 Viktória B-Béres1*
Viktória B-Béres1* Zsuzsanna Kókai1
Zsuzsanna Kókai1 Gábor Várbíró1Gulperizat Mustazhapova1,2,3Zoltán Csabai4,5Bálint Pernecker4Gábor Borics1
Gábor Várbíró1Gulperizat Mustazhapova1,2,3Zoltán Csabai4,5Bálint Pernecker4Gábor Borics1 István Bácsi6Pál Boda1
István Bácsi6Pál Boda1
- 1Centre for Ecological Research, Institute of Aquatic Ecology, Department of Tisza Research, Debrecen, Hungary
- 2Department of Biochemical Engineering, University of Debrecen, Debrecen, Hungary
- 3Department of Ecology, University of Debrecen, Debrecen, Hungary
- 4Department of Hydrobiology, University of Pécs, Pécs, Hungary
- 5Department of Botany and Zoology, Masaryk University, Brno, Czechia
- 6Department of Hydrobiology, University of Debrecen, Debrecen, Hungary
Climate change is putting increasing pressure on flowing waters. Drastic water level fluctuations in rivers or drying up of small and medium-sized streams all contribute to the biodiversity crisis threatening freshwater ecosystems. Benthic diatoms are important elements of biofilm in small streams. However, knowledge on the relationship between benthic diatoms and flow intermittence is incomplete, especially in regions recently impacted by recurrent drying. Thus, we investigated benthic diatom flora of small intermittent, hilly streams in the warm temperate region of Europe (the Pannonian Ecoregion). Our hypotheses were addressed to compositional changes, biodiversity loss and diatom-based ecological assessment. The results revealed clear flow intermittence-induced differences in taxa and trait composition of diatoms. Altogether six species for the dry phase and three species in the aquatic phase were identified as indicative ones by using indicator value analyses. In contrast to water regime induced changes in assemblages, there was a seasonal overlap in taxa and trait composition. During the study period, the drying up of streams did not result in significant biodiversity loss either at taxa or trait levels. Functional dispersion, however, reduced significantly by summer. Overall, neither the hydrological regime nor seasonal changes had a significant effect on diatom-based quality indices, except for the Rott trophic index (TID index). The TID index values were significantly lower in dry phases than in aquatic ones. These results suggested that the drying up of streams has a very complex influence on benthic diatoms. It seems that taxonomical and functional redundancy can reduce the negative impact of short-time flow intermittence on assemblages. As a practical benefit, the results are the first to support the use of diatom-based quality indices in the assessment of flow intermittence in the temperate region.
Introduction
Over the past decades, biodiversity loss has become a major problem facing humanity and as it is constantly worsening it threatens ecosystems functioning and human well-being (Díaz et al., 2006; He et al., 2019; Crabot et al., 2021; Thompson et al., 2021). Although freshwaters are one of the most diverse ecosystems, integrity loss of communities here is an increasing trend that poses a challenge to biodiversity conservation (Dudgeon et al., 2006; Reid et al., 2019). Climate change and its impact on freshwater biodiversity is one of the 12 threats named by Reid et al. (2019) endangering the entire ecosystem from specimen to community level resulting in, inter alia, reduction in cell/body size, changes in phenology, changes in species distribution, spread of invasive or non-native species or even disease outbreaks (see more in Reid et al., 2019). Rapid and extreme changes in water regime, increasing water temperature, drying of small water bodies and nutrient concentration during drying are just some of the influences that can result in irreversible transformations in freshwater ecosystems (see more in Magand et al., 2020).
