Abstract
Improving composting efficiency by adding domestic garbage and analyzing the associated microbial mechanisms constitute the foundation for promoting the utilization of agricultural waste. The present study investigated composting efficiency and the underlying fungal mechanisms associated with different domestic garbage amendments using industrial-scale aerobic composting. The results indicated that the amendment of domestic garbage significantly improved composting efficiency, as evidenced by elevated temperatures, higher germination index (GI) values, and increased cellulose and hemicellulose transformation. Fungal community composition was found to differ according to both the composting stage (PERMANOVA, F = 14.03, p < 0.01) and treatment (PERMANOVA, F = 5.92, p < 0.01). The functional profiles of the fungal community, derived from the bioinformatic tool FUNGuild, showed that the activities of typical functional guilds—symbiotrophic (r = −0.913) and pathogenic (r = −0.926) fungi—were significantly negatively correlated with compost maturity, while saprotrophic fungi (r = 0.947) were positively correlated with composting maturity. Therefore, high-efficiency composting could be considered a process characterized by the gradual decline of symbiotrophic and pathogenic fungi, accompanied by their replacement with saprotrophic fungi. These putative functional guilds were influenced by key environmental factors, such as the C/N ratio, TOC, and cellulose content, thereby playing a significant role in enhancing composting efficiency. The greenhouse pot experiments with maize seedlings exhibited that the composting end product derived from domestic garbage amendments significantly promoted growth. Taken together, this study provides deeper insight into how functional ecological groups within the fungal community contribute to enhanced composting performance.
1 Introduction
Globally, the growing production of organic matter leads to serious environmental toxicity and economy-related challenges (Sukholthaman and Sharp, 2016). Among these, the increasing volume of rural domestic garbage has been recognized as a potential source of environmental issues and health hazards, as it contains pathogenic microbes, parasite ova, and organic micropollutants (Mao et al., 2020). Domestic garbage is mainly composed of kitchen waste, leaves, and crop residues, according to a survey conducted in typical Chinese rural regions. The annual output of domestic garbage in China was approximately 21 million tons in 2017 and is predicted to reach 40 million tons by 2025 (Shen, et al., 2023). At present, the main disposal methods for domestic garbage are sanitary landfill (53.0%) and artificial incineration (43.8%) (Yue et al., 2025). This garbage is stacked randomly, affecting the appearance of the village and polluting the ecological environment. From the perspectives of sustainable agriculture, circular economy, and environmental protection, composting offers clear advantages over current management strategies, including reducing the use of chemical fertilizers and preventing pollution. In this context, integrating composting technology with agricultural production has become a key approach to achieving sustainable development (Ayilara et al., 2020).
In response to this trend, aerobic composting has been universally recognized as an ecological method to manage large-scale waste due to its ability to hygienize and produce humified end products (Torrijos et al., 2021). Aerobic composting efficiency could be influenced by operational factors such as the C/N ratio (Wu et al., 2017), nutrient availability (Ge et al., 2022), and especially the addition of bulking material (Juarez et al., 2015). For example, amendments such as sewage sludge (Yuan et al., 2016), sawdust (Chiumenti, 2015), and biochar (Du et al., 2019) have been widely utilized with dairy manure to achieve effective composting. However, in the context of practical investigation in China, the impact of domestic garbage on the efficiency of aerobic composting has not been evaluated, and the extent to which this initial material impacts dynamic variations in microbial communities requires further research.
Composting can be considered a controlled microbiological fermentation process, in which the microbiome plays a critical role in the mineralization and biodegradation of organic materials. Fungal activities play critical roles in the biodegradation of lignocellulosic materials due to their tolerance of high temperatures, thereby contributing to composting maturity (Qiao et al., 2019). These participating fungal consortia generally proceed to experience variations in both taxonomy and ecological function according to the predominant physicochemical and nutritional characteristics of the composting stages (Gu et al., 2017). This includes a mesophilic stage characterized by the inhibition of thermotolerant microorganisms, a thermophilic stage marked by concentrative decomposition and proliferation of thermophilic microorganisms, and a mature stage characterized by an adversely mesophilic microbial consortium. As major determinants of environmental conditions, raw material properties are regarded as key drivers of fungal community succession (Onwosi et al., 2017). While the taxonomic patterns of composting fungi have been well characterized, there remains limited knowledge regarding the actual function of the fungal community due to the high variety of ecological guilds.
Previous reports indicate that fungal communities in composting systems are highly diverse assemblages composed of multiple ecologically functional groups (Hu et al., 2019). Meanwhile, growing evidence suggests that the ecological functions of fungal communities, rather than their taxonomic patterns, are compactly associated with the environmental conditions of aerobic composting (Liu et al., 2023). For example, Qiao et al. (2020) suggested that dynamic variations in pathogenic fungi directly impact aerobic composting efficiency under different C/N ratios. However, specific knowledge regarding how fungal functional groups interact with composting efficiency remains limited. In this study, the FUNGuild database was utilized as an online annotation tool to deduce the functional guilds of the identified fungal taxa based on their trophic modes (e.g., pathotroph, symbiotroph, saprotroph) (Louca et al., 2016). This approach has been utilized to decipher fungal trophic composition in various environments, such as the ocean (Shi et al., 2020), soil (Zhu et al., 2022), and especially composting systems (Kracmarova et al., 2022). It is considered a vital biological indicator of composting end-product quality due to its sensitivity to environmental variations and prompt responses (Zhang et al., 2020). Therefore, a comprehensive characterization and understanding of the fungal community and the mechanisms that enhance composting efficiency are essential for producing high-quality compost products.
To unravel the temporal functional patterns of the fungal community in response to domestic garbage amendments during aerobic composting, two contrasting initial material ratios were applied. The overall goals of this research were to: (1) explore the efficacy of differing additional ratios of domestic garbage; (2) infer the impact of different initial material ratios on fungal ecological function and its relationship with compost maturity; and (3) elucidate potential interactions between environmental conditions and functionally diverse fungal guilds according to composting stages.
