 Benoît Bertrand
Benoît Bertrand Andres Mauricio Villegas Hincapié
Andres Mauricio Villegas Hincapié Lison Marie1,2
Lison Marie1,2- 1CIRAD, UMR IPME, Montpellier, France
- 2IPME, Université de Montpellier, IRD, CIRAD, Montpellier, France
- 3Impact Partners Fundation, Manizales, Colombia
So far, the main Arabica coffee breeding programmes in Latin America have focused on the selection of varieties adapted to intensive full-sun farming systems. Meanwhile, little attention has been paid to breeding varieties specifically adapted to shade, which is the main characteristic of agroforestry systems (AFS). Yet the specialty coffee sector is currently expanding and seeking specific sensory qualities related to exotic varieties and no breeding programme exists to create new varieties for this type of market. Two trials were set up: one in full sun and the other under shade. F1 hybrids and their parents (pure lines and Ethiopian accessions) were studied in a factorial-crossing design to measure tree volume, yield (3 years), bean size, the bean NIRS signature and the final cupping score. Bean size and the final cupping score seemed to be relatively unaffected by shading. Strong maternal heritability was observed for bean size. In the trials, F1 hybrids produced “75–80%” more than the maternal lines and “40–50%” more than the male parents in the shade trial and in the full-sun trial, respectively. By choosing the F1 hybrid, it is possible to increase productivity under both shade and full sun while simultaneously achieving good sensory qualities. Selecting a F1 hybrid for “specialty coffee” seems exceptional. This raises a fundamental issue concerning the maternal heritability of seed biochemical composition. We conclude that selection under shade is essential for the selection of varieties adapted to AFS.
Introduction
Coffee agroforestry systems (AFS) in which coffee is grown in association with other trees on the same plot of land are widespread in many tropical countries. When AFS are properly managed, they can buffer climatic fluctuations, and benefit from biological and economic synergies, leading to sustainable land management and higher and more stable incomes for local stakeholders (mainly smallholder farmers) (DaMatta, 2004). Under shade, coffee beans are denser and far more flavor intense, notably with fine acidity and a pleasant aroma (Muschler, 2001). Unfortunately, coffee AFS productivity can be up to 15–30% lower than in full-sun systems (Vaast et al., 2006). It is very difficult to maintain the ideal level of shade as regulating shade by pruning the branches of the shade trees is expensive, meaning the shade percentage is usually more than 40%. Another important reason put forward to explain the lower productivity of coffee in AFS is the fact that both dwarf and tall varieties developed for intensive full-sun systems are used even though they are not suitable for AFS cropping conditions (Bertrand et al., 2011).
In the present study, we are currently at the breeding stage, in which, like in most genetic breeding programmes, we are selecting pre-candidates we will then transplant to a farming environment to analyze G × E interactions, including interactions between coffee trees and shade trees. Interactions with shade trees are complex because, in addition to shade effects, evapotranspiration and root interactions must also be taken into consideration. In the first stage of breeding, it is difficult to compare small numbers accurately while also accounting for the effects of competition with shade trees. However, it is possible to mimic the effects of shading by using artificial shade. The first question we aim to answer is, is it possible to select genotypes that produce the equivalent quantity and quality under a high shade percentage (>50%), as in full sun?
Of the large volumes of coffees produced under AFS and full-sun systems, an ever-increasing quantity is being sold as “specialty coffee.” The Association of Fine Coffees (SCA, https://sca.coffee/), defines specialty coffee based on product quality (green beans, roasted beans or the prepared beverage) and by the quality of life that coffee can deliver to everyone involved in its cultivation (Rhinehart, 2009). Specialty coffees can be distinguished from mainstream coffee based on a variety of factors including quality, sustainability, and closer relationships with growers (Bacon et al., 2008). Unfortunately, the productivity of farms that produce so-called specialty coffees is not included. Specialty coffee roasters have understood that the choice of genotype has a huge impact on sensory quality and that old varieties such as Geisha or Typica or Bourbon are becoming keys to the market for the highest quality specialty coffees. Unfortunately, the varieties that produce the best coffees are also relatively unproductive or very susceptible to coffee rust and are therefore not very profitable for farmers (McCook and Vandermeer, 2015). On the other hand, the exceptional quality of a very small number of varieties is recognized. The second question we aimed to answer is, can new varieties be bred for the specialty market that combine exceptional quality and good productivity in shade and/or in full-sun conditions ?
To answer these questions, we conducted two trials, one in shady and one in full-sun conditions. To our knowledge, this is the first time that selection in full sun and selection under shade has been compared in coffee species but also in other species. We evaluated whether the performances were equivalent in the two trials. We used the same two factorial designs to compare the performance of pure Colombian lines, wild accessions from Ethiopia and F1 hybrids. A strong heterosis effect (20–70% increase in yield) has been observed when F1 hybrids were obtained by crossing dwarf American pure line varieties with wild Ethiopian accessions in full sun (Bertrand et al., 2005). In the present study, we wanted to check if heterosis is of the same amplitude in the shade. We assessed whether the selection was comparable between the two factorial trials and produced the same selected genotypes with constraints linked to three specifications. The first specification reflects the requirements of an intensive cultivation system close to that defined in the green revolution (small tree volume, large bean size and high productivity per hectare). Based on the different orientations in plant breeding proposed by Lammerts van Bueren et al. (2018), this orientation would be considered as “trait-based breeding.” The second specifications reflect the requirements of the Specialty Coffee association (https://sca.coffee/). This orientation would be considered “corporate-based breeding,” resulting in globally adapted? adaptable? cultivars that produce the highest profit. The third specifications reflect the requirements of agroforestry (productivity per hectare, tree volume, bean size, and beverage quality). This system would be considered as “Ecosystem-based breeding” aimed at developing varieties adapted to agroforestry, illustrated by the European Breedcafs project (https://www.breedcafs.eu/). The Breedcafs approach is based on the need to increase productivity without harming the environment and ensuring the sustainability of the agroecosystems (Campbell et al., 2014; Struik and Kuyper, 2014). The experimental design we used and results we obtained allow us to draw key conclusions regarding the selection of candidate varieties suitable for use in different farming systems and targeting different markets. Based on these conclusions, we make some recommendations for the organization of the coffee sector regarding breeding.
Materials and Methods
Planting Material and Crossing Design
Our study was conducted using a factorial crossing design involving crosses between wild and cultivated parents. Hybrids were obtained by crossing three dwarf cv. Castillo pure lines (Alvarado-Alvarado et al., 2005a), with 10 tall wild Ethiopian forms (hereafter referred to as “Ethiopian accessions”) pre-selected for yield and cup quality in the coffee collection established in Colombia. Crosses were established by directed pollination. Each cv. Castillo line received pollen from wild Ethiopian accessions. Table 1 lists the 28 F1 hybrids evaluated, including three maternal lines and 10 male parents.
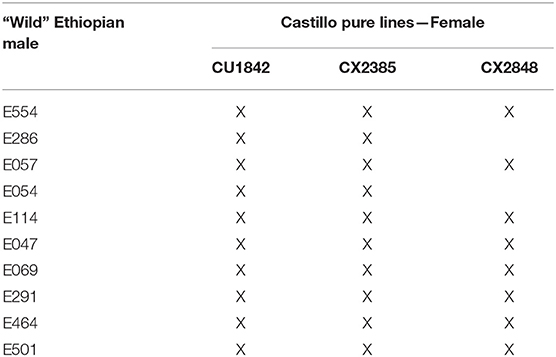
Table 1. Genotypes studied: 28 F1 hybrids (dwarf), 3 maternal pure lines (dwarf), 10 Ethiopian accessions (tall).
Two trials (hereafter referred to as “shade” and “sun”) were conducted with the same design, one in full sun and the other one in a plot under a net allowing only 55% of the photosynthetic photon flux density. It is technically complicated to work in AFS conditions in the first stage of the breeding programme, which involves choosing from a large number of crosses those that can be used in the following selection stage. In our study, we mimicked the effect of an agroforestry system by reducing light intensity, which is one of the main effects of AFS, using artificial shading. Another difficulty we had to solve is that it is not possible to compare dwarf and tall genotypes in the same plot. In both trials, dwarf genotypes, female parents and hybrids were compared in one sub-plot and Ethiopian tall parents were grown together in another sub-plot. The number of trees per genotype was 10 to 20 plants with total randomization within each sub-plot.
The plants were grown at the Naranjal experimental station in the municipality of Chinchiná—Caldas—Colombia (4° 58 ′17 45 “N latitude and 75° 39 ′09, 88″ W longitude), at 1,381 m asl. The minimum, average, and maximum temperatures were 17.4, 21.2, and 26.8°C, respectively. The average relative humidity was 84.2%, and the average annual precipitation was 3,568 mm.
The plantation densities were 1.3 m × 1.5 m (i.e., 5,128 trees.ha−1) for the dwarf plants and 1.5 m x 1.5 m (i.e., 4,444 trees.ha−1) for the tall plants (i.e., the Ethiopian accessions). The soils in which the plants were grown received 1,000 kg.ha−1year of N-P-K-Ca-Mg (18-3-10-8-0.5) and 250 kg.ha−1year of N. The plants were grown with zero pesticide applications.
Traits Observed
The coffee tree canopy volume was estimated at 24 months by comparing the shape of the tree to a cone. The radius (r) in cm, calculated by taking the average of the two largest plagiotropic branches, and the total height of the tree in cm, was used to estimate the conical volume V (cm3) = 1/3 × π × r2 × h (Bryant and Kothmann, 1979).
Yield was measured in kg/tree of fresh berries. Yield was estimated over three growing seasons (2013–2016).
