 Hongjie Yang1,2
Hongjie Yang1,2 Peishi Jiang
Peishi Jiang Yi Sun
Yi Sun Lingyi Chen
Lingyi Chen Siwei Zhu
Siwei Zhu- 1School of Medicine, Nankai University, Tianjin, China
- 2Department of Colorectal Surgery, Tianjin Union Medical Center, The First Affiliated Hospital of Nankai University, Tianjin, China
- 3The Institute of Translational Medicine, Tianjin Union Medical Center of Nankai University, Tianjin, China
- 4Tianjin Institute of Coloproctology, Tianjin, China
- 5State Key Laboratory of Medicinal Chemical Biology, Tianjin Key Laboratory of Protein Sciences, Frontiers Science Center for Cell Responses, National Demonstration Center for Experimental Biology Education and College of Life Sciences, Nankai University, Tianjin, China
- 6Department of Oncology, Tianjin Union Medical Center, The First Affiliated Hospital of Nankai University, Tianjin, China
Objective: This study aimed to investigate the role of TMEM59L in colorectal cancer (CRC) and its interaction with the TGF-β/Smad signaling pathway.
Methods: We analyzed the correlation between TMEM59L expression levels and patient survival, as well as its impact on the TGF-β/Smad signaling pathway, using data from The Cancer Genome Atlas (TCGA). Additionally, transwell, CCK-8, EdU, and colony formation assays were conducted to assess the effects of TMEM59L on CRC cell migration, invasion, and proliferation. Gene silencing and overexpression, along with specific inhibitors/agonists, were used to validate the involvement of TMEM59L in the regulation of the TGF-β/Smad signaling pathway.
Results: We found that high TMEM59L expression was associated with poor patient survival and TGF-β pathway activation. After si-TMEM59L treatment, the migration and invasion abilities of CRC cells were reduced, while cell proliferation remained affected to a lesser extent. Additionally, the levels of TGF-β protein were decreased, and the phosphorylation of Smad2/3 was reduced. In vivo, TMEM59L knockdown reduced metastatic potential as demonstrated by decreased fluorescence intensity, while overexpression of TMEM59L increased metastatic potential, which was reversed by TGF-β inhibition.
Conclusion: TMEM59L may promote CRC metastasis by enhancing cell migration and invasion, with minimal impact on cell proliferation, potentially through the TGF-β/Smad signaling pathway.
Introduction
Colorectal cancer (CRC) is one of the most common digestive system cancers worldwide, accounting for 10.0% of all cancer cases (1). As the burden of colorectal cancer increases, the identification of effective targeted therapy targets has become increasingly important. TMEM (Transmembrane proteins) are widely distributed proteins in cell membranes with multiple transmembrane domains. They play a key role in tumor progression, influencing processes like cell proliferation, migration, and invasion (2, 3). TMEM45A (4), TMEM45B (5), TMEM48 (6), TMEM98 (7), TMEM119 (8), and TMEM168 (9), proteins are considered oncogenes in certain types of cancer because they contribute to increased cell proliferation, migration, invasion, and EMT. In contrast, proteins such as TMEM106A (10), TMEM170B (11), and TMEM88 (12) are considered tumor suppressors, as they reduce cell proliferation, migration, invasion, and EMT in some cancer types.
TMEM59L was first discovered in 1999 and was initially considered a protein expressed specifically in the brain (13). Research on the mechanism of this protein has primarily focused on the nervous system (14–16). In 2022, the link between TMEM59L and cancers was first discovered. Since then, numerous studies have found that TMEM59L is associated with the survival of patients with cancers (17–19). However, these studies are primarily based on bioinformatics analyses from databases, and there is currently a lack of cell and animal experimental research. Bioinformatics studies have reported that TMEM59L is associated with the activation of the TGF-β/signaling pathway (17). This study aims to explore the effects of TMEM59L on CRC tumor cells and its relationship with the TGF-β signaling pathway through cell and animal experiments. To our knowledge, this is the first study to systematically validate the function of TMEM59L in colorectal cancer through comprehensive in vitro and in vivo experiments, providing a theoretical foundation for developing novel anti-metastatic therapeutic strategies.
Materials and methods
Bioinformatics analysis
The RNA-seq gene expression data and corresponding clinical information of colorectal cancer patients were obtained from The Cancer Genome Atlas (TCGA) database, and the transcriptional levels of TMEM59L mRNA were extracted. Using the optimal cutoff algorithm, the cutoff point that minimized the Log-rank test p-value was determined to classify patients into high and low TMEM59L expression groups. Kaplan-Meier survival curves were then plotted to assess the relationship between TMEM59L expression levels and patient survival rates. Additionally, gene set enrichment analysis (GSEA) was performed to examine whether gene expression in the high TMEM59L expression group was significantly enriched in TGF-β-related signaling pathways. To further investigate the clinical relevance of TMEM59L, we analyzed its expression distribution across pathological stages (I-IV) using violin plots with trend analysis. Multivariable Cox regression analysis was performed using a stepwise approach, progressively adjusting for age, gender, and pathological stage to evaluate the independent prognostic value of TMEM59L expression.
Cell lines and cell culture
The human colorectal cancer cell lines LoVo and HCT116 were obtained from ATCC. The human colorectal cancer cell line HCT116-luc and four derived stable cell lines were obtained from Shanghai Zhong qiao xin zhou biotechnology. The stable cell lines included: TMEM59L knockdown cells (HCT116-luc#hTMEM59L[shRNA], referred to as TMEM59L_KD) with corresponding scramble shRNA control (HCT116-luc#Scramble_shRNA, referred to as Control_KD), and TMEM59L overexpression cells (HCT116-luc#hTMEM59L, referred to as TMEM59L_OE) with corresponding vector control (HCT116-luc#Control, referred to as Control_OE). These cell lines were generated using the Neon™ transfection system (Cat# MPK5000) and selected using hygromycin resistance. All cell lines were validated by quantitative PCR using specific primers for TMEM59L and GAPDH as internal control. The experimental operation was carried out in strict accordance with the instructions on the official website. The supernatant was removed by digestion and centrifugation of the target cells growing logarithmically in good condition (suspended cells could be collected directly), and the cell precipitate was obtained by re-suspension counting with PBS and 1×106 cells were taken into the EP tube for centrifugation. The cells were suspended with electrolysis buffer, and the plasmids were added and thoroughly mixed. Input the optimal power parameters, power the above mixture, and power the EGFP control group under the same parameters. The cells in each group after electrotransfer were transferred to the corresponding culture vessel and cultured in the incubator, and the cell status was observed after 24h, and the electrotransfer efficiency was judged according to the EGFP control group. 48h after cell culture, the target cells after electrotransfer were screened with the screening drug corresponding to the scheme. After the drug screening cycle, the drug-containing medium was removed and replaced with fresh medium, and the cell state was restored after culture for 24-48h.) Cell lines were maintained in RPMI-1640 medium (Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal calf serum (Invitrogen). Cells were cultured at 37 °C with 5% CO2. Antibiotics are purchased from shanghai yesen biotechnology.
