 Vera Manageiro1,2
Vera Manageiro1,2 Raquel Romão1
Raquel Romão1 Inês Barata Moura1
Inês Barata Moura1 Daniel A. Sampaio3Luís Vieira3Eugénia Ferreira1,2the Network EuSCAPE-Portugal
Daniel A. Sampaio3Luís Vieira3Eugénia Ferreira1,2the Network EuSCAPE-Portugal Manuela Caniça1,2*
Manuela Caniça1,2*- 1National Reference Laboratory of Antibiotic Resistances and Healthcare Associated Infections, Department of Infectious Diseases, National Institute of Health Dr. Ricardo Jorge, Lisbon, Portugal
- 2Centre for the Studies of Animal Science, Institute of Agrarian and Agri-Food Sciences and Technologies, University of Oporto, Oporto, Portugal
- 3Innovation and Technology Unit, Department of Human Genetics, National Institute of Health Dr. Ricardo Jorge, Lisbon, Portugal
In Portugal, the epidemiological stage for the spread of carbapenemase-producing Enterobacteriaceae (CPE) increased from sporadic isolates or single hospital clones (2010–2013), to hospital outbreaks, later. Here we report data from a 6-month study performed under the European Survey on Carbapenemase-Producing Enterobacteriaceae (EuSCAPE). During the study period, 67 isolates (61 Klebsiella pneumoniae and 6 Escherichia coli) non-susceptible to carbapenems were identified in participant hospital laboratories. We detected 37 blaKPC–type (including one new variant: blaKPC–21), 1 blaGES–5, and 1 blaGES–6 plus blaKPC–3, alone or in combination with other bla genes. Bioinformatics analysis of the KPC-21-producing E. coli identified the new variant blaKPC–21 in a 12,748 bp length plasmid. The blaKPC–21 gene was harbored on a non-Tn4401 element, presenting upstream a partial ISKpn6 (ΔISKpn6/ΔtraN) with the related left IR (IRL) and downstream a truncated Tn3 transposon. PFGE and MLST analysis showed an important diversity, as isolates belonged to distinct PFGE and STs profiles. In this study, we highlighted the presence of the high-risk clone E. coli sequence-type (ST) 131 clade C/H30. This worldwide disseminated E. coli lineage was already detected in Portugal among other antibiotic resistance reservoirs. This study highlights the intra- and inter-hospital spread and possible intercontinental circulation of CPE isolates.
Introduction
Carbapenems, a class of β-lactam antibiotics with wide activity, are often the antimicrobials of last resort to treat infections associated to extended-spectrum β-lactamase (ESBL)- or plasmid-mediated AmpC (PMAβ)-producing Enterobacteriaceae isolates (Papp-Wallace et al., 2011; Rodríguez-Baño et al., 2018). Unfortunately, carbapenem non-susceptible Enterobacteriaceae (CNSE) have been reported worldwide mainly because of the acquisition of carbapenemase-encoding genes (Potter et al., 2016; Codjoe and Donkor, 2018). Since the first description of a carbapenemase-producing Enterobacteriaceae (CPE) in Europe in the 1990s, a large variety of carbapenemases has been identified in each of the four Ambler molecular classes, mainly the KPC-type (class A), VIM-, IMP-, and NDM-types (class B), and OXA-48-type (class D) (Grundmann et al., 2017; Logan and Weinstein, 2017). CPE isolates are usually resistant to many other β-lactam and non-β-lactam antibiotics, leading to multi-resistant isolates.
In Portugal, the epidemiological stage for the spread of CPE increased from sporadic isolates or single hospital clones, from April 2010 to February 2013, to sporadic hospital outbreaks later (Albiger et al., 2015; Manageiro et al., 2015b,c). Here we report data from a 6-month prevalence study performed under the European Survey on Carbapenemase-Producing Enterobacteriaceae (EuSCAPE) with the collaboration of different Portuguese Laboratories.
Materials and Methods
Bacterial Isolation, Antibiotic Susceptibility, and Molecular Characterization
This study included a total of 104 clinical isolates (94 Klebsiella pneumoniae and 10 Escherichia coli) collected from November 2013 to April 2014 in 10 Portuguese hospitals. The first ten consecutive and non-replicated CNSE isolates obtained during this period, in each hospital, from blood, lower respiratory tract secretions, urine, puncture fluids, and wound secretions, of single patients, were sent to the National Reference Laboratory, in Lisbon, and were considered. Successive carbapenem-susceptible isolates of the same species were also preserved as controls whenever possible, accordingly to EuSCAPE protocol (Grundmann et al., 2017). Overall, 67 CNSE (61 K. pneumoniae and 6 E. coli) and 37 controls (33 K. pneumoniae and 4 E. coli) were analyzed.
In the context of the EuSCAPE study, all data were anonymized and collected in accordance with the European Parliament and Council decisions on the epidemiological surveillance and control of communicable disease in the European Community (Eur-Lex-31998D2119, 1998; Eur-Lex-32000D0096, 2000).
Antibiotic Susceptibility and Molecular Characterization of Antimicrobial Resistance
Antimicrobial susceptibility was performed by disk diffusion method for 15 antibiotics (Table 1), and by broth microdilution method for tigecycline and colistin, using EUCAST guidelines1. Clinical isolates with resistance or with decreased susceptibility to ertapenem were considered presumptively CPE. Isolates were considered multidrug resistant when presenting reduced susceptibility to three or more structurally unrelated antibiotics.
