 Caio S. Takiya1
Caio S. Takiya1 Caroline M. Ylioja1Amanda Bennett1Melissa J. Davidson1Maggie Sudbeck1
Caroline M. Ylioja1Amanda Bennett1Melissa J. Davidson1Maggie Sudbeck1 Tryon A. Wickersham2Michael J. VandeHaar3
Tryon A. Wickersham2Michael J. VandeHaar3 Barry J. Bradford1*
Barry J. Bradford1*- 1Department of Animal Sciences and Industry, Kansas State University, Manhattan, KS, United States
- 2Department of Animal Science, Texas A&M University, College Station, TX, United States
- 3Department of Animal Science, Michigan State University, East Lansing, MI, United States
Ruminants can convert feeds unsuitable and unpalatable for humans into milk and meat, and thereby play a key role in food security. Milk production efficiency is usually calculated as the ratio between nutrients secreted in milk and nutrient intake, but this metric does not address concerns about human/livestock feed competition. Our objective was to evaluate effects of diets composed of ecological leftovers (ECO; industrial by-products and feed produced on land unsuitable for human food production) on dairy cattle productivity compared with traditional diets used in the U.S. We also sought to estimate human-edible (HE) nutrient recovery rate (HE inputs vs. milk nutrients) in different scenarios: thrift (all potentially HE ingredients counted as such), choice (ingredients rarely consumed by humans considered not HE), and land use (land used for forage production could be used to grow corn and soybeans for direct human consumption). Experiment 1 evaluated effects of an ECO diet (ECO1), incorporating wheat straw and by-products, on performance of 12 mid-lactation cows in a crossover design with 20-d periods. Experiment 2 evaluated effects of a different ECO diet (ECO2), using winter crop forage and by-products with or without rumen-protected Lys and Met (ECO2-AA), on performance of 12 late-lactation cows in a 3 × 3 Latin square design with 21-d periods. Both ECO diets were compared to lactation diets typical in North America (CON). Although ECO1 decreased feed efficiency (milk yield ÷ feed intake), both feed intake and milk yield were maintained for primiparous cows. ECO1 increased the HE recovery of metabolizable energy (ME) and protein relative to CON1 across all food system scenarios. In Experiment 2, ECO diets significantly decreased feed intake and milk yield, and in the thrift scenario, recovery of ME and protein were worsened by ECO2. All diets resulted in a positive net recovery of HE digestible essential amino acids, and ECO diets further improved their recovery. In conclusion, several factors affect recovery of HE nutrients fed to dairy cows, including dietary composition, land use, and human food system assumptions. Depending on these factors, ECO diets can either improve or reduce the efficiency of converting HE nutrients from feeds into milk.
Introduction
Growing world population, lack of arable land for agriculture expansion, energy demands (bio-fuels), and food waste may threaten global food security in coming decades. Food waste along the supply chain from the producer to the consumer is estimated at 40% in the U.S. (Hall et al., 2009; Guners, 2017). The world's population is expected to grow from 7.6 to 10 billion between 2017 and 2067 (United Nations, 2017) while the global demand for milk is expected to increase by 48% between 2005 and 2050 (Alexandratos and Bruinsma, 2012). In addition, global life expectancy (Zijdeman and Ribeira da Silva, 2014), adult height (NCD Risk Factor Collaboration, 2016), and body mass index (Finucane et al., 2011) have consistently increased over the years, thus affecting the overall maintenance nutrient requirements of the human population. A larger and wealthier global population will demand not only more food but also other goods and services that require land, water, energy, and minerals for their production. In the past 60 years, growth in demand for food has been met primarily by steady increases in agricultural productivity; however, there are worrying signs that productivity gains are leveling off. For instance, the global rate of yield increase of cereal crops has steadily declined—dropping from 3.2%/year in 1960 to 1.5% in 2000 (FAO Food Agriculture Organization of the United Nations, 2009). Thus, new agricultural technologies that can reinvigorate productivity gains and approaches to enhance agricultural/food system efficiency are considered critical to meeting global food security goals.
Studies have suggested that net food production could be increased by shifting crop production away from bioenergy, livestock feed, and other non-food applications (Foley et al., 2011). These authors estimated that shifting major crops directly to human consumption would add over a billion tons to global food production or the equivalent of 3 × 1015 kcal annually, a 49% increase (Foley et al., 2011). Food security concerns, though, extend beyond caloric needs of the growing world population; in fact, amino acids and vitamins are more likely to be limiting for global health than energy (Wu et al., 2014; Chaudhary et al., 2018). As animal-source foods are high in protein, they should not be evaluated primarily as energy sources. In addition to the high protein content found in animal-source foods, protein quality of beef and milk is 1.40–1.87 times greater than human-edible (HE) plant protein inputs used in the dairy industry (Ertl et al., 2016a). Furthermore, some scenarios do not evaluate best practices in assessing the contribution of livestock to the agricultural system. In applied animal nutrition, feed processing methods (Giuberti et al., 2014), genetic modifications (Ferraretto and Shaver, 2015), and metabolic modifiers (McGuffey, 2017) have been consistently used to improve feed efficiency in livestock.
Ruminant rations are composed of a diversity of feed ingredients, of which some could be used as food for humans. Mottet et al. (2018) estimated that only 4–5% of feed consumed by domestic ruminants globally is human-edible (HE), a much smaller share compared with feed consumed by monogastric livestock (83%). Both beef and dairy industries have a major impact on the food system because they can convert feeds unsuitable and unpalatable for humans into high-quality, protein-rich products (Tedeschi et al., 2015), with relatively high concentration and bioavailability of vitamin B12 (Matte et al., 2012). Feed efficiency of livestock is classically defined as the production of animal products (milk, meat, eggs, etc.) divided by the amount of feed ingested. An alternative measurement of feed efficiency in livestock is the calorie/protein retention proposed by Fry et al. (2018), which estimates the percentage from the total calorie and protein consumed by animals that is available for human consumption. These measurements, however, can be misleading because ruminants produce protein-rich meat and milk from fibrous feeds not suitable for many other species, including humans. Therefore, ruminants could be considered more efficient in terms of supporting the human food supply if they consume feeds unsuitable for human nutrition and do not displace human food production from arable land.
In contrast to ruminant systems that rely almost exclusively on grazing non-arable land, highly productive intensive ruminant systems such as those predominant in the U.S. utilize a greater proportion of HE grains and protein sources. Although intensive systems may decrease efficiency on a HE basis, they also enable the use of byproduct feeds in carefully balanced rations. North American dairy rations contain 20–30% by-product feedstuffs (Tricarico, 2016), and thus provide an important avenue for converting these materials into nutrient-dense foods for human consumption. Thus, use of by-product feeds in intensive systems can enhance efficiency in the overall food system. Despite the challenges of feeding by-products (i.e., risk of mycotoxin contamination, variation in nutrient composition between batches, etc.), studies have demonstrated that diets with relatively high proportions of by-products can maintain or even improve ruminant performance (Bradford and Mullins, 2012). Dairy cows fed by-products in place of cereals and pulses had similar dry matter intake (DMI) and milk yield compared with cows fed conventional diets, and the by-product diet increased the conversion ratio of undesirable food for humans into milk (Ertl et al., 2015). More recently, dairy goats fed a diet wherein the concentrate portion was comprised entirely of by-products exhibited improvements in N utilization and feed efficiency and produced less methane in comparison with a traditional diet containing corn and sunflower meal (Romero-Huelva et al., 2017). These experiments support the concept that feeding by-products to ruminants can decrease feed costs, improve the environmental sustainability of milk production (VandeHaar and St-Pierre, 2006), and support high levels of productivity, if utilized carefully (Bradford and Mullins, 2012). Furthermore, Schader et al. (2015) modeled a food system in which animals were fed zero human-edible feeds (grassland and by-products) and predicted reductions in greenhouse emissions, pesticide use intensity, freshwater use, soil erosion potential, and N- and P-surplus while providing enough food for the 2050 reference scenario from Alexandratos and Bruinsma (2012). Thus, feeding animals with ingredients not edible for human consumption likely has a positive impact not only on global food security, but also on the environment.
Feeding animals with resources not suitable for human consumption, such as by-products and grass from marginal land unsuited for crop production, is referred to as producing livestock on “ecological leftovers” (ECO; Garnett, 2009). The ECO concept follows a set of normative principles including (1) arable land should primarily be designated to plant-based food for humans, (2) animals should be fed biomass unwanted by or unsuitable for humans, and (3) semi-natural grassland should be used for livestock, if grazing can be justified by reasons other than meat and milk production (e.g., biodiversity or soil conservation). Although there are many studies evaluating the replacement of cereals and pulses with by-products in ruminant diets, the literature lacks data evaluating the effects of full ECO diets on dairy cow performance and HE nutrient recovery. The objective of this study was to evaluate impacts of different ECO diets on performance of mid- and late-lactation Holstein cows compared with a conventional diet used in the U.S.; we hypothesized that cows fed carefully-formulated ECO diets would maintain similar milk production while improving HE nutrient recovery (amount HE nutrients recovered per HE nutrients fed). Furthermore, we provide an alternative approach to estimating HE nutrient recovery rate for dairy cows.
Materials and Methods
Two experiments were carried out between March and May 2015 (Experiment 1) and March and May 2017 (Experiment 2) at the Dairy Teaching and Research Center of Kansas State University (KSU), under the approval of the KSU Institutional Animal Care and Use Committee. In both experiments, cows were housed in a tie-stall barn containing rubber beds covered with wood shavings, and individual feed bunks and waterers.
Experiment 1
Twelve dairy cows (six primiparous and six multiparous) after peak lactation (154 ± 20 days post-partum and 42.6 ± 5.4 kg/d milk yield at the start of experiment) were randomly assigned to treatment sequence (within parity) in a crossover design. Periods lasted 20 d; the first 17 d allowed for diet adaptation and d 18–20 were used for data collection and sampling. Treatments were a conventional diet containing 31% by-product feeds (CON1) or a diet with 95% by-product feeds (ECO1; Table 1). Cows were fed diets twice daily as total mixed rations (TMR) for ad libitum intake. Feedstuff and TMR samples were collected, composited, and analyzed for chemical composition (Table 2) using wet chemistry methods by the Dairy One Forage Laboratory (Ithaca, NY), including dry matter (method 930.15, AOAC), ash (method 942.05, AOAC), crude fat (ether extraction method 2003.05, AOAC), acid detergent fiber (method 973.18, AOAC), neutral detergent fiber (NDF) adding sodium sulfite and alpha-amylase to the detergent (Van Soest et al., 1991), and crude protein [CP; methods 990.03 (animal feed) and 992.23 (cereal grain), AOAC]. Net energy of lactation was calculated according to the National Research Council (2001) and non-fiber carbohydrate (NFC) was calculated as: NFC = 100 – (ash-free NDF% + CP% + crude fat% + ash%). Total mixed ration samples were assessed for particle size distribution (Heinrichs and Kononoff, 2013).
