 Liangzhe Yang1,2
Liangzhe Yang1,2 Kui Wu
Kui Wu- 1Hubei Institute of Geosciences, Hubei Geological Bureau, Wuhan, China
- 2Hubei Key Laboratory of Resource and Ecological Environment Geology, Hubei Geological Bureau, Wuhan, Hubei, China
This study reports the first discovery of bivalves (family Alatoconchidae) from the Middle Permian Maokou Formation in Jingmen City, Hubei Province, South China. These newly identified fossil localities bridge a significant geographic gap in alatoconchid distribution, providing new insights into their paleobiogeography and paleoecology. Their frequent occurrence in warm, shallow-marine environments supports the view that South China provided an ideal ecological setting for these organisms during the Guadalupian. Furthermore, the absence of alatoconchids in younger strata suggests their extinction was closely linked to the Guadalupian-Lopingian boundary mass extinction event.
1 Introduction
As the final period of the Paleozoic Era, the Permian witnessed numerous global-scale biological evolution and environmental upheavals (Henderson et al., 2020). During this time, the southern Gondwana continent collided with the northern Laurasia continent, forming the supercontinent Pangaea, which had a profound impact on Earth’s systems (Scotese, 2009). Alongside this continental assembly, large-scale volcanic eruptions occurred and significantly influenced the Earth environment (Shen et al., 2019). Earlier eruptions of large igneous provinces (LIP) in the Early-Middle Permian are thought to have caused the end of the Late Palazeozoic Ice Age, which lasted for about 100 million years (Jurikova et al., 2025). Later eruptions, including the Emeishan LIP in the Middle Permian and the Siberian LIP at the end of the Permian, are thought to have triggered two mass extinctions separately, namely, the Capitanian (end-Guadalupian) and the end-Permian mass extinctions (Burgess et al., 2014; Stanley, 2016; Huang et al., 2025). The latter event, which wiped out 81%–94% of marine species, was the most severe mass extinction of the Phanerozoic (Stanley, 2016). During the end-Permian mass extinction, larger-bodied clades among conodonts, foraminifers, bivalves, and brachiopods experienced higher extinction rates (Dal Corso et al., 2022). Although lower in overall extinction rate, the Capitanian mass extinction had a severe impact on shallow-marine taxa, incuding fusulinacean foraminifers, corals, bivalves, and ammonoids (Wignall et al., 2009; Bond et al., 2010).
Like the end-Permian mass extinction, the Capitanian mass extinction also exhibited body-size selectivity among marine clades, including the extinction of the “tropical trio” (Verbeekinidae, Alatoconchidae, Waagenophullidae) (Isozaki, 2011; Feng et al., 2020). As one of the most distinctive fossils of the Permian, bivalves of the family Alatoconchidae are known for their aberrant morphology and exceptional size, with some individuals reaching lengths of approximately 1 m (e.g., Isozaki and Ajinović, 2009; Chen et al., 2018; Isozaki, 2024). The exceptional size of alatoconchids, compared to other bivalves, might have been an adaptation to a specific ecological niche, possibly linked to symbiosis and increased metabolic efficiency in nutrient-rich, shallow-marine environments (Isozaki and Ajinović, 2009; Asato et al., 2017; Chen et al., 2021). These bivalves, characterized by wing-like flanges and a two-layered shell structure, have been interpreted as epifaunal suspension feeders potentially engaging in photo- and/or chemo-symbiosis (Asato et al., 2017; Chen et al., 2018). They are typically found in biostromes forming “alatoconchid beds,” which can be several meters thick (Chen et al., 2018; Isozaki, 2024). However, many of these fossils were often recrystallized, deformed or broken due to sediment compaction, making them difficult to distinguish from the surrounding strata.
Since their initial description in Japan, Alatoconchidae fossils have been reported from 14 localities worldwide, including recently discovered occurrences in Kesennuma, northeastern Japan, and Partizansk, southern Primorye, Russia (Isozaki, 2024). Among these, South China−located near the paleoequator and characterized by widespread carbonate platforms development−has emerged as a major biogeographic center for alatoconchid proliferation during the Guadalupian (Yancey and Boyd, 1983). More than thirty occurrences have been reported from South China, particularly concentrated in the Dian-Qian-Gui Basin and the Upper Yangtze region (Chen et al., 2018). These fossils are typically preserved within bioclastic limestone or shell-rich beds and have often been found in association with other reef-building organisms such as fusulinids and corals. Due to their unique morphology, many early finds were mistakenly identified as large phylloid algae before more detailed morphological and microstructural studies confirmed their bivalve affinities. Reported specimens show considerable variation in size and preservation state, ranging from large, nearly complete shells to small, fragmented remains embedded in matrix. Despite these extensive discoveries, only a single locality has been reported from the Lower Yangtze region, and, until now, no occurrences had been documented from the Middle Yangtze region, leaving a notable geographic gap in the distribution of Alatoconchidae across South China (Chen et al., 2018). This study reports the first confirmed records of alatoconchid bivalves from two new localities in Jingmen City, Hubei Province−within the Middle Yangtze region−thus filling this important gap. Their sedimentary context suggests they thrived in a shallow-marine setting, consistent with previous occurrences in the Upper Yangtze and Dian-Qian-Gui regions. The discovery not only extends the paleogeographic range of Alatoconchidae but also raises new questions regarding their dispersal mechanisms and ecological adaptations. Furthermore, the findings contribute to a better understanding of the environmental conditions preceding the Capitanian mass extinction, shedding light on the possible biotic responses to this event.
