 Anthony Loor-Giler1,2
Anthony Loor-Giler1,2 Camila Sanchez-Castro3†
Camila Sanchez-Castro3† Marcela Robayo-Chico3
Marcela Robayo-Chico3 Byron Puga-Torres4Silvana Santander-Parra5†
Byron Puga-Torres4Silvana Santander-Parra5† Luis Nuñez5,6*
Luis Nuñez5,6*- 1Laboratorios de Investigación, Dirección General de Investigación, Universidad de las Américas (UDLA), Quito, Ecuador
- 2Facultad de Ciencias Veterinarias, Universidad de Buenos Aires, Buenos Aires, Argentina
- 3Facultad de Ingeniería y Ciencias Aplicadas, Carrera de Ingeniería en Biotecnología, Universidad de Las Américas (UDLA), Quito, Ecuador
- 4Facultad de Medicina Veterinaria y Zootecnia, Universidad Central del Ecuador, Quito, Ecuador
- 5Facultad de Ciencias de la Salud, Carrera de Medicina Veterinaria, Universidad de Las Américas, Quito, Ecuador
- 6One Health Research Group, Facultad de Ciencias de la Salud, Universidad de Las Américas, Quito, Ecuador
In Ecuador, the consumption of raw milk is fundamental, nonetheless it also presents risks due to the presence of Salmonella, a pathogen responsible for foodborne diseases (FBD). This study evaluated the prevalence of Salmonella spp. in raw milk from the provinces of Pichincha and Manabí, identifying the Salmonella enterica serovars Typhi, Paratyphi, Enteritidis and Typhimurium, in order to determine the risks associated with FBD. For this study, 600 raw milk samples were analyzed using a specific qPCR assay for Salmonella enterica, in parallel, bacterial isolation of the strains was performed according to ISO 6579-1:2017, and serotyped using endpoint PCR assays. Finally, the 16S rDNA of some isolates was sequenced and the identification of these isolates was confirmed. A prevalence of 37.5% (n = 225 positive samples) was obtained, where the climatic season had a significant influence on Salmonella enterica contamination, being higher in the warm season. In the analysis of serovars, Salmonella Typhimurium and Salmonella Enteritidis predominated with 62 and 60 samples respectively, followed by Salmonella Paratyphi with 32 samples and Salmonella Typhi with only 28 samples. A phylogenetic relationship was found between the 16 s samples sequences of this study and those previously reported in NCBI. The high prevalence of Salmonella enterica in raw milk intended for consumption in Ecuador represents a significant public health risk due to its high pathogenic potential. It is therefore essential to improve biosafety practices among livestock producers and implement sanitary control measures to reduce the risk posed by this pathogen in raw milk and similar food products.
1 Introduction
In Ecuador, the consumption of dairy products has deep cultural roots, especially in rural areas, where production is a pillar of the economy and tradition (Vranješ et al., 2015; Puga-Torres et al., 2022). Raw milk and its derivatives are appreciated for their taste and their high nutritional value, as they contain high proportions of macronutrients (proteins, fats and carbohydrates), vitamins and minerals, which, in areas where food is limited, represent an indispensable basis of nutrition for these communities (Burgess, 2014; Thorning et al., 2016). Those same hyper-nutritious content is what makes raw milk the ideal medium for the growth of pathogenic microorganisms such as: Salmonella spp., Escherichia coli, Listeria monocytogenes, Campylobacter jejuni, Staphylococcus aureus, and others (Quigley et al., 2013; Williams et al., 2023; Antoszewska et al., 2024). Among these, Salmonella spp. stands out as one of the main causes of foodborne diseases (FBD) worldwide, so its presence in unpasteurized dairy products represents a public health problem (Ohl and Miller, 2001; DeAngelis, 2024). According to the World Health Organization (WHO), 1.9 billion people suffer from infectious enteritis each year—resulting in 715,000 deaths—with one third of these cases linked to contaminated food, and Salmonella species responsible for 9% of foodborne diarrheal diseases (WHO, 2018; Pakbin et al., 2022).
Salmonella enterica, a member of the genus Salmonella and the Enterobacteriaceae family, is a gram-negative, frequently intracellular, bacillary pathogen (Cabello et al., 1993). It has a genome of approximately 5 Mb and in some serovars of Salmonella enterica subsp. enterica, a ~94 kb large virulence plasmid (pSLT) that contributes to pathogenesis through genes such as those in the spv operon. While pSLT is characteristic of serovars like Typhimurium, other clinically relevant serovars, such as Typhi lack this plasmid and instead rely on distinct virulence factors, including chromosomally encoded pathogenicity islands and, in the case of S. Typhi, the Vi capsular antigen (Johnson et al., 2018; Hiley et al., 2019; Knodler and Elfenbein, 2019). This subspecies includes most of the serotypes associated with infections in humans and animals, including the most common ones: Salmonella Typhi, Salmonella Typhimurium, Salmonella Enteritidis, and Salmonella Paratyphi (Knodler and Elfenbein, 2019). Salmonellosis is the name given to infection caused by bacteria of the genus Salmonella, which affects the gastrointestinal tract, although in certain cases it can lead to systemic disease (Terrier and Martinez, 2006). Based on disease presentation, symptoms and epidemiological characteristics, Salmonella enterica serotypes are classified as typhoidal, with Salmonella Typhi and Salmonella Paratyphi as the most distinctive, and non-typhoidal, such as Salmonella Typhimurium and Salmonella Enteritidis as the most distinguishing in regions such as North and South America, as well as Europe, according to WHO and European Food Safety Authority (EFSA) surveillance data; and may differ in other regions depending on local epidemiological patterns and food production systems (Knodler and Elfenbein, 2019; Daigle, 2021).
In Ecuador, the presence of Salmonella in food, especially meat, is widely documented, and its association with multidrug-resistant strains has been highlighted, which represents a growing risk to public health. In these, Typhimurium stood out as the most common serovar and the resistance genes CTX (for cephalosporins) and KPC-2 were found (Mejía et al., 2020; Mejia et al., 2021; Vinueza-Burgos et al., 2023; Villavicencio et al., 2024). Infections with non-typhoidal serovars in humans produce an episode of enterocolitis lasting 4–7 days, causing abdominal pain, anorexia, nausea, vomiting, watery or mucoid diarrhea, rarely with blood (Ohl and Miller, 2001; Knodler and Elfenbein, 2019). Susceptibility to infection with these strains is highest in infants, elderly and immunocompromised persons (Chen et al., 2025). Most of the serovars classified as non-typhoid have a veterinary impact, especially in poultry and livestock, because they cause severe enteritis symptoms, with high mortality (Al-Adwani et al., 2013; Shaji et al., 2023; Aworh et al., 2024). In contrast, typhoid salmonellosis is severe, characterized by systemic spread of the pathogen, invading extraintestinal tissues (Cabello et al., 1993; Baqir et al., 2025). During infection for up to 30 days, in addition to the symptoms present in non-typhoid salmonellosis, weakness and pink skin patches on the extremities are present (Bhandari, 2024). Mortality due to typhoid salmonellosis is close to 10% in cases of non-resolution of infection with antibiotics, which increases in low-income countries (Pieters et al., 2018).
Controlling the occurrence of FBDs is a critical public health concern, small and sporadic outbreaks of salmonellosis have been described during the last century, mainly caused by contaminated food, with 52% of non-typhi and 37% of typhi serovars (Savage, 1929; Nelson and Snider, 1946; Ohl and Miller, 2001; Zizza et al., 2024). In Pennsylvania, USA 2007, 29 cases of salmonellosis were determined by consumption of cheese formulated from raw milk (Centers for Disease Control and Prevention (CDC), 2007). In Italy, in 2015, in relation to several cases of FBDs, high levels of contamination and non-identified bacterial counts were identified in raw milk for daily consumption (Chiarlone et al., 2025). Recently, in Poland, 50% contamination by pathogenic microorganisms was identified in organic food of animal origin, where 8% corresponded to Salmonella enterica (Sosnowski et al., 2024). In 2021, the Ministry of Public Health of Ecuador (MPS) identified 5,872 cases of undiagnosed food-borne infections, which could be related to the presence of bacteria, such as Salmonella enterica, in raw milk for daily consumption (MSP, 2021).
Taking into account the considerations outlined above and previously reported for Salmonella spp. in Ecuador, it is necessary to identify this bacteria with high zoonotic potential in raw milk, that contaminate high consumption and production foods, such as raw milk, which could cause FBDs. The present study aims to determine the prevalence of Salmonella spp. in raw milk from the two provinces with the highest production in Ecuador (Pichincha and Manabí), using isolation with selective media and molecular identification through a qPCR assay from the enriched milk; in addition, the presence of the serotypes Typhi, Paratyphi, Enteritidis and Typhimurium were determined from the bacterial isolates, thus determining the risk according to the type of salmonellosis that they would cause.
