 Markus Schmidt
Markus Schmidt Günter Seyfried
Günter Seyfried Uliana Reutina
Uliana Reutina Zeki Seskir
Zeki Seskir Eduardo R. Miranda
Eduardo R. Miranda- 1Biofaction KG, Vienna, Austria
- 2Institute for Technology Assessment and Systems Analysis, Karlsruhe Institute of Technology, Karlsruhe, Germany
- 3School of Art, Design and Architecture, University of Plymouth, Plymouth, United Kingdom
This study investigates whether it is possible to simulate quantum entanglement with theoretical memristor models, physical memristors (from Knowm Inc.) and slime molds Physarum polycephalum as bioelectric components. While the simulation with theoretical memristor models has been demonstrated in the literature, real-world experiments with electric and bioelectric components had not been done so far. Our analysis focused on identifying hysteresis curves in the voltage-current (I-V) relationship, a characteristic signature of memristive devices. Although the physical memristor produced I-V diagrams that resembled more or less hysteresis curves, the small parasitic capacitance introduced significant problems for the planned entanglement simulation. In case of the slime molds, and unlike what was reported in the literature, the I-V diagrams did not produce a memristive behavior and thus could not be used to simulate quantum entanglement. Finally, we designed replacement circuits for the slime mold and suggested alternative uses of this bioelectric component.
1 Introduction
This paper focuses on slime mold-based bioelectronics and an attempt to simulate quantum computing with bioelectric components. The synthesis of biology, electronics, and quantum mechanics explores the convergence of disparate fields, offering insights of the challenges of bioelectronics and quantum computational paradigms. Through this interdisciplinary lens, we uncover the difficulties at the intersection of nature-inspired algorithms and quantum computing theory.
We started with the following assumptions: i) slime molds can (sometimes) act as memristors, ii) memristors can be used to simulate quantum circuits. Based on these assumptions, we wanted to experimentally find out if slime molds could be used to simulate quantum circuits classically.
In the first part of the manuscript, we introduce the memristor as a relatively new electronic component, the slime mold Physarum polycephalum as an experimental organism, and a summary of how quantum entanglement could be simulated with memristors. In the results section, we present the outcomes of our experiments, examine the electric characteristics of slime molds as potential biomemristors, and finally, discuss alternative bioelectric applications for the slime mold.
1.1 Memristors
A memristor is a non-linear electronic device that is a resistor with a memory, hence the name memristor. It is the fourth fundamental two-terminal circuit element. The other three are the resistor, defined by a relationship between voltage (v) and current (i); the inductor, defined by a relationship between flux
While the memristor was theoretically predicted in the early 1970s (Chua, 1971), it was only in 2008 when a team of scientists from Hewlett-Packard labs built the first operational monolithic memristor made by sandwiching a thin film of titanium dioxide between platinum electrodes (Strukov et al., 2008). In addition to providing the physical operating mechanism (Strukov et al., 2008), they also showed a pinched hysteresis loop (which looks like a tilted figure eight), which is the hallmark of all memristors. In contrast to a regular resistor, the memristor has a dynamic relationship between current (I) and voltage (V), which means that the electrical resistance (R = U/I) changes because of past voltages. Following the invention of the physical memristor, additional two-terminal circuit elements with memory were described, namely the memcapacitor and the meminductor (Di Ventra et al., 2009). Memfractors or fractional memory elements have interpolated characteristics between a memcapacitor, a memristor, and a meminductor (Abdelouahab et al., 2014).
Memristive devices could potentially be an alternative for CMOS-based logic computation as they do not require a separate data storage and computation (von Neumann) architecture (Luo et al., 2020; Lehtonen et al., 2010; Chattopadhyay and Rakosi, 2011).
They can carry out fundamental Boolean logic operations (Lehtonen et al., 2010) where the same devices serve simultaneously as logic gates and memory that use resistance (instead of voltage or charge) as the physical state variable (Ielmini and Milo, 2017; Borghetti et al., 2010). Studies in recent years have shown that we can expect viable memristor-based non-von Neumann hardware solutions for deep neural networks, artificial intelligence and edge computing (Yao et al., 2020; Jebali et al., 2024; Aguirre et al., 2024; Huang et al., 2024).
Sparse coding with memristor networks is of broad interest for applications in signal processing, computer vision, object recognition and neurobiology (Sheridan et al., 2017). The connection between memristors and neurobiology is of particular interest. Memristors have for example been deployed to create artificial neurons and carry out neuromorphic computing (Snider, 2003; Snider et al., 2011; Park et al., 2022).
The learning behavior of slime molds (Physarum polycephalum) has also been modeled with memristors (Pershin et al., 2009).
1.2 Slime mold Physarum polycephalum as an experimental organism and possible memristor
Physarum polycephalum is a slime mold found in cool, moist and dark environments, for example, in the woods, on decaying leaves and tree bark (Figure 1). It is a single eukaryotic cell with many heads; hence the term ‘polycephalum’. It is typically yellow and visible to the naked eye. This organism feeds through a process called phagocytosis. It coats its food in enzymes, allowing specific nutrients to be ingested, leaving behind a mass of unwanted material. In the laboratory, oat flakes are used to feed it (Figure 1).

Figure 1. Physarum polycephalum on decaying a tree bark (a). Physarum polycephalum culture in a Petri dish feeding on oat flakes (b).
Physarum polycephalum exhibits a complex life cycle, but the point of interest here is its plasmodium stage, when the organism actively searches for food. As it does so, it grows a network of protoplasmic tubes that rhythmically contract and expand, producing shuttle streaming of its internal fluid.
The organism is straightforward to culture in Petri dishes (Figure 1) and it is relatively easy to prompt it to grow specific topologies of protoplasmic tubes by placing food and repellents (e.g. salt) strategically on the dish. The ability to manipulate its growth patterns has underpinned the early stages of research into building Physarum polycephalum-based machines to realize tasks deemed as computational: e.g., the organism was prompted to find the shortest path to a target destination through a maze (Adamatzky, 2015b).
The focus of our research is the electrical behavior of Physarum polycephalum. Its intracellular activity produces fluctuating levels of electrical potential. Typically, this is in the range of ± 50 mV, displaying oscillations at periods between 50s and 200s, with amplitudes ranging from 5 mV to 40 mV (Kishimoto, 1958; Adamatzky and Jones, 2011; Adamatzky, 2014; Boussard et al., 2021; Wohlfarth-Bottermann, 1979). Interestingly, electrical current can be relayed through its protoplasmic tube, and this prompts it to behave like an electronic component.
Several research papers have found that Physarum polycephalum has the ability to solve mazes (Nakagaki et al., 2000), remember and anticipate events (Saigusa et al., 2008), actively balance its diet (Bonner, 2010) and grow in a way that matches efficient road networks (Adamatzky and Jones, 2010). Researchers in the field of unconventional computing are quite fascinated with the ways P. polycephalum can be used in electronics and computing, one review paper lists 25 different applications (Adamatzky, 2015b).
In the literature, slime molds have also been reported to sometimes have memristive properties (Gale et al., 2016). Gale et al. wrote, for example, that they tested 11 slime molds of which “2 exhibited good memristance curves, the other 9 exhibited open curves” (Gale et al., 2014). Other studies reported no memristive behavior in slime mold (Schmidt et al., 2025).
1.3 Simulating quantum circuits with memristors
For the development of quantum computing, considering its associated costs, limited availability, and high error rates, not every experiment can or should run on a quantum computer. This is why quantum simulators and classical simulators of quantum computers are essential parts of the quantum computing development efforts. The Gottesman-Knill theorem (Aaronson and Gottesman, 2004) states that a quantum circuit consisting solely of CNOT, Hadamard, and phase gates can be classically efficiently simulated. Therefore, in addition to the quantum hardware that can simulate quantum computers, there is a wide range of quantum circuits that can run operations efficiently on classical hardware without the need for a quantum hardware. Below, a range of available quantum simulators and proposals are covered.
Memristors were reported to efficiently simulate certain quantum circuits containing Hadamard and CNOT gates, which combined can be used to create entanglement (Karafyllidis et al., 2018; Fyrigos et al., 2018; Karafyllidis et al., 2019). According to these authors, memristors can be used for Qubit state representation and simulating quantum computers. Here we provide a very short summary of how this is done, see the original papers for more detailed explanations.
Since the qubit is the elementary module of a quantum computing system (Fyrigos et al., 2018) it has been suggested that the first step towards a memristor-based one is to model the representation of a qubit by memristors. They suggested a model where the state of the qubit was mapped to a 3-D memristance space. Starting out with the representation of a qubit
The quantum state
Using the Bloch sphere and its coordinate system, Equation 2 can be represented as Equation 3:
where
Where we now have mapped three factors of a “qubit’s basis state (one for the real part of each state and one for the imaginary part of
A group of three memristors can be used to represent the qubit state, which, using Equation 5, becomes Equation 6:
In other words, each line in Equation 5 represents one dimension in a 3D space that represents a qubit Bloch sphere. For each dimension one memristor is needed, so we need three memristors to simulate one qubit, and the simulation requires that each memristor is presented with the voltage input specified in Equation 5 (see corresponding Figure 4 in the Results section). For further details on the mathematics behind this, see Karafyllidis et al. (2019).
1.4 Aim of the study
Here we draw on recent work that suggests that: A) slime molds (sometimes) behave like memristors. B) (simulated) memristors can be used to simulate quantum entanglement. When A and B are true, it follows that slime molds can be used to simulate quantum entanglement and thus a key feature of quantum computers.
The first aim of the study was to examine the claim that slime molds are indeed behaving like memristors, as there seem to be conflicting statements in the literature, and if they do, if they could simulate a quantum entanglement?
The second aim was to create a replacement circuit to describe the electric behavior of slime molds and discuss other potential bioelectric uses of P. polycephalum.
2 Methodology
2.1 Cultivation of Physarum polycephalum
In the cultivation of P. polycephalum for our experiments, we initiated the process by establishing a robust stock culture in 55 mm Petri dishes. Subsequently, new cultures were systematically developed in 50 mL conical centrifuge tubes. The cultures in the Petri-dishes and the tubes were sealed in sterile polypropylene bags with 0.2
2.1.1 Culture setup
To provide the necessary humidity and nutrients for growth, each tube was filled with a nutrient-free agar medium, sterilized oat flakes placed on top of it. The oat flakes were sterilized for 30 min at 120 °C in a Lacor 22 L pressure cooker. The design involved the inclusion of a 3 mm aperture positioned at the 32.5 mL markings within each centrifuge tube. This orifice functioned as a designated site for the integration of an electro-conductive collar, in parts modeled after the design of Braund and Miranda (2017).