The incidence of rust per plant was assessed in years 2 and 3 by visual inspection using a scale of 0–4, where 0 = absence of lesions; 1= sporulating lesions reaching 1 to 5% of the total leaf area; 2 to 3 = gradual increase in the number of diseased branches with sporulating lesions, and 4 = more than 50% of the leaf area affected (very susceptible cultivars may have lost their leaves before the observation date).
Samples of healthy ripe cherries belonging to each genotype were handpicked during the highest production period, between July and September 2014, in four harvest sessions. The cherries were then processed individually using the wet method (de-pulping, fermentation and drying) to obtain at least 200 g of green coffee beans with “11–12%” final moisture content. The green coffee samples were sieved (mesh size 14 to 20) according to the Colombian technical standard NTC 5248 (Instituto Colombiano de Normas Técnicas y Certificación [Instituto Colombiano de Normas Técnicas y Certificación (ICONTEC), 2004]. Beans smaller than sieve mesh size 15 were discarded along with defective beans. For each tree, we evaluated the size of the green beans from 16 to 20, as exportable coffee must be at least 16/64 inches).
The sensory evaluation was performed according to the Specialty Coffee Association (SCA) cupping protocol by the Coffee Growers Committee tasting panel in Valle del Cauca (Colombia), composed of five Q-Grader tasters. To avoid fatigue, a maximum of 7 samples were compared at a time. A standard session consisted of six sets, with seven samples per set and three cups of each sample. The standard session was performed twice in randomized order for blind analysis. The final cupping scores of each sample were obtained by summing the individual scores in the fragrance/aroma, flavor, aftertaste, acidity, body, balance, sweetness, uniformity, cleanliness, overall, and defects categories.
NIRS Phenotyping
The NIRS technique can be used to analyse coffee products as it can extract considerable information on the biochemical composition of the product. In a previous study (Posada et al., 2009), we showed that NIRS-based inter-variety relationships were stable across environments. In the present study, we tested whether the Euclidean distances between the NIRS signatures of the genotypes were the same under shade and in full sun.
NIR reflectance spectra were collected using a scanning monochromator NIR systems spectrophotometer (model 6500, Perstorp Analytical Inc., 1201 Tech Road Silver Spring, MD, USA) controlled by NIRS2 (4.0) software (Intrasoft Intl., LLC, RD109, Sellers Lane, Port Matilda, PA, USA). The analyses were performed on 50 gm aliquots of green coffee after grinding to <0.5 mm. Each sample represented one tree and each tree was analyzed. For each sample, an NIR spectrum was acquired in reflectance (R) mode, where R represents the reflectance energy in the 4,000–9,090 cm−1 range in 7 cm−1 steps. The log (1/R) absorbance spectrum was obtained by the mean of those measurements and comparison with the reference. The mean quadratic error, estimated from two subsamples (two distinct samplings of the same sample) based on the raw spectrum (log 1/R), was under 300 μabs, which is below the manufacturer's specifications and indicated satisfactory repeatability of the spectral measurement. Given these results, a single spectrum was acquired per sample.
Data Analysis
JMP, XLSTAT, and R software packages were used for the statistical analyses.
Comparison of Hybrids and Parental Populations
To evaluate the performances of the three populations (Ethiopian accessions, Castillo pure lines and F1 hybrid population), data for each population grown under shade and in full sun were analyzed using one-way analysis of variance (ANOVA), followed by a Tukey's test at P ≤ 0.05.
Comparison of Genotype Intra Trial and Inter Trial Variability
The performances of the genotypes in each trial were evaluated by ANOVA. To compare inter-trial performance, we calculated the correlation between the two trials using the LSmeans calculated for each genotype, and to compare the performance of the same genotype in the two trials, we used the non-parametric Wilcoxon rank-sum test.
Heritability Estimations
Analyses of variance of the full factorial mating design using a two-way cross-classification model were carried out according to the Henderson III procedure for shade and sun conditions. The R package lme4 was used (Bates et al., 2015). We used the following random effects model:
where μ is the general mean, Mi is the random effect of the ith female, Fj is the random effect of the jth male, (MF)ij is the female x male interaction, and Eijkis the within-family deviation of the kth individual within the(ij)th female x male combination. We estimated the variance components (including maternal and paternal variance) and narrow-sense heritability for the main traits (yield, final cupping score, bean size).
The estimated phenotypic variance is free from any genetic model and is given by:
As usual, the estimates of narrow-sense and broad-sense heritability are given by:
and
Stability of the Genotype Performances Across Plots
Chemometric processing initially consisted of a principal component analysis (PCA) based on the spectra on the 4,000–9,090 cm−1 segment. Factorial scores of principal components (PCs) showing an eigenvalue higher than 1 were used to create clusters using the Euclidean distance and the k-means algorithm (Posada et al., 2009). We tested the number of clusters to be created, i.e., from 2 for sun to 6 for shade conditions, to test the robustness of the variety types? We compared two approaches to test the stability of the performances of the coffee variety across plots, the final cupping score and the NIRS signature. The pseudo r of correlation was calculated by comparing the two Euclidean distance matrices using a Mantel test, along with comparison of the associated probabilities. The procedure that gave the highest pseudo r associated with the greatest probability was considered to be the most efficient.
Selection Based on Farming System Specifications
In the first approach, selection was based on specifications for intensive full-sun farming systems (which correspond to the conventional specifications used by the main coffee breeding programme in Latin America). Selection was based on three traits: tree volume, yield and bean size. In the second approach, selection was based on the final cupping score and productivity, which is the main trait of interest for the specialty coffee market. In the third approach, selection was based on AFS specifications with the following threshold: yield under shade at least equivalent to the best pure line grown in full sun; a final cupping score higher than 83/100; a tree volume lower than 40% of the largest pure line grown in full sun; a green bean size at least equivalent to the performance of the maternal line producing the smallest beans.
Clustering Genotypes in the Two Plots for Further Selection in Different Farming Systems
Rather than select the genotypes, we clustered the genotypes based on the Euclidean distance between varieties based on the yield, bean size, tree volume and the NIRS signature values. The information of the NIRS spectra was reduced by using the scores of genotypes on the main first factor resulting from a discriminant analysis of the six main PCs.
Results
Performance of Parent Populations and Hybrids
The performances of the three populations were monitored in the shade and full-sun trials. The average production under shade was more than 40% lower than in full sun.
In both tests, the hybrids gave the best performance (Figure 1). They produced 75–80% more than the maternal lines and 40–50% more than the male parents in the shade and sun trials, respectively.
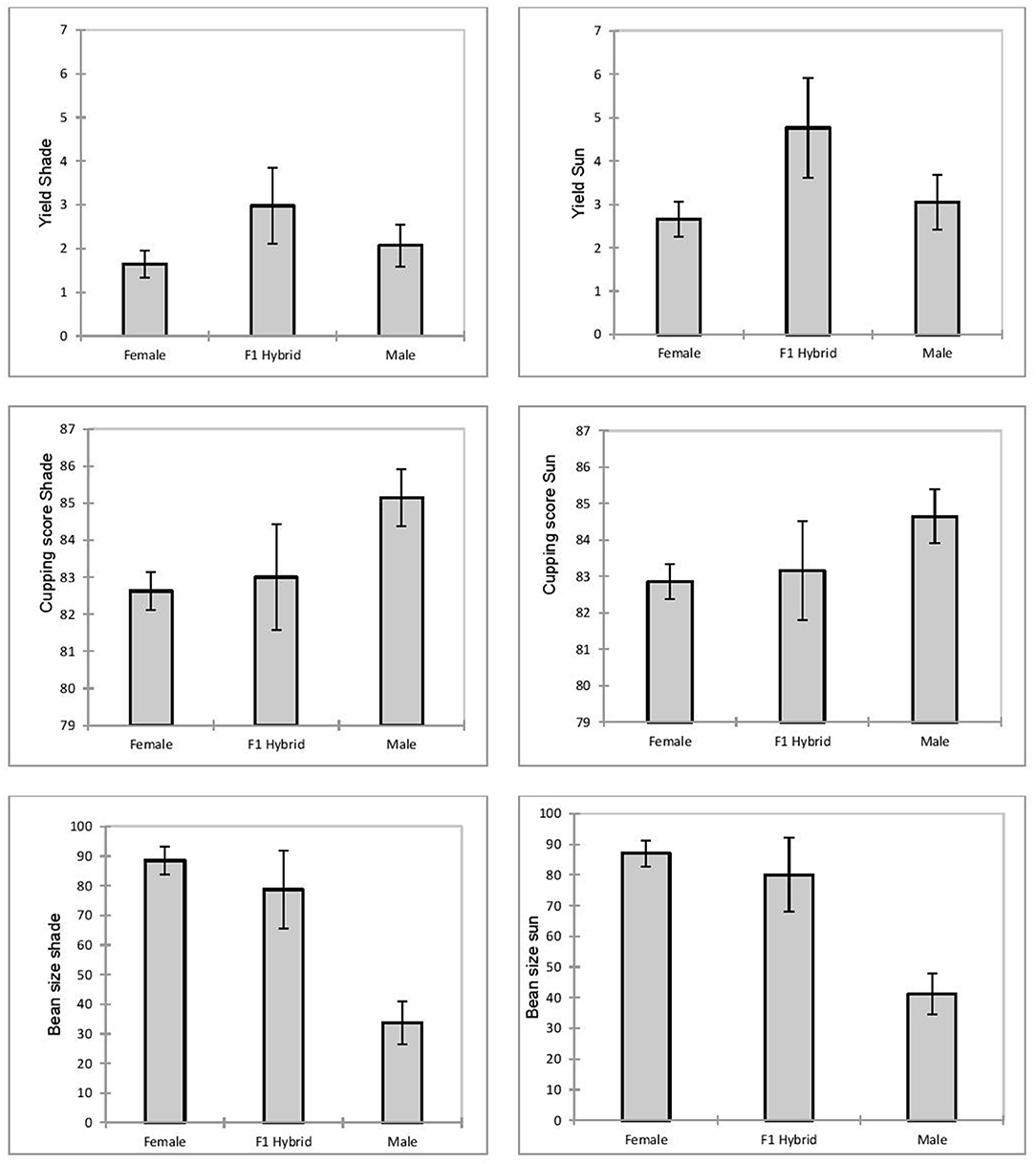
Figure 1. Comparison of yield (expressed as kg of green coffee beans per plant), final cupping score (out of 100), bean size (percentage of bean size 16–20), in female, male and F1 hybrids in shade and full-sun plots.
