Abstract
The Cretaceous Period, marked by global events such as volcanic activity, oceanic anoxic episodes, and the end-Cretaceous mass extinction, has been extensively studied in marine records. However, terrestrial Cretaceous systems remain understudied except in regions like northeastern China. Dinosaur eggs, abundant in Upper Cretaceous terrestrial strata, provide critical insights into paleoenvironments, climate, and biotic evolution. The Qinglongshan site in Yunyang District, Shiyan, Hubei Province, preserves thousands of semi-exposed, three-dimensionally intact dinosaur eggs with minimal deformation, offering a rare opportunity to study nesting behavior and environmental dynamics. Preliminary studies classified these eggs as Placoolithus tumiaolingensis (Dendroolithidae), but their chronostratigraphic context remains poorly constrained, hindering regional correlations. This study addresses this research gap by applying Laser Ablation Multi-Collector Inductively Coupled Plasma Mass Spectrometer (LA-MC-ICP-MS) U-Pb dating to biogenic calcite samples from egg - bearing horizons, and the test results indicate an depositional age (DA) of 85.91 ± 1.74 Ma. The results aim to establish a robust chronological framework for the Qinglongshan egg assemblage for the first time, enhancing understanding of Late Cretaceous terrestrial ecosystems in China’s interior and their response to global environmental changes. This study underscores the potential of dinosaur egg fossils as proxies for reconstructing the “Cretaceous World” in terrestrial settings.
1 Introduction
The Cretaceous period has garnered significant attention due to a series of major global events, including widespread volcanic activity, oceanic anoxic events, superchrons, biotic radiation, and mass extinctions (Jones and Jenkyns, 2001; Bralower et al., 1994; Cronin et al., 2001; Walliser, 1996; Leckie et al., 2002). However, most studies have focused on marine strata and fossil records from the Cretaceous. In contrast, research on terrestrial Cretaceous systems remains limited, except in regions such as northeastern China, where studies have achieved relatively higher resolution (Xi et al., 2019). In other areas, critical environmental-biological events in terrestrial Cretaceous settings are often difficult to define due to challenges in stratigraphic correlation or incomplete fossil records (Zhao et al., 2013).
Dinosaur eggs are a common fossil type in Upper Cretaceous terrestrial strata. They exhibit diverse microscopic shell structures and nesting patterns, and have been discovered in various sedimentary environments (Horner and Makela, 1979; Cousin et al., 1994; Mikhailov et al., 1994; Zhao, 2003; Sander et al., 2008). These fossils provide critical insights into regional stratigraphic correlation, paleoenvironmental reconstruction, and paleoclimate studies (Zhao et al., 2015). In recent years, research on dinosaur egg fossils, their burial chronologies, and associated environmental contexts has emerged as an important approach to understanding the terrestrial “Cretaceous World” (He et al., 2013; Zhao et al., 2017; Han et al., 2022).
Abundant Late Cretaceous dinosaur egg fossils were discovered in Qinglongshan (Qinglong mountain), Yunyang District, Shiyan, Hubei Province (Zhou, 1998). Through conservation efforts, thousands of these eggs are now semi-exposed in situ within their original stratigraphic layers. Most eggs exhibit minimal deformation, preserved as intact three-dimensional structures. Their unique arrangement and well-defined nesting patterns indicate limited post-depositional disturbance, while the continuous egg-bearing section reveals dozens of fossilized egg layers. A preliminary study by Zhang et al. (2018) classified the majority of Qinglongshan eggs as a single oospecies, Placoolithus tumiaolingensis, which belongs to the oofamily Dendroolithidae—a group characterized by highly porous egg-shells with branched eggshell units. However, the dinosaur species responsible for laying these eggs remains unidentified. Thus, Qinglongshan represents an understudied dinosaur nesting site, offering critical insights into dinosaur reproductive behavior and the Late Cretaceous climate, environmental dynamics, and biotic evolution in China’s interior regions.
Dinosaur egg dating generally relies on indirect methods, as fossilized eggs often lack sufficient radioactive isotopes for direct dating. Constraints are achieved by dating volcanic rocks or ash layers around the eggs using U-Pb or K-Ar/Ar-Ar methods to determine the age of zircon and feldspar. Relative dating methods like biostratigraphy are also used, comparing other fossils in the same rock layer with global stratigraphic profiles to estimate the eggs’ age. Paleomagnetic dating helps identify the stratum and constrain the eggs’ age (e.g., Li et al., 2010 used magnetostratigraphy to show that the Xixia Basin dinosaur eggs are no later than 83 Ma). However, these methods have indirect errors, such as volcanic eruptions occurring before or after the eggs, or surrounding minerals being affected by later geological processes.
Similarly, dinosaur bone dating faces similar challenges. But advances in carbonate U-Pb absolute dating have enabled in-situ dating of dinosaur bones. Qi et al. (2024) used LA-ICP-MS to U-Pb date early diagenetic calcite in Jurassic sauropod bone cavities from the Sichuan Basin, South China, obtaining an age of 165.3 ± 3.6/5.6 Ma, consistent with the maximum depositional age of 165.8 ± 1.0 Ma from detrital zircons in surrounding rocks, indicating rapid post-mortem diagenesis.
Nevertheless, the absence of reliable chronostratigraphic data for the egg-bearing horizons at Qinglongshan has significantly hindered regional stratigraphic correlation and sedimentological analysis. To address this gap, our study employs LA-MC-ICP-MS U-Pb dating on calcite samples recently identified within the dinosaur eggs. The results aim to establish a robust chronological framework and enhance the comprehensive understanding of the temporal and environmental context of this significant dinosaur egg assemblage.
2 Geological setting and sample
The east-west trending Qinling Orogenic Belt traversing central China has under-gone three major tectonic evolutionary stages since the Neoproterozoic: ocean-continent plate subduction orogeny, continent-continent plate subduction-collision orogeny, and intracontinental orogeny (Zhang et al., 2019). From the Late Jurassic to the Cretaceous, the Qinling Orogenic Belt experienced extensional deformation that propagated from its core to the margins. Localized extension along pre-existing NWW-trending faults formed a series of faulted basins such as Xixia, Xichuan, and Yunyang (Figure 1). These basins were filled with thick, predominantly coarse-clastic deposits comprising alluvial fan, fan-delta, and lacustrine facies sediments (Liu and Zhang, 2008).
FIGURE 1

(A) Spatial distribution of Cretaceous basins in the Qinling Orogen (modified after Chen and Santosh, 2014; Dong et al., 2021). (B) Simplified geological map of Yunyang area. (C) Simplified geological map of Qinglongshan area. (D) Stratigraphic column of Qinglongshan region, the Upper Cretaceous is divided into three groups (modified after Guan et al., 1997). The dinosaur egg samples were collected from the lower Gr.