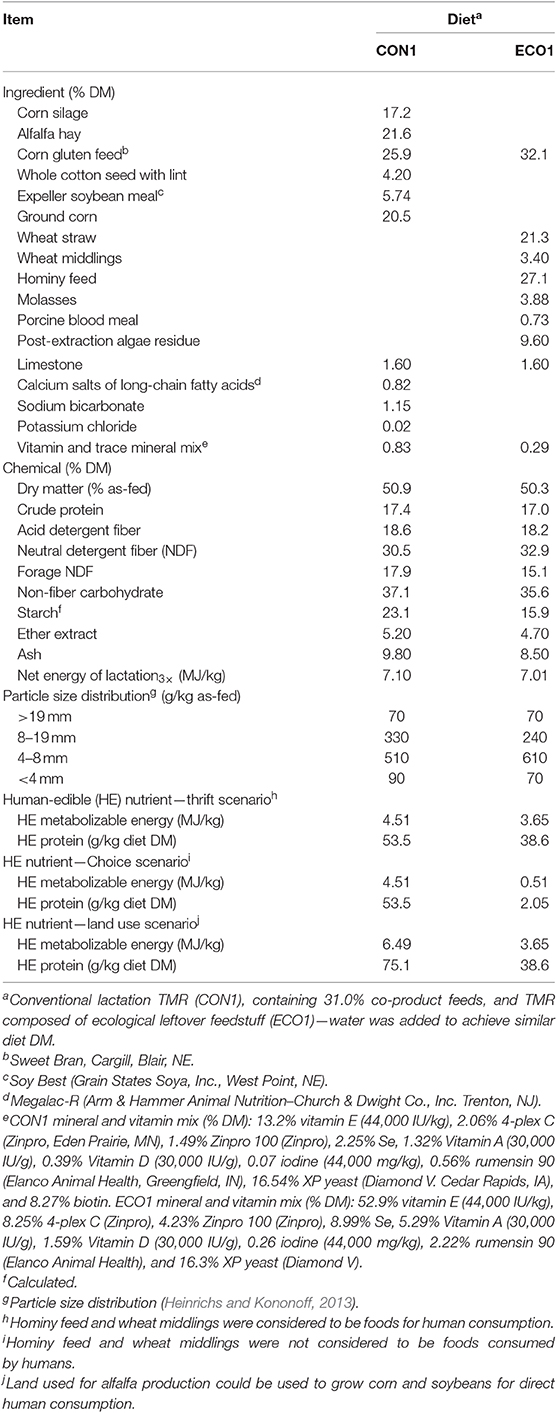
Table 1. Ingredients and chemical composition of diets provided in Experiment 1.
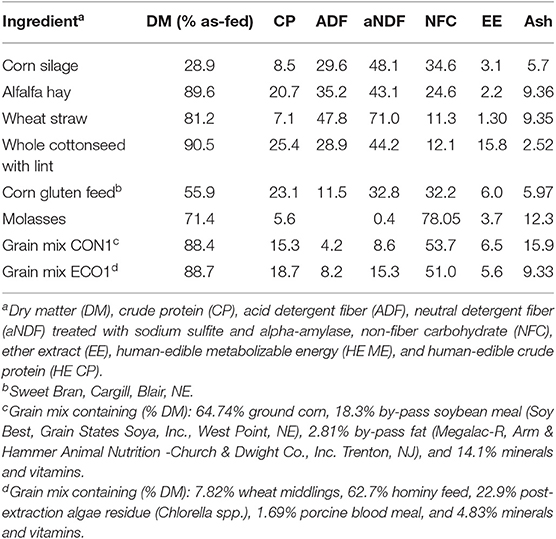
Table 2. Chemical composition of feeds used in Experiment 1 (% of DM, otherwise stated).
Feed and refusals were weighed daily to calculate DMI over each sampling period. On d 20, blood samples were collected from coccygeal vessels into evacuated tubes (6 mL K3EDTA Vacutainer tubes; Becton Dickinson, Franklin Lakes, NJ) before the morning feeding and analyzed for plasma glucose concentration by a colorimetric kit (kit #439–90901; Wako Chemicals USA Inc., Richmond, VA), and plasma insulin concentration by a bovine-specific sandwich ELISA (#10–1201–01; Mercodia AB, Uppsala, Sweden). Absorbance was measured on a spectrophotometer (PowerWave XS; BioTek Instruments Inc., Winooski, VT) and calculations were performed using Gen5 software (BioTek Instruments Inc.).
Cows were milked thrice daily, and milk samples were collected on the last 3 days of each experimental period at nine consecutive milkings in plastic vials with preservative (Microtabs II, Advanced Instruments, Norwood, MA) and analyzed by DHIA Laboratories (Manhattan, KS) for milk fat, true protein, lactose, and urea N concentrations as well as somatic cell count (SCC) as described in Ylioja et al. (2018). Somatic cell linear score (SCLS) was calculated as: SCLS = log2 (SCC/100) + 3 (Shook, 1993). Body weight and body condition score were recorded at the beginning and end of each period (Wildman et al., 1982).
Data were submitted to analysis of variance using the MIXED procedure of SAS 9.4 (SAS Institute, Cary, NC) to account for the fixed effects of diet, parity, and their interaction, as well as the random effects of cow and period. Residual normality was verified. When diet by parity interaction was significant, differences among least square means were analyzed using the Tukey option. Significance was declared at P ≤ 0.05 and tendencies at 0.05 < P ≤ 0.10. One primiparous cow was removed from analysis due to refusal to consume the ECO1 diet.
Experiment 2
Twelve second lactation dairy cows (231 ± 40 days post-partum and 34.4 ± 7.2 kg/d milk yield at the start of the experiment) were assigned to a 3 × 3 Latin square design experiment balanced for carryover effects. Adaptation to diets was allowed for 17 d and 4 d were used for data collection and sampling. Cows were blocked (n = 4) according to fat-corrected milk yield and days in milk, and randomly assigned to treatment sequence within block. Treatments (Table 3) were: (1) a conventional TMR for lactating cows (CON2) containing 25.7% by-product feeds; (2) a TMR comprised entirely of ECO feedstuffs (ECO2); and (3) ECO2 with top-dressed rumen-protected lysine and methionine [ECO2-AA; 77 g/d AjiPro-L (Ajinomoto, Chicago, IL) and 45 g/d MetaSmart (Adisseo, Antony, France)]. Cows were milked and fed twice daily (0530 and 1,600 h). The CON2 TMR was mixed before the morning feeding in a horizontal mixer wagon, whereas ECO2 TMR was prepared before the afternoon feeding in a stationary horizontal mixer. Spent coffee grounds were acquired from three coffee shops every other day and stored in plastic buckets at room temperature until mixed with other feeds. Amino acids were top-dressed and mixed into the top 2/3 of the TMR for each cow at both feedings for ECO2-AA. CON2 was formulated according to the National Research Council (2001). ECO2 and ECO2-AA were formulated using Formulate2 (Diet Formulation Systems LCC, Visalia, CA) based on the National Research Council (2001) model and requirements. Chemical composition of feeds is shown in Table 4.
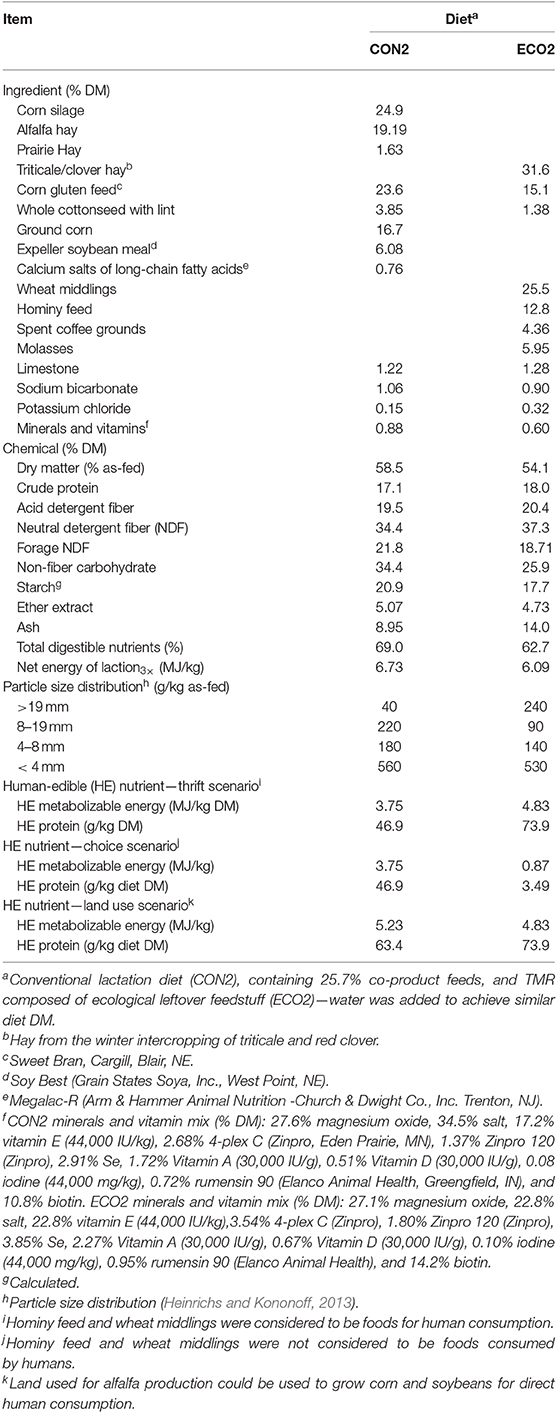
Table 3. Ingredient, chemical composition, and particle size distribution of diets provided in Experiment 2.
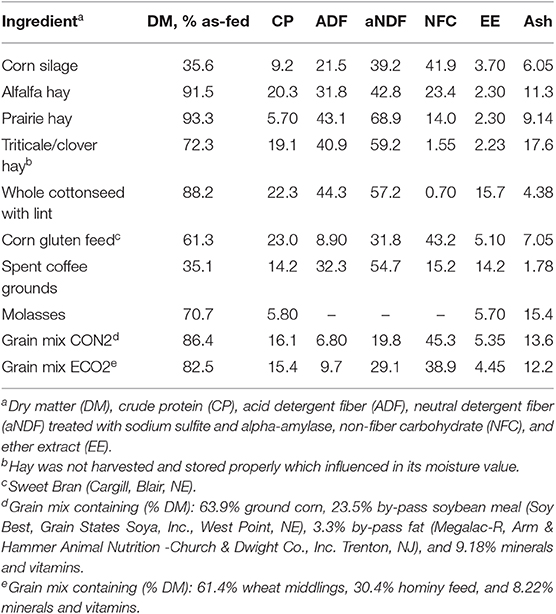
Table 4. Chemical composition of feeds used in Experiment 2 (% of DM unless otherwise labeled).
Feed and refusals were weighed daily, and feeding rates targeted 10–15% refusals. During the last 4 d of each period, TMR samples were collected to assess particle size distribution and chemical composition as described earlier. Milk samples were collected during every milking of the last 4 d of each period, and analyzed by MQT Lab Services (Kansas City, MO) using the same methods described in Experiment 1. Body weight and body condition score were recorded at the start and end of each period.