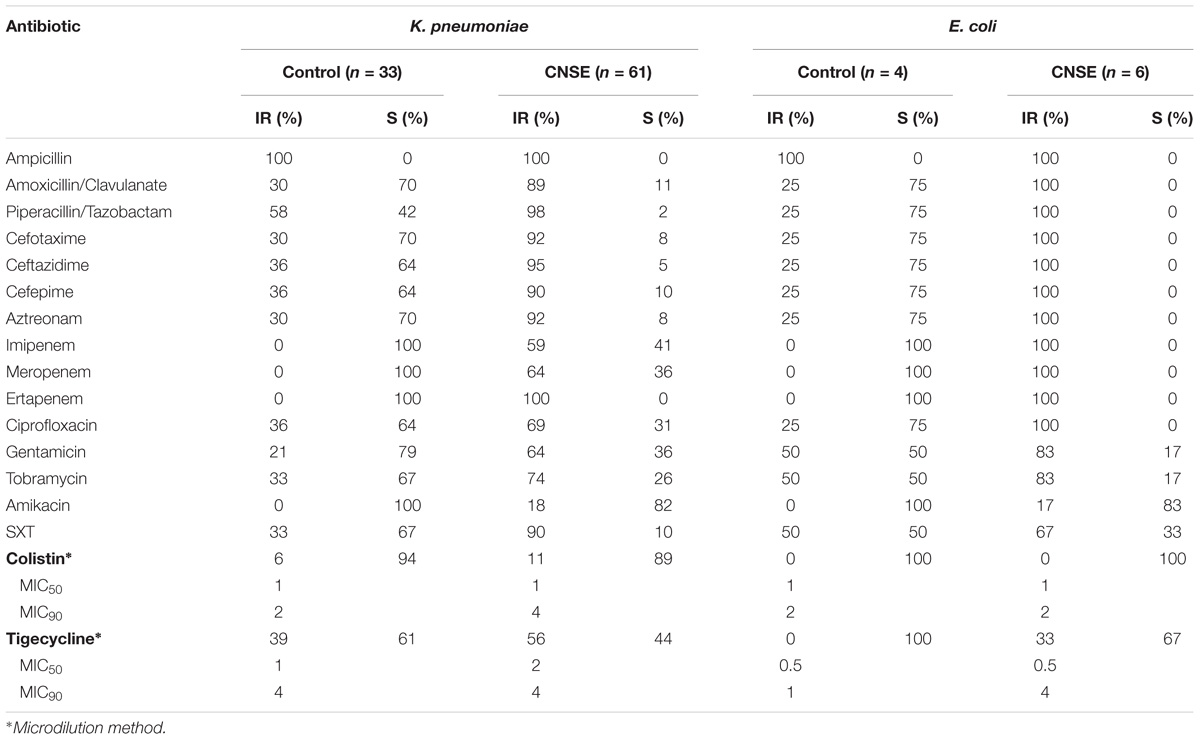
TABLE 1. Antimicrobial susceptibility of 67 (61 K. pneumoniae and 6 E. coli) CNSE and 37 (33 K. pneumoniae and 4 E. coli) control isolates.
PCR and sequencing were applied to detect and identify the main CPE (blaKPC and blaGES from class A; blaIMP, blaV IM, and blaNDM from class B; and blaOXA–48 from class D)-, ESBL (blaTEM, blaSHV, blaOXA, blaCTX–M) – and PMAβ (blaCMY, blaMOX, blaFOX, blaLAT, blaACT, blaMIR, blaDHA, blaMOR, blaACC)-encoding genes, as previously described (Manageiro et al., 2015b). Plasmid-mediated colistin resistance-encoding genes (mcr-type) were also investigated (Manageiro et al., 2017).
Transfer Experiments
Transferability of blaKPC–21 from E. coli UR19829 was performed by broth mating out assays using sodium azide-resistant E. coli J53 as a recipient strain, and by transformation, as previously described (Manageiro et al., 2015b, 2017).
Molecular Typing
Clonal relatedness of 67 CNSE isolates was investigated by pulsed-field gel electrophoresis (PFGE) as previously described (Manageiro et al., 2017). Genetic diversity of the K. pneumoniae (n = 10, i.e., 1 representative of each PFGE cluster) and E. coli (n = 10) isolates was investigated by multilocus sequence typing (MLST) (Manageiro et al., 2015b). E. coli sequence type (ST) subclones were also analyzed on the basis of the E. coli fimH gene (Manageiro et al., 2015a).
Genomic Characterization of KPC-21-Producing E. coli
KPC-21-producing E. coli was genotypically characterized by whole-genome sequencing (WGS) as previously described (Manageiro et al., 2017). The assembled contigs were analyzed and studied for the presence of antibiotic resistance- and virulence-encoding genes, multi-locus sequence types, fim type, serotype, plasmid replicon types, and insertion sequences (ISs) using bioinformatics tools from the Center for Genomic Epidemiology2 and ISsaga (Varani et al., 2011).
The pUR19829-KPC21 plasmid structure was constructed based on the genetic organization of the closest plasmid sequences obtained by BLASTn, provided by NCBI3, followed by contig neighbor’s prediction from assembly information.
Statistical Analysis
OpenEpi software, version 3.01 was used for statistical analysis (Sullivan et al., 2009). Fisher exact test was used to assess differences in clinical and epidemiological risk factors for control and CNSE-carrying patients. One-tailed P values of ≤0.05 were considered to be statistically significant. Associations were determined by calculation of odds ratios with 95% confidence intervals.