The Qinglongshan area, located in the western part of the Yunyang Basin, is dominated by red terrigenous clastic continental deposits primarily composed of alluvial fan facies. These deposits unconformably overlie Precambrian metamorphic rocks. Guan et al. (1997) subdivided this red-bed sequence into three lithological units from bottom to top (Figure 1D). The lower unit is distinguished from the middle and upper units by its composition of breccias, breccia-bearing siltstones to fine sandstones, and abundant dinosaur egg fossils, the dinosaur eggs collected in this study were obtained from this stratum. Zhou et al. (1998) assigned the lower unit to the Late Cretaceous based on dinosaur egg fossil studies, correlating it with the Gaogou Formation in the Xichuan (Xixia) Basin. According to Guan et al. (1997) and Zhou et al. (1998), the Yunyang Qinglongshan Formation and the Gaogou Formation in the Xixia Basin are equivalent. Pan et al. (2007) identified the Xixia Basin as the Late Cretaceous Coniacian (89.8–86.3 Ma) through dinosaur egg fossils, small dinosaur bone fossils, and some fossil fragments of bivalves, gastropods, ostracods, charophytes, plant pollen, and plant fossils. However, the parataxonomic validity of reported oogenera such as Faveoloolithus, Dictyoolithus, and Prismatoolithus has been questioned (Zhao et al., 2015), and issues persist in the subdivision of Dendroolithidae (Zhang et al., 2018).
The carbonate samples in this study were collected from a calcite-filled dinosaur egg fossil preserved at Site No. 1 of the Tumiaoling Dinosaur Egg Fossil Locality in Qinglongshan. The sample consists of monomineralic calcite with well-developed crystals and coarse grain size. The sampled egg is embedded within breccia-bearing siltstone of the lower unit and forms part of a large clutch containing 28 eggs. The morphological characteristics of these eggs and the clutch align closely with Placoolithus tumiaolingensis as described by Zhang et al. (2018). The sampled oblate-spheroid egg shows consistent morphology with others in the clutch, displaying regular spacing without evidence of compressive deformation (Figure 2A). Two additional eggs in the clutch also exhibit calcite infilling. While the sampled egg suffered partial shell damage during collection, the other two calcite-filled eggs retain nearly intact external shells and contain no mixed sediments or shell fragments internally. In contrast, the remaining 26 eggs in the clutch show varying degrees of shell fragmentation, with some retaining only the lower approximately 1/2–3/4 of their shells. Their interiors are filled with breccia-bearing siltstone containing shell fragments, lithologically identical to the host rock (Figures 1A, 2B). These observations suggest that the calcite crystallization process occurred under conditions of intact eggshell preservation, likely postdating or contemporaneous with the fossilization of the dinosaur eggs. The sampling criteria were as follows: the outer surface of the dinosaur eggshell was highly intact, and inside the egg, there were almost no other mixed sediments or eggshell fragments except for calcite crystals, mainly based on purity and homogeneity. In addition, since the dinosaur egg preservation site has been built into a museum, we only collected two fragments from a broken dinosaur egg according to the above principles (Zhang et al., 2018).
FIGURE 2

Calcite dating samples from the eggs. (A) Sampled dinosaur egg fossil; (B) Egg clutch containing the sampled dinosaur egg fossil (red arrow indicates the egg fossil sampled in this study; blue arrows mark two additional calcite-filled egg fossils within the clutch).
The emended oogenus Placoolithus is characterized by the following unique combination of characters:oblate egg shape; an equatorial plane circular or sub-circular with long axis of 120–170 mm and short axis of 117–159 mm (Figure 2); eggshell thickness of 1.31–2.40 mm. Moreover, symmetric or asymmetric branches of eggshell unit usually appear in the middle part of the eggshell, and occasionally near the outer surface of the eggshell (Zhang et al., 2018). The eggshell units of the eggs usually contain two symmetrical branches. Most of the eggshell units branch in the middle part of the eggshell, while the others divide near the inner surface of the eggshell. Sometimes, one of the two branches subdivides into two smaller branches near the outer surface of eggshell (Figure 3).
3 Dating methods
LA-MC-ICP-MS carbonate U-Pb dating was performed to obtain the absolute age of the dinosaur eggs in the Isotope Laboratory of the Institute of Global Environmental Change, Xi’an Jiaotong University, using the method similar to that described in Wang et al. (2022), Wang et al. (2025), and Niu et al. (2025). The egg fossil samples were fixed in a 25 mm epoxy round target and polished to about 15,000 mesh. Note that the cross section of the eggshell should be exposed. Deionized water ultrasound was used to remove possible surface contamination on the target (Figure 4).
The samples and reference materials (RMs) are put into the S155 sample cell (Laurin Technics 155, aerosol dispersion volume <1 cm3), then laser denudation is performed using the 193 nm ArF LA system (RESOlution LR), and the generated aerosols go directly to the Multi-Collector Inductively Coupled Plasma Mass Spectrometer (MC-ICP-MS, Neptune XT) for measurement. Glass reference materials NIST616 and NIST614 are used to adjust the signal and determine the parameters (see Table 1 for details). Every 8 samples were interspersed with NIST614 glass and carbonate RM samples of known age (Nuriel et al., 2021; Hill et al., 2016; Wang et al., 2022; Wang et al., 2025), for instrument signal drift and matrix effect correction, respectively.
TABLE 1
| MC-ICP-MS (Neptune XT) | |
|---|---|
| RF power | 1,300 W |
| Cooling gas flow rate | 16 L/min |
| Auxiliary gas flow rate | 0.8 L/min |
| Argon make-up gas flow rate | 0.9 L/min |
| Nitrogen gas flow rate | 0.6 mL/min |
| Interface cones | Jet sample cone + X skimmer cone |
| Instrument resolution | Low, ∼400 |
| Analysis mode | Static |
| Detection system | 9 Faraday collectors +8 ICs |
| Collector configuration | IC1B(207Pb), IC2(206Pb), IC6(208Pb), IC7(232Th), IC8(235U) |
| Integration time | 0.131 s, one block of 15,000 cycles |
| Laser ablation system (RESOlution LR + S155) | |
|---|---|
| Laser type | ArF excimer laser |
| Wavelength | 193 nm |
| Pulse length | 20 ns |
| Energy density | 2 J/cm2 |
| Ablation mode | Single hole drilling for 25 s, 10 cleaning pulses |
| Spot diameter | 100 (120) μm for signal (cleaning) |
| Repetition rate | 10 Hz |
| Helium carrier gas flow rate | 0.45 L/min |
Operating parameters for U-Pb measurements by LA-MC-ICP-MS.
