–1007. doi: 10.2307/2261181
Politi, N., Hunter, M. Jr., and Rivera, L. (2010). Availability of cavities for avian cavity nesters in selectively logged subtropical montane forests of the Andes. For. Ecol. Manag. 260, 893–906. doi: 10.1016/j.foreco.2010.06.009
R Core Team (2015). R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing.
Remm, J., and Lõhmus, A. (2011). Tree cavities in forests – the broad distribution pattern of a keystone structure for biodiversity. For. Ecol. Manag. 262, 579–585. doi: 10.1016/j.foreco.2011.04.028
Robitaille, A., and Saucier, J.-P. (1998). Paysages régionaux du québec méridional. Québec, QC: Les Publications Du Québec.
Saint-Germain, M., Drapeau, P., and Buddle, C. M. (2007). Host-use patterns of saproxylic phloeophagous and xylophagous Coleoptera adults and larvae along the decay gradient in standing dead black spruce and aspen. Ecography 30, 737–748. doi: 10.1111/j.2007.0906-7590.05080.x
Simard, S., Martin, K., Vyse, A., Larson, B., Messier, C., Puettmann, K., et al. (2013). “Meta-networks of fungi, fauna and flora as agents of complex adaptive systems,” in Managing forests as complex adaptive systems: Building resilience to the challenge of global change, eds C. Messier, K. J. Puettmann, and K. D. Coates (New York, NY: Routledge), 133–164.
Spies, T. A., Ripple, W. J., and Bradshaw, G. A. (1994). Dynamics and pattern of a managed coniferous forest landscape in Oregon. Ecol. Appl. 4, 555–568. doi: 10.2307/1941957
Stokland, J. N., Siitonen, J., and Jonsson, B. G. (2012). Biodiversity in dead wood. New York, NY: Cambridge University Press. doi: 10.1017/CBO9781139025843
Tews, J., Brose, U., Grimm, V., Tielbörger, K., Wichmann, M. C., Schawger, M., et al. (2004). Animal species diversity driven by habitat heterogeneity/diversity: The importance of keystone structures. J. Biogeogr. 31, 79–92. doi: 10.1046/j.0305-0270.2003.00994.x
Trudeau, C., Imbeau, L., Drapeau, P., and Mazerolle, M. J. (2011). Site occupancy and cavity use by the northern flying squirrel in the boreal forest. J. Wild. Manag. 75, 1646–1656.
Wesolowski, T., and Martin, K. (2018). “Tree holes and hole nesting birds in European and North American forests,” in Ecology and conservation of forest birds, Chap. 4, eds G. Mikusiński, J. Roberge, and R. Fuller (Cambridge: Cambridge University Press), 79–134. doi: 10.1017/9781139680363
Wiebe, K. L., and Moore, W. S. (2020). “Northern Flicker (Colaptes auratus), version 1.0,” in Birds of the world, ed. P. G. Rodewald (Ithaca, NY: Cornell Lab of Ornithology). doi: 10.2173/bow.norfli.01
Keywords: boreal mixedwood forests, old forest stands, cavity-using vertebrate communities, nest webs, ecological networks complexity, resilience and stability of cavity-using vertebrate communities, natural forests structural development
Citation: Cadieux P, Drapeau P, Ouellet-Lapointe U, Leduc A, Imbeau L, Deschênes R and Nappi A (2023) Old forest structural development drives complexity of nest webs in a naturally disturbed boreal mixedwood forest landscape. Front. For. Glob. Change 6:1084696. doi: 10.3389/ffgc.2023.1084696
Received: 30 October 2022; Accepted: 12 January 2023;
Published: 27 January 2023.
Edited by:
Andre Arsenault, Natural Resources Canada, Canadian Forest Service, Atlantic Forestry Centre, CanadaReviewed by:
Manuela Branco, University of Lisbon, PortugalAlan Feest, University of Bristol, United Kingdom
Copyright © 2023 Cadieux, Drapeau, Ouellet-Lapointe, Leduc, Imbeau, Deschênes and Nappi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pierre Drapeau,  ZHJhcGVhdS5waWVycmVAdXFhbS5jYQ==
ZHJhcGVhdS5waWVycmVAdXFhbS5jYQ==