Nucleotide Sequence Accession Number
The new blaKPC–21 nucleotide sequence was submitted to the NCBI GenBank Database with accession number NG_049254 and the complete plasmid sequence of pUR19829-KPC21 with accession number MH133192.
Results and Discussion
During the study period, 67 isolates (61 K. pneumoniae and 6 E. coli) CNSE were identified in nine of the 10 Hospital Laboratories, with a non-susceptibility rate for meropenem and imipenem of 64 and 59%, respectively, for K. pneumoniae, and of 100% for E. coli. As expected, when compared with the control isolates, CNSE presented higher level of non-susceptibility to all antibiotic classes tested (Table 1). Colistin and tigecycline MIC50 values for CNSE were similar than those obtained for control isolates. Eleven out of the 104 (16.3%) isolates were colistin resistant, without the presence of the plasmid-mediated mcr-1 or mcr-2 gene. However, MCR-1 determinant was already identified in different reservoirs in Portugal, such as vegetables, animals and humans (Jones-Dias et al., 2016; Beyrouthy et al., 2017; Kieffer et al., 2017).
Thirty-eight (56.7%) isolates (36 K. pneumoniae, 2 E. coli) were confirmed to be CPE; we identified 36 blaKPC–type (including one new variant: blaKPC–21), 1 blaGES–5, and 1 blaGES–6 plus blaKPC–3, alone or in combination with other bla genes (Supplementary Figure S1). The remaining 29 isolates were non-susceptible to carbapenems possibly due to porins deficiency with association of PMAβ (CMY-2 and DHA-1) and/or ESBL (mainly CTX-M-15) production (Martínez-Martínez, 2008).
The new blaKPC–21 gene differed from blaKPC–2 by one point mutation that leads to the amino acid substitution Trp105Arg; this position is involved in the binding and maintaining of the KPC catalytic activity (Papp-Wallace et al., 2010). In silico typing revealed an KPC-21-producing E. coli belonging to ST131 clade C/H30, associated with the fimbriae-encoding fimH allele 30, which become the most dominant lineage since the 2000s (Nicolas-Chanoine et al., 2014; Pitout and DeVinney, 2017). Moreover, bioinformatics analysis of the KPC-21-producing E. coli identified this variant in a 12,748 bp length plasmid, with a mean coverage of 580-fold and GC content of 58.5% (Figure 1).
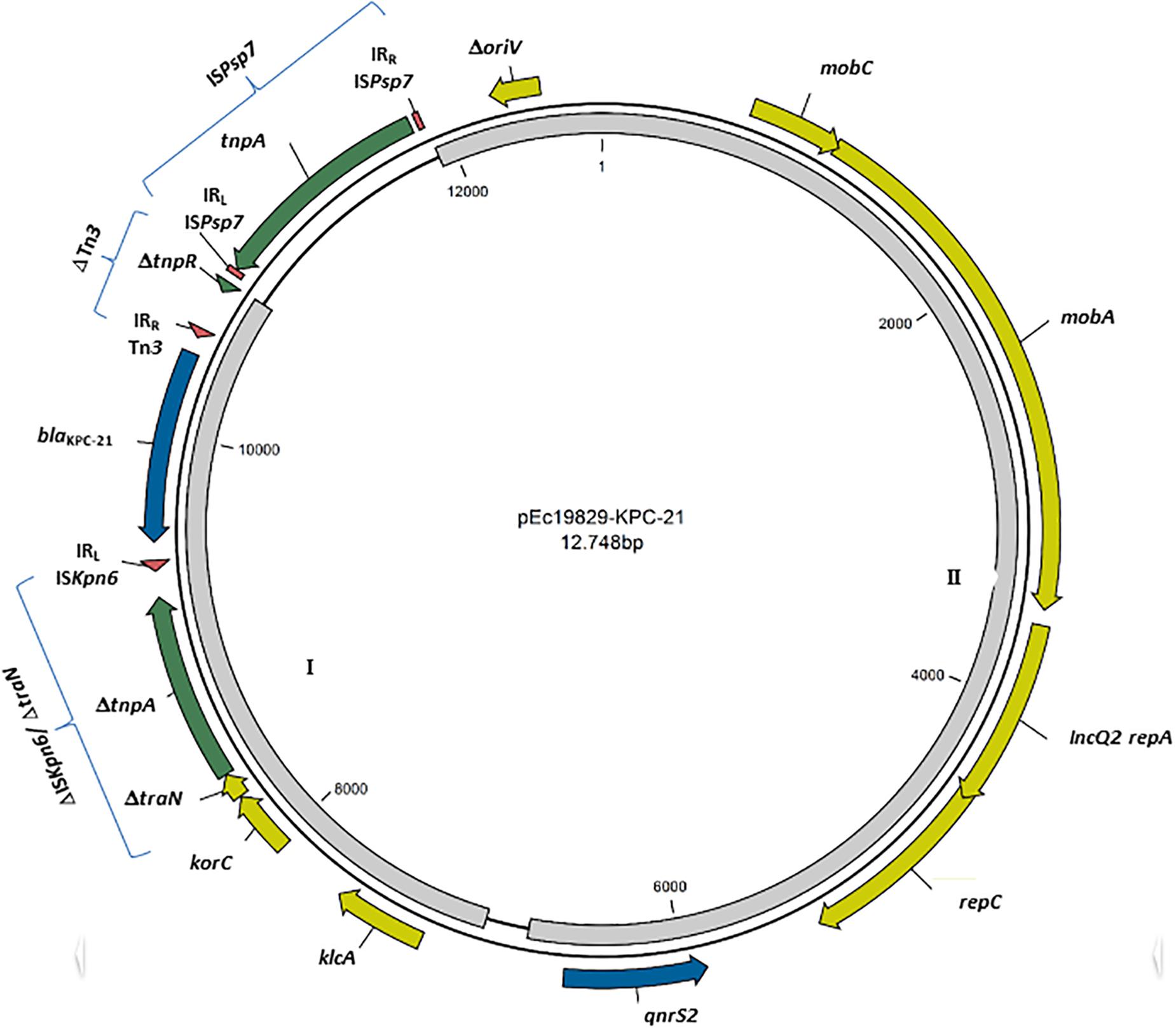
FIGURE 1. Schematic representation of KPC-21-harboring plasmid (pUR19829-KPC21). Genes are denoted by arrows. Blue, antibiotic resistance genes; Green, mobile genetic elements; Yellow, plasmid mobilization genes and replication origin. Right and left inverted repeats (IRR and IRL) are indicated as red triangles. Gray regions 1: 99.97% of identity with KX756453, and Gray region 2: >99.9% sequence identity to KT896499.
