 Céline Rochais
Céline Rochais Tasmin L. Rymer
Tasmin L. Rymer Neville Pillay
Neville Pillay- 1School of Animal, Plant and Environmental Sciences, University of the Witwatersrand, Johannesburg, South Africa
- 2College of Science and Engineering, James Cook University, Cairns, QLD, Australia
- 3Center for Tropical Environmental and Sustainability Sciences, James Cook University, Cairns, QLD, Australia
Linking cognitive performance with fitness outcomes, measured using both reproductive and survival metrics, of free-living animals is crucial for understanding the evolution of cognition. Although there is increasing evidence showing a link between cognitive traits and reproductive success metrics, studies specifically exploring the link between cognition and survival are scarce. We first explore which cognitive traits related to survival have been investigated in free-living animals. We also discuss the challenges associated with investigating the links between cognition and survival. We then review studies that specifically consider survival of animals of known cognitive abilities that are either free-living or in captivity and later released into the wild. We found nine studies exploring cognitive traits in wild populations. The relationships between these cognitive traits and survival were equivocal. We found a further nine studies in captive-reared populations trained to predator cues and later released into the wild. Training to recognize predator cues was correlated with increased survival in the majority of studies. Finally, different individual intrinsic characteristics (i.e., age, body condition, personality, sex) showed varied effects between studies. We argue that finding ecologically relevant cognitive traits is crucial for gaining a better understanding of how selection impacts certain cognitive traits, and how these might contribute to an individual’s survival. We also suggest possible standardized, easy to implement, cognitive tests that can be used in long-term studies, which would generate large sample sizes, take into account intrinsic characteristics, and provide an opportunity to understand the mechanisms, development and evolution of cognition.
Introduction
Cognition refers to the processes by which animals collect, retain and use information from their environment (Shettleworth, 2010). An array of cognitive functions impact an animal’s behavior in the wild, including perception, attention, learning, memory, decision-making and executive functions (flexibility, categorization, problem-solving; Cauchoix and Chaine, 2016). Individuals vary in their cognitive abilities, which likely influences how they react to changes in their environment (Mazza et al., 2018). Consequently, individual variation in cognitive ability is a key component of fitness (Morand-Ferron et al., 2015; Pritchard et al., 2016), which is broadly defined as “the capacity of organisms to survive and reproduce in their environment” (Hua and Bromham, 2018). However, the relationship between cognition and fitness, both survival and reproductive success, is likely to differ between species, and may depend on the physical/social environment and/or life-history traits. Thus, exploring individual variation in cognition, and its relationship with different fitness components, could aid in understanding how and why cognitive abilities have evolved (Boogert et al., 2018).
Several studies have investigated how fitness, estimated using reproductive success metrics, relates to cognition (Supplementary Appendix 1). Surprisingly, few studies have focused on the link between survival, another important fitness proxy, and cognition. Reproductive success and survival are inherently linked since individuals with greater survival also increase their chances of having higher lifetime reproductive success. Furthermore, there are many competing demands for resources that may influence the decision making process on whether to promote growth or reproduction, whether to invest in protection and survival, or to find an optimal balance between these competing interests (Abram et al., 2021), all of which likely depend on cognitive processes. In addition, many studies state that cognitive traits (e.g., learning) can impact survival (e.g., Pravosudov and Roth, 2013), yet the majority of these studies do not specifically test whether this is the case. One explanation could be that measuring survival itself is particularly challenging.
In free-living animals, it is often difficult to discern whether an individual has died or has merely dispersed from the area of capture (e.g., Huebner et al., 2018). Indeed, the only definitive assessment of survival is observed mortality, which is often not observed directly in nature, and ultimately leads to missing values in datasets. Calculating survival is therefore more complicated than assessing reproductive success, and likely requires data-intensive capture-mark-recapture, which could be also influenced by individual personality (e.g., neophobia), and modeling that account for the possibility of dispersal. In addition, species-specific life history traits, particularly for survival, make comparative studies difficult. For example, some studies use juvenile survival (e.g., Sonnenberg et al., 2019), whereas others use adult survival (e.g., Maille and Schradin, 2016; Huebner et al., 2018). Nevertheless, survival analyses are used in other ecological studies (e.g., Santos et al., 2015; MacLeod et al., 2018), regardless of these limitations, and using data loggers could help to determine survival (e.g., African striped mice, Rhabdomys pumilio, Maille and Schradin, 2016; Vuarin et al., 2019), which begs the question of why studies relating cognition to survival are not more common. While studying the links between cognition and survival is challenging, one approach that could be adopted is to first measure the performance of individuals in one cognitive test over multiple presentations (i.e., time) or use several cognitive tests to evaluate several cognitive functions (Rowe and Healy, 2014). Thereafter, studies could explore how cognitive ability correlates with a particular survival metric such as “survivor or non-survivor” or “days of longevity” (e.g., Huebner et al., 2018). Using several survival measures should be considered since only one measure could be misleading through overestimation or underestimation (Boogert et al., 2018).
Although greater cognitive performance is associated with fitness benefits, increasing evidence of natural within-species variation in cognitive ability indicates that cognition is not cost-free (reviewed in Thornton and Lukas, 2012). For example, in fruit flies (Drosophila melanogaster) tested in the laboratory, learning performance was negatively correlated with longevity (Lagasse et al., 2012), physiological response to food restriction (Mery and Kawecki, 2005) and larval competitive ability (Mery and Kawecki, 2003). In addition, whereas some cognitive traits may be positively or negatively associated with fitness, not all cognitive traits may affect fitness depending on their ecological relevance. For example, song repertoire size, an indirect predictor of various reproductive success measures, in male song sparrows (Melospiza melodia) is positively correlated with inhibitory control (detour reaching performance) but not with learning performance in a motor or color discrimination task (Boogert et al., 2011).
The different outcomes described above also indicate many challenges in linking cognitive traits and fitness. For example, the relationships between cognitive abilities and survival may vary depending on the study design (e.g., laboratory vs. studies in the wild, sample size, chosen cognitive and survival metrics) and individual characteristics of particular species (Huebner et al., 2018), such as sex (Maille and Schradin, 2016). For example, a positive relationship between learning (aversive conditioning) and survival post-release in the wild occurs in black-tailed prairie dogs (Cynomys ludovicianus, Shier and Owings, 2006) but not in greater bilbies (Macrotis lagopus, Moseby et al., 2012). This could be a species-specific response, but it could also be due to differences in study design; Shier and Owings (2006) trained prairie dogs with only a single cue, whereas Moseby et al. (2012) trained bilbies with multiple cues. Survival is also age-dependent, generally decreasing with increasing age in several species (Lemaître and Gaillard, 2020). Senescence is a widespread phenomenon that is especially pronounced in cognitive abilities but it may be delayed in species with strong selection on cognitive abilities, such as food-caching species (Heinen et al., 2021). Personality traits also influence cognitive abilities (Sih and Del Giudice, 2012), fitness outcomes (Collins et al., 2019) and add challenges to measuring survival (e.g., neophobia may decrease recapture success and may bias estimates of survival). Currently, links between cognition and survival have generally only been studied in a small number of free-living species, often focusing on a single cognitive trait (Huebner et al., 2018), and not taking into account intrinsic characteristics such as sex, age and personality, making generalizations on the relationship between cognition and survival challenging.