As climate change, water abstraction or even land use resulted in a shift of perennial watercourses becoming intermittent ones in the last 5 decades, small streams all around the world are increasingly threatened by flow cessation (Messager et al., 2021). According to the latest IPCC report (IPCC, 2021), there is high probability of increase in extreme climatic events, including severe droughts will occur in the near future. These events will result in an increase in the number of intermittent streams and the duration of non-flow periods. In the Mediterranean regions characterized by an intra-annual moisture deficit, a number of intermittent streams can be considered as natural elements of the landscape (Sabater et al., 2016; Acuña et al., 2017). In contrast, the intermittent shift of formerly perennial streams to an intermittent state is a relatively new phenomenon in the warm temperate region, where the summers are hot and there are no significant seasonal differences in precipitation1 (B-Béres et al., 2019; Várbíró et al., 2020). Independently of the historical background of intermittent streams, i.e., drying is a new phenomenon or it dates back to the ancient times, these watercourses constitute a very large fraction of river networks and their number is probably underestimated (Sabater et al., 2016).
Intermittency is a strong pressure on organisms. Populations living in intermittent streams are either able to adapt to dry conditions or they disperse to a new area or become extinct from the given ecosystem (Falasco et al., 2016a; Sabater et al., 2016; Stubbington et al., 2017). Thus, these watercourses can be usually considered as lower diversity areas during the aquatic phase compared to permanent waters. However, appearance of lentic and terrestrial species in assemblages during intermittence can result in higher taxonomical and functional variability here (Stubbington et al., 2017). But intermittence is a strong environmental pressure that induces diversity changes during drying of streams. The replacement of taxa is quite intense then, the appearance of lentic and terrestrial taxa in assemblages first results in an increase of diversity in pools (Stubbington et al., 2017; Novais et al., 2020). But finally the increase in biotic interactions within these assemblages and the ongoing drying of pools lead to the decrease in biodiversity (Stubbington et al., 2017).
Freshwater benthic diatoms are one of the most abundant algal groups in the biofilm of streams (Pan et al., 1999), thus they play a key role in these ecosystems as primary producers and also as a food source for herbivores (Kireta et al., 2012). Since they are sensitive indicators of environmental stressors (McCormick and Cairns, 1994), they provide valuable information on the ecological quality of waters (Water Framework Directive – EC, 2000). In addition, in the last few years, an increasing number of studies have shown that benthic diatoms respond sensitively to drying of waters (Novais et al., 2014, 2020; Falasco et al., 2016a; Piano et al., 2017; B-Béres et al., 2019; Stubbington et al., 2019; Várbíró et al., 2020; Tornés et al., 2021). Previous studies pointed out that diatom indices can be useful metrics comparing the ecological quality of intermittent streams in different phases (Barthès et al., 2015; Novais et al., 2020). In a comprehensive study, Novais et al. (2020) validated that values of diatom-based metrics from dry and from previous aquatic phases are comparable and showed no significant differences. There is no doubt, however, that drying of streams results in compositional modifications within assemblages (Novais et al., 2014, 2020; Falasco et al., 2016a; Piano et al., 2017; B-Béres et al., 2019; Várbíró et al., 2020; Tornés et al., 2021). Flow cessation and drying of streams create very harsh conditions for benthic algae and finally result in a decrease in biodiversity (Sabater et al., 2017; B-Béres et al., 2019; Novais et al., 2020; Várbíró et al., 2020; Tornés et al., 2021). Although taxonomical or functional redundancy seems to be able to balance this negative effect (Novais et al., 2020; Lukács et al., 2021), latest studies suggest that this phenomenon is highly dependent on the duration of the dry periods not only in the case of diatoms (Tornés et al., 2021), but also in macroinvertebrate assemblages (Crabot et al., 2021). Others pointed out that the time scale of dry periods is crucial in determining the extent of compositional changes. Drying of streams has a stronger negative influence on benthic diatom assemblages in regions where the flow intermittence is a new phenomenon than in regions with historical occurrence of intermittent streams (Várbíró et al., 2020). Our knowledge of flow intermittence – benthic diatom relationship, however, is incomplete in regions recently impacted by recurrent drying. It is especially true in the Pannonian Ecoregion, the largest part of which belongs to Hungary. Approximately 10,000 watercourses are registered here, but less than 10% of them are monitored regularly by the Water Framework Directive-related (WFD) National Biomonitoring System2 (Stubbington et al., 2018). Very small streams with a catchment area less than 10 km2 are not included in this biomonitoring system. Thus, we have no information on how many of these small streams are affected by drying in the region. Our knowledge of how these streams contribute to the maintenance of biodiversity of benthic flora is also incomplete. Therefore, we started to study benthic diatom assemblages of these very small intermittent streams in the last few years focusing first on the southern hilly part of the region. Our main aim was to investigate the impact of drying up on stream diatom assemblages. We hypothesized the following:
(H1) Assemblages composition – Dry and aquatic phases of streams can be characterized by compositional differences at both taxonomical and trait levels. It is also assumed that indicator taxa could be identified in both phases.