2 Geological setting and methods
During the Permian, the supercontinent Pangaea was assembled, and the rapid injection of carbon dioxide into the atmosphere marked the end of the Late Paleozoic Ice Age (Jurikova et al., 2025). However, the South China Block was not incorporated into Pangaea; instead, it was positioned in the eastern part of the Paleo-Tethys Ocean (Scotese, 2009; 2021; Henderson et al., 2020). Furthermore, being located near the equator, the South China Block experienced a generally warm and humid climate after the Late Paleozoic Ice Age, leading to the deposition of extensive coal- and gas-bearing strata in different environmental settings (Shen et al., 2019).
Coinciding with the waning of the Late Paleozoic Ice Age, widespread massive chert deposits formed in South China, likely influenced by contemporaneous tectonic and volcanic activity (Zhang et al., 2023). The Middle Permian strata of the South China Block are primarily represented by the Gufeng and Maokou Formations. The Gufeng Formation consists predominantly of shale and chert, whereas the Maokou Formation is mainly composed of thick-bedded limestone interbedded with chert nodules (Shen et al., 2019; Zhang et al., 2023). In Hubei Province, which is part of the Middle Yangtze region, the Maokou Formation exhibits massive limestone with abundant chert nodules, with the depositional environment deepening northward (Figure 1; Shen et al., 2019). During the Guadalupian, the Maokou Formation in central Hubei was deposited on a shallow tropical carbonate ramp, characterized by bioclastic grainstones and well-oxygenated micrites (Scotese, 2009; Shen et al., 2019). This stable, shallow-marine setting supported abundant benthic communities—crinoids, fusulinids, phylloid algae—and enabled the in situ preservation of our disarticulated alatoconchid shell fragments. Overlying the Maokou Formation, the Upper Permian Wujiaping Formation represents one of the best-developed Permian sedimentary successions in South China, displaying diverse lithofacies ranging from fluvial settings near the highland margin to swamp, shallow marine, and deep-water basins (Shen et al., 2019; Hou et al., 2020). In Hubei Province, the Wujiaping Formation is commonly subdivided into the Longtan and Xiayao Formations. The Longtan Formation consists of thin-bedded argillaceous shales containing abundant spherical pyrite, while the Xiayao Formation comprises medium-to thick-bedded limestone and marlstone (Shen et al., 2019). A parallel unconformity developed between the Maokou and Wujiaping (or Longtan) Formations due to the Dongwu movement, which significantly influenced the sedimentary framework of South China (Shen et al., 2019; Hou et al., 2020).
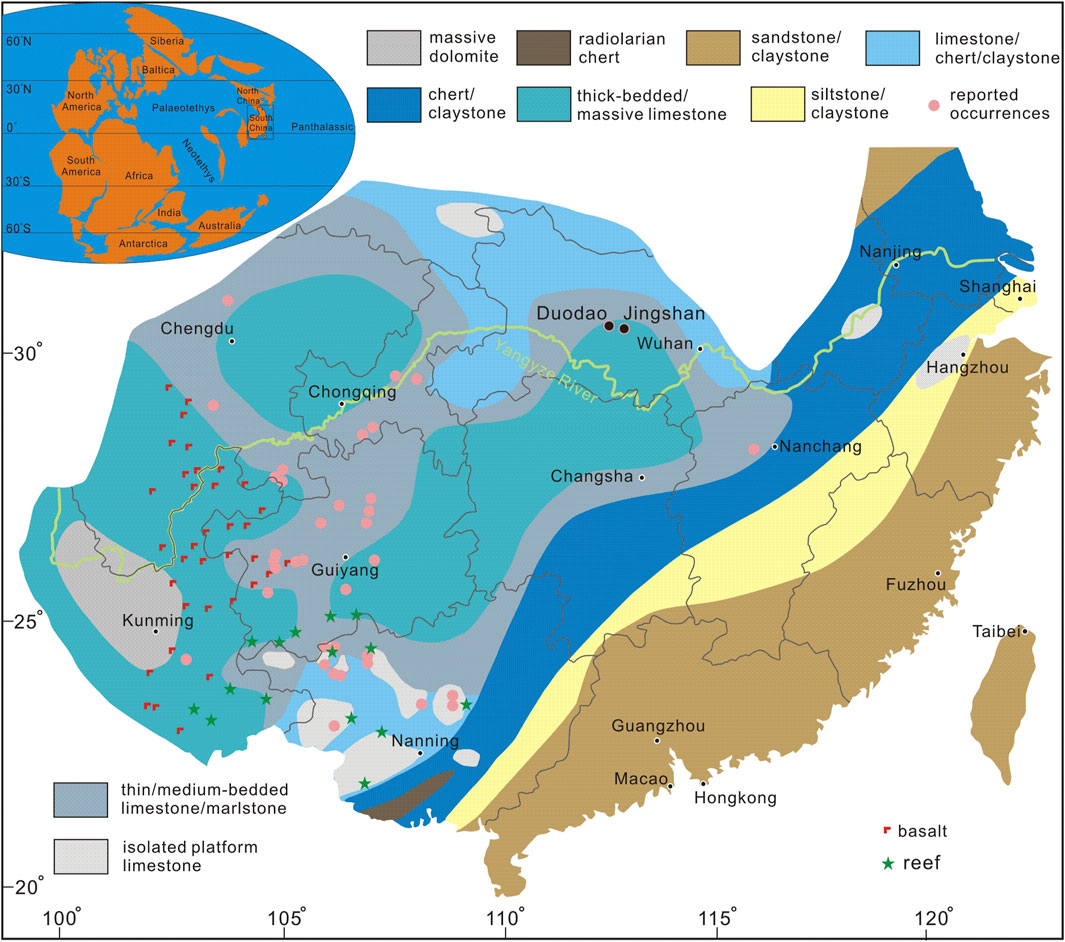
Figure 1. Late Guadalupian lithofacies of South China with occurrences of Alatoconchidae (modified from Chen et al., 2018).
Jingmen City, located in central Hubei Province, preserves Middle and Upper Permian strata that exhibit characteristics similar to those observed elsewhere in the province. However, the strata in this region are notably steeply dipping to overturned due to regional tectonic deformation, leading to complex bedding of the Longtan Formation (Figure 2). Further, due to its loose and easily weathered lithology, the Longtan Formation in this area is often poorly exposed.