Analysis of variance was performed using the Mixed procedure of SAS 9.4 (SAS Institute) in a model including the fixed effects of diet and block and the random effects of period and cow within block. Normality of residuals was verified. Differences among least square means were evaluated using the Tukey option. Significance was declared at P ≤ 0.05 and tendencies at 0.05 < P ≤ 0.10.
Human-Edible Nutrient Recovery Calculations
Maximum HE ME and protein contents of cattle diets were estimated based on sugar, starch, true protein, and fat concentrations in corn grain (including grain in silage), hominy feed, soybean meal products, wheat middlings, and molasses. Other feedstuffs were considered unsuitable for human consumption (such as spent coffee grounds). Metabolizable energy content of feeds and milk were calculated based on Atwater calorie factors (Atwater and Bryant, 1900): 4 kcal/g of starch or sugar, 9 kcal/g of fat, and 4 kcal/g of protein (Table 5). Corn silage was considered to contain 45.0% corn grain (DM basis), and the corn grain was assumed to contain 66% starch and 9.5% protein (DM basis), and 50% moisture. Soybean meal product (Soy Best, Grain States Soya Inc., West Point, NE) was estimated to contain 6.6% fat (DM basis), 49% protein (DM basis), and 11% moisture. Hominy feed and wheat middlings nutrient values were retrieved from the National Research Council (2001). We calculated HE nutrient recovery in milk in three scenarios; the first considered hominy feed and wheat middlings suitable for human consumption (thrift scenario), the second considered hominy feed and wheat middlings as unlikely to be consumed by humans (choice scenario), and the third was similar to the thrift scenario but considered that the land used for alfalfa hay production could be used to grow soybeans and corn (land use scenario).
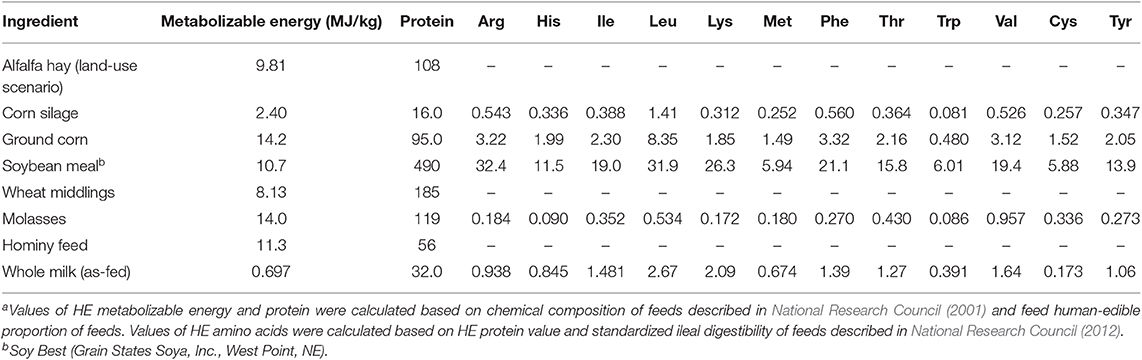
Table 5. Human-edible (HE) nutrient values used for calculations (DM basis; g/kg unless otherwise labeled)a.
For the land use scenario, a blend of corn and dehulled soybeans (82 and 18% on DM basis, respectively) offering a ME:protein ration similar to milk (21.8 kcal/g protein; 3.5% fat, 3.2% protein, and 4.8% lactose) was evaluated in place of alfalfa. Thus, one hectare used for alfalfa hay production could be used to grow 0.57 ha of corn and 0.43 ha of soybeans. The annual yields (on DM basis) of alfalfa hay (7,263 kg/ha), corn (7,125 kg/ha), and soybeans (2,240 kg/ha), retrieved from Kansas 2017 Annual Crop Production Summary (Bounds, 2018), were used to determine the potential HE ME and protein displaced by alfalfa hay production. Soybean yield was discounted 7.3% (Perkins, 1995) to account for inedibility of hulls. Employing these assumptions, the displaced HE nutrient potential of alfalfa hay was estimated at 9.63 MJ/kg and 103 g protein/kg alfalfa.
To account for amino acid recovery, we estimated the HE digestible amino acid inputs and outputs using the feed total amino acid content and standardized ileal digestibility (SID) values from the National Research Council (2012) ingredient library, with the exception of milk values that we retrieved from INRA-CIRAD-AFZ (2002). Standardized ileal digestibility of amino acids in pigs can be used when values in humans are not available (FAO, 2013). Recoveries of HE nutrients in milk were calculated as the ratio between nutrients secreted in milk and the corresponding HE nutrient intake by cattle, where a value of 2 would indicate that for every HE unit (energy, protein, or amino acid) the cow consumes, she would secrete 2 units in milk. Detailed calculations for HE nutrient displacement by alfalfa, HE SID content of feeds, and HE nutrient recovery can be found in Supplementary Material.
Results
Experiment 1
Cows fed ECO1 had decreased feed efficiency in terms of milk yield (P = 0.025) per kg of DMI compared with CON1 (Table 6). Feeding ECO1 instead of CON1 to cows also reduced yields of milk fat (P = 0.001) and milk protein (P = 0.020) and tended to decrease (P = 0.084) lactose yield. On the other hand, ECO1 reduced (P < 0.001) milk urea N concentration and increased (P < 0.01) blood glucose concentration compared with CON1. As expected, primiparous cows had lesser (P ≤ 0.001) DMI, SCLS, and body weights compared to multiparous cows. The effect of diet on milk yield differed by parity, where the ECO1 diet decreased milk yield (interaction P = 0.045) only in multiparous cows.
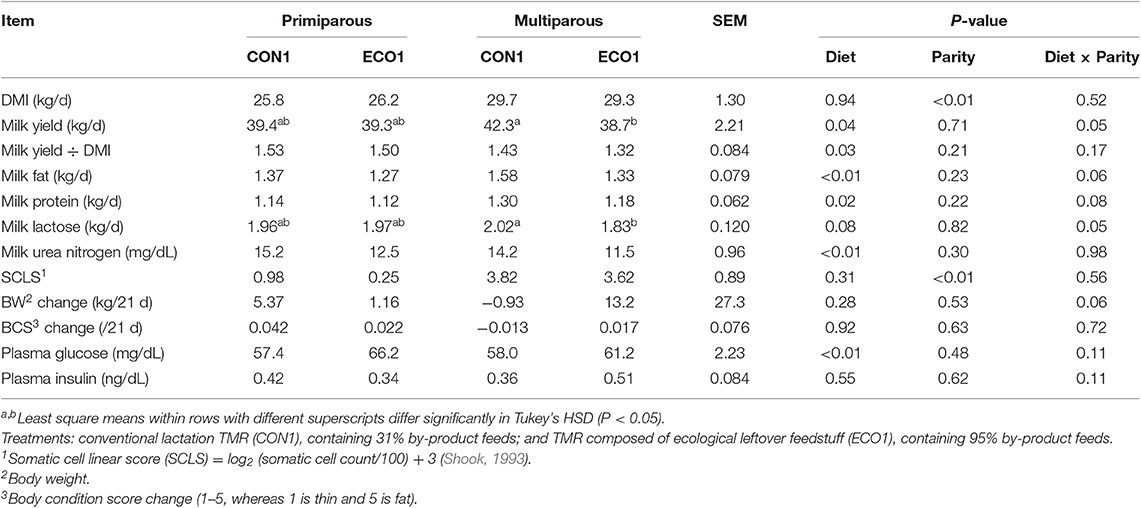
Table 6. Performance and blood plasma metabolites of mid-lactation cows fed a conventional lactation diet or a by-product-based diet (Experiment 1).
In all three calculated scenarios, the ECO1 diet decreased (P < 0.01, Table 7) HE ME inputs by 27–119 MJ/d and HE protein inputs by 0.37–1.53 kg/d compared with CON1 diet. In the choice scenario, HE inputs differed by parity for the CON1 diet only, with multiparous cows consuming more than primiparous. However, the overall effects of diet were similar in magnitude; ECO1 had 90% lower ME inputs and 95% lower HE protein inputs than CON1. The land use scenario had outcomes similar to the thrift scenario, although CON1 HE input values for ME and protein were greater than in the thrift scenario so that reductions in HE inputs with ECO1 were even greater. In all food system scenarios, cows fed ECO1 had greater HE ME and protein recoveries than those fed CON1 (P < 0.01), although the magnitude of the difference varied greatly based on the scenario (Figure 1). For thrift scenario and land use scenarios, recoveries of HE energy and protein were 0.6–0.94 for CON1 and increased by 5–50% for ECO1. However, for choice scenario ECO1 increased recoveries of energy and protein by 700–2,000%. Human-edible digestible amino acid recoveries were analyzed in the choice scenario. Cows fed the ECO1 diet had greater (P < 0.01) recoveries of all amino acids evaluated in this study (Arg, His, Ile, Leu, Lys, Met, Phe, Thr, Trp, Val, Cys, and Tyr; Table 8).
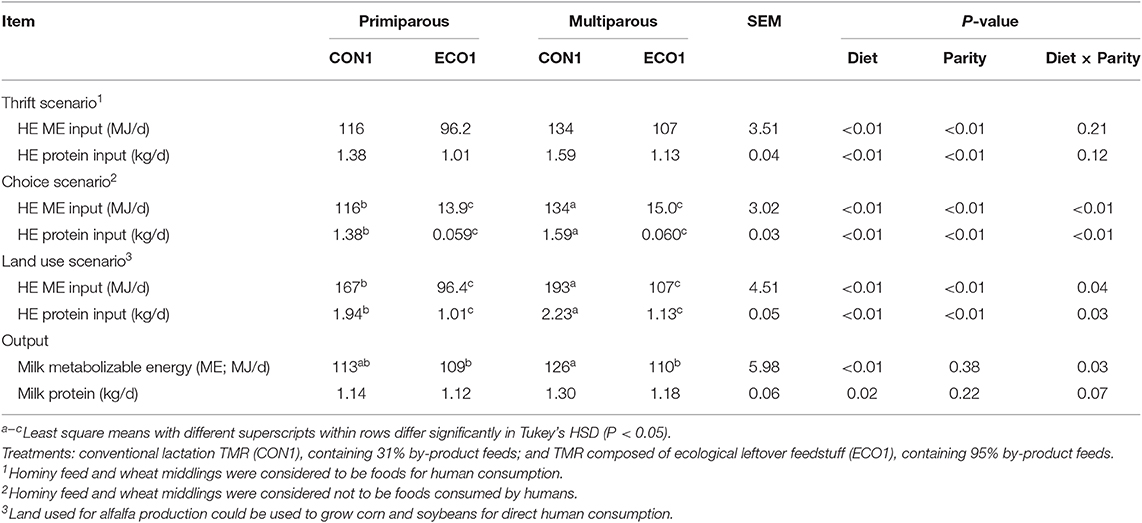
Table 7. Human-edible (HE) nutrient inputs and outputs of mid-lactation cows fed a conventional lactation diet or a by-product-based diet (Experiment 1).