(H2) Biodiversity – Drying of streams coincides with the loss of diversity.
(H3) Diatom-based assessment of water quality – Only slight differences in diatom-based quality are assumed between the different water regimes.
Materials and Methods
Sampling Area and Environmental Parameters
A total of 30 diatom samples were collected from 13 sampling sites on 8 very small intermittent streams in spring and summer in 2019 (Figure 1 and Supplementary Table 1). The streams are located in the southern part of Hungary within the Mecsek Mountains. The local climate is quite special here since Mediterranean impacts can also prevail beside continental ones resulting in a moderately cool and moderately dry climate in the region (Ács et al., 2015). Thus, this area can be characterized by 650–750 mm of mean annual precipitation, 7–10°C of mean annual temperature and more than 2000 h of total annual sunshine duration (Ács et al., 2015; Mezősi, 2017).
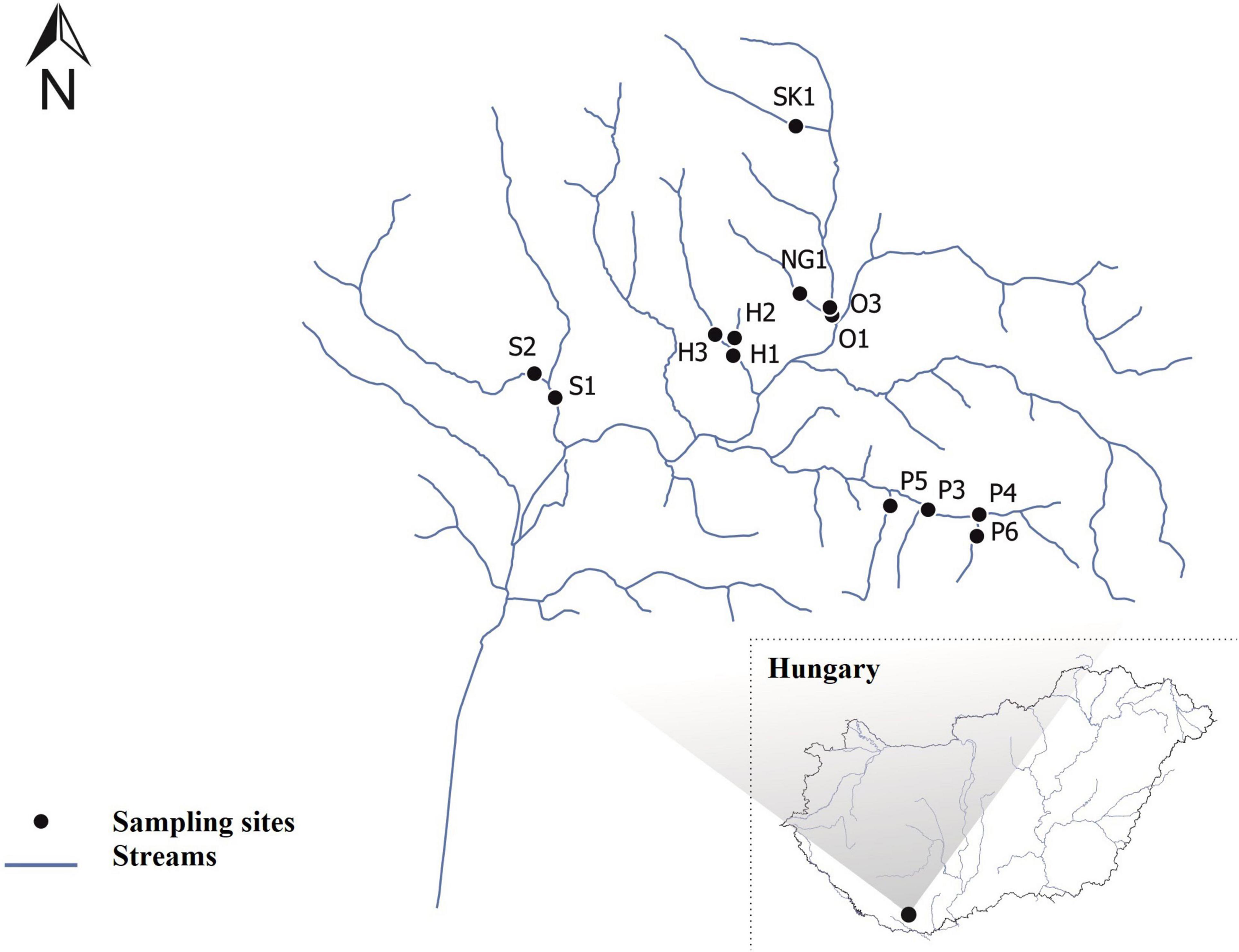
Figure 1. Map of the study area in the Pannonian Ecoregion (Hungary), the watercourses are marked with blue lines. The sampling sites were marked with black circles. Abbreviations of sampling sites are found in Supplementary Table 1.
The catchment areas of each studied stream are less smaller than 10 km2. They are hilly streams, their bedrock is calcareous or red sandstone (Petőczi stream) and the sediment size is coarse to medium. In all but one sampling site (Szentkatalin), the cover of aquatic macrovegetation is also negligible. Most of the streams run through forests; where characteristic tree taxa are mostly beech, oaks, poplars, birch and robinia.
