 Alois Honek
Alois Honek Jiří Skuhrovec
Jiří Skuhrovec Zdenka Martinkova1
Zdenka Martinkova1 Peter Zach
Peter Zach- 1Crop Research Institute, Prague, Czechia
- 2Institute of Forest Ecology, Slovak Academy of Sciences, Zvolen, Slovakia
In 2016–2019, seasonal changes in the abundance of the harlequin ladybird Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae) were established in the city center of Prague, Central Europe, and in its outskirts. Adults were sampled from lime trees (Tilia spp.) at regular intervals throughout the growing season. The abundance of H. axyridis paralleled the course of abundance of its prey, the aphid Eucallipterus tiliae L., which peaks either early or late in the season. As a result, the seasonal dynamics of H. axyridis were unimodal, with a peak in the early (late June—early July of 2017 and 2019) or late (late July—mid-September of 2016 and 2018) period of the season. In the early period, there was a small (1–4 days) difference in the timing of the peak of H. axyridis between the city center and the outskirts. In the late period, the peak occurred significantly earlier (by 13–21 days) in the city center due to the warmer climate there than in the outskirts. The difference in the timing of the population peak between both locations disappeared after recalculating the calendar to thermal time (number of day degrees above 10.6°C thresholds elapsed from the end of H. axyridis hibernation). The warm mesoclimate of the city center advances the seasonal dynamics of H. axyridis, contributing to the success of this invasive species in urban habitats.
Introduction
In Central Europe, Harmonia axyridis (Pallas) (Coleoptera, Coccinellidae) is a dominant invasive ladybird, although it is only a recently established component of aphidophagous coccinellid communities (Roy et al., 2016). This species occurs in urban (Adriaens et al., 2008; Purse et al., 2015; Mukwevho et al., 2017), agricultural (Musser and Shelton, 2003; Lucas et al., 2007; Riddick, 2017), and forest areas (Vandereycken et al., 2012) on trees, herbs and crops populated by a variety of aphid species. This adaptability to diverse habitats is due to the plasticity of its seasonal cycle. Overwintering adults leave the hibernation sites in April. During the vegetative season, 2–3 incomplete generations of offspring may be produced, depending on temperature. In the autumn, adults of all generations enter dormancy and overwinter together (Honek et al., 2019b). Dispersion of adults into and using host stands for reproduction is opportunistic since adults always occupy stands with high prey abundance (Honek et al., 2019b). Due to its plastic life cycle and high adaptability to local conditions, H. axyridis is a suitable subject for studying the effects of urban and rural environments on the life cycle (Dudek et al., 2015; Sloggett, 2017; Honek et al., 2018b).
Among the characteristics that differentiate urban and rural areas is a difference in their mesoclimate, which is typically warmer in city centers than in the outskirts (Bonan, 2002). As temperature determines the rate of development of exotherms, we hypothesize that differences in mesoclimate affect the pace of life of H. axyridis and predict that (i) the peak abundance will occur earlier in the city center than in the outskirts and (ii) this difference will level off if population development is plotted against thermal time. This hypothesis was tested in 2016–2019, when populations of H. axyridis were sampled from Tilia sp. infested with aphids in the city center and on the outskirts of Prague.
Materials and Methods
Sampling
The abundance of H. axyridis was recorded in 2 areas situated at ∼10 km distance in the western part of Prague, Central Europe. The beetles were sampled at 8 sites on the outskirts (situated between 50.081–50.094 N and 14.263–14.336 E) and at 4 sites in the city center (50.072–50.075 N, 14.454–14.465 E). The tree stands at particular sites consisted of ≥ 5 lime trees (Tilia cordata Mill. or Tilia platyphyllos Scop.) infested with the aphid Eucallipterus tiliae (L.) (Sternorrhyncha: Aphididae). This aphid species has a single peak of abundance, the timing of which varies from year to year (Dixon and Barlow, 1979; Dahlsten et al., 1999).
The lime trees were sampled throughout the whole period when H. axyridis adults were present on the lime trees (May to October) at 10-day intervals. Beetles were swept from the canopy up to a height of approximately 3 m. Sampling was standardized using a standard sweep net (35 cm diameter, 140 cm handle) operated by J.S. (city center) and A.H. (outskirts). All the days on which samples were collected were sunny and calm. The sampling (80–200 sweeps at each site) was performed, and H axyridis abundance (number of adults caught per 100 sweeps) was established between 08.00 and 18.00 h. The abundance of aphids was classified according to an earlier elaborated scale (Honek et al., 2019b) into 4 degrees as follows based on the aphid counts: (1) no aphids, (2) < 0.002 aphids per leaf (aphids in swept material but no individuals found on 500 leaves), (3) ≥ 0.002 to 1.0 aphids per leaf, and (4) > 1.0 aphids per leaf. The difference in urbanization at the outskirts and city center areas was quantified as the percentage of the surrounding area (a circle with a 500-m radius centered at each sampling site) covered by impervious surfaces (buildings, streets, parking places, etc.) (Honek et al., 2018b). These values were 39 ± 3.0% in the outskirts and 71 ± 5.6% in the city center and were determined using maps available at https://en.mapy.cz.