In this review, we provide the methods and results obtained from cognitive studies of (i) different species tested directly in the wild, (ii) free-living species temporarily held and tested in a field laboratory and later returned to the wild, and (iii) captive-reared species later reintroduced to the wild. In these studies, individual cognitive traits were measured and related to specific survival metrics as a proxy of fitness. Our aims were to: (1) investigate whether there is a positive, negative or absence of a relationship between cognition and survival in animals in nature or studied in captivity but released into the wild, and (2) discuss the challenges of studying this relationship in a natural environment. Finally, we suggest some experimental approaches to studying cognition in free-living species to enable us to more accurately assess the relationship between cognition and survival.
Methods
Literature Survey
We first conducted a thorough search of the literature using the ISI Web of Science. We started with 32 broad (e.g., cognition or cognitive trait) and specific (e.g., “working memory” or “spatial learning”) keywords (Figure 1). Each of these keyword searches was then refined with five additional keywords (survival, mortality, longevity, death and fitness; Figure 1). These searches resulted in a total of 277,438 hits. Therefore, we further refined each search by including first the keyword “animal” (Figure 1). If this did not reduce the number of hits to a manageable number (e.g., 60), we further refined the search by including each of the following taxonomic groups: mammal, primate, bird, reptile, amphibian, fish, insect and invertebrate (Figure 1). This resulted in a total of 6,637 hits. Articles were discarded if they did not reach three criteria (Figure 1): (1) studies had to be conducted on animals that were free-living or were wild strains that had been brought into captivity for a short period for testing and then released (e.g., studies conducted on laboratory strains, such as rats, mice and fish, were not considered). (2) Studies had to explicitly evaluate survival (e.g., some studies merely mentioned survival as a potential benefit but did not actually test this). (3) Studies had to explicitly evaluate the relationship between the cognitive trait and survival.
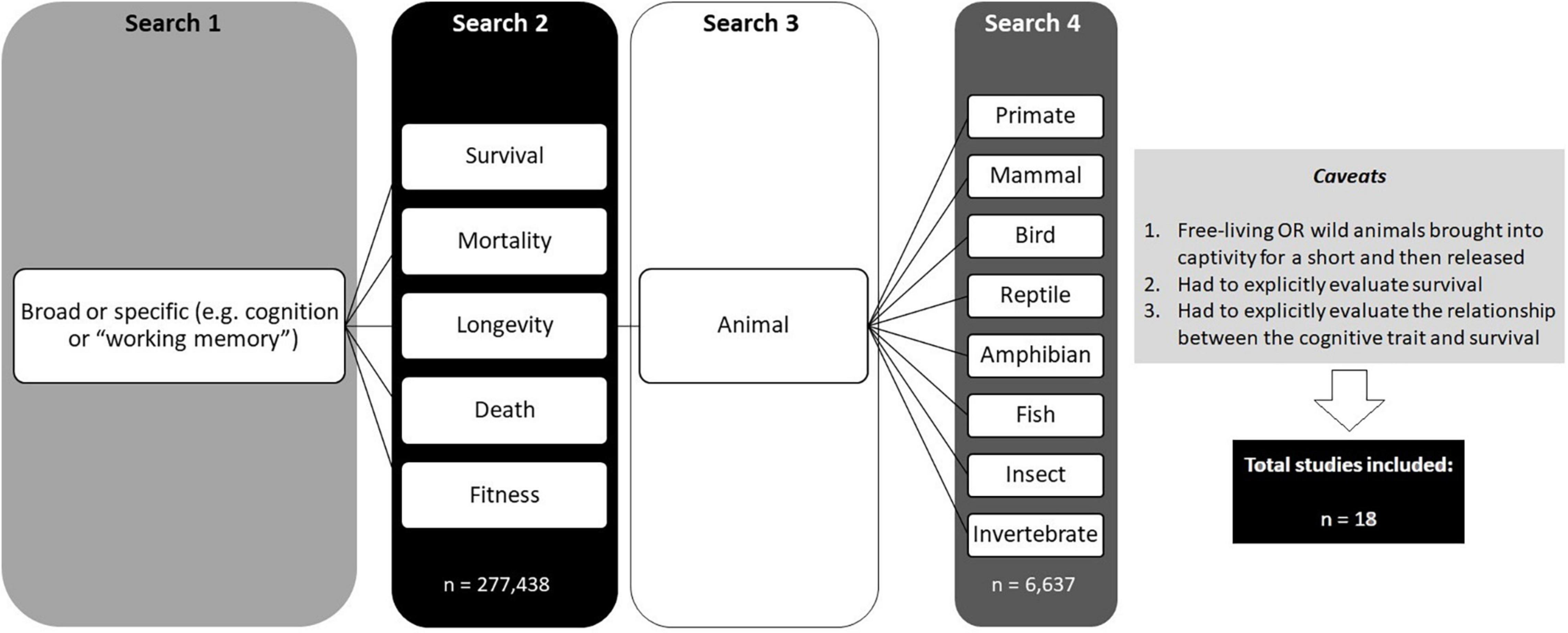
Figure 1. Flow diagram showing the literature search process and the criteria used to include/exclude papers in the review.
Results
We found nine studies linking cognition and survival in free-living animals (Table 1) and nine studies in captive reared animals reintroduced to the wild (Table 2). Different cognitive tasks were used in each study as appropriate for the study species. For example, a Barnes maze was used for spatial learning and memory in striped mice (Maille and Schradin, 2016), whereas “spatial arrays” were used for spatial learning and memory in mountain chickadees (Poecile gambeli, e.g., Sonnenberg et al., 2019). In addition, tasks were based either on artificial stimuli (e.g., color discrimination learning, lever pulling problem-solving, Madden et al., 2018) or natural stimuli (e.g., attention toward a predator, memory to find a shelter in a predator-mimicking situation, e.g., Ferrari et al., 2014). Finally, different individual intrinsic characteristics (i.e., age, body condition, personality, sex) were included as co-variates that could explain the link between cognition and survival. However, these varied between studies.