In this study, altogether 11 samples were collected from dry streambed. Hereinafter they will be referred to as the dry phase. There was flowing or standing water in the streambed in 19 sampling events. After we proved that there were no differences in composition and in diversity and also in ecological status indices between these two conditions (Supplementary Table 2; see details in Statistical analyses subchapter) we handled these 19 sampling events together as an aquatic phase.
At sampling sites where surface water could be found in the streambed (aquatic phase), a total of 10 physical and chemical parameters were measured. Water temperature (T - °C), pH and conductivity (COND - μS cm–1) were measured in the field with a portable-multi-parameter digital meter (HQ30d, Germany). The water samples were kept at 4°C in a cooler bag during transportation to the laboratory for further analyses. Nutrient and chloride contents were determined according to international and national standards (NO3–-N in mg L–1 – MSZ 1484-13:2009, 2009; NO2–-N in μg L–1 – MSZ 1484-13:2009, 2009; NH4+-N in μg L–1 – MSZ ISO 7150-1:1992, 1992; PO43–-P in mg L–1 – MSZ EN ISO 6878:2004, 2004 and Cl– in mg L–1 – MSZ 1484-15:2009, 2009). Total suspended solids (TSS in mg L–1 – MSZ 12750-6:1971, 1971) and total dissolved solids (TDS in mg L–1 – Németh, 1998) were measured according to national standard and proposal. Based on the nutrient concentrations and other physical and chemical parameters of the streams, the anthropogenic impact on them was negligible (Supplementary Table 3).