2 Materials and methods
2.1 Thermophilic aerobic composting experimental design
The composting windrows were established at an industrial scale (length * width * height = 8 m * 2.6 m * 1.4 m) using mushroom residue, cow manure, and domestic garbage as the initial materials (Table 1). Fresh cow manure and domestic garbage were collected from the factory of Xinde Agricultural Industry, China. The mushroom residue was composed of corncobs, rice and corn straws, and sawdust and was obtained after mushroom production. Two composting treatments were arranged as follows: (1) DG, with domestic garbage amendments (C/N = 25.04); (2) NG, with no domestic garbage amendments (C/N = 29.23).
Table 1
| Composting material | pH | Moisture content (%) | Total C (g/kg) | Total N (g/kg) | Total P (g/kg) | Total K (g/kg) | C/N ratio | Ratio of the materials | |
|---|---|---|---|---|---|---|---|---|---|
| DG | NG | ||||||||
| Domestic garbage | 8.26 | 38.31 | 269.85 | 16.70 | 12.14 | 10.81 | 16.15 | 2000 | 0 |
| Mushroom residue | 6.57 | 61.97 | 356.28 | 8.21 | 4.61 | 3.58 | 43.40 | 4,000 | 4,000 |
| Cow manure | 8.05 | 74.92 | 316.47 | 18.79 | 16.43 | 11.09 | 16.84 | 2000 | 2000 |
Physicochemical characteristics of the raw materials and the ratio of the materials (kg, dry-weight based).
The initial moisture content of the raw material mixtures was approximately 60%, and they were arranged in stacked windrows. During the composting time, the average pore O2 concentration was 15% by volume under an aeration frequency of 0.5 L/min/kg of organic matter in the composting piles. This region had an average temperature of 14.5°C during the composting period. After thoroughly mixing, the compost blend was stirred every 5 days throughout the composting process.
For every treatment, four analytical replicates were sampled at 0, 1, 5, 9, 14, and 24 days at specific intervals according to temperature variations during composting. To assure representativeness, each pile was sampled at three heights (the lower, middle, and upper sections: 10–20 cm, 45–55 cm, 105–125 cm, respectively) and then blended uniformly to create a composite sample. Subsequently, the subsample was divided into two sections: one for the analysis of physicochemical indicators and the other stored at −70°C for subsequent DNA extraction.
2.2 Measurement methods
The temperature of the compost pile and the surrounding environment was monitored at 10 a.m., using a real-time temperature sensor. The pH value was determined using a pH meter (Shanghai Leici Portable, PHS-25) after horizontally shaking the dry samples with deionized water (1:10, w/v). The germination index (GI) value was measured using cress seed (Lepidium sativum L.) to evaluate the toxicity and maturity of the composting product. The C/N ratio, NH4+-N, NO3−-N, water-soluble carbon (WSC), and lignocellulose-related index were measured following the methods described by Qiao et al. (2019).
2.3 DNA extraction, PCR amplification, and sequencing
Frozen compost samples (5.0 g) were homogenized under liquid nitrogen. Total genomic DNA was extracted from 0.25 g of the composite sample utilizing a commercial kit (Thermo Fisher, SCIENTIFIC) following the manufacturer’s protocol. The primers ITS1F (5-GGAAGTAAAAGTCGTAACAAGG-3) and ITS 2 (5-ATGCTGCGTCTCTTCGATGC-3) were used to amplify the internal transcribed spacer 1 (ITS1) region (Claesson et al., 2009).
PCR amplification was performed using a 7,500 PCR System (Foster City, CA, USA). The reaction system contained SYBR ® Premix Ex TaqTM (2 ×) 10.0 μL, front-end primer (10 μm) 0.4 μL, back-end primer (10 μM) 0.4 μL, ROX Reference Dye II(50 ×)0.4 μL, DNA template 2.0 μL, and sterilized double-distilled water 6.8 μL. The PCR reaction procedure were 30 s at 95°C, followed by 30 cycles of 95°C for 5 s and 60°C for 60 s. All amplicons were amplified in triplicate, pooled to a final concentration of 10 nM, and sequenced using the Illumina MiSeq platform, USA.
2.4 Sequence data processing
Sequences with read lengths less than 200 bp, ambiguous base calls, and those that mismatched with the barcode were removed using FLASH V1.2.7 and QIIME 2. The average neighbor algorithm was applied to define OTUs based on sequence similarity with a threshold of 97% (Qiao et al., 2019). After quality filtering of ambiguous and singleton short OTUs, clustering yielded 2,338,256 passing fungal ITS rRNA gene sequences from 48 samples. The number of high-quality sequences per composting sample ranged from 23,123 to 67,477, with an average of 48,713. Altogether, 2,007 OTUs were obtained after the sequences were normalized to an equal depth of 23,123 sequences.
For the analysis of fungal taxa, each representative sequence was assigned based on the UNITE ITS database (Nilsson et al., 2018).
2.5 The greenhouse experiment to evaluate growth-promoting ability
To evaluate the effects of the composting product on plant growth, a maize greenhouse experiment was performed from May to July 2024. A total of four treatments were applied: (1) CK: control, without fertilizer; (2) CF: chemical fertilizer; (3) DG: composting product with domestic garbage amendments; (4) NG: composting product without domestic garbage amendments.
The greenhouse experimental soil was a clay loamy Typic Hapli-Stagnic Anthrosol. The basic properties were as follows: pH 6.78, organic matter content 15.17 g/kg, total nitrogen content 3.27 g/kg, total phosphorus content 3.58 g/kg, available phosphorus content 31.5 g/kg, and available potassium content 221 g/kg. This region had an average temperature of 22.1°C, with natural sunlight and watering every two days during the greenhouse experiment.
The composting end product was applied at 2.0% (DW, W/W) per pot containing 3.0 kg of soil mass. The pot dimensions were as follows: top diameter 17 cm, bottom diameter 13 cm, and height 18 cm. To maintain consistent nutrient levels, all treatments received the same proportions of nitrogen, phosphorus, and potassium. For each treatment, four analytical replicates were set up. Uniform maize seedlings at the 2–3 leaf stage were selected and transplanted, followed by regular watering management. Typical economic indicators, including plant height, stem diameter, leaf area, fresh and dry weights, and relative chlorophyll content (SPAD), were measured 45 days after the start of the pot experiment.