Air temperature at 2 m above the ground was used to calculate the thermal time. The data for the outskirts were taken from the meteorological station of the Crop Research Institute (50.085N, 14.298E), and the data for the city center were taken from the meteorological station Klementinum (50.086N, 14.416E). The thermal time (expressed in day degrees dd) for H. axyridis was calculated as the sum of effective temperatures (°C) above the 10.6°C lower development threshold (Honek et al., 2018a) starting on 20 April, the date H. axyridis left the hibernation sites (Honek et al., 2018a). The thermal time (expressed in day degrees dd) for E. tiliae was calculated as the sum of effective temperatures (°C) above the 5.0°C lower development threshold starting from the beginning of the year (Dixon et al., 1997; Brabec et al., 2014).
Data Analysis
For each year and site, the Julian day was determined when the maximum abundance of H. axyridis (population peak) was observed. Each year, the average terms of maximum abundance were calculated for the city center and the outskirts as the arithmetic mean (± SE) of the terms of population peaks at particular sites. Differences were tested using t-tests or (if the normality test failed) Mann-Whitney tests. Calculations were performed using SigmaStat 3.5 (Systat Software Inc, 2006).
For each site and year the relative frequency of H. axyridis was calculated so that the maximum abundance was rated 1 and the abundance on other dates as a proportion of this maximum. This data plotted against time (Julian day) was fitted by the Asymmetric Double Sigmoidal (ADS) function. Calculations were performed using TableCurve 2D (Systat Software Inc, 2002).
Results