2 Methods
2.1 Sampling
In order to determine prevalence data, the sample number was calculated using the formula: , according to the parameters previously described for an unknown population (Rodríguez del Águila and González-Ramírez, 2014), where: n = required sample size; Pexp = expected prevalence; d = desired absolute precision (0.05 o 5%); z = statistic for a level of confidence (95%) = 1.96. The minimum number of total raw milk samples required would then be 384. A total of 600 raw milk samples were collected between July 2022 and June 2023 from two provinces of Ecuador (Pichincha = 300 and Manabí = 300), with approximately 50 samples collected per province per season using a simple random sampling during a climatic year. According to the National Institute of Meteorology and Hydrology (INAMHI), samples will be classified as “warm” if collected between June and September, a period characterized by drought and high temperatures of approximately 30°C; and “rainy” if collected between October and May, a period characterized by abundant rainfall and low temperatures of approximately 22°C. Additionally, the size of the producer will be taken into account, where according to the Ministerial Agreement No. 095 (MAG, 2022), those who own between 1 and 50 head of cattle will be classified as “small producers,” and those who own between 50 and 200 head of cattle will be classified as “medium producers.” Sample collection was carried out based on NTE INEN ISO 707 and Standard ISO 7218 [Instituto Ecuatoriano de Normalización (INEN), 2015]. Samples were collected in sterile containers and placed at 4°C until they were transported to the Research Laboratories of the Universidad de Las Americas (UDLA), where they were subjected to bacteriology and molecular processes necessary for the identification of Salmonella spp. All the processes carried out in this research were in accordance with the guidelines and approval of the Committee on the Care and Use of Laboratory and Domestic Animal resources of the Agency of Regulation and Control of Phytosanitary and Animal Health of Ecuador (AGROCALIDAD), under number #INT/DA/019.
2.2 Bacteria isolation and DNA extraction from pre-enrichment raw milk
The isolation of Salmonella spp. from raw milk samples was carried out in accordance with ISO 6579-1:2017 (Instituto Ecuatoriano de Normalización (INEN), 2015). Samples were placed in Buffered Peptone Water (BPW), as a pre-enrichment method, and incubated for ~19 h at 200 rpm. After incubation, a 1 mL aliquot of the pre-enrichment was taken and subjected to DNA extraction using the GT Reagent in a phenol/chloroform based method, according to the previously described protocol (Dalla-Costa et al., 2017). In parallel, a portion of the enrichment was placed in Modified Semi-Solid Rappaport-Vassiliadis (MSRV) and incubated at 40°C for at least 24 h. Taking the greyish colored growth, it was seeded on Xylose-Lysine-Deoxycholate (XLD) agar and incubated at 37°C. After at least 19 h, red colonies with a black center or totally black colonies were picked and placed in 200uL of 1x TE Buffer, and DNA was extracted using a boiling-based method. Samples were frozen at −20°C overnight, subsequently heated to 95°C in a dry bath, centrifuged at 12,000 RCF for 10 min to pellet cellular debris, and 100uL of supernatant was in a new tube. DNA samples were placed at −20°C until use (Dashti and Dashti, 2009).
2.3 Standard curve construction
To prepare the calibration curve, the segment of the ttr locus corresponding to the qPCR assay to be used was amplified using the ttr-6 and ttr-4 primers (Table 1), the 2X Promega GoTaq Green Master Mix enzyme kit (Promega, Madison, WI, USA) according to the manufacturer’s instructions and applying the same thermocycling temperatures described previously (Malorny et al., 2004), and confirmed by SANGER sequencing (described in section 2.6). For this purpose, DNA extracted from Salmonella Typhimurium strain ATCC 14028 was used. The PCR product was purified with ExoSAP-IT™ Express PCR product Cleanup Enzyme (Applied Biosystems, Santa Clara, CA 95051, USA) and subsequently quantified on Nano Drop equipment (Thermo Fisher Scientific, California, CA, USA). Using the web-based DNA Copy Number and Dilution Calculator, the volumes needed to create a standard of 109 copies of genetic material were determined. Then, 10 serial dilutions were performed to 1 copy of genetic material to determine the sensitivity of the assay.
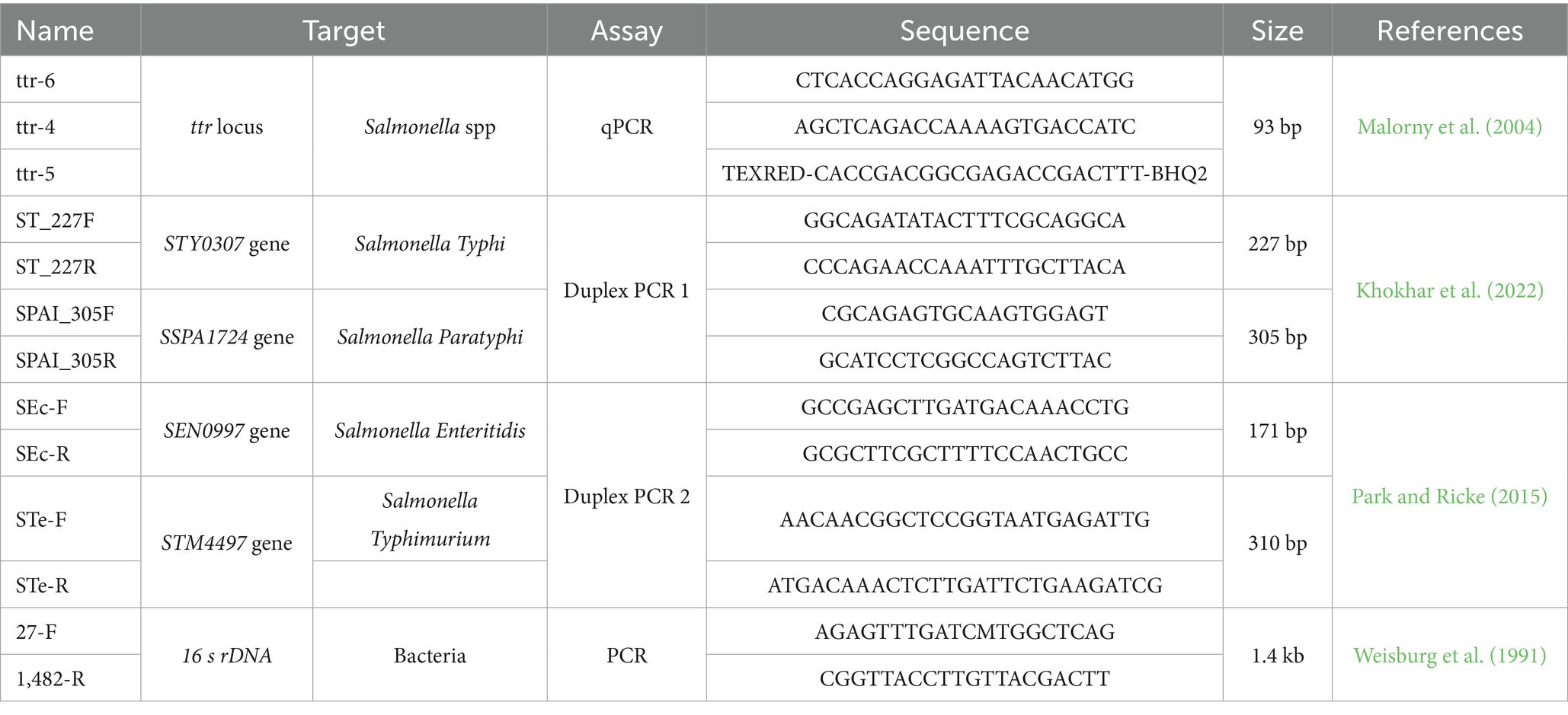
Table 1. Primers and probes used in this study.
2.4 Detection of Salmonella spp. by qPCR
To determine the presence of Salmonella spp in the raw milk samples, and to confirm the veracity of the bacterial isolates, an adapted single-plex qPCR assay was run with the previously described primers and probes for ttr locus, changing FAM fluorophore to TexRED and quencher to BHQ2 (Table 1). The assay was run using TaqMan Universal Master Mix II, with UNG, 0.2 μM of each primer, 0.1 μM of each probe, 1 μL of extracted DNA and made up to 10 μL volume with UltraPure™ DNase/RNase-Free Distilled Water. The qPCR reactions were run on the CFX96 Touch Real-Time PCR Detection System thermal cycler (Bio Rad Laboratories, Inc., Hercules, CA 94547, USA). The reactions were run under the protocol: 1 cycle at 50°C for 2 minutes for the inactivation of UNG, 1 cycle at 95°C for 5 minutes for the initial denaturation, followed by 45 cycles at 95°C for 15 seconds for denaturation, 60°C for 45 seconds for reading and annealing, and 72°C for 30 seconds for extension. The assay was performed on DNA extracted from pre-enriched raw milk and from bacterial isolates of each sample. All reactions were run in duplicate, and each run included a positive control (Salmonella Typhimurium ATCC 14028) and a negative control (ddH₂O).
2.5 Identification of Salmonella enterica serovars Typhi, Paratyphi, Enteritidis and Typhimurium
The presence serovars of Typhi and Paratyphi using the duplex PCR 1 previously described protocol (Khokhar et al., 2022), and Enteritidis and Typhimurium using the duplex PCR 2 previously described protocol (Table 1) (Park and Ricke, 2015) were identified in DNA samples extracted from isolated strains of Salmonella spp. PCR reactions were run using 2X Promega GoTaq Green Master Mix (Promega, Madison, WI, USA), 0.4 μM of each primer, 1 μL of DNA and made up to 10 μL volume with UltraPure™ DNase/RNase-Free Distilled Water. Thermocycling protocols were run according to protocols previously described (Park and Ricke, 2015; Khokhar et al., 2022). The amplified samples were subjected to horizontal electrophoresis on 2% agarose gel stained with SYBR Safe DNA Gel Strain (Thermo Fisher Scientific) and compared with a 50 bp molecular weight marker (Thermo Fisher Scientific). Gels were analyzed using the ChemiDoc XRS + analysis system (Bio Rad) and identified based on molecular weight according to each target.