2.6 Statistical analysis
All statistical analyses were conducted using R (v4.4.4, 2024). Non-normal data were standardized when necessary to meet the assumptions of normality and homoscedasticity for statistical analyses. For alpha diversity, the richness and diversity of the observed OTUs were calculated using the Chao 1 and Shannon indices, respectively. The fungal community compositions across the composting treatments and processes were visualized using PCoA, PERMANOVA, and ANOSIM based on the Bray–Curtis dissimilarity. Spearman correlations between the abundance of fungal functional guilds and composting maturity were conducted. In addition, fungal functional mode analysis was performed using FUNGuild (Nguyen et al., 2016). SEM was performed to unravel the impacts of the critical environmental conditions, fungal functional guilds, and composting maturity.
3 Results and discussion
3.1 Variational patterns of physicochemical and biological properties
Composting efficiency mainly refers to the degree of maturity achieved by organic matter in compost after mineralization and humification processes. A basic requirement for the safe application of composting products to soil is their stability. In the present study, composting maturity was monitored using relevant factors, including temperature, water-soluble substances, lignocellulose content, and biological indicators.
Both composting piles exhibited a typical composting profile, including mesophilic (20–45°C), thermophilic (45–70°C), and mature stages (Figure 1). Compared to the ambient temperature, the composting temperature in the DG and NG treatments rapidly increased to over 50°C on day 1. Then, DG reached the thermophilic stage with a peak temperature of 73.6°C on day 3, compared to NG, which reached a maximum of 68.3°C on day 5. In addition, DG maintained temperatures above 55.0°C for more consecutive days than NG in the thermophilic stage. The significance analysis also indicated that DG (p < 0.01) maintained significantly higher temperatures than NG, supporting more efficient composting for the elimination of pathogens and parasites in DG, which aligns well with the findings of Xie et al. (2021). This might be attributed to the domestic garbage amendments, which resulted in a more ideal C/N ratio of 25.04, thereby promoting the proliferation of active indigenous microorganisms in the raw composting materials (Kulikowska, 2016).
Figure 1

Temperature variations in the piles DG and NG and the ambient environment during the composting process. Arrows on the x-axis indicate the sampling days.
Based on the temperature variations, two sampling points were selected within each of the three typical stages (mesophilic, thermophilic, and mature) to represent the composting process. The pH values of the two treatments showed an increasing trend, peaking on day 5, followed by a decreasing trend as composting progressed (Figure 2A). A decreasing tendency in the C/N ratio was observed in both piles throughout the three composting stages. The final C/N ratios in both piles were below 20, indicating that their composts had reached an acceptable standard of maturity (Petric et al., 2015). It is widely accepted that the GI is the most sensitive and reliable biological indicator to reflect phytotoxicity inhibitory effects and the degree of compost maturity (Mahapatra et al., 2022). The GI of both piles gradually increased until the mature stage. A significantly higher GI level was observed in DG (91.0%) than in NG (81.7%) when composting ended, suggesting lower phytotoxicity in DG (Yang et al., 2021). If the GI is greater than 50%, the compost is considered to be basically decomposed, while a GI value of 85% indicates full decomposition (Kong et al., 2024). Accordingly, the DG treatment demonstrated significantly better compost maturity. Taken together, our results indicated that the adjustment of domestic garbage was optimal for active composting.
Figure 2

Variations in physicochemical and biological properties. (A) pH; (B) C/N ratio; (C) germination index (GI); (D) ammonium nitrogen (NH4+-N); (E) nitrate nitrogen (NO3−-N); (F) water-soluble carbon content (WSC); (G) lignin; (H) cellulose; (I) hemicellulose. The legend shown in A applies to the others.
For the water-soluble substances, the NH4+-N content rapidly increased in DG, reaching a peak value of 6.53 mg/g on day 1, compared to a peak of 6.56 mg/g in NG on day 5. This increase was related to the ammonification process, which involves the conversion of organic nitrogen into ammonia during humification. Thereafter, the NH4+-N content decreased due to NH3 volatilization at high temperatures, immobilization into nitrogenous compounds by microbes, and the transformation of NH4+-N to NO3−-N. The NO3−-N content in the DG and NG piles showed a similar increasing trend over time (p < 0.05). The DG treatment led to a significantly higher reduction in WSC of 42.62% compared to NG (22.34%) at the end of composting. This suggests that the degradation and incorporation of WSC by microbial activity were greater than the amount of newly synthesized soluble carbon (Liu et al., 2018). That is, the WSC substances were primarily biodegraded during the mesophilic and thermophilic stages. Thereafter, a relative balance in the WSC substances revealed that microorganisms utilized these carbon sources to degrade and alter the most resistant carbon fractions of domestic garbage since composting is also a synthesis process (Hu et al., 2019).
The lignin content fluctuated to 7.49 and 7.85% in the DG and NG piles, respectively, at the end of composting. Accordingly, the cellulose content decreased by 65.11 and 61.53% in the DG and NG treatments on day 24, while the hemicellulose content decreased by 51.6 and 93.6% in the DG and NG piles, respectively. Given the highly complex nature of composting materials and their compositions, previous reports have shown considerable variability regarding the initial content of fibrous compounds (Tian et al., 2017). Specifically, the amount of lignin exhibited a slight increase on day 5, differing from the reports of Zhao et al. (2024) who studied aerobic composting of rice straws, tree branches, and pine needles. This difference may be because domestic garbage contains more carbonaceous materials, such as carbohydrates, fats, and amino acids, which degrade quickly during the mesophilic stage, thereby increasing the relative content of lignin. Cellulose was transformed principally during the thermophilic stage, while hemicellulose degradation occurred from the start of composting until day 14. Considering the importance of organic compound humification in microbial metabolism for active composting, domestic garbage amendments could promote the decomposition of lignocellulosic materials and thereby contribute to compost maturity.
Overall, these observations indicate that the amendment of domestic garbage significantly accelerated composting efficiency. This is mainly due to the high content of easily biodegradable organic matter in domestic garbage, which creates a favorable environment for microbial reproduction In addition, domestic garbage can create a suitable and relatively loose composting structure (Chen et al., 2020), which facilitates ventilation and oxygen circulation within the compost pile, thereby maintaining a high-temperature environment and promoting the humification of organic matter. Given the significance of domestic garbage as a bulking material, the composting process provides an environmentally sustainable method for its disposal. Although the amendment of domestic garbage achieves high composting efficiency, it poses some risks due to volatile organic compounds, pathogen carriers, and leachate pollution, when considering the temporal variability of domestic garbage.