Dissemination of blaKPC has been mainly supported by the horizontal transfer of Tn4401-type transposon, which harbors tnpA encoding a transposase, tnpR encoding resolvase, and two insertion sequence elements (ISKpn7 and ISKpn6) bracketing the blaKPC gene (Cuzon et al., 2011). In this study, the blaKPC–21 gene was harbored on a non-Tn4401 element (Chen et al., 2014), presenting upstream a partial ISKpn6 (ΔISKpn6/ΔtraN) with the related left IR (IRL) and downstream truncated Tn3 transposon downstream (Figure 1). This region has 99.97% of identity with pKP1194a, a plasmid carried by a hospital-associated KPC-2-producing K. pneumoniae isolated in Brazil (Accession number KX756453) (Figure 1- gray region I); this suggest an intercontinental circulation of isolates and mobile genetic elements (MGE), and the consequent need of concerted actions against the spreading of antibiotic resistance, at a worldwide level. The pUR19829-KPC21 enclosed also an intact ISPsp7 element, an insertion sequence from IS30 family, firstly described in Pseudomonas spp. (Szuplewska et al., 2014). Furthermore, the pUR19829-KPC21 backbone contained a region coding for plasmid replication (IncQ2 repA, repC), and mobilization (mobA, mobC), showing >99.9% sequence identity to the corresponding regions of pKPSH169, a 7.7 Kbp qnrS2-harboring IncQ plasmid identified in municipal wastewater treatment facilities in Israel (Accession number KT896499) (Figure 1- gray region II); this similarity highlights the high level of promiscuity of isolates between clinical settings and environment, where both reservoirs play a role in the antibiotic resistance dissemination (Stokes and Gillings, 2011). However, the lack of conjugative elements or an oriT region, associated with the presence of a truncated oriV region (Figure 1) suggests that pUR19829-KPC21 plasmid is nonmobilizable (Smillie et al., 2010). This fact is corroborated by the absence of a successful plasmid conjugation or transformation.
The variables used in the evaluation of risk factors for infection or colonization of patients with CNSE or control isolates are present in Table 2. When compared to the 37 control strains, only ESBL-production and the patient admission at a hospital in the center of Portugal were significantly associated with CNSE isolates in the period of the study. In the era of ESBL-producing Enterobacteriaceae, the antibiotic regimens suggested for severe health-associated infections are necessarily based on carbapenems (Rodríguez-Baño et al., 2018). Unfortunately carbapenem use has being described as a risk factor for CPE acquisition, only preceded by the use of medical devices (van Loon et al., 2018). In addition, the present study attests that Portugal, during the period of the study, has a different CNSE geographical distribution with the center of Portugal significantly associated with carbapenem non-susceptibility. This fact corroborates previous studies which indicated that in Portugal, in 2015, only sporadic isolates or single hospital cases were described (Albiger et al., 2015).