2.6 16S rDNA sequencing
Ten DNA samples were randomly selected from the pool of raw milk isolates Excel’s RAND function, and subjected to 16S rDNA amplification to ensure representative variability (Weisburg et al., 1991). The protocol was run using 2X Promega GoTaq Green Master Mix enzyme (Promega, Madison, WI, USA), 0.3 μM of primers 27F and 1482R (Table 1), 1 μL of DNA extracted from the isolated bacteria and made up to 10 μL volume with UltraPure™ DNase/RNase-Free Distilled Water. PCR products were subjected to horizontal 1.2% agarose gel electrophoresis and stained with SYBR™ Safe DNA Gel Strain (Thermo Fisher Scientific) and compared with a 100 bp molecular weight marker (Thermo Fisher Scientific). Confirmed amplicons with a molecular weight of ~1,450 were purified with ExoSAP-IT™ Express PCR product Cleanup (Applied Biosystems, Santa Clara, CA 95051, USA) according to the manufacturer’s instructions. The purified samples were sequenced forward and reverse using BigDye® Terminator v3.1 Cycle Sequencing Kit (Thermo Fisher Scientific) and the sequence reaction was read on the ABI 3500 Series Genetic Analyzer (Applied Biosystems). Electropherograms were analyzed using Geneious software version 10.2.31 and compared with other Salmonella spp. sequences previously deposited in the GenBANK, randomly selected to differentiate the sequences of Salmonella bongori and Salmonella enterica. Both sets of sequences were aligned using the Clustal X algorithm in ClustalX 2.1 (Larkin et al., 2007). A phylogenetic tree was then constructed using the MEGA X program (Kumar et al., 2016), using the Neighbor joining statistical method and the p-distance substitution model and phylogeny test bootstrap model with 1,000 bootstrap to differentiate the species of the Salmonella genus.
2.7 Statistical analysis
A descriptive statistic of the data obtained was constructed differentiating the presence of Salmonella spp. based on the origin of the samples, the size of the producer, and the climatic time of collection. Additionally, the differentiation of the isolated serovars was classified under these same parameters (Supplementary material). A heat map was constructed to discriminate the proportion of positives for each serovar studied in each month of collection corresponding to a climatic time using RStudio Software V2022.12.0, which was also used to determine the prevalence of Salmonella spp. in raw milk in Ecuador. The normality of the data was determined using a Shapiro–Wilk test. As the data were not normally distributed, Z-test tests were used to determine whether there were any statistically significant differences in the prevalence of Salmonella spp. between provinces (Pichincha vs. Manabí), producer sizes (small vs. medium) and climatic seasons (warm vs. rainy); and Cochran’s Q test for serovars. The significance threshold for all tests was set at p < 0.05.
3 Results
3.1 Standard curve and sensitivity
The standard curve generated with the 9 serial dilutions in base 10 generated a curve with an efficiency of 97.6% and a correlation coefficient of 0.999 (Supplementary material). Samples with a Ct of <38 (equivalent to one copy) were considered positive. Since all points on the curve showed amplification, a limit of detection (LoD) of up to one copy of bacterial genetic material was established.
3.2 Detection of Salmonella spp. in pre-enrichment milk by qPCR
Of the 600 raw milk samples analyzed in this study, the prevalence of Salmonella spp. was 37.5% (Table 2). Among the data studied, only differences between warm and rainy seasons (and not the parameters of size and locality) were found; showing that climatic season is a significant factor influencing Salmonella contamination rates in raw milk, regardless of province or producer size (Supplementary material). These data, in terms of climatic season, showed higher prevalence in warm weather, with 96.00% of the total positive samples in both provinces and for both producer sizes sampled (n = 216/225). Smallholder farms showed the highest prevalence of Salmonella spp. (81.78%, n = 184), compared to medium-sized producers. Particularly by province, Pichincha showed a slightly higher positivity for this bacterium with 55% of the total samples (Figure 1). Significant differences were identified only in the presence of Salmonella spp. based on the collection time (climate), where those classified as warm always showed a higher positivity ratio (p < 0.001). Based on the groups generated by these three parameters, the highest positivity rate was found in the group of samples collected from small producers in Pichincha during the warm season; with similar percentages in the same province and climate, but in medium-sized producers, no significant differences with the other specific groups (p > 0.05). Bacterial isolation of Salmonella spp., subsequently confirmed by qPCR, showed 94.67% agreement (213 samples) with direct detection from enriched milk and yielded a Cohen’s kappa of 0.96, indicating a high level of concordance between both methods despite the greater sensitivity of the latter.
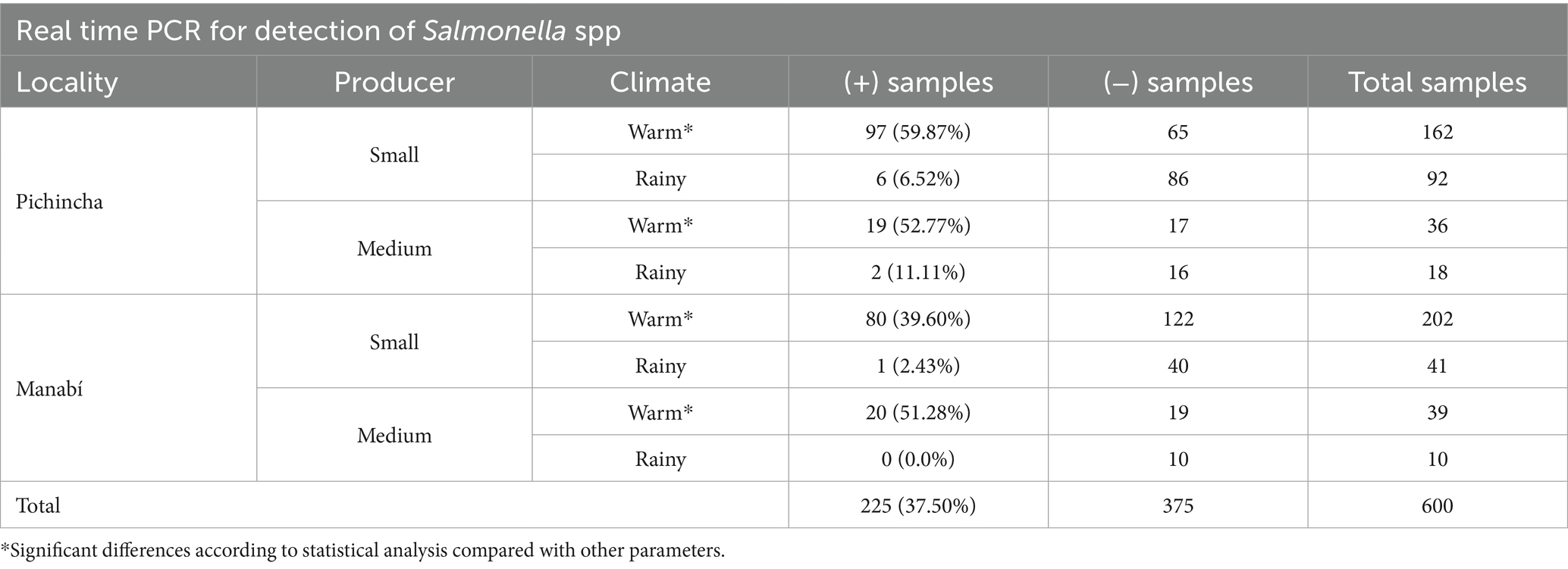
Table 2. Distribution of positives values of Salmonella spp. classified by locality, size of producer and climate.