3.2 Succession of α- and β-diversity of the fungal community
For α-diversity analyses, the number of sequences for each sample was rarefied to 23,123 sequences, which captured more than 97% of the observed OTU richness and diversity. Overall, the data support previous findings on fungal community diversity and richness succession during composting, which initially increase and then decrease over time (López-González et al., 2015). During the thermophilic stage, the DG treatment resulted in significantly higher richness and higher diversity on day 5 compared to NG (Figure 3A). This may be attributed to the amendment of domestic garbage increasing the supply of organic carbon substrates, thereby allowing thermophilic organisms to reflect the biochemical evolution of composting in terms of growth-supporting substrates and ecological niches. Intriguingly, the increase in fungal community diversity closely corresponded with the temperature during the mesophilic stage (Figure 3B).
Figure 3

Variations in the richness (Chao 1, A) and diversity (Shannon, B) indices. PCoA ordination based on PERMANOVA showing significant associations between community composition and both composting treatment and time (C). For (A,B) the asterisks indicate significant differences (* p < 0.05, **p < 0.01, *** p < 0.001). For (C) samples from each sampling day are represented by colored symbols.
PCoA was conducted to visualize differences in the fungal community throughout the composting developmental process across the two treatments (Figure 3C). Fungal community composition was observed to be distinct according to both the composting stage (PERMANOVA, F = 14.03, p < 0.01) and treatment (PERMANOVA, F = 5.92, p < 0.01). The composting stage was identified as the most influential factor determining fungal community composition, in line with previous reports (Qiao et al., 2020). This may be due to variations in environmental conditions that occur during each composting phase. In particular, the present result indicated a relatively short composting duration, which is likely due to the higher content of easily decomposable carbonaceous substances in domestic garbage. Regarding the composting treatment, the initial fungal communities formed aggregated clusters during the mesophilic stage, followed by significant variations in the thermophilic (PERMANOVA, F = 12.19, p < 0.01) and mature (PERMANOVA, F = 8.41, p < 0.001) phases. Intriguingly, fungal communities in DG on day 5 during the thermophilic stage were statistically similar to those in NG on day 9, suggesting a separate transitional stage present in NG. These observations indicate differences in fungal community reorganization and physiological responses between different composting materials and phases. Previous studies have reported that variations in composting fungal community composition are driven by alterations in material composition and micro-environmental conditions that occur at each stage (Jiang et al., 2020). Generally, organisms indicate non-random repercussions to resource availability due to different nutrient requirements (Duan et al., 2019). During composting progression, certain microorganisms reduce their metabolic activity to achieve dormancy, whereas others enhance their environmental adaptability to thrive in a better habitat. Different micro-environmental conditions associated with each composting stage were observed in the present investigation and were identified as the most influential factor driving fungal community composition (Giling et al., 2019). Material composition was identified as the secondary factor contributing to community variation. The amendment of domestic garbage mainly altered the C/N ratio, which was considered one of the most critical factors impacting composting maturity. As such, the initially favorable C/N ratio in the domestic garbage-amended treatment provided greater nutrient availability for fungal growth compared to the NG treatment. Consequently, the domestic garbage amendment led to an appropriate adjustment of the C/N ratio, which impacted the α- and β-diversity of the fungal community and achieved significantly higher composting efficiency.
3.3 Linking the typical functional guild patterns with composting maturity
Functional profiles of the fungal community were generated using the bioinformatic tool FUNGuild (Figure 4), and over 67.8% of high-throughput sequences were classified into three functional guilds based on their trophic modes, namely saprotrophic, pathogenic, and saprotrophic fungi. Only FUNGuild assignments with high confidence rankings (“probable”and“highly probable”) were included in the present analysis. Consequently, 33.2% of high-throughput reads were not unassigned. However, one limitation of this method is that the trophic modes assigned to fungal ecological guilds through FUNGuild are predictions and not direct measurements. In addition, the use of the ITS1F-ITS2 primer favored the phyla Ascomycota and Basidiomycota, while potentially underrepresenting Zygomycota, Chytridiomycota, and thermophilic fungi in the composting piles, which may partially affect the putative results.
Figure 4

Variation in the relative abundances of symbiotrophic (A), pathogenic (B), and saprotrophic (C) fungi, and the correlation between their relative abundances and composting maturity (D–F).
The relative abundance of putative symbiotrophic fungi tended to decrease significantly from day 0 (20.3%) to day 9 (2.3%), whereas they remained low but relatively stable during the mature stage (Figure 4A). The DG treatment, amended with DW, (1.6%) contained a lower proportion of symbiotrophs than NG (3.1%). The anaerobic animal endosymbiont fungal genus Orpinomyces is only abundant during the mesophilic stage, as it needs to receive nutrients from composting piles and is more sensitive to environmental disturbance than other microorganisms (Awasthi et al., 2018). Ultimately, the DW amendment treatment DG experienced higher temperatures and thus increased disturbance, whereas NG led to a more diverse, symbiotroph-enriched fungal community.
Pathogenic fungi showed a similar successional pattern to symbiotrophs, as their relative abundances decreased from 29.6 to 2.2% when composting ended (Figure 4B). In the raw materials, NG (25.0%) contained significantly fewer pathogenic fungi than DG (34.2%), likely due to the higher proportion of diverse DW in the DG treatment. As composting progressed, pathogenic fungi decreased sharply in DG, especially during the thermophilic stage, with both treatments revealing balanced variations by day 14. Overall, the finished compost product exhibited a substantial reduction in pathogens compared to the initial material of composting, with DG harboring significantly lower pathogen abundance than NG. Composting has been characterized as a process that gradually eliminates pathogenic fungi and increases saprotrophs (Gruninger et al., 2014). Consistent with this, the overall relative abundance of saprotrophic fungi increased from 6.5% in the raw materials to 34.6% at the end of composting (Figure 4C). A significantly higher relative abundance of saprotrophic fungi was observed in DG (39.2%) compared to NG (30.1%). This identified trade-off relationship between saprotrophic and pathogenic fungi during the composting process was confirmed by a previous study (Wang et al., 2018), supporting the idea that temperature could be one of the influential determinants during the elimination of pathogens in the composting process. Accordingly, the composition of the initial composting materials also influenced the variation in pathogenic and saprotroph fungi. Saprotrophic fungi can be efficient decomposers that promote carbon mineralization processes and may play a critical role in the accumulation of refractory carbon. Therefore, it is reasonable to suggest that saprotrophic fungi are also influenced by the type of organic matter, reflecting the composition of the composting materials. To conclude, the dynamic variations observed indicate that aerobic composting can be seen as a gradual succession of saprotrophic fungi, rather than pathogenic fungi. This finding highlights the successional patterns of pathogenic fungi and has important implications for achieving highly effective composting.