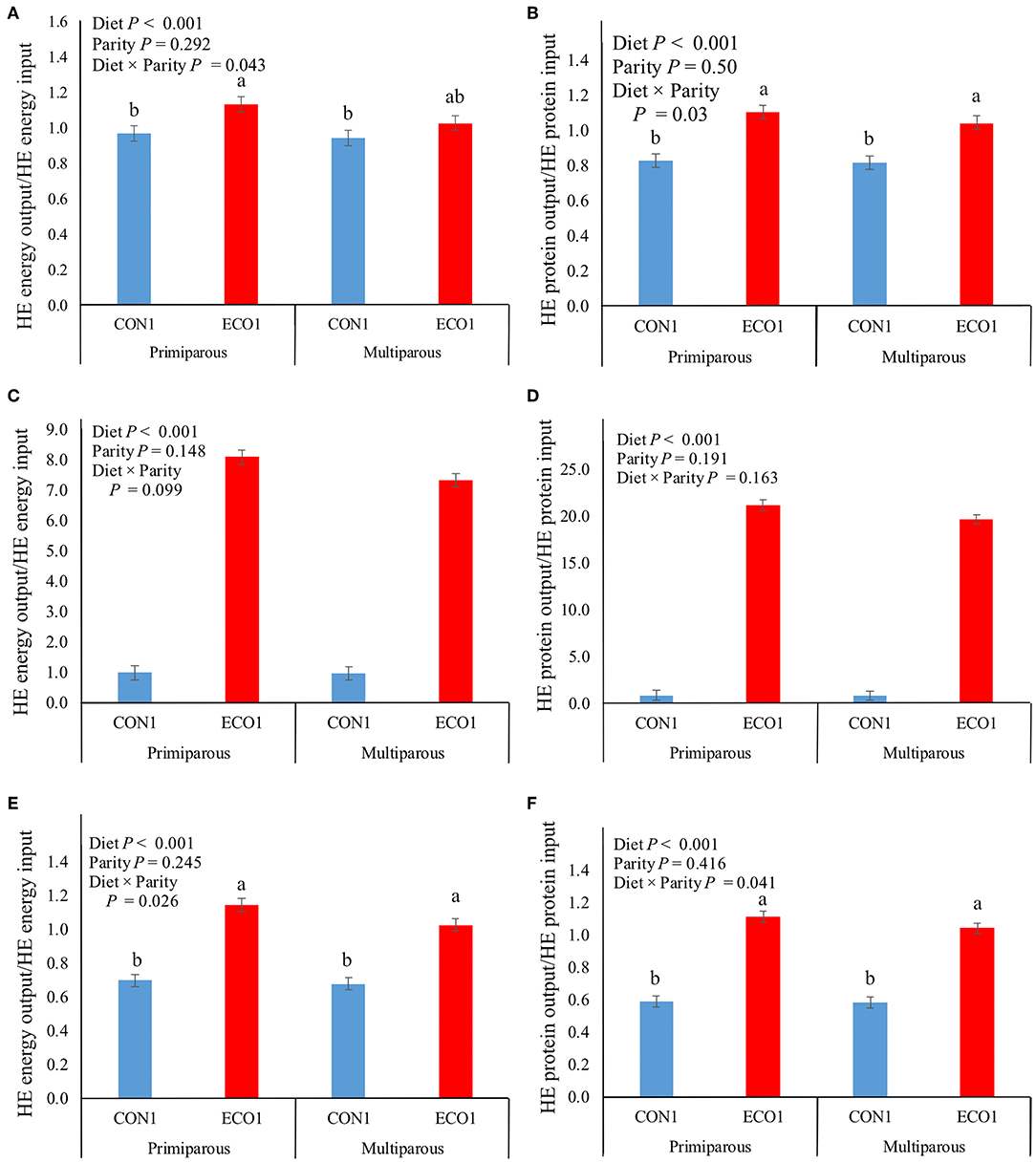
Figure 1. Human-edible (HE) metabolizable energy (left panels) and protein (right panels) conversion rate of mid-lactation cows fed a conventional lactation diet or a co-product-based diet (Experiment 1) in a thrift scenario (A,B), a choice scenario (C,D), or a land use scenario (E,F). A recovery value of 2 would indicate that for every HE unit (either MJ or protein) the cow consumes, she would produce 2 HE unit in milk. Thrift scenario considered that hominy feed and wheat middlings were edible by humans, whereas choice scenario considered that hominy feed and wheat middlings were not edible by humans. Land use scenario calculations were made based on thrift scenario with additional consideration that the land used for alfalfa production could have been used to grow corn and soybeans for direct human consumption. Treatments: conventional lactation TMR (CON1), containing 31% by-product feeds; and TMR composed of ecological leftover feedstuff (ECO1), containing 95% by-product feeds. Letters above columns describe treatment by parity differences (P < 0.05; Tukey's Honestly Significant Difference test). Error bars are SE.
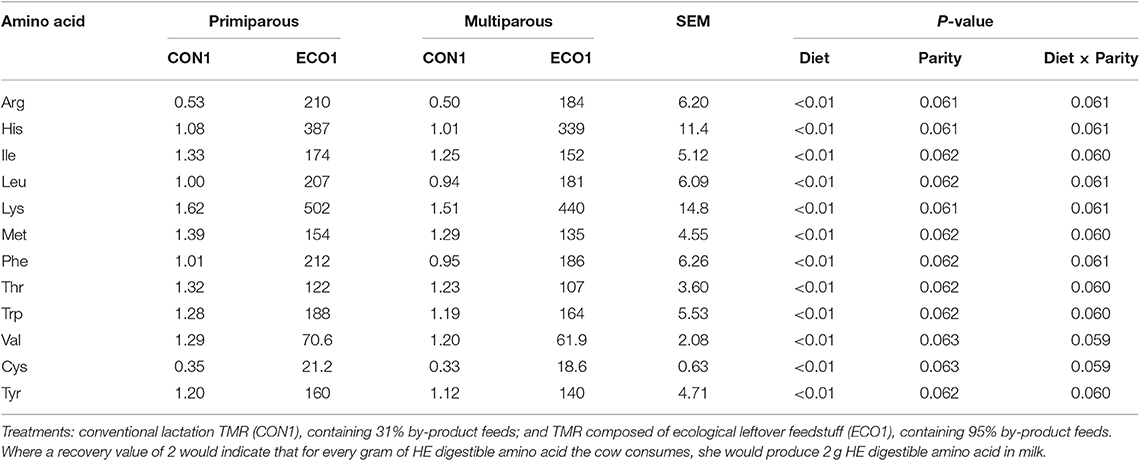
Table 8. Human-edible (HE) digestible amino acid recovery (g output/g input) in milk of mid-lactation cows fed a conventional lactation diet or a by-product-based diet (Experiment 1, Choice Scenario).
Experiment 2
Cows fed ECO2 or ECO2-AA diets consumed less DM (P < 0.01) compared with those fed CON2 (Table 9). ECO2 and ECO2-AA likewise decreased (P < 0.01) milk yield of cows. Feed efficiency (milk yield ÷ DMI) was similar between cows fed CON2 and ECO2 but was reduced (P = 0.02) when cows were fed ECO2-AA. In general, ECO2 and ECO2-AA diets reduced the production of milk fat, protein, and lactose. Cows fed ECO2 diets also had greater (P < 0.01) SCLS relative to those fed the CON2 diet.
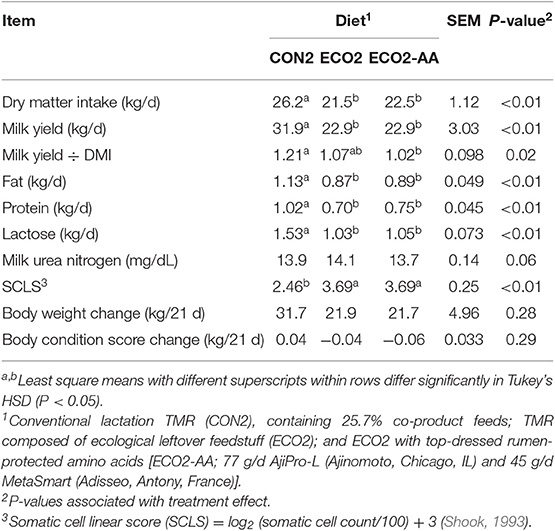
Table 9. Performance of late-lactating cows fed a conventional lactation diet or by-product-based diets (Experiment 2).
Feeding cows with ECO2 and ECO2-AA diets reduced (P < 0.01) outputs of HE ME and protein in milk in comparison with CON2 diet (Table 10). In the thrift scenario, ECO2 diets had no effect on HE ME inputs, and they increased (P < 0.01) HE protein input. In the choice scenario, cows fed ECO2 diets had lesser (P < 0.01) HE ME and protein inputs compared with those fed CON2 diet. In the land use scenario, cows fed ECO2 diets had lesser (P < 0.01) HE ME input but did not affect HE protein inputs. Recovery of HE ME and protein in milk were decreased (−30 and −40% on average, respectively; P < 0.01) by ECO2 diets under the thrift scenario (Figures 2A,B). On the other hand, in choice scenario (Figures 2C,D) ECO2 diets improved (P < 0.01) the recovery of HE ME and protein in comparison with CON2 diet. In the land use scenario (Figures 2E,F), recovery of HE ME was not affected by diets and recovery of HE protein was worsened (P < 0.01) by ECO2 diets. In the choice scenario, HE ME and HE protein recoveries increased by 360 and 950%, respectively. Similar to observations in Experiment 1, ECO2 diets in a choice scenario improved (P < 0.01) the recovery of all amino acids evaluated (Table 11), with no detectable differences between ECO2 and ECO2-AA.
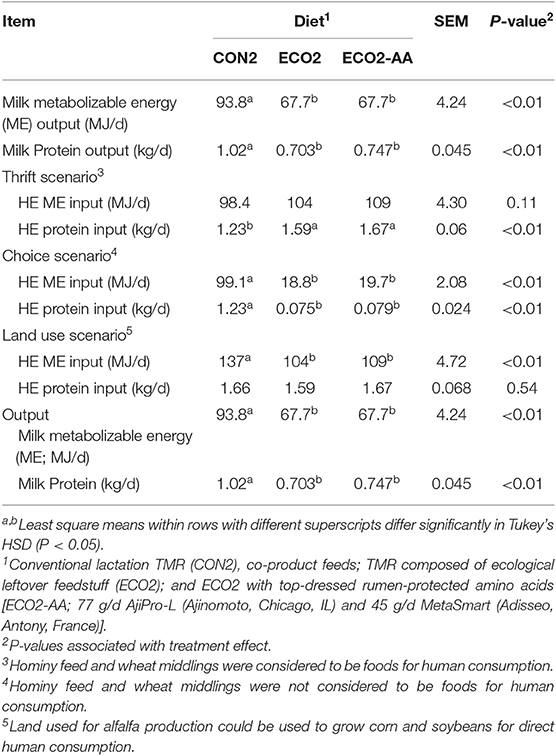
Table 10. Human-edible (HE) nutrients inputs and outputs of late-lactation cows fed a conventional lactation diet or by-product-based diets (Experiment 2).