In vivo metastasis experiments
Animal study was approved by the independent ethics committee of Tianjin jinke biotechnology. Nude mice (HFK Bio-Technology, Beijing, China) were maintained in accordance with the ethics standards of the World Medical Association (Declaration of Helsinki). Cells were injected to the caudal vein of 16–19 g female nude mice as a 100 µL suspension (5 × 105 cells). After 28 days, mice were sacrificed and lungs were stripped for analysis. D-luciferin potassium salt D (40902ES01) was purchased from shanghai Yesen Biotechnology (Shanghai, China). Bioluminescence imaging was performed in the afternoon on days 0, 7, 14, 21, and 28 post-cell injection. For imaging, potassium luciferin was administered at a dose of 150 mg/kg by intraperitoneal injection. The potassium luciferin was fully dissolved in sterile PBS without Mg2+ and Ca2+, and filtered through a 0.22-micron microporous membrane. Imaging was performed at the peak fluorescence intensity, 10 minutes after the luciferin injection. The TGF-β pathway inhibitor SB-431542 was purchased from Yesen Biotechnology (Shanghai, China) and administered at a dose of 10 mg/kg on days 7, 14, 21, and 28 post-cell injection. It was fully dissolved in 5% DMSO and administered by intraperitoneal injection in the morning, prior to the afternoon imaging sessions.
Hematoxylin-eosin staining
Lungs were immersed in 4% paraformaldehyde for 24 h and transferred to 60% ethanol. Individual lobes of lung biopsy material were placed in processing cassettes, dehydrated through the serial alcohol gradients, and embedded in paraffin. Before immunostaining, 5 μm thick lung tissue sections were dewaxed in xylene, rehydrated through decreasing concentrations of ethanol, and washed in PBS. Then sections were stained with hematoxylin and eosin. After staining, sections were dehydrated through increasing concentrations of ethanol and xylene.
Proliferation assay
Primary cells were seeded in 96-well plates and treated as needed. Cell viability was measured 24\48\72\96 hours after treatment cessation using the Cell-Counting Kit 8 (CCK-8) (40203ES80) from shanghai yesen biotechnology. Plates were incubated for 1.5 hours at 37°C and absorbance was measured at 450nm using the Infinite 200 Tecan i-control plate reader machine. The CCK-8 assay was performed in at least 3 replicates for each experimental condition.
Cells were seeded in a confocal fluorescence microscope special dish and treated with siTMEM59L or NC for 48 h. Cells proliferation was tested using EdU Cell Proliferation Kit (40276ES76) from shanghai yesen biotechnology with Alexa Fluor 594 in accordance with the operating instructions for processing. Cell nuclei were co-stained with Hoechst 33258. After that, EdU-positive cells were monitored with a fluorescence microscope (Olympus Corporation, Tokyo, Japan).
To observe the capacity changes of single cells to form a colony, cells were inoculated treated with siTMEM59L or NC for 48 h. The fresh RPMI1640 medium was used every 3 days to replace the medicated culture medium, and cells were cultured for 10 days. Then, the cellular colonies were immobilized, treated with crystal violet solution and the numbers of colonies were counted.
Cell migration and invasion assays
Cell migration assay was performed in 24-well CIM plates (BD Biosciences, CA, USA). Briefly, 1-3 × 104 cells per well were seeded in serum-free medium in the upper compartment of the CIM plates. Serum-complemented medium was added to the lower compartment of the chamber. After 24 h incubation, cells that passed through the septum were fixed with cold methanol and stained with crystal violet. The average number of migrated cells in four random microscopic fields was counted. Cell invasion assay was performed in 24-well CIM plates coated with matrigel. Other steps were identical to those of cell migration assay.
Western blot analysis
Cells were homogenized in loading buffer (0.1 M Tris-HCl, pH 6.8, 1% SDS, 10% β-mercaptoethanol, 11% glycerol), separated by SDS-PAGE and electro-blotted to the nitrocellulose membranes, then blocked with 5% non-fat milk in TBS for 1.5 h at room temperature. The membranes were incubated with the indicated primary antibodies at 4 °C overnight. After washing three times with TBST, membranes were probed by horseradish peroxidase-labeled secondary antibodies for 45 min at room temperature. Protein bands were visualized by enhanced chemiluminescence. Anti-GAPDH (60004) was from Proteintech (Chicago, IL, USA). The secondary antibodies were purchased from Abcam (Cambridge, UK). Phospho-Smad2/3 (Thr8) (31132ES50), Phospho-Smad2 (Ser467) (31131ES50), Smad2 (31058ES50), TGF-β1 (30013ES50) was purchased from Yeasen Biotechnology (Shanghai, China). TMEM59L (PA5-72688) was purchased from Invitrogen (Thermo Fisher Scientific, Waltham, MA, USA).
RNA interference
Small interfering RNAs (siRNAs) were provided by GenePharma (Shanghai, China) and transfected into cells with siRNA Mate (GenePharma) following the provider’s instructions.
Interference sequences used were:
TMEM59L #1 -sense:
5’- GCGUGGAAGCCUAUGUGAATT -3’;
TMEM59L #1 -antisense:
5’- UUCACAUAGGCUUCCACGCTT-3’;
TMEM59L #2 -sense:
5’- GCAAUGACCUUGUCAACUCTT -3’;
TMEM59L #2 -antisense:
5’- GAGUUGACAAGGUCAUUGCTT -3’;
TMEM59L #3 -sense:
5’- GGCCAAGGUGGAGUCUGAATT -3’;
TMEM59L #3 -antisense:
5’- UUCAGACUCCACCUUGGCCTT -3’;
TMEM59L #4 -sense:
5’- GCACAAGGGCUUCAUGAUGTT -3’;
TMEM59L #4 -antisense:
5’- CAUCAUGAAGCCCUUGUGCTT -3’;
Negative control -sense:
5’- UUCUCCGAACGUGUCACGUTT -3’;
Negative control -antisense:
5’ -ACGUGACACGUUCGGAGAATT -3’.
Statistical analysis
Bioinformatics analysis was performed using R (version 4.3.3), and other statistical analyses were done using Python (version 3.9.7). The unpaired two-tailed t-test was used for in vitro study and vivo study. A two-sided p < 0.05 was considered statistically significant. *** p < 0.001; ** p < 0.01; * p < 0.05; N.S. p > 0.05.
Result
High TMEM59L expression predicts poor survival and is linked to TGF-β pathway enrichment in colorectal cancer
Based on the Kaplan-Meier survival analysis, patients with high TMEM59L expression showed significantly poorer survival rates compared to those with low TMEM59L expression (Figure 1A). The GSEA results for the high TMEM59L expression group indicate significant enrichment in the KEGG TGF-β signaling pathway (Figure 1B). Analysis of TMEM59L expression across pathological stages revealed a significant increasing trend (Spearman rho = 0.242, p < 0.001), with mean expression levels progressively rising from Stage I (0.321 ± 0.304) to Stage IV (0.743 ± 0.777) (Figure 1C). Multivariable Cox regression analysis showed that while TMEM59L was significantly associated with survival in univariate analysis (HR = 1.474, 95% CI: 1.082-2.008, p = 0.014). TMEM59L remained significant after adjusting for age and gender but lost significance after stage adjustment (HR = 1.145, p = 0.457), suggesting the prognostic effect may be mediated by tumor stage (Figure 1D).