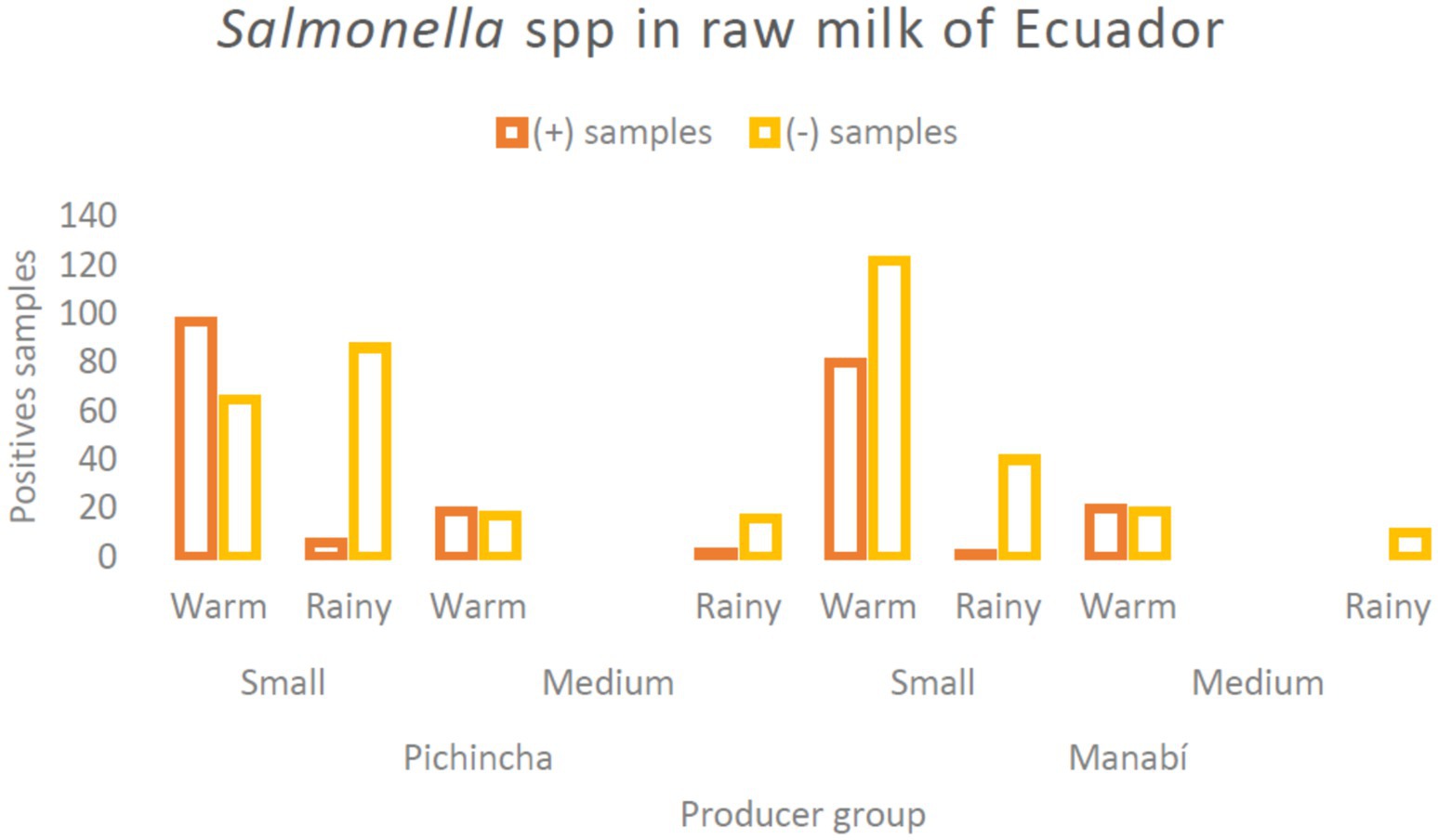
Figure 1. Bar chart showing positive Salmonella samples in raw milk from Ecuador. It compares small and medium producing groups during warm and rainy seasons in Pichincha and Manabí. Positive samples are indicated in orange, while negative samples are in yellow.
3.3 Identification of isolated serotypes Typhi, Paratyphi, Enteritidis and Typhimurium
Of the isolated samples, Salmonella Enteritidis and Salmonella Typhimurium were the most common serovars with 27.5 and 26.6% of the isolates, respectively, showing significant differences with Salmonella typhi (p < 0.001), which was the least present 12.44% (Table 3). Of the analyzed isolates, 19.11% could not be classified in any of these serovars.
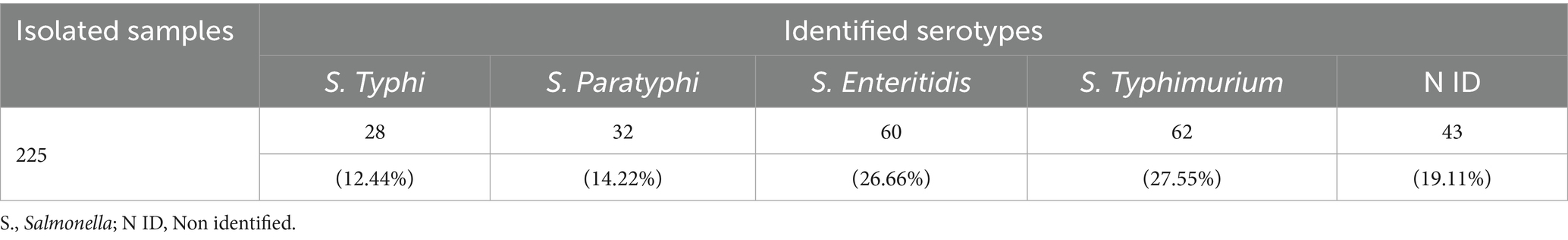
Table 3. Analysis of PCR results for serovars in identified Salmonella spp. samples.
In relation to the climatic season in which the samples were collected, it was found that Salmonella Paratyphi, Salmonella Typhimurium and Salmonella Typhi had significant differences in terms of the positives found in months classified as warm and those classified as rainy (p < 0.001), being predominant in the first case. However, this behavior was visibly observed in only two of the months (Figure 2), so the high number of samples collected and subsequently found to be positive in the warm season could be a statistical bias. On the other hand, the serovar Salmonella enteritidis showed predominance in the months of October and February, corresponding to rainy, and in September, corresponding to warm (Figure 2); although maintaining similar values in both cases (Supplementary material). No positive samples were found in March and November.
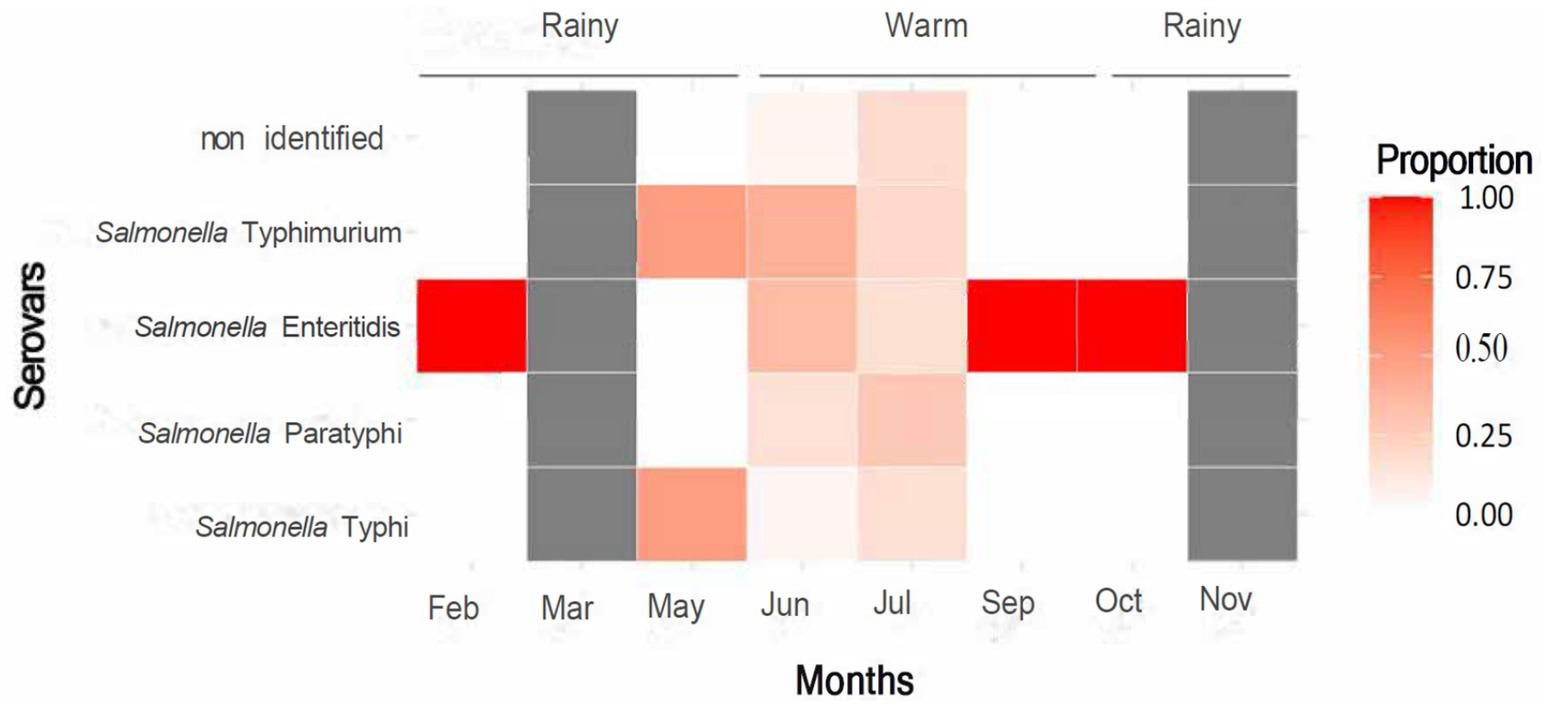
Figure 2. Heatmap showing the proportion of various Salmonella serovars across months and seasons. Darker shades indicate higher proportions. Salmonella Enteritidis peaks in February and September. Seasons are labeled as rainy and warm. A color scale on the right indicates proportion levels from zero (light) to one (dark red).
3.4 Analysis of 16S rDNA sequences
The phylogenetic tree generated formed two general clades differentiated by containing the two species of the genus Salmonella. The first clade contains the Salmonella bongori sequences obtained from GenBank, and the other contains the Salmonella enterica sequences obtained in this study and those collected from NCBI distributed throughout the study (Figure 3). As we expected, the sequences obtained in this study showed a similarity of more than 98% with the sequences previously deposited in GenBank [Supplementary material (Matrix of Nucleotides)], showing their generic closeness and correspondence with this species.