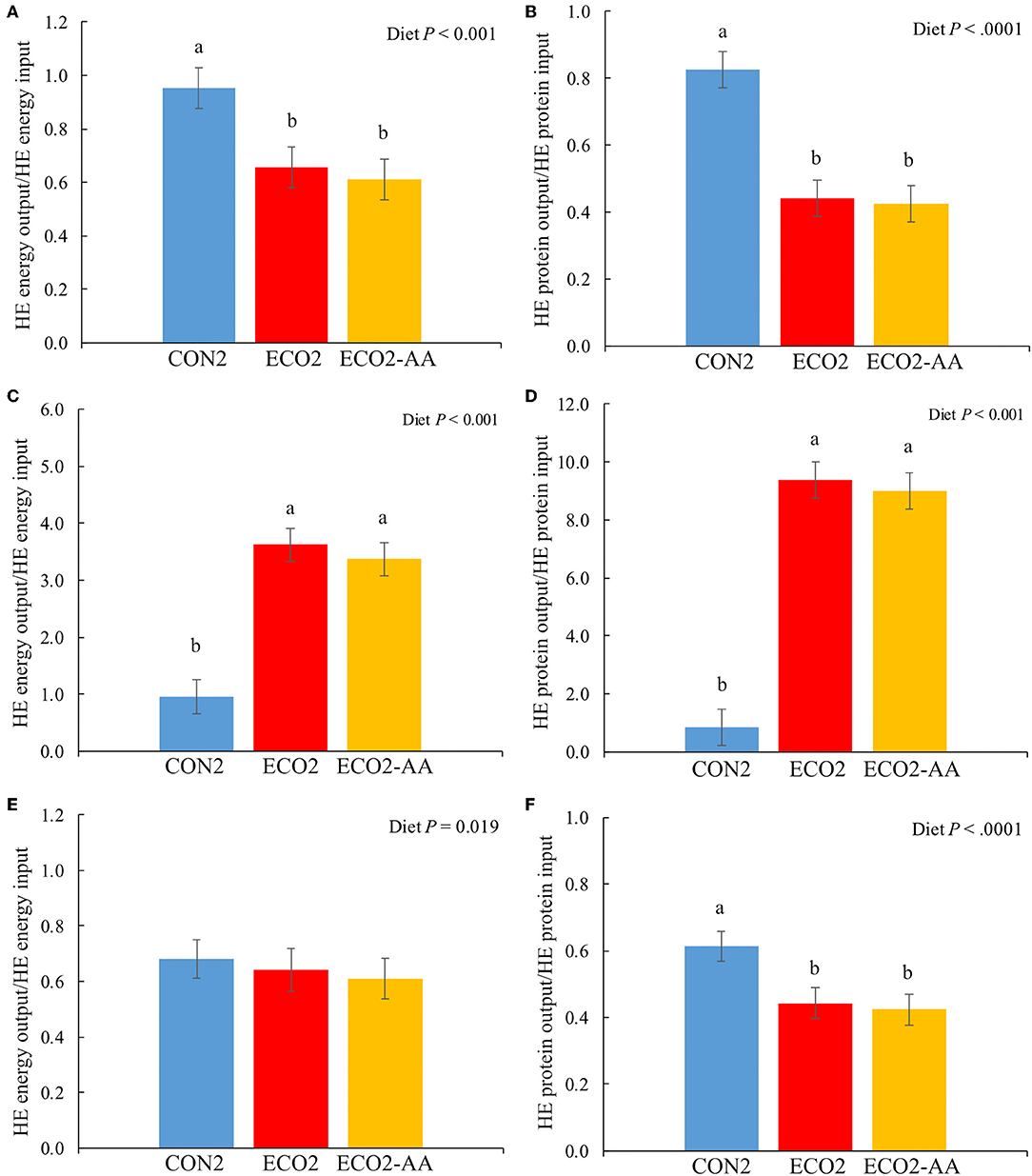
Figure 2. Human-edible (HE) metabolizable energy (top panels) and protein (bottom panels) conversion rate of mid-lactation cows fed a conventional lactation diet or a co-product-based diet (Experiment 2) in a thrift scenario (A,B), in a choice scenario (C,D), or in a land use scenario (E,F), where a recovery value of 2 would indicate that for every HE unit (either MJ ME or protein) the cow consumes, she would produce 2 HE unit in milk. Thrift scenario considered that hominy feed and wheat middlings were edible by humans, whereas choice scenario considered that hominy feed and wheat middlings were not edible by humans. Land use scenario calculations were made based on thrift scenario with additional consideration that the land used for alfalfa production could be used to grow corn and soybeans for direct human consumption. Conventional lactation TMR (CON2), co-product feeds; TMR composed of ecological leftover feedstuff (ECO2); and ECO2 with top-dressed rumen-protected amino acids [ECO2-AA; 77 g/d AjiPro-L (Ajinomoto, Chicago, IL) and 45 g/d MetaSmart (Adisseo, Antony, France)]. ME stands for metabolizable energy. Letters above columns describe treatment differences (P < 0.05; Tukey's Honestly Significant Difference test). Error bars are SE.
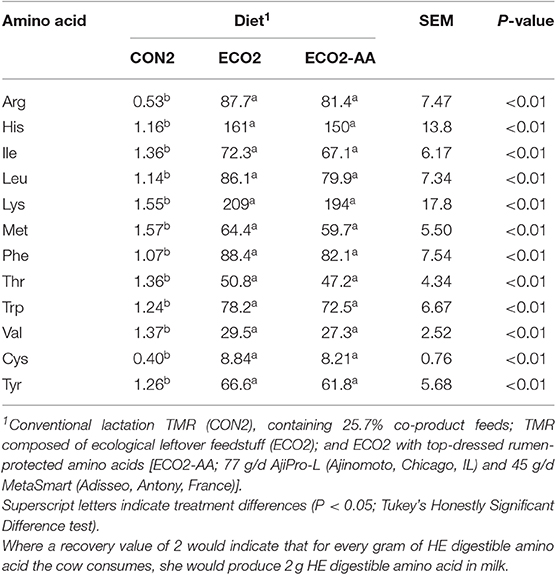
Table 11. Human-edible (HE) digestible essential amino acid recovery (g output/g input) in milk of mid-lactation cows fed a conventional lactation diet or by-product-based diets (Experiment 2, Choice Scenario).
Discussion
Dairy Cattle Productivity
The hypothesis of the current study was that cows fed ECO diets would maintain milk production while improving the HE nutrient conversion rate by dairy cows. Neither ECO1 nor ECO2 strategies were able to maintain equivalent milk yield in all cows. Furthermore, this study showed evidence that a dairy cow diet composed of by-products does not necessarily improve the HE nutrient conversion rate in dairy cows, as the outcome is dependent on parity, feed ingredients, and assumptions made in calculating the value of feed ingredients for human consumption.
In Experiment 1, cows had similar DMI across diets, but the ECO1 treatment decreased milk yield in multiparous cows and reduced overall feed efficiency in all cows, regardless of parity. Decreased milk yield might be attributed to decreased nutrient digestibility for ECO1 compared with CON1. Replacing alfalfa with wheat straw (on forage NDF basis) in diets of lactating cows decreased fiber digestibility, milk yield, and milk fat content (Poore et al., 1991). Anderson and Hoffman (2014) reported that wheat straw NDF digestibility (48 h in vitro) may range from 21 to 49% (39.0% average) whereas corn silage and alfalfa hay fiber digestibility ranges from 58 to 67% (Ivan et al., 2005) and 50 to 56% (Sulc et al., 2016), respectively. Oba and Allen (1999) summarized data from several experiments to determine the influence of fiber digestibility among forages with similar fiber and crude protein content on dairy cow performance. These authors reported that a one-unit increase in forage fiber digestibility was associated with a 0.17 kg/day increase in DM intake and a 0.25 kg/day increase in fat-corrected milk yield. High-productivity dairy cattle systems are generally based on dedicated production of highly digestible forages, and this represents a significant challenge in attempting to avoid displacing arable land from human food crop production.
The responses to ECO1 differed by parity, and this interaction provides some insights into the likely cause of the decline in milk yield when feeding ECO diets. Multiparous cows produce more milk and thus have greater nutrient requirements than primiparous cows, and in this parity group, ECO1 apparently failed to provide adequate nutrients (either from poor digestibility or an imbalance in nutrient profile) to maintain the high level of milk production. On the other hand, primiparous cows, which produce more moderate yields, maintained productivity when fed ECO1. Not surprisingly, cows with greater levels of productivity are less flexible in terms of diet formulation.
As in the current study, Karlsson et al. (2018) reported similar DMI and a marginal decrease (32.1 vs. 30.8 kg/d, P = 0.06) in milk yield when replacing cereal grains and soybean meal with sugar beet pulp and distillers' grains. In contrast, Pang et al. (2018) fed cows a diet composed of grass silage with either a by-product or grain concentrate and found a trend for decreased DMI (P = 0.06) for the group fed the by-product concentrate with no differences in milk yield. Co-products often provide less rumen-degradable protein and more rumen undegradable protein in comparison with the original feed (Bradford and Carpenter, 2017). Moreover, feeds that are excessively heated may exhibit relatively high concentrations of Maillard products, thus decreasing protein digestibility. Although not explored in Experiment 1, the formulation software highlighted a possible Met deficiency for dairy cows fed ECO1 (estimated supply of 55 g/d intestinally-available Met vs. estimated requirement of 59 g/d to maintain yield of control group; National Research Council, 2001), which is often considered to be the first-limiting amino acid for milk production in dairy cattle (Schwab and Broderick, 2017). It is important to highlight that it is difficult to meet nutrient requirements with ECO ingredients alone because by-products often lack valuable nutrients that have been removed for other purposes.
Based on the formulation software report and results from Experiment 1, the second experiment evaluated whether supplementing a by-product diet with rumen-protected amino acids would maintain milk production levels of cows. However, due to differences in feed availability, a modified formulation was used. Contrasting with experiment 1, cows fed ECO2 diets, regardless of amino acid supplements, had a substantial decrease in both DMI (−4.7 kg/d) and milk yield (−9.0 kg/d). This large negative impact on performance of cows can likely be attributed to the forage source used in this study. Supplementing ECO2 with amino acids tended to decrease milk urea N concentration, suggesting an improvement in N utilization. However, the triticale/clover hay had high crude protein content and high moisture content that favored spoilage and reductions in feed intake. These problems again highlight the challenges associated with avoiding the dedication of land toward growing high-quality forages production for dairy cattle.