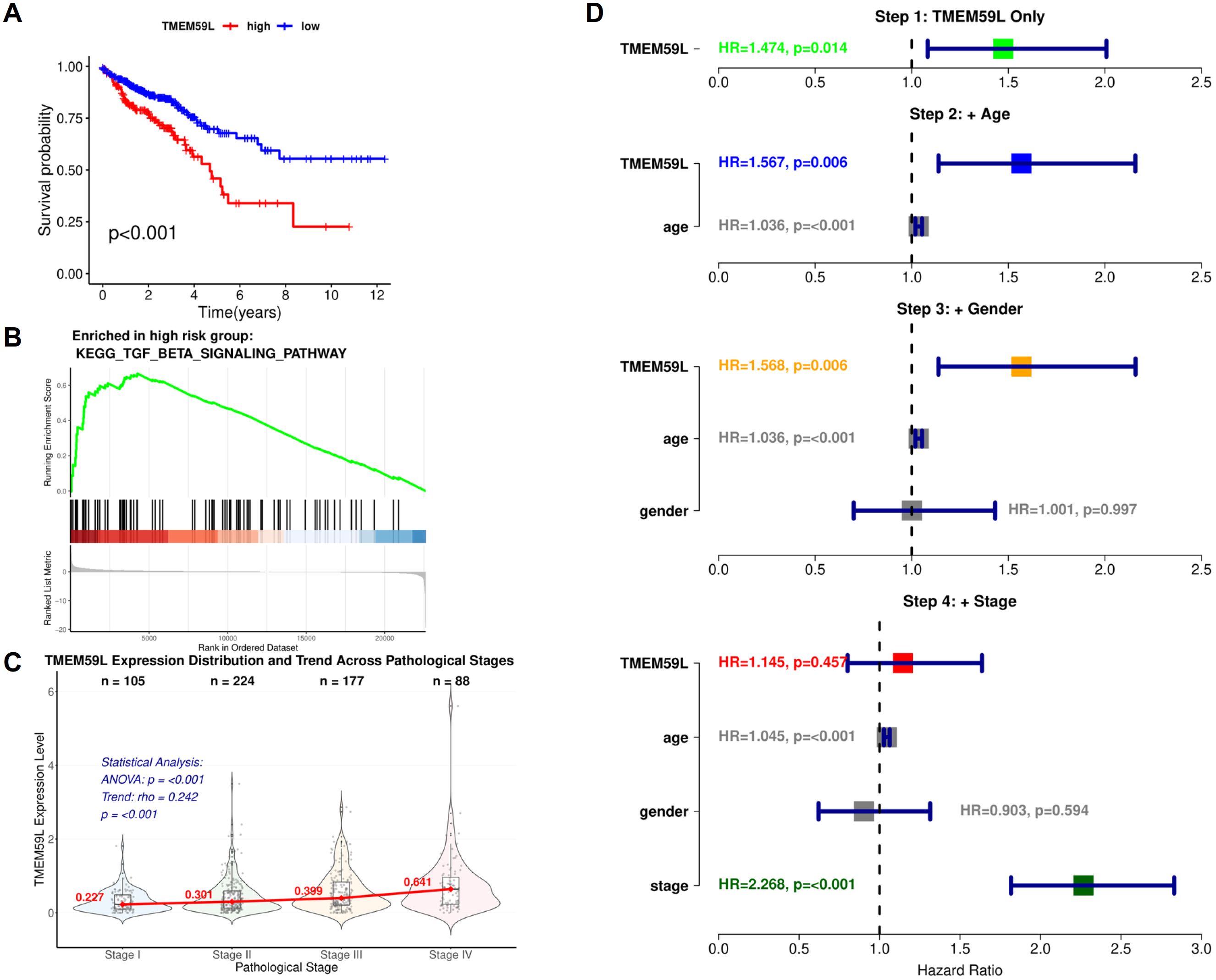
Figure 1. Bioinformatics analysis of TMEM59L. (A) Kaplan-Meier Survival Analysis of Colorectal Cancer Patients Stratified by TMEM59L Expression Levels. (B) GSEA Enrichment Plot of the KEGG TGF-β Signaling Pathway in TMEM59L High-Expression Group. (C) Violin Plot of TMEM59L Expression Distribution Across Pathological Stages (I-IV) with Trend Analysis. (D) Forest Plot of Stepwise Cox Regression Analysis Adjusting for Age, Gender, and Pathological Stage.
Knockdown of TMEM59L inhibits metastasis of colorectal cancer cells in vivo and in vitro
Quantitative PCR validation confirmed successful generation of stable cell lines. TMEM59L_KD showed 92% reduction in mRNA expression (0.08-fold relative to Control_KD), while TMEM59L_OE demonstrated 384-fold increase in TMEM59L mRNA levels compared to Control_OE.We firstly treated two colorectal cancer cell lines HCT116 and LoVo with siRNA-TMEM59L with four different sites small interfering RNA and the siRNA interference effect was effective (Figure 2A). The complete, unprocessed Western blot images are provided in Supplementary Files 1–4. In the following experiment, we selected two siRNAs for each cell line for further experiments. We then performed migration and invasion assays. Knockdown of TMEM59L in colorectal cancer cells inhibited the migration and invasion abilities of both LoVo and HCT116 cells (Figure 2B). To evaluate the influence of TMEM59L knocked down on cancer cell motility in vivo, we injected TMEM59L_KD cells into caudal vein of nude mice and fluorescence was observed continuously for 4 weeks, and fluorescence photographs were taken once a week. TMEM59L_KD cells had poor transfer capacity. At the beginning, the fluorescent intensity of nude mice in both groups was similar, and then the fluorescent intensity was detected every seven days. The fluorescent intensity of the control group gradually became stronger than that of the experimental group, and by day 14, the difference was statistically significant (Figure 3A). The Hematoxylin-eosin staining of sectioned lung tissues also showed that HCT116-luc#shTMEM59L cells groups had less metastases (Figure 3B). These results suggest that TMEM59L knocked down inhibit motility of a subset of colorectal cancer cells in vitro and in vivo.
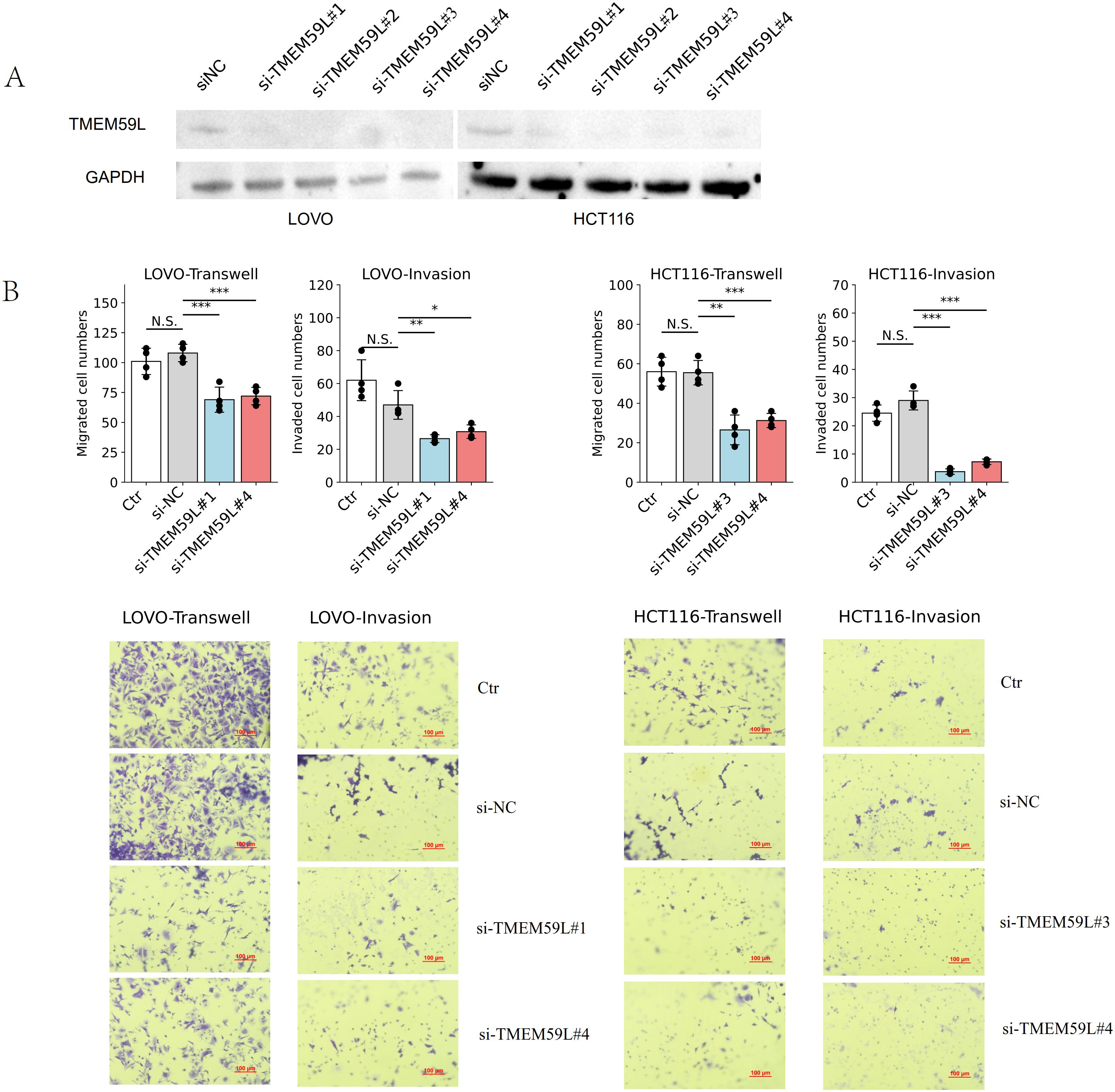
Figure 2. Effects of TMEM59L on cell motility in vitro. (A) Validation of the efficiency of TMEM59L knockdown after transfection with different siRNAs for 48 h (B) TMEM59L knockdown inhibited cell motility. After siRNA transfection for 48 h, cells were subjected to migration or invasion assay for 24 h. *p < 0.05; **p < 0.01; ***p < 0.001.