The Iolite software (Paton et al., 2011) is used for background deduction and downhole effect correction, as well as calculations of the isotope ratios, errors, correlation coefficients and other information required for the Tera-Wasserburg diagram (Tera and Wasserburg, 1972). Carbonate RM samples ASH15 (Nuriel et al., 2021) and SB19-2 (Wang et al., 2022; Wang et al., 2025) were used to determine the correction coefficient of U/Pb ratio and U content. U-Pb age results were calculated using IsoplotR (Vermeesch, 2018). The final reported error of age and (207Pb/206Pb)0 are 2SE, including the dispersion of sample points and the error of glass and carbonate RMs. To ensure the precision and reproducibility, standard ASH15 and SB19-2 was performed as unknowns in the course. And the ages we got (2.954 ± 0.022 Ma, 1.099 ± 0.009 Ma) are within uncertainty of the published results (2.965 ± 0.011 Ma and 1.099 ± 0.011 Ma). We adopted a test-while-selecting approach. We analyzed possible proper spots by checking Pb isotope ratios, especially 207Pb/206Pb in real-time during testing, and chose test points near areas with low 207Pb/206Pb ratios or similar features. Compared to U content, we deem the radiogenic Pb proportion more crucial.
4 Dating results
The microscopic characteristics of the eggshell and the white part are obviously different (Figure 4, Figure 5A). The white part is relatively pure calcite. The eggshell showed a mixture of secondary calcite and primary carbonate minerals, and its structure is similar to Zhang et al. (2018). It may be that the pores in the eggshell were subsequently filled with calcite. U-Pb LA-MC-ICP-MS tests were conducted on both the white and eggshell parts. The results (Figure 5B) showed that the white part contained a small amount of Pb, which basically did not contain U. The eggshell part had a certain amount of U (ca. 0.5 ug/g) and therefore detectable radiogenic Pb, among which the calcisfilled part of the eggshell contained little or no U, and the signal fluctuated greatly, which may be caused by the surrounding pollution.
Considering the U and Pb concentration, we only calculated the U-Pb age using points in the eggshell with relatively stable signals. The U-Pb dating results are shown in Figure 5C. The 2 eggshells got same ages within the error, finally we use all the data together to get the combined age of 85.91 ± 1.74 Ma (Figure 5C).
FIGURE 3

Photos of dinosaur eggshell under the microscope. The eggshell unit is slender and symmetrical (rarely asymmetrical), and is divided into two branches in the middle part of the eggshell. The compact layer accounts for 1/10–1/4 of the eggshell.
FIGURE 4

The scanned images (A) of the dinosaur egg samples after fixed into the epoxy resin (diameter: 25 mm), as well as the stitched images (B) from microscopic scanning.
FIGURE 5
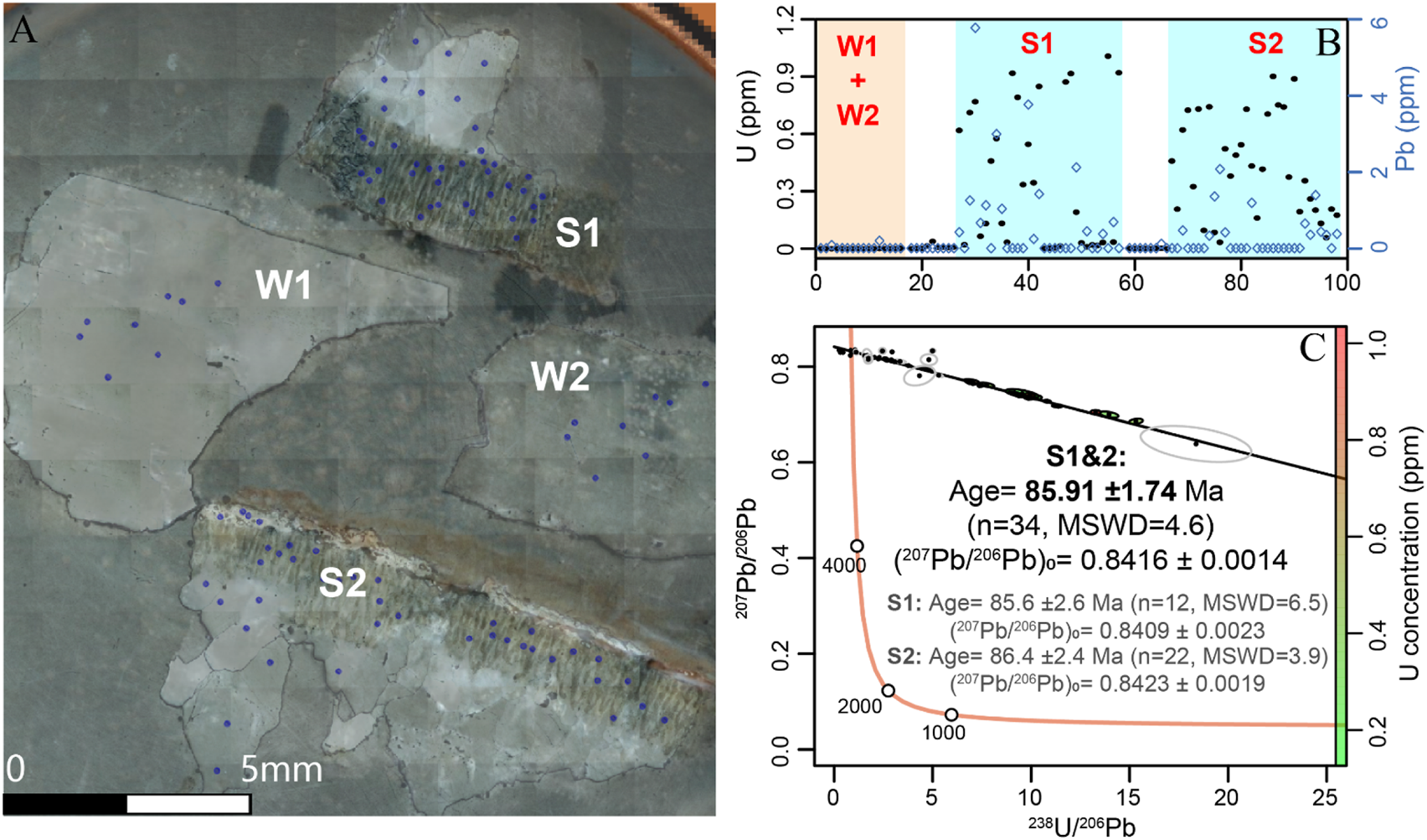
The dinosaur egg fossil image and the U-Pb results of the eggshells. (A) Microscopic image of fossil dinosaur eggs, S for eggshell while W means the white part. 1 and 2 come from different egg fossil fragments. The blue points show the laser ablation analysis positions; (B) U (black dots) and Pb (blue squares) concentration of the samples based on the LA results, the low value before the S1 and S2 are white parts near the shells. (C) The T-W diagram U-Pb age results of the eggshell parts using IsoplotR (Vermeesch, 2018). The right color bar shows the U concentration. Different part ages are showed in the T-W diagram. The hollow ellipse means the points are not used during the age calculation because of the signal instability or the low U content.