Human-Edible Nutrient Recovery
Bywater and Baldwin (1980) discussed the rationale of calculating animal efficiency based on total feed inputs vs. inputs of HE energy and protein, since the ultimate concern is total human food production. For instance, returns of HE digestible energy and protein varied from 57 to 128% and 96 to 276%, respectively, depending on the feeding strategy adopted in some studies (Baldwin et al., 1992; Oltjen and Beckett, 1996). More recently, the term edible feed conversion ratio was introduced by Wilkinson (2011) to relate HE nutrient inputs and outputs in animal products, and this approach has been used to estimate HE energy and protein recovery in milk in several studies (Ertl et al., 2015; Karlsson et al., 2018; Pang et al., 2018). However, this concept estimates the proportion of crops and crop by-products that are potentially suitable for human consumption in the UK, rather than considering the nutrient composition of each feedstuff as consumed by humans. For instance, cereal by-products, oilseed meals (excluding soybean meal), and other by-products (molasses, candy waste, spent coffee, and others) were assigned a HE nutrient coefficient of 0.2 (meaning the ingredients are considered 20% human edible), whereas soybean meal and cereal and pulse grains were assigned to a HE nutrient coefficient of 0.8. This concept would consider molasses and spent coffee grounds to contain the same nutritional value for humans. In addition, the cereal portion in silages is not considered as a HE nutrient by Wilkinson (2011). Corn silage is the main forage source for confined dairy cows in much of the world, and failure to consider the corn grain portion in plants as a HE food (before ensiling) would ultimately overestimate HE nutrient recovery of the diet. In addition, the above calculations ignored the fact that land used for growing forages could in some cases be used for growing crops to be consumed directly by humans. Vandehaar (1998) calculated the efficiency of land use by considering the HE value of corn and soybeans on a protein- and energy-equivalent basis to milk to account for this possibility, postulating that a combination of corn and soybeans (similar to blends of other grains and legume seeds) would be the most effective way to feed people with limited land resources. Therefore, alternative methods to determine HE nutrient value of feeds should be considered to more precisely describe the efficiency of HE feed conversion. As an alternative approach, to describing efficiency of HE feed utilization, we calculated energy, protein, and digestible amino acid content of feeds in cattle diets that could potentially be used for human consumption relative to secretion of the respective nutrients in milk.
Experiment 1 showed evidence that increased inclusion of by-product feeds in the diet improves the HE nutrient recovery in milk of cows, which has also been observed by others (Ertl et al., 2015; Karlsson et al., 2018; Pang et al., 2018). However, Experiment 1 demonstrated an average increase in HE recovery of just 13% in terms of energy and 22% in terms of protein (thrift scenario), whereas previous experiments have shown recoveries ranging from 367 to 495% in energy and 267 to 441% in protein (Ertl et al., 2015; Karlsson et al., 2018; Pang et al., 2018)—suggesting that calculations strategies (i.e., HE coefficients) heavily influence conclusions regarding HE nutrient recovery in milk. For instance, in the land use scenario of Experiment 1, the average increase in HE recovery was around 50 and 40% in terms of energy and protein, respectively. (Ertl et al., 2016a) calculated the HE conversion rate (based on minimum or maximum potential recoveries rates of HE energy and protein from the respective feedstuffs) from other studies that fed by-product-based concentrates to cows (Voelker and Allen, 2003; Dann et al., 2014). The authors reported no net HE energy or protein production when calculations used minimum HE nutrient coefficients; however, when calculating the conversion rate using maximum HE nutrient coefficients, authors reported net production of HE nutrients by lactating cows (Ertl et al., 2016b). It is important to note that the thrift scenario arguably represents an extreme, incongruous situation—one where human populations have become more open to consuming foods currently avoided, but where by-products (from biofuel production, for example) remain available for cattle diets. However, this scenario could arise through food technologies that enhance the incorporation of products like wheat middlings in human foods rather than out of desperation for food sources.
To demonstrate that HE nutrient conversion rate is highly dependent on model assumptions, we also estimated HE conversion values with the assumption that wheat middlings and hominy feed are unlikely to be consumed by humans (choice scenario). Although wheat middlings can be incorporated into certain human foods (i.e., pasta; Kaur et al., 2012), it is extensively marketed as a by-product feed for livestock, and its competitive price (Capehart et al., 2018) in relation to other energy and protein sources demonstrates its low perceived desirability in the human food supply. Hominy feed is a by-product from the manufacture of pearl hominy, hominy grits, or corn meal, and it is rarely used for human nutrition. Note that hominy feed is different from hominy food, where the former is a by-product of dry corn milling and the latter is a food made from kernels of corn soaked in alkali solution that removes hull and germ. The HE recovery outcome varied drastically depending on the HE values of these two ingredients. The proportional increase in HE nutrient conversion rate was much greater for ME (8.0-fold) and protein (24.8-fold) in the choice scenario compared with the thrift scenario (ME 1.13-fold and protein 1.25-fold) in Experiment 1, and for Experiment 2, the recovery of both ME and protein improved from a HE nutrient loss in the thrift scenario to a HE nutrient gain in the choice scenario.
Some approaches to calculating HE conversion rate of nutrients ignore forage displacement entirely, but it is more reasonable that only non-arable forage land (i.e., upland pastures) should be excluded in calculations. For instance (Ertl et al., 2015), did not consider any “HE value” for alfalfa hay when estimating the HE conversion rate, potentially underestimating the HE inputs—since crops for human food production could be planted instead of alfalfa (acknowledging the complexity of this question in mountainous Austria). By incorporating the HE opportunity cost of land used for alfalfa in our calculations, the calculated HE energy recovery for CON diets dropped from ~96 to 60%, and from 84 to 62% for HE protein recovery. It is therefore quite meaningful to consider land-use tradeoffs and the extent to which forage production could be shifted to human food crops in a given locale. Shortfalls with the land use approach are that fertilizer is needed to grow corn, row crops usually increase erosion, and the protein quality from the blend of corn and soybeans is not as great as milk protein.
In areas that are seasonally unsuitable for grain production but have moderate climates, some forage crops can grow well (Wilke and Snapp, 2008) while improving soil physical, chemical, and biological properties (Fageria et al., 2005). We utilized winter cover crop forage (triticale and clover hay) produced off-season on the same land used for a food-producing crop in a dual-cropping system (Salmon et al., 2004) in Experiment 2. It is noteworthy that winter cover crops may have either negative or positive effects on subsequent corn yield, depending on agricultural practices adopted. A meta-analysis of winter cover crops under agricultural practices in the USA and Canada showed that a bi-culture of winter cover crops increased corn yield by 21%; legume cover crops had an overall positive effect on corn yield, whereas grass winter cover crops had a neutral effect (Miguez and Bollero, 2005). Although winter cover crops are usually planted by farmers due to potential forage harvest and economic benefits, cover crops also protect soil from erosion, protect water quality (reduce losses of pesticides, nutrients, or sediments from fields), and promote carbon sequestration and nitrogen fixation (Dabney et al., 2001). On the other hand, cover crops involve additional inputs and risks. For instance, winter forage conditions in many regions are not ideal for crop dry-down at harvest (hence the moisture content of triticale/hay used in this experiment). Cover crops also require additional costs compared with a single crop system, must be planted when time and labor are limited, may increase risk of diseases, and can reduce soil moisture (Dabney et al., 2001).
To account for protein quality, we calculated the recovery of HE feed digestible amino acids in milk. As mentioned earlier, ruminants can convert non-protein nitrogen or protein with poor amino acid profile into a high-quality protein that is better able to meet human amino acids requirements (Virtanen, 1966). Grains tend to have poorer amino acid profile and digestibility in comparison with animal-based products (Wu, 2016). Thus, cereal-based diets are able to meet total protein requirements but may fail to meet essential amino acid requirements (Bouis et al., 2011). Furthermore, removing animals from US agriculture would likely lead to deficiencies in supplies of several nutrients for the US population, including Ca, essential fatty acids, and vitamins A and B (White and Hall, 2017). Vandehaar (1998) estimated a 20% greater nutritional value in the protein from milk compared to protein from a mix of corn and soybeans, due to both protein availability and amino acid profile. Ritchie et al. (2018) estimated a 20.6% deficiency in average Lys requirement across the global food system if meat and dairy products were removed from supply chains. The contrast between Lys content in plant- and animal-based diets has been described elsewhere (FAO/WHO/UNU, 2007; Swaminathan et al., 2012). In Experiment 1, calculations from the choice scenario on amino acid recovery showed that feeding ECO1 greatly increased the milk recovery of all amino acids, including digestible Lys, which was secreted in milk at 471× the amount of HE Lys that cows consumed. Because molasses was considered the only HE ingredient in the choice scenario, relatively high values of digestible amino acids recovery in milk were expected. Interestingly, the CON1 diet also supported a net gain in HE supply of most amino acids, with the exception of Arg and Cys. For example, milk Lys secretion by cows fed CON1 was 291× greater than the amount of HE Lys consumed by those cows. In agreement with the current study, Karlsson et al. (2018) described improved net production of several human-digestible amino acids (His, Ile, Leu, Lys, Thr, Trp, Val, Phe + Tyr) when replacing corn and soybean meal with by-products. Furthermore, the greatest improvement in net production of digestible amino acids was observed for Lys (Karlsson et al., 2018).
In Experiment 2, HE conversion rate was not necessarily improved by ECO diets. In the thrift scenario, ECO diets worsened HE ME and protein conversion, whereas in the choice scenario, ECO diets improved HE ME and protein conversion rates. The experiment 2 outcomes (at least the thrift scenario) oppose those reported in several studies (Ertl et al., 2015; Karlsson et al., 2018), mainly because of the negative impact of ECO diets on milk yield—which was not observed in the studies cited earlier. Although HE conversion rate was not necessarily improved by ECO diets, note that these diets theoretically do not use arable land and may present a higher efficiency of land use even with cows producing less milk during the lactation. For instance, Vandehaar (1998) estimated that milk protein produced by a dairy cow yielding 10,000 kg of milk per year when fed a diet without by-products requires more than twice the land base as a nutritionally equivalent supply of protein and energy from corn and soybeans produced for direct human consumption. On the other hand, a cow producing just 5,000 kg of milk annually while consuming 33% by-products increases the efficiency of land use to 76% relative to a corn and soybean system (Vandehaar, 1998). Decreasing the utilization or increasing efficiency of land use for crops is of great interest for sustainability of food systems (Peters et al., 2016).
It is important to note that our approach carefully accounts for HE inputs and outputs of specific nutrients in particular productive stages and food system scenarios but does not consider the feed required during non-productive stages of the cow's life nor the HE value of the cow carcass after culling. These are not trivial components of the overall dairy system; Thoma et al. (2013) attributed 13% of dairy farm greenhouse gas emissions to beef production, and Tichenor (2015) suggested that this may be an underestimate. Nevertheless, dietary strategies similar to those evaluated herein can also contribute to net HE protein production in beef systems (Baber et al., 2018), and it is likely that our comparisons would hold for net beef + milk production, assuming ECO diets were employed across a farm. Future studies employing a life-cycle assessment using our approach would provide a more comprehensive estimation of net production of HE nutrients by the dairy industry.