Spearman correlations between composting maturity and the relative abundances of typical functional groups were calculated to identify putative functional fungal groups (Figures 4D–F). The relative abundances of symbiotrophic and pathogenic fungi were significantly negatively correlated with composting maturity, whereas saprotrophic fungi showed a positive correlation. Interestingly, the functional fungal groups of symbiotrophic (r = −0.913, p < 0.001), pathogenic (r = −0.926, p < 0.001), and saprotrophic (r = 0.947, p < 0.001) fungi in DG showed stronger correlations with the composting stage than those in NG. While previous studies have mainly addressed the dynamic pattern of fungal functional group diversity and richness (Chang et al., 2020), fewer studies have focused on the response of functional guilds to composting maturity. In the present study, the results indicated that composting maturity could impact fungal community functional traits, which may serve as bioindicators of the composting environment. A key implication of the present study is that Spearman correlations are merely theoretical calculations; therefore, the causal mechanisms between composting maturity and the relative abundances of typical functional groups require experimental verification.
3.4 Direct and indirect interactions between fungal functional guilds and composting maturity
Understanding the interactions between environmental conditions and fungal functional guilds provides information regarding potential biological interactions, habitat relationships, and shared physiology (Wu et al., 2022). In the present study, temperature, C/N ratio, NH4+-N, cellulose, and TOC were first identified as the most influential predictors of fungal functional guilds based on random forest models. Among these, temperature showed the highest mean predictor importance (MPI) for the GI. SEM analysis further indicated that environmental factors during composting regulate composting efficiency by influencing the succession of typical fungal functional traits (Figure 5). The effect of the DG amendment on key compost properties varied throughout the composting process. During the mesophilic and thermophilic stages, DG primarily influenced fungal functional traits through changes in temperature and the C/N ratio. This was mainly due to the appropriate amendment of DG, which achieved optimized carbon and nutrient availability (Wang et al., 2020), thereby facilitating the fermentation process by triggering the succession of fungal functional guilds.
Figure 5
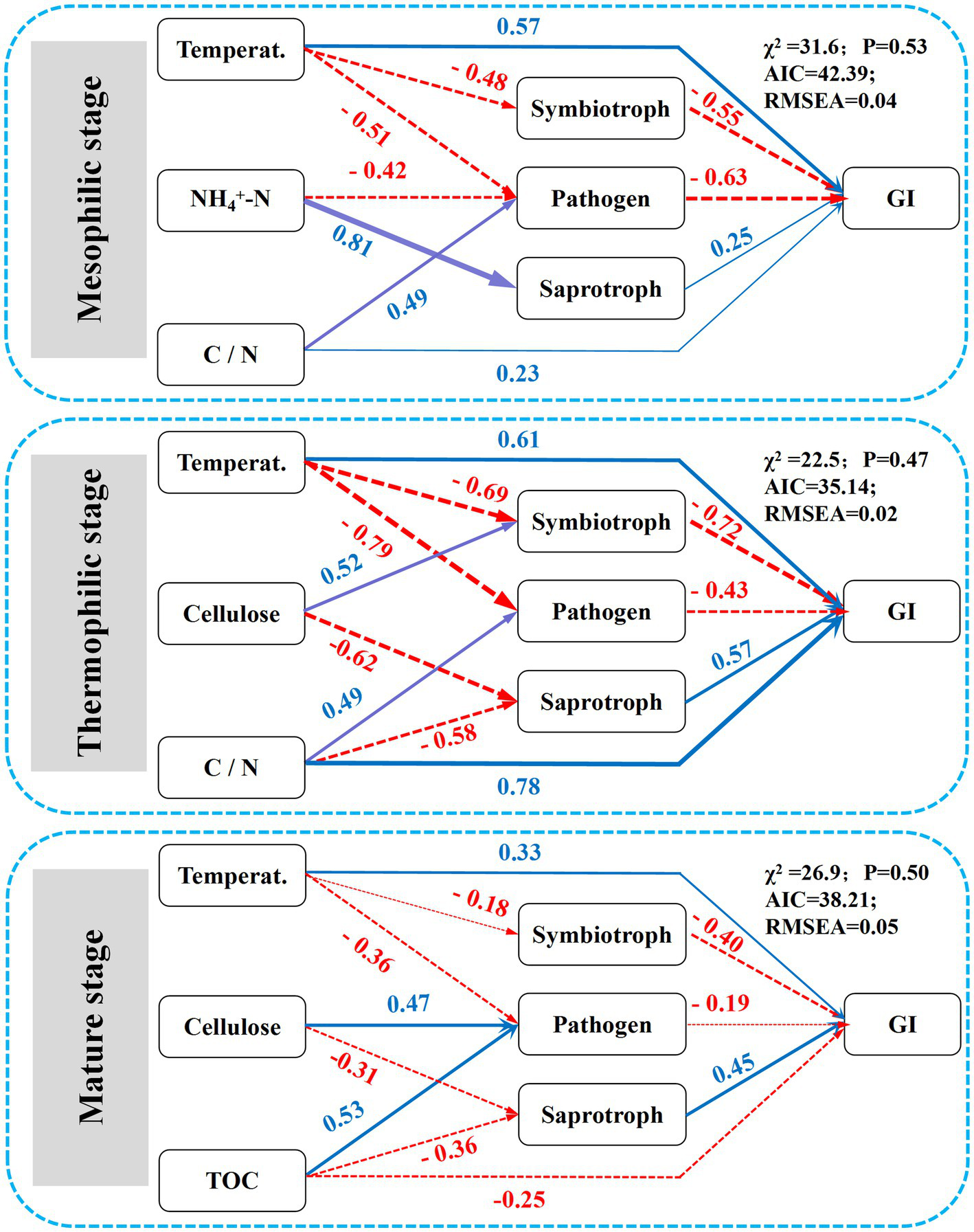
SEM illustrating the effects of the domestic garbage amendment and fungal functional guilds on composting maturity (GI value). R2 values represent the percentage of the explained variance. Temperat. means temperature. Numbers on the arrows are standardized path coefficients, illustrating their variance by the predictors. Solid blue arrows represent positive relationships, while dashed red arrows indicate negative relationships.