Each year, the course of change in the abundance of H. axyridis adults was unimodal. The term of peak abundance varied among years. In both areas, abundance peaked early in 2017 (22 June and 23 June) and 2019 (5 July and 9 July) and late in 2016 (29 August and 19 September) and 2018 (15 July and 28 July), with the peak occurring earlier every year in the city center than in the outskirts (Figure 1). The interval dividing the population peak in the city center and the population peak in the outskirts increased with the time that had elapsed since the beginning of the season (Figure 2). The difference was statistically significant in the years when abundance peaked late (2016 and 2018) (Table 1). The obvious difference on Julian days leveled off after the calendar time was transformed to thermal time (except 2017, when the population peak occurred very early). Differences in the timing of the population peak thus arose due to differences in the thermal conditions of the city center and the outskirts.
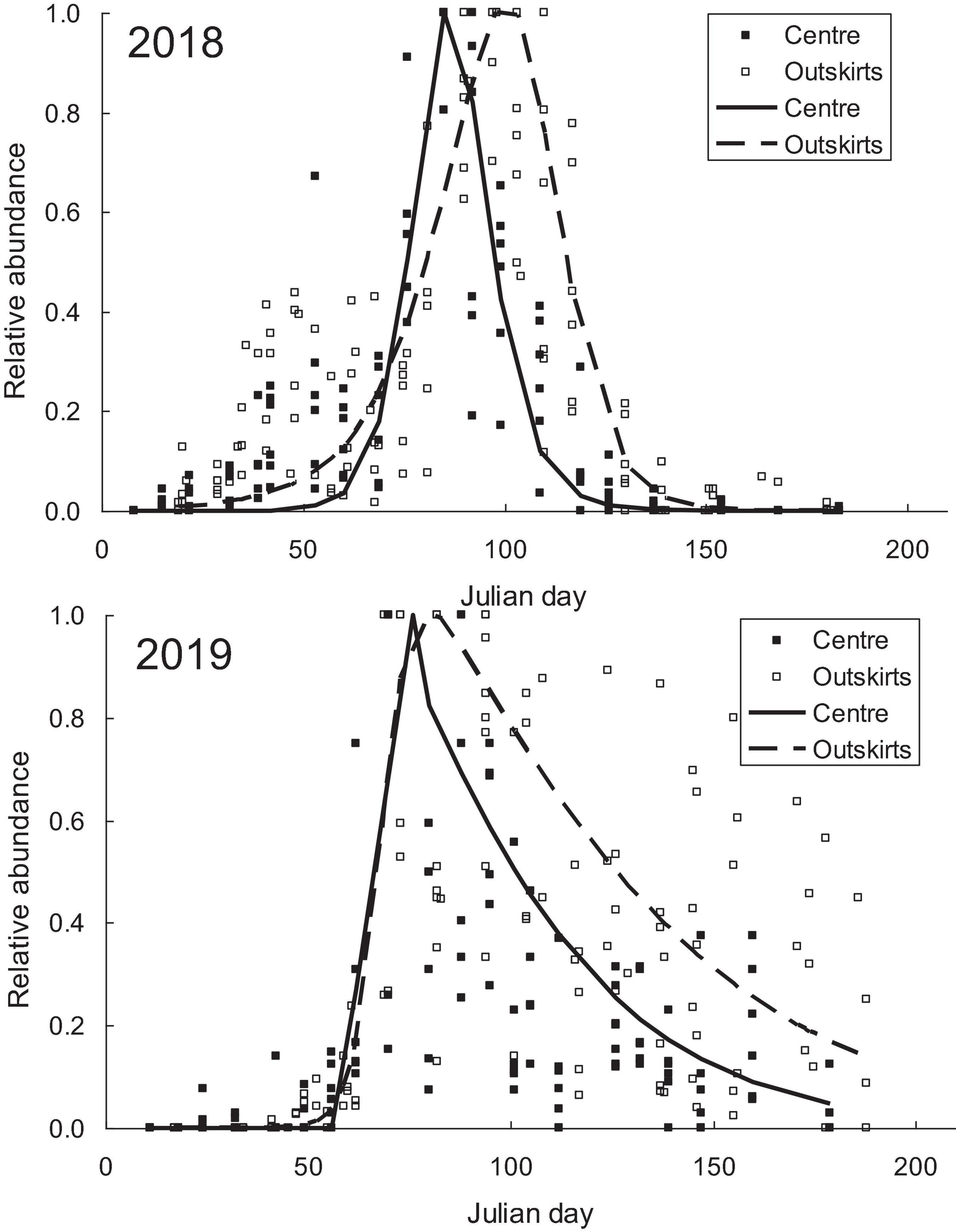
Figure 1. Changes in the relative abundance of Harmonia axyridis in the city center and the outskirts of Prague plotted against calendar time (Julian days from leaving their hibernacula on 20 April). The maximum abundance at each locality is scored 1. The data were fitted to the asymmetric double sigmoidal (ADS) equation. The figure shows example of a year when population peak occurred late in the season (2018) and of a year when population peak occurred early in the season (2019).
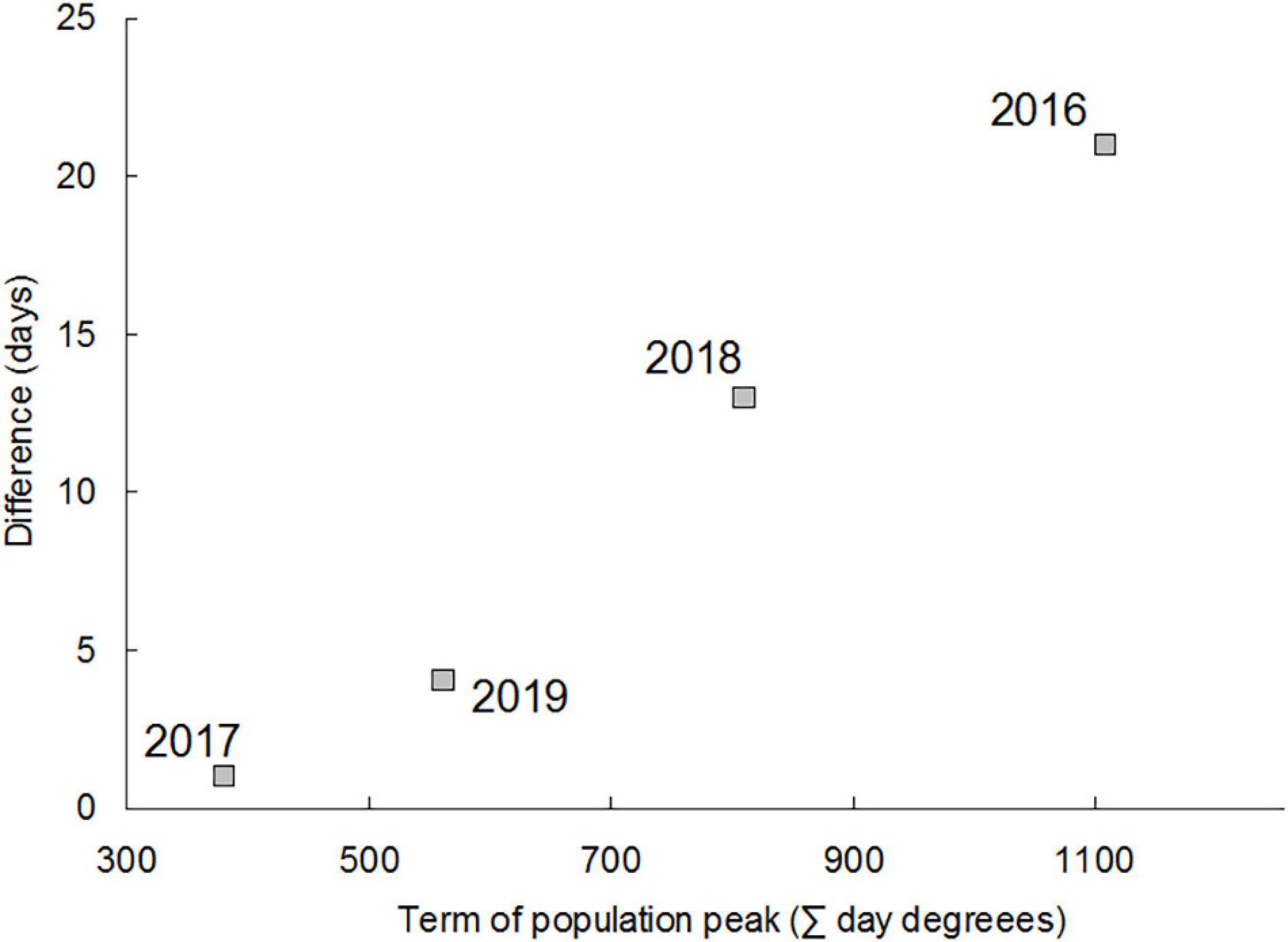
Figure 2. Difference (days) between the term of the population peak in the city center and the term of the population peak in the outskirts of Prague plotted against the thermal time (day degrees) when the maximum abundances were observed (average of the thermal time value in the city center and the outskirts).