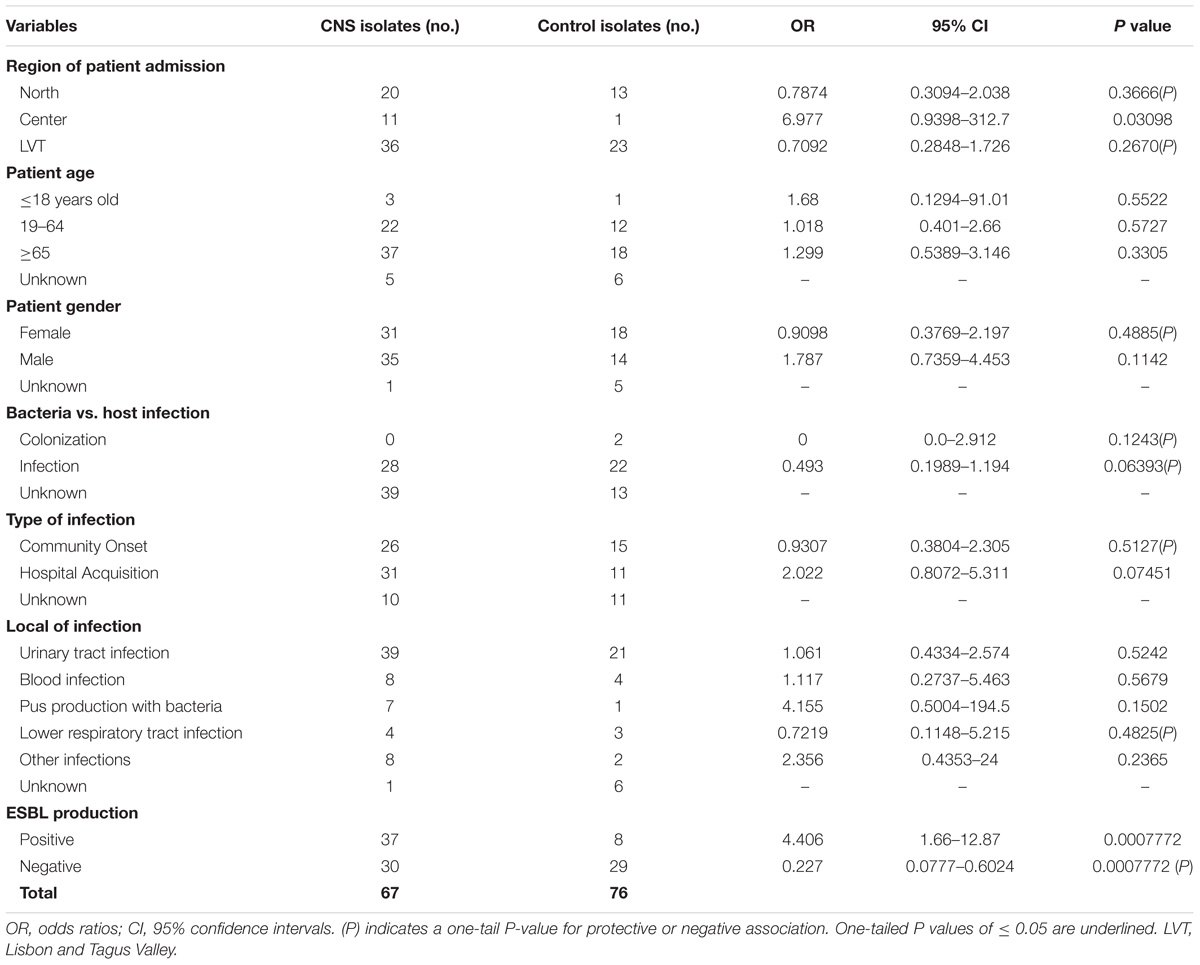
TABLE 2. Evaluation of risk factors for patients with infections caused by carbapenem susceptible or CNSE bacteria.
PFGE and MLST analysis showed an important diversity, with isolates belonging to distinct PFGE and STs (Supplementary Figure S1). With respect to K. pneumoniae (Supplementary Figure S1A), a total of 10 clusters and 25 unique PFGE profiles were generated using XbaI, indicating the that the circulating clones in that period were genetically diverse. However, carbapenemase-producing K. pneumoniae isolates were more clonal (six PFGE clusters including 69.4% of these isolates) than non-carbapenemase-producing K. pneumoniae (four PFGE clusters including 50.0% of these isolates). As shown in Supplementary Figure S1, both CNSE species showed intra- and inter-hospital spread (e.g., PFGE clusters KpI and KpIX), with some hospital-specific clones (e.g., PFGE clusters KpIV and KpVIII). However, as also showed in Spain in other EuSCAPE study (Esteban-Cantos et al., 2017), the carbapenem-non-susceptible K. pneumoniae population was more clonal than the carbapenem-susceptible population (data not shown). Ten different MLSTs were detected among carbapenemase-producing (ST14, ST15, ST45, ST231, and ST1513) and non-carbapenemase-producing (ST11, ST17, ST348, and ST395) K. pneumoniae isolates. At our knowledge, this is the first description of ST17, ST395, and ST1513 K. pneumoniae in Portugal (Manageiro et al., 2015b; Rodrigues et al., 2016; Vubil et al., 2017). Noteworthy, the GES-5 enzyme was detected in a ST231 K. pneumoniae isolate as previously reported in Portugal, but in the same hospital, which shows its capacity to maintain in clinical settings due to the selection pressure of this environment (Manageiro et al., 2015b). Furthermore, ST45 was recently the cause of a hospital-based outbreak caused by multidrug-resistant, KPC-3- and MCR-1-producing K. pneumoniae in Portugal (Mendes et al., 2018).
The high-risk clone carbapenemase-positive K. pneumoniae ST258 was not detected in this study or among clinical carbapenemase-producing K. pneumoniae isolates in Portugal (Manageiro et al., 2015b; Rodrigues et al., 2016; Vubil et al., 2017). However, concerning carbapenem-non-susceptible E. coli, besides the six different PFGE unique profiles, the isolates belongs all but two (ST405-fimH27 and ST23-fimH35) to the ST131 clade C/H30 high-risk clone disseminated worldwide (Supplementary Figure S1B) (Woodford et al., 2011; Pitout and DeVinney, 2017). Noteworthy, this clone was already detected in Portugal among other antibiotic resistance reservoirs, such as in an E. coli strain isolated from a dolphin housed at a Zoo Park (Manageiro et al., 2015a); in dogs and cats with urinary tract infection (Marques et al., 2018); and in E. coli strains from wastewater and gulls (Varela et al., 2015). Again, this shows that clinical settings and different environmental compartments may be considered communicating vessels through which bacteria and resistance genes are able to flow (Stokes and Gillings, 2011).