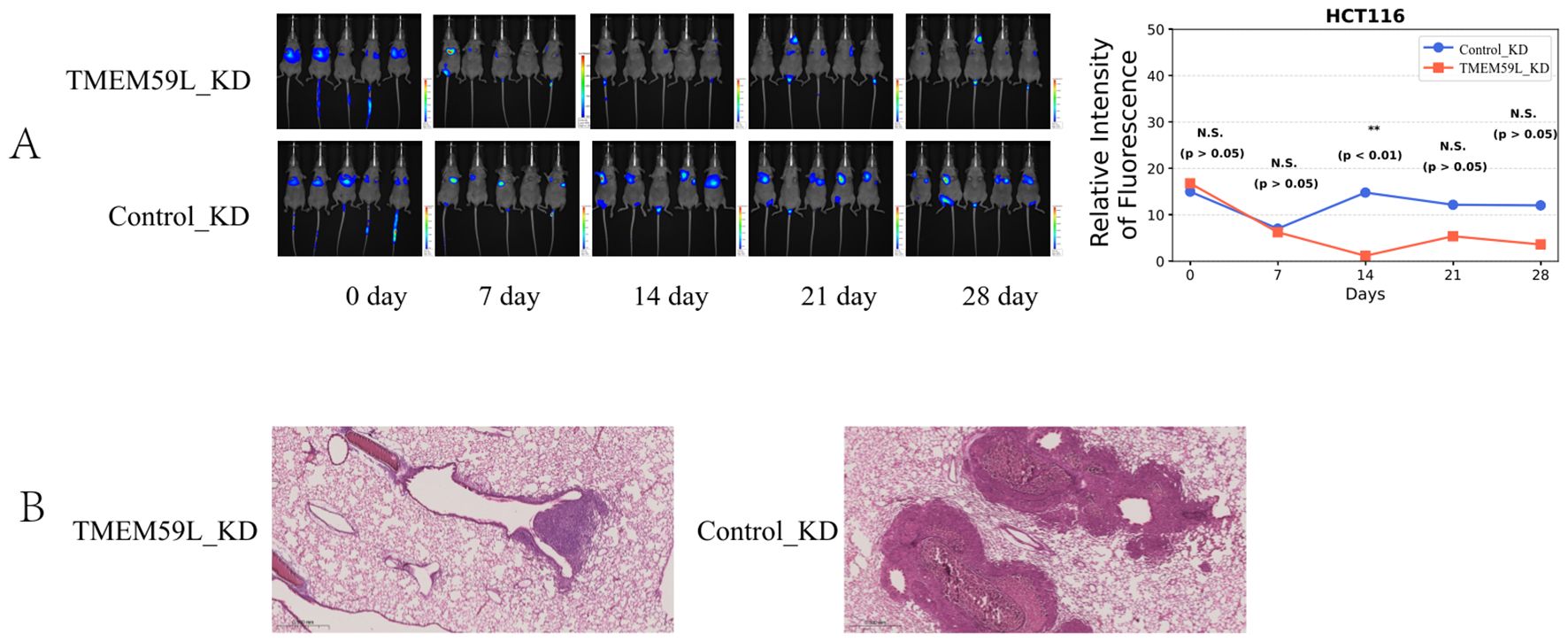
Figure 3. Effects of TMEM59L on cell motility in vivo. (A) Live bioluminescence imaging of nude mice following HCT116 cell injection. Bioluminescence signals were obtained using an IVScope 8200 imaging system at 0, 7, 14, 21, and 28 days post-treatment. Relative intensity of fluorescence in two groups of nude mice. (B) Hematoxylin-eosin staining of the lung tissues of mice. **p < 0.01.
Effect of TMEM59L on cell growth and proliferation
To investigate whether TMEM59L could influence cell growth in HCT116 and LoVo cells, si-TMEM59L was used to treat HCT116 and LoVo cells, and detect the cell viability at different time points (24, 48, 72 and 96 h) by CCK-8 assay. The results show that si-TMEM59L did not have a significant positive or negative influence on cell viability in most groups of HCT116 and LoVo cells. Only in HCT116 cells treated with si-TMEM59L#3 at 96 hours was an increase in cell viability observed. (Figure 4A). Furthermore, the influence of cell growth by TMEM59L was tested by cloning formation and EdU incorporation assays. After si-TMEM59L treatment, the colony numbers showed no statistically significant difference (N.S.) across all groups in both LoVo and HCT116 cells (Figure 4B). Similarly, after si-TMEM59L treatment, the percentage of EdU-positive cells, which indicates actively proliferating cells, did not exhibit any significant changes in either LoVo or HCT116 cells (Figure 4C).