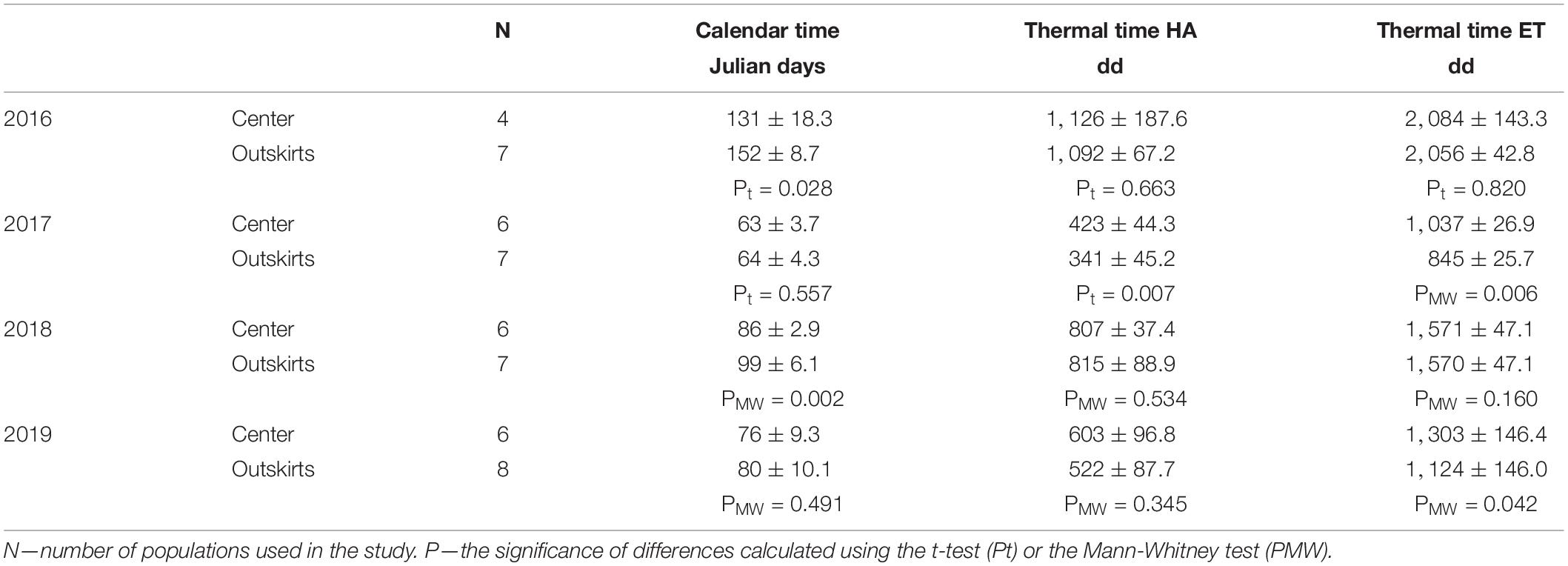
Table 1. Timing of the peak density of Harmonia axyridis in calendar time (Julian days since the release of hibernacula set at 20 April), thermal time calculated using thermal constants of H. axyridis as the number of day degrees (dd) above the 10.6°C threshold (Thermal time HA), and thermal time calculated using thermal constants for E. tiliae as the number of day degrees (dd) above the 5.0°C thermal threshold (Thermal time ET).