2.1.2 Electro-conductive collar design and 3D printing
The electro-conductive collar, with an outer diameter of 4 mm and an inner diameter of 1.5 mm, was designed and 3D printed using a Prusa MK3 printer with the RepRapper Conductive PLA.
2.1.3 PVC tube connection setup
The experimental setup further involved the connection of two centrifuge tubes through a PVC tube with a length of 15 mm, to facilitate the growth of the slime mold through the PVC tube, connecting the two electro-conductive collars (adapters). These collars were linked to electrodes, allowing for the application of electric currents through the plasmodium, see Figure 2.

Figure 2. The slime mold measurements in the test tube set up. Overall view and details of the tube with the black electric conductive rings.
2.2 Electric measurements of Physarum polycephalum
2.2.1 Electrode connection and current measurement
To assess the response of P. polycephalum to electric currents, we utilized a Rigol DG1022 function generator to generate the currents under investigation. Simultaneous measurement of both volts and amperes became a crucial aspect of our methodology. This approach was deemed indispensable because of the sensitivity of the experimental system; even minimal imprecision in the alignment of measuring points could result in misleading data, potentially yielding erroneous hysteresis curves (Schmidt et al., 2025). The slime mold was seeded in one of the two vertical containers, with food placed in the other one. The slime grew to the other container via the horizontal transparent tube that was connected to the black conductive contacts where measurements were carried out. In the beginning the slime mold grew like a net (see Figure 2 upper right) but after some days the different strands consolidated to a single tube (see Figure 2 lower right).
2.2.2 Measurement instrumentation
For the simultaneous measurement/calculation of volts and amperes, we employed the Diligent Analog Discovery 3 in conjunction with a measuring circuit fitted onto a breadboard. Analog Discovery 3 allows for the synchronous measurement of 2 separate Voltage channels. In addition to measuring the applied Voltage from the Rigol DG1022 function generator, the Ampere values were calculated by measuring the Voltage at a serial resistance measured at 150 K Ω, using Ohm’s law we calculated I=U/R. This setup allowed us to precisely capture and record the electrical responses of P. polycephalum during the experiments, using the Digilent Waveform3 software as a logging device. Additionally, we conducted a second A control measurement using a Rigol DM3056e digital multimeter (which had a different measurement timing and was only used to control the resulting values of the Diligent Analog Discovery 3) (Schmidt et al., 2025). The Analog Discovery 3 was regularly calibrated according to the manufacturer’s calibration guidelines (Arduino, 2023). These calibration procedures ensured stable and reproducible results throughout the experimental period.
2.2.3 Environmental conditions and slime mold viability
We report laboratory conditions based on a few separate measurements. These measurements indicated an average room temperature of 23,6 °C and an average relative humidity of 44,5%, measured using a DHT22 sensor connected to an Arduino Uno R4 (Arduino, 2023; SparkFun, 2010).
Slime mold viability was not used as an exclusion criterion during the experiments; we measured all available cultures, including those showing signs of dormancy (sclerotia formation) or reduced activity. However, for a dedicated control group measurement, we used cultures that had been cultivated on 3 April 2024 and re-measured on 19 August 2025 (503 days), by which time they were most likely dead, as indicated by their sclerotic appearance.
The study consisted of two main segments of measurements that included slime mold cultures. The main measurements, conducted from 25 January 2024, to 3 April 2024, and included 289 observations across 38 distinct slime mold cultures. Prior to this, the test measurements occurred over 14 days between 2 June 2023, and 22 December 2023, involving 195 observations from over 20 different cultures, including 44 controls such as office plants, cacti, and dummy resistors. Additional 23 comparative measurements were done from August 19-20, 2025, of which 4 were with slime molds, resulting in a total of 293 slime mold measurements. The living slime mold cultures ranged in age from 4 to 25 days, providing a fairly large dataset for analysis. To assess baseline electrical behavior of non-biological and non-Physarum samples and to identify possible setup artifacts, we performed a set of control and comparison measurements. These included distilled water (VDE 0510 compliant), Viennese tap water, and a 1% NaCl solution prepared from 100 g distilled water (VDE 0510) and 1 g NaCl. We also tested an empty tube from our setup (PVC tube with 3D-printed adapters made from RepRapper conductive PLA) as well as the same tube filled with the 1% NaCl solution.
Additional biological controls included a Trichocereus macrogonus var. Pachanoi cactus, a basil plant (Ocimum basilicum), hard cheese and vegan coconut yoghurt. These measurements were acquired with the same instrumentation and wiring as used for the P. polycephalum experiments, employing identical sinus waveforms and logging procedures, allowing direct comparison of signal levels and potential hysteresis-like artifacts across biological, non-biological, and hardware-only baselines. See Table 1 and for more details, Table 2.
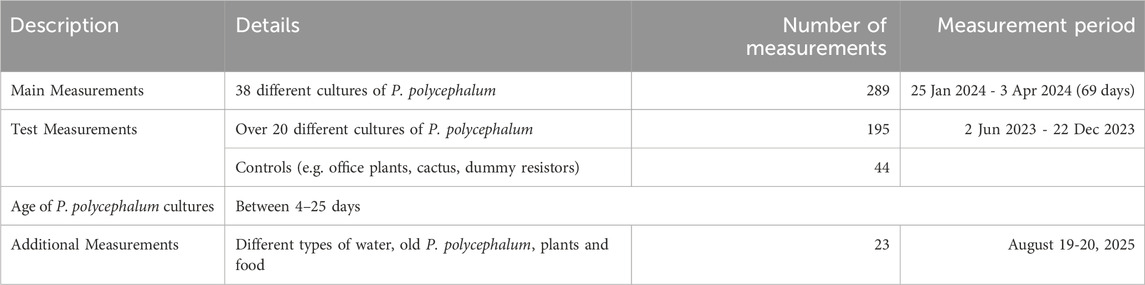
Table 1. Summary of electric measurements with Physarum polycephalum.
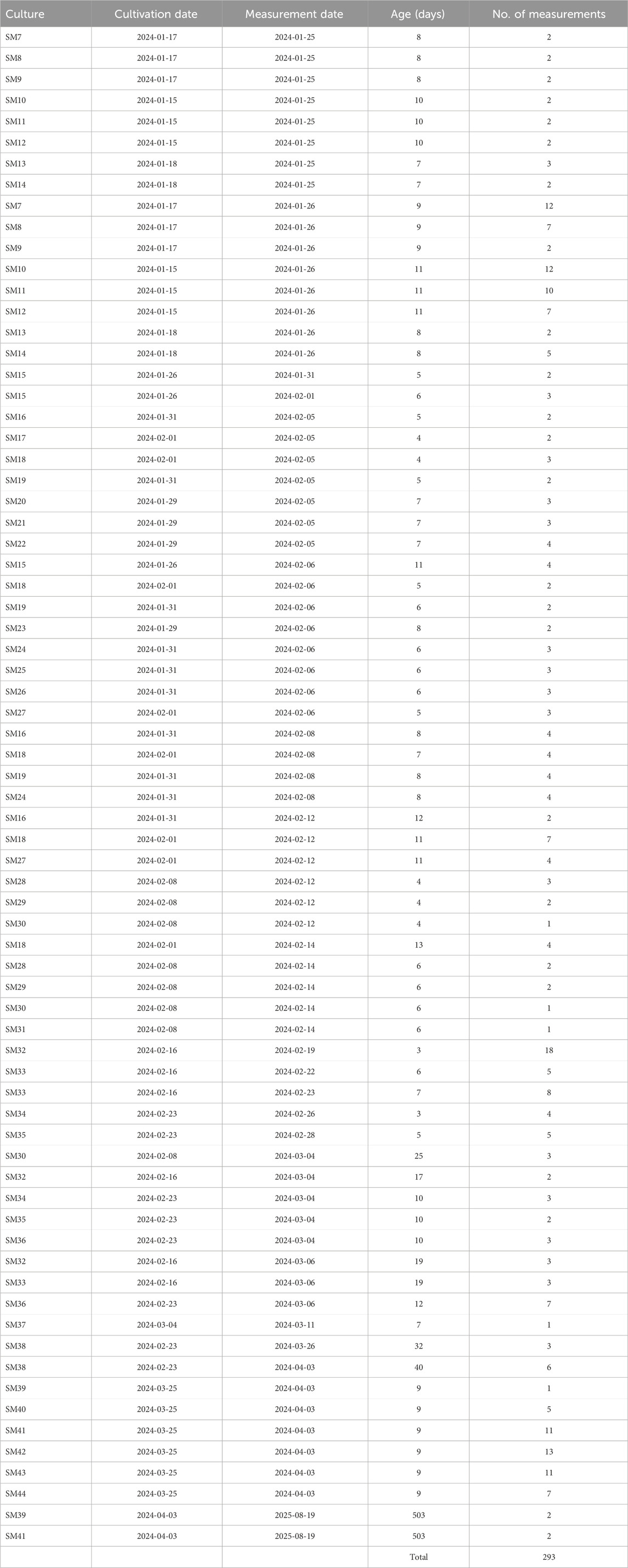
Table 2. Details of electric slime mold measurements.
The datasets generated and analyzed during the current study are available in the Mendeley repository: https://data.mendeley.com/datasets/ghm9gvwwvb/1.
2.3 Electric measurements of physical memristor
As a comparison, we conducted the experiments not only with the slime molds but also with a real physical memristor, the Knowm Inc. 1 × 16 array memristor chip.
In our investigation, we used the Memristor Discovery platform, consisting of the Memristor Discovery board v.2 integrated with the Diligent Digilent Analog Discovery 3 for data collection and analyses.
After connecting the Knowm 1 × 16 array memristor chip onto the Memristor Discovery board we conducted standard experiments provided by the Memristor Discovery software to familiarize ourselves with the setup and operation of the system.