Figure 2. Outcrop photos of the Duodao section. (A) the underlying shallow-grey limestone with chert nodules and the biostrome (outlined by red dash line); (B) the overlying shallow-grey limestone with chert nodules and the biostrome (outlined by red dash line); (C) the biostrome with large amounts of bivalve shells; (D) the biostrome with chert nodules; (E) the whole view of the Duodao section; red dash line outlines the biostrome.
To investigate the occurrence of alatoconchid fossils in the Permian carbonate strata of central Hubei, a systematic paleontological survey was conducted in the Jingmen region. Fieldwork focused on fresh rock exposures, with particular attention given to fossil-bearing horizons. Two key fossiliferous sites were identified: the Duodao section and the Jingshan section (Figure 1). These newly discovered specimens were found within carbonate-dominated deposits of the Middle Permian and occur as disarticulated but well-preserved shell fragments, indicating minimal post-mortem transport. The stratigraphic context of these fossils was documented, and collected samples were subjected to taxonomic identification. Accompanying fossil assemblages were also recorded to establish a biostratigraphic framework. Fossil specimens were examined using a ZEISS Axioscope microscope equipped with an Axiocam 508 camera for high-resolution imaging. Additionally, thin sections were prepared for petrographic analysis to assess the microstructure and diagenetic history of the fossils. All collected specimens are permanently housed in the collections of the Hubei Institute of Geosciences, with specimen numbers available upon request.
The findings from these investigations contribute to the understanding of the spatial distribution of alatoconchid fossils in South China. The discovery of these fossils from two previously unreported localities in Jingmen significantly expands their known range in the Middle Yangtze region, offering new insights into their paleoecological and paleogeographic significance during the Permian.
3 Results
The Duodao section (E: 112°10′43.48″, N: 31°02′26.57″) consists of a relatively continuous Permian carbonate succession, dominated by thick-bedded limestone interspersed with siliceous nodules. This section is exposed along a country road (Figure 2), with several hundred meters of limestone containing chert nodules belonging to the Qixia and Maokou Formations discontinuously exposed below it.
Beneath the alatoconchid biostrome—located in the carbonate-dominated beds of the Middle Permian Maokou Formation—in the Duodao section, a diverse fossil assemblage is present, including bivalves, brachiopods, rugose corals, calcareous algae, gastropods, bryozoans, and crinoids. Alatoconchid specimens were identified within bioclastic limestone in the upper part of the section, where no other fossils were found except for crinoid fragments (Figure 3). Although most alatoconchid specimens are fragmented due to diagenesis, several retain diagnostic morphological features, including remnants of the characteristic wing-like flanges and shell layering.