The best dating results were obtained from biogenic calcite, whereas abiogenic calcite yielded poor results. This might be due to the pore structures in abiogenic calcite, which enhance permeability and deviate from a semi-closed system (Figure 6). The following figure shows the laser spot positions for the S2 fragment sample. The orange square spots, which provide consistent ages, are located on the dark calcite visible under a microscope and are primarily biogenic calcite resulting from the biomineralization of amniotes. These correspond to the colored ellipses in the right-side S2 of Figure 5. The orange box spots correspond to the uncolored ellipses in the right-side S2 of Figure 5. The two-half - filled squares correspond to the two colorless ellipses close to the sides of the line in Figure 5.
FIGURE 6

The three spot positions for dinosaur egg (S2) carbonate LA-MC-ICP-MS are shown. Red X spots are on the calcite in the protein part, which cannot yield ages. Orange square spots, which give consistent ages, are on the dark calcite under a microscope and are mainly biogenic calcite from the biomineralization of amniotes. Orange box spots give poor ages and mainly show translucent, white calcite with pore structures. All other mineralization (from physicochemical processes without biotic precipitation) is “abiogenic calcite” (Zhang et al., 2025).
In this study, we only obtained a small number of samples, but achieved very good results. What we can confirm is that a large amount of data are distributed along the regression line (Figure 5), rather than being scattered on both sides, which proves that there is no significant loss of U. Moreover, we conducted repeat tests on two sets of samples and obtained consistent ages within the margin of error. This fully demonstrates that, at least on the dinosaur eggshells we collected, this age is very reliable and is highly consistent with the stratigraphic age.
5 Discussion
5.1 The concordance of U-Pb ages of Qinglongshan dinosaur eggshells with stratigraphy and the climatic background
Based on the evidence presented in this study, and after examining the microscopic and laser-ablation images of the Qinglongshan dinosaur eggshells, we are reasonably confident that the carbonate U–Pb ages obtained from the eggshell represent a depositional age (DA). As shown in Figure 6 of the paper, the vast majority of our laser spots were placed within biogenic calcite—the structural component of the eggshell. Microscopic inspection reveals no visible fractures, fissures, or other disturbances in these areas; any such features appear to have formed during or very shortly after burial, and the resulting ages are internally consistent. In contrast, the white, translucent, pore-filling abiogenic calcite is clearly a later diagenetic overgrowth, and the ages obtained from these domains deviate markedly from the regression line.
The stratigraphy containing dinosaur egg fossils in Qinglongshan has not yielded other paleontological fossils with chronostratigraphic significance, nor have isotopic dating studies of other minerals been conducted. The Yunyang Basin lacks volcanic ash for zircon U-Pb dating, and thus direct comparisons cannot be made. Therefore, the U-Pb age data obtained from calcite within the dinosaur eggs in this study provide the first chronological constraints with absolutely radioactive dating for the formation of these fossils in Qinglongshan. The results indicate that some dinosaur egg fossils in this area formed at 85.91 ± 1.74 Ma or earlier, corresponding to the late Coniacian or early Santonian stage of the Late Cretaceous. According to Guan et al. (1997) and Zhou et al. (1998), the Yunyang Qinglongshan Formation and the Gaogou Formation in the Xixia Basin are equivalent. The egg assemblage from the Taohe Basin in Xichuan (Xixia) is characterized by the presence of clutch-type eggs and some elongate eggs, with the dominant oogenera at this horizon being the highly porous Dendroolithus, the flattened Placoolithus and the alveolar Faveoloolithus. The assemblage from the Yunxian–Yunxi Basin is likewise marked by an abundance of Dendroolithus, accompanied by occasional spherical forms such as Paraspheroolithus. Although the nomenclature applied to these ootaxa still varies, it is evident that the composition and character of the egg faunas from both the Taohe Basin and the Yunxian–Yunxi Basin closely match those of the Zhaoying (Gaogou) Formation in the Xiping–Chimei Basin, all being distinguished by the predominance of Dendroolithus. Pan et al. (2007) primarily based on dinosaur egg fossils, small dinosaur bone fossils, and fossil fragments of bivalves, gastropods, ostracods, charophytes, plant pollen, and plant fossils, identified the Xixia Basin as the Late Cretaceous Coniacian (89.8–86.3 Ma). This stratigraphic classification has been widely accepted and is consistent with our test result of 85.91 ± 1.74 Ma (Figure 1).
The oofamily Dendroolithidae primarily includes two oogenera: Dendroolithus and Placoolithus. Fossils of this oofamily have been reported not only in Qinglongshan (Yunyang Basin) but also in Anlu, Xichuan, Xixia, and Tiantai (Zhao et al., 2015). Zhao and Li (1998) described Dendroolithus wangdianensis from the lower Gong’anzhai Formation in Anlu, establishing the oofamily Dendroolithidae. Based on ostracod and charophyte fossils, they assigned the host strata to the early Late Cretaceous, predating the Wangshi Group in the Jiaolai Basin and the Nanxiong Group in the Nanxiong Basin. Zhao and Zhao (1998) reported Placoolithus taohensis from the Majiacun Formation and Dendroolithus xichuanensis from the Gaogou Formation in the Xichuan Basin. Through comparative studies, they concluded that these egg-bearing strata were contemporaneous with the lower Gong’anzhai Formation in Anlu, also dating to the early Late Cretaceous. Fang et al. (1998) documented four dendroolithid oospecies in the Xixia Basin: Dendroolithus furcatus and Dendroolithus dendriticus from the Zoumagang Formation, and Dendroolithus sanlimiaoensis and Dendroolithus zhaoyingensis from the Zhaoying Formation. Notably, Dendroolithus zhaoyingensis was later synonymized with Dendroolithus dendriticus by Zhao et al. (2015). Subsequent studies on bivalve, gastropod, and ostracod fossils from the Dendroolithidae-bearing strata in the Xixia Basin consistently suggest a Coniacian–Campanian age (Fang et al., 1998; Chen et al., 2007; Pan et al., 2007; Cao and Wang, 2011), corresponding to the mid–late Late Cretaceous.