Another caveat of this study is that we focused on a single aspect of sustainability—the utility of dairy cattle in an efficient food system, focusing primarily on arable land as a constraint. There are, of course, many other aspects of sustainability to be considered in implementing ECO diet strategies on dairy farms, not least the profitability of the farm. It is likely that the ECO diets we tested would decrease the gross cost of diet ingredients compared to CON diets, though the lack of commercial markets for some ingredients prevented us from evaluating this. In fact, very few of the ingredients we utilized could be fed industry-wide due to supply constraints; lactating dairy cows in the U.S. consume more than 200,000 metric tons of feed annually. Regardless, the 28% decline in milk yield with ECO2 would almost certainly reduce revenue far more than feed costs, leaving little to cover fixed costs (i.e., infrastructure, labor, energy), and decreasing milk output per unit of these inputs. As is often the case, focusing on maximizing one narrow aspect of sustainability will likely harm sustainability in other ways.
In summary, HE nutrient conversion rate should be carefully analyzed, as values are highly dependent on whether individual feeds are considered HE and on the calculations performed (fixed coefficient for a group of ingredients vs. variable coefficients according to feed composition). Furthermore, ECO diets can be formulated using a wide range of ingredients depending on their availability and cost. As pointed out throughout this paper, diets with a relatively high inclusion of by-products can either improve, reduce, or maintain productive performance of animals. Under some scenarios, ECO diets do not necessarily improve HE feed conversion rate in dairy cows, and they will likely decrease feed efficiency of cows in terms of milk yield ÷ DMI. Regardless of diet, parity, and productive stage, cows typically generate a net return of HE essential amino acids in milk. These findings contribute to our understanding of how dairy production contributes to a sustainable global food system in alignment with competing environmental and development goals.
Conclusion
In this study, we showed that even with conventional diets on modern North American farms, the dairy industry is relatively efficient at using human-edible foods to produce milk and particularly in generating a net increase in digestible essential amino acids. Moreover, we showed that inclusion of ecological leftovers can be used to effectively decrease the use of human-edible foods, especially if foods that are not desirable by humans are not consumed by humans. This increase in human-edible nutrient recovery can occur even if milk production per cow declines.
Data Availability Statement
All datasets generated for this study are included in the article/Supplementary Material.
Ethics Statement
The animal study was reviewed and approved by Kansas State University Institutional Animal Care and Use Committee.
Author Contributions
CT, CY, TW, and BB contributed to the overall design and supervised research. CY and MS carried out the Experiment 1 and acquisition of data. CT, AB, and MD carried out Experiment 2 and acquisition of data. CT, CY, MV, TW, and BB designed and performed analyses on HE nutrient recovery, drafted and revised the first version of the manuscript. All authors reviewed and approved the final manuscript.
Funding
This work was a contribution (no. 20-053-J) from the Kansas Agricultural Experiment Station (Manhattan, KS), supported by USDA National Institute of Food and Agriculture (Washington, DC) Hatch project 1018048.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank Ajinomoto (Chicago, IL) and Adisseo (Antony, France) for amino acid product donations and Ajinomoto for partially funding Experiment 2. Authors would also like to thank Mike Scheffel and employees of the Kansas State University Dairy Teaching and Research Center (Manhattan, KS) for their assistance in completing this study. CT appreciates the scholarship (88881.170046/2018-01) provided by the Coordination for the Improvement of Higher Education Personnel (CAPES, Brasília, Brazil).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsufs.2019.00114/full#supplementary-material
References
Alexandratos, N., and Bruinsma, J. (2012). World Agriculture Towards 2030/2050: The 2012 Revision. Available online at: www.fao.org/economic/esa (accessed May 29, 2019).
Anderson, T., and Hoffman, P. (2014). Nutrient Composition of Straw Used in Dairy Cattle Diets. Available online at: https://fyi.extension.wisc.edu/forage/files/2014/01/StrawFOF.pdf (accessed May 29, 2019).
Atwater, W. O., and Bryant, A. P. (1900). The Availability and Fuel Values of Food Materials. Connecticut (Storrs) Agricultural Experiment Station 12th Annual Report.
Baber, J. R., Sawyer, J. E., and Wickersham, T. A. (2018). Estimation of human-edible protein conversion efficiency, net protein contribution, and enteric methane production from beef production in the United States. Transl. Anim. Sci. 2, 439–450. doi: 10.1093/tas/txy086
Baldwin, R. L., Donovan, K. C., and Beckett, J. L. (1992). “An update on returns on human edible input in animal agriculture,” in Proceedings California Animal Nutrition (Fresno, CA), 97–111.
Bouis, H. E., Eozenou, P., and Rahman, A. (2011). Food prices, household income, and resource allocation: socioeconomic perspectives on their effects on dietary quality and nutritional status. Food Nutr. Bull. 32, S14–S23. doi: 10.1177/15648265110321S103
Bounds, D. (2018). News Release National Agricultural Statistics Service Kansas 2017 Annual Crop Production Summary. Manhattan, NY. Available online at: http://usda.mannlib.cornell.edu/usda/nass/CropProdSu//2010s/2018/CropProdSu-01-12-2018.pdf (accessed May 21, 2019).
Bradford, B. J., and Carpenter, A. J. (2017). “Utilization of by-product and co-product feeds,” in Large Dairy Herd Management, ed D. K. Beede (Champaign, IL: American Dairy Science Association), 739–750. Available online at: https://www.adsa.org/ (accessed May 29, 2019). doi: 10.3168/ldhm.0854
Bradford, B. J., and Mullins, C. R. (2012). Invited review: strategies for promoting productivity and health of dairy cattle by feeding nonforage fiber sources. J. Dairy Sci. 95, 4735–4746. doi: 10.3168/jds.2012-5393
Bywater, A. C., and Baldwin, R. L. (1980). “Alternative strategies in food-animal production,” in Animals, Feed, Food and People: An Analysis of the Role of Animals in Food Production. (Boulder, CO: Westview Press for the American Association for the Advancement of Science), 1–30. Available online at: https://www.cabdirect.org/cabdirect/abstract/19801872772 (accessed May 29, 2019).
Capehart, T., Liefert, O., and Olson, D. (2018). Corn Disappearance Down as Quarterly Stocks Come in High. Available online at: https://www.ers.usda.gov/webdocs/publications/88426/fds-18d.pdf?v=43207 (accessed May 29, 2019).
Chaudhary, A., Gustafson, D., and Mathys, A. (2018). Multi-indicator sustainability assessment of global food systems. Nat. Commun. 9:848. doi: 10.1038/s41467-018-03308-7
Dabney, S. M., Delgado, J. A., and Reeves, D. W. (2001). Using winter cover crops to improve soil and water quality. Commun. Soil Sci. Plant Anal. 32, 1221–1250. doi: 10.1081/CSS-100104110
Dann, H. M., Tucker, H. A., Cotanch, K. W., Krawczel, P. D., Mooney, C. S., Grant, R. J., et al. (2014). Evaluation of lower-starch diets for lactating Holstein dairy cows. J. Dairy Sci. 97, 7151–7161. doi: 10.3168/jds.2014-8341
Ertl, P., Knaus, W., and Zollitsch, W. (2016a). An approach to including protein quality when assessing the net contribution of livestock to human food supply. Animal 10, 1883–1889. doi: 10.1017/S1751731116000902
Ertl, P., Zebeli, Q., Zollitsch, W., and Knaus, W. (2015). Feeding of by-products completely replaced cereals and pulses in dairy cows and enhanced edible feed conversion ratio. J. Dairy Sci. 98, 1225–1233. doi: 10.3168/jds.2014-8810
Ertl, P., Zebeli, Q., Zollitsch, W., and Knaus, W. (2016b). Feeding of wheat bran and sugar beet pulp as sole supplements in high-forage diets emphasizes the potential of dairy cattle for human food supply. J. Dairy Sci. 99, 1228–1236. doi: 10.3168/jds.2015-10285
Fageria, N. K., Baligar, V. C., and Bailey, B. A. (2005). Role of cover crops in improving soil and row crop productivity. Commun. Soil Sci. Plant Anal. 36, 2733–2757. doi: 10.1080/00103620500303939
FAO (2013). Dietary protein quality evaluation in human nutrition. Report of an FAQ expert consultation. FAO Food Nutr. Pap. 92, 1–66. Available online at: http://www.fao.org/ag/humannutrition/35978-02317b979a686a57aa4593304ffc17f06.pdf
FAO Food and Agriculture Organization of the United Nations (2009). “How to feed the world in 2050,” in How to Feed the World in 2050 (Rome: FAO). Available online at: http://www.fao.org/wsfs/forum2050/wsfs-forum/en/ (accessed June 07, 2019).
FAO/WHO/UNU (2007). Protein and Amino Acid Requirements in Human Nutrition: Report of a Joint FAO/WHO/UNU Expert Consultation. Geneva: The World Health Organization.
Ferraretto, L. F., and Shaver, R. D. (2015). Effects of whole-plant corn silage hybrid type on intake, digestion, ruminal fermentation, and lactation performance by dairy cows through a meta-analysis. J. Dairy Sci. 98, 2662–2675. doi: 10.3168/jds.2014-9045
Finucane, M. M., Stevens, G. A., Cowan, M. J., Danaei, G., Lin, J. K., Paciorek, C. J., et al. (2011). National, regional, and global trends in body-mass index since 1980: systematic analysis of health examination surveys and epidemiological studies with 960 country-years and 9·1 million participants. Lancet 377, 557–567. doi: 10.1016/S0140-6736(10)62037-5
Foley, J. A., Ramankutty, N., Brauman, K. A., Cassidy, E. S., Gerber, J. S., Johnston, M., et al. (2011). Solutions for a cultivated planet. Nature 478, 337–342. doi: 10.1038/nature10452
Fry, J. P., Mailloux, N. A., Love, D. C., Milli, M. C., and Cao, L. (2018). Feed conversion efficiency in aquaculture: do we measure it correctly? Environ. Res. Lett. 13:024017. doi: 10.1088/1748-9326/aaa273
Garnett, T. (2009). Livestock-related greenhouse gas emissions: impacts and options for policy makers. Environ. Sci. Policy 12, 491–503. doi: 10.1016/j.envsci.2009.01.006
Giuberti, G., Gallo, A., Masoero, F., Ferraretto, L. F., Hoffman, P. C., and Shaver, R. D. (2014). Factors affecting starch utilization in large animal food production system: a review. Starch 66, 72–90. doi: 10.1002/star.201300177
Guners, D. (2017). Wasted: How America is Losing Up to 40 Percent of Its Food From Farm to Fork to Landfill. Washington, DC: Natural Resources Defense Council, Issue Paper R10-05A.
Hall, K. D., Guo, J., Dore, M., and Chow, C. C. (2009). The progressive increase of food waste in America and its environmental impact. PLoS ONE 4:e7940. doi: 10.1371/journal.pone.0007940
Heinrichs, J., and Kononoff, P. (2013). Evaluating Particle Size of Forages and Tmrs Using the New Penn State Forage Particle Separator. University Park, PA. Available online at: www.das.psu.edu/teamdairy/ (accessed May 29, 2019).
INRA-CIRAD-AFZ (2002). The INRA-CIRAD-AFZ Tables. Available online at: https://www.feedtables.com/content/tables (accessed May 29, 2019).