Figure 3. Close-up views of the Alatoconchid bivalves from the Duodao section; (A) and (B) are mostly fragments of the bivalves, including different parts; (C) the edge of the flanges; (D) the anterior part of the shell and near the peak.
The Jingshan section (E: 112°50′54.55″, N: 30°53′39.02″) is composed of grayish-white, thick-bedded limestone exposed along a village road. Fossil fragments found in fallen rock samples exhibit morphological characteristics consistent with alatoconchid-bearing layers from other regions. Alatoconchid material occurs as disarticulated shell fragments, most measuring 2–8 cm in length and preserving the characteristic prismatic layer and thin foliate laminae. Although many fragments show moderate recrystallization and edge chipping, several specimens retain distinct flange morphologies and shell layering (Figure 4). However, the source rock of these samples has not been identified. Among the recovered specimens, Verbeekina sp., a fusulinid index fossil indicative of the Middle Permian, was identified. Additionally, crinoid fragments were found associated with alatoconchid shells (Figure 4).

Figure 4. Alatoconchid bivalves from the Jingshan section; (A) Biostrome with large amounts of broken shells of Alatoconchid bivalves; (B) shell fragments of Alatoconchid bivalves, and co-occurred crinoids and fisulinids; (C) close up view of the part in the red box of (B), showing the details of the external prismatic layer and the internal recrystallized part; (D) co-occurred crinoids and fisulinids (Verbeekina sp.) with Alatoconchidae; (E) close up view of the part in the red box of (D).
All shell fragments from the Duodao and Jingshan sections are tightly embedded in the surrounding limestone, making their extraction impractical. The bivalve shells from both sections are highly fragmented, with most specimens measuring less than 10 cm in length and 1–5 mm in thickness, though the largest recovered specimen, from the Jingmen section, reaches approximately 15 cm in length (Figure 3). Shell thickness is generally less than 3 mm, indicating a high aspect ratio between width and thickness.
One of the most distinctive features of these shells is the tight folding along their margins (Figures 3A–C). A specimen from the Duodao section exhibits a pronounced U-shaped extension along the lateral edge of the valve (Figure 3C). Another articulated specimen preserves both valves and the dorsal niche between them (Figure 3D), which further supports its identification as an alatoconchid. Unfortunately, shell fragments from the Jingshan section are highly fragmented but occur in significantly greater abundance (Figure 4A). One specimen from this section exhibits similar dimensions and shell morphology to those from the Duodao section (Figures 4B, C), suggesting a comparable taxonomic affinity. The recovered shell fragments from the Jingshan section are predominantly disarticulated and embedded within the limestone matrix, making complete extraction challenging. The measurable fragments range from 3 to 5 cm in length and 1.0–5.2 mm in thickness. These fragments exhibit the characteristic broad, wing-like flanges and thin shell walls typical of Alatoconchidae. The shell microstructure consists of an outer prismatic layer and an inner foliated layer, each approximately 3 mm thick. These features are consistent with those described in previous studies of Alatoconchidae specimens.
4 Discussion
4.1 Taxonomic identification and comparison with Alatoconchidae
Phylum Mollusca Linnaeus, 1758.
Class Bivalvia Linnaeus, 1758.
Order Pteriida Newell, 1965.
Superfamily Ambonychioidea Miller, 1877.
Family Alatoconchidae Termier et al., 1973.
Genus et. sp. indet.
Diagnosis (from Yancey and Boyd, 1983): Large to very large equivalved shells with elongate or circular outline in dorsal view; shell wall thick, with massive infilling of umbonal cavity, but thin on leading edge, beaks commonly terminal; umbonal carina alate, compressed dorsoventrally and laterally produced, resulting indistinctive wing-like flanges formed by reflexed valve wall; very large flat ventral surface, bounded by umbonal carina; ligament duplivincular, byssal groove present, but non-functional in adults of some species; outer shell layer composed of very large simple prisms set perpendicular to shell surface.
Materials: Specimens collected from the Maokou Formation at the Duodao and Jingshan sections, Jingmen City, Hubei Province, South China. All specimens are preserved as internal and external molds and are generally fragmentary.
Description and remark: These fragments exhibit valves that are dorsoventrally compressed yet strongly laterally extended into broad, wing-like flanges, a hallmark of Alatoconchidae morphology. The shells measure 3–5 mm in total thickness and reveal a distinctive double-layered microstructure comprising an outer prismatic calcite layer and an inner foliated layer, each averaging approximately 3 mm in thickness. Notably, hinge regions lack true hinge teeth, consistent with other members of Alatoconchidae and distinguishing them from hinge-toothed Pteriida families. Alatoconchidae are further diagnosed by their U-shaped isoclinal folds along the valve margins, forming characteristic “wings” that are absent in other Pteriida. In contrast, extant Pteriidae (pearl and winged oysters) possess well-developed hinge dentition, nacreous inner layers, and robust shells exceeding 5 mm thickness. Pectinidae exhibit strong radial ribs, equilateral outlines, and auricles near the hinge—features lacking in our specimens. Other extinct Pteriida families such as Ambonychiidae and Inoceramidae lack the pronounced lateral flanges and exhibit differing shell layering, often with thicker shells and pseudo-plicate ornamentation.