The open-source nature and available documentation of the Memristor Discovery software, allowed tinkering the experimental parameters to match those used in our previous slime mold experiments, by changing the code of the software in the Eclipse, version 4.30, development environment. Specifically, we adjusted the waveform characteristics to obtain the three different input signals shown in 5.
We documented the graphical outputs using the save function to record pictures of the behavior of the memristors using our parameters. Additionally, the Memristor Discovery software allowed the export of numerical data in the form of. csv spreadsheets which were used for further analysis.
2.4 Software based electric circuit simulation
Additionally, we conducted experiments using software simulations of memristors using the LTspice software. This simulation environment enabled the incorporation of various memristor models into its library, allowing modeling different memristor characteristics. Within the LTspice software, MacOs version 17.2.2, we constructed a virtual version of our experimental circuit, mirroring the setup utilized in our slime mold experiments. This virtual representation allowed us to simulate the behavior of memristors under conditions like those encountered in our physical experiments. We generated the same three input signals as in the previous experiments using. pwl files. We conducted simulations to observe the response of the virtual memristors, to compare the electrical behavior of memristors and the physiological responses of slime molds. Documentation was retrieved using the data export function within the LTspice software (Schmidt et al., 2025).
2.5 Data processing
2.5.1 Data inclusion and file parsing
CSV files were ingested from the experimental repository. We analyzed only files that met two criteria recorded in an external metadata table (Excel): (i) the waveform was a sinus, and (ii) the total length of the recorded measurement was
2.5.2 Robust de-spiking and zero-phase smoothing
To filter artifacts while preserving the phase relationship between
1. Hampel filter (robust de-spiker). Outliers were replaced by the local median using a Hampel identifier with half-window
2. Savitzky–Golay smoothing (zero-phase). We smoothed
2.5.3 Feature extraction
All features were computed on the conditioned signals.
2.5.4 Metric-wise decisions and final classification
For interpretability, each metric was mapped to a qualitative label (RC-like, memristor-like, ambiguous/mixed) using empirically set thresholds. The final label was derived from the following four criteria: PCA_ecc, Ellipse_RMSE, Pinch_Ratio, and Width@V0. A simple majority of consistent votes (at least 2 agreeing and no conflicting majority) yielded the final class; otherwise the label was ambiguous/mixed.
2.5.5 Thresholds used in this study
The following thresholds were used in our analysis, see Table 2.
2.5.6 Quality controls and edge cases
Features that could not be computed robustly (e.g., no
3 Results
3.1 Are slime molds memristors and can they simulate quantum entanglement?
When applying the three different voltage sources described in Equation 5 to: A) the LTSpice Memristor simulation, B) the physical Knowm memristor and C) the slime molds (2 examples shown here: slime mold nr. 36 and slime mold nr.32) we measured the currents as shown in Figure 3. The data shown in Figure 3 can also be represented as the corresponding I-V-diagrams, as shown in Figure 4. While the simulated memristor shows the expected inclined “figure eight” shape which is pinched at the 0-0 values (for voltage and current), the Knowm memristor as well as the slime molds produce different I-V-diagrams. In the case of the Knowm memristor a “figure eight” might only be recognized for positive voltages, in which case the curve starts with a high memristance (resistance) and above a certain threshold, the voltage switches to a low memristance state that is maintained until the voltage reverts to 0 (or into the negatives). No such behavior is observable in the negative voltage range, where a high memristance is observed all the way through. The slime molds are still different, as most slime molds tested do not produce a pinched hysteresis curve, but an open ellipsoid shape (see slime mold 36 in Figure 4). In some cases, an ellipsoid shape was observed but with more pointed ends at a slightly different angle (see slime mold 32 in Figure 4). Both slime molds shown here do not exhibit a dominant memristive characteristic.
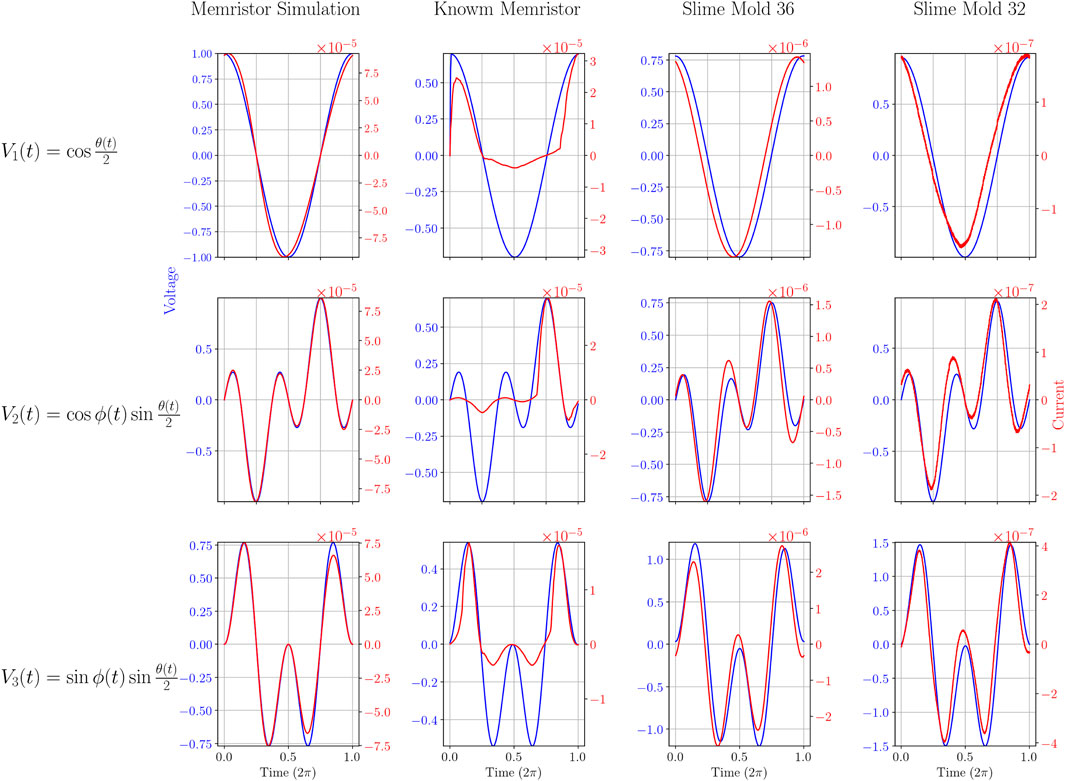
Figure 3. Voltage and Current curves over time of the four different experiments for each of the three different input signals. x- axis: time from 0 to
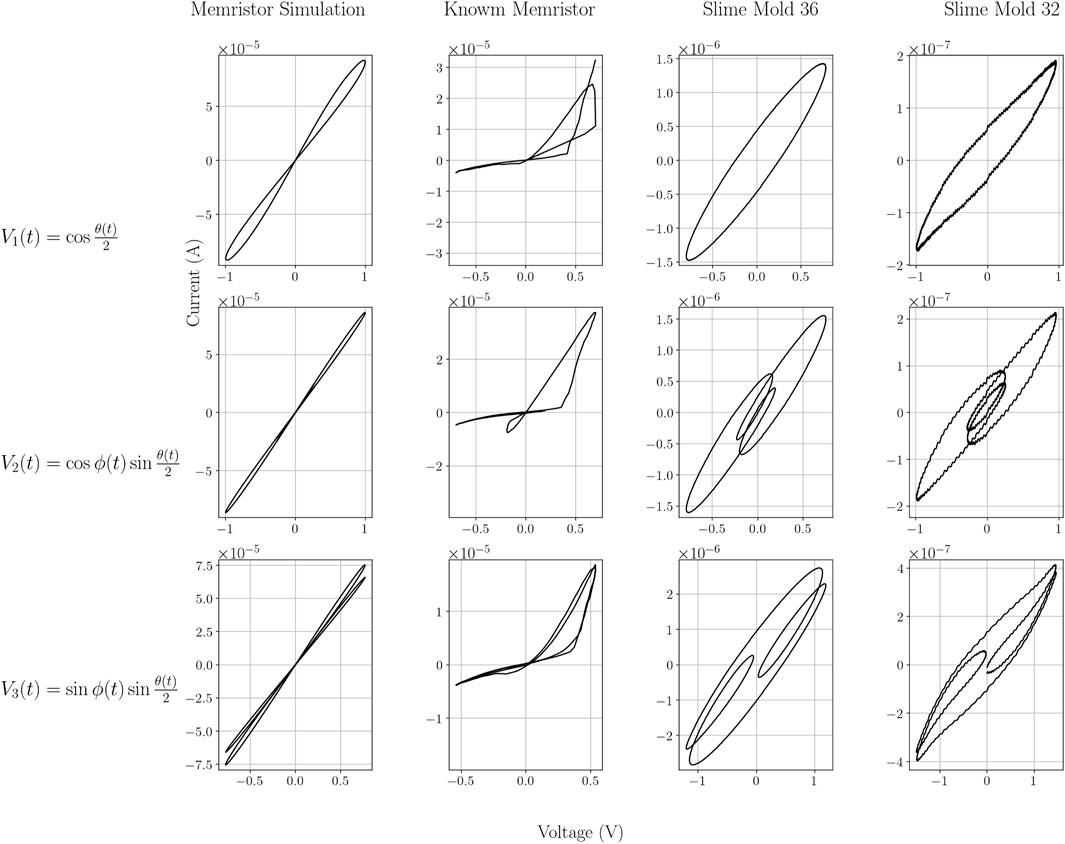
Figure 4. Current-voltage characteristic of the four different experiments for each of the three different input signals. x- axis: voltage V, y-axis: current
For the slime molds, the curves differ from cycle to cycle. The biggest shift is from the first to the consecutive cycles, as the I-V curve starts at (or close to) the 0-0 point. Within the first cycle the curve then adopts a shape that is repeated for future cycles, although a slight change can be observed within each cycle, see Figure 5. In our analysis, the first cycles were never used, but the second.
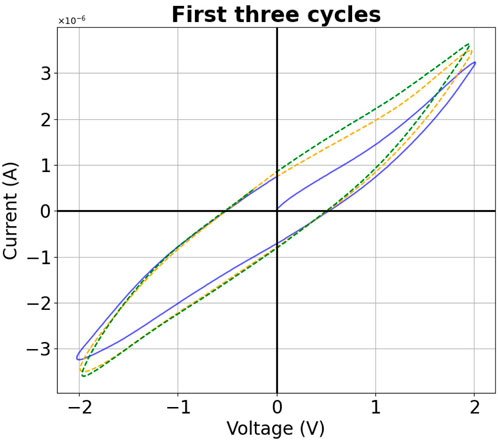
Figure 5. Color coded first three cycles of a typical slime mold measurement.