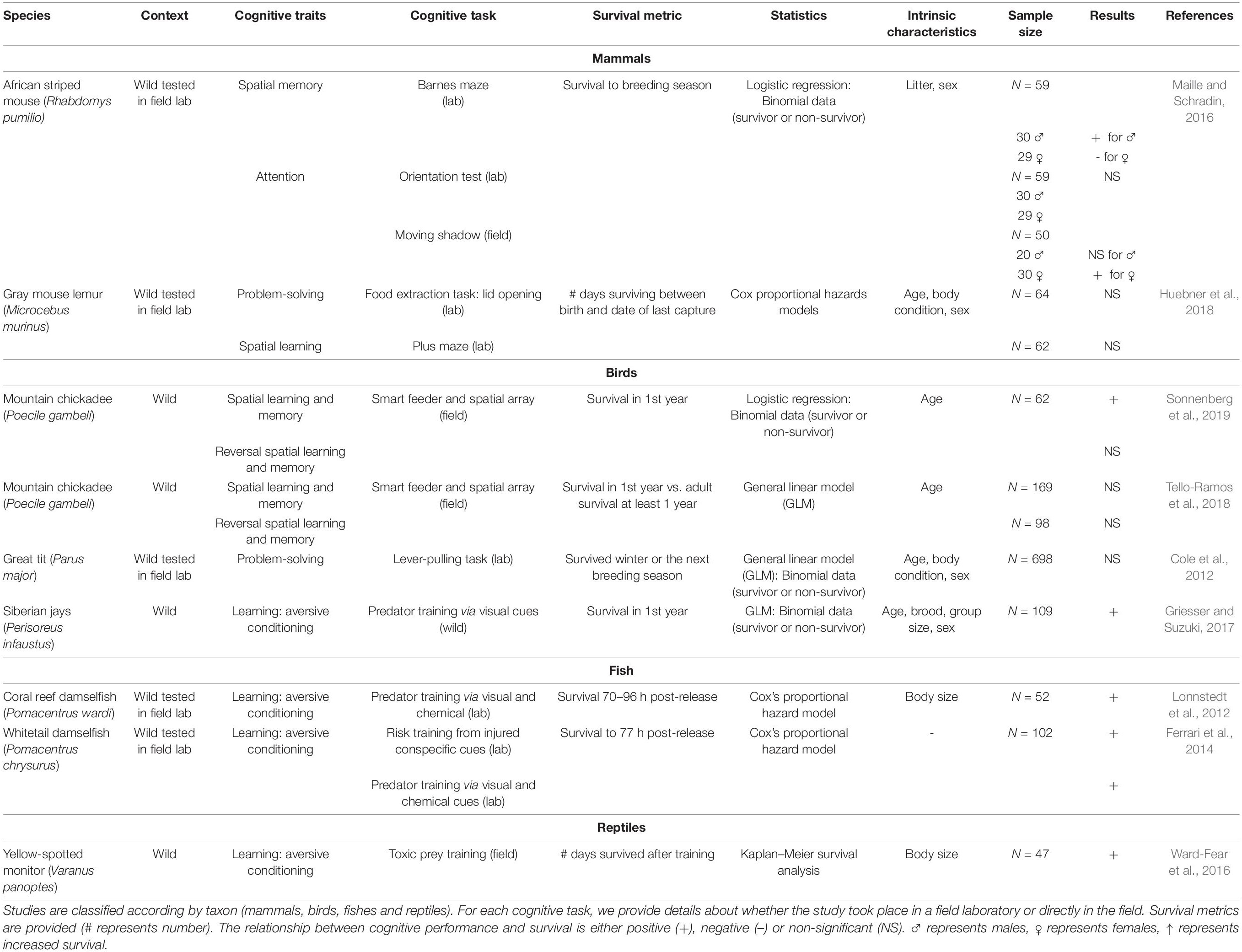
Table 1. Overview of studies linking cognitive performance and survival in free-living animal populations.
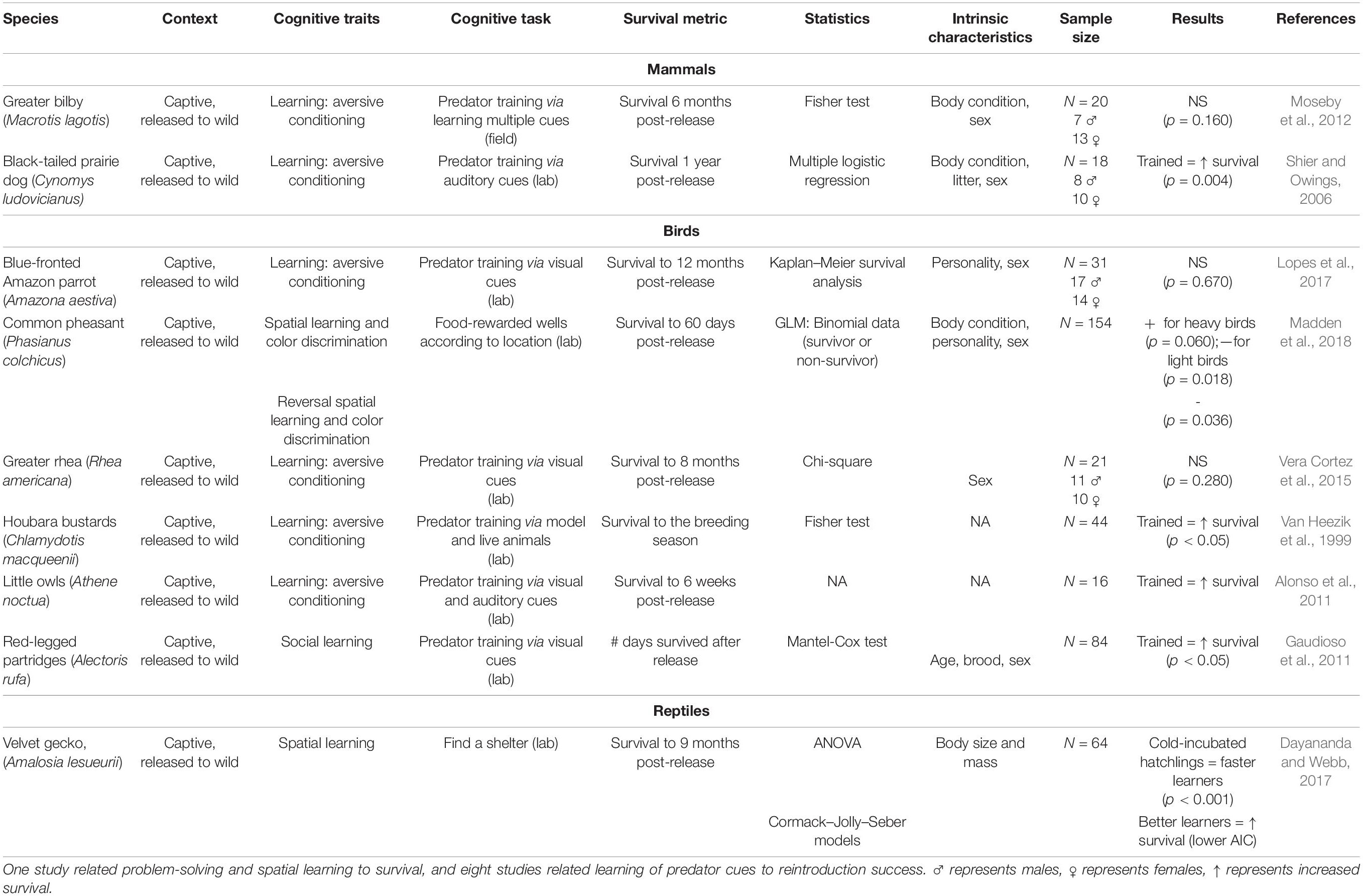
Table 2. Overview of studies linking cognitive abilities in captive animals and their reintroduction success.