Compared with previously described Alatoconchidae species, such as Shikamaia sp., the specimens from the Duodao and Jingshan sections are notably smaller, with a maximum recorded length of ∼15 cm, whereas most reported alatoconchid specimens exceed 30 cm and may reach up to 1 m in length (Asato et al., 2017). However, similar-sized Alatoconchidae specimens have been described from other regions, including Japan (Isozaki, 2024) and South China (Chen et al., 2018). This suggests that the observed size variation may be influenced by preservation conditions, ontogenetic stages, or paleoenvironmental factors rather than taxonomic differences. Additionally, environmental factors such as water depth and substrate conditions may have played a role in constraining their growth.
The recovered specimens exhibit several morphological features consistent with the Alatoconchidae family. Notably, the shells are dorsoventrally compressed with broad, wing-like flanges, a characteristic trait of Alatoconchidae. The shell microstructure reveals a double-layered composition: an outer prismatic layer and an inner foliated layer, each approximately 3 mm thick. Despite the fragmented nature of the specimens, the preservation of key morphological traits, including the U-shaped lateral shell extensions and thin, layered structure, supports their identification as alatoconchids. While complete hinge structures are not preserved in our specimens, the observed features—such as the shell’s flattened morphology, the presence of wing-like flanges, and the specific shell microstructure—are consistent with those described in genera like Shikamaia and Saikraconcha. However, due to the absence of complete specimens, we refrain from assigning our material to a specific genus or species within Alatoconchidae.
4.2 Paleoecology and depositional environment
The alatoconchid bivalves discovered in the Duodao and Jingshan sections provide valuable insights into the paleoecology and depositional environment of the Middle Permian carbonate successions in central Hubei. Alatoconchids have been widely interpreted as epifaunal suspension feeders that may have engaged in symbiotic relationships to support their large body size (Isozaki and Ajinović, 2009; Asato et al., 2017; Chen et al., 2018). Their association with biostromes in shallow-marine carbonate platforms suggests they thrived in nutrient-rich, well-oxygenated waters with sufficient light penetration to sustain potential photosymbiosis (Isozaki, 2024).
As crinoids are stenohaline organisms with a high demand for oxygen and are typically absent from low-oxygen settings, their presence, therefore, indicates oxygen-rich conditions conducive to sustaining diverse benthic communities (Kammer and Ausich, 2006). The newly discovered alatoconchid fossils in central Hubei exhibit a strong association with crinoid fragments, suggesting a stable, carbonate-dominated ecosystem. The presence of fusulinids, rugose corals, and calcareous algae in the lower parts of the sections further supports the interpretation of a shallow-marine, reef-associated environment. The scarcity of other benthic organisms within the alatoconchid-bearing layers may indicate periodic environmental fluctuations, such as variations in nutrient availability or episodic sediment influx, which could have influenced community composition. The Alatoconchidae specimens from the Maokou Formation exhibit several taphonomic features indicative of rapid burial and low-energy depositional conditions. The shell fragments are generally well-preserved, with minimal signs of abrasion or bioerosion, suggesting limited transport and a relatively stable post-mortem environment. The presence of articulated shell pairs and the lack of significant fragmentation further support rapid burial events that protected the remains from extensive physical or biological disturbance. Additionally, the surrounding matrix is composed of fine-grained limestone, consistent with a low-energy marine setting conducive to the preservation of delicate shell structures.
Notably, the alatoconchid fossils from the Jingshan section were found within fallen rock samples, making it difficult to determine their exact stratigraphic context. However, their preservation and association with fusulinid index fossils indicate a Middle Permian age, consistent with previous reports from South China (Chen et al., 2018). The measurable fragments (3–5 cm in length and 1.0–5.2 mm in thickness) observed in these specimens, compared with larger alatoconchid fossils from other regions, may reflect differences in local environmental conditions or ontogenetic stages at the time of fossilization.
The Middle Permian strata in the Duodao and Jingshan sections are dominated by thick-bedded limestones interspersed with siliceous nodules, indicative of deposition in a shallow-marine, platform setting. The occurrence of alatoconchid biostromes within these limestones suggests they thrived in areas with relatively low sedimentation rates, allowing the development of stable carbonate buildups. The presence of chert nodules within the Maokou Formation may reflect periodic silica precipitation, potentially influenced by volcanic or hydrothermal activity (Shen et al., 2019; Zhang et al., 2023).