Using the data shown in Figure 4, applying Ohm’s law R = U/I results in the memristance/resistance curves shown in Figure 6. Althoughthe model (simulation) exhibits a variation of the resistance between a (positive) lower
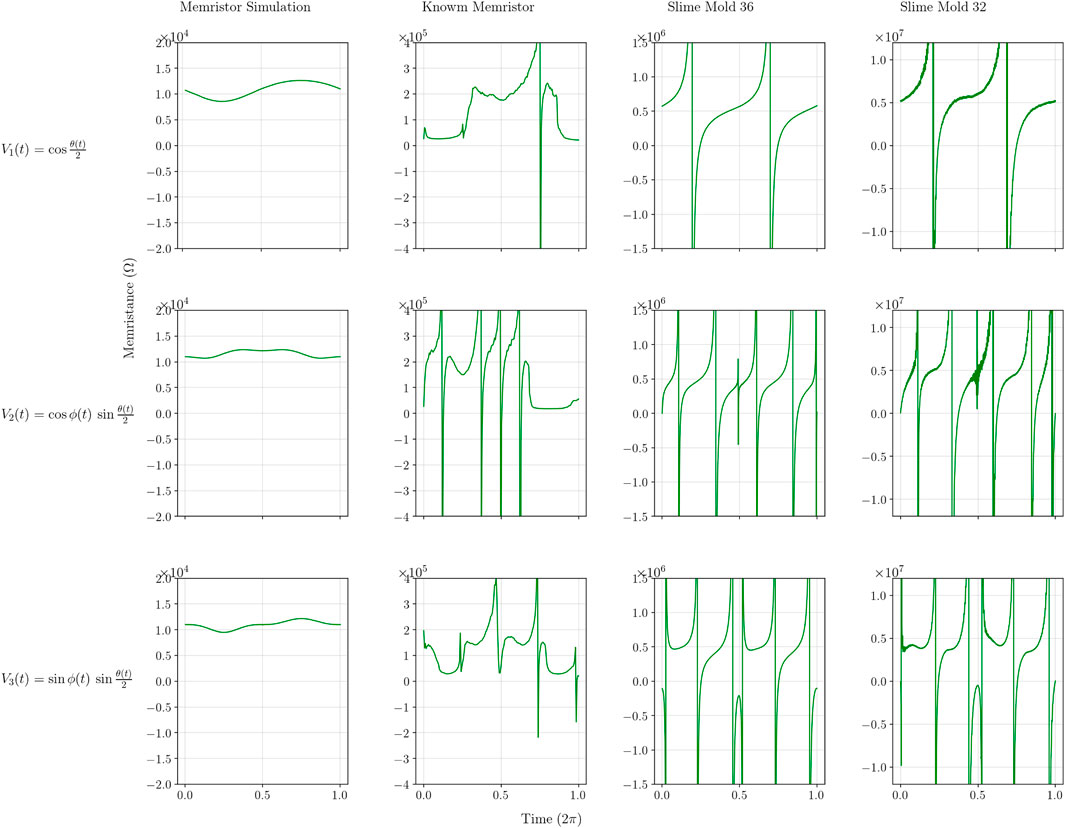
Figure 6. Memristance (Ohm) curves of the four different experiments for each of the three different input signals.
Neither the physical nor the biological components behave like ideal memristors due: a) to asymmetry between positive and negative voltages and small parasitic capacitance (physical component); and (b) no or small memristive effect and capacitance (biological component). The physical component, the Knowm memristor curve appears less smooth compared to the memristor simulation curve. The two biological component curves appear even less smooth. They exhibit sharp changes in resistance/impedance and several fluctuations even reaching negative values, indicating a behavior unlike the memristor simulation.
3.2 Slime mold electric characterization over a wider voltage and frequency range
We tested a wider range of voltages and frequencies as input signals to the slime molds to find out if a memristance would appear. We applied up to 11 different sinus inputs between 5 mV and 2 V. None of the Voltages tested resulted in pinched “8” shaped curves, all were elliptical. Results with lower voltages showed two specific outcomes. First, the slime molds themselves produce a low electric output, in these cases approximately 20 mV (or −20 mV depending on the polarization) and less than 10
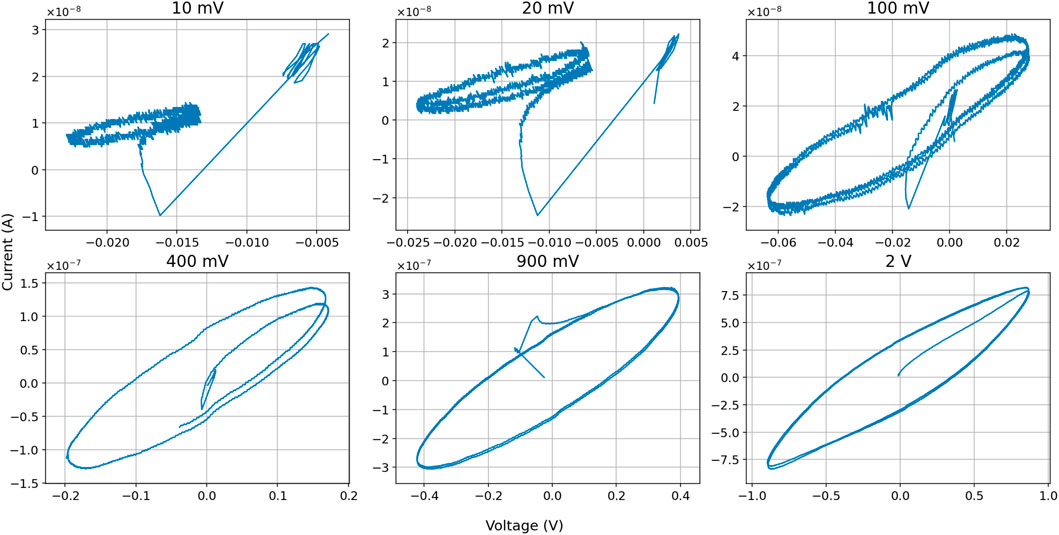
Figure 7. I-V diagrams of different sinus input voltages, including first cycle. Example shows results in slime mold nr. 43.
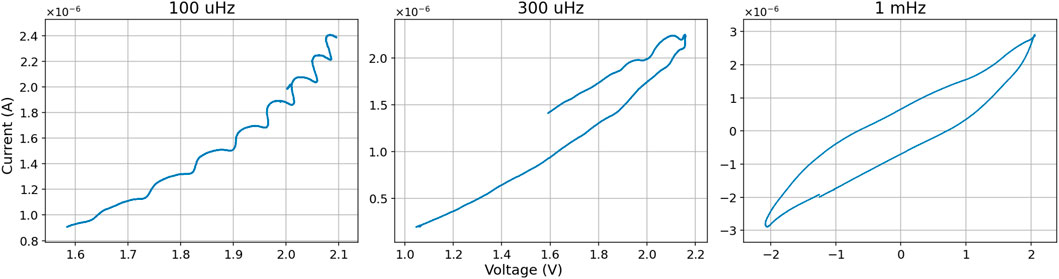
Figure 8. I-V diagrams of different very low sinus input frequencies. Example shows results in slime mold nr. 35.
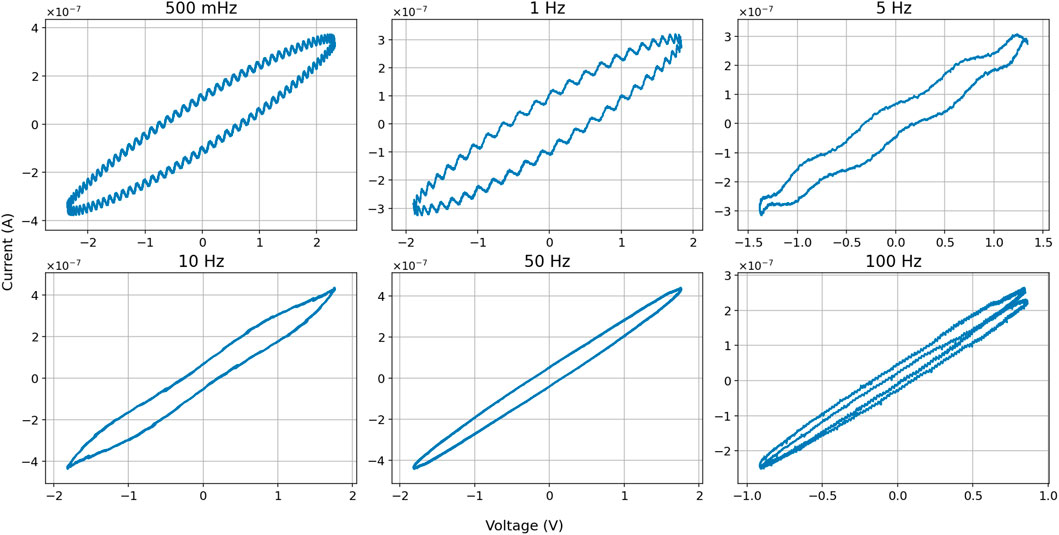
Figure 9. I-V diagrams of different “higher” sinus input frequencies. Example shows results in slime mold nr. 32.
Finally, we also measured the voltage and current of seven different slime molds with no external input. These measurements show that there is a small intrinsic electrical output of the slime molds that may change over time. In some cases (e.g. SM30 and SM32) the measured voltage was below 10 mV and did not change much over time). In another case (SM34) the output was relatively stable below −2 mV but had some periodicity (about 10 cycles in 1,000 s). Yet other measurements (e.g. SM18, SM35 and SM36) started with a higher voltage up to an absolute value of 159 mV and gradually moved towards a value closer to zero. SM35 also showed periodicity on top of the discharge, with about 8 cycles in 1,000 s, see Figure 10.