Literature Survey: Cognition and Survival in Wild Populations
In the nine studies on the relationship between several cognitive traits (i.e., problem-solving, spatial and reversal learning, spatial memory) and survival in wild populations, four (three on birds, and one on a reptile; Table 1) measured the traits in situ, and five (two on mammals, one on birds, and two on fish; Table 1) measured the traits in a field laboratory before returning the individuals back to the wild. All studies training individuals to predator cues found a 100% positive relationship between training/learning and survival (N = 5; Table 1). Studies testing spatial learning and memory found contrasting results (Npositive relationship = 2, but one only with males; N non–significant relationship = 2). One study testing attention found a positive relationship only with females. Studies testing problem-solving (N = 2) or reversal spatial learning (N = 2) did not find any significant relationship with survival.
Literature Survey: Cognition and Survival in Captive Reared Populations
We found one study on the common pheasant (Phasianus colchicus) that aimed to relate the performance of two cognitive traits (i.e., discriminative, spatial and reversal learning) with survival when these pheasants were later reintroduced to the wild (Table 2). The results showed a negative relationship between survival and reversal learning, that is, pheasants that were slow to reverse previously learned associations were more likely to survive, and a positive relationship between discriminative learning and survival in heavy but not light birds (Madden et al., 2018). Eight studies (two on mammals, five on birds, and one on reptiles; Table 2) aimed to train captive-bred animals to predator-prey stimuli to increase species reintroduction success later. Five studies found a positive relationship between training and survival, whereas three studies did not find any significant relationship.
Discussion
We found nine studies specifically focusing on the survival of free-living animals of known cognitive abilities and nine studies exploring survival of captive-reared species later reintroduced to the wild. We found that study designs varied widely in sample size, chosen cognitive traits and survival metrics, and in the inclusion of intrinsic characteristics such as sex, age, and personality. It is challenging practically to link cognition and fitness proxies in free-living animals for two reasons. First, it is difficult to find cognitive traits that are aligned with the ecology of the study species, that are biologically relevant, and that are likely to explain survival across species. Second, in some species, it is difficult to distinguish between dispersal and death to be able to evaluate survival, an often overlooked fitness metric.
The first challenge when studying the effects of cognition on survival is finding appropriate cognitive traits to test in relation to fitness proxies. For example, Huebner et al. (2018) suggested that the absence of a relationship between problem-solving and spatial learning performance and survival in gray mouse lemurs (Microcebus murinus) shows that survival might not be predicted by the specific cognitive abilities addressed in the study. Some cognitive traits might be under selection in some species compared to others based on the ecology of that species (e.g., memory in food caching birds, Sonnenberg et al., 2019; problem-solving in non-migratory birds, Sol et al., 2016). Thus, one solution is to examine cognitive traits related to a particular hypothesis for an increase or decrease in survival, and hence choose an ecologically relevant cognitive trait to disentangle what selective pressures are acting on it (Boogert et al., 2018). Alternatively, assessing diverse cognitive functions in free-living animals, including perception, learning, memory (short and long-term), attention, decision-making and executive functions (flexibility, categorization, problem-solving), will help us understand whether cognitive performance in a particular context is associated with fitness benefits and/or costs in different species (Cauchoix and Chaine, 2016). For example, attentional processes will impact an animal’s behavior in the wild in many different situations, and it has been suggested that attention might explain the relationship between general cognitive ability and survival skills in several species (Matzel et al., 2020). Ultimately, it would be beneficial to find candidate cognitive traits that are likely to explain survival across species.
Some studies merely mentioned, but did not actually test for, survival as a potential benefit, whereas others clearly explore these relationships. For example, the wealth of literature on the cognitive ecology of food-storing in birds that has emerged over the last 30 years suggests that studying spatial memory abilities and associated neural structures in food-hoarding animals that rely on food caching and later retrieval of caches is relevant to understanding overwintering survival (reviewed in Pravosudov and Roth, 2013). Some meta-analysis studies also showed a positive relationship between brain size and innovative propensity as a measure of cognition and survival (Sol et al., 2005, 2007). Nevertheless, only two recent studies in birds empirically tested the relationship between spatial memory abilities and survival per se (Tello-Ramos et al., 2018; Sonnenberg et al., 2019). Tello-Ramos et al. (2018) found no significant relationship between survival to the first year and spatial and reversal learning performances, whereas Sonnenberg et al. (2019) found increased survival in individuals performing better in spatial learning and memory tasks. Considering that an array of cognitive abilities could impact an animal’s behavior in the wild, it should be noted that some cognitive traits, such as perception or decision-making, and some executive functions (e.g., categorization) have not been studied, either in relation to survival (Table 1) or to reproductive success (Supplementary Appendix 1).