Sample Collection, Preparation and Identification
In flowing conditions, sampling was done according to the European standard (EN 13946, 2014). In pools, when more than one pool was detected, composite samples were collected with the proviso that at least 5 cobbles should be used as substrate per pool. In the dry phase, 5–10 cobbles were sampled from the middle of the streambed. Independently of the aquatic states, when cobbles were not found in the streambed, we used the most characteristic substrates for collecting diatom samples (pebbles, emergent or submerged macrophytes, branches). Substrates were cleaned by toothbrushes and diatom samples were preserved by Lugol’s solution according to the recommendation of European standard (EN 13946, 2014). Diatom valves were prepared by hot hydrogen-peroxide method (EN 13946, 2014) and valves were embedded into Naphrax® (EN 14407, 2014). At least 400 valves were counted and identified using a Leica DMRB microscope (1000–1600-fold magnification). The identification of diatoms was carried out using up-to-date and relevant literatures: Krammer and Lange-Bertalot (1997a,b, 2004a,b), Potapova and Hamilton (2007); Bey and Ector (2013), Stenger-Kovács and Lengyel (2015).
Data Processing and Statistical Analyses
Diatom taxa were classified into 27 trait categories including cell size according to Berthon et al. (2011), guild, life-form and attachment type according to Rimet and Bouchez (2012) and length and width (L/W) ratio according Stenger-Kovács et al. (2018); Supplementary Table 4). We applied moisture preference of taxa proposed by van Dam et al. (1994) updated in 2011 (Supplementary Table 4). Additionally, taxa were referred to as pioneer according to Rimet and Bouchez (2012; Supplementary Table 4). The number of categories within traits were 5, 4, 3, 3, 6, and 5, respectively; Table 1.
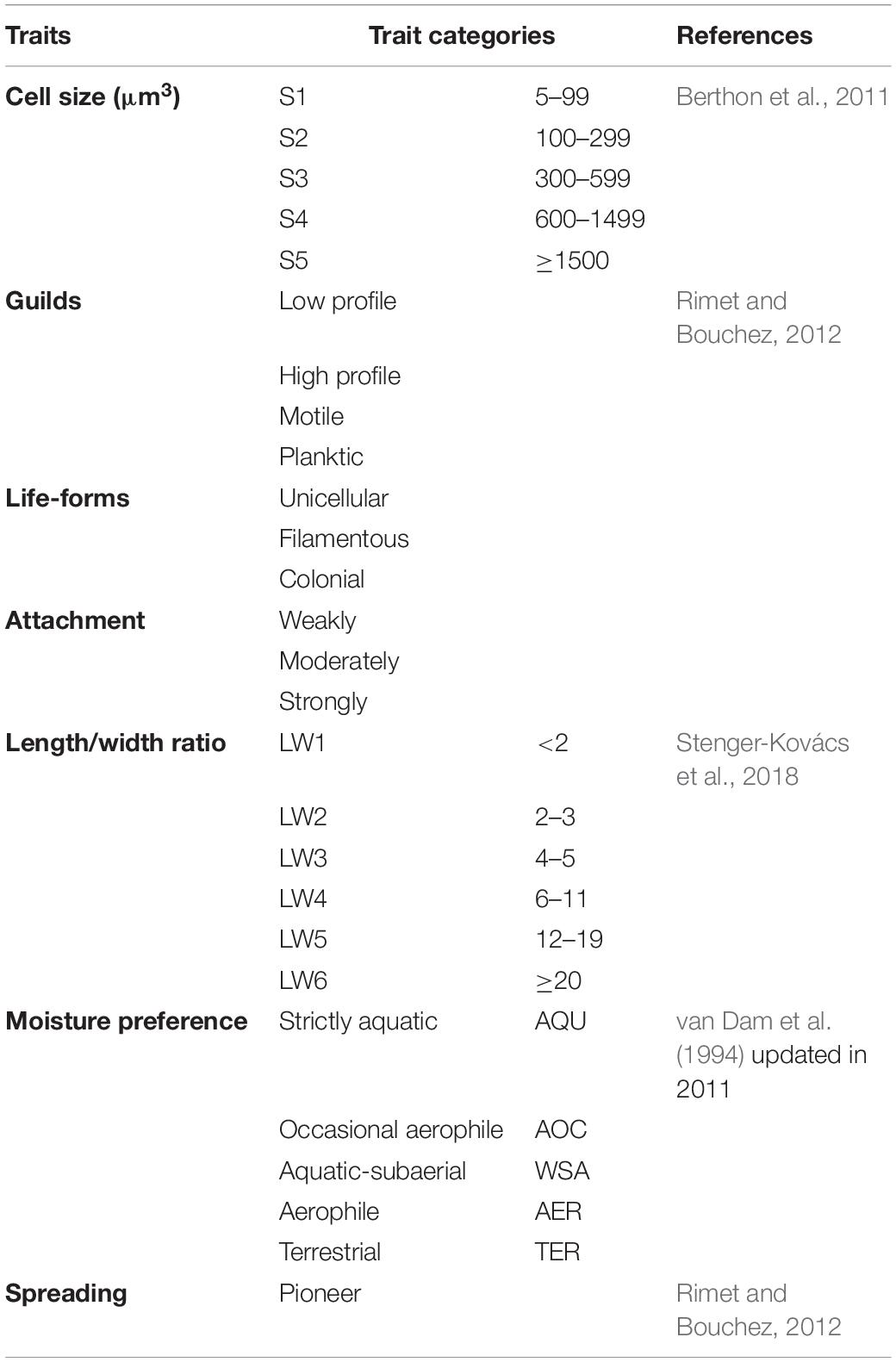
Table 1. Applying functional traits and their categories and key references.
The number of taxa observed in the samples was considered as taxa richness (Taxa_S). Effective Shannon’s H values (Jost, 2006) was calculated to compare the “true diversities” of assemblages (Beck and Schwanghart, 2010; Stuart-Smith et al., 2013; Morris et al., 2014). Effective Shannon’s H is the exponential of Shannon’s H. Taxa richness and Shannon’s H were calculated by using the Past software package (version 4.08; Hammer et al., 2001).