3.5 Pot experiment to evaluate the growth effect of the composting product on plant seedlings
The aerobic composting end products were used as cultivation media to evaluate their effect on maize seedling growth. Major growth indicators, including plant height, stem diameter, leaf area, fresh and dry weights, and SPAD, were monitored 45 days after transplanting into pots (Table 2). It should be noted that the CK treatment showed significantly lower growth indicators, indicating that the end product of DG had a significant growth-promoting effect. Plant height in the DG treatment (20.96 cm) increased by 9.79 and 11.31% compared to NG (19.09 cm) and CF (18.83 cm). Although DG increased stem diameter, leaf area, and SPAD, these differences were not statistically significant. Specifically, the fresh and dry shoot biomass values of DG were also significantly higher than those of the other treatments, whereas no significant differences were observed between NG and CF. Overall, these typical seedling growth indices revealed that the composting end product of DG was effective in promoting plant growth. Generally, better maturity of the composting end product means that organic matter can be hydrolyzed into smaller molecules, thereby improving fertilizer utilization (Wu et al., 2020) and mitigating the content of harmful substances in crops. In addition, the composting end product of DG improved soil structure and increased soil permeability, thereby effectively regulating plant physiological metabolism to promote crop growth compared to the NG and CF treatments (Wang et al., 2023). Therefore, the greenhouse experiments with maize seedlings showed that the amendment of domestic garbage significantly increased composting maturity and promoted plant growth. It is worth mentioning that including more typical plants and an appropriate timeframe should be considered in future greenhouse studies to better elucidate the effects of the end product on plant growth.
Table 2
| Treatment | Plant height (cm) | Stem diameter (mm) | Leaf area (cm2) | Fresh weight (g) | Dry weight (g) | SPAD |
|---|---|---|---|---|---|---|
| CK | 9.64 ± 0.72c | 2.83 ± 0.28c | 47.29 ± 1.96c | 1.64 ± 0.21c | 0.11 ± 0.03c | 27.84 ± 0.97b |
| CF | 18.83 ± 0.38b | 5.70 ± 0.26ab | 81.36 ± 2.82a | 4.60 ± 0.19b | 0.51 ± 0.03b | 30.84 ± 0.97a |
| NG | 19.09 ± 0.42b | 5.87 ± 0.20ab | 83.66 ± 4.26a | 4.59 ± 0.16b | 0.63 ± 0.02b | 32.36 ± 1.15a |
| DG | 20.96 ± 0.39a | 5.96 ± 0.14a | 83.90 ± 5.17a | 4.94 ± 0.23a | 0.78 ± 0.11a | 33.68 ± 1.05a |
Effects of the composting end product on maize seedling growth.
4 Conclusion
This study demonstrated that domestic garbage amendment could lead to distinct fungal metabolic succession, contributing to high-efficiency amoebic composting. The relative abundances of typical functional groups, including symbiotrophic and pathogenic fungi, were significantly negatively correlated with composting maturity, whereas saprotrophic fungi were positively correlated. Thus, thermophilic aerobic composting can be viewed as a process of gradual succession of symbiotrophic and pathogenic fungi, rather than saprotrophic fungi. Overall, this study clarifies that the variational succession of putative functional fungi significantly contributes to high-efficiency composting. The study suffers from limitations associated with the temporal variability of domestic garbage composition, which should be addressed in future studies. In addition, the induction of fungal functional guilds remains a topic for future research aimed at elucidating their role in sustainable strategies for promoting plant growth.
Statements
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
CQ: Funding acquisition, Investigation, Methodology, Writing – original draft. LB: Data curation, Formal analysis, Writing – original draft. QZ: Investigation, Methodology, Writing – review & editing. XY: Methodology, Project administration, Resources, Writing – review & editing. LR: Conceptualization, Writing – review & editing. WW: Project administration, Supervision, Validation, Writing – review & editing. JW: Resources, Software, Validation, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by the National Natural Science Foundation of China (32402677), the Anhui Provincial Natural Science Foundation (2108085QC126), the Anhui Province Scientific Research Project of Colleges and Universities (2022AH051643), the University Synergy Innovation Program of Anhui Province (GXXT-2023-102), the Research and Development of Rice–Wheat (Oil) Mechanization and Intelligent Production Management Technology and Equipment (2022yfd2301402-3), and the Talent Introduction Project of Anhui Science and Technology University (Zhyj202003).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsufs.2025.1600462/full#supplementary-material
References
1
AwasthiS. K.WongJ.LiJ.WangQ.ZhangZ.KumarS.et al. (2018). Evaluation of microbial dynamics during postconsumption food waste composting. Bioresour. Technol.251, 181–188. doi: 10.1016/j.biortech.2017.12.040
2
AyilaraM. S.OlanrewajuO. S.BabalolaO. O.OdeyemiO. (2020). Waste management through composting: challenges and potentials. Sustain. For.12:4456. doi: 10.3390/su12114456
3
ChangF.JiaF.LvR.ZhenL.LiY.WangY. (2020). Changes in structure and function of bacterial and fungal communities in open composting of Chinese herb residues. Can. J. Microbiol.66, 194–205. doi: 10.1139/cjm-2019-0347
4