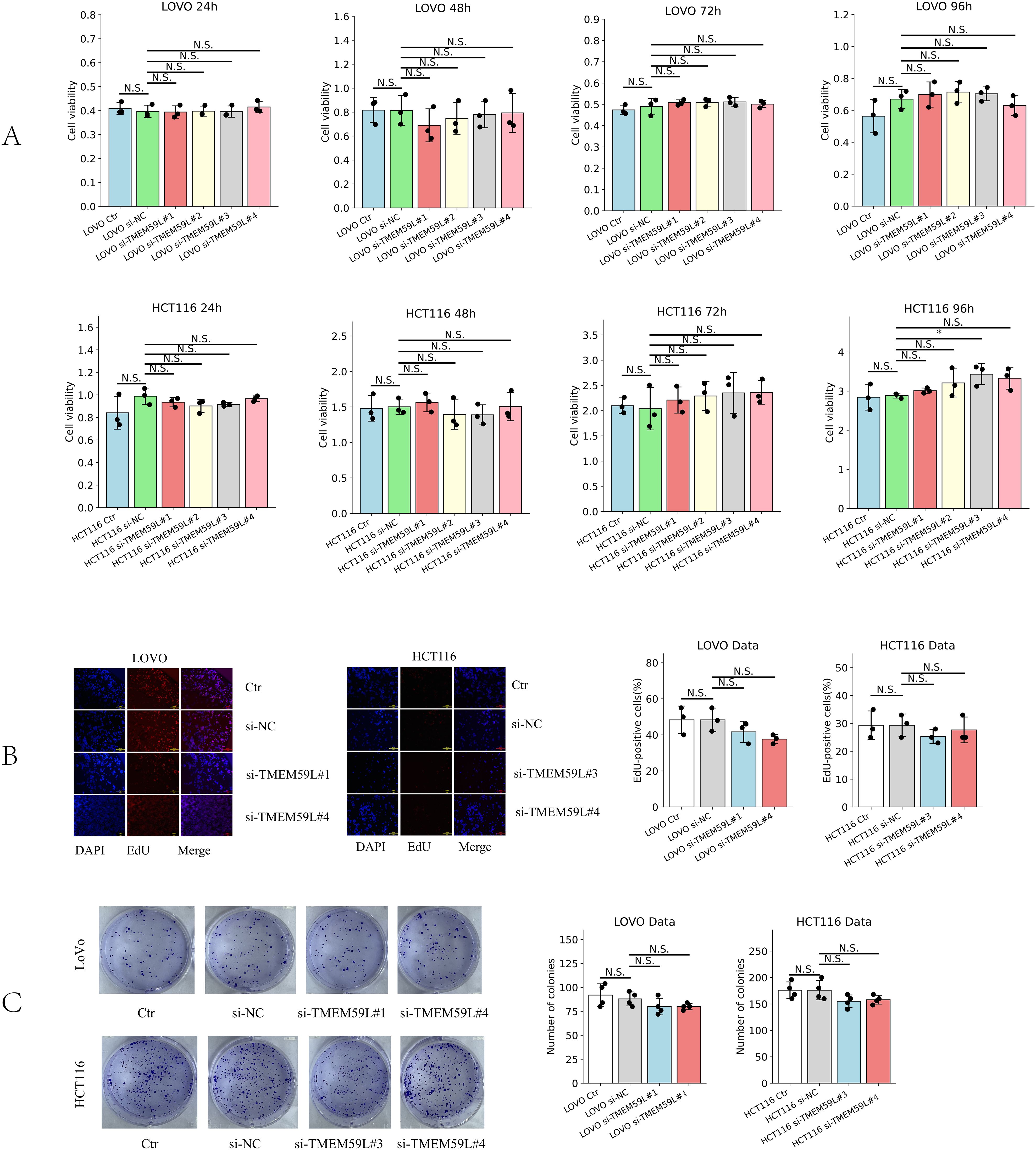
Figure 4. Effect of TMEM59L on cancer cell proliferation. (A) Effect of TMEM59L on cancer cell proliferation detected by CCK-8 assay. (B) Effect of TMEM59L on cancer cell proliferation detected by EdU incorporation assays. (C) HCT116 and LoVo cells were incubated with siRNA-TMEM59L or NC for 10 days, and the number of colonies were calculated.
TGF-β and Smad signaling pathway mediates TMEM59L expression
Data analysis prompted TMEM59L exerting a key influence in cancer development may associated TGF-β signaling (17). Our enrichment analysis of the latest TCGA data reached the same conclusion, so we conducted experimental exploratory verification. After siRNA TMEM59L interference, we found that the TGF-β signaling pathway may play a vital role. TGF-β pathway is involved in activation of Smad signaling pathway (20). We examined the roles of TGF-β and Smad signaling pathway. Treatment with interfering the expression of TMEM59L strongly inhibite TGF-β and inhibited phosphorylation levels of Smad2/3 in HCT116 cells (Figure 5A), but this phenomenon was not obvious in LoVo cells (Figure 5B). The complete, unprocessed Western blot images are provided in Supplementary Files 5–9. SRI-011381 is TGF-β agonist, in HCT116 cells, after treating in SRI-011381, TMEM59L interference induced inhibited phosphorylation levels of Smad2/3 was abrogated (Figure 5C). The complete, unprocessed Western blot images are provided in Supplementary Files 10–14. In animal experiments, To evaluate the impact of TMEM59L overexpression (TMEM59L_OE) on in vivo motility and the relationship between SB431542 and TMEM59L, we divided mice into four groups: Ctr+DMSO, Ctr+SB431542, TMEM59L_OE+DMSO, and TMEM59L_OE+SB431542. Fluorescence images were taken weekly and observed continuously for 4 weeks. The study found that, starting from day 14, the fluorescence intensity in the TMEM59L_OE+DMSO group was higher than that in the Ctr+DMSO group. Furthermore, starting from day 7, the fluorescence intensity in the TMEM59L_OE+SB431542 group was reduced compared to the TMEM59L_OE+DMSO group (Figure 6A). The fluorescence intensity in the TMEM59L_OE+SB431542 group was significantly reduced compared to the TMEM59L_OE+DMSO group on day 14 and day 21 (p < 0.05) (Figure 6B). Hematoxylin-eosin staining of sectioned lung tissues also showed that the SB431542 TMEM59L_OE+SB431542 group had fewer metastatic lesions compared to the TMEM59L_OE+DMSO group (Figure 6C).
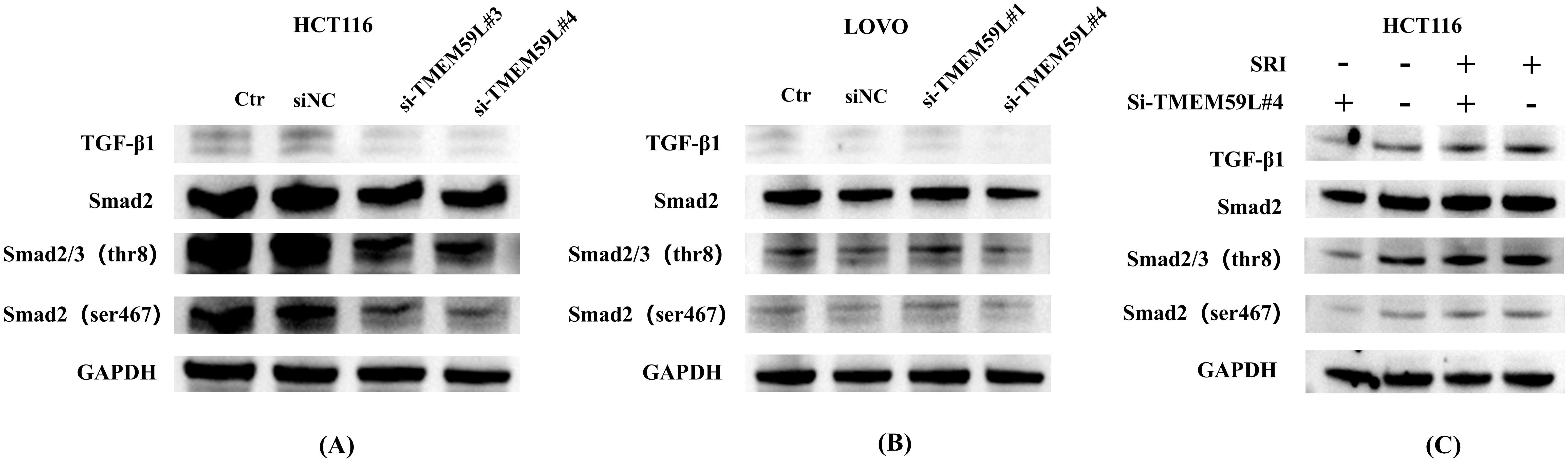
Figure 5. TGF-β pathway mediates TMEM59L expression in vitro. (A) Western blot analysis of Smad phosphorylation and TGF-β expression in HCT116 cells transfected with TMEM59L-specific siRNA for 48 hours. The same GAPDH loading control is shown for all groups in panel (A, B) Western blot analysis of Smad phosphorylation and TGF-β expression in LoVo cells transfected with TMEM59L-specific siRNA for 48 hours. The same GAPDH loading control is shown for all groups in panel (B, C) Western blot analysis of Smad phosphorylation and TGF-β expression in HCT116 cells pretreated with SRI-011381 for 1 hour, followed by TMEM59L-specific siRNA treatment for 48 hours. The same GAPDH loading control is shown for all groups in panel (C).