The depositional environment of the alatoconchid-bearing strata appears to have been influenced by regional tectonics and sea-level fluctuations. The South China Block, situated in the eastern Paleo-Tethys Ocean during the Permian, experienced a warm and humid climate following the waning of the Late Paleozoic Ice Age (Scotese, 2009; Henderson et al., 2020). This climatic transition likely contributed to the extensive development of carbonate platforms, providing a suitable habitat for alatoconchids and associated marine organisms.
In summary, the alatoconchid fossils from central Hubei represent an important addition to the known distribution of this group in South China. Their occurrence in shallow-marine carbonates, along with associated fossil assemblages, highlights the ecological significance of these bivalves and their role in Middle Permian marine ecosystems. Further analysis of their paleoecological adaptations and depositional context will contribute to a better understanding of the environmental conditions preceding the Capitanian mass extinction and the potential factors that influenced alatoconchid distribution and extinction.
4.3 Paleobiogeographic significance and implications for the Capitanian mass extinction
Alatoconchid bivalves were widely distributed during the Permian, primarily inhabiting low-latitude, tropical to subtropical regions along the Paleo-Tethys Ocean. Their occurrences have been documented across 12 regions worldwide, including East Asia (China, SW Japan, Malaysia, W. Philippines, Thailand, and Myanmar), the Middle East (Afghanistan, Oman, and Iran), southern Europe (Croatia), northern Africa (Tunisia), and northwestern North America (Alaska) (Isozaki, 2024). Most of these fossil localities are associated with shallow-marine shelf carbonates, reflecting the environmental preferences of alatoconchids for warm, stable carbonate platform settings. Notably, three occurrences (SW Japan, W. Philippines, and Alaska) have been identified in accreted paleo-atoll carbonates originally deposited in mid-oceanic domains of the Permian Panthalassa Ocean. This suggests that alatoconchids were not strictly confined to shallow continental shelves but could also inhabit isolated carbonate platforms within deeper marine settings.
The discovery of alatoconchid fossils in central Hubei significantly expands their known geographic range within South China. Their presence in the Maokou Formation, along with fusulinids, crinoids, and corals, suggests that the extensive carbonate platforms of the South China Block provided a favorable habitat for these organisms. The fossil-bearing strata, characterized by bioclastic limestones with Verbeekina sp. and crinoid remains, indicate deposition in a shallow-marine setting under warm climatic conditions with moderate to high energy levels. The association of alatoconchid fragments with crinoids and other benthic invertebrates aligns with previous reports from the Upper Yangtze and Dian-Qian-Gui regions, reinforcing their preference for carbonate platform environments during the Guadalupian (Chen et al., 2018).
The Capitanian mass extinction (∼260 Ma) was one of the most severe biotic crises of the Permian, significantly impacting marine ecosystems (Henderson et al., 2020). Recent studies suggest that widespread marine extinctions during this period were closely linked to major volcanic episodes associated with the Emeishan Large Igneous Province (ELIP) in South China (Wignall et al., 2009; Bond et al., 2010; Huang et al., 2025). These eruptions triggered global environmental changes, including ocean acidification, anoxia, and elevated sea surface temperatures, all of which likely contributed to the decline of carbonate-producing organisms.
As large suspension feeders dependent on stable marine conditions, alatoconchid bivalves may have been particularly vulnerable to these environmental disruptions. Their extinction coincides with the loss of other reef-associated taxa, including fusulinids, rugose corals, and calcareous algae, suggesting that deteriorating water quality and declining carbonate saturation played a role in their disappearance (Shen et al., 2019; Song et al., 2023; Huang et al., 2025). The abrupt disappearance of alatoconchid fossils near the Capitanian-Wuchiapingian boundary supports the hypothesis that they were among the victims of the Capitanian crisis (Isozaki and Ajinović, 2009; Chen et al., 2021).
The observed dominance of disarticulated shell fragments within the alatoconchid-bearing strata may indicate post-mortem transport or reworking, suggesting that these organisms experienced ecological stress prior to their extinction. Given previous interpretations of alatoconchids as potential photo- or chemo-symbiotic suspension feeders (Asato et al., 2017; Isozaki, 2024), further analysis of sedimentary facies and geochemical conditions could help clarify their ecological role and response to environmental perturbations.
The demise of alatoconchids reflects the broader instability of Middle Permian marine ecosystems under the influence of large-scale volcanic activity. The Emeishan volcanism not only caused widespread extinctions but also disrupted carbonate platform development, leading to regional shifts in sedimentary environments (Bond et al., 2010; Song et al., 2023; Huang et al., 2025). The loss of alatoconchids, along with other carbonate-dependent taxa, underscores the vulnerability of suspension-feeding organisms to volcanic-driven oceanic crises.