The main components of diversity metrics as functional richness (FRich), functional evenness (FEve), functional divergence (FDiv) and functional dispersion (FDis) were calculated by using “FD” R software package in R (Laliberté and Legendre, 2010; Laliberté et al., 2014; version 3.5.2; R Core Team, 2019).
To identify the indicative species of sites’ groups with similar drying characteristics, we performed indicator value analysis based on relative abundances of taxa (IndVal; Dufrêne and Legendre, 1997). For the calculation of these values, R package “indicspecies” was used (v1.7.9; De Cáceres and Legendre, 2009).
Altogether four indices were calculated to assess the diatom-based ecological quality of streams: Specific pollution sensitivity index (IPS; Coste, 1982), Rott’s trophic index (TID; Rott et al., 1999), Rott’s saprobic index (SID; Rott et al., 1997) and the Hungarian phytobenthos metric (IPSITI; Várbíró et al., 2012). While SID can assess organic load, TID is sensitive for trophic level. The IPS and IPSITI aim to assess both trophic and saprobic states. All of them are based on the weighted average equation of Zelinka and Marvan (1961) modified by Coste (1982).
pj: relative abundance of counting units of “j” taxon in sample.
sj: sensitivity of “j” taxon (optimum).
vj: tolerance/indicator value of “j” taxon.
Diatom indices IPS, SID, and TID were calculated by using OMNIDIA software package (version 5.2; Lecointe et al., 2003), while ISPITI is the arithmetic mean of these three indices. The final values of indices vary between 1 (worse quality) to 20 (best quality).
In the first step, we applied Mann-Whitney test for testing significant differences in diversity and diatom metrics between flowing and standing conditions. We also performed Non-metric Multi-dimensional Scaling (NMDS) with Bray-Curtis distance by applying CANOCO 5.0 software package to compare the taxa and trait composition of streams in flowing and standing aquatic conditions (ter Braak and Šmilauer, 2002). These tests proved that there were no differences in either the studied metrics or the composition of these streams. Thus, as we mentioned above, these two conditions were merged and sampling events were handled as aquatic phase in further analyses.