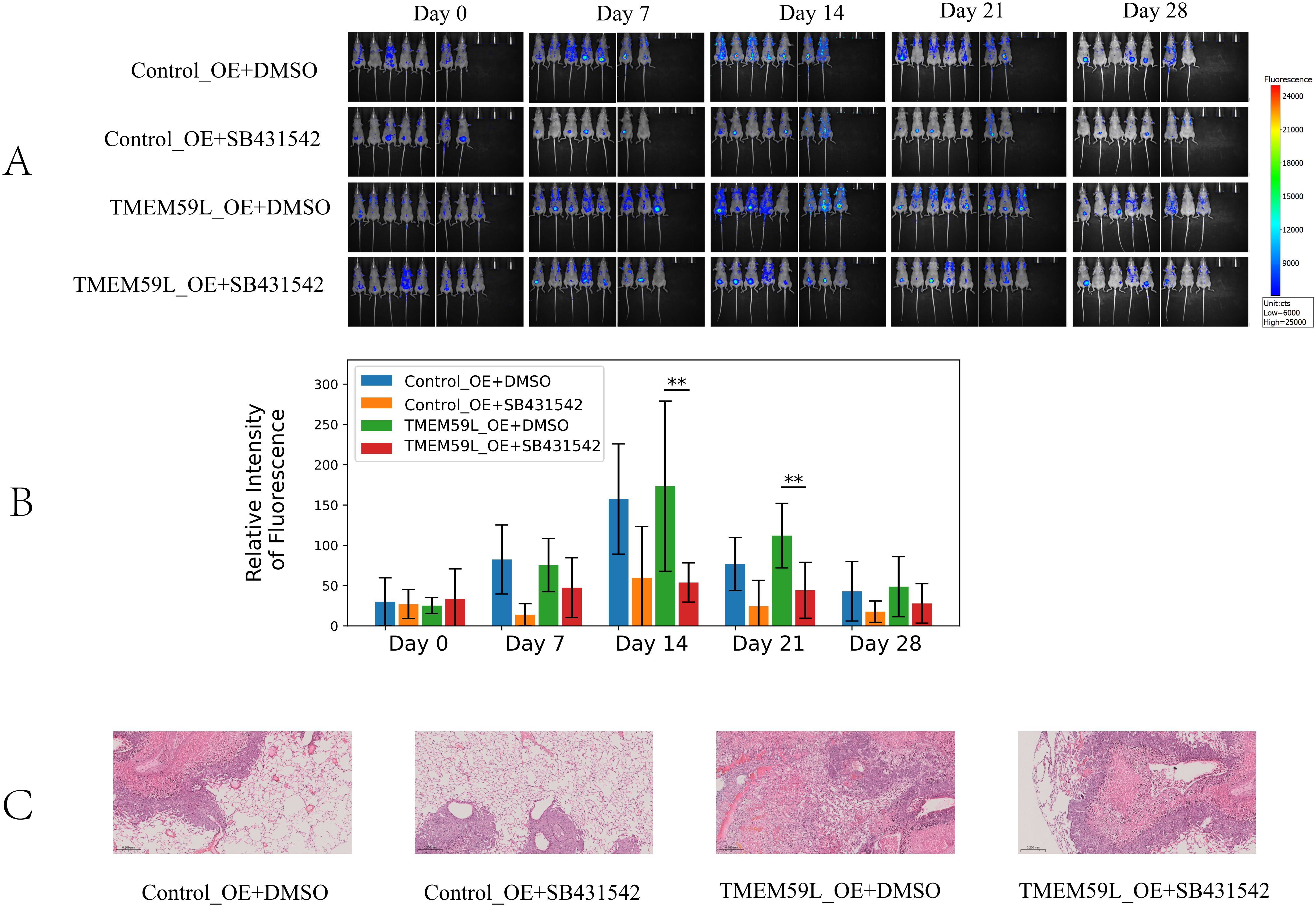
Figure 6. TGF-β pathway mediates TMEM59L expression in vivo. (A) Live bioluminescence imaging of nude mice following HCT116 cell injection. Bioluminescence signals were obtained using an IVScope 8200 imaging system at 0, 7, 14, 21, and 28 days post-treatment. Relative intensity of fluorescence in two groups of nude mice. (B) Relative intensity of fluorescence in two groups of nude mice. (C) Hematoxylin-eosin staining of the lung tissues of mice. **p < 0.01.
Discussion
TMEM59L first appeared in a 1999 article, which reported the discovery of a gene encoding a novel protein, brain-specific membrane-associated protein (BSMAP), now known as TMEM59L. Northern blot analysis showed that BSMAP mRNA is preferentially and highly expressed in the brain, with the protein predicted to be a type I membrane glycoprotein, potentially involved in the functions of the central nervous system (13). In 2014, Aoki et al. (21) reported that TMEM59L is a gene associated with axon growth and is downstream of Islet2a, playing a role in the development of sensory neurons. In 2016, Kobayashi et al. (22) reported that knockdown of TMEM59L significantly reduced insulin secretion from MIN6c4 cells under glucose and/or KCl stimulation, but did not significantly alter the cellular insulin content. However, overexpression of TMEM59L increased insulin secretion. In 2017, Zheng et al. (14) reported that TMEM59L is primarily localized to the Golgi and endosomes. It can interact with autophagy-related proteins ATG5 and ATG16L1, and its overexpression triggers autophagy, promoting caspase-dependent cell apoptosis. Knockdown of TMEM59L reduces anxiety and depression in mice. In 2020, Rutledge et al (23). reported that TMEM59L, along with other genes such as Hs3st3a1 and Hs3st3b1, is involved in the ureteric branching program, which plays a role in the branching morphogenesis of the ureter during kidney development. In 2024, Yuan et al (15). reported that TMEM59L is one of the neuronal marker genes and is highly expressed in differentiated neuronal cell clusters, suggesting that it may be involved in the differentiation process of adipose-derived stromal cells induced into neurons, suggesting its potential as a marker for neurodevelopmental processes.
The relationship between TMEM59L and cancers was first reported in 2022. In that year, Kołat et al. (24) reported that TMEM59L may act as a target of the AP-2 transcription factor and be involved in cancer progression. In the same year, a study by Chang Shi et al. (17) discovered that high TMEM59L expression is associated with shorter survival in patients with various cancers, while higher TMEM59L methylation levels correlate with longer survival. In 2023, Yang et al (18) reported that TMEM59L is a key marker gene for predicting lymph node metastasis in CRC patients and further confirmed that its high expression is closely associated with shortened overall survival in CRC patients. In 2024, Liu et al (19). also found an association between high TMEM59L expression and shortened overall survival in CRC patients.
Our study demonstrates that high TMEM59L expression in CRC is linked to poor survival and significant enrichment of the TGF-β signaling pathway. Kaplan-Meier analysis revealed that patients with high TMEM59L expression had worse survival compared to those with low expression, supporting its role in cancer progression. GSEA further confirmed enrichment of the TGF-β pathway in the high TMEM59L group, suggesting that TMEM59L may influence tumor biology and contribute to CRC metastasis via this pathway.