The seed produced by females and F1 hybrids were significantly larger than those produced by Ethiopian males in both shade and sun trials. However, no significant differences between females and F1 hybrids were observed in the two trials. The tree volume of the male parents appeared to was much larger than that of the hybrids or female lines. There was no significant difference in tree volume between the female lines and F1 hybrids in the shade and sun trials; nor was there any difference in tree volume between female lines (dwarfs) and F1 hybrids (also dwarfs) in the two conditions.
Surprisingly, female cultivars that had been specially selected for rust resistance showed significantly more symptoms than the Ethiopian genotypes or F1 hybrids. The final cupping score of males was significantly higher (P < 0.01) than the scores of F1 hybrids and female lines in both the shade and sun trials. No significant differences were found between hybrids and females in the two trials.
Comparison of Genotypic Performance Between the Two Trials
The linear regression for bean size genotypes under shade relative to the bean size in full sun was significant (P < 0.001) and highly predictive, with an r2 of 0.68. For yield, the regression was significant (P < 0.01), with an r2 of 0.42, even though this cannot be considered as predictive. We compared the performance of each genotype between shade and full sun (Data sheet 1) using the non-parametric Wilcoxon rank-sum test. Most genotypes produced significantly higher yields in full sun than under shade. We identified five genotypes whose performance did not differ significantly in the two conditions (E464, E057, E054, E047, and CX2385 × E291). The last two produced the same quantity of cherries in the shade as in the sun. Concerning the size of the beans, the performances of the great majority of the genotypes were identical in full sun and under shade. Seven genotypes (E501, E291, E286, E114, E054, E057, and CX2385 × E057) showed a significant difference in favor of sun. Finally, the Ethiopian E047 and E554 produced significantly larger seeds under shade than in the full sun. The tree volume of all the genotypes was identical under shade and in full sun, except for the hybrid CX2848 × E114 whose tree volume was larger under full sun and the majority of the Ethiopian genotypes (E 464, E069, E291, E057, E114, E286, E047, E054) whose tree volumes were bigger when grown under shade (data not shown). Regarding the final cupping score, four genotypes scored better in full sun (CU1842, CU1842 x E464, CU1842 × E501, and the Ethiopian E501), while five genotypes grown under shade (E464, E069, E047, and the hybrids CX1842 × E069 and CX2385 × E069) often received higher scores.
Heritability of Main Traits
The regression of the mid-parent on hybrids for bean size was significant under shade (r2 = 0.25) and null in full sun (r2 = 0.06). However, we observed a strong maternal effect of this trait with a maternal heritability of between h2 = 0.69 and 0.37, respectively, for shade and sun.
For the final cupping score, we observed a significant r2 under shade and a non-significant r2 in full sun (0.25 and 0.15, respectively) between the mid-parent performances and the hybrid performances. Female and male heritabilities for the final cupping score were close to zero, ranging between h2 = 0.04 and 0.10 in full sun and under shade. Finally, the regression for the mid-parent regarding observed hybrid yield was null under shade and in full sun. The maternal or paternal heritabilities for yield were close to zero (h2 female = 0.00 and 0.11, respectively, under shade and in full sun and h2 male = 0.07 under shade and 0.23 in full sun). For yield, the mid-parent value did not appear to be a useful guide for the selection of parents to test combinations of hybrids.
Comparison of the Stability of the NIRS Genotypic Signature Under the Two Conditions
The PCA based on all the NIRS spectra in the sun trials produced a similar pattern. The first axis (PC1) separated the majority of the Ethiopian parents from the maternal parents (Castillo pure lines) (Supplementary Figure 2). The F1 hybrids were located midway between the two parental populations in full sun or under “shade.”
We then performed a K-means analysis based on the first 15 PCs. We established three robust clusters corresponding to the three sub-populations (namely Ethiopian accessions, Castillo lines and F1 hybrids) confirming the PCA findings. We then used discriminant analysis to explain and predict the membership of observations according to their sub-groups. The first 15 factors that explained more than 99% of the total variance and had eigenvalues of over one were used to calculate the discriminant function models. We obtained a well-classified rate of around 90%, i.e., 91% per type in full sun and 90.3% under shade. We found same clustering in three subgroups for the NIRS signature. This stability shows that the two environmental conditions (sun and shade) did not significantly affect the interrelations between the NIRS signature of the genotypes.
Comparison of the Stability of the Euclidean Distance Matrices Under the Two Conditions
We compared the two distance matrices, sun and shade, using a Mantel test on the Euclidean matrix distances between genotypes using the factorial scores of the four PCs calculated in (A) on the phenotype values for yield, bean size, tree volume and final cupping score; in (B) on the phenotype values for yield, bean size, tree volume and NIRS signature. For the NIRS spectra, we used the genotype scores on the main first factor derived from a discriminant analysis of the 15 main PCs. We showed that the pseudo r of correlations obtained by comparing the two distance matrices based on a Mantel test and p-value was highly significant in both cases. Hence, the relative distances between genotypes were highly stable in both conditions when we used phenotyping on the four target traits (yield, bean size, tree volume and final cupping score) or an “extract” of the NIRS signature instead of sensory analysis. Use of the NIRS signature produced a higher result than that obtained using sensory analysis since the r obtained was higher (pseudo r = 0.801 using the NIRS signature vs. r = 0.716 using the final cupping scores).
Selection for Full Sun
The selection of the best candidates was based on a set of specifications whose priorities differ depending on the farming system. From previous results, it appeared? Previous result showed? that the female varieties, which are among the best lines currently disseminated in Colombia, were distinguished from the other genotypes by the remarkable size of their seeds. Indeed, this was a key criterion put forward by breeders in Colombia, the other criterion being tree volume, to adapt the varieties to an intensive full-sun system, which is the predominant farming system in Colombia. The three female varieties had a small volume, i.e., a trait that is very highly selected by breeders in Latin America. Highlighting this trait along with bean size revealed two distinct groups: females and F1 hybrids on the one hand, and male genotypes on the other in the full sun plot and the shade plot (Figure 2). Applying the same selection to both tests produced different results. Under shade, we selected the three pure lines and six F1 hybrids and under full sun, the same three pure lines and 10 F1 hybrids. Five of the hybrids were the same and five were different. In the full-sun selection scenario, the F1 hybrids (which inherited the dominant rust resistance genes from their maternal parent) produced 60–80% more than the best female line (CU1842). Regarding the final cupping score, four F1 hybrids had higher scores than the best maternal lineage (CU1842). In particular, the F1 hybrid “CU1842 × E047” which produced 40% more than the best female line, with a comparable tree volume and with a final score of over 85 and a bean size over 85%, appears to be exceptional.