Comparing the taxa and trait composition in dry and aquatic phases, and also in spring and summer, four NMDSs with Bray-Curtis distance were performed. Here, we used the relative abundances of taxa and the community weighted mean (CWM) matrix, in which the main values of traits in the community were weighted by the relative abundances of the taxa matrix.
One-way ANOVA tests were used to compare the diversity characteristics and also diatom-based water qualities between dry and aquatic phases and also between the two seasons. The fixed factors were the phase-type or the season, while the dependent variables were taxa_S, effective Shannon’s H, FRich, FEve, FDiv, FDis and the diatom indices as IPS, TID, SID, and IPSITI.
Results
Taxa Composition
A total of 208 diatom taxa were identified in the samples: 206 to species and 2 to genus level (Supplementary Table 5).
According to the taxonomy-based NMDS analysis (stress value: 0.089), there was only partial overlap in diatom taxa pools between dry phase and aquatic phase (Figure 2A). While altogether 106 taxa were recorded in both water types, 24 taxa were found exclusively during dry phase and 78 taxa were identified exclusively during aquatic phase (Supplementary Table 5). The different hydrological regime of small streams explained 82.17% of the variance in the taxa composition of benthic diatom assemblages for the first two canonical axes (Figure 2A). The eigenvalues of the first and second axes were 0.577 and 0.245. The diatom taxa Diadesmis contenta, Meridion circulare, Planothidium lanceolatum, Reimeria sinuata and R. uniseriata characterized the assemblages of streams with dry streambed, while Cocconeis species, Navicula and Nitzschia sensu lato species, Gomphonema pumilum and Halamphora species as H. montana and H. normanii characterized the assemblages of streams where water was present (Figure 2A). Diadesmis contenta, Meridion circulare, Navicula veneta, Frustulia vulgaris, Luticola ventricosa and Nitzschia liebetruthii were significant indicators of dry-phase assemblages (p < 0.05), while Gomphonema pumilum, Gomphonema angustatum and Navicula lanceolata were indicative of aquatic phase (p < 0.05, in case of G. pumilum: p < 0.01).