Ivan, S. K., Grant, R. J., Weakley, D., and Beck, J. (2005). Comparison of a corn silage hybrid with high cell-wall content and digestibility with a hybrid of lower cell-wall content on performance of Holstein cows. J. Dairy Sci. 88, 244–254. doi: 10.3168/jds.S0022-0302(05)72682-5
Karlsson, J., Spörndly, R., Lindberg, M., and Holtenius, K. (2018). Replacing human-edible feed ingredients with by-products increases net food production efficiency in dairy cows. J. Dairy Sci. 101, 7146–7155. doi: 10.3168/jds.2017-14209
Kaur, G., Sharma, S., Nagi, H. P. S., and Dar, B. N. (2012). Functional properties of pasta enriched with variable cereal brans. J. Food Sci. Technol. 49, 467–474. doi: 10.1007/s13197-011-0294-3
Matte, J. J., Guay, F., and Girard, C. L. (2012). Bioavailability of vitamin B 12 in cows' milk. Br. J. Nutr. 107, 61–66. doi: 10.1017/S0007114511002364
McGuffey, R. K. (2017). A 100-year review: metabolic modifiers in dairy cattle nutrition. J. Dairy Sci. 100, 10113–10142. doi: 10.3168/jds.2017-12987
Miguez, F. E., and Bollero, G. A. (2005). Review of corn yield response under winter cover cropping systems using meta-analytic methods. Crop Sci. 45, 2318–2329. doi: 10.2135/cropsci2005.0014
Mottet, A., Teillard, F., Boettcher, P., De' Besi, G., and Besbes, B. (2018). Review: domestic herbivores and food security: current contribution, trends and challenges for a sustainable development. Animal 12, s188–s198. doi: 10.1017/S1751731118002215
National Research Council (NRC) (2001). Nutrient Requirements of Dairy Cattle. Washington, DC: National Academies Press.
National Research Council (NRC) (2012). Nutrient Requirements of Swine. Washington, DC: National Academies Press.
NCD Risk Factor Collaboration (NCD-RisC) (2016). A century of trends in adult human height. Elife 5:e13410. doi: 10.7554/eLife.13410
Oba, M., and Allen, M. S. (1999). Evaluation of the importance of the digestibility of neutral detergent fiber from forage: effects on dry matter intake and milk yield of dairy cows. J. Dairy Sci. 82, 589–596. doi: 10.3168/jds.S0022-0302(99)75271-9
Oltjen, J. W., and Beckett, J. L. (1996). Role of ruminant livestock in sustainable agricultural systems. J. Anim. Sci. 74:1406. doi: 10.2527/1996.7461406x
Pang, D., Yan, T., Trevisi, E., and Krizsan, S. J. (2018). Effect of grain- or by-product-based concentrate fed with early- or late-harvested first-cut grass silage on dairy cow performance. J. Dairy Sci. 101, 7133–7145. doi: 10.3168/jds.2018-14449
Perkins, E. G. (1995). “Composition of soybeans and soybean products,” in Practical Handbook of Soybean Processing and Utilization, ed D. R. Erickson (Urbana, IL: AOCS Press, 9–28.
Peters, C. J., Picardy, J., Darrouzet-Nardi, A. F., Wilkins, J. L., Griffin, T. S., and Fick, G. W. (2016). Carrying capacity of U.S. Agricultural land: ten diet scenarios. Elem. Sci. Anthr. 4:000116. doi: 10.12952/journal.elementa.000116
Poore, M. H., Moore, J. A., Swingle, R. S., Eck, T. P., and Brown, W. H. (1991). Wheat straw or alfalfa hay in diets with 30% neutral detergent fiber for lactating Holstein cows. J. Dairy Sci. 74, 3152–3159. doi: 10.3168/jds.S0022-0302(91)78500-7
Ritchie, H., Reay, D. S., and Higgins, P. (2018). Beyond calories: a holistic assessment of the global food system. Front. Sustain. Food Syst. 2:57. doi: 10.3389/fsufs.2018.00057
Romero-Huelva, M., Ramírez-Fenosa, M. A., Planelles-González, R., García-Casado, P., and Molina-Alcaide, E. (2017). Can by-products replace conventional ingredients in concentrate of dairy goat diet? J. Dairy Sci. 100, 4500–4512. doi: 10.3168/jds.2016-11766
Salmon, D. F., Mergoum, M., and Gómez Macpherson, H. (2004). “Triticale production and management,” in Tricale Improvement and Production, eds H. G. Macpherson, M, Mergoum (Rome: FAO), 27–36. Available online at: http://www.fao.org/3/a-y5553e.pdf (accessed May 29, 2019).
Schader, C., Muller, A., Scialabba, Nel-H, Hecht, J., Isensee, A., Erb, K. H., et al. (2015). Impacts of feeding less food-competing feedstuffs to livestock on global food system sustainability. J. R. Soc. Interface 12:20150891. doi: 10.1098/rsif.2015.0891
Schwab, C. G., and Broderick, G. A. (2017). A 100-year review: protein and amino acid nutrition in dairy cows. J. Dairy Sci. 100, 10094–10112. doi: 10.3168/jds.2017-13320
Shook, G. E. (1993). Genetic improvement of mastitis through selection on somatic cell count. Vet. Clin. North Am. Food Anim. Pract. 9, 563–577. doi: 10.1016/S0749-0720(15)30622-8
Sulc, R. M., Parker, A., Albrecht, K., Cassida, K., Hall, M., Min, D. H., et al. (2016). “Low lignin-alfalfa: wide rea field test results,” in Proceedings Western Alfalfa and Forage Symposium (Reno, NV).
Swaminathan, S., Vaz, M., and Kurpad, A. V. (2012). Protein intakes in India. Br. J. Nutr. 108, S50–S58. doi: 10.1017/S0007114512002413
Tedeschi, L. O., Muir, J. P., Riley, D. G., and Fox, D. G. (2015). The role of ruminant animals in sustainable livestock intensification programs. Int. J. Sustain. Dev. World Ecol. 22, 452–465. doi: 10.1080/13504509.2015.1075441
Thoma, G., Popp, J., Nutter, D., Shonnard, D., Ulrich, R., Matlock, M., et al. (2013). Greenhouse gas emissions from milk production and consumption in the United States: a cradle-to-grave life cycle assessment circa 2008. Int. Dairy J. 31, S3–S14. doi: 10.1016/j.idairyj.2012.08.013
Tichenor, N. (2015). Role of dairy in the carbon footprint of US beef. Proc. Natl. Acad. Sci. U.S.A. 112, E820–E821. doi: 10.1073/pnas.1421941112
Tricarico, J. M. (2016). “Role of dairy cattle in converting feed to food,” in Tri-State Dairy Nutrition Conference, 18-20 April 2016, Fort Wayne, Indiana, U.S.A. 25th Anniversary, 49–57. Available online at: https://www.cabdirect.org/cabdirect/abstract/20163250358 (accessed May 29, 2019).
United Nations (2017). World Population Prospects: The 2017 Revision, Key Finding and Advance Tables. Available online at: https://esa.un.org/unpd/wpp/Publications/Files/WPP2017_KeyFindings.pdf (accessed May 29, 2019).
Van Soest, P. J., Robertson, J. B., and Lewis, B. A. (1991). Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, 3583–3597. doi: 10.3168/jds.S0022-0302(91)78551-2
Vandehaar, M. J. (1998). Efficiency of nutrient use and relationship to profitability on dairy farms. J. Dairy Sci. 81, 272–282. doi: 10.3168/jds.S0022-0302(98)75576-6
VandeHaar, M. J., and St-Pierre, N. (2006). Major advances in nutrition: relevance to the sustainability of the dairy industry. J. Dairy Sci. 89, 1280–1291. doi: 10.3168/jds.S0022-0302(06)72196-8
Virtanen, A. I. (1966). Milk production of cows on protein-free feed. Science 153,1603–1614. doi: 10.1126/science.153.3744.1603
Voelker, J. A., and Allen, M. S. (2003). Pelleted beet pulp substituted for high-moisture corn: 1. Effects on feed intake, chewing behavior, and milk production of lactating dairy cows. J. Dairy Sci. 86, 3542–3552. doi: 10.3168/jds.S0022-0302(03)73959-9
White, R. R., and Hall, M. B. (2017). Nutritional and greenhouse gas impacts of removing animals from US agriculture. Proc. Natl. Acad. Sci. U.S.A. 114, E10301–E10308. doi: 10.1073/pnas.1707322114
Wildman, E. E., Jones, G. M., Wagner, P. E., Boman, R. L., Trout, H. F., and Lesch, T. N. (1982). A dairy cow body condition scoring system and its relationship to selected production variables in high producing Holstein dairy cattle. J. Dairy Sci. 65, 495–501. doi: 10.3168/jds.S0022-0302(82)82223-6
Wilke, B. J., and Snapp, S. S. (2008). Winter cover crops for local ecosystems: linking plant traits and ecosystem function. J. Sci. Food Agric. 88, 551–557. doi: 10.1002/jsfa.3149
Wilkinson, J. M. (2011). Re-defining efficiency of feed use by livestock. Animal 5, 1014–1022. doi: 10.1017/S175173111100005X
Wu, G. (2016). Dietary protein intake and human health. Food Funct. 7, 1251–1265. doi: 10.1039/C5FO01530H
Wu, G., Fanzo, J., Miller, D. D., Pingali, P., Post, M., Steiner, J. L., et al. (2014). Production and supply of high-quality food protein for human consumption: sustainability, challenges, and innovations. Ann. N. Y. Acad. Sci. 1321, 1–19. doi: 10.1111/nyas.12500
Ylioja, C., Abney-Schulte, C., Stock, R., and Bradford, B. (2018). Effects of fat supplementation to diets high in nonforage fiber on production responses of midlactation dairy cows. J. Dairy Sci. 101, 6066–6073. doi: 10.3168/jds.2017-13991
Zijdeman, R., and Ribeira da Silva, F. (2014). Life Expectancy at Birth (Total)—Clio-Infra Dataverse. Amsterdam: IISH Dataverse. Available online at: https://clio-infra.eu/Indicators/LifeExpectancyatBirthTotal.html (accessed June 20, 2019).
Keywords: arable land, by-product, feed efficiency, net food production, sustainability
Citation: Takiya CS, Ylioja CM, Bennett A, Davidson MJ, Sudbeck M, Wickersham TA, VandeHaar MJ and Bradford BJ (2019) Feeding Dairy Cows With “Leftovers” and the Variation in Recovery of Human-Edible Nutrients in Milk. Front. Sustain. Food Syst. 3:114. doi: 10.3389/fsufs.2019.00114
Received: 30 August 2019; Accepted: 28 November 2019;
Published: 19 December 2019.
Edited by:
Kathleen L. Hefferon, Cornell University, United StatesReviewed by:
Alexander Nikolov Hristov, Pennsylvania State University (PSU), United StatesWilhelm Knaus, University of Natural Resources and Life Sciences Vienna, Austria
Copyright © 2019 Takiya, Ylioja, Bennett, Davidson, Sudbeck, Wickersham, VandeHaar and Bradford. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Barry J. Bradford, YmpicmFkQG1zdS5lZHU=