Studies in cognitive conservation biology (i.e., individuals being aversively conditioned in captivity with olfactory and/or visual stimuli from predators) could be relevant for demonstrating the adaptive value of learned information. Some studies showed a higher proportion of population survival in captive-bred animals trained to recognized predator stimuli (Table 2). One study in captivity did not train animals with predator cues but tested two cognitive traits. Madden et al. (2018) related discriminative, spatial and reversal learning performance of captive common pheasants with survival when these pheasants were later reintroduced to the wild; individuals that were slow to reverse previously learned associations (reversal learning task) were more likely to survive to 4 months old (Madden et al., 2018). We included studies on individuals reared in captivity in the results of our review but we should be cautious about the interpretation of fitness consequences of natural variation in learning ability (Morand-Ferron, 2017). Indeed, animals reared in captivity are not confronted with the same environmental challenges compared to their free-living counterparts, particularly if they are retained in captivity from an early age. They are often limited in their food diversity, and/or may be food deprived, forcing participation in a test. In addition, captive animals may be limited by space, again potentially forcing participation, or increasing stress that might impact the results. Consequently, results relating cognition to survival in animals reared in captivity should be treated with caution since they may not reflect true patterns seen in free-living animals.
The second challenge when studying the effects of cognition on survival will be to accurately distinguish between dispersal and death, which will permit evaluation of survival in some species. Calculating survival likely requires data-intensive capture-mark-recapture and modeling that account for the possibility of dispersal. Thus, survival could be better estimated in species with no breeding dispersal. For example, food-caching Parids, such as mountain chickadees, are permanent residents that have just one post-natal dispersal movement during their first year of life, enabling the study of test birds for several years (Sonnenberg et al., 2019; Heinen et al., 2021). However, some authors have managed to assess the relationship between cognition and survival in species showing dispersal. For example, Maille and Schradin (2016) found that wild male African striped mice that performed better in a long-term spatial memory task survived for longer. Survival was measured as the number of days alive before the onset of the breeding season and hence before long-distance dispersal. A mouse was considered to have died when it had not been trapped or observed for at least two consecutive months. Survival could also be estimated in individuals where one sex does not disperse. However, this will lead to biased assumptions about how and why cognitive abilities have evolved in a species (Boogert et al., 2018). Furthermore, dispersal and survival are often inter-linked (e.g., an animal that disperses from the natal nest has a higher probably of being predated than one that does not, Clobert et al., 2009). It would be interesting to investigate differences in cognitive abilities between dispersing and non-dispersing species in the same taxonomic group. This could highlight specific cognitive traits that will enhance survival according to the presence/absence of dispersal in phylogenetically closely related species, allowing us to better understand the adaptive value of cognitive traits.
Intrinsic characteristics, such as age, sex and personality, are not always known or considered in studies related to cognition and survival in the wild. These intrinsic characteristics can impact inter-individual variation in cognitive performance and survival. For example, in striped mice, faster attention toward a predator stimulus was positively linked with survival in females but not males, yet male, but not female, survival co-varied positively with better spatial memory of shelter locations (Maille and Schradin, 2016). Survival is also strongly age-dependent (Lemaître and Gaillard, 2020). Therefore, it is important to control for age either during data collection or statistically by taking into account age as a covariate in statistical models. Senescence in specific cognitive traits could also be delayed in species with strong selection on certain cognitive traits, such as in food-caching species, and hence could influence survival outcomes (Heinen et al., 2021). Personality can also influence key behaviors for survival in the wild (e.g., Aplin et al., 2013). Bold animals can adopt risky behaviors toward predators but will also be more willing to explore their environment and increase their chances of finding food or mates (MacPherson et al., 2017). Thus, individual differences in personality might drive individual differences in cognitive abilities, and together they might drive responsiveness to environmental change (Griffin et al., 2015) and ultimately survival. From a practical point of view, personality traits add challenges to accurate survival measurements. For example, neophobia may influence recapture success, and hence may bias estimates of survival. We suggest that studies attempting to link survival or other fitness proxies (such as reproductive success) to cognition should always consider other possible influencing factors, such as sex, age, or personality.
It is also challenging to find the appropriate task to test a particular cognitive trait. For example, individual variation in problem-solving performance could be related to motivational variation rather than cognitive mechanisms per se (Van Horik and Madden, 2016). This leads to issues of either capturing animals and testing them in a field laboratory for short-term tests or testing them directly in the field under spontaneous, but less controlled, conditions (e.g., with the possible impact of social learning). Furthermore, assessing the survival of wild animals of known cognitive abilities requires tracking the same individuals in space and over time (Tello-Ramos et al., 2018). The cognitive test should then be standardized between studies and easily applied to be used permanently for long term studies (e.g., Morand-Ferron et al., 2015; Cauchoix et al., 2017). Automated operant devices placed directly in the field could be appropriate for cognitive tests, since animals can participate in a task spontaneously, increasing the probability of generating a large sample size (Morand-Ferron et al., 2016). One could argue that experimental protocols involving novel objects may elicit avoidance in neophobic individuals (Stuber et al., 2013) or those that have better access to other resources (e.g., adults compared to juveniles, Morand-Ferron et al., 2011). However, a longer duration of the experiment could lead to a familiarization to the devices, allowing novelty responses to reduce with time, thereby eliminating personality biases, such as has been observed in great tits (Parus major, Morand-Ferron et al., 2015). In addition, the device could be adapted to different species while still assessing the same cognitive response. For example, automated devices have been used in studies of passerine species but are difficult to create and modify. Thus, creating simple, ecologically relevant tasks, such as mimicking a predator shape in order to measure attention, setting a novel object in the field to measure habituation (basis of learning), or documenting the technique and time required for nest building as a measure of problem-solving could also be relevant in studying the evolution of cognition (Keagy et al., 2011).
Standardized and easy to use tests will help to generate sample sizes with better statistical power. Generally, an acceptable statistical power is only associated with very large sample sizes, which are often difficult, if not impossible, to obtain in the wild. The practical challenge will then be to conduct long term studies over several years, such as in Cole et al. (2012). However, to reach a sample size with better statistical power in the wild would likely require several years of study, particularly for species that are rare or cryptic. Conducting studies across years comes with additional potential confounding effects, such as seasonal changes, weather effects (e.g., temperature and humidity changes), unexpected catastrophic events (e.g., bushfires or floods) and human impacts (e.g., deforestation and land degradation).