Discussion
Mesoclimate Factors That Affect Population Development
Divergence in the quality of urban (downtown area) and rural (outskirt area) environments is well established and is associated with a variety of environmental characteristics. Urban centers differ from the outskirts in terms of excessive light pollution, decreased humidity, modified schedules of precipitation and variations in wind direction and speed (Bonan, 2002). The differences increase with the size of the town and are appreciable in cities whose human population exceeds 1 million (Karl et al., 1988), as is the case for Prague. Urban conditions affect vegetation (Pyšek, 1998), its insect phytophagans and their predators (Bolger et al., 2000; McIntyre, 2000; Pickett et al., 2001), as well as their pathogens (Dudek et al., 2017a, b). Apart from light pollution, divergent ambient temperature is the most important difference between urban and rural environments. The warm temperature of urban areas compared to that of rural areas accelerates the rate of life processes of exotherm organisms. Differences in their duration are compensated when calendar time is converted to thermal time (Trudgill et al., 2005).
The agreement between the term of the population peak in the city center and the term of the population peak in the outskirts resulting from this transformation (Table 1) indicates that the timing of peaks depends on the sum of a definite amount of heat. Increased ambient temperature accelerates the life processes of H. axyridis and shifts the peak of its seasonal cycle of abundance if sufficient prey is available to support an acceleration. This is a necessary consequence of the influence of urban centers on populations of exothermic organisms.
The effect of ambient temperature could be modified by thermoregulation, which can shift body temperature against ambient temperature, albeit slightly. A decrease in body temperature can be achieved by searching for and hiding in cool places. Coccinellids living in the city center on trees surrounded by buildings and roads that are severely heated in the sunshine (Parlow, 2011) have little chance of decreasing their pace of life. Additionally, they have no reason to decrease their body temperatures because the ambient temperature remains mostly below the upper temperature threshold of 30°C (Honek et al., 2018a). The temperature maxima in the city center exceeded this limit by 14 times (2016) to 29 times (2019), in most cases (64%) by less than 2°C.
An increase in body temperature above ambient temperature is realized by basking in the sunshine (Sloggett and Honek, 2012; Sloggett, 2021). The capacity to increase body temperature also exists in H. axyridis (Michie et al., 2010), but evidence of thermoregulation under natural conditions is indirect. This evidence consists of the geographic variation of the ratio of dark and light color forms that changes in parallel with climate (Dobzhansky, 1924; Komai, 1956). The mechanisms affecting morph distribution include differences in thermoregulation, assortative mating and winter mortality of color morphs (Osawa and Nishida, 1992; Su et al., 2009). This geographic variability is important in some parts of the native area of H. axyridis; however, in recently colonized regions of Europe, this phenomenon cannot be reliably demonstrated, as a significant geographic or temporal variation in proportional representation of dark and light forms was not established (Honek et al., 2020a), and thus, the effect of thermoregulation on the life cycle of H. axyridis cannot be reliably demonstrated. Thus thermoregulation likely does not significantly alter the effect of ambient temperature on the population dynamics of H. axyridis in the city center and its outskirts.
Ultimate Cause of Variation in the Timing of the Seasonal Peak of Abundance
In Tilia, the change in abundance of adult H. axyridis parallels that of the abundance of its prey, the monophagous anholocyclic aphid Eucallipterus tiliae (Honek et al., 2019a). Aphid abundance estimated in a period of 10–20 days around the peak of H. axyridis abundance (Alois Honek and Jiri Skuhrovec, unpublished) exceeded the threshold density of 1 aphid per leaf area necessary for the start of H. axyridis oviposition (Honek et al., 2020b). The dependence of H. axyridis on its prey determines the early (2017 and 2019) or late (2016 and 2018) timing of the population peaks in particular years. The annual variation in the timing of the peak abundance of E. tiliae is well known (Dixon and Barlow, 1979; Dahlsten et al., 1999), likely determined by the size of the overwintering population. The population peak occurs early in the vegetative season if hibernating eggs are abundant; if not, the population peak occurs late in the season (Dixon and Barlow, 1979). As a consequence, the population peaks of E. tiliae in the city center and the outskirts in particular years occurred approximately in the same period of the season. The ultimate cause of the timing of the H. axyridis population peak is the population dynamics of its prey, E. tiliae, which does not depend on the thermal conditions of the season.