ChenS.HuangJ.XiaoT.GaoJ.BaiJ.LuoW.et al. (2020). Carbon emissions under different domestic waste treatment modes induced by garbage classification: case study in pilot communities in Shanghai, China. Sci. Total Environ.717:137193. doi: 10.1016/j.scitotenv.2020.137193
5
ChiumentiA. (2015). Complete nitrification-denitrification of swine manure in a full-scale, non-conventional composting system. Waste Manag.46, 577–587. doi: 10.1016/j.wasman.2015.07.035
6
ClaessonM. J.O'SullivanO.WangQ.NikkiläJ.MarchesiJ. R.SmidtH.et al. (2009). Comparative analysis of pyrosequencing and a phylogenetic microarray for exploring microbial community structures in the human distal intestine. PLoS One4:e6669. doi: 10.1371/journal.pone.0006669
7
DuJ.ZhangY.QuM.YinY.FanK.HuB.et al. (2019). Effects of biochar on the microbial activity and community structure during sewage sludge composting. Bioresour. Technol.272, 171–179. doi: 10.1016/j.biortech.2018.10.020
8
DuanY.AwasthiS. K.LiuT.ChenH.ZhangZ.WangQ.et al. (2019). Dynamics of fungal diversity and interactions with environmental elements in response to wheat straw biochar amended poultry manure composting. Bioresour. Technol.274, 410–417. doi: 10.1016/j.biortech.2018.12.020
9
GeM.ShenY.DingJ.MengH.ZhouH.ZhouJ.et al. (2022). New insight into the impact of moisture content and pH on dissolved organic matter and microbial dynamics during cattle manure composting. Bioresour. Technol.344:126236. doi: 10.1016/j.biortech.2021.126236
10
GilingD. P.BeaumelleL.PhillipsH. R.CesarzS.EisenhauerN.FerlianO.et al. (2019). Niche for ecosystem multi-functionality in global change research. Glob. Change Biol.25, 763–774. doi: 10.1111/gcb.14528
11
GruningerR. J.PuniyaA. K.CallaghanT. M.EdwardsJ. E.YoussefN.DagarS. S.et al. (2014). Anaerobic fungi., phylum Neocallimastigomycota: advances in understanding their taxonomy, life cycle, ecology, role and biotechnological potential. FEMS Microbiol. Ecol.90, 1–17. doi: 10.1111/1574-6941.12383
12
GuW.LuS.TanZ.XuP.XieK.LiX.et al. (2017). Fungi diversity from different depths and times in chicken manure waste static aerobic composting. Bioresour. Technol.239, 447–453. doi: 10.1016/j.biortech.2017.04.047
13
HuT.WangX.ZhenL.GuJ.ZhangK.WangQ.et al. (2019). Effects of inoculating with lignocellulose-degrading consortium on cellulose-degrading genes and fungal community during cocomposting of spent mushroom substrate with swine manure. Bioresour. Technol.291:121876. doi: 10.1016/j.biortech.2019.121876
14
JiangX.DengL.MengQ.SunY.HanY.WuX.et al. (2020). Fungal community succession under influence of biochar in cow manure composting. Environ. Sci. Pollut. Res.27, 9658–9668. doi: 10.1007/s11356-019-07529-1
15
JuarezM. F.Pr ahauserB.WalterA.InsamH.Franke-WhittleI. H. (2015). Co-composting of biowaste and wood ash, influence on a microbially driven-process. Waste Manag.46, 155–164. doi: 10.1016/j.wasman.2015.09.015
16
KongY.ZhangJ.ZhangX.GaoX.YinJ.WangG.et al. (2024). Applicability and limitation of compost maturity evaluation indicators: a review. Chem. Eng. J.489:151386. doi: 10.1016/j.cej.2024.151386
17
KracmarovaM.UhlikO.StrejcekM.SzakovaJ.CernyJ.BalikJ.et al. (2022). Soil microbial communities following 20 years of fertilization and crop rotation practices in the Czech Republic. Environ. Microbiome17, 1–18. doi: 10.1186/s40793-022-00406-4
18
KulikowskaD. (2016). Kinetics of organic matter removal and humification progress during sewage sludge composting. Waste Manag.49, 196–203. doi: 10.1016/j.wasman.2016.01.005
19
LiuQ.HeX.WangK.LiD. (2023). Biochar drives humus formation during composting by regulating the specialized metabolic features of microbiome. Chem. Eng. J.458:141380. doi: 10.1016/j.cej.2023.141380
20
LiuH.ZhongX.HuangY.QiaoC.ShaoC.LiR.et al. (2018). Production of free amino acid and short peptide fertilizer from rapeseed meal fermentation using Bacillus flexus NJNPD41 for promoting plant growth. Pedosphere28, 261–268. doi: 10.1016/S1002-0160(18)60012-8
21
López-GonzálezJ. A.Del Carmen Vargas-GarcíaM.LópezM. J.Suárez-EstrellaF.del Mar JuradoM.MorenoJ. (2015). Biodiversity and succession of mycobiota associated with agricultural lignocellulosic waste-based composting. Bioresour. Technol.187, 305–313. doi: 10.1016/j.biortech.2015.03.124
22
LoucaS.ParfreyL. W.DoebeliM. (2016). Decoupling function and taxonomy in the global ocean microbiome. Science353, 1272–1277. doi: 10.1126/science.aaf4507
23
MahapatraS.AliM. H.SamalK. (2022). Assessment of compost maturity-stability indices and recent development of composting bin. Energy Nexus6:100062. doi: 10.1016/j.nexus.2022.100062
24
MaoH.WangK.WangZ.PengJ.RenN. (2020). Metabolic function, trophic mode, organics degradation ability and influence factor of bacterial and fungal communities in chicken manure composting. Bioresour. Technol.302:122883. doi: 10.1016/j.biortech.2020.122883
25
NguyenN. H.SongZ.BatesS. T.BrancoS.TedersooL.MenkeJ.et al. (2016). FUNGuild: an open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol.20, 241–248. doi: 10.1016/j.funeco.2015.06.006
26
NilssonR. H.LarssonK. H.TaylorA. F. S.Bengtsson-PalmeJ.JeppesenT. S.SchigelD.et al. (2018). The UNITE database for molecular identification of fungi: handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res.47, D259–D264. doi: 10.1093/nar/gky1022
27
OnwosiC. O.IgbokweV. C.OdimbaJ. N.EkeI. E.NwankwoalaM. O.IrohI. N.et al. (2017). Composting technology in waste stabilization: on the methods, challenges and future prospects. J. Environ. Manag.190, 140–157. doi: 10.1016/j.jenvman.2016.12.051