Some taxa appear to be good study candidates for obtaining adequate sample sizes for linking fitness proxies and cognitive abilities. The few studies conducted on mammals indicate that rodents may be appropriate because of their small body size, short longevity, ease of study and ability to obtain large sample sizes in general (e.g., Maille and Schradin, 2016). Short longevity can add a practical advantage to measuring survival. For example, in striped mice, it is possible to measure several cognitive traits during their 1-year lifespan. Similarly, among birds, most work has been done on short-lived passerines, generating large sample sizes (e.g., Cole et al., 2012). Fish could also be promising candidates for future studies. For example, experimental studies on guppies (Poecilia reticulata) showed that individuals with larger brains had improved cognitive abilities (spatial learning tested in a sample of N = 450; Kotrschal et al., 2013), and large-brained females had higher survival compared to small-brained females (N = 794; Kotrschal et al., 2015), which was related to predation pressure in a wild population (N = 160; Kotrschal et al., 2017). Some other model species relating cognition and fitness in the wild are absent in the literature. For example, cephalopods, known for their complex cognitive abilities such as flexible decision-making and future planning (Billard et al., 2020), could be relevant models of study. Some insect species could be candidates for obtaining adequate sample sizes, and linking cognitive performance and fitness (e.g., Raine and Chittka, 2008); however, it may be challenging to study them in the wild and measure their survival. Overall, more studies are needed on survival in a diverse range of free-living animal species of known cognitive abilities. This will allow for comparative studies, highlight potential differences in which cognitive traits are important for survival in some taxa but not in others, and finally will allow us to better understand the factors shaping the evolution of animal cognition.
Because there is no standardized approach to link survival and cognition, the differing results of previous studies could also indicate that the outcome depends on the conditions under which fitness proxy metrics are assessed. Depending on the study species, survival can be evaluated in days from birth to death, in days from birth to dispersal (e.g., Maille and Schradin, 2016), or as seasonal or annual survival vs. death (e.g., Sonnenberg et al., 2019). Madden et al. (2018) determined a threshold for survival at 60 days in common pheasants by using a putative survival curve based on previous observations and according to anthropogenic activity (pheasant shooting). Studies in fish monitored survival from 70 to 96 h post release (Lonnstedt et al., 2012; Ferrari et al., 2014). These different measurements suggest that it is essential to consider a species’ characteristics to determine the relevant survival metric to use as a fitness proxy. It is also important to potentially define the baseline longevity of a population and then measure deviations related to cognition to generate continuous data rather than simple binary (yes/no) outcomes and, hence, explore how survival relates to the evolution of cognition.
Conclusion
Linking survival and experimental measures of cognitive performance is challenging in free-living species, and clearly more research is needed. Practical challenges for studying wild animals include: (i) proper selection of a cognitive task that will test for ecologically relevant cognitive traits and will be likely to explain the relationship between cognition and survival across species; and (ii) choice of a fitness metric, and if it is possible, consideration of several fitness proxies (e.g., individual survival, clutch size, fledgling number), depending on the species’ life history. Moreover, generating large sample sizes to reach an acceptable statistical power, and accounting for individual intrinsic characteristics, such as age, sex and personality, should be considered in cognition studies to partition the direct selection on cognition from other selection pressures. Finding appropriate methods and protocols adapted to wild species will highlight positive, negative or no relationships between cognition and survival, and hence disentangle benefits or costs of cognition. Linking fitness and cognitive ability might explain intra- and/or inter-species variation in a comparative and integrated evolutionary framework.
Author Contributions
All authors listed have made a substantial, direct, and intellectual contribution to the work, and approved it for publication.
Funding
This research was supported by a fellowship (to CR) by the University of the Witwatersrand.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
This study was made possible by the administrative and technical support of the Succulent Karoo Research Station (registered South African NPO 122-134). We would like to thank reviewers for providing valuable comments on previous versions of the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.729546/full#supplementary-material
References
Abram, F., Arcari, T., Guerreiro, D., and O’Byrne, C. (2021). Evolutionary trade-offs between growth and survival: the delicate balance between reproductive success and longevity in bacteria. Adv. Microb. Physiol. 79, 133–162. doi: 10.1016/bs.ampbs.2021.07.002
Alonso, R., Orejas, P., Lopes, F., and Sanz, C. (2011). Pre-release training of juvenile little owls Athene noctua to avoid predation. Anim. Biodivers. Conserv. 34, 389–393.
Aplin, L. M., Farine, D. R., Morand-Ferron, J., Cole, E. F., Cockburn, A., and Sheldon, B. C. (2013). Individual personalities predict social behaviour in wild networks of great tits (Parus major). Ecol. Lett. 16, 1365–1372. doi: 10.1111/ele.12181
Billard, P., Schnell, A. K., Clayton, N. S., and Jozet-Alves, C. (2020). Cuttlefish show flexible and future-dependent foraging cognition. Biol. Lett. 16:20190743. doi: 10.1098/rsbl.2019.0743
Boogert, N. J., Anderson, R. C., Peters, S., Searcy, W. A., and Nowicki, S. (2011). Song repertoire size in male song sparrows correlates with detour reaching, but not with other cognitive measures. Anim. Behav. 81, 1209–1216. doi: 10.1016/j.anbehav.2011.03.004
Boogert, N. J., Madden, J. R., Morand-Ferron, J., and Thornton, A. (2018). Measuring and understanding individual differences in cognition. Phil. Trans. R. Soc. B Biol. Sci. 373, 20170280. doi: 10.1098/rstb.2017.0280