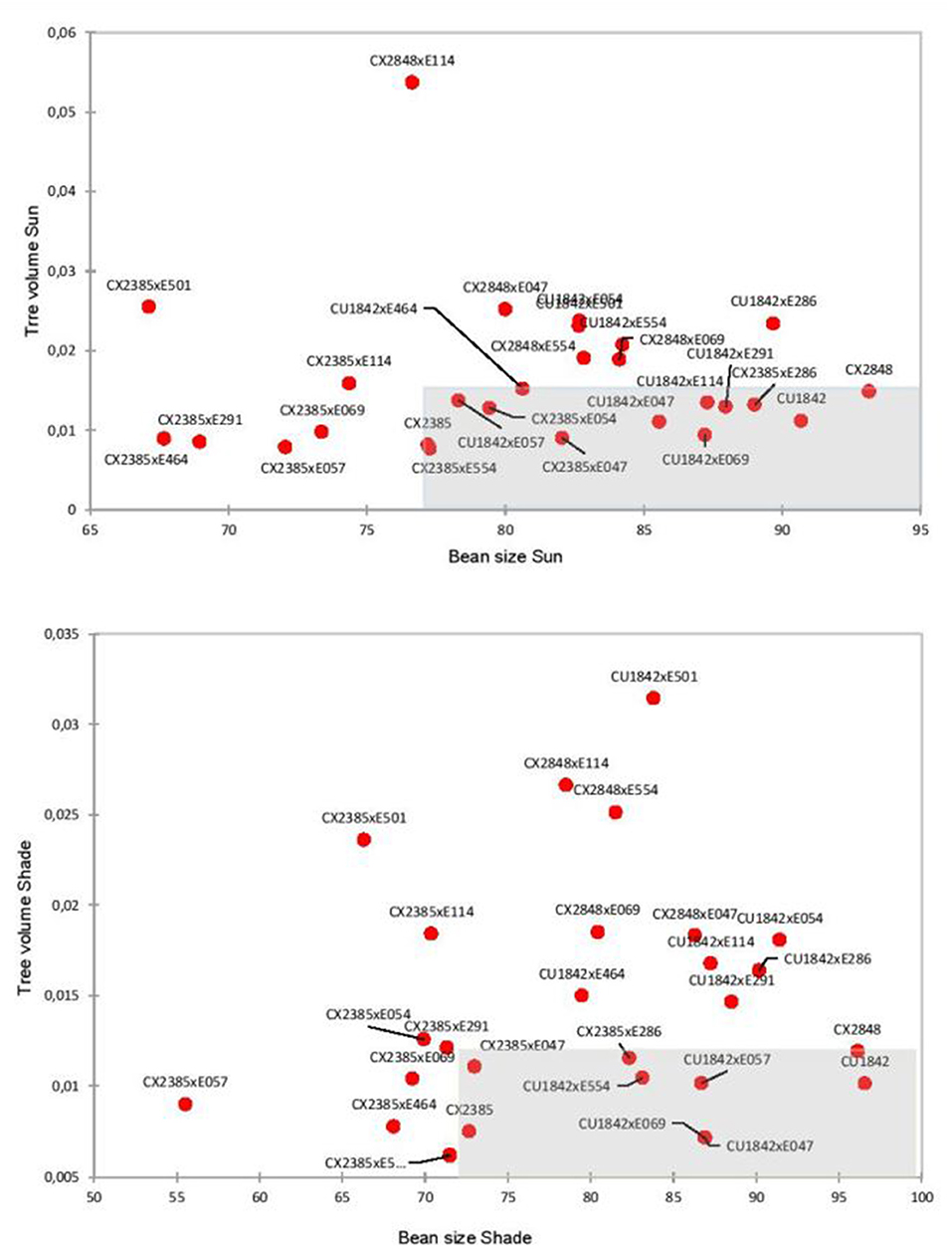
Figure 2. Selection for a “full-sun” farming system. Scatterplots showing tree volume and bean size. The F1 hybrid population and the population of female lines (three components of cv. Castillo) are represented. The genotypes inside the rectangle were selected because they produced similar performances in terms of tree volume and bean size to those of the maternal lines. Upper graph, selection in the full-sun plot, lower graph, in the shade plot.
Selection for Specialty Coffee
We used the Specialty Coffee Association's definition to define specialty coffee specifications. Through cupping, coffee tasters assess a coffee score and determine whether it is a specialty grade quality. For breeders, this means tracking down genotypes that produce the highest cup quality scores. In this definition, all the other traits are secondary. However, we considered that productivity is also an important criterion for farmers, so we included it in the two figures that deal with the selection of specialty coffee candidate varieties.
It is known that coffee quality is closely linked to environmental conditions and particularly to elevation (Bertrand et al., 2012). In the present study, selection was made at an elevation that could not generate very high scores (i.e., over 90), and we only selected genotypes with a final cupping score of over 85. Applying the same selection in full sun and under shade, produced quite different results. Three Ethiopian genotypes (E501, E286, E057) and three F1 hybrids stood out when selected in the sun trial (Figure 3A). These three F1 hybrids all had the same mother (CU1842). The productivity of the Ethiopian E286 was of the same order of magnitude as that of the F1 hybrids. The E286 genotype produces more than 4.5 kg of cherries per tree. This “wild” genotype can therefore produce a lot of very good coffee in full sun conditions. However, productivity has to be related to hectares. The tree volume was 3–4-fold higher. In practice, this would mean that the planting density would have to be reduced twofold (1,800–2,500 trees/ha for E286 compared to 4,000–5,000 trees/ha for the CU1842 × E047 F1 hybrid). One would thus expect half the productivity per hectare compared to the best hybrids (respectively, 2,000 kg vs. 4,800 kg of green coffee).
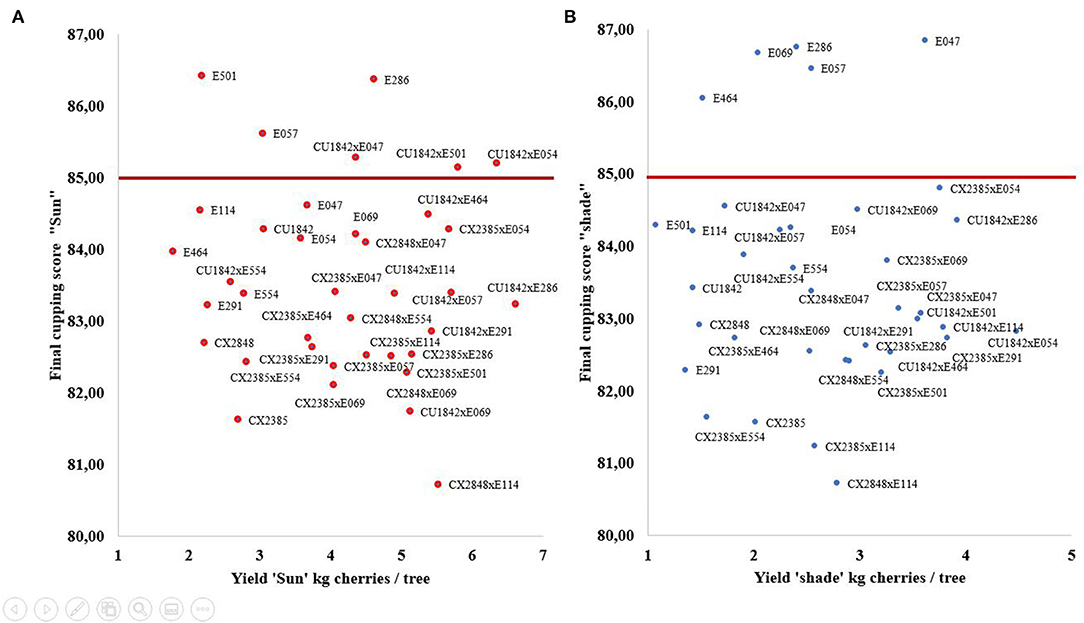
Figure 3. Selection for “specialty coffee.” Scatterplots showing the final cupping score and yield. The genotypes inside the rectangle were selected because they produced higher performances than the threshold (blue line) in terms of the final cupping score. (A) selection in the full-sun trial; (B) in the “shade” trial.
In the shade plot, only five Ethiopian genotypes were selected (Figure 3B). The E047 genotype produces more than 3.5 kg of cherries per tree. This “wild” genotype can therefore simultaneously produce a lot of coffee and very good coffee under shady conditions. Again, it is important to link this productivity to hectares. One would expect half the productivity per hectare compared to the best hybrids (respectively, 1,600 kg vs. 3,200 kg of green coffee).
Selection for “AFS”
There are no recognized specifications for this farming system. We believe it would be important to select the most productive genotypes, with large bean sizes, the best sensory scores and volumes compatible with crop densities that would allow high yields per hectare. Farmers who grow coffee in AFS are in competition with growers who cultivate coffee in full-sun conditions because coffee purchase prices do not take agroforestry into account. We thus set the yield threshold at the yield level of the best Castillo lineage grown in full sun, i.e., genotypes selected in the shade plot would have to produce at least as much as the best Castillo pure line grown in the full sun plot. For the final cupping score, as the best Castillo lineage achieved a final cupping score of 83.42 under shade, we set the threshold at 83. Regarding the seed size, the performance had to be at least that of the worst Castillo line. Regarding tree volume, we set a tree volume under shade that could not be more than 40% bigger than that of the largest female in full sun.
When this selection was applied, we retained two F1 hybrids whose characteristics are listed in Table 2. In shade conditions, the two selected hybrids (CX2385 × E054 and CU1842 × E286) produced, respectively, 22 and 2% more than the best pure line in full sun conditions. Both hybrids have a cup quality similar to that of the pure line. The size of the beans produced by one of the hybrids is smaller than the pure line while the size of the beans produced by the other is comparable with the pure line.
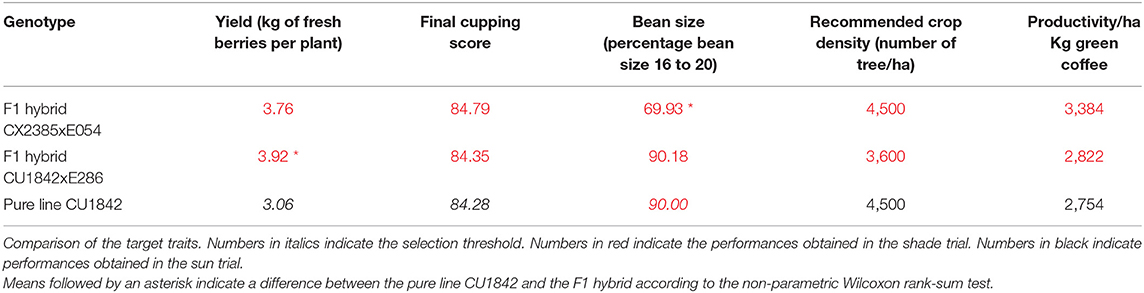
Table 2. Performances in the shade plot of hybrids selected under shade, compared to the performances of the best maternal pure lines in full sun.
Clustering Genotypes for Further Selection in Suitable Farming Systems
At this early stage of preselection, the breeder can consider that the aim is to cluster tested genotypes rather than select them, which may be useful to subsequently dispatch the groups to coffee breeding hubs and/or to multi-location variety trials to test genotype x environment interactions. Genotypes may be selected across all traits by pooling them in groups based on common traits. This helps avoid an a priori bias. We tested this approach in both the sun and shade trials. This enabled us to determine if it would be useful to practice two types of pre-selection (under shade or in full sun) at this stage of selection. Clustering was based on yield, bean size, tree volume and the NIRS signature. The NIRS signature can be considered a preferable to costly sensory analysis at this stage of selection and non-selection context. To reduce the NIRS signature at one variable, we used the scores of the genotypes for the main first factor resulting from a discriminant analysis of the 15 main PCs.