In summary, the newly discovered alatoconchid fossils from central Hubei provide critical paleobiogeographic evidence for the distribution of these bivalves within the Paleo-Tethys Ocean. Their extinction during the Capitanian crisis highlights the susceptibility of carbonate-dependent organisms to large-scale environmental changes and reinforces the role of volcanic-induced stress in shaping Permian marine ecosystems. Further research on alatoconchid paleoecology and extinction dynamics will enhance our understanding of the mechanisms driving biotic crises during the Permian.
5 Conclusion
The discovery of alatoconchid bivalves in the Middle Permian Maokou Formation of Jingmen City, Hubei Province, expands the known paleogeographic range of this taxon and underscores the ecological significance of South China as a key habitat during the Guadalupian. Their absence in younger strata reinforces the hypothesis that alatoconchids were among the marine organisms severely impacted by the Guadalupian-Lopingian boundary crisis. This finding not only contributes to a more comprehensive understanding of Permian marine biodiversity and extinction dynamics but also highlights the potential for further fossil discoveries in Hubei and adjacent regions. Future research integrating detailed stratigraphic, geochemical, and paleoenvironmental analyses will be essential to elucidate the ecological constraints and evolutionary history of alatoconchids in relation to broader Permian biotic events.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
LY: Writing – original draft, Writing – review and editing. KW: Writing – review and editing, Writing – original draft. WW: Writing – review and editing. BY: Writing – review and editing. DY: Writing – review and editing. SY: Writing – review and editing. HY: Writing – review and editing. YC: Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. Hubei Provincial Natural Science Foundation (2024AFD394; 2023AFD218)Science and Technology Special Fund of Hubei Geological Bureau(KJ2024-5, KJ2025-4).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that Gen AI was used in the creation of this manuscript. language polishing.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Asato, K., Kase, T., Ono, T., Sashida, K., and Agematssu, S. (2017). Morphology, systematics and paleoecology of Shikamaia, aberrant permian bivalves (Alatoconchidae: Ambonychioidea) from Japan. Paleontological Res. 21, 358–379. doi:10.2517/2017pr002
Bond, D. P. G., Hilton, J., Wignall, P. B., Ali, J. R., Stevens, L. G., Sun, Y. D., et al. (2010). The Middle Permian (Capitanian) mass extinction on land and in the oceans. Earth-Science Rev. 102, 100–116. doi:10.1016/j.earscirev.2010.07.004
Burgess, S. D., Bowring, S., and Shen, S. Z. (2014). High-precision timeline for Earth's most severe extinction. Proc. Natl. Acad. Sci. U. S. A. 111 (9), 3316–3321. doi:10.1073/pnas.1317692111
Chen, F. Y., Xue, W. Q., Yan, J. X., and Meng, Q. (2021). The implications of the giant bivalve family Alatoconchidae for the end-Guadalupian (Middle Permian) extinction event. Geol. J. 56, 6073–6087. doi:10.1002/gj.4151
Chen, F. Y., Xue, W. Q., Yan, J. X., Wignall, P. B., Meng, Q., Luo, J. X., et al. (2018). Alatoconchids: giant permian bivalves from South China. Earth-Science Rev. 179, 147–167. doi:10.1016/j.earscirev.2018.01.012
Dal Corso, J., Song, H. J., Callegaro, S., Chu, D. L., Sun, Y. D., Hilton, J., et al. (2022). Environmental crises at the Permian–Triassic mass extinction. Nat. Rev. Earth & Environ. 3 (3), 197–214. doi:10.1038/s43017-021-00259-4
Feng, Y., Song, H. J., and Bond, D. P. G. (2020). Size variations in foraminifers from the early permian to the late triassic: implications for the guadalupian–lopingian and the permian–triassic mass extinctions. Paleobiology 46, 511–532. doi:10.1017/pab.2020.37
Henderson, C. M., Shen, S. Z., Gradstein, F. M., and Agterberg, F. P. (2020). “The permian period,” in Geologic time scale 2020, 875–902.