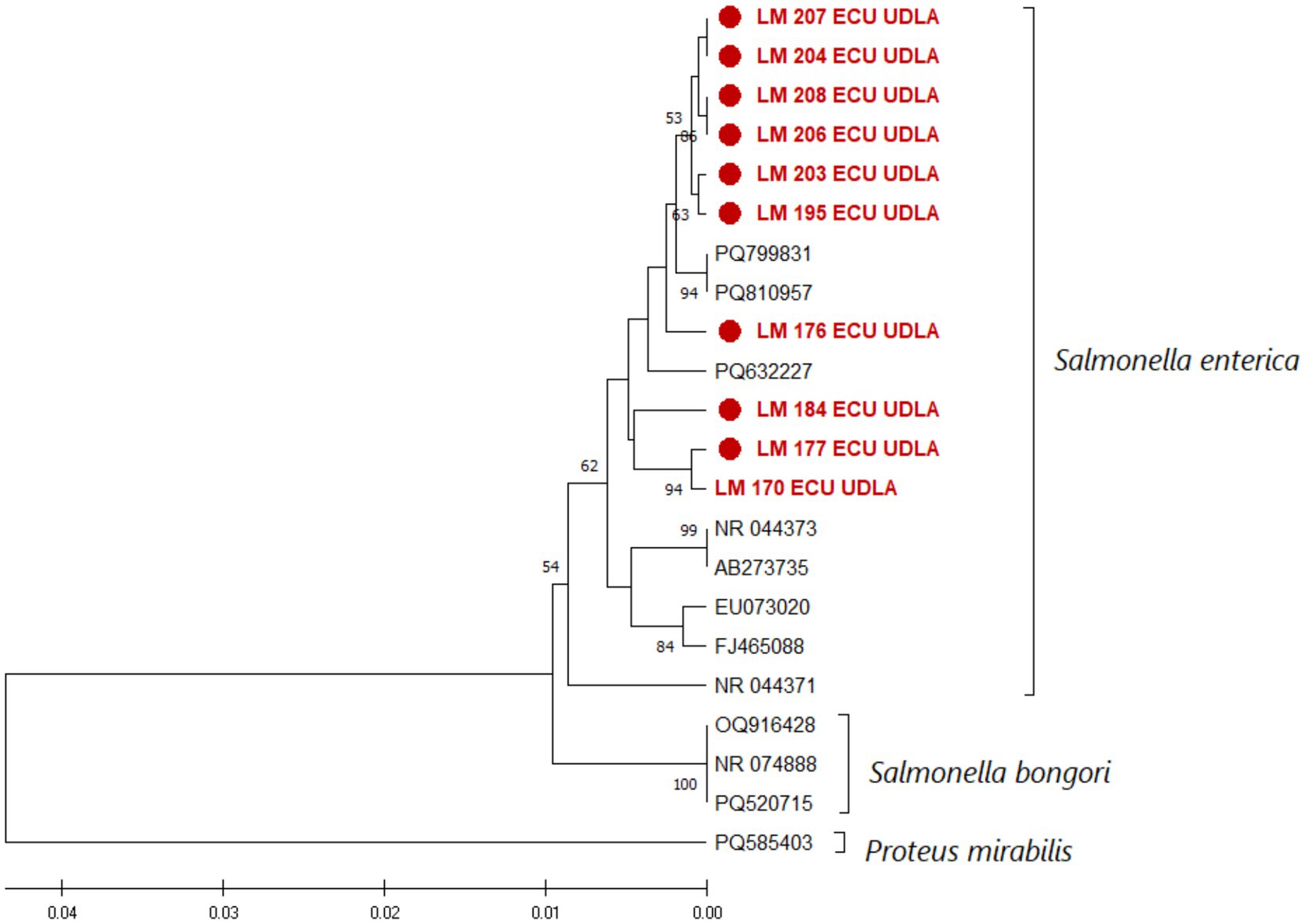
Figure 3. Phylogenetic relationships between the Salmonella Enterica sequences obtained here and other sequences collected from NCBI based on part of the 16S rDNA gene nucleotide sequence. The sequences were aligned in ClustalX 2.1. The phylogenetic tree was constructed using the MEGA X software package. Numbers along the branches refer to bootstrap values for 1,000 replicates. The scale bar represents the number of substitutions per site. A Proteus mirabilis 16S sequence was used as an outgroup. The sequences obtained in this study are shown in red and are marked with ●.
4 Discussion
Microbiological contamination of raw milk is a relevant problem due to its rich nutrient composition, which makes it a favorable medium for the growth of pathogenic microorganisms (Rampling, 1996; Quigley et al., 2013; Antoszewska et al., 2024). Among these, Salmonella genus stands out as an agent of concern, both because of its pathogenic potential and its ability to spread through contaminated food (WHO, 2018; Bhandari, 2024). This problem has significant public health implications, especially in contexts where raw milk is consumed without adequate heat treatment (Vranješ et al., 2015). The use of PCR-based diagnostic methods for the early detection of contaminants in raw milk and other media, without the need for bacterial isolation, is presented as a highly efficient alternative with greater sensitivity and specificity (Malorny et al., 2004; Lopes et al., 2018; Demirci et al., 2019). This study shows the adaptation of a previously described qPCR protocol. The protocol was validated by constructing a standard curve and performing a sensitivity assay. These tests showed that the protocol can detect as little as one copy of genetic material (Supplementary Figure 1). The protocol’s efficiency meets the standards set forth in the Minimum Information for Publication of Quantitative Real-Time PCR Experiments (MIQE) guidelines (Bustin et al., 2009). With this method, the presence of Salmonella spp. in raw milk used for consumption in Ecuador was demonstrated (Table 1).
A prevalence of 37.5% of Salmonella spp. was identified in the analyzed samples. The high presence of this pathogen in raw milk is presented as a risk for Ecuadorian public health, particularly in the populations that consume it. This value is consistent with previous studies in nearby regions that report the presence of this bacterium contaminating more than 30% of dairy products. This was particularly true in production systems that had inadequate hygiene (Dewey-Mattia et al., 2018; Barac et al., 2024). High contamination rates can be associated to different extra-insecure factors, such as inadequate practices during milking, storage and transport, as well as the lack of control systems (Agarwal et al., 2012). The data suggest a possible link between Salmonella spp. contamination and warmer seasons, though this may be influenced by other unmeasured factors (Table 1). This is similar to what has been previously described in studies conducted in Egypt, Ethiopia, Spain and Colombia, where bacterial proliferation in raw milk (highlighting the appearance of Salmonella species) more than doubled during the summer (warm seasons), compared to the cold season (Sospedra et al., 2009; Omar et al., 2018; Tobar-Delgado et al., 2020; Admasu et al., 2024). This may be due to the affinity of Salmonella spp. which, in spite of having the capacity to grow between 5° and 47°C, has a higher growth rate at temperatures above 30°C (Cabello et al., 1993; Daigle, 2021). It is essential to enhance control measures and bio-sanitary monitoring aimed at specific pathogens that tend to proliferate more rapidly during the warmer seasons.
The predominance of serotypes Enteritidis (26.66%) and Typhimurium (27.55%) isolates for Salmonella spp. (Table 3) is concordant with studies previously conducted in China, Brazil and Ethiopia where these serovars of Salmonella enterica were the most common in samples of raw milk and its derivatives (Degenhardt et al., 2023; Wang et al., 2023a; Beyene et al., 2024). In 2018, a study on the sources of Salmonella spp. contamination, which also highlighted these serovars as the most commonly found, indicated that foods of animal origin are the main source of Salmonella spp. isolation (42.76% of cases) (Ferrari et al., 2019). These cases of contamination were mainly associated to noncompliance with good animal farming practices, including deficiencies in milking, storage and transportation of milk (Bedassa et al., 2023; Wang et al., 2023b; Antoszewska et al., 2024). A higher number of Salmonella spp. positive samples was found in small producers, although this may reflect sampling bias or confounding variables (Table 1). In 2021, it was described that small farmers in Ecuador have deficiencies in the implementation of good animal farming practices, which may be influencing the proliferation of this microorganism (Lowenstein et al., 2016). To respond to these deficiencies, regulatory agencies such as AGROCALIDAD should establish monitoring and control measures to reduce the high rates of contamination and consumption of contaminated foods that pose a public health risk, in addition to previously indicated guidelines (AGROCALIDAD, 2012).
The presence of the serovars Typhi and Paratyphi (Table 3), although found to a lesser extent than non-typhi serovars, are a public health concern because they are causative agents of typhoid and paratyphoid fever (Bhandari, 2024). As these serovars are only associated with human infections and have no animal reservoirs, their presence in raw milk is most likely due to human contamination, although contamination during sample collection cannot be ruled out (Agarwal et al., 2012; Soyer et al., 2013; LeBoa et al., 2023). While contamination of raw milk with typhoid serovars is uncommon, recent studies have demonstrated the presence of Salmonella Typhi in raw milk. This presence has been attributed to inadequate hygienic conditions during handling and storage (Kaur et al., 2018; Bai et al., 2019). The worrying appearance of these pathogens is a wake-up call to regulatory entities to raise quality standards and sanitary control in small and medium sized dairy farms to avoid future outbreaks associated with these bacteria. When differentiating the serovars of Salmonella enterica based on the climatic season of collection, a peculiar pattern was observed, in which the few samples found in the months grouped as “rainy” correspond mostly to serovar enteritidis. Heat stress, increased fly activity, and enhanced biofilm formation in warm conditions can all contribute to greater Salmonella spp. prevalence by promoting shedding, transmission, and environmental persistence (Traub-Dargatz et al., 2006; Borges et al., 2018; Shahanaz et al., 2025). Since the serovars studied here share similar growth capacities in terms of temperature, there is not enough evidence to associate it to any specific phenomenon (Cabello et al., 1993; Daigle, 2021). The differentiation method is applied for bacterial isolates, therefore it is necessary to use assays capable of detecting the simultaneous presence of more than one serovar in the same sample (such as enriched milk) by applying qPCR methods like the one already used in this study to identify the presence of Salmonella spp. directly from milk (Postollec et al., 2011).