The burial age of dendroolithid egg fossils in Qinglongshan, determined in this study, falls within the mid-Late Cretaceous, aligning with the age range of Xixia Basin dendroolithid egg fossils, but slightly postdating their first appearances in Anlu and Xichuan. Paleogeographically, the Xichuan, Yunyang, and Xixia basins are closely clustered, with the Xichuan Basin sandwiched between the other two. This indicates that Dendroolithidae - producing dinosaur populations probably gathered in the Xichuan Basin during the early Late Cretaceous. Subsequently, during the mid-late Late Cretaceous, new distribution patterns of these populations in the Yunnan and Xixia basins to the south and north emerged.
Fang et al. (2000) and Fang et al. (2003) reported the discovery of Dendroolithus cf. dendriticus, Dendroolithus guoqingsiensis, and Dendroolithus shuangtangensis from the Chichengshan Formation in the Tiantai Basin, Zhejiang Province. However, Dendroolithus guoqingsiensis and Dendroolithus shuangtangensis were subsequently revised by Wang et al. (2013) as Parafaveoloolithus guoqingsiensis and Similifaveoloolithus shuangtangensis, respectively. He et al. (2013) through zircon U-Pb isotopic dating of tuffaceous interbeds in the red beds of the Tiantai Basin, constrained the depositional age of the Chichengshan Formation to between 91.2 and 94.4 Ma, approximately corresponding to the Turonian stage (early Late Cretaceous). This age range differs significantly from the burial age of the dendroolithid egg fossils recovered from Qinglongshan in this study and is also older than the strata bearing dendroolithid egg fossils in the Xixia Basin. Therefore, the presence of Dendroolithidae in the Tiantai Basin remains questionable. Further compar-ative studies are necessary to verify the ootaxonomic validity of Dendroolithus cf. dendriticus reported from the Tiantai Basin and Dendroolithus dendriticus from the Xixia Basin.
It is also worth noting that Zhou (1998) reported a member of the Spheroolithidae from the Qinglongshan area—Paraspheroolithus irenensis. This oospecies has not been reported from either the Tiantai or Xixia basins, yet it is among the most common taxa in the slightly younger Jiaolai Basin (Wang and Wang, 2024). This implies that the dinosaur egg-bearing strata at Qinglongshan may also include deposits from the late Late Cretaceous. Whether the Qinglongshan assemblage represents a transitional group between those of the Xixia and Jiaolai basins remains to be determined and will require further ootaxonomic studies and more precise stratigraphic and geochronological data.
Wang et al. (2012) preliminarily established a chronological sequence of dinosaur egg fossil assemblages from the Late Cretaceous of China, dividing them—from oldest to youngest—into four major assemblages represented by the Tiantai Basin, Xixia Basin, Jiaolai Basin, and Nanxiong Basin, respectively. Based on the above analysis, the Qinglongshan dinosaur egg assemblage may exhibit certain affinities with those of the Xixia and Tiantai basins. However, several oofamilies reported from both Tiantai and Xixia, including Macroelongatoolithidae, Prismatoolithidae, Dictyoolithidae, and Faveoloolithidae, have not been identified at Qinglongshan. Although the Qinglongshan site yields a large number of dinosaur eggs, the diversity at the oogenus and oospecies levels is relatively low, suggesting a highly specific paleoenvironment in the Yunyang Basin that may have favored reproduction by a limited range of dinosaur taxa.
When examining the Earth’s climate during the Late Cretaceous period, we may also uncover important clues. Comparison of global bottom-water temperatures and latitudinal thermal gradients suggests that global climate changed from a warm greenhouse state during the late Albian through late Cenomanian to a hot greenhouse phase during the latest Cenomanian through early Campanian, then to cool greenhouse conditions during the mid-Campanian through Maastrichtian (Huber et al., 1995; Huber et al., 2002). According to Linnert et al. (2014), the Late Cretaceous “greenhouse” climate underwent a transition from one of the warmest intervals of the past 140 million years to a cooler regime. Notably, a pronounced cooling event (from 35°C to 28°C) occurred around 84 million years ago (87–82 Ma), Notable variations have also been observed in the isotopic signatures (Linnert et al., 2014). In addition, Bornemann et al. (2008) provided evidence for glaciation during the mid-Cretaceous through stable isotope analysis, further supporting the notion of a progressive cooling trend during the Late Cretaceous. This transition from a warm to a cooler climate was likely to have had a significant impact on dinosaur diversity (Bornemann et al., 2008). The global temperature drop in the Late Cretaceous may have affected the number and oogenus distribution of dinosaur eggs in Qingshanshan. However, it is yet to be determined whether the size of the Yunyang Basin, a relatively small basin, decisively impacted the diversity of dinosaur egg fossils, as the lifestyle of dinosaurs themselves could influence such diversity.
5.2 The potential future application value of U-Pb ages of dinosaur eggshells
The U-Pb absolute dating of dinosaur egg fossils has a revolutionary significance for stratigraphy and basin research. Dinosaur egg fossils are widespread in China, particularly in the central and southern regions. There are many fault-subsidence basins rich in Late Cretaceous continental clastic rocks, such as the Xixia, Xichuan, Nanxiong, Jiaolai, Tiantai, Heyuan, Sanshui, and Ganzhou basins, which all contain abundant dinosaur egg fossils. Currently, only the Tiantai Basin has been dated to 98–91 Ma through volcanic ash - based dating of its sedimentary rocks. Other basins lack absolute age data. Thus, the Tiantai Basin’s dinosaur egg fossil assemblage, dominated by Macroelongatoolithidae, Faveoloolithidae, and Dictyoolithidae, is assigned to the Cenomanian—Turonian. In contrast, the Nanxiong Basin’s assemblage, represented by Elongatoolithidae, Ovaloolithidae, and Stalicoolithidae, belongs to the latest Late Cretaceous. Based on the Tiantai and Nanxiong basins, Wang et al. (2012) established a Late Cretaceous dinosaur egg fossil evolution sequence. This sequence allows for indirect stratigraphic age estimation of egg-bearing basins. For instance, the Xixia Basin’s fossil assemblage, including Macroelongatoolithidae, Faveoloolithidae, Dictyoolithidae, and Dendroolithidae, is equivalent to or slightly younger than the Tiantai Basin, dating to the early - mid Late Cretaceous (Turonian—Santonian). The Jiaolai Basin’s assemblage, comprising Spheroolithidae, Stalicoolithidae, Elongatoolithidae, Ovaloolithidae, and Dictyoolithidae, postdates the Xixia Basin but predates the Nanxiong Basin, corresponding to the mid - late Late Cretaceous (Santonian—Campanian). Nonetheless, conclusions from biostratigraphic correlation require absolute age dating for higher precision. Similarly, other dinosaur egg bearing basins need absolute age verification in addition to biostratigraphic dating. Overall, dinosaur egg fossil dating is broadly applicable for determining the age of Late Cretaceous continental basins in China and can significantly contribute to resolving their geological age-related issues.