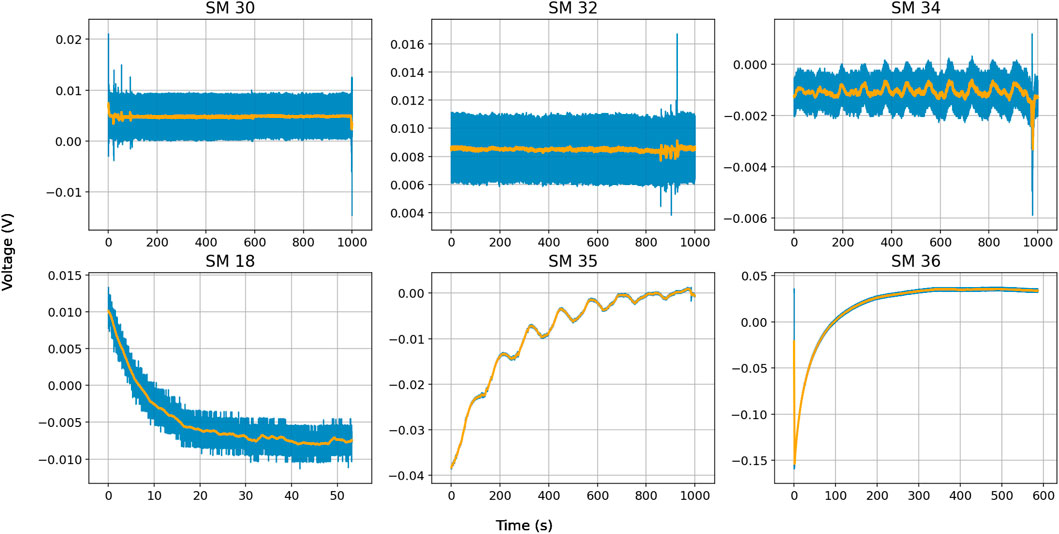
Figure 10. Slime mold electric output without external inputs. Blue line represents actual measurements, orange line is a moving average (N = 41).
3.3 I-V curve shape analysis
We analyzed all I-V measurements with a sinus input and a length below 1,000 s to find out whether they rather resemble an elliptical curve shape or a pinched figure eight. While the first points towards an RC like characteristic, the latter one is a clear and indicative sign of a memristor.
We analyzed
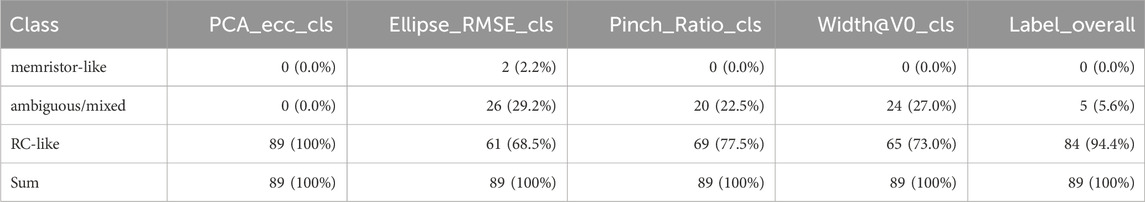
Table 3. Per-metric and overall classification counts across the analyzed dataset
At the shape level, PCA_ecc indicated RC-consistent, elongated ellipses for all traces (89/89, 100%). The ellipse-fit residual classified 61 traces as RC-like (68.5%), 26 as ambiguous (29.2%), and 2 as memristor-like (2.2%). Pinch-based evidence identified 69 RC-like (77.5%), 20 ambiguous (22.5%), and 0 memristor-like (0.0%). The zero-voltage width metric labeled 65 RC-like (73.0%), 24 ambiguous (27.0%), and 0 memristor-like (0.0%). Aggregating the four criteria by simple majority yielded an overall RC-like label for 84 traces (94.4%), with the remainder 5 (5.6%) ambiguous; no trace reached a memristor-like majority. Ambiguous outcomes typically reflected disagreement between pinch/width (weak memristive signatures) and ellipse-based criteria, or edge cases with scarce
Given the results of slime molds do not show memristive behavior, no further attempts were made to simulate a quantum entanglement with these components. The artifacts, including large positive and negative values up to infinity impeded further steps.
3.4 Inactive desiccated slime mold and other measurements
We carried out measurements with two sinus frequencies (7.76 mHz and 23,39 mHz) and an amplitude of 4 V (plus/minus 2 V) in two slime molds that were already 1 year 3 months and 15 days inside their test tube container. Visually the had turned brown (initially yellow/white) and appeared desiccated. The resulting I-V curves. These old slime molds produced curves that were not dissimilar to recently cultivated ones, see Figure 11.
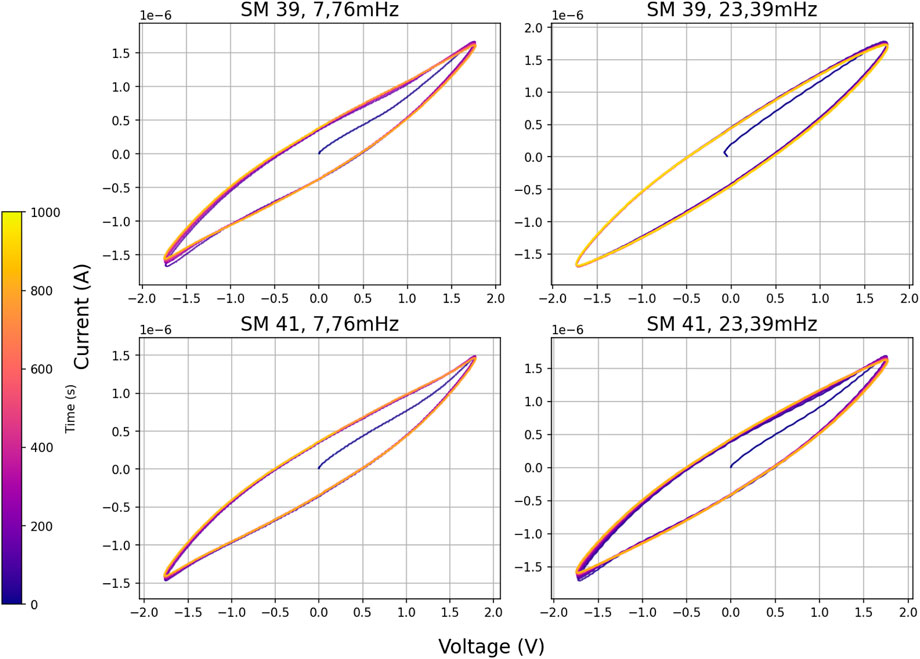
Figure 11. I-V diagrams of old slime molds >15 months, measured over 1,000 s.
Measurement with two frequencies (7.76 mHz and 23,39 mHz) for different types of water (destilled, Viennese tap water, 1% NaCl, were measured in a plastic container with electrodes 5 cm aparts, and 1% NaCl in the same test tube were we measured the slime molds), plants (cactus and basil) and food (hard cheese and yoghurt) were carried out as a comparison to the slime molds, see Figure 12. In the water samples, the degree of ions/salt in the water changes the curves significantly. Destilled water showed two pinched points in their loop, salt water was an open curve, Viennese tap water was a relatively narrow open curve, and salt water in the test tube was also more narrow than in the wider plastic container. In the plants, basil resulted in open curves, as did the cactus but which had a more pronounced non-linearity. The two tested food objects were somewhere in the middle between salt water and the cactus.
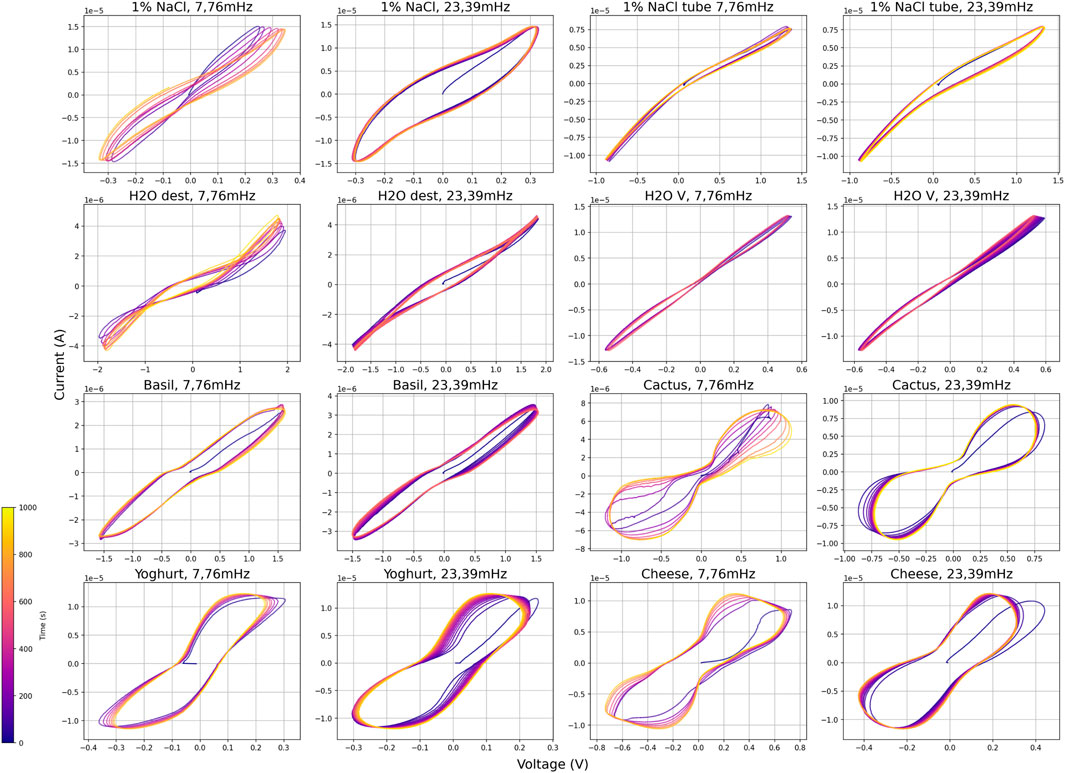
Figure 12. Set of I-V diagrams of comparative measurements, 1,000 s duration.