Given that it was not possible to assign a serovar in 19.11% of the samples positive for Salmonella spp. (Table 3), the need for future studies that address serovars that are fundamental for both animal health and possible zoonotic events is highlighted. These unidentified samples may correspond to a wide variety of serovars, among them: Salmonella Dublin, mainly associated with infections in cattle, Salmonella Newport, associated with fatal cases due to food consumption, or Salmonella Heidelberg, found in dairy products in a few cases (Clegg et al., 1983; Self et al., 2023; Velasquez-Munoz et al., 2024). Considering account that about 2,500 serotypes of Salmonella enterica have been proposed, where <100 are responsible for human infections; the analysis of all these is only feasible using metagenomic sequencing techniques (Cabello et al., 1993; Nichols et al., 2020). Given the serious pathogenic effect of typhoid serovars, and that serovars Enteritidis and Typhimurium comprise up to 75% of salmonellosis cases, these are a priority for diagnosis (WHO, 2018). It is important to note that in 2019, typhoid Salmonella serovars revealed 110,000 associated deaths around the world, while, despite their lower pathogenicity, 155,000 deaths from non-typhi Salmonella are estimated each year (Majowicz et al., 2010; WHO, 2023). There is no reliable data on mortality due to salmonellosis in Ecuador, however in 2023 a study in Colombia indicated that Salmonella spp. continued to be a cause of mortality due to foodborne infection (Rosso et al., 2023). It is important to assess the impact on public health of Salmonella spp. contamination in food, both as reported in this study and possibly present in other foods.
Analyzing the 16S gene sequences of the 10 bacterial isolates, as expected, all the sequences obtained clustered with sequences of Salmonella enterica previously reported in the NCBI (Figure 3). These results reaffirm the presence of this pathogen in the selected raw milk samples. Among the sequenced samples, those for which the serovar could not be identified also showed high levels of similarity with the other sequences obtained (Supplementary material). Nevertheless, the use of 16S to assign a serovar is not reliable, and more specific techniques such as qPCR or NGS are necessary (Weisburg et al., 1991; Postollec et al., 2011). Although the sequencing of bacterial isolates serves as a form of specific identification of bacteria genus, its high cost and execution time (including the bacterial isolation process) make it inefficient compared to updated methods such as qPCR assays. Early and specific detection allows adequate control and containment measures to minimize the risk to public health that contaminants such as Salmonella spp. represent. Further research is required to implement simultaneous detection and quantification of prevalent serovars directly from contaminated materials, to assess the extent of the issue in agriculture, hence reducing costs and application time.
5 Conclusion
The presence of Salmonella spp. in raw milk highlights the significant public health risks associated with its consumption, particularly in regions where proper heat treatment is not practiced. The application of qPCR methods demonstrated high sensitivity and specificity. This enabled the direct detection of Salmonella spp. without the need for time-consuming bacterial isolation. The identification of serovars associated with both animal and human contamination underscores the importance of improving hygiene practices during milking, storage, and transportation. Strengthening regulatory surveillance and adopting molecular diagnostic tools are critical steps to reduce contamination rates and prevent foodborne illnesses. These findings provide valuable evidence to influence in national food safety policies and support the development of targeted surveillance programmes aimed at reducing the risk of Salmonella transmission through raw milk.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material.
Ethics statement
The animal study was approved by Committee on the Care and Use of Laboratory and Domestic Animal resources of the Agency of Regulation and Control of Phytosanitary and Animal Health of Ecuador (AGROCALIDAD), under number INT/DA/019. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
AL-G: Data curation, Formal analysis, Writing – original draft, Conceptualization, Investigation. CS-C: Data curation, Methodology, Writing – review & editing. MR-C: Investigation, Writing – review & editing, Methodology. BP-T: Conceptualization, Writing – review & editing, Methodology. SS-P: Conceptualization, Writing – review & editing. LN: Writing – review & editing, Investigation, Conceptualization, Methodology, Funding acquisition, Project administration, Validation.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was supported by Universidad de Las Américas, Quito -Ecuador (Grant number: VET.LNN.21.02).
Acknowledgments
We are grateful to the farmers who allowed us to collect samples and to the students of the Universidad Central de Ecuador who collaborated in the process.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsufs.2025.1593266/full#supplementary-material
Footnotes
References
Admasu, H. N., Bedassa, A., Tessema, T. S., Kovac, J., Vipham, J. L., and Woldegiorgis, A. Z. (2024). Seasonal variation of Salmonella enterica prevalence in milk and cottage cheese along the dairy value chain in three regions of Ethiopia. Food Safety and Risk 11:2. doi: 10.1186/s40550-024-00108-4
Agarwal, A., Awasthi, V., Dua, A., Ganguly, S., Garg, V., and Marwaha, S. (2012). Microbiological profile of milk: impact of household practices. Indian J. Public Health 56, 88–94. doi: 10.4103/0019-557X.96984
AGROCALIDAD. (2012). Guía de buenas prácticas pecuarias de producción. Available online at: https://www.agrocalidad.gob.ec/ (accessed September 28, 2024).
Al-Adwani, S. R., Crespo, R., and Shah, D. H. (2013). Production and evaluation of chicken egg-yolk-derived antibodies against Campylobacter jejuni colonization-associated proteins. Foodborne Pathog. Dis. 10, 624–631. doi: 10.1089/fpd.2012.1313
Antoszewska, A., Maćkiw, E., Kowalska, J., Patoleta, M., Ławrynowicz-Paciorek, M., and Postupolski, J. (2024). Microbiological risks of traditional raw cow’s Milk cheese (Koryciński cheeses). Food Secur. 13:1364. doi: 10.3390/foods13091364
Aworh, M. K., Nilsson, P., Egyir, B., Owusu, F. A., and Hendriksen, R. S. (2024). Rare serovars of non-typhoidal Salmonella enterica isolated from humans, beef cattle and abattoir environments in Nigeria. PLoS One 19:e0296971. doi: 10.1371/journal.pone.0296971
Bai, Y., Cui, Y., Suo, Y., Shi, C., Wang, D., and Shi, X. (2019). A rapid method for detection of Salmonella in Milk based on extraction of mRNA using magnetic capture probes and RT-qPCR. Front. Microbiol. 10:770. doi: 10.3389/fmicb.2019.00770
Baqir, S. M., Sharma, N., Lutaf, A., Ghitan, M., and Lin, Y. S. (2025). Typhoid fever as a cause of liver failure in the United States: a case report. Case Rep. Gastrointest. Med. 2025:3087201. doi: 10.1155/crgm/3087201
Barac, D., Mansour, H., Awad, S., Ghazy, M. A., and Abdel-Mawgood, A. (2024). Prevalence of non-typhoidal salmonellae in the retail chicken meat in Alexandria, Egypt. Indian J. Microbiol. 64, 1680–1684. doi: 10.1007/s12088-024-01210-z
Bedassa, A., Nahusenay, H., Asefa, Z., Sisay, T., Girmay, G., Kovac, J., et al. (2023). Prevalence and associated risk factors for Salmonella enterica contamination of cow milk and cottage cheese in Ethiopia. Food Safety Risk 10:2. doi: 10.1186/s40550-023-00101-3
Beyene, A. M., Alemie, Y., Gizachew, M., Yousef, A. E., Dessalegn, B., Bitew, A. B., et al. (2024). Serovars, virulence factors, and antimicrobial resistance profile of non-typhoidal Salmonella in the human-dairy interface in Northwest Ethiopia: a one health approach. PLoS Negl. Trop. Dis. 18:e0012646. doi: 10.1371/journal.pntd.0012646
Bhandari, J. (2024). Typhoid fever. StatPearls. Available online at: https://www.ncbi.nlm.nih.gov/books/NBK557513/?utm_source=chatgpt.com (accessed January 27, 2025).
Borges, K. A., Furian, T. Q., Souza, S. N., Menezes, R., Tondo, E. C., Salle, C. T. P., et al. (2018). Biofilm formation capacity of Salmonella serotypes at different temperature conditions. Pesqui. Vet. Bras. 38, 71–76. doi: 10.1590/1678-5150-pvb-4928
Burgess, K. (2014). Milk and dairy products in human nutrition (2013), by E.Muehlhoff, a.Bennett and D.McMahon, food and agriculture organisation of the United Nations (FAO), Rome. E-ISBN: 978-92-5-107864-8 (PDF). Available on web-site (publications-sales@fao.Org). Int. J. Dairy Technol. 67, 303–304. doi: 10.1111/1471-0307.12124
Bustin, S. A., Benes, V., Garson, J. A., Hellemans, J., Huggett, J., Kubista, M., et al. (2009). The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 55, 611–622. doi: 10.1373/clinchem.2008.112797
Cabello, F., Hormaeche, C., Mastroeni, P., and Bonina, L. (1993). Biology of Salmonella. Boston, MA: Springer US.