The Nanxiong Basin features a complete stratigraphic section spanning the K - Pg boundary and is a key site for studying dinosaur extinction. However, the exact K - Pg boundary position remains debated. Traditionally, it was placed at the upper contact of the Pingling Formation, where dinosaur egg fossils disappear (Zhao et al., 1991). However, Zhao et al. (2017) proposed a slightly lower boundary based on spore and pollen fossil studies, suggesting some dinosaurs survived into the Paleogene. Absolute age dating of dinosaur egg fossils from this section could resolve this dispute and clarify whether dinosaurs went completely extinct by the Late Cretaceous.
Admittedly, this study still has certain deficiencies. For example, the collected dinosaur egg samples are somewhat homogeneous, and the number of laser spots on the eggshell fragments used for the two dating results could be increased to further minimize dating errors. However, given the excellent reproducibility of the two ages and the strong consistency of the data from biogenic calcite, with no evident diagenesis or uranium loss, the ages obtained via LA-MC-ICP-MS U-Pb dating on eggshell biogenic calcite align well with the relative stratigraphic ages. This offers a valuable reference for future U-Pb dating of dinosaur eggshell carbonates. The significant diagenesis issue identified in recent studies of dinosaur egg fossil from Gobi Desert, Mongolia using the carbon and oxygen isotopes (Graf et al., 2018) requires further investigation in Qinglongshan dinosaur eggshells, particularly in distinguishing between biogenic and abiogenic calcite. Traditional high-resolution sampling methods for carbon and oxygen isotopes from speleothems (Chen et al., 2022; Cheng et al., 2016) may be inadequate. Instead, laser-based techniques should be employed for carbon and oxygen isotope analysis of dinosaur eggshells.
In summary, this study presents the first reproducible absolute age obtained from U-Pb dating of dinosaur eggshell carbonates in China from Qinglongshan. Although some outstanding research groups around the world have already carried out preliminary studies and attempts on U-Pb dating of dinosaur eggshells, (Kim et al., 2024; Niespolo et al., 2024), which has made many useful explorations on the U-Pb element migration mechanism of dinosaur egg fossilization, the U uptake issue, and carbonate U-Pb dating. Our achievement still holds significant implications and expectations for research on dinosaur evolution and extinction, as well as environmental changes on Earth during the Late Cretaceous, which strongly promotes the absolute dating of dinosaur egg fossils.
6 Conclusion
Qinglongshan area of the Yunyang Basin in Hubei Province, central China, is rich in dinosaur egg fossils. In this study, we employed LA-MC-ICP-MS U-Pb dating on two parts of a single dinosaur eggshell and obtained reproducible age results. The data from the biogenic calcite indicate that the dinosaur eggs in the Yunyang Basin date back to 85.91 ± 1.74 Ma, placing them in the Late Cretaceous period. This era was marked by a significant global temperature decline. The application of U-Pb dating to dinosaur eggshells may represent a major advance in the study of dinosaur evolution.
Statements
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author contributions
QC: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review and editing. XC: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review and editing. JW: Data curation, Formal Analysis, Methodology, Software, Validation, Writing – review and editing. BZ: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review and editing. SZ: Funding acquisition, Investigation, Methodology, Supervision, Validation, Writing – review and editing. YN: Methodology, Validation, Writing – review and editing. GW: Formal Analysis, Investigation, Methodology, Validation, Writing – review and editing. KH: Formal Analysis, Investigation, Methodology, Resources, Writing – review and editing. WZ: Formal Analysis, Investigation, Methodology, Resources, Writing – review and editing. DY: Investigation, Writing – review and editing. JL: Investigation, Writing – review and editing. YZ: Investigation, Resources, Writing – review and editing. GC: Investigation, Resources, Writing – review and editing. ML: Formal Analysis, Investigation, Writing – review and editing. HC: Methodology, Resources, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by the Commonweal Geological Survey Project of Shaanxi Province (202507 to XC; 202310 to QC), the Natural Science Foundation of Hubei Province (2024AFD380 to BZ), National Natural Science Foundation of China (42272020 to SZ), Science and Technology Innovation Project of Laoshan Laboratory (LSKJ202203300), the Scientific Research Project of Hubei Geological Bureau (KJ2023-2,KJ2025-3 to BZ), Shaanxi Provincial Key Research and Development Plan General Project-Social Development Field (2024SF-YBXM-561 to QC), the Opening Fund of State Key Laboratory of Loess and Quaternary Geology (SKLLQG2209 to XC), the Shaanxi Science and Technology Association Youth Talent Lifting Program (20230538 to XC).
Acknowledgments
We express our gratitude to the editors and reviewers for their dedication and hard work.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/feart.2025.1638838/full#supplementary-material
References
1
Bornemann A. Norris R. D. Friedrich O. Beckmann B. Schouten S. Damsté J. S. S. et al (2008). Isotopic evidence for glaciation during the Cretaceous supergreenhouse. Science319 (5860), 189–192. 10.1126/science.1148777
2
Bralower T. J. Arthur M. A. Leckie R. M. Sliter W. V. Allard D. J. Schlanger S. O. (1994). Timing and paleoceanography of Oceanic dysoxia/anoxia in the Late Barremian to early Aptian (early Cretaceous). Palaios9, 335–369. 10.2307/3515055
3
Cao M. Z. Wang D. Y. (2011). Late Cretaceous non-marine Ostracoda from the Dinosaur egg-bearing strata of the Xixia Basin, Henan, China. Acta Micropalaeontologica Sin.28, 98–105. 10.3969/j.issn.0001-6616.2007.03.004
4
Chen J. H. Wang D. Y. Feng J. C. Fu G. H. Zhu S. G. (2007). Late Cretaceous Non-Marine Bivalves from the Dinosaur egg bearing Strata of Xixia Basin. Henan46, 299–313.
5
Chen Q. Cheng X. Cai Y. Luo Q. Zhang J. Tang L. et al (2022). Asian summer monsoon changes inferred from a stalagmite δ18O record in Central China during the last Glacial Period. Front. Earth Sci.10, 863829. 10.3389/feart.2022.863829
6
Chen Y. J. Santosh M. (2014). Triassic tectonics and mineral systems in the Qinling Orogen, central China. Geol. J.49 (4-5), 338–358. 10.1002/gj.2618
7
Cheng H. Edwards R. L. Sinha A. Spötl C. Yi L. Chen S. et al (2016). The Asian monsoon over the past 640,000 years and ice age terminations. nature534 (7609), 640–646. 10.1038/nature18591
8
Cousin R. Breton G. Fournier R. (1994). Dinosaur egg laying and nesting in France. Dinosaur Eggs and babies. New York: Cambridge University Press, 37–46.