We obtained three groups in the two plots (Figure 4), a group with male parents, another with the majority of the hybrids, and a final group with the Castillo pure lines and some hybrids. The male parents grouped together both under shade and in full sun. This cluster remained stable in any environment. The “F1 hybrid cluster” and the “pure line cluster” differed substantially under shade and in full sun conditions.
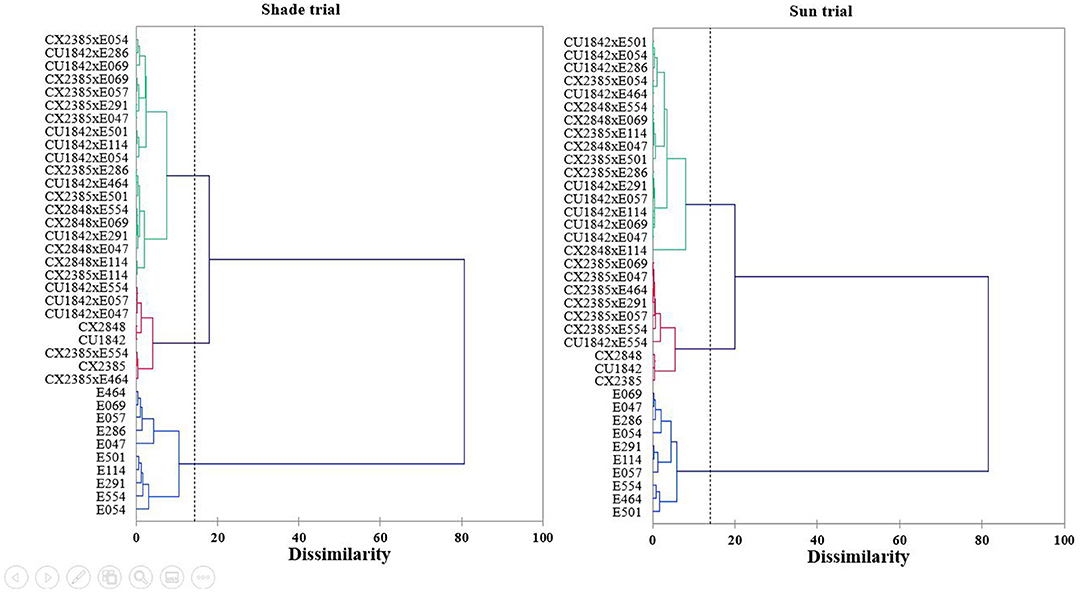
Figure 4. Hierarchical cluster analysis (Euclidean distance, Ward's grouping method) resulting from the discriminant analysis of the 38 varieties studied based on yield, bean size, tree volume and NIRS signature, under shade and in full sun conditions. The upper graph corresponds to shade conditions and the lower graph to full-sun conditions.
The majority of hybrids (16) formed the same group under both conditions, indicating comparable performance in terms of productivity, seed size, tree volume and biochemical composition. However, in full sun conditions, seven hybrids grouped with the Castillo lines while four hybrids grouped with the Castillo lines in the shade. Only three hybrids (CX2385 x E464, CU1842 × E554 and CX2385 × E554) clustered with the Castillo lines in both conditions, suggesting that the performances in terms of yield, bean size and biochemical composition of those F1 hybrids are close to the maternal pure line whatever the environment. Five hybrids changed clusters depending on the environment. These genotypes are therefore particularly adapted to one or the other of the two environments.
Discussion
Evidence of the effects of increasing atmospheric levels of greenhouse gases has become clearer and more convincing (IPCC, 2013, 2014). After very alarmist studies that predicted drastic drops in coffee production (Bunn et al., 2015; Ovalle-Rivera et al., 2015), a recent study claims that coffee crops could survive climate change and global warming to a greater extent than previously estimated (DaMatta et al., 2019). It is therefore very difficult to predict the genotypes that should be used to best adapt to climate change.
For the selection of varieties adapted to climate change and global warming, the solution is undoubtedly to select genotypes for a wide range of scenarios at the same time by testing future varieties in as many environments as possible (i.e., by favoring genotype x environment studies). However, in tree breeding, in the first stage, genotypes are tested in a single environment to avoid increasing costs. The risk is to bias this first stage by selecting genotypes adapted to this particular environment. Here we chose to carry out the first stage of the selection in full sun and using artificial shading to mimic one of the main effects of AFS systems.
The Performances of the Cultivated and “Wild” Accessions Differed Markedly
We distinguished the following genotypes according to their origin and mode of reproduction. It was extremely interesting to compare the group of so-called wild Ethiopian genotypes with the three cultivated lines selected in Colombia. The three latter lines are clearly representative of the breeding initiatives that have been underway in Latin America for more than 50 years in response to two imperatives, reduce the size of the plant to enable a higher density per hectare and to select varieties that are resistant to orange rust.
The average tree volume of the lines from Colombia was 4- to 5-fold smaller than the original accessions from Ethiopia, illustrating the effectiveness of selection efforts. This reduction in tree volume enabled cultivation densities of 4,500 to 6,000 trees/ha compared to only 2,500 to 3,000 for the Ethiopian accessions. The reduction in plant volume is one of the pillars of the Green Revolution in cereals (Elias et al., 2012). Arabica coffee is no exception to this rule. In Arabica, the reduction in volume is based on a single gene that confers dwarfism. Dwarfism has also had substantial benefits in fruit tree production, enabling higher yields, and facilitating harvesting in orchards (Battistini and Battistini, 2005). In the case of Arabica, productivity per plant has not been increased as the productivity of the Castillo pure line was lower than that of the Ethiopian, both under shade and in full sun conditions. It is therefore productivity per hectare that has increased simply because of plant dwarfism, thereby confirming the findings of Moncada et al. (2019), who suggest that the yield potential of various Ethiopian accessions exceeds that of any pure line variety cultivated in Colombia today.
Regarding quality, the Castillo lines had large beans, which is a characteristic sought after by coffee buyers, whereas the Ethiopian accessions had much smaller bean sizes that are not compatible with the usual commercial standards. However, many Ethiopians accessions do have large beans. It turns out that the selection of the Ethiopian accessions used in this study did not take this criterion into account. In comparison, the homozygous lines of Castillo have very large seeds. In fact, selection on this trait can be highly effective, as its maternal heritability is high.
Ethiopian accessions in our study produce small beans but remarkable sensory quality. Indeed we provide evidence that bean size is not correlated with sensory quality, rather, it is the weight of 100 healthy green beans (g) that is correlated with sensorial quality (Marie et al., 2020).
On the other hand, the sensory quality of the Castillo pure lines appeared to be quite good: one of the lines (CU1842) had a final score over 84 both in full sun and under shade. There was no mention of off-type flavors during the tasting (data not shown), which confirmed that breeders have been able to circumvent the negative effects of introgression of chromosome fragments that have been reported in some Arabica varieties derived from the Timor hybrid (Bertrand et al., 2003). However, we noted that the selection resulted in a sensory quality inferior to that of most of the Ethiopian accessions. Commercial varieties produced and released in Colombia to date have been selected mainly for their high yield and resistance to leaf rust, while cup quality has been evaluated by discarding progenies whose cup quality differs from that of Typica, Bourbon and Caturra. There is therefore considerable room for improvement of this agronomic trait.
The Performances of the F1 Hybrids Are Far Beyond Exceed the Productivity of Both Parents
The performance of the F1 hybrids was remarkable in terms of productivity. They were much superior to the Ethiopian parents or cultivated lines, both in full sun and under shade. This confirmed our previous findings (Bertrand et al., 2011; Marie et al., 2020) in studies carried out using a factorial design or in farmers' plots. The very high productivity of the F1 hybrids is due to the heterosis between the two parental populations (Bertrand et al., 2005). Although Arabica is a self-pollinated organism, a 30–80% heterosis level seems particularly high for an autogamous species, especially since a recent study (Scalabrin et al., 2020) showed that this species was responsible for upsetting a very recent allo-polyploidization event on the evolutionary scale (10,000 to 20,000 years ago), and since this species has been found to have extremely low levels of variation as a consequence of the allopolyploidization event. Unsurprisingly, it was not possible to predict the heterosis by the value of the average parent. The heritability of the yield, estimated in both environments, was close to zero. The genetic and molecular basis of heterosis remains elusive (Schnable and Springer, 2013). For a species as little polymorphic as the Arabica, our hypothesis favors complementarity of the parents in the regulation systems.
Bean Size and Sensory Quality Are Maternal-Like for Other Traits
The bean size of the F1 hybrids was close to that of the maternal lines. We observed a strong maternal effect for this trait in both full sun and shade conditions. The F1 hybrids generally inherited the best characteristics of the Castillo maternal lines. The seed size could be quite accurately predicted under shade based on the value of the maternal genotype. Finally, the tree volume of F1 hybrids was also close to that of the maternal lines. The dwarfism gene in cv. Caturra was dominant and was expressed in all the F1 hybrids.
In terms of sensory quality, the results obtained for the F1 hybrids were quite close to those of the maternal lines, but with greater variability, suggesting that this trait has selection potential. This is consistent with the findings of a previous study (Bertrand et al., 2006). There was no apparent heterosis for this character, but it would be hard to predict it based on the average value of both parents and heritability was not significant. The fact that the F1 hybrids were closer to the maternal lines than to the Ethiopian accessions suggests that, like bean size, maternal heritability was strong for this trait. However, the fact that the hybrids grouped between the two parental groups based on the NIRS signatures strongly suggests that biochemical composition is predominantly additive (Supplementary Figure 2). We now need to confirm whether the direction of the cross has a strong impact on sensory quality.