Hou, Z. S., Fan, J. X., Henderson, C. M., Yuan, D. X., Shen, B. H., Wu, J., et al. (2020). Dynamic palaeogeographic reconstructions of the wuchiapingian stage (lopingian, late permian) for the South China Block. Palaeogeogr. Palaeoclimatol. Palaeoecol. 546, 109667. doi:10.1016/j.palaeo.2020.109667
Huang, H., Deng, C. Z., Grasby, S. E., Cawood, P. A., Hou, M. C., Yang, C. C., et al. (2025). Mercury evidence of Emeishan volcanism driving the mid-Capitanian (Middle Permian) extinction. Geol. Soc. Am. Bull. 137, 1177–1184. doi:10.1130/b37796.1
Isozaki, Y. (2024). Additional occurrence of capitanian (guadalupian, permian) gigantic bivalve Alatoconchidae from NE Japan and Primorye (far East Russia): paleobiogeographical implication to NE segment of greater South China. Bull. Natl. Mus. Nat. Sci. 50, 27–37. doi:10.50826/bnmnsgeopaleo.50.0_27
Isozaki, Y., and Ajinović, D. (2009). End-Guadalupian extinction of the Permian gigantic bivalve Alatoconchidae: end of gigantism in tropical seas by cooling. Palaeogeogr. Palaeoclimatol. Palaeoecol. 284, 11–21. doi:10.1016/j.palaeo.2009.08.022
Isozaki, Y., Aljinović, D., and Kawahata, H. (2011). The guadalupian (permian) kamura event in European tethys. Palaeogeogr. Palaeoclimatol. Palaeoecol. 308, 12–21. doi:10.1016/j.palaeo.2010.09.034
Jurikova, H., Garbelli, C., Whiteford, R., Reeves, T., Laker, G. M., Liebetrau, V., et al. (2025). Rapid rise in atmospheric CO2 marked the end of the late palaeozoic Ice age. Nat. Geosci. 18, 91–97. doi:10.1038/s41561-024-01610-2
Kammer, T. W., and Ausich, W. I. (2006). The “age of crinoids”: a mississippian biodiversity spike coincident with widespread carbonate ramps. Palaios 21, 238–248. doi:10.2110/palo.2004.p04-47
Linnaeus, C. (1758). Systema naturae, per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio decima, reformata. Tomus I. Laurentii Salvii, 823.
Miller, S. A. (1877). The American palaeozoic fossils; A catalogue of the genera and species, with names of authors, dates, places of publication, groups of rocks in which found, and the etymology and significant of the words, and an introduction devoted to the stratigraphical geology of the palaezoic rocks, xv + 333. Cincinnati: The author.
Scotese, C. R. (2009). “Late Proterozoic plate tectonics and paleogeography: a tale of two supercontinents, Rodinia and Pannotia,”. Global Neoproterozoic petroleum systems: the emerging potential in North Africa. Editors J. Craig, J. Thurow, A. Whitman, and Y. Abutarruma (Geological Society of London Special Publication), 326, 67–83.
Scotese, C. R. (2021). An atlas of phanerozoic paleogeographic maps: the seas come in and the seas go out. Annu. Rev. Earth Planet. Sci. 49, 679–728. doi:10.1146/annurev-earth-081320-064052
Shen, S. Z., Zhang, H., Zhang, Y., Yuan, D. X., Chen, B., He, W. H., et al. (2019). Permian integrative stratigraphy and timescale of China. Sci. China Earth Sci. 62, 154–188. doi:10.1007/s11430-017-9228-4
Song, H. Y., Algeo, T. J., Song, H. J., Tong, J. N., Wignall, P. B., Bond, D. P. G., et al. (2023). Global oceanic anoxia linked with the Capitanian (Middle Permian) marine mass extinction. Earth Planet. Sci. Lett. 610, 118128. doi:10.1016/j.epsl.2023.118128
Stanley, S. M. (2016). Estimates of the magnitudes of major marine mass extinctions in earth history. Proc. Natl. Acad. Sci. U. S. A. 113 (42), E6325-E6334–E6334. doi:10.1073/pnas.1613094113
Termier, H., Termier, G., and Lapparent, A. F. (1973). Grands bivalves récifaux du Permien supérieur de l`Afghanistan central. Ann. la Société Géologique Nord 93, 75–80.
Wignall, P. B., Védrine, S., Bond, D. P. G., Wang, W., Lai, X. L., Ali, J. R., et al. (2009). Facies analysis and sea-level change at the Guadalupian–Lopingian Global Stratotype (Laibin, South China), and its bearing on the end-Guadalupian mass extinction. J. Geol. Soc. 166 (4), 655–666. doi:10.1144/0016-76492008-118
Yancey, T. E., and Boyd, D. W. (1983). Revision of the Alatoconchidae: a remarkable family of Permian bivalve. Journal of. Paleontology 26, 497–520.
Keywords: Alatoconchidae, Middle Yangtze area, Guadalupian (Middle Permian), Hubei, Maokou Formation
Citation: Yang L, Wu K, Wang W, Yang B, Yang D, Yuan S, Yu H and Cui Y (2025) First record of alatoconchid bivalves from the Middle Permian Maokou Formation in central Hubei, South China. Front. Earth Sci. 13:1601733. doi: 10.3389/feart.2025.1601733
Received: 28 March 2025; Accepted: 29 April 2025;
Published: 15 May 2025.
Edited by:
Harry Dowsett, United States Department of the Interior, United StatesReviewed by:
Nursufiah Sulaiman, Universiti Malaysia Kelantan, MalaysiaXin Sun, China University of Geosciences Wuhan, China
Copyright © 2025 Yang, Wu, Wang, Yang, Yang, Yuan, Yu and Cui. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kui Wu, a3Vpd3VAY3VnLmVkdS5jbg==