9
Cronin M. Tauxe L. Constable C. Selkin P. Pick T. (2001). Noise in the quiet zone. Earth Planet. Sci. Lett.190, 13–30. 10.1016/s0012-821x(01)00354-5
10
Dong Y. Sun S. Santosh M. Zhao J. Sun J. He D. et al (2021). Central China orogenic belt and amalgamation of East Asian continents. Gondwana Res.100, 131–194. 10.1016/j.gr.2021.03.006
11
Fang X. S. Lu L. W. Cheng Z. W. Zou Y. P. Pang Q. Q. Wang Y. M. et al (1998). Cre-taceous egg fossils in Xixia, Henan. Beijing: Geology Press, 1–125.
12
Fang X. S. Lu L. W. Jiang Y. G. Yang L. F. (2003). Cretaceous fossil eggs from the Tiantai basin of Zhejiang, with a discussion on the extinction of dinosaurs. Geol. Bull.07, 512–520. 10.12097/gbc.20030798
13
Fang X. S. Wang Y. Z. Jiang Y. G. (2000). On the Late Cretaceous fossil eggs of Tiantai, Zhejiang. Geol. Rev.01, 105–112.
14
Graf J. Tabor N. J. Ferguson K. Winkler D. A. Lee Y. N. May S. et al (2018). Diagenesis of dinosaur eggshell from the Gobi Desert, Mongolia. Palaeogeogr. Palaeoclimatol. Palaeoecol.494, 65–74. 10.1016/j.palaeo.2017.11.011
15
Guan K. N. Zhou X. G. Ren Y. F. Xu S. Q. (1997). A preliminary study on the upper Cretaceous strata and dinosaur egg fossils in the Qinglongshan area, Yunxian County, Hubei Province. Earth Sci.6, 7–11.
16
Han F. Wang Q. Wang H. Zhu X. Zhou X. Wang Z. et al (2022). Low dinosaur biodiversity in central China 2 million years prior to the end-cretaceous mass extinction. Proc. Natl. Acad. Sci.119, e2211234119. 10.1073/pnas.2211234119
17
He H. Wang X. Wang Q. Jiang S. Cheng X. Zhang J. et al (2013). SIMS zircon U–Pb dating of the Late Cretaceous dinosaur egg-bearing red deposits in the Tiantai Basin, southeastern China. J. Asian Earth Sci.62, 654–661. 10.1016/j.jseaes.2012.11.015
18
Hill C. A. Polyak V. J. Asmerom Y. P. Provencio P. (2016). Constraints on a Late Cretaceous uplift, denudation, and incision of the Grand Canyon region, southwestern Colorado Plateau, USA, from U-Pb dating of lacustrine limestone. Tectonics35, 896–906. 10.1002/2016tc004166
19
Horner J. R. Makela R. (1979). Nest of juveniles provides evidence of family structure among dinosaurs. Nature282, 296–298. 10.1038/282296a0
20
Huber B. T. Hodell D. A. Hamilton C. P. (1995). Middle–Late Cretaceous climate of the southern high latitudes: stable isotopic evidence for minimal equator-to-pole thermal gradients. Geol. Soc. Am. Bull.107 (10), 1164–1191. 10.1130/0016-7606(1995)107<1164:mlccot>2.3.co;2
21
Huber B. T. Norris R. D. MacLeod K. G. (2002). Deep-sea paleotemperature record of extreme warmth during the Cretaceous. Geology30 (2), 123–126. 10.1130/0091-7613(2002)030<0123:dsproe>2.0.co;2
22
Jones C. E. Jenkyns H. C. (2001). Seawater strontium isotopes, oceanic anoxic events, and seafloor hydrothermal activity in the Ju-rassic and Cretaceous. Am. J. Sci.301, 112–149. 10.2475/ajs.301.2.112
23
Kim N. Niespolo E. M. Varricchio D. (2024). Testing dinosaur eggshells for U-Pb dating. AGU24.
24
Leckie R. M. Bralower T. J. Cashman R. (2002). Oceanic anoxic events and plankton evolution: biotic response to tectonic forcing during the mid‐cretaceous. Paleoceanography17, 13–29. 10.1029/2001pa000623
25
Li Z. Y. Huang B. C. Fang X. S. (2010). Magnetostratigraphic study and its geological significance of the dinosaur egg-bearing section in Xixia Basin, Central China. Chin. J. Geophys.53, 874–887.
26
Linnert C. Robinson S. A. Lees J. A. Bown P. R. Pérez-Rodríguez I. Petrizzo M. R. et al (2014). Evidence for global cooling in the Late Cretaceous. Nat. Commun.5 (1), 4194. 10.1038/ncomms5194
27
Liu S. F. Zhang G. W. (2008). Evolution and geodynamics of basin/mountain systems in East Qinling- Dabieshan and its adjacent regions, China. Geol. Bull. China27, 1943–1960. 10.12097/gbc.20081201
28
Mikhailov K. E. Sabath K. Kurzanov S. (1994). “Eggs and nests from the Cretaceous of Mongolia,” in Dinosaur eggs and babies. Editors CarpenterK.HirschK. F.HornerJ. R. (Cambridge: Cambridge University Press), 88–115.
29
Niespolo E. Kim N. Kracht O. Akhtar A. (2024). Trace element profiling in Cenozoic avian eggshells and implications for U-daughter dating. AGU24.
30
Niu X. Wang J. Kang L. Li Y. Zhang H. Dong X. et al (2025). Millennial-scale climate variability of the Asian summer monsoon over the last 690,000 years: insights from cave records. Sci. Bull.70, 1513–1522. 10.1016/j.scib.2025.02.011
31
Nuriel P. Wotzlaw J.-F. Ovtcharova M. Vaks A. Stremtan C. Šala M. et al (2021). The use of ASH-15 flowstone as a matrix-matched reference material for la-ser-ablation U-Pb geochronology of calcite. Geochronology3, 35–47. 10.5194/gchron-3-35-2021
32
Pan H. Z. Wang D. Y. Feng J. C. Fu G. H. Zhu S. G. (2007). Late Cretaceous non-marine Gastropoda from the Dinosaur egg-bearing strata of Xixia Basin, Henan. Acta Palaeontol. Sin.46, 293–298.