Centers for Disease Control and Prevention (CDC) (2007). Salmonella typhimurium infection associated with raw milk and cheese consumption--Pennsylvania, 2007. MMWR Morb. Mortal Wkly. Rep. 56, 1161–1164.
Chen, S. S.-Y., Lin, T. T.-A., Chiang, Y.-L., Chen, C.-Y., Lin, W.-J., and Chang, R. (2025). Nontyphoidal salmonellosis is associated with an increased risk of stroke: insights from multinational real-world data. Int. J. Stroke 20:717. doi: 10.1177/17474930251313717
Chiarlone, S. A., Gori, A., Ravetta, S., Armani, A., Guardone, L., Pedonese, F., et al. (2025). Microbiological analysis conducted on raw Milk collected during official sampling in Liguria (north-West Italy) over a ten-year period (2014–2023). Animals 15:286. doi: 10.3390/ani15020286
Clegg, F., Chiejina, S., Duncan, A., Kay, R., and Wray, C. (1983). Outbreaks of Salmonella Newport infection in dairy herds and their relationship to management and contamination of the environment. Vet. Rec. 112, 580–584. doi: 10.1136/vr.112.25.580
Dalla-Costa, L. M., Morello, L. G., Conte, D., Pereira, L. A., Palmeiro, J. K., Ambrosio, A., et al. (2017). Comparison of DNA extraction methods used to detect bacterial and yeast DNA from spiked whole blood by real-time PCR. J. Microbiol. Methods 140, 61–66. doi: 10.1016/j.mimet.2017.06.020
Dashti, A. A., and Dashti, H. (2009). Heat treatment of bacteria: a simple method of DNA extraction for molecular techniques. Kuwait Med J 41, 117–122. Available at: https://www.academia.edu/14010095/Heat_Treatment_of_Bacteria_A_Simple_Method_of_DNA_Extraction_for_Molecular_Techniques
DeAngelis, D. (2024). Salmonella is everywhere right now—Here’s how to protect yourself from foodborne illness. London: University of London.
Degenhardt, R., Carvalho, M. M., Voidaleski, M. F., Daros, G. F., Guaragni, A., de Melo Pereira, G. V., et al. (2023). Brazilian artisanal colonial cheese: characterization, microbiological safety, and survival of Salmonella enterica serovar Enteritidis during ripening. Braz. J. Microbiol. 54, 2129–2135. doi: 10.1007/s42770-023-01022-1
Demirci, M., Yigin, A., Altun, S., Uysal, H., Saribas, S., and Kocazeybek, B. (2019). Salmonella Spp. and Shigella Spp. detection via multiplex real-time PCR and discrimination via MALDI-TOF MS in different animal raw milk samples. Niger. J. Clin. Pract. 22, 1083–1090. doi: 10.4103/njcp.njcp_596_18
Dewey-Mattia, D., Manikonda, K., Hall, A. J., Wise, M. E., and Crowe, S. J. (2018). Surveillance for foodborne disease outbreaks — United States, 2009–2015. MMWR Surveill. Summ. 67, 1–11. doi: 10.15585/mmwr.ss6710a1
Ferrari, R. G., Rosario, D. K. A., Cunha-Neto, A., Mano, S. B., Figueiredo, E. E. S., and Conte-Junior, C. A. (2019). Worldwide epidemiology of Salmonella serovars in animal-based foods: a meta-analysis. Appl. Environ. Microbiol. 85:591. doi: 10.1128/AEM.00591-19
Hiley, L., Graham, R. M. A., and Jennison, A. V. (2019). Genetic characterisation of variants of the virulence plasmid, pSLT, in Salmonella enterica serovar typhimurium provides evidence of a variety of evolutionary directions consistent with vertical rather than horizontal transmission. PLoS One 14:e0215207. doi: 10.1371/journal.pone.0215207
Instituto Ecuatoriano de Normalización (INEN) (2015). NTE INEN ISO 707: Leche y productos lácteos. Orientaciones para el muestreo (ISO 707:2008, IDT). Ecuador: Instituto Ecuatoriano de Normalización (INEN).
Johnson, R., Mylona, E., and Frankel, G. (2018). Typhoidal Salmonella: distinctive virulence factors and pathogenesis. Cell. Microbiol. 20:e12939. doi: 10.1111/cmi.12939
Kaur, A., Das, R., Nigam, M. R., Elangovan, R., Pandya, D., Jha, S., et al. (2018). Rapid detection device for Salmonella typhi in Milk, juice, water and calf serum. Indian J. Microbiol. 58, 381–392. doi: 10.1007/s12088-018-0730-4
Khokhar, F., Pickard, D., Dyson, Z., Iqbal, J., Pragasam, A., John, J. J., et al. (2022). Multiplex PCR assay to detect high risk lineages of Salmonella Typhi and Paratyphi a. PLoS One 17:e0267805. doi: 10.1371/journal.pone.0267805
Knodler, L. A., and Elfenbein, J. R. (2019). Salmonella enterica. Trends Microbiol. 27, 964–965. doi: 10.1016/j.tim.2019.05.002
Kumar, S., Stecher, G., and Tamura, K. (2016). MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 33, 1870–1874. doi: 10.1093/molbev/msw054
Larkin, M. A., Blackshields, G., Brown, N. P., Chenna, R., McGettigan, P. A., McWilliam, H., et al. (2007). Clustal W and Clustal X version 2.0. Bioinformatics 23, 2947–2948. doi: 10.1093/bioinformatics/btm404
LeBoa, C., Shrestha, S., Shakya, J., Naga, S. R., Shrestha, S., Shakya, M., et al. (2023). Environmental sampling for typhoidal salmonellas in household and surface waters in Nepal identifies potential transmission pathways. PLoS Negl. Trop. Dis. 17:e0011341. doi: 10.1371/journal.pntd.0011341
Lopes, A. T. S., Albuquerque, G. R., and Maciel, B. M. (2018). Multiplex real-time polymerase chain reaction for simultaneous quantification of Salmonella spp., Escherichia coli, and Staphylococcus aureus in different food matrices: advantages and disadvantages. Biomed. Res. Int. 2018, 1–12. doi: 10.1155/2018/6104015
Lowenstein, C., Waters, W. F., Roess, A., Leibler, J. H., and Graham, J. P. (2016). Animal husbandry practices and perceptions of zoonotic infectious disease risks among livestock keepers in a rural parish of Quito, Ecuador. Am. Soc. Trop. Med. Hygiene 95, 1450–1458. doi: 10.4269/ajtmh.16-0485
MAG (2022). Acuerdo Ministerial No.095. Ministerio de Agricultura y Ganadería. Available online at: https://www.agricultura.gob.ec/ (accessed September 30, 2024).
Majowicz, S. E., Musto, J., Scallan, E., Angulo, F. J., Kirk, M., O’Brien, S. J., et al. (2010). The global burden of nontyphoidal Salmonella gastroenteritis. Clin. Infect. Dis. 50, 882–889. doi: 10.1086/650733
Malorny, B., Paccassoni, E., Fach, P., Bunge, C., Martin, A., and Helmuth, R. (2004). Diagnostic real-time PCR for detection of Salmonella in food. Appl. Environ. Microbiol. 70, 7046–7052. doi: 10.1128/AEM.70.12.7046-7052.2004
Mejía, L., Medina, J. L., Bayas, R., Salazar, C. S., Villavicencio, F., Zapata, S., et al. (2020). Genomic epidemiology of Salmonella Infantis in Ecuador: from poultry farms to human infections. Front Vet Sci 7:547891. doi: 10.3389/fvets.2020.547891
Mejia, L., Vela, G., and Zapata, S. (2021). High occurrence of multiresistant Salmonella Infantis in retail meat in Ecuador. Foodborne Pathog. Dis. 18, 41–48. doi: 10.1089/fpd.2020.2808
MSP (2021). Subsistema de vigilancia SIVE- Alerta Enfermedades transmitidas por agua y alimentos. Available online at: https://www.salud.gob.ec/ (accessed September 28, 2024).
Nelson, K., and Snider, G. E. (1946). A minor epidemic of Salmonella infection. Va. Med. Mon. 73, 160–167.