Portugal was one of the EuSCAPE participating countries that presented higher proportions of KPC-positive K. pneumoniae (Grundmann et al., 2017). The percentage of carbapenem non-susceptible K. pneumoniae was low in invasive infections in the study period [2.4%, EARS-Net 2013]4. However, although the consumption of carbapenems has declined by 13.3% between 2012 and 2016 (PPCIRA, 2017), Portugal is reporting since 2013 a significant increasing trend of carbapenem non-susceptible K. pneumoniae [6.4%, EARS-Net 2016]4. The number of inter-institutional transmission is also increasing (Glasner et al., 2013; Albiger et al., 2015), being K. pneumoniae the principal cause of bacterial health-associated infections in Portugal, as in other European countries (ECDC, 2013). Of concern is the fact that KPC-producing organisms cause infections with high morbidity and mortality (Porreca et al., 2018; Rodríguez-Baño et al., 2018). These results reinforces that reducing antibiotic use alone is likely insufficient for reversing resistance (Lopatkin et al., 2017). We strongly believe that the chain of transmission of isolates and genes in clinical settings will be reduced or broken, especially with containment measures rigorously implemented and followed at local level.
Members of the Network EuScape-Portugal
North region: C.H. São João E.P.E. (J.T. Guimarães), H. Braga (C. Iglesias), C.H. Póvoa de Varzim e Vila do Conde E.P.E. (F. Fonseca). Centre region: C.H.U. de Coimbra/Covões (H. Oliveira), C.H.U. de Coimbra/HUC (L. Boaventura), C.H. Médio Tejo E.P.E. (Ana Soares). Lisbon and Tagus Valley region: C.H. Oeste Norte E.P.E. (A. Vicente). H. Garcia de Orta E.P.E. (J. Diogo), C.H. Lisboa Ocidental E.P.E. (E. Gonçalves), C.H. Lisboa Central E.P.E. (M. Pinto).
Author Contributions
VM designed the study, performed the molecular experiments, bioinformatics analysis, analyzed the data, and wrote the manuscript. RR, IBM, and EF performed the microbiological and molecular experiments, and analyzed the data. DAS and LV performed Illumina genome sequencing experiments. The Network EuSCAPE-Portugal participants acquired laboratory data. MC designed the study, wrote and reviewed the manuscript. All authors read and approved the final manuscript.
Funding
VM was supported by Fundação para a Ciência e a Tecnologia (FCT) fellowship (Grant SFRH/BPD/ 77486/2011), financed by the European Social Funds (COMPETE-FEDER) and national funds of the Portuguese Ministry of Education and Science (POPH-QREN). The authors thank FCT for Project grant UID/MULTI/00211/2013.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to thank Rafael Graça for his technical assistance.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.02834/full#supplementary-material
FIGURE S1 | Pulsed-field gel electrophoresis (PFGE) dendrogram and genetic relatedness of 61 K. pneumoniae (A) and 6 E. coli (B) CNSE isolates. Isolate number, hospital code, year of isolation, carbapenems antibiotic susceptibility, detected carbapenemases, extended-spectrum β-lactamases (ESBL), inhibitor resistant SHV (IRS), and plasmid-mediated AmpC (PMAβ), Multilocus sequence typing (MLST) for selected isolates and PFGE profile types are shown. These profiles, from 0001 to 0035, were defined as forming clusters KpI to KpX, for K. pneumoniae, and from 0001 to 0006 for E. coli. For E. coli isolates, fim-type is also shown.
Footnotes
- ^ http://www.eucast.org/clinical_breakpoints/
- ^ https://cge.cbs.dtu.dk/services/
- ^ http://blast.ncbi.nlm.nih.gov/Blast.cgi
- ^ https://ecdc.europa.eu/
References
Albiger, B., Glasner, C., Struelens, M. J., Grundmann, H., Monnet, D. L., and The European Survey of Carbapenemase-Producing Enterobacteriaceae (EuSCAPE) Working Group (2015). Carbapenemase-producing Enterobacteriaceae in Europe: assessment by national experts from 38 countries, May 2015. Euro Surveill. 20:30062. doi: 10.2807/1560-7917.ES.2015.20.45.30062
Beyrouthy, R., Robin, F., Lessene, A., Lacombat, I., Dortet, L., Naas, T., et al. (2017). MCR-1 and OXA-48 in vivo acquisition in KPC-producing Escherichia coli after colistin treatment. Antimicrob. Agents Chemother. 61:e02540-16. doi: 10.1128/AAC.02540-16
Chen, L., Mathema, B., Chavda, K. D., DeLeo, F. R., Bonomo, R. A., and Kreiswirth, B. N. (2014). Carbapenemase-producing Klebsiella pneumoniae: molecular and genetic decoding. Trends Microbiol. 22, 686–696. doi: 10.1016/j.tim.2014.09.003
Cuzon, G., Naas, T., and Nordmann, P. (2011). Functional characterization of Tn4401, a Tn3-based transposon involved in blaKPC gene mobilization. Antimicrob. Agents Chemother. 55, 5370–5373. doi: 10.1128/AAC.05202-11
ECDC (2013). Point Prevalence Survey of Healthcare Associated Infections and Antimicrobial use in European Acute Care Hospitals, 2011-2012. Stockholm: European Centre for Disease Prevention and Control.