Proximate Cause of the Difference in Timing of the Population Peaks in the City Center and the Outskirts
The magnitude of the difference between the H. axyridis population peaks in the city center and the outskirts increases with the time that has elapsed since the beginning of the season. The difference in the timing of the population peak levels after the recalculation of the calendar time to thermal time, which passes faster in warm city centers than in colder outskirts. The question remains whether the main factor of the origin of this difference is the temperature effect on the development of the aphid population or, alternatively, is the main factor directly the effect on the predator population? Using our data, this question cannot be reliably resolved, but the assumption of a direct effect on H. axyridis populations appears more probable. The effect on H. axyridis is significant because temperatures are below the maximum temperature for development, which is approximately 30°C (Auad et al., 2014; Barahona-Segovia et al., 2016) and in this range ambient temperature affects the rate of development of H. axyridis populations positively. Temperature also affects the rate of development of aphid populations, but in a different way than in H. axyridis. This is because the development threshold for aphid populations is 5°C (Honek and Kocourek, 1990; Honek, 1996), and temperature maximum is then ≤ 25°C (Dixon et al., 2009). The temperature in summer often exceed the temperature maxima for aphids, and in this range, does not influence the rate of development of E. tiliae positively. It can therefore be assumed that the main reason for the difference in population peak timing is the effect of temperature on populations of H. axyridis and not the effect of temperature on the population development of prey.
Significance of the Flexibility of Timing of the H. axyridis Life Cycle
This 4-year study enabled us to determine the causes of annual and local variability in the timing of the population peak of adult H. axyridis. In particular years, the peak occurred early or late in the season depending on variations in prey abundance. The results corroborated the hypothesis that differences in mesoclimate affect the pace of life of H. axyridis and supported the predictions (i) that the increased temperature of the city center, compared to that in the outskirts, advanced the onset of the population peak and (ii) that this difference will level off if population development is plotted against thermal time. Minor differences in the timing of the population peak were due to differences in the mesoclimate in the Prague city center and the outskirts, and the magnitude of the difference increased with the length of time that had elapsed since the beginning of the season. The flexibility of the timing of population development contributes to the success of H. axyridis invasion into the non-native area of Central Europe because it enables the effective utilization of prey resources whose location varies in space and time.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Author Contributions
AH and JS designed the experiments and collected data. ZM coordinate all work. AH and PZ analyzed the data and performed the statistical analyses. All authors wrote and edited the manuscript.
Funding
The work was supported by the program VES19 INTER-COST No. MSMT-15739/2019-6 (MŠMT ČR), Grant No. QK 1910281 from the National Agency for Agricultural Research (NAZV) of the Czech Republic and the grant VEGA 2/0032/19 of the Scientific Grant Agency of Ministry of Education, Science, Research and Sport of the Slovak Republic.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Ms Helena Uhlířová, Jana Kohoutová, and Hana Smutná for their excellent technical assistance.
References
Adriaens, T., San Martin y Gomez, G., and Maes, D. (2008). Invasion history, habitat preferences and phenology of the invasive ladybird Harmonia axyridis in Belgium. Biocontrol 53, 69–88. doi: 10.1007/s10526-007-9137-6
Auad, A. M., Santos, J. C., and Fonseca, M. G. (2014). Effects of temperature on development and survival of Harmonia axyridis (Coleoptera: Coccinellidae) and Chrysoperla externa (Neuroptera: Chrysopidae) fed on Rhopalosiphum padi (Hemiptera: Aphididae). Fla. Entomol. 97, 1353–1363. doi: 10.1653/024.097.0409
Barahona-Segovia, R. M., Grez, A. A., and Bozinovic, F. (2016). Testing the hypothesis of greater eurythermality in invasive than in native ladybird species: from physiological performance to life-history strategies. Ecol. Entomol. 41, 182–191. doi: 10.1111/een.12287
Bolger, D. T., Suarez, A. V., Crooks, K. R., Morrison, S. A., and Case, T. J. (2000). Arthropods in urban habitat fragments in southern California: area, age, and edge effects. Ecol. Appl. 10, 1230–1248. doi: 10.1890/1051-0761(2000)010[1230:AIUHFI]2.0.CO;2
Bonan, G. (2002). Ecological Climatology: Concept and Application. Cambridge: Cambridge University Press.
Brabec, M., Honěk, A., Pekár, S., and Martinkova, Z. (2014). Population dynamics of aphids on cereals: digging in the time-series data to reveal population regulation caused by temperature. PLoS One 9:e106228. doi: 10.1371/journal.pone.0106228
Dahlsten, D. L., Zuparko, R. L., Hajek, A. E., Rowney, D. L., and Dreistadt, S. H. (1999). Long-term sampling of Eucallipterus tiliae (Homoptera: Drepanosiphidae) and associated natural enemies in a northern California site. Environ. Entomol. 28, 845–850. doi: 10.1093/ee/28.5.845
Dixon, A. F. G., and Barlow, N. D. (1979). Population regulation in the lime aphid. Zool. J. Linnean Soc. 67, 225–237. doi: 10.1111/j.1096-3642.1979.tb01113.x
Dixon, A. F. G., Hemptinne, J. L., and Kindlmann, P. (1997). Effectiveness of ladybirds as biological control agents: patterns and processes. Entomophaga 42, 71–83. doi: 10.1007/BF02769882
Dixon, A. F. G., Honek, A., Keil, P., Kotela, M. A. A., Sizling, A. S., and Jarosik, V. (2009). Relationship between the maximum and minimum thresholds for development in insects. Funct. Ecol. 23, 257–264. doi: 10.1111/j.1365-2435.2008.01489.x
Dobzhansky, T. (1924). Die geographische und individuelle Variabilität von Harmonia axyridis Pall. in ihren Wechselbeziehungen. Biol. Zentralbl. 44, 401–421.