Nichols, M., Stevenson, L., Koski, L., Basler, C., Wise, M., Whitlock, L., et al. (2020). Detecting national human enteric disease outbreaks linked to animal contact in the United States of America. Revue Scientifique et Technique de l’OIE 39, 471–480. doi: 10.20506/rst.39.2.3098
Ohl, M. E., and Miller, S. I. (2001). Salmonella: a model for bacterial pathogenesis. Annu. Rev. Med. 52, 259–274. doi: 10.1146/annurev.med.52.1.259
Omar, D., Alashmawy, M., and Ramadan, H. (2018). Occurrence and PCR identification of Salmonella spp. from milk and dairy products in Mansoura, Egypt. Int. Food Res. J. 25, 446–452. Available at: http://www.ifrj.upm.edu.my/25%20(01)%202018/(59).pdf
Pakbin, B., Rossen, J. W. A., Brück, W. M., Montazeri, N., Allahyari, S., Dibazar, S. P., et al. (2022). Prevalence of foodborne and zoonotic viral pathogens in raw cow milk samples. FEMS Microbiol. Lett. 369:fnac108. doi: 10.1093/femsle/fnac108
Park, S. H., and Ricke, S. C. (2015). Development of multiplex PCR assay for simultaneous detection of Salmonella genus, Salmonella subspecies I, Salm. Enteritidis, Salm. Heidelberg and Salm. Typhimurium. J. Appl. Microbiol. 118, 152–160. doi: 10.1111/jam.12678
Pieters, Z., Saad, N. J., Antillón, M., Pitzer, V. E., and Bilcke, J. (2018). Case fatality rate of enteric fever in endemic countries: a systematic review and Meta-analysis. Clin. Infect. Dis. 67, 628–638. doi: 10.1093/cid/ciy190
Postollec, F., Falentin, H., Pavan, S., Combrisson, J., and Sohier, D. (2011). Recent advances in quantitative PCR (qPCR) applications in food microbiology. Food Microbiol. 28, 848–861. doi: 10.1016/j.fm.2011.02.008
Puga-Torres, B., Aragón Vásquez, E., Ron, L., Álvarez, V., Bonilla, S., Guzmán, A., et al. (2022). Milk quality parameters of raw Milk in Ecuador between 2010 and 2020: a systematic literature review and Meta-analysis. Food Secur. 11:3351. doi: 10.3390/foods11213351
Quigley, L., O’Sullivan, O., Stanton, C., Beresford, T. P., Ross, R. P., Fitzgerald, G. F., et al. (2013). The complex microbiota of raw milk. FEMS Microbiol. Rev. 37, 664–698. doi: 10.1111/1574-6976.12030
Rodríguez del Águila, M., and González-Ramírez, A. (2014). Sample size calculation. Allergol Immunopathol. 42, 485–492. doi: 10.1016/j.aller.2013.03.008
Rosso, F., Rebellón-Sánchez, D. E., Llanos-Torres, J., Hurtado-Bermudez, L. J., Ayerbe, L., Suárez, J. H., et al. (2023). Clinical and microbiological characterization of Salmonella spp. isolates from patients treated in a university hospital in South America between 2012–2021: a cohort study. BMC Infect. Dis. 23:625. doi: 10.1186/s12879-023-08589-y
Savage, W. G. (1929). Unsolved problems in Salmonella food poisoning. Proc. R. Soc. Med. 22, 277–285. doi: 10.1177/003591572902200302
Self, J. L., Judd, M. C., Huang, J., Fields, P. I., Griffin, P. M., and Wong, K. K. (2023). Epidemiology of salmonellosis among infants in the United States: 1968–2015. Pediatrics 151:56140. doi: 10.1542/peds.2021-056140
Shahanaz, E., Zwally, K. M., Powers, C., Lyons, B., Kaufman, P., Athrey, G., et al. (2025). Flies as vectors of foodborne pathogens through food animal production: factors affecting pathogen and antimicrobial resistance transmission. J. Food Prot. 88:100537. doi: 10.1016/j.jfp.2025.100537
Shaji, S., Selvaraj, R. K., and Shanmugasundaram, R. (2023). Salmonella infection in poultry: a review on the pathogen and control strategies. Microorganisms 11:2814. doi: 10.3390/microorganisms11112814
Sosnowski, M., Wieczorek, K., and Osek, J. (2024). Identification of bacterial pathogens in organic food of animal origin in Poland. Food Secur. 13:3526. doi: 10.3390/foods13213526
Sospedra, I., Rubert, J. V., Soler, C., Soriano, J. M., and Mañes, J. (2009). Microbial contamination of Milk and dairy products from restaurants in Spain. Foodborne Pathog. Dis. 6, 1269–1272. doi: 10.1089/fpd.2009.0337
Soyer, Y., Richards, J., Hoelzer, K., Warnick, L. D., Fortes, E., Mcdonough, P., et al. (2013). Antimicrobial drug resistance patterns among cattle- and human-associated Salmonella strains. J. Food Prot. 76, 1676–1688. doi: 10.4315/0362-028X.JFP-13-018
Terrier, B., and Martinez, V. (2006). Salmonelosis. EMC - Tratado de Medicina 10, 1–6. doi: 10.1016/S1636-5410(06)70402-0
Thorning, T. K., Raben, A., Tholstrup, T., Soedamah-Muthu, S. S., Givens, I., and Astrup, A. (2016). Milk and dairy products: good or bad for human health? An assessment of the totality of scientific evidence. Food Nutr. Res. 60:32527. doi: 10.3402/fnr.v60.32527
Tobar-Delgado, M. E., Rodríguez-Bautista, J. L., Patiño-Burbano, R. E., Argote-Vega, F. E., Castro-Rincón, E., and Mogollón-García, H. D. (2020). Quality of raw milk in the highland tropics of Nariño, Colombia. Agron. Mesoamer. 22, 695–707. doi: 10.15517/am.v31i3.40892
Traub-Dargatz, J. L., Ladely, S. R., Dargatz, D. A., and Fedorka-Cray, P. J. (2006). Impact of heat stress on the fecal shedding patterns of Salmonella enterica typhimurium DT104 and Salmonella enterica Infantis by 5-week-old male broilers. Foodborne Pathog. Dis. 3, 178–183. doi: 10.1089/fpd.2006.3.178
Velasquez-Munoz, A., Castro-Vargas, R., Cullens-Nobis, F. M., Mani, R., and Abuelo, A. (2024). Review: Salmonella Dublin in dairy cattle. Front Vet Sci 10:1331767. doi: 10.3389/fvets.2023.1331767
Villavicencio, F., Albán, V., Satán, C., Quintana, H., Enríquez, W., Jaramillo, K., et al. (2024). Salmonella enterica Serovar Infantis KPC-2 Producer: First Isolate Reported in Ecuador. Microb. Drug Resist. 30, 502–508. doi: 10.1089/mdr.2024.0072
Vinueza-Burgos, C., Hidalgo-Arellano, L., Gómez-Coronado, C., Medina-Santana, J. L., and Cevallos-Almeida, M. (2023). Prevalence, serovars, and risk factors associated with the presence of Salmonella in pork sold in public markets in Quito, Ecuador. F1000Res 12:1367. doi: 10.12688/f1000research.138671.1
Vranješ, A. P., Popović, M., and Jevtić, M. (2015). Raw milk consumption and health. Srp. Arh. Celok. Lek. 143, 87–92. Available at: http://srpskiarhiv.rs/global/pdf/articles-2015/januar-februar/RawMilkConsumptionandHealth.pdf
Wang, J., Zhu, X., Zhao, Y., Liu, H., Zhang, Z., Yan, L., et al. (2023a). Prevalence and antimicrobial resistance of Salmonella and ESBL E. coli isolated from dairy cattle in Henan Province, China. Prev. Vet. Med. 213:105856. doi: 10.1016/j.prevetmed.2023.105856
Wang, J., Zhu, X., Zhao, Y., Xue, Y., Zhang, Z., Yan, L., et al. (2023b). Risk factors associated with Salmonella in dairy cattle farms in Henan and Hubei provinces, China. Anim. Dis. 3:20. doi: 10.1186/s44149-023-00085-9
Weisburg, W. G., Barns, S. M., Pelletier, D. A., and Lane, D. J. (1991). 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 173, 697–703. doi: 10.1128/jb.173.2.697-703.1991
WHO. (2018). Salmonella (non-typhoidal). Available online at: https://www.who.int/news-room/fact-sheets/detail/salmonella-(non-typhoidal) (accessed January 26, 2025).
WHO (2023). Typhoid fever. Available online at: https://www.who.int/es/news-room/fact-sheets/detail/typhoid?utm_source=chatgpt.com (accessed February 1, 2025).
Williams, E. N., Van Doren, J. M., Leonard, C. L., and Datta, A. R. (2023). Prevalence of Listeria monocytogenes, Salmonella spp., Shiga toxin-producing Escherichia coli, and Campylobacter spp. in raw milk in the United States between 2000 and 2019: a systematic review and meta-analysis. J. Food Prot. 86:100014. doi: 10.1016/j.jfp.2022.11.006
Keywords: Salmonella spp., raw milk, Salmonella enterica serovars, contamination, risk of infection
Citation: Loor-Giler A, Sanchez-Castro C, Robayo-Chico M, Puga-Torres B, Santander-Parra S and Nuñez L (2025) High contamination of Salmonella spp. in raw milk in Ecuador: molecular identification of Salmonella enterica serovars Typhi, Paratyphi, Enteritidis and Typhimurium. Front. Sustain. Food Syst. 9:1593266. doi: 10.3389/fsufs.2025.1593266
Edited by:
John J. Maurer, Virginia Tech, United StatesReviewed by:
Alejandro Piña-Iturbe, Pontificia Universidad Católica de Chile, ChileChelea Matchawe, Institute of Medical Research and Studies of Medicinal Plants (IMPM), Cameroon
Copyright © 2025 Loor-Giler, Sanchez-Castro, Robayo-Chico, Puga-Torres, Santander-Parra and Nuñez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luis Nuñez, ZmFiaWFubjdAeWFob28uZXM=
†ORCID: Camila Sanchez-Castro, https://orcid.org/0009-0001-6248-7681
Silvana Santander-Parra, https://orcid.org/0000-0002-8609-4399