Esteban-Cantos, A., Aracil, B., Bautista, V., Ortega, A., Lara, N., Saez, D., et al. (2017). The carbapenemase-producing Klebsiella pneumoniae population is distinct and more clonal than the carbapenem-susceptible population. Antimicrob. Agents Chemother. 61:e02520-16. doi: 10.1128/AAC.02520-16
Eur-Lex-31998D2119 (1998). Decision Number 2119/98/EC of the European Parliament and of the Council of 24 September 1998: Setting up a Network for the Epidemiological Surveillance and Control of Communicable Diseases in the Community. Available at: http://data.europa.eu/eli/dec/1998/2119/oj
Eur-Lex-32000D0096 (2000). Commission Decision Number 2000/96/EC of 22 December 1999 on the Communicable Diseases to be Progressively Covered by the Community Network Under Decision No 2119/98/EC of the European Parliament and of the Council. Available at: https://eur-lex.europa.eu/eli/dec/2000/96(1)/oj
Glasner, C., Albiger, B., Buist, G., Tambiæ Andraseviæ, A., Canton, R., Carmeli, Y., et al. (2013). Carbapenemase-producing Enterobacteriaceae in Europe: a survey among national experts from 39 countries, February 2013. Euro Surveill. 18:20525. doi: 10.2807/1560-7917.ES2013.18.28.20525
Grundmann, H., Glasner, C., Albiger, B., Aanensen, D. M., Tomlinson, C. T., Andraseviæ, A. T., et al. (2017). Occurrence of carbapenemase-producing Klebsiella pneumoniae and Escherichia coli in the European survey of carbapenemase-producing Enterobacteriaceae (EuSCAPE): a prospective, multinational study. Lancet Infect. Dis. 17, 153–163. doi: 10.1016/S1473-3099(16)30257-2
Jones-Dias, D., Manageiro, V., Ferreira, E., Barreiro, P., Vieira, L., Moura, I. B., et al. (2016). Architecture of class 1,2, and 3 integrons from Gram-negative bacteria recovered among fruits and vegetables. Front. Microbiol. 7:1400. doi: 10.3389/fmicb.2016.01400
Kieffer, N., Aires-de-Sousa, M., Nordmann, P., and Poirel, L. (2017). High rate of MCR-1-producing Escherichia coli and Klebsiella pneumoniae among pigs, Portugal. Emerg. Infect. Dis. 23, 2023–2029. doi: 10.3201/eid2312.170883
Logan, L. K., and Weinstein, R. A. (2017). The epidemiology of carbapenem-resistant Enterobacteriaceae: the impact and evolution of a global menace. J. Infect. Dis. 215, S28–S36. doi: 10.1093/infdis/jiw282
Lopatkin, A. J., Meredith, H. R., Srimani, J. K., Pfeiffer, C., Durrett, R., and You, L. (2017). Persistence and reversal of plasmid-mediated antibiotic resistance. Nat. Commun. 8:1689. doi: 10.1038/s41467-017-01532-1
Manageiro, V., Clemente, L., Graça, R., Correia, I., Albuquerque, T., Ferreira, E., et al. (2017). New insights into resistance to colistin and third-generation cephalosporins of Escherichia coli in poultry, Portugal: novel blaCTX-M-166 and blaESAC genes. Int. J. Food Microbiol. 263, 67–73. doi: 10.1016/j.ijfoodmicro.2017.10.007
Manageiro, V., Clemente, L., Jones-Dias, D., Albuquerque, T., Ferreira, E., and Canica, M. (2015a). CTX-M-15-producing Escherichia coli in Dolphin, Portugal. Emerg. Infect. Dis. 21, 2249–2251. doi: 10.3201/eid2112.141963
Manageiro, V., Ferreira, E., Almeida, J., Barbosa, S., Simões, C., Bonomo, R. A., et al. (2015b). Predominance of KPC-3 in a survey for carbapenemase-producing Enterobacteriaceae in Portugal. Antimicrob. Agents Chemother. 59, 3588–3592. doi: 10.1128/AAC.05065-14
Manageiro, V., Sampaio, D. A., Pereira, P., Rodrigues, P., Vieira, L., Palos, C., et al. (2015c). Draft genome sequence of the first NDM-1-producing Providencia stuartii isolated in Portugal. Genome Announc. 3:e01077-15. doi: 10.1128/genomeA.01077-15
Marques, C., Belas, A., Franco, A., Aboim, C., Gama, L. T., and Pomba, C. (2018). Increase in antimicrobial resistance and emergence of major international high-risk clonal lineages in dogs and cats with urinary tract infection: 16 year retrospective study. J. Antimicrob. Chemother. 73, 377–384. doi: 10.1093/jac/dkx401
Martínez-Martínez, L. (2008). Extended-spectrum beta-lactamases and the permeability barrier. Clin. Microbiol. Infect. 14(Suppl. 1), 82–89. doi: 10.1111/j.1469-0691.2007.01860.x
Mendes, A. C., Novais, Â, Campos, J., Rodrigues, C., Santos, C., Antunes, P., et al. (2018). mcr-1 in carbapenemase-producing Klebsiella pneumoniae with hospitalized patients, Portugal, 2016–2017. Emerg. Infect. Dis. 24, 762–766. doi: 10.3201/eid2404.171787
Nicolas-Chanoine, M. H., Bertrand, X., and Madec, J. Y. (2014). Escherichia coli ST131, an intriguing clonal group. Clin. Microbiol. Rev. 27, 543–574. doi: 10.1128/CMR.00125-13
Papp-Wallace, K. M., Endimiani, A., Taracila, M. A., and Bonomo, R. A. (2011). Carbapenems: past, present, and future. Antimicrob. Agents Chemother. 55, 4943–4960. doi: 10.1128/AAC.00296-11
Papp-Wallace, K. M., Taracila, M., Wallace, C. J., Hujer, K. M., Bethel, C. R., Hornick, J. M., et al. (2010). Elucidating the role of Trp105 in the KPC-2 β-lactamase. Protein Sci. 19, 1714–1727. doi: 10.1002/pro.454
Pitout, J. D. D., and DeVinney, R. (2017). Escherichia coli ST131: a multidrug-resistant clone primed for global domination. F1000Res. 6:F1000 Faculty Rev-195. doi: 10.12688/f1000research.10609.1
Porreca, A. M., Sullivan, K. V., and Gallagher, J. C. (2018). The Epidemiology, evolution, and treatment of KPC-producing organisms. Curr. Infect. Dis. Rep. 20:13. doi: 10.1007/s11908-018-0617-x
Potter, R. F., D’Souza, A. W., and Dantas, G. (2016). The rapid spread of carbapenem-resistant Enterobacteriaceae. Drug Resist. Updat. 29, 30–46. doi: 10.1016/j.drup.2016.09.002
PPCIRA (2017). Programa de Prevenção e Controlo de Infeções e de Resistência aos Antimicrobianos 2017. Lisboa: Direção-Geral da Saúde.