Partial Rust Tolerance
Castillo varieties that have been specially bred for rust resistance were found to have a higher incidence of the disease than accessions from Ethiopia. Ethiopian accessions carry resistance genes (SH1, SH2, SH4, SH5) that are all overcome by the rust strains found in Colombia. Colombia lineages (including cv. Castillo) derived from the Timor hybrid, which bears SH6 to SH9 genes, are expected to have incidence levels close to zero. The levels achieved here showed that the fungus was able to overcome the resistance genes SH6 to SH9 in all three lines. In fact, several varieties developed in Brazil using different sources of rust resistance are today completely susceptible or only partially resistant (Zambolim, 2016). The situation is the same throughout Latin America where the rust races have overcome resistance genes deployed in the varieties derived from the Timor hybrid. This is becoming a major concern in the coffee sector. In this study, we were not surprised to find that the vertical resistance of the three lineages of Castillo has been overcome. It is known that once this so-called vertical resistance is overcome, a good dose of partial resistance may remain. According to Alvarado-Alvarado et al. (2005b), over 80% of the genotypes involved in the Castillo variety have incomplete resistance and the three lines chosen for our study shared this feature. Surprisingly, we found that this partial resistance was lower in the Castillo lines than in the Ethiopian accessions and in the F1 hybrids.
How should we interpret the greater tolerance of Ethiopian parents and F1 hybrids to rust compared to homozygous lines? Is it due to persistent heterozygosity in the Ethiopians and a fortiori in the F1 hybrids? Is it due to a better leaf/fruit balance, as observed by Echeverria-Beirute et al. (2018) and or better plant health (“good plant health,” a concept revisited by Döring et al. (2012). We developed this health concept in a recent work (Toniutti et al., 2017; Echeverria-Beirute et al., 2018). Indeed, the incidence of rust in susceptible varieties increases with the fruit load relative to total tree biomass (Avelino et al., 2006; Echeverria-Beirute et al., 2018). This suggests that breeding of homozygous lines for productivity and dwarfism could lead to the creation of varieties with reduced plant health.
Comparison of Performances Under Shade and Full-Sun Conditions
The shade imposed on the plants in our study was homogeneous and strong. The coffee yield was reduced by 40% when more than 55% of the natural light was filtered out. In a study with a similar design in which 45% of the natural light was filtered out, shade was reported to reduce coffee tree productivity by 18% (Vaast et al., 2006). Jaramillo-Botero et al. (2010) found that trees under 48% shading yielded 20% less than trees grown in full-sun conditions. In Brazil, coffee productivity was found to be reduced by 15% in coffee plantations of Grevillea robusta trees grown under shade (Baggio et al., 1997). In natural conditions, full-sun coffee production was 30% higher than that of shaded coffee in Costa Rica, while in Nicaragua full-sun coffee production was equal to that of shaded coffee (Haggar et al., 2011). Coffea arabica is an understorey plant in its area of origin. By nature, this species tolerates high levels of shade and shade cannot be considered as a suboptimal environment. However, in the present study, the impact of shading on production levels was high probably because our artificial shading was both strong and uninterrupted.
Bean size and final cupping score seemed to be? was? relatively unaffected by artificial shading. There were disparities between genotypes, suggesting there is room for selection. The accessions from Ethiopia were the most sensitive to light regimes. This result was out of line with what has been observed in other studies where shade positively affected bean size as well as beverage quality (Vaast et al., 2006). This difference could stem from the fact that Vaast et al. only worked on one genotype.
Bean Size and NIRS Signature Are Stable Characters While Yield and Above All Sensorial Scores Are Less Stable
We observed high stability in the genotype classifications of bean size, meaning selection for bean size would produce the same results if carried out under shade or in full-sun conditions. The fact that shading is known to reduce the ambient temperature (Ehrenbergerová et al., 2017; Da Silva et al., 2018) by about 4°C at the leaf surface implies that bean size selection is not sensitive to shading or temperature. Using the NIRS signature showed revealed high stability in the distances between varieties regardless of the extent of shading, which is consistent with our previous findings (Posada et al., 2009). We found a correlation between the yields of genotypes grown in the shade and in the sun, but the correlation was not strong enough to predict the productivity of a genotype in the shade by knowing its performance in full sun (and vice versa). Selection for green coffee yield therefore depends on light intensity and/or temperature. We found no correlation between genotype ranking and the final cupping score between the two environments. Unfortunately, this is a time-consuming and expensive method, especially as it involved a large number of samples at this pre-selection stage. If we assume that the biochemical composition of the grain reflects its sensory quality, as we observed high stability using the NIRS, we can hypothesize that the absence of a correlation between final cupping score in the two environments was due to the lack of precision inherent to the sensorial test. However, NIRS measures the biochemical composition of the grain, i.e., the aroma precursors, not volatile compounds. Two future lines of research will therefore be conducted and the findings compared to those obtained using the sensorial method. First, it would be interesting to test whether a NIRS signature can advantageously replace sensory measurements. In this system, the distance from well-known varieties will be evaluated. Selection will be based on the candidate's distance from well-known varieties. When the distance is short, this indicates that the varieties are close to each other and are therefore considered to belong to the same “sensorial universe.” In a second experiment, it could also be considered that differences between Arabica genotypes are mainly due to volatile compounds. In this case, we would routinely use SPME-GC-MS analysis to classify the genotypes in different groups based on their volatile compounds. We would then a analyse the convergence of the three methods (i.e., using classical sensory analysis, NIRS and SPME-GC-MS) and choose the most repeatable and cheapest method.
Does Selection in Full Sun or Under Shade Lead to the Same Results? The Answer Depends on the Specifications
Specifications dictated objectively or subjectively by the market, producers and/or political authorities are the result of implicit or explicit choices based on rational or irrational interests. Breeders try to fully respect the stakeholders' specifications.
In the present study, we opted to illustrate selection specifications corresponding to three main farming systems which produce coffees for three types of markets corresponding to three “breeding orientations” as defined by Lammerts van Bueren et al. (2018). We showed that the selection of genotypes according to specialty coffee specifications resulted in the selection of coffees only of Ethiopian origin that were remarkable for their sensory quality under shade conditions, and three Ethiopian accessions and three F1 hybrids under full-sun conditions. In this “corporate-based breeding” approach, it is clear that selection in full sun does not produce the same results as in the shade. We therefore strongly recommended from the first stage of selection on, to test the genotypes both under shade and in full sun. On the other hand, for the sensory qualities to be fully expressed, we recommend carrying out this selection at a high altitude. The Ethiopian accessions tested in this study were collected at high altitude in Ethiopia. They are therefore suitable for cultivation at high altitudes, and this is where they will best express their sensory qualities. It seems that there could be a specific adaptation of some of these genotypes to shade or on the contrary, to full light at high altitudes (see for example, the remarkable performance of the E286 genotype in full sun compared to its poor performance under shade). It would be interesting to test for this specific adaptation in several dozen genotypes. The Ethiopian accessions, on the other hand, are rarely adapted to low altitude conditions with higher temperatures (personal observations). One defect we identified is their small bean size. By selecting on a wider genetic basis, it will probably be possible to find Ethiopian accessions of excellent quality with large beans. It would also be useful to breed 100% wild Ethiopian varieties by crossing accessions from Ethiopia to obtain F1 hybrids that are more productive than the Ethiopian accessions and have an excellent sensorial profile. This work has already been done in Ethiopia itself (Opile and Agwanda, 1993; Bellachew, 1997) but unfortunately the material is not yet available for other producing countries.
The main problem in selecting Ethiopian lines or Ethiopian F1 hybrids concerns tree volume. The volumes of the selected trees vary 2- to 4-fold compared to the Castillo lines or F1 hybrids. This means that the crop densities and consequently yields per hectare are reduced accordingly. This would be acceptable for farmers if the coffee were to be sold at 1.5 or twice the coffee price set by the New York Stock Exchange to compensate for the producers' loss of income. The specialty coffee market needs to consider this limitation in order to continue to secure sustainable supplies of specialty coffee.
A very large part of the current world coffee orchard is composed of dwarf varieties (small tree volume) well-adapted to full sun (i.e., Catuai, Caturra, IAPAR59, CR95, Castillo®, Marsellesa®, Obata, etc.), with mainstream sensory quality and large beans for the pure lines that make up the Castillo multi-line variety. Our starting point was the observation that selection for full sun must first focus on the volume of the plant to maximize production per hectare. We have shown that it is much better to use F1 hybrids, which produce considerably more with the same plant volume. These hybrids also had the same sensory and physical qualities as the maternal lines. In a recent article (Marie et al., 2020), we showed that two F1 hybrids (Centroamericano-H1 and Mundo Maya-H16) with a volume only 20% higher than that of Marsellesa® or Caturra (one of the most widely cultivated varieties) produced 50% higher yields than Marsellesa® or Caturra. In the present study, 10 F1 hybrids produced 60–85% more than the best female line (CU1842) with the same tree volume (i.e., resulting in the same cultivation density). The results of this study confirm previous reports: substantial genetic progress can be expected from the dissemination of F1 hybrids (obtained by crossing American dwarf x wild Ethiopian varieties) in Arabica farming systems based on the principles of the first Green Revolution (full-sun dwarf varieties, mainstream cup quality). The question of whether during selection, the trees should also be tested for their ability to produce under shade does not arise.