Cauchoix, M., and Chaine, A. S. (2016). How can we study the evolution of animal minds? Front. Psychol. 7:358. doi: 10.3389/fpsyg.2016.00358
Cauchoix, M., Hermer, E., Chaine, A. S., and Morand-Ferron, J. (2017). Cognition in the field: comparison of reversal learning performance in captive and wild passerines. Sci. Rep. 7:12945. doi: 10.1038/s41598-017-13179-5
Clobert, J., Le Galliard, J. F., Cote, J., Meylan, S., and Massot, M. (2009). Informed dispersal, heterogeneity in animal dispersal syndromes and the dynamics of spatially structured populations. Ecol. Lett. 12, 197–209. doi: 10.1111/j.1461-0248.2008.01267.x
Cole, E. F., Morand-ferron, J., Hinks, A. E., and Quinn, J. L. (2012). Cognitive ability influences reproductive life history variation in the wild. Curr. Biol. 22, 1808–1812. doi: 10.1016/j.cub.2012.07.051
Collins, S. M., Hatch, S. A., Elliott, K. H., and Jacobs, S. R. (2019). Boldness, mate choice and reproductive success in Rissa tridactyla. Anim. Behav. 154, 67–74. doi: 10.1016/j.anbehav.2019.06.007
Dayananda, B., and Webb, J. K. (2017). Incubation under climate warming affects learning ability and survival in hatchling lizards. Biol. Lett. 13:20170002. doi: 10.1098/rsbl.2017.0002
Ferrari, M. C. O., Mccormick, M. I., Meekan, M. G., and Chivers, D. P. (2014). Background level of risk and the survival of predator-naive prey : can neophobia compensate for predator naivety in juvenile coral reef fishes? Proc. R. Soc. B Biol. Sci. 282:20142197. doi: 10.1098/rspb.2014.2197
Gaudioso, V. R., Sánchez-García, C., Pérez, J. A., Rodríguez, P. L., Armenteros, J. A., and Alonso, M. E. (2011). Does early antipredator training increase the suitability of captive red-legged partridges (Alectoris rufa) for releasing? Poult. Sci. 90, 1900–1908. doi: 10.3382/ps.2011-01430
Griesser, M., and Suzuki, T. N. (2017). Naive juveniles are more likely to become breeders after witnessing predator mobbing. Am. Nat. 189, 58–66. doi: 10.1086/689477
Griffin, A. S., Guillette, L. M., and Healy, S. D. (2015). Cognition and personality: an analysis of an emerging field. Trends Ecol. Evol. 30, 207–214. doi: 10.1016/j.tree.2015.01.012
Heinen, V. K., Pitera, A. M., Sonnenberg, B. R., Benedict, L. M., Branch, C. L., Bridge, E. S., et al. (2021). Specialized spatial cognition is associated with reduced cognitive senescence in a food-caching bird. Proc. R. Soc. B Biol. Sci. 288:20203180. doi: 10.1098/rspb.2020.3180
Hua, X., and Bromham, L. (2018). Fitness in Encyclopedia of Ecology, 2nd Edn, Volume 3. Netherlands: Elsevier, 102–109. doi: 10.1016/B978-0-12-409548-9.09361-1
Huebner, F., Fichtel, C., and Kappeler, P. M. (2018). Linking cognition with fitness in a wild primate: fitness correlates of problem- solving performance and spatial learning ability. Phil. Trans. R. Soc. B Biol. Sci. 373:20170295. doi: 10.1098/rstb.2017.0295
Keagy, J., Savard, J. F., and Borgia, G. (2011). Complex relationship between multiple measures of cognitive ability and male mating success in satin bowerbirds. Anim. Behav. 81, 1063–1070. doi: 10.1016/j.anbehav.2011.02.018
Kotrschal, A., Buechel, S. D., Zala, S. M., Corral-Lopez, A., Penn, D. J., and Kolm, N. (2015). Brain size affects female but not male survival under predation threat. Ecol. Lett. 18, 646–652. doi: 10.1111/ele.12441
Kotrschal, A., Deacon, A. E., Magurran, A. E., and Kolm, N. (2017). Predation pressure shapes brain anatomy in the wild. Evol. Ecol. 31, 619–633. doi: 10.1007/s10682-017-9901-8
Kotrschal, A., Rogell, B., Bundsen, A., Svensson, B., Zajitschek, S., Brännström, I., et al. (2013). Artificial selection on relative brain size in the guppy reveals costs and benefits of evolving a larger brain. Curr. Biol. 23, 168–171. doi: 10.1016/j.cub.2012.11.058
Lagasse, F., Moreno, C., Preat, T., and Mery, F. (2012). Functional and evolutionary trade-offs co-occur between two consolidated memory phases in Drosophila melanogaster. Proc. R. Soc. B Biol. Sci. 279, 4015–4023. doi: 10.1098/rspb.2012.1457
Lemaître, J.-F., and Gaillard, J.-M. (2020). Senescence in the Wild: Theory and Physiology in Encyclopedia of Biomedical Gerontology. Netherlands: Elsevier, 185–192.
Lonnstedt, O. M., Mccormick, M. I., Meekan, M. G., Ferrari, M. C. O., and Chivers, D. P. (2012). Learn and live: predator experience and feeding history determines prey behaviour and survival. Proc. R. Soc. B Biol. Sci. 279, 2091–2098. doi: 10.1098/rspb.2011.2516
Lopes, A. R. S., Rocha, M. S., Junior, M. G. J., Mesquita, W. U., Silva, G. G. G. R., Vilela, D. A. R., et al. (2017). The influence of anti-predator training, personality and sex in the behavior, dispersion and survival rates of translocated captive-raised parrots. Glob. Ecol. Conserv. 11, 146–157. doi: 10.1016/j.gecco.2017.05.001
MacLeod, K. J., Sheriff, M. J., Ensminger, D. C., Owen, D. A. S., and Langkilde, T. (2018). Survival and reproductive costs of repeated acute glucocorticoid elevations in a captive, wild animal. Gen. Comp. Endocrinol. 268, 1–6. doi: 10.1016/j.ygcen.2018.07.006
MacPherson, B., Mashayekhi, M., Gras, R., and Scott, R. (2017). Exploring the connection between emergent animal personality and fitness using a novel individual-based model and decision tree approach. Ecol. Inform. 40, 81–92. doi: 10.1016/j.ecoinf.2017.06.004
Madden, J. R., Langley, E. J. G., Whiteside, M. A., Beardsworth, C. E., and Van Horik, J. O. (2018). The quick are the dead: pheasants that are slow to reverse a learned association survive for longer in the wild. Phil. Trans. R. Soc. B Biol. Sci. 373:20170297. doi: 10.1098/rstb.2017.0297
Maille, A., and Schradin, C. (2016). Survival is linked with reaction time and spatial memory in African striped mice. Biol. Lett. 12, 20160346. doi: 10.1098/rsbl.2016.0346
Matzel, L. D., Patel, H. M., Piela, M. C., Manzano, M. D., Tu, A., and Crawford, D. W. (2020). General cognitive ability predicts survival-readiness in genetically heterogeneous laboratory mice. Front. Ecol. Evol 8:531014. doi: 10.3389/fevo.2020.531014
Mazza, V., Eccard, J. A., Zaccaroni, M., Jacob, J., and Dammhahn, M. (2018). The fast and the flexible: cognitive style drives individual variation in cognition in a small mammal. Anim. Behav. 137, 119–132. doi: 10.1016/J.ANBEHAV.2018.01.011
Mery, F., and Kawecki, T. J. (2003). A fitness cost of learning ability in Drosophila melanogaster. Proc. R. Soc. B Biol. Sci. 270, 2465–2469.