Rodrigues, C., Bavloviè, J., Machado, E., Amorim, J., Peixe, L., and Novais, Â (2016). KPC-3-producing Klebsiella pneumoniae in Portugal linked to previously circulating non-CG258 lineages and uncommon genetic platforms (Tn4401d-IncFIA and Tn4401d-IncN). Front. Microbiol. 7:1000. doi: 10.3389/fmicb.2016.01000
Rodríguez-Baño, J., Gutiérrez-Gutiérrez, B., Machuca, I., and Pascual, A. (2018). Treatment of infections caused by extended-spectrum-beta-lactamase-, AmpC-, and carbapenemase-producing Enterobacteriaceae. Clin. Microbiol. Rev. 31:e00079-17. doi: 10.1128/CMR.00079-17
Smillie, C., Garcillán-Barcia, M. P., Francia, M. V., Rocha, E. P., and de la Cruz, F. (2010). Mobility of plasmids. Microbiol. Mol. Biol. Rev. 74, 434–452. doi: 10.1128/MMBR.00020-10
Stokes, H. W., and Gillings, M. R. (2011). Gene flow, mobile genetic elements and the recruitment of antibiotic resistance genes into Gram-negative pathogens. FEMS Microbiol. Rev. 35, 790–819. doi: 10.1111/j.1574-6976.2011.00273.x
Sullivan, K. M., Dean, A., and Soe, M. M. (2009). OpenEpi: a web-based epidemiologic and statistical calculator for public health. Public Health Rep. 124, 471–474. doi: 10.1177/003335490912400320
Szuplewska, M., Ludwiczak, M., Lyzwa, K., Czarnecki, J., and Bartosik, D. (2014). Mobility and generation of mosaic non-autonomous transposons by Tn3-derived inverted-repeat miniature elements (TIMEs). PLoS One 9:e105010. doi: 10.1371/journal.pone.0105010
van Loon, K., Voor In ‘t Holt, A. F., and Vos, M. C. (2018). A systematic review and meta-analyses of the clinical epidemiology of carbapenem-resistant Enterobacteriaceae. Antimicrob. Agents Chemother. 62:e01730-17. doi: 10.1128/AAC.01730-17
Varani, A., Siguier, P., Gourbeyre, E., Charneau, V., and Chandler, M. (2011). ISsaga is an ensemble of web-based methods for high throughput identification and semi-automatic annotation of insertion sequences in prokaryotic genomes. Genome Biol. 12:R30. doi: 10.1186/gb-2011-12-3-r30
Varela, A. R., Manageiro, V., Ferreira, E., Guimarães, M. A., da Costa, P. M., Canica, M., et al. (2015). Molecular evidence of the close relatedness of clinical, gull and wastewater isolates of quinolone-resistant Escherichia coli. J. Glob. Antimicrob. Resist. 3, 286–289. doi: 10.1016/j.jgar.2015.07.008
Vubil, D., Figueiredo, R., Reis, T., Canha, C., Boaventura, L., and Da Silva, G. J. (2017). Outbreak of KPC-3-producing ST15 and ST348 Klebsiella pneumoniae in a Portuguese hospital. Epidemiol. Infect. 145, 595–599. doi: 10.1017/S0950268816002442
Keywords: carbapenemase-producing Enterobacteriaceae, KPC-21, EuSCAPE, Portugal, Klebsiella pneumoniae, Escherichia coli
Citation: Manageiro V, Romão R, Moura IB, Sampaio DA, Vieira L, Ferreira E, the Network EuSCAPE-Portugal and Caniça M (2018) Molecular Epidemiology and Risk Factors of Carbapenemase-Producing Enterobacteriaceae Isolates in Portuguese Hospitals: Results From European Survey on Carbapenemase-Producing Enterobacteriaceae (EuSCAPE). Front. Microbiol. 9:2834. doi: 10.3389/fmicb.2018.02834
Received: 30 June 2018; Accepted: 05 November 2018;
Published: 27 November 2018.
Edited by:
Patrícia Poeta, Universidade de Trás-os-Montes e Alto Douro, PortugalReviewed by:
Marie-Cecile Ploy, University of Limoges, FranceAlfonso Soler-Bistue, Instituto de Investigaciones Biotecnológicas (IIB-INTECH), Argentina
Copyright © 2018 Manageiro, Romão, Moura, Sampaio, Vieira, Ferreira, the Network EuSCAPE-Portugal and Caniça. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Manuela Caniça, bWFudWVsYS5jYW5pY2FAaW5zYS5taW4tc2F1ZGUucHQ=