Dudek, K., Dudek, M., and Tryjanowski, P. (2015). Wind turbines as overwintering sites attractive to an invasive lady beetle, Harmonia axyridis Pallas (Coleoptera: Coccinellidae). Coleopts Bull. 69, 665–669. doi: 10.1649/0010-065X-69.4.665
Dudek, K., Huminska, K., Wojciechowicz, J., and Tryjanowski, P. (2017a). Metagenomic survey of bacteria associated with the invasive ladybird Harmonia axyridis (Coleoptera: Coccinellidae). Eur. J. Entomol. 114, 312–316. doi: 10.14411/eje.2017.038
Dudek, K., Sienkiewicz, P., Gwiazdowicz, D. J., and Tryjanowski, P. (2017b). A “clean” alien species? Parasites of the invasive ladybird Harmonia axyridis (Coleoptera: Coccinellidae). Eur. J. Entomol. 114, 350–354. doi: 10.14411/eje.2017.044
Honek, A. (1996). The relationship between thermal constants for insect development: a verification. Acta Soc. Zool. Bohem. 60, 115–152.
Honek, A., and Kocourek, F. (1990). Temperature and development time in insects: a general relationship between thermal constants. Zool. Jahrb. Abt. Syst. Geog. Biol. Tiere 117, 401–439.
Honek, A., Brabec, M., Martinkova, Z., Dixon, A. F. G., Pekár, S., and Skuhrovec, J. (2019a). Factors determining local and seasonal variation in abundance of Harmonia axyridis (Coleoptera: Coccinellidae) in Central Europe. Eur. J. Entomol. 116, 93–103. doi: 10.14411/eje.2019.011
Honek, A., Brown, P. M. J., Martinkova, Z., Skuhrovec, J., Brabec, M., Burgio, G., et al. (2020a). Factors determining variation in colour morph frequencies in invasive Harmonia axyridis populations. Biol. Invasions 22, 2049–2062. doi: 10.1007/s10530-020-02238-0
Honek, A., Martinkova, Z., and Pekar, S. (2020b). Threshold aphid population density for starting oviposition in Harmonia axyridis. Biocontrol 65, 425–432. doi: 10.1007/s10526-020-10019-w
Honek, A., Martinkova, Z., Dixon, A. F. G., Skuhrovec, J., Roy, H. E., Brabec, M., et al. (2018a). Life cycle of Harmonia axyridis (Coleoptera: Coccinellidae) in central Europe. Biocontrol 63, 241–252. doi: 10.1007/s10526-017-9864-2
Honek, A., Martinkova, Z., and Strobach, J. (2018b). Effect of aphid abundance and urbanization on the abundance of Harmonia axyridis (Coleoptera: Coccinellidae). Eur. J. Entomol. 115, 703–707. doi: 10.14411/eje.2018.069
Honek, A., Martinkova, Z., Roy, H. E., Dixon, A. F. G., Skuhrovec, J., Pekar, S., et al. (2019b). Differences in the phenology of Harmonia axyridis and native coccinellids on plants in central Europe (Coleoptera: Coccinellidae). Environ. Entomol. 48, 80–87. doi: 10.1093/ee/nvy173
Karl, T. R., Diaz, H. F., and Kukla, G. (1988). Urbanization: its detection and effect in the United States climate record. J. Clim. 1, 1099–1123. doi: 10.1175/1520-0442(1988)001<1099:UIDAEI>2.0.CO;2
Lucas, E., Vincent, C., Labrie, G., Chouinard, G., Fournier, F., Pelletier, F., et al. (2007). The multicolored asian lady beetle Harmonia axyridis (Coleoptera: Coccinellidae) in Quebec agroecosystems ten years after its arrival. Eur. J. Entomol. 104, 737–743. doi: 10.14411/eje.2007.094
McIntyre, N. E. (2000). Ecology of urban arthropods: a review and a call to action. Ann. Entomol. Soc. Am. 93, 825–835. doi: 10.1603/0013-8746(2000)093[0825:EOUAAR]2.0.CO;2
Michie, L. J., Mallard, F., Majerus, M. E. N., and Jiggins, F. M. (2010). Melanic through nature or nurture: genetic polymorphism and phenotypic plasticity in Harmonia axyridis. J. Evol. Biol. 23, 1699–1707. doi: 10.1111/j.1420-9101.2010.02043.x
Mukwevho, V. O., Pryke, J. S., and Roets, F. (2017). Habitat preferences of the invasive harlequin ladybeetle Harmonia axyridis (Coleoptera: Coccinellidae) in the western Cape Province, South Africa. Afr. Entomol. 25, 86–97. doi: 10.4001/003.025.0086