In the third type of selection, we opted to select varieties for their adaptation to shade conditions. Indeed, even in Brazil (the world's leading coffee-producing country) where cultivation under full sun largely predominates, some voices which suggest that AFS may represent a better management system for biodiversity and soil conservation are finally being heard. The main shortcoming of AFS is lower productivity compared with the intensive full-sun system. In a previous multi-location trial in an agroforestry system (Bertrand et al., 2011), hybrid yields were shown to be 58% higher than those of the American cultivars. In the present study, we have shown that F1 hybrid varieties are far more productive than others under shade. It is therefore possible to select hybrids that produce in the shade as well as the best pure line varieties grown in full sun. This finding is a powerful argument to encourage coffee growers who use the full-sun cropping system to change to AFS cropping systems, especially in regions where temperatures are expected to increase in climate scenarios. For the selection of genotypes adapted to agroforestry systems, we advise proceeding as we did in this study (i.e., selecting the genotypes that produce as much under shade as the best controls in full sun). For this farming system to be profitable, the productivity levels indeed need to approach those of farming systems in full sun. However, this advantage will disappear the day growers in full-sun adopt F1 hybrids. To really progress, it will be necessary to select hybrids which produce as much in full sun as under heavy shade. Data sheet 1 shows that this is the case for the hybrid CX2385 × E291. Indeed, although the volume of the tree is not modified by shading and the number of nodes formed does not change either (data not shown), the number of flower buds does. How does light modify the number of flower buds? Is it the quantity or quality of light that has an effect? Are there any allelic variations for this trait? At a time when global warming is accelerating and agroforestry is perceived as a mitigation solution, it is urgent to launch research on this subject.
In both conditions (full-sun or shade), by opting for F1 hybrids rather than Ethiopian accessions it would be possible to increase crop densities and productivity per hectare while at the same time achieving good sensory qualities. However, being unable to achieve the quality of the Ethiopian varieties under shade and the fact of selecting only three hybrids under full-sun conditions is a matter of concern. Selecting a genotype with both remarkable sensory quality and F1 hybrid productivity per hectare appears to be exceptional? unlikely? for the time being.
The results we obtained also raise questions concerning the type of varieties. The first question concerns maternal heritability of cup quality. Should Ethiopian accessions be used as maternal parents? Indeed, the majority of F1 hybrids created in Latin America up to now were bred with an American pure line as mother and an Ethiopian accession as father. The heritability of good cup quality is perhaps mainly maternal, as is the case with bean size.
Can we imagine creating exceptional pure lines that combine both the productivity of dwarf F1 hybrids and the quality of the best “wild” Ethiopian accessions? This raises the question of the heritability of characters and the nature of heterosis in Arabica. Can it be fixed in the homozygous state? The path is long and hazardous. The nature of heterosis in an allopolyploid plant is still very poorly understood (Solhaug et al., 2016) but we suspect that it not very “fixable.” To create high performance lines for productivity and quality, a very broad genetic base is needed to have a chance of selecting the best candidates. This strategy is only used in Ethiopia, the primary centre of diversity of the species, since the genetic diversity available there is very high. In other countries which do not have access to very wide diversity, in other words, all the other countries in the world, this strategy will probably quickly reach its limits.
So, if the F1 hybrid is the best type of variety to adapt to different farming systems and to climate change, the main problem to be solved is that of seed reproduction (Etienne et al., 2018). We have shown that this obstacle can be overcome by using male sterility, which makes it possible to produce F1 hybrid seeds at a price compatible with high-density cultivation (Georget et al., 2019). The transfer of male sterility genes to good progenitors should now be a research priority for the Arabica breeder community.
Conclusion
Although in this study, we did explore the main farming systems, others remain to be explored. Coffee harvest mechanization in Brazil is based on tall varieties (Mundo-Novo, Bourbon) which were not covered in the present study. We believe that tall F1 hybrids could be tested advantageously in mechanized systems where they may perform better than Mundo Novo and Bourbon. In the future, we could also test a selection based on both tree volume and productivity. In our opinion, there is no doubt that hybrids between tall American varieties and Ethiopian accessions will be more productive than their own parents.
It is also up to agronomists to develop new farming systems, especially to address the Anthropocene crisis. We could imagine, for instance, designing ultra-intensified farming systems with super-dwarf plant formats. Indeed, dwarfism genes exist that are even more pronounced than the Caturra gene (i.e., Laurina). This would allow greenhouse cultivation and, in theory, the cultivation of Arabica coffees outside the intertropical zone (Djerrab et al., 2020) to escape global warming.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Author Contributions
BB is the writer of the paper. AV established the trial, organized the observations, and collected and interpreted the results. LM statistically analyzed the results. J-CB helped to revise the document and to interpret the results. All authors contributed to the article and approved the submitted version.
Funding
This research was funded by CENICAFE, MINCIENCIAS, National Agricultural Science, Technology and Innovation Program, Agreement RC-0148-2013, code 2251-569-33518 and by the BREEDCAFS project (EUROPEAN COMMISSION under the Horizon 2020—Research and Innovation Programme, H2020-SFS-2016-2, Grant Agreement Number: 727934).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Valle del Cauca coffee growers committee for cup tasting and CENICAFE (Centro Nacional de Investigaciones de Café- Cenicafé Sede Planalto, km. 4 vía Chinchiná-Manizales. Manizales (Caldas) – Colombia, for technical support in the field.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsufs.2021.709901/full#supplementary-material
References
Alvarado-Alvarado, G., Posada-Suárez, H., and Cortina-Guerrero, H. (2005a). Castillo: Nueva Variedad de Café Con Resistencia a La Roya. Avances Ténicos Cenicafe, 337. Available online at: https://www.cenicafe.org/es/publications/avt0337.pdf.
Alvarado-Alvarado, G., Posada-Suárez, H., and Cortina-Guerrero, H. (2005b). La variedad castillo: una variedad de café Coffea arabica L. con elevada productividad y amplia resistencia a enfermedades. Fitotecnia Colombiana 8:21.
Avelino, J., Zelaya, H., Merlo, A., Pineda, A., Ordoñez, M., and Savary, S. (2006). The intensity of a coffee rust epidemic is dependent on production situations. Ecol. Modell. 197, 431–447. doi: 10.1016/j.ecolmodel.2006.03.013
Bacon, C., Ernesto Méndez, V., Flores Gómez, M., Stuart, D., and Díaz Flores, S. (2008). Are sustainable coffee certifications enough to secure farmer livelihoods? The millenium development goals and Nicaragua's fair trade cooperatives. Globalizations 5, 259–274. doi: 10.1080/14747730802057688
Baggio, A., Caramori, P., Androcioli Filho, A., and Montoya, L. (1997). Productivity of southern brazilian coffee plantations shaded by different stockings of Grevillea robusta. Agrofor. Syst. 37, 111–120. doi: 10.1023/A:1005814907546
Bates, D., Mächler, M., Bolker, B., Steven, C., and Walker, S. (2015). Fitting linear mixed-effects models using Lme4. J. Stat. Softw. 67:51. doi: 10.18637/jss.v067.i01
Battistini, A., and Battistini, G. (2005). VICTOR®: a semi-dwarfing cherry rootstock for dry conditions. Acta Hortic. 667, 189–190. doi: 10.17660/ActaHortic.2005.667.27
Bellachew, B. (1997). “Arabica coffee breeding in Ethiopia,” in Proceedings of the 17th Colloquium of International Coffee Science Association, ed. ASIC (Nairobi), 406–414.