3.5 A replacement circuit for slime molds and alternative potential bioelectric uses of Physarum polycephalum
We modeled several potential replacement circuits of the slime mold(s) in LT Spice to generate a similar output as shown in Figure 4. The most similar results were found in the circuit with a capacitor and a resistor in series followed by a parallel capacitor and resistor. Further parametric refinement of this promising replacement circuit yielded very similar results in LT Spice as measured in the slime mold (SM36). The replacement circuit is shown in Figure 13 and the two corresponding I-V curve sets in Figure 14. The slime mold is not a totally passive device. A voltage meter can measure a voltage between 20 and 40 mV when connected directly to the slime mold. The measured voltage reduced slightly during the measurement.
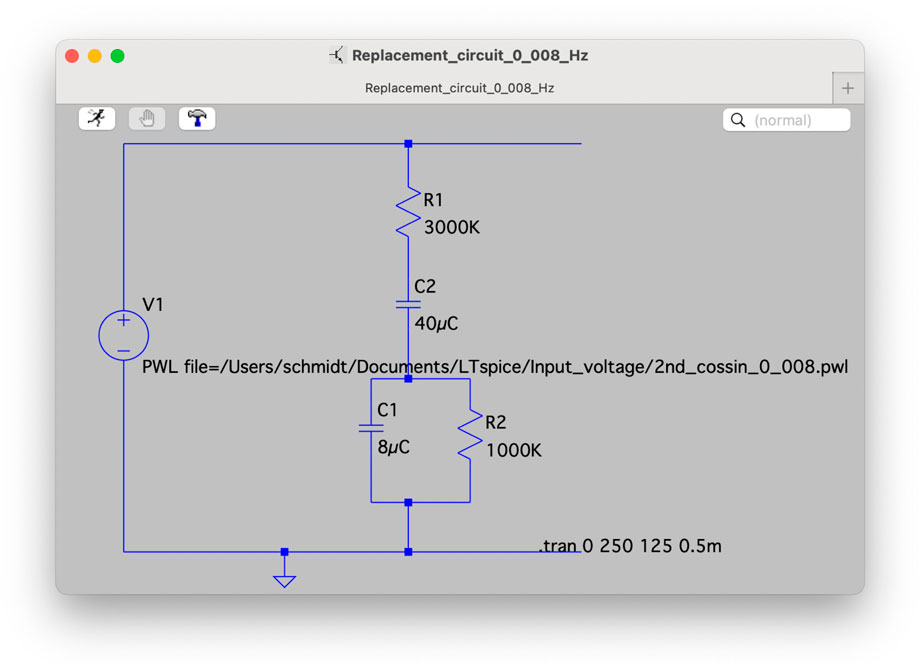
Figure 13. Final slime mold replacement circuit containing only resistors and capacitors (Schmidt et al., 2025)
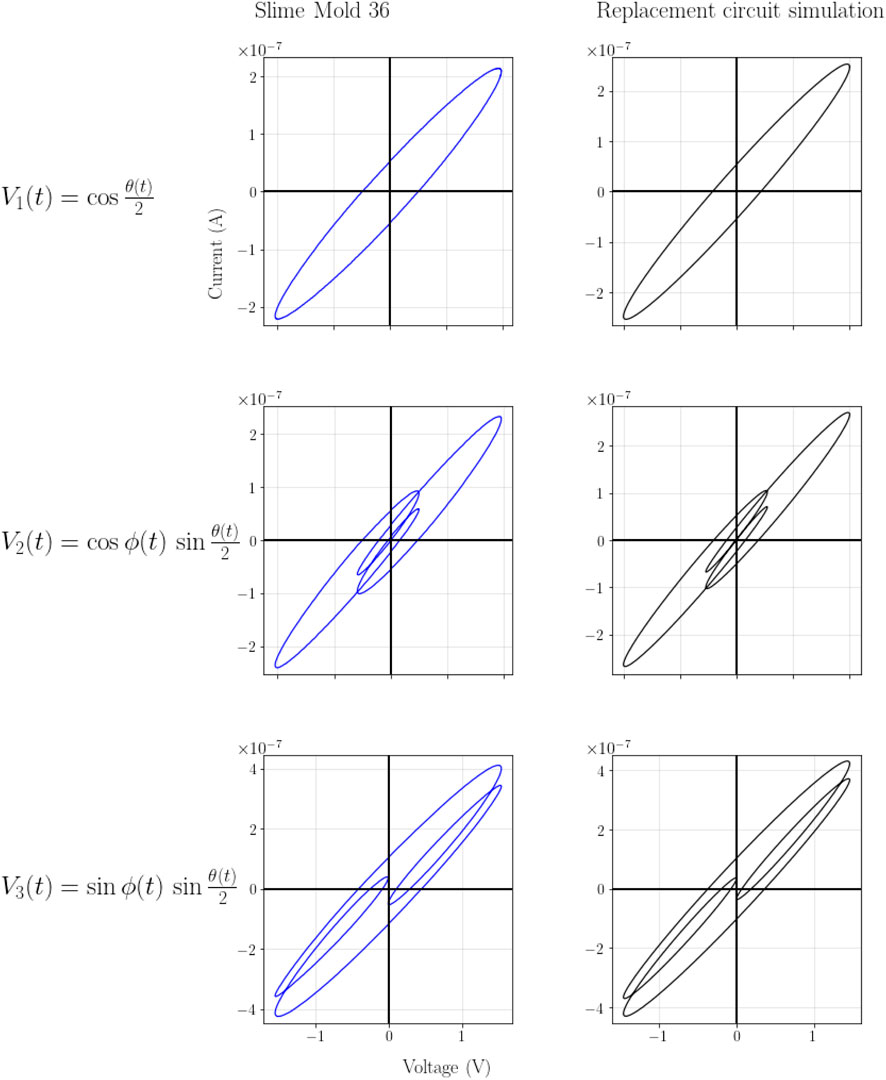
Figure 14. Comparison of the three resulting IV curves between slime mold and the replacement ciurcuit.
Using the slime mold as a “pseudo-capacitor” we applied it within some electronic circuits such as a low and high pass filter. In a low-pass filter, the cutoff frequency and level of attenuation naturally depend on the leading resistor. If we choose the resistor as
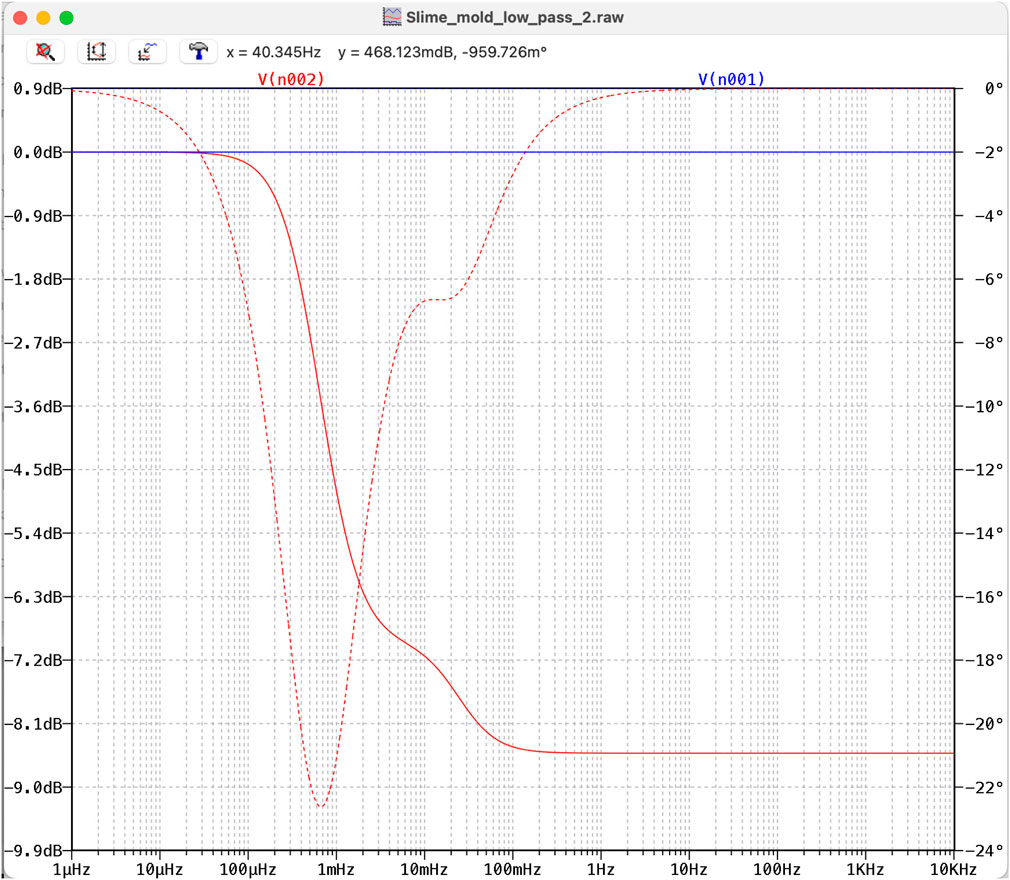
Figure 15. Bode diagram of a low pass filter with a slime mold instead of a capacitor. Red line: attenuation); red dotted line: phase shift.