Mery, F., and Kawecki, T. J. (2005). A cost of long-term memory in Drosophila. Science 308:1148. doi: 10.1126/science.1111331
Morand-Ferron, J. (2017). Why learn? Curr. Opin. Behav. Sci. 16, 73–79. doi: 10.1016/j.cobeha.2017.03.008
Morand-Ferron, J., Cole, E. F., and Quinn, J. L. (2016). Studying the evolutionary ecology of cognition in the wild: a review of practical and conceptual challenges. Biol. Rev. 91, 367–389. doi: 10.1111/brv.12174
Morand-Ferron, J., Cole, E. F., Rawles, J. E. C., and Quinn, J. L. (2011). Who are the innovators? Behav. Ecol. 22, 1241–1248. doi: 10.1093/beheco/arr120
Morand-Ferron, J., Hamblin, S., Cole, E. F., Aplin, L. M., and Quinn, J. L. (2015). Taking the operant paradigm into the field: associative learning in wild great tits. PLoS One 10:e0133821. doi: 10.1371/journal.pone.0133821
Moseby, K. E., Cameron, A., and Crisp, H. A. (2012). Can predator avoidance training improve reintroduction outcomes for the greater bilby in arid Australia? Anim. Behav. 83, 1011–1021. doi: 10.1016/j.anbehav.2012.01.023
Pravosudov, V. V., and Roth, T. C. (2013). Cognitive ecology of food hoarding: the evolution of spatial memory and the hippocampus. Annu. Rev. Ecol. Evol. Syst. 44, 173–193.
Pritchard, D., Hurly, T., Tello-Ramos, M., and Healy, S. (2016). Why study cognition in the wild (and how to test it)? J. Exp. Anal. Behav. 105, 41–55. doi: 10.1002/jeab.195
Raine, N. E., and Chittka, L. (2008). The correlation of learning speed and natural foraging success in bumble-bees. Proc. R. Soc. B Biol. Sci. 275, 803–808. doi: 10.1098/rspb.2007.1652
Rowe, C., and Healy, S. D. (2014). Measuring variation in cognition. Behav. Ecol. 25, 1287–1292. doi: 10.1093/beheco/aru090
Santos, C. D., Cramer, J. F., Pâraû, L. G., Miranda, A. C., Wikelski, M., and Dechmann, D. K. N. (2015). Personality and morphological traits affect pigeon survival from raptor attacks. Sci. Rep. 5:15490. doi: 10.1038/srep15490
Shettleworth, S. J. (2010). Cognition, Evolution and Behavior, 2nd Edn. New York, USA: Oxford University Press.
Shier, D. M., and Owings, D. H. (2006). Effects of predator training on behavior and post-release survival of captive prairie dogs (Cynomys ludovicianus). Biol. Conserv. 132, 126–135. doi: 10.1016/J.BIOCON.2006.03.020
Sih, A., and Del Giudice, M. (2012). Linking behavioural syndromes and cognition: a behavioural ecology perspective. Phil. Trans. R. Soc. B Biol. Sci. 367, 2762–2772. doi: 10.1098/rstb.2012.0216
Sol, D., Duncan, R. P., Blackburn, T. M., Cassey, P., and Lefebvre, L. (2005). Big brains, enhanced cognition, and response of birds to novel environments. Proc. Natl. Acad. Sci. U.S.A. 102, 5460–5465. doi: 10.1073/pnas.0408145102
Sol, D., Sayol, F., Ducatez, S., and Lefebvre, L. (2016). The life-history basis of behavioural innovations. Phil. Trans. R. Soc. B Biol. Sci. 371:20150187. doi: 10.1098/rstb.2015.0187
Sol, D., Székely, T., Liker, A., and Lefebvre, L. (2007). Big-brained birds survive better in nature. Proc. R. Soc. B Biol. Sci. 274, 763–769. doi: 10.1098/rspb.2006.3765
Sonnenberg, B. R., Branch, C. L., Pitera, A. M., Bridge, E., and Pravosudov, V. V. (2019). Natural selection and spatial cognition in wild food-caching mountain chickadees. Curr. Biol. 29, 670–676. doi: 10.1016/j.cub.2019.01.006
Stuber, E. F., Araya-Ajoy, Y. G., Mathot, K. J., Mutzel, A., Nicolaus, M., Wijmenga, J. J., et al. (2013). Slow explorers take less risk: a problem of sampling bias in ecological studies. Behav. Ecol. 24, 1092–1098. doi: 10.1093/beheco/art035
Tello-Ramos, M. C., Branch, C. L., Pitera, A. M., Kozlovsky, D. Y., Bridge, E. S., and Pravosudov, V. V. (2018). Memory in wild mountain chickadees from different elevations: comparing first-year birds with older survivors. Anim. Behav. 137, 149–160. doi: 10.1016/J.ANBEHAV.2017.12.019
Thornton, A., and Lukas, D. (2012). Individual variation in cognitive performance : developmental and evolutionary perspectives. Phil. Trans. R. Soc. B Biol. Sci. 367, 2773–2783. doi: 10.1098/rstb.2012.0214
Van Heezik, Y., Seddon, P. J., and Maloney, R. F. (1999). Helping reintroduced houbara bustards avoid predation: effective anti-predator training and the predictive value of pre-release behaviour. Anim. Conserv. 2, 155–163. doi: 10.1111/j.1469-1795.1999.tb00061.x
Van Horik, J. O., and Madden, J. R. (2016). A problem with problem solving: motivational traits, but not cognition, predict success on novel operant foraging tasks. Anim. Behav. 114, 189–198. doi: 10.1016/j.anbehav.2016.02.006
Vera Cortez, M., Valdez, D. J., Navarro, J. L., and Martella, M. B. (2015). Efficiency of antipredator training in captive-bred greater rheas reintroduced into the wild. Acta Ethol. 18, 187–195. doi: 10.1007/s10211-014-0206-4
Vuarin, P., Pillay, N., and Schradin, C. (2019). Elevated basal corticosterone levels increase disappearance risk of light but not heavy individuals in a long-term monitored rodent population. Horm. Behav. 113, 95–102. doi: 10.1016/J.YHBEH.2019.05.001
Keywords: cognitive ecology, reproductive success, sample size, survival, fitness
Citation: Rochais C, Rymer TL and Pillay N (2022) Challenges in Linking Cognition and Survival: A Review. Front. Ecol. Evol. 10:729546. doi: 10.3389/fevo.2022.729546
Received: 23 June 2021; Accepted: 31 January 2022;
Published: 25 February 2022.
Edited by:
Blandine Françoise Doligez, Centre National de la Recherche Scientifique (CNRS), FranceReviewed by:
Jorge Cassinello, Experimental Station of Arid Zones (CSIC), SpainPierre Bize, University of Aberdeen, United Kingdom
Copyright © 2022 Rochais, Rymer and Pillay. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Céline Rochais, Y2VsaW5lLnJvY2hhaXNAZ21haWwuY29t