Bertrand, B., Alpizar, E., Lara, L., SantaCreo, R., Hidalgo, M., Quijano, J., et al. (2011). Performance of Coffea Arabica F1 hybrids in agroforestry and full-sun cropping systems in comparison with American pure line cultivars. Euphytica 181, 147–158. doi: 10.1007/s10681-011-0372-7
Bertrand, B., Boulanger, R., Dussert, S., Ribeyre, F., Berthiot, L., Descroix, F., et al. (2012). Climatic factors directly impact the biochemical composition and the volatile organic compounds fingerprint in green Arabica coffee bean as well coffee beverage quality. Food Chem. 135, 2575–2583. doi: 10.1016/j.foodchem.2012.06.060
Bertrand, B., Etienne, H., Cilas, C., Charrier, A., and Baradat, P. (2005). Coffea arabica hybrid performance for yield, fertility and bean weight. Euphytica 141, 255–262. doi: 10.1007/s10681-005-7681-7
Bertrand, B., Guyot, B., Anthony, F., and Lashermes, P. (2003). Impact of the Coffea canephora gene introgression on beverage quality of C. arabica. Theoret. Appl. Genet. 107, 387–394. doi: 10.1007/s00122-003-1203-6
Bertrand, B., Vaast, P., Alpizar, E., Etienne, H., Davrieux, F., and Charmetant, P. (2006). Comparison of bean biochemical composition and beverage quality of Arabica hybrids involving Sudanese-Ethiopian origins with traditional varieties at various elevations in Central America. Tree Physiol. 26, 1239–1248. doi: 10.1093/treephys/26.9.1239
Bryant, F., and Kothmann, M. (1979). Variability in predicting edible browse from crown volume. J. Range Manage. 32:144. doi: 10.2307/3897560
Bunn, C., Läderach, P., Guillermo, J., Jimenez, P., Montagnon, C., and Schilling, T. (2015). Multiclass classification of agro-ecological zones for Arabica coffee: an improved understanding of the impacts of climate change. PLoS ONE 10:16. doi: 10.1371/journal.pone.0140490
Campbell, B., Thornton, P., Zougmor,é, R., van Asten, P., and Lipper, L. (2014). Sustainable intensification: what is its role in climate smart agriculture? Curr. Opin. Environ. Sustain. 8, 39–43. doi: 10.1016/j.cosust.2014.07.002
Da Silva, N., Da, F., Gomes Morinigo, K., De França Guimarães, N., De Souza Gallo, A., De Souza, M., et al. (2018). Shade trees spatial distribution and its effect on grains and beverage quality of shaded coffee trees. J. Food Qual. 2018:7909467. doi: 10.1155/2018/7909467
DaMatta, F. (2004). Ecophysiological constraints on the production of shaded and unshaded coffee: a review. Field Crops Res. 86, 99–114 doi: 10.1016/j.fcr.2003.09.001
DaMatta, F., Rahn, E., Läderach, P., Ghini, R., and Ramalho, J. (2019). Why could the coffee crop endure climate change and global warming to a greater extent than previously estimated? Clim. Change 152, 167–178. doi: 10.1007/s10584-018-2346-4
Djerrab, D., Bertrand, B., Breitler, J.-C., Léran, S., Dechamp, E., Campa, C., et al. (2020). Photoperiod-dependent transcriptional modifications in key metabolic pathways in Coffea arabica. Tree Physiol. 41, 302–316. doi: 10.1093/treephys/tpaa130
Döring, T., Pautasso, M., Finckh, M., and Wolfe, M. (2012). Concepts of plant health - reviewing and challenging the foundations of plant protection. Plant Pathol. 61, 1–15. doi: 10.1111/j.1365-3059.2011.02501.x
Echeverria-Beirute, F., Murray, S., Klein, P., Kerth, C., Miller, R., and Bertrand, B. (2018). Rust and thinning management effect on cup quality and plant performance for two cultivars of Coffea arabica L. J. Agric. Food Chem. 66, 5281–5292. doi: 10.1021/acs.jafc.7b03180
Ehrenbergerová, L., Šenfeldr, M., and Habrová, A. (2017). Impact of tree shading on the microclimate of a coffee plantation: a case study from the Peruvian Amazon. Bois Forets Tropiques 4, 13–22. doi: 10.19182/bft2017.334.a31488
Elias, A., Busov, V., Kosola, K., Ma, C., Etherington, E., Shevchenko, O., et al. (2012). Green revolution trees: semidwarfism transgenes modify gibberellins, promote root growth, enhance morphological diversity, and reduce competitiveness in hybrid poplar. Plant Physiol. 160, 1130–1144. doi: 10.1104/pp.112.200741
Etienne, H., Breton, D., Breitler, J.-C., Bertrand, B., Déchamp, E., Awada, R., et al. (2018). Coffee somatic embryogenesis: how did research, experience gained and innovations promote the commercial propagation of elite clones from the two cultivated species? Front. Plant Sci. 9:1630. doi: 10.3389/fpls.2018.01630
Georget, F., Marie, L., Alpizar, E., Courtel, P., Bordeaux, M., Hidalgo, J., et al. (2019). Starmaya: the first Arabica F1 coffee hybrid produced using genetic male sterility. Front. Plant Sci. 10:1344. doi: 10.3389/fpls.2019.01344
Haggar, J., Barrios, M., Bolaños, M., Merlo, M., Moraga, P., Munguia, R., et al. (2011). Coffee agroecosystem performance under full sun, shade,conventional and organic management regimes in Central America. Agrofor. Syst. 82, 285–301. doi: 10.1007/s10457-011-9392-5
Instituto Colombiano de Normas Técnicas y Certificación (ICONTEC). (2004). Norma Técnica Colombiana. ntc 5248. Café verde. Análisis del Tamaño. Tamizado Manual.
IPCC (2013). “Climate change 2013: the physical science basis,” in Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, eds T.F. Stocker, D. Qin, G.-K. Plattner, M. Tignor, S. K. Allen, J. Boschung, A. Nauels, Y. Xia, V. Bex, and P. M. Midgley (Cambridge; New York, NY: Cambridge University Press), 1535.
IPCC (2014). “Climate change 2014: mitigation of climate change,” in Contribution of Working Group III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, eds O., Edenhofer, R. Pichs-Madruga, Y. Sokona, E. Farahani, S. Kadner, K. Seyboth, A. Adler, I. Baum, S. Brunner, P. Eickemeier, B. Kriemann, J. Savolainen, S. Schlömer, C. von Stechow, T. Zwickel, and J. C. Minx (Cambridge; New York, NY: Cambridge University Press).
Jaramillo-Botero, C., Silva Santos, R., Prieto Martinez, H., Cecon, P., and Pereira Fardin, M. (2010). Production and vegetative growth of coffee trees under fertilization and shade levels. Sci. Agric. 67, 639–645. doi: 10.1590/S0103-90162010000600004
Lammerts van Bueren, E., Struik, P., van Eekeren, N., and Nuijten, E. (2018). Towards resilience through systems-based plant breeding. A review. Agron. Sustain. Dev. 38:42. doi: 10.1007/s13593-018-0522-6
Marie, L., Abdallah, C., Campa, C., Courtel, P., Bordeaux, M., Navarini, L., et al. (2020). G x E Interactions on yield and quality in Coffea arabica: new F1 hybrids outperform American cultivars. Euphytica 216:17. doi: 10.1007/s10681-020-02608-8
McCook, S., and Vandermeer, J. (2015). The big rust and the red queen: long-term perspectives on coffee rust research. Phytopathology 105, 1164–1173. doi: 10.1094/PHYTO-04-15-0085-RVW
Moncada, P., Del, M., Cortina, H., and Alarcón, R. (2019). Cup quality and yield evaluation of the Ethiopian germplasm collection of coffea arabica L. J. Agric. Rural Res. 3, 100–126. Available online at: http://aiipub.com/journals/jarr-190919-010082/
Muschler, R. (2001). Shade improves coffee quality in a sub-optimal coffee-zone of Costa Rica. Agrofor. Syst. 51, 131–139. doi: 10.1023/A:1010603320653
Opile, W., and Agwanda, C. (1993). Propagation and distribution of cultivar Ruiru 11. Kenya Coffee 58, 1496–1508.
Ovalle-Rivera, O., Läderach, P., Bunn, C., Obersteiner, M., and Schroth, G. (2015). Projected shifts in Coffea arabica suitability among major global producing regions due to climate change. PLoS ONE 10:e0124155. doi: 10.1371/journal.pone.0124155
Posada, H., Ferrand, M., Davrieux, F., Lashermes, P., and Bertrand, B. (2009). Stability across environments of the coffee variety near infrared spectral signature. Heredity 102, 113–119. doi: 10.1038/hdy.2008.88
Rhinehart, R. (2009). What Is Specialty Coffee? Specialty Coffee Association of America. Available online at: https://scaa.org/?page=RicArtp1
Scalabrin, S., Toniutti, L., Di Gaspero, G., Scaglione, D., Magris, G., Vidotto, M., et al. (2020). A single polyploidization event at the origin of the tetraploid genome of Coffea arabica is responsible for the extremely low genetic variation in wild and cultivated germplasm. Sci. Rep. 10, 1–13. doi: 10.1038/s41598-020-61216-7
Schnable, P. S., and Springer, N. (2013). Progress toward understanding heterosis in crop plants. Annu. Rev. Plant Biol. 64, 71–88. doi: 10.1146/annurev-arplant-042110-103827
Solhaug, E. M, Ihinger, J., Jost, M., Gamboa, V., Marchant, B., et al. (2016). Environmental regulation of heterosis in the allopolyploid Arabidopsis suecica. Plant Physiol. 170, 2251–2263. doi: 10.1104/pp.16.00052
Struik, P., and Kuyper, T. (2014). Sustainable intensification in agriculture: the richer shade of green. A review. Agron. Sustain. Dev. 37, 1237–1257. doi: 10.1007/s13593-017-0445-7
Toniutti, L., Breitler, J.-C., Etienne, H., Campa, C., Doulbeau, S., Urban, L., et al. (2017). Influence of environmental conditions and genetic background of Arabica Coffee (C. arabica L) on Leaf Rust (Hemileia vastatrix) pathogenesis. Front. Plant Sci. 8:2025. doi: 10.3389/fpls.2017.02025
Vaast, P., Bertrand, B., Perriot, J.-J., Guyot, B., and Génard, M. (2006). Fruit thinning and shade improve bean characteristics and beverage quality of coffee (Coffea arabica L.) under optimal conditions. J. Sci. Food Agric. 86, 197–204. doi: 10.1002/jsfa.2338
Keywords: Coffea arabica, farming systems, agroforestry systems, specialty coffee, F1 hybrids, Dwarf American pure lines, wild Ethiopian accessions
Citation: Bertrand B, Villegas Hincapié AM, Marie L and Breitler J-C (2021) Breeding for the Main Agricultural Farming of Arabica Coffee. Front. Sustain. Food Syst. 5:709901. doi: 10.3389/fsufs.2021.709901
Received: 14 May 2021; Accepted: 30 July 2021;
Published: 23 August 2021.
Edited by:
Paulo Mazzafera, State University of Campinas, BrazilReviewed by:
Aejaz Ahmad Dar, Sher-e-Kashmir University of Agricultural Sciences and Technology of Jammu, IndiaRodrigo Rocha, Brazilian Agricultural Research Corporation (EMBRAPA), Brazil
Copyright © 2021 Bertrand, Villegas Hincapié, Marie and Breitler. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Benoît Bertrand, YmVub2l0LmJlcnRyYW5kQGNpcmFkLmZy