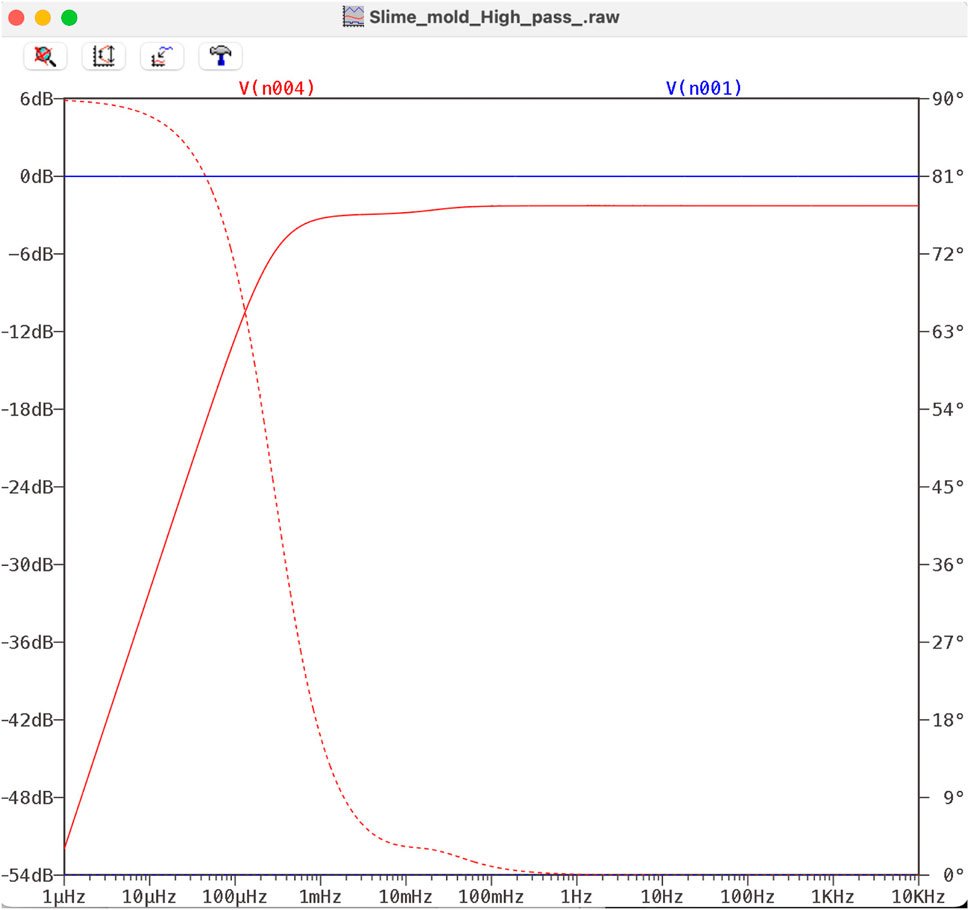
Figure 16. Bode diagram of a high pass filter with a slime mold instead of a capacitor. Red line: attenuation); red dotted line: phase shift.
The red dotted lines in Figures 15, 16 show the phase shift of these particular examples, which indicates that slime molds may also be deployed in phase shift circuits.
4 Discussion
4.1 Absence of pinched hysteresis in slime molds under tested conditions
The potential for bio-inspired materials, such as slime molds, to behave like a memristor would be of great interest due to their natural adaptability and self-organizing properties.
To find out whether or not slime molds behave like memristors, we followed the experimental works reported in (Gale et al., 2014; Braund and Miranda, 2017; Miranda and Braund, 2017; Adamatzky, 2015a). These works suggested that slime mold can behave like a memristor: a passive two-terminal electronic devices which resistance depends on the history of voltage and current that has passed through them, resulting in a distinctive pinched hysteresis loop in their I-V characteristics. However, our repeated measurements, showed that in our set-up, the slime mold does not behave like a memristor. Not a single slime mold produced a pinched hysteresis curve. While some slime mold experiments resulted in slightly S-shaped curves (see slime mold B), all of them were rather elliptical (with sinus or cosines inputs). The elliptical shape, we believe, is caused by a capacitance that is created by the cell membrane and internal cell fluids that act as electrolytes. This is comparable to the electric behavior in plants. When electrodes are inserted into a plant, they interact with the plant’s internal ionic fluids and cellular structures, creating a measurable capacitance. This phenomenon occurs because plant tissues contain conductive ionic fluids, such as sap, which facilitate the movement of electrical charges (Gao et al., 2024). Additionally, the cell membranes within the plant act as dielectric barriers, separating different conductive regions within the tissue. These biological components form an equivalent parallel resistance-capacitance circuit when connected to electrodes, allowing for the storage and separation of charges. As a result, the electrical capacitance of the system should never be zero. The capacitance measured in a plant-electrode system is influenced by various factors, including the plant’s moisture content, the type of plant tissue, and the positioning of the electrodes. For instance, a study by Dalton (1995) demonstrates that the presence of biological fluids and cell membranes ensures that the capacitance value is always non-zero, reflecting the plant’s intrinsic electrical properties.
These effects prevent the curve from passing through the origin (0,0), indicating that it might not be possible to simulate a quantum computer in a simple setup, where just electrodes are implanted without further mitigation. The slime mold is not a plant, of course, and one of its unique features is that it lacks internal cell membranes, which means that it appears like a single cell organism (with thousands of cell nuclei). This specific unique internal Bauplan, however, does not change the fact that organisms in general are (also) capacitors. See Atkinson et al. (2023) for an overview of biological capacitors in bioelectronics.
Our results showed, in all of the datasets, elliptical I-V relationships. A slight saddle, seen in some of the measurements, suggests some non-linear behavior, but not the classic hysteresis loop of a memristor. The absence of a pinched hysteresis loop suggested only weak or no memristive behavior under the tested conditions.
Why are our findings different from those reported in the literature? First, the reported memristive effect does not seem to appear in all experiments. Gale et al. Gale et al. (2016), for example wrote: “Of the eleven samples in batch 1, two exhibited good memristance curves (as in they had pinched hysteresis), the other nine exhibited open curves” and “For batch 2, which were measured at a larger voltage range (between
4.2 Slime mold as device for other electrical circuits
The replacement circuit we created (see Figure 13) consists entirely of R and C components, and is similar, although not identical, to plant equivalent electrical circuits outlined by Van Haeverbeke et al. (2023)(see their Table 4). An additional 10-20 mV voltage source that (may) fall off continuously seems a good description of the slime molds without the nonlinear “saddles” by first approximation. Although these circuits do not represent or contain memristors, they might still be used as sub-circuits for various other types of circuits. The configuration suggests it could be useful in a number of circuits, for example a Low-Pass Filter Circuit. A low-pass filter allows low-frequency signals to pass while attenuating high-frequency signals. Usually such a subcircuit could be part of audio circuits, analog-to-digital conversion stages, or any application where it is necessary to remove high-frequency noise. Like the Low-Pass filter, the voltage divider with frequency dependence, a circuit that may serve as a frequency-dependent voltage divider, where the output voltage depends on the input frequency. This circuit can be used in circuits where frequency-based signal conditioning is required. Another option to implement the slime mold would be a Phase Shift Network that could be used to introduce a phase shift in a signal, depending on how the resistor and slime molds are connected. Phase shift networks are used in phase-shift oscillators and certain types of filters. Similarly to the low pass is the high pass filter circuit that allows high frequencies to pass while blocking low frequencies. In principle, the slime mold could also be used in several other circuits, such as RC Integrator Circuit, a RC Differentiator Circuit, a RC Time Constant, or a Snubber Circuit. Previous work has also shown that slime molds can improve microbial fuel cells when applied to the cathode side Taylor et al. (2015). This and other potential bioelectric applications seem interesting for future bioelectric deployment of the slime mold, despite it not showing memristive behavior.
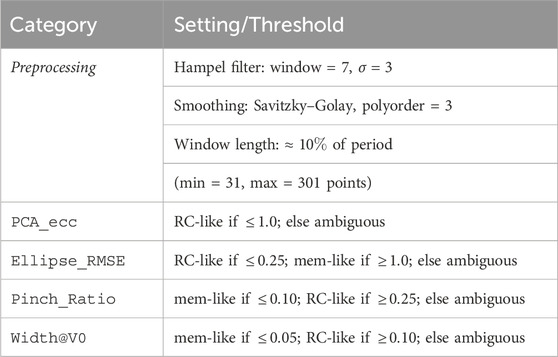
Table 4. Preprocessing settings and heuristic thresholds used for classification. Thresholds define ranges for memristor-like, RC-like, or ambiguous labels per metric.
5 Conclusion
Simulating quantum entanglement with an ideal memristor model has been demonstrated in the literature. In this work we intended to carry out this simulation using simulated memristor models, real physical/electric (Knowm Inc. memristor) and slime molds (Physarum polycephalum). Our experiments demonstrate that the biological component, Physarum polycephalum which has been described in the literature to exhibit memristive behavior turned out not to show any (significant) memristive characteristics due to its inherent capacitance. While some publications claim that slime mold could have memristive or memcapacitive characteristics, neither were detected in our experiments. Instead, we showed that slime molds could very well be described, by first approximation, with replacement circuits consisting only of resistors and capacitors. Some of the slime molds did show a slight non-linearity when going over a certain voltage threshold, but none could be characterized clearly as a memristor or a memcapacitor.
The inherent capacitance of slime mold (that is also seen in other organisms (Van Haeverbeke et al., 2023), seems to be based on the cytoplasma electrolyte, and is a characteristic that is practically impossible to change. Thus, according to our results, any electric circuit requiring a memristive behavior cannot be designed with slime molds alone. The inherent capacitance of the slime mold defies the claim that slime molds are (bio-)memristors. It remains to be seen if future experiments will identify a methodology to grow slime molds without a capacitance.
While slime molds are not memristors they can be - in a first approximation - described with an adequate replacement circuit consisting of resistors and capacitors. This RC replacement network is versatile and could be integrated as a sub-circuit in various analog circuits, including high- and low-pass filters, timing circuits, signal processing units, and phase shift networks. While slime molds could not be used to simulate quantum entanglement, they may still have a future as bioelectronic components for different types of applications.
Data availability statement
The datasets presented in this study can be found in the Mendeley repository: https://data.mendeley.com/datasets/ghm9gvwwvb/1.
Author contributions
MS: Conceptualization, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review and editing. GS: Data curation, Formal Analysis, Methodology, Software, Visualization, Writing – original draft, Writing – review and editing. UR: Investigation, Methodology, Writing – review and editing. ZS: Formal Analysis, Validation, Writing – review and editing. EM: Conceptualization, Investigation, Methodology, Resources, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. MS, GS, and UR acknowledge funding from the European Union “Mi-Hy” Project 101114746 and KIT funding for “Kunst und Wissenschaft: kreative Stoerung, Innovationsmotor oder Aesthetisierung der Forschung? - Neue Herausforderung fuer die Technikfolgeabschätzung (TA)”.