33
Paton C. Hellstrom J. Paul B. Woodhead J. Hergt J. (2011). Iolite: freeware for the visualisation and processing of mass spectrometric data. J. Anal. Atomic Spectrom.26, 2508. 10.1039/c1ja10172b
34
Qi L. Hou M. Mulder J. A. Cawood P. A. Guo Y. Wu S. et al (2024). In situ U-Pb dating of Jurassic dinosaur bones from Sichuan Basin, South China. Geology52, 216–221. 10.1130/g51872.1
35
Sander P. M. Peitz C. Jackson F. D. Chiappe L. M. (2008). Upper Cretaceous titanosaur nesting sites and their implications for sau-ropod dinosaur reproductive biology. Palaeontogr. A284, 69–107. 10.1127/pala/284/2008/69
36
Tera F. Wasserburg G. J. (1972). U-Th-Pb systematics in three Apollo 14 basalts and the problem of initial Pb in lunar rocks. Earth Planet. Sci. Lett.14, 281–304. 10.1016/0012-821x(72)90128-8
37
Vermeesch P. (2018). IsoplotR: a free and open toolbox for geochronology. Geosci. Front.9, 1479–1493. 10.1016/j.gsf.2018.04.001
38
Walliser O. H. (1996). Global events and event stratigraphy in the Phanerozoic.
39
Wang J. Niu X. Kang L. Cheng H. (2025). U-Th and U-Pb geochronology of Quaternary carbonates. Natl. Sci. Rev., nwaf078. 10.1093/nsr/nwaf078
40
Wang J. Niu X. W. Kang L. (2022). U-Pb dating of Quaternary speleothems using LA& ID MC-ICPMS. Quat. Sci.42, 1410–1419. 10.11928/j.issn.1001-7410.2022.05.14
41
Wang Q. Wang X. L. (2024). New progress on the Study of dinosaur eggs in Laiyang, Shandong. J. Jilin Univ.54, 38–45.
42
Wang Q. Zhao Z. K. Wang X. L. Zhang S. K. Jiang Y. G. (2013). New forms of dictyoolithids from the Tiantai Basin, Zhejiang Province of China and a parataxonomic revision of the dictyoolithids. Vertebr. Palasiat.51, 43–54.
43
Wang X. L. Wang Q. Jiang S. X. Cheng X. Zhang J. L. Zhao Z. K. et al (2012). Dinosaur egg faunas of the Upper Cretaceous terrestrial red beds of China and their stratigraphical significance. J. Stratigr.36, 400–416. 10.19839/j.cnki.dcxzz.2012.02.017
44
Xi D. Wan X. Li G. (2019). Cretaceous integrative stratigraphy and timescale of China. Sci. China Earth Sci.62, 256–286. 10.1007/s11430-017-9262-y
45
Zhang S. Choi S. Kim N. H. Xie J. Park Y. Plümper O. et al (2025). Biogenic origin of secondary eggshell units in dinosaur eggshells elucidates lost biomineralization process in maniraptoran dinosaurs. Sci. adv.11 (22), eadt1879. 10.1126/sciadv.adt1879
46
Zhang G. W. Guo A. L. Dong Y. P. Yao A. P. (2019). Rethinking of the Qinling orogen. J. Geomechanics25, 746–768. 10.12090/j.issn.1006-6616.2019.25.05.064
47
Zhang S. K. Yang Z. R. Li Z. Q. Hu Y. G. (2018). New dinosaur egg material from Yunxian, Hubei Province,China resolves the clas-sification of dendroolithid eggs. Acta Palaeontol. Pol.63, 671–678. 10.4202/app.00523.2018
48
Zhao Z. K. (2003). The nesting behavior of Troodontid Dinosaurs. Vertebr. Palasiat., 157–168.
49
Zhao H. Zhao Z. K. (1998). Dinosaur eggs from Xichuan Basin, Henan Province. Vertebr. Palasiatioa36, 282–296.
50
Zhao Z. K. Jie Y. E. Qiang W. (2017). Dinosaur extinction and subsequent mammalian recovery during the Cretaceous-Paleogene (K/Pg) transition in the Nanxiong Basin. Chin. Sci. Bull.62, 1869–1881. 10.1360/n972016-00886
51
Zhao Z. K. Li Z. C. (1998). A new structural type of the dinosaur eggs from Anlu County, Hubei Province. Vertebr. Palasiatioa26, 107–164.
52
Zhao Z. K. Wang Q. Zhang S. K. (2015). Vertebrata Paleontologia Sinica: volume two: amphibia, reptilia, aves: book seven: Di-nosaur eggs. Beijing: Science Press, 1–168.
53
Zhao Z. K. Ye J. Li H. M. Zhao Z. H. Yan Z. (1991). Extinction of the dinosaurs across the cretaceous-tertiary boundary in nanxiong Basin, Guangdong Province. Vertebr. Palasiat.29, 1–20.
54
Zhao Z. K. Zhang S. K. Wang Q. Wang X. (2013). Dinosaur diversity during the transition between the middle and late parts of the Late Cretaceous in eastern Shandong Province, China: evidence from dinosaur eggshells. Chin. Sci. Bull.58, 4663–4669. 10.1007/s11434-013-6059-9
55
Zhou X. (1998). Dinosaur eggs of the Late Cretaceous from the Qinglongshan region, Yunxian county, Hubei province. Hubei Geol. Min. Resour.12, 1.
56
Zhou X. G. Ren Y. F. Xu S. G. Guan K. N. (1998). Dinosaur eggs of the Late Cretaceous from the Qinglongshan Region, Yunxian County, Hubei Province. Hubei Geol. and Mineral Resour.12, 1–10.
Summary
Keywords
Yunyang dinosaur eggs, carbonate U-Pb dating, geological age, Late Cretaceous, LA-MC-ICP-MS
Citation
Chen Q, Cheng X, Wang J, Zhao B, Zhang S, Ning Y, Wang G, He K, Zhang W, Yu D, Li J, Zou Y, Chen G, Li M and Cheng H (2025) Geological age of the Yunyang dinosaur eggs revealed by in-situ carbonate U-Pb dating and its scientific implications. Front. Earth Sci. 13:1638838. doi: 10.3389/feart.2025.1638838
Received
31 May 2025
Accepted
16 July 2025
Published
11 September 2025
Volume
13 - 2025
Edited by
Peter David Roopnarine, California Academy of Sciences, United States
Reviewed by
Ryan T Tucker, Stellenbosch University, South Africa
Guangyan Zhou, China University of Geosciences Wuhan, China
Updates
Copyright
© 2025 Chen, Cheng, Wang, Zhao, Zhang, Ning, Wang, He, Zhang, Yu, Li, Zou, Chen, Li and Cheng.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bi Zhao, kamp_m@163.com
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.