28
PetricI.AvdihodžićE.IbrićN. (2015). Numerical simulation of composting process for mixture of organic fraction of municipal solid waste and poultry manure. Ecol. Eng.75, 242–249. doi: 10.1016/j.ecoleng.2014.12.003
29
QiaoC.PentonC. R.LiuC.ShenZ.OuY.LiuZ.et al. (2019). Key extracellular enzymes triggered high-efficiency composting associated with bacterial community succession. Bioresour. Technol.288:121576. doi: 10.1016/j.biortech.2019.121576
30
QiaoC.PentonC. R.LiuC.TaoC.DengX.OuY.et al. (2020). Patterns of fungal community succession triggered by C/N ratios during composting. J. Hazard. Mater.401:123344. doi: 10.1016/j.jhazmat.2020.123344
31
ShenY.ZhuT.KumarR.KumarA.ChenS. (2023). Domestic garbage classification and incentive-based policies in China: an empirical Analysis. Water.154: 4074. doi: 10.3390/w15234074
32
ShiM.ZhaoY.ZhuL.SongX.TangY.QiH.et al. (2020). Denitrification during composting: biochemistry, implication and perspective. Int. Biodeterior. Biodegrad.153:105043. doi: 10.1016/j.ibiod.2020.105043
33
SukholthamanP.SharpA. A. (2016). System dynamics model to evaluate effects of source separation of municipal solid waste management: a case of Bangkok, Thailand. Waste Manag.52, 50–61. doi: 10.1016/j.wasman.2016.03.026
34
TianX.YangT.HeJ.ChuQ.JiaX.HuangJ. (2017). Fungal community and cellulose-degrading genes in the composting process of Chinese medicinal herbal residues. Bioresour. Technol.241, 374–383. doi: 10.1016/j.biortech.2017.05.116
35
TorrijosV.DopicoD. C.SotoM. (2021). Integration of food waste composting and vegetable gardens in a university campus. J. Clean. Prod.315:128175. doi: 10.1016/j.jclepro.2021.128175
36
WangX.KongZ.WangY.WangM.LiuD.ShenQ. (2020). Insights into the functionality of fungal community during the large scale aerobic co-composting process of swine manure and rice straw. J. Environ. Manag.270:110958. doi: 10.1016/j.jenvman.2020.110958
37
WangY.LiuZ.HaoX.WangZ.WangZ.LiuS.et al. (2023). Biodiversity of the beneficial soil-borne fungi steered by Trichoderma-amended biofertilizers stimulates plant production. NPJ Biofilms Microbiomes9:46. doi: 10.1038/s41522-023-00416-1
38
WangK.YinX.MaoH.ChuC.TianY. (2018). Changes in structure and function of fungal community in cow manure composting. Bioresour. Technol.255, 123–130. doi: 10.1016/j.biortech.2018.01.064
39
WuX.AmanzeC.YuZ.LiJ.LiuY.ShenL.et al. (2022). Evaluation of fungal community assembly and function during food waste composting with Aneurinibacillus sp. LD3 inoculant. Bioresour. Technol.363:127923. doi: 10.1016/j.biortech.2022.127923
40
WuS.ShenZ.YangC.ZhouY.LiX.ZengG.et al. (2017). Effects of C/N ratio and bulking agent on speciation of Zn and cu and enzymatic activity during pig manure composting. Int. Biodeterior. Biodegrad.119, 429–436. doi: 10.1016/j.ibiod.2016.09.016
41
WuN.XieS.ZengM.XuX.LiY.LiuX.et al. (2020). Impacts of pile temperature on antibiotic resistance, metal resistance and microbial community during swine manure composting. Sci. Total Environ.744:140920. doi: 10.1016/j.scitotenv.2020.140920
42
XieG.KongX.KangJ.SuN.FeiJ.LuoG. (2021). Fungal community succession contributes to product maturity during the co-composting of chicken manure and crop residues. Bioresour. Technol.328:124845. doi: 10.1016/j.biortech.2021.124845
43
YangY.WangG.LiG.MaR.KongY.YuanJ. (2021). Selection of sensitive seeds for evaluation of compost maturity with the seed germination index. Waste Manag.136, 238–243. doi: 10.1016/j.wasman.2021.09.037
44
YuanJ.ChadwickD.ZhangD.LiG.ChenS.LuoW.et al. (2016). Effects of aeration rate on maturity and gaseous emissions during sewage sludge composting. Waste Manag.56, 403–410. doi: 10.1016/j.wasman.2016.07.017
45
YueJ.ChenS.WengZ. (2025). Rural household garbage sorting for sustainable development: contributing to substantial health improvements in China. Sustainability17:4255. doi: 10.3390/su17104255
46
ZhangC.GaoZ.ShiW.LiL.ZhouB. (2020). Material conversion, microbial community composition and metabolic functional succession during green soybean hull composting. Bioresour. Technol.316:123823. doi: 10.1016/j.biortech.2020.123823
47
ZhaoJ.ZhangY.CongH.ZhangC.WuJ. (2024). Quantifying the contribution of lignin to humic acid structures during composting. Chem. Eng. J.492:152204. doi: 10.1016/j.cej.2024.152204
48
ZhuD.SethupathyS.GaoL.NawazM. Z.ZhangW.JiangJ.et al. (2022). Microbial diversity and community structure in deep-sea sediments of South Indian Ocean. Environ. Sci. Pollut. Res.29, 45793–45807. doi: 10.1007/s11356-022-19157-3
Summary
Keywords
domestic garbage, aerobic composting, composting efficiency, fungal community, functional guild
Citation
Qiao C, Bao L, Zhang Q, Yiqin X, Ren L, Wu W and Wang J (2025) Fungal functional metabolism succession contributes to product efficiency during co-composting of domestic garbage and cow manure. Front. Sustain. Food Syst. 9:1600462. doi: 10.3389/fsufs.2025.1600462
Received
28 March 2025
Accepted
22 May 2025
Published
06 June 2025
Volume
9 - 2025
Edited by
Çağrı Akyol, Ghent University, Belgium
Reviewed by
Ana Vanessa Aguilar-Paredes, Scuola Sant'Anna di Studi Avanzati, Italy
Chang Yuan, China Agricultural University, China
Updates
Copyright
© 2025 Qiao, Bao, Zhang, Yiqin, Ren, Wu and Wang.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lantian Ren, 1049642413@qq.com
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.