Musser, F. R., and Shelton, A. F. (2003). Factors altering the temporal and within-plant distribution of coccinellids in corn and their impact on potential intra-guild predation. Environ. Entomol. 32, 575–583. doi: 10.1603/0046-225X-32.3.575
Osawa, N., and Nishida, T. (1992). Seasonal variation in elytral colour polymorphism in Harmonia axyridis (the ladybird beetle): the role of non-random mating. Heredity 69, 297–307. doi: 10.1038/hdy.1992.129
Parlow, E. (2011). “Urban climate,” in Urban Ecology, ed. J. Niemelä (Oxford: Oxford University Press), 31–44. doi: 10.1093/acprof:oso/9780199563562.003.0005
Pickett, S. T. A., Cadenasso, M. L., Grove, J. M., Nilon, C. H., Pouyat, R. V., Zipperer, W. C., et al. (2001). Urban ecological systems: linking terrestrial ecological, physical, and socioeconomic components of metropolitan areas. Annu. Rev. Ecol. Syst. 32, 127–157. doi: 10.1146/annurev.ecolsys.32.081501.114012
Purse, B. V., Comont, R., Butler, A., Brown, P. M. J., Kessel, C., and Roy, H. E. (2015). Landscape and climate determine patterns of spread for all colour morphs of the alien ladybird Harmonia axyridis. J. Biogeogr. 42, 575–588. doi: 10.1111/jbi.12423
Pyšek, P. (1998). Alien and native species in Central European urban floras: a quantitative comparison. J. Biogeogr. 25, 155–163. doi: 10.1046/j.1365-2699.1998.251177.x
Riddick, E. W. (2017). Spotlight on the positive effects of the ladybird Harmonia axyridis on agriculture. Biocontrol 62, 319–330. doi: 10.1007/s10526-016-9758-8
Roy, H. E., Brown, P. M. J., Adriaens, T., Berkvens, N., Borges, I., Clusella Trullas, S., et al. (2016). The harlequin ladybird, Harmonia axyridis: global perspectives on invasion history and ecology. Biol. Invasions 18, 997–1044. doi: 10.1007/s10530-016-1077-6
Sloggett, J. J. (2017). Harmonia axyridis (Coleoptera: Coccinellidae): smelling the rat in native ladybird declines. Eur. J. Entomol. 114, 455–461. doi: 10.14411/eje.2017.058
Sloggett, J. J. (2021). Aphidophagous ladybirds (Coleoptera: Coccinellidae) and climate change: a review. Insect Conserv. Diver. doi: 10.1111/icad.12527
Sloggett, J. J., and Honek, A. (2012). “Genetic studies,” in Ecology and Behavior of the Ladybird Beetles, eds I. Hodek, H. F. van Emden, and A. Honěk (Hoboken, NJ: Blackwell Publishing), 13–53. doi: 10.1002/9781118223208.ch2
Su, W., Michaud, J. P., Zhi, Z., Fan, Z., and Shuang, L. (2009). Seasonal cycles of assortative mating and reproductive behaviour in polymorphic populations of Harmonia axyridis in China. Ecol. Entomol. 34, 483–494. doi: 10.1111/j.1365-2311.2008.01075.x
Systat Software Inc (2002). TableCurve 2D 5.01 for Windows User’s Manual. Point Richmond, CA: SYSTAT.
Trudgill, D. L., Honek, A., Li, D., and VanStraalen, N. M. (2005). Thermal time – concepts and utility. Ann. Appl. Biol. 146, 1–14. doi: 10.1111/j.1744-7348.2005.04088.x
Keywords: urban habitats, invasive, ladybirds, prey, temperature, mesoclimate, seasonal dynamics
Citation: Honek A, Skuhrovec J, Martinkova Z, Kulfan J, Jauschova T and Zach P (2021) Warm Mesoclimate Advances the Seasonal Dynamics of Harmonia axyridis in Urban Habitats. Front. Ecol. Evol. 9:725397. doi: 10.3389/fevo.2021.725397
Received: 15 June 2021; Accepted: 07 September 2021;
Published: 30 September 2021.
Edited by:
Danny Haelewaters, Ghent University, BelgiumReviewed by:
Piotr Tryjanowski, Poznań University of Life Sciences, PolandAhmad Pervez, Radhey Hari Govt. Post Graduate College, India
Ted Cottrell, United States Department of Agriculture (USDA), United States
Copyright © 2021 Honek, Skuhrovec, Martinkova, Kulfan, Jauschova and Zach. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jiří Skuhrovec, amlyaXNsYXZza3Vocm92ZWNAZ21haWwuY29t