Acknowledgments
We would also like to thank Satvik Venkatesh and Andrew Adamatzky for helpful feedback on their previous slime mold electronic measurements and memfractance.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. Generative AI was used to aid in the preparation of python scripts to analyse the data. Careful checking of the code and results were done manually to confirm its validity.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aaronson, S., and Gottesman, D. (2004). Improved simulation of stabilizer circuits. Phys. Rev. A 70, 052328. doi:10.1103/PhysRevA.70.052328
Abdelouahab, M.-S., Lozi, R., and Chua, L. (2014). Memfractance: a mathematical paradigm for circuit elements with memory. Int. J. Bifurcation Chaos 24, 1430023. doi:10.1142/s0218127414300237
Adamatzky, A. (2014). Slime mould electronic oscillators. Microelectron. Eng. 124, 58–65. doi:10.1016/j.mee.2014.04.022
Adamatzky, A. (2015a). Slime mould processors, logic gates and sensors. Philos. Trans. A Math. Phys. Eng. Sci. 373, 20140216. doi:10.1098/rsta.2014.0216
Adamatzky, A. (2015b). Twenty-five uses of slime mould in electronics and computing - survey. Int. J. Unconv. Comput. 11, 449–471.
Adamatzky, A., and Jones, J. (2010). Road planning with slime mould: if physarum built motorways it would route m6/m74 through newcastle. Int. J. Bifurcation Chaos 20, 3065–3084. doi:10.1142/S0218127410027568
Adamatzky, A., and Jones, J. (2011). On electrical correlates of physarum polycephalum spatial activity: can we see physarum machine in the dark? Biophysical Rev. Lett. 06, 29–57. doi:10.1142/s1793048011001257
Aguirre, F., Sebastian, A., Le Gallo, M., Song, W., Wang, T., Yang, J. J., et al. (2024). Hardware implementation of memristor-based artificial neural networks. Nat. Commun. 15, 1974. doi:10.1038/s41467-024-45670-9
Atkinson, J. T., Chavez, M. S., Niman, C. M., and El-Naggar, M. Y. (2023). Living electronics: a catalogue of engineered living electronic components. Microb. Biotechnol. 16, 507–533. doi:10.1111/1751-7915.14171
Bonner, J. T. (2010). Brainless behavior: a myxomycete chooses a balanced diet. Proc. Natl. Acad. Sci. U. S. A. 107, 5267–5268. doi:10.1073/pnas.1000861107
Borghetti, J., Snider, G. S., Kuekes, P. J., Yang, J. J., Stewart, D. R., and Williams, R. S. (2010). Memristive switches enable stateful logic operations via material implication. Nature 464, 873–876. doi:10.1038/nature08940
Boussard, A., Fessel, A., Oettmeier, C., Briard, L., Dobereiner, H. G., and Dussutour, A. (2021). Adaptive behaviour and learning in slime moulds: the role of oscillations. Philos. Trans. R. Soc. Lond B Biol. Sci. 376, 20190757. doi:10.1098/rstb.2019.0757
Braund, E., and Miranda, E. (2017). On building practical biocomputers for real-world applications: receptacles for culturing slime mould memristors and component standardisation. J. Bionic Eng. 14, 151–162. doi:10.1016/s1672-6529(16)60386-4
Chattopadhyay, A., and Rakosi, Z. (2011). “Combinational logic synthesis for material implication,” in 2011 IEEE/IFIP 19th international conference on VLSI and system-on-chip (Hong Kong, China: ieee). doi:10.1109/VLSISoC.2011.6081665
Chua, L. (1971). Memristor: the missing circuit element. IEEE Trans. Circuit Theory 18, 507–519. doi:10.1109/tct.1971.1083337
Chua, L. (2018). How we predicted the memristor. Nat. Electron. 1, 322. doi:10.1038/s41928-018-0074-4
Dalton, E. N. (1995). In-situ root extent measurements by electrical capacitance methods. Plant Soil 173, 157–165. doi:10.1007/bf00155527
Di Ventra, M., Pershin, Y., and Chua, L. (2009). Circuit elements with memory: memristors, memcapacitors, and meminductors. Proc. IEEE 97, 1717–1724. doi:10.1109/jproc.2009.2021077
Fyrigos, I.-A., Ntinas, V., Karafyllidis, I., Sirakoulis, G., Karakolis, P., and Dimitrakisc, P. (2018). Early approach of qubit state representation with memristors.
Gale, E., Adamatzky, A., and de Lacy Costello, B. (2014). Slime mould memristors. BioNanoScience 5, 1–8. doi:10.1007/s12668-014-0156-3
Gale, E., Aadamatzky, A., and de Lacy Costello, B. (2016). On the memristive properties of slime mould. Springer 21 (5), 75–90. doi:10.1007/978-3-319-26662-6_4
Gao, C., Gu, Y., Liu, Q., Lin, W., Zhang, B., Lin, X., et al. (2024). All plant-based compact supercapacitor in living plants. Small 20, e2307400. doi:10.1002/smll.202307400
Huang, Y., Ando, T., Sebastian, A., Chang, M.-F., Yang, J., and Xia, Q. (2024). Memristor-based hardware accelerators for artificial intelligence. Nat. Rev. Electr. Eng. 1, 286–299. doi:10.1038/s44287-024-00037-6
Ielmini, D., and Milo, V. (2017). Physics-based modeling approaches of resistive switching devices for memory and in-memory computing applications. J. Comput. Electron 16, 1121–1143. doi:10.1007/s10825-017-1101-9
Jebali, F., Majumdar, A., Turck, C., Harabi, K. E., Faye, M. C., Muhr, E., et al. (2024). Powering ai at the edge: a robust, memristor-based binarized neural network with near-memory computing and miniaturized solar cell. Nat. Commun. 15, 741. doi:10.1038/s41467-024-44766-6
Karafyllidis, I., Sirakoulis, G., and Dimitrakis, P. (2018). “Representation of qubit states using 3d memristance spaces: a first step towards a memristive quantum simulator,” in Proceedings of the 14th IEEE/ACM international symposium on nanoscale architectures (Athens, Greece: NANOARCH ’18), 163–168. doi:10.1145/3232195.3232197
Karafyllidis, I., Sirakoulis, G., and Dimitrakis, P. (2019). Memristive quantum computing simulator. IEEE Trans. Nanotechnol. 18, 1015–1022. doi:10.1109/tnano.2019.2941763
Kishimoto, U. (1958). Rhythmicity in the protoplasmic streaming of a slime mold, physarum polycephalum. i. a statistical analysis of the electric potential rhythm. J. Gen. Physiol. 41, 1205–1222. doi:10.1085/jgp.41.6.1205
Lehtonen, E., Poikonen, J. H., and Laiho, M. (2010). Two memristors suffice to compute all boolean functions. Electron. Lett. 46, 230–231. doi:10.1049/el.2010.3407
Luo, L., Dong, Z., Duan, S., and Lai, C. (2020). Memristor-based stateful logic gates for multi-functional logic circuit. IET Circuits, Devices Syst. 14, 811–818. doi:10.1049/iet-cds.2019.0422
Miranda, E., and Braund, E. (2017). A method for growing bio-memristors from slime mold. J. Vis. Exp., e56076. doi:10.3791/56076
Nakagaki, T., Yamada, H., and Toth, A. (2000). Maze-solving by an amoeboid organism. Nature 407, 470. doi:10.1038/35035159
Park, S. O., Jeong, H., Park, J., Bae, J., and Choi, S. (2022). Experimental demonstration of highly reliable dynamic memristor for artificial neuron and neuromorphic computing. Nat. Commun. 13, 2888. doi:10.1038/s41467-022-30539-6
Pershin, Y., La Fontaine, S., and Di Ventra, M. (2009). Memristive model of amoeba learning. Phys. Rev. E 80, 021926. doi:10.1103/physreve.80.021926
Saigusa, T., Tero, A., Nakagaki, T., and Kuramoto, Y. (2008). Amoebae anticipate periodic events. Phys. Rev. Lett. 100, 018101. doi:10.1103/PhysRevLett.100.018101
Schmidt, M., Seyfried, G., Reutina, U., Seskir, Z., and Miranda, E. R. (2025). Electrical characterization of the alleged bio-memristor physarum polycephalum. MRS Adv. doi:10.1557/s43580-025-01198-8
Sheridan, P. M., Cai, F., Du, C., Ma, W., Zhang, Z., and Lu, W. D. (2017). Sparse coding with memristor networks. Nat. Nanotechnol. 12, 784–789. doi:10.1038/nnano.2017.83
Snider, G. (2003). “Molecular-junction-nanowire-crossbar-based neural network,”. United States: HP Inc Hewlett Packard Enterprise Development LP.
Snider, G., Amerson, R., Carter, D., Abdalla, H., Qureshi, M., Leveille, J., et al. (2011). From synapses to circuitry: using memristive memory to explore the electronic brain. Computer 44, 21–28. doi:10.1109/mc.2011.48
Strukov, D. B., Snider, G. S., Stewart, D. R., and Williams, R. S. (2008). The missing memristor found. Nature 453, 80–83. doi:10.1038/nature06932
Taylor, B., Adamatzky, A., Greenman, J., and Ieropoulos, I. (2015). Physarum polycephalum: towards a biological controller. Biosystems 127, 42–46. doi:10.1016/j.biosystems.2014.10.005
Tim, T. V. f. (2021). Physarum polycephalum sur souche. Image available on Wikimedia Commons. Licensed under CC BY-SA 4.0.
Van Haeverbeke, M., De Baets, B., and Stock, M. (2023). Plant impedance spectroscopy: a review of modeling approaches and applications. Front. Plant Sci. 14, 1187573. doi:10.3389/fpls.2023.1187573
Wohlfarth-Bottermann, K. E. (1979). Oscillatory contraction activity in physarum. J. Exp. Biol. 81, 15–32. doi:10.1242/jeb.81.1.15
Keywords: living electronics, memristor, slime mold, biocapacitance, biocomputing
Citation: Schmidt M, Seyfried G, Reutina U, Seskir Z and Miranda ER (2025) From simulation to reality: experimental analysis of a quantum entanglement simulation with slime molds (Physarum polycephalum) as bioelectronic components. Front. Soft Matter 5:1588404. doi: 10.3389/frsfm.2025.1588404
Received: 05 March 2025; Accepted: 02 September 2025;
Published: 08 October 2025.
Edited by:
Michael Taynnan Barros, University of Essex, United KingdomReviewed by:
Milan Cabarkapa, University of Kragujevac, SerbiaPanagiotis Mougkogiannis, University of the West of England, United Kingdom
Copyright © 2025 Schmidt, Seyfried, Reutina, Seskir and Miranda. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Markus Schmidt, c2NobWlkdEBiaW9mYWN0aW9uLmNvbQ==; Zeki Seskir, emVraS5zZXNraXJAa2l0LmVkdQ==