Cell migration and invasion are key processes for tumor cells to acquire metastatic potential. In in vitro cell experiments, enhanced migration and invasion are typically associated with epithelial-mesenchymal transition (EMT) (25, 26), cell adhesion and detachment (27), activation of specific signaling pathways (such as TGF-β signaling pathway) (28), and immune microenvironment modulation (29). This study is the first to investigate the association between TMEM59L and cancers through cell and animal experiments. We found that knockdown of TMEM59L in HCT116 and LoVo cells significantly inhibited their migration and invasion, both in vitro and in vivo. Regarding cell proliferation and growth, knockdown of TMEM59L did not significantly affect cell viability, colony formation, or EdU incorporation assays in most cell lines, suggesting that TMEM59L may not directly regulate CRC cell growth under these experimental conditions. However, we did observe an increase in cell viability in HCT116 cells 96 hours after treatment with si-TMEM59L#3, suggesting that TMEM59L may play a role in suppressing tumor cell viability. The selective effect of TMEM59L on cell migration and invasion, with minimal impact on proliferation, represents an interesting functional profile that warrants further investigation.
In the early stages of cancer, TGF-β protein exerts a cancer-suppressive effect by inducing cell cycle arrest and apoptosis. During cancer progression, cancer cells gradually develop resistance and begin to secrete TGF-β on their own (30). The aberrant activation of the TGF-β signaling pathway can promote tumor cell migration, invasion, and EMT, as well as immune evasion, thereby driving tumor aggressiveness and metastasis (31). We found that TMEM59L knockdown significantly inhibited TGF-β/Smad signaling. Treatment with the TGF-β agonist SRI-011381 reversed the inhibitory effects of TMEM59L knockdown on Smad phosphorylation, suggesting that TMEM59L regulates metastasis via the TGF-β signaling pathway. In vivo, TMEM59L overexpression significantly promoted metastasis, as indicated by increased fluorescence intensity. Treatment with the TGF-β receptor inhibitor SB431542 reduced metastatic potential in the overexpression groups as demonstrated by decreased fluorescence intensity, further confirming the involvement of the TGF-β pathway in TMEM59L-mediated metastasis. Our study is the first to reveal the role of TMEM59L in colorectal cancer metastasis through animal and cell experiments. While our study demonstrates that TMEM59L modulates TGF-β signaling, the precise molecular mechanism remains to be elucidated. Previous studies have shown that TMEM59L is primarily localized to the Golgi and endosomes and can interact with autophagy-related proteins (14). Given this subcellular localization and our observation of concurrent reduction in TGF-β protein levels and Smad2/3 phosphorylation following TMEM59L knockdown, TMEM59L may regulate TGF-β signaling through intracellular trafficking or protein processing mechanisms. However, the specific molecular interactions require further investigation.
This study has several limitations. First, while we demonstrate that TMEM59L modulates TGF-β signaling, the precise molecular mechanism remains unclear—we cannot determine whether TMEM59L affects TGF-β transcription, protein secretion, or receptor signaling. Second, the tail vein injection model has inherent limitations as it bypasses the early stages of metastasis including local invasion and intravasation, therefore not fully recapitulating the complete spontaneous metastatic process that occurs in clinical settings. Third, the differential TGF-β pathway response to TMEM59L knockdown between HCT116 and LoVo cells remains unexplained and requires further investigation to determine the molecular basis for this cellular heterogeneity.
Conclusions
In conclusion, our findings suggest that TMEM59L plays a pivotal role in CRC metastasis, potentially through the modulation of the TGF-β/Smad signaling pathway. TMEM59L promotes CRC metastasis by enhancing cell migration and invasion, with minimal impact on cell proliferation. These results highlight the potential of TMEM59L as a therapeutic target for inhibiting CRC metastasis, particularly through the modulation of the TGF-β/Smad pathway. Future research directions should include detailed mechanistic studies to elucidate how TMEM59L regulates TGF-β signaling, investigation of TMEM59L function in additional cancer models, and exploration of the molecular basis for cellular heterogeneity in pathway responses.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Ethics statement
Ethical approval was not required for the study involving humans in accordance with the local legislation and institutional requirements. Written informed consent to participate in this study was not required from the participants or the participants’ legal guardians/next of kin in accordance with the national legislation and the institutional requirements. The animal study was approved by Ethics committee of Tianjin jinke biotechnology. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
HY: Investigation, Methodology, Writing – original draft. JL: Conceptualization, Writing – review & editing. PJ: Writing – review & editing. YS: Writing – review & editing. LC: Writing – review & editing. SZ: Project administration, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This research was funded by the National Science Foundation of China (No. 11774256 and No.12174204), the Tianjin Key Medical Discipline (Specialty) Construction Project (No. TJYXZDXK-044A), the Joint Scientific Research Project between Nankai University and Tianjin Union Medical Center (No. 2016rmnk002), the Foundation of the Committee on Science and Technology of Tianjin (Grant Number 24JCZDJC01340), and the Hospital-level Scientific Research Fund of Tianjin Union Medical Center (grant number 2025YJZD003).
Acknowledgments
The authors would like to thank TCGA project for providing the publicly available data used in this study. We also express our gratitude to all patients and researchers involved in TCGA for their invaluable contributions.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. During the preparation of this work, the author used ChatGPT for language polishing. After using this tool, the author reviewed and edited the content as needed and take full responsibility for the content of the published article.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fonc.2025.1674849/full#supplementary-material
Abbreviations
CRC, Colorectal Cancer; si-TMEM59L, Small interfering RNA targeting TMEM59L; TMEM59L_KD, TMEM59L knockdown stable cell line; Control_KD, Scramble shRNA control stable cell line; TMEM59L_OE, TMEM59L overexpression stable cell line; Control_OE, Vector control stable cell line; TCGA, The Cancer Genome Atlas; CCK-8, Cell Counting Kit-8; EdU, 5-Ethynyl-2’-deoxyuridine; KEGG, Kyoto Encyclopedia of Genes and Genomes; TMEM, Transmembrane Protein; GSEA, Gene Set Enrichment Analysis; NC, Negative Control.
References
1. Eng C, Yoshino T, Ruíz-García E, Mostafa N, Cann CG, O’Brian B, et al. Colorectal cancer. Lancet. (2024) 404:294–310. doi: 10.1016/S0140-6736(24)00360-X
2. Schmit K and Michiels C. TMEM proteins in cancer: A review. Front Pharmacol. (2018) 9:1345. doi: 10.3389/fphar.2018.01345
3. Herrera-Quiterio GA and Encarnación-Guevara S. The transmembrane proteins (TMEM) and their role in cell proliferation, migration, invasion, and epithelial-mesenchymal transition in cancer. Front Oncol. (2023) 13:1244740. doi: 10.3389/fonc.2023.1244740
4. Jiang H, Chen H, Wan P, Liang M, and Chen N. Upregulation of TMEM45A promoted the progression of clear cell renal cell carcinoma in vitro. J Inflammation Res. (2021) 14:6421–30. doi: 10.2147/JIR.S341596
5. Zhao LC, Shen BY, Deng XX, Chen H, Zhu ZG, and Peng CH. TMEM45B promotes proliferation, invasion and migration and inhibits apoptosis in pancreatic cancer cells. Mol Biosyst. (2016) 12:1860–70. doi: 10.1039/C6MB00203J
6. Jiang XY, Wang L, Liu ZY, Song WX, Zhou M, and Xi L. TMEM48 promotes cell proliferation and invasion in cervical cancer via activation of the Wnt/β-catenin pathway. J Recept Signal Transduct Res. (2021) 41:371–7. doi: 10.1080/10799893.2020.1813761
7. Mao M, Chen J, Li X, and Wu Z. siRNA-TMEM98 inhibits the invasion and migration of lung cancer cells. Int J Clin Exp Pathol. (2015) 8:15661–9.
8. Sun T, Bi F, Liu Z, and Yang Q. TMEM119 facilitates ovarian cancer cell proliferation, invasion, and migration via the PDGFRB/PI3K/AKT signaling pathway. J Transl Med. (2021) 19:111. doi: 10.1186/s12967-021-02781-x
9. Xu J, Su Z, Ding Q, Shen L, Nie X, Pan X, et al. Inhibition of proliferation by knockdown of transmembrane (TMEM) 168 in glioblastoma cells via suppression of wnt/β-catenin pathway. Oncol Res. (2019) 27:819–26. doi: 10.3727/096504018X15478559215014
10. Liu J and Zhu H. TMEM106A inhibits cell proliferation, migration, and induces apoptosis of lung cancer cells. J Cell Biochem. (2019) 120:7825–33. doi: 10.1002/jcb.28057
11. Li M, Han Y, Zhou H, Li X, Lin C, Zhang E, et al. Transmembrane protein 170B is a novel breast tumorigenesis suppressor gene that inhibits the Wnt/β-catenin pathway. Cell Death Dis. (2018) 9:91. doi: 10.1038/s41419-017-0128-y
12. Zhao X, Li G, Chong T, Xue L, Luo Q, Tang X, et al. TMEM88 exhibits an antiproliferative and anti-invasive effect in bladder cancer by downregulating Wnt/β-catenin signaling. J Biochem Mol Toxicol. (2021) 35:e22835. doi: 10.1002/jbt.22835
13. Elson GC, de Coignac AB, Aubry JP, Delneste Y, Magistrelli G, Holzwarth J, et al. BSMAP, a novel protein expressed specifically in the brain whose gene is localized on chromosome 19p12. Biochem Biophys Res Commun. (1999) 264:55–62. doi: 10.1006/bbrc.1999.1481
14. Zheng Q, Zheng X, Zhang L, Luo H, Qian L, Fu X, et al. The neuron-specific protein TMEM59L mediates oxidative stress-induced cell death. Mol Neurobiol. (2017) 54:4189–200. doi: 10.1007/s12035-016-9997-9
15. Yuan X, Li W, Liu Q, Long Q, Yan Q, and Zhang P. Genomic characteristics of adipose-derived stromal cells induced into neurons based on single-cell RNA sequencing. Heliyon. (2024) 10:e33079. doi: 10.1016/j.heliyon.2024.e33079
16. Ma Z, Li W, Zhuang L, Wen T, Wang P, Yu H, et al. TMEM59 ablation leads to loss of olfactory sensory neurons and impairs olfactory functions via interaction with inflammation. Brain Behav Immun. (2023) 111:151–68. doi: 10.1016/j.bbi.2023.04.005
17. Shi C, Zhang L, Chen D, Wei H, Qi W, Zhang P, et al. Prognostic value of TMEM59L and its genomic and immunological characteristics in cancer. Front Immunol. (2022) 13:1054157. doi: 10.3389/fimmu.2022.1054157
18. Yang H, Liu J, Jiang P, Li P, Zhou Y, Zhang Z, et al. An analysis of the gene expression associated with lymph node metastasis in colorectal cancer. Int J Genomics. (2023) 2023:9942663. doi: 10.1155/2023/9942663
19. Liu Q and Liao L. Identification of macrophage-related molecular subgroups and risk signature in colorectal cancer based on a bioinformatics analysis. Autoimmunity. (2024) 57:2321908. doi: 10.1080/08916934.2024.2321908
20. Wang Q, Xiong F, Wu G, Wang D, Liu W, Chen J, et al. SMAD proteins in TGF-β Signalling pathway in cancer: regulatory mechanisms and clinical applications. Diagn (Basel Switzerland). (2023) 13:2769. doi: 10.3390/diagnostics13172769
21. Aoki M, Segawa H, Naito M, and Okamoto H. Identification of possible downstream genes required for the extension of peripheral axons in primary sensory neurons. Biochem Biophys Res Commun. (2014) 445:357–62. doi: 10.1016/j.bbrc.2014.01.193
22. Kobayashi M, Yamato E, Tanabe K, Tashiro F, Miyazaki S, and Miyazaki J. Functional analysis of novel candidate regulators of insulin secretion in the MIN6 mouse pancreatic β Cell line. PloS One. (2016) 11:e0151927. doi: 10.1371/journal.pone.0151927
23. Rutledge EA and McMahon AP. Mutational analysis of genes with ureteric progenitor cell-specific expression in branching morphogenesis of the mouse kidney. Dev Dyn. (2020) 249:765–74. doi: 10.1002/dvdy.157
24. Kołat D, Kałuzińska Ż, Bednarek AK, and Płuciennik E. Prognostic significance of AP-2α/γ targets as cancer therapeutics. Sci Rep. (2022) 12:5497. doi: 10.1038/s41598-022-09494-1
25. Carter P and Kang Y. Tumor heterogeneity and cooperating cancer hallmarks driven by divergent EMT programs. Cancer Res. (2024) 85:12–4. doi: 10.1158/0008-5472.CAN-24-4309
26. Zhang X, Zhang X, Li M, Jiao S, and Zhang Y. Monitoring partial EMT dynamics through cell mechanics using scanning ion conductance microscopy. Anal Chem. (2024) 96:14835–42. doi: 10.1021/acs.analchem.4c02612
27. Haake SM, Rios BL, Pozzi A, and Zent R. Integrating integrins with the hallmarks of cancer. Matrix Biol. (2024) 130:20–35. doi: 10.1016/j.matbio.2024.04.003
28. Xiao L, Li Q, Chen S, Huang Y, Ma L, Wang Y, et al. ADAMTS16 drives epithelial-mesenchymal transition and metastasis through a feedback loop upon TGF-β1 activation in lung adenocarcinoma. Cell Death Dis. (2024) 15:837. doi: 10.1038/s41419-024-07226-z
29. Zhou HJ, Mu BX, Wen MC, Zhao Q, Li Y, Zhao WX, et al. Yiqi Huayu Jiedu Decoction reduces colorectal cancer liver metastasis by promoting N1 neutrophil chemotaxis. Front Immunol. (2025) 16:1530053. doi: 10.3389/fimmu.2025.1530053
30. Shi Y and Massagué J. Mechanisms of TGF-beta signaling from cell membrane to the nucleus. Cell. (2003) 113:685–700. doi: 10.1016/S0092-8674(03)00432-X
Keywords: colorectal neoplasms, TMEM59L, TGF-β signaling pathway, metastasis, therapeutic target
Citation: Yang H, Liu J, Jiang P, Sun Y, Chen L and Zhu S (2025) The role of TMEM59L in colorectal cancer progression and its interaction with the TGF-β/Smad pathway. Front. Oncol. 15:1674849. doi: 10.3389/fonc.2025.1674849
Received: 23 August 2025; Accepted: 10 November 2025; Revised: 04 November 2025;
Published: 25 November 2025.
Edited by:
Zhongqiu Wang, Affiliated Hospital of Nanjing University of Chinese Medicine, ChinaReviewed by:
Richa Shrivastava, Birla Institute of Technology and Science, IndiaBakrim Saad, Université Ibn Zohr, Morocco
Copyright © 2025 Yang, Liu, Jiang, Sun, Chen and Zhu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Siwei Zhu, emh1c2l3ZWlAMTYzLm5ldA==