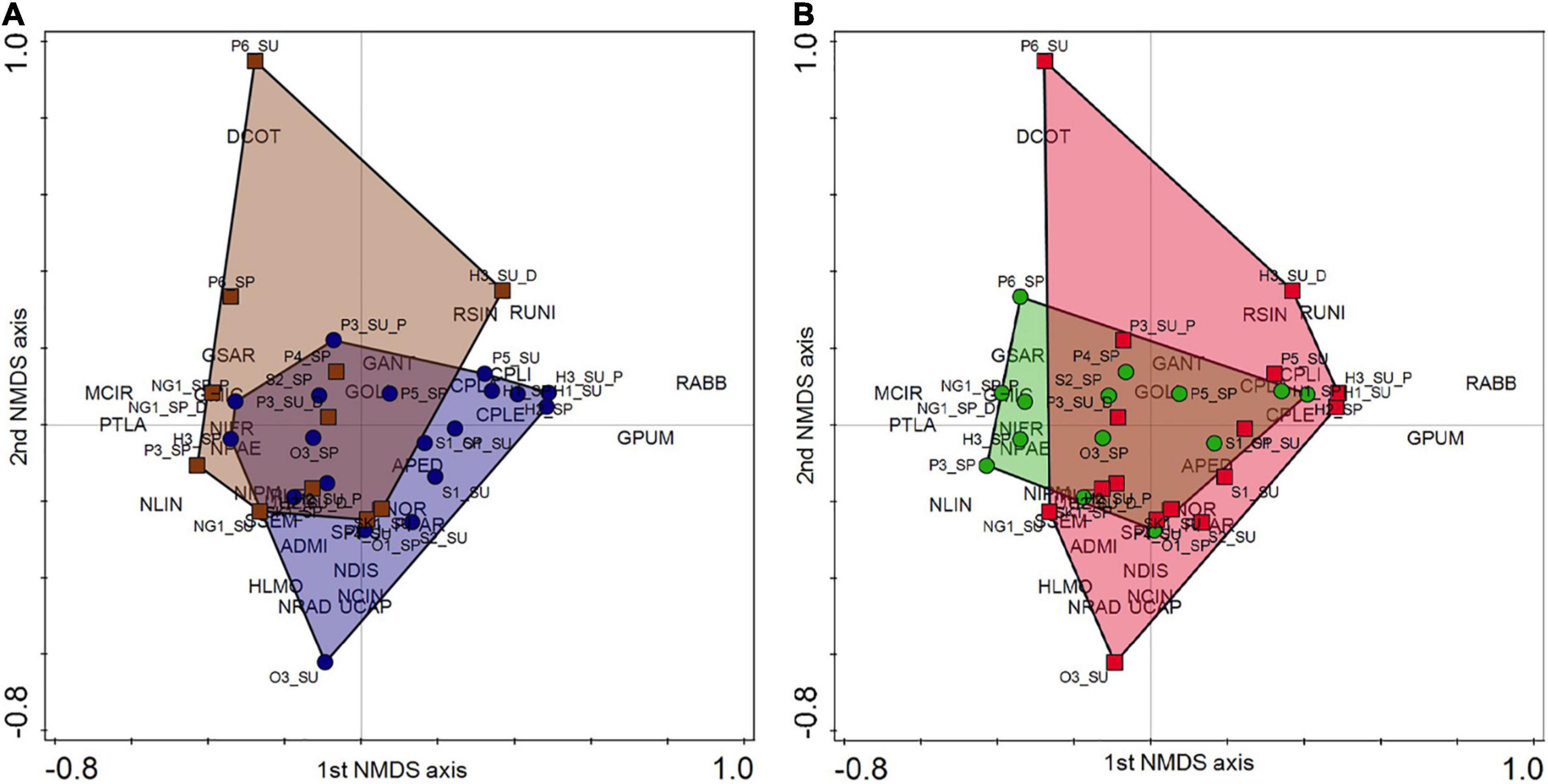
Figure 2. Results of the species-based Non-metric Multi-dimensional Scaling (NMDS) analysis of benthic diatom assemblages. Four letter codes indicate the 208 taxa identified in the samples (see list and codes in Supplementary Table 5). (A) Dry phase is marked with brown squares (their covered area is brown). Aquatic phase is marked with blue circles (their covered area is blue). (B) Spring period is marked with green circles (their covered area is green). Summer period is marked with red squares (their covered area is red).
According to the NMDS analysis, a considerable seasonal overlap in taxa composition was found (Figure 2B). From those taxa, which were recorded in spring, 39 taxa were not found in summer. In contrast, altogether 74 taxa were recorded exclusively in summer. Some of them were dominant in summer such as Cocconeis placentula, Halamphora normanni, Placoneis paraelginensis and Sellaphora pseudopupula.
Trait Composition
Based on the trait composition of diatom assemblages the NMDS (stress value: 0.088) analysis revealed clear differences between dry phase and aquatic phase (Figure 3A). The eigenvalues of NMDS were 0.559 and 0.238, while the hydrological regime and trait composition correlation was 79.74 with the first two canonical axes. The dry phase was characterized by trait categories as high profile guild, S2 size, colonial, moderate or weak attachments and aerophilic, but surprisingly, non-aerophilic category was also characteristic here. In contrast, low profile guild, small (S1) or medium size (S3), unicellular, pioneer and more or less rounded forms (LW1 and LW2) were dominant in aquatic phase. In addition, here, WSA (aquatic-subaerial) and AOC (occasional aerophile) spreading trait groups characterized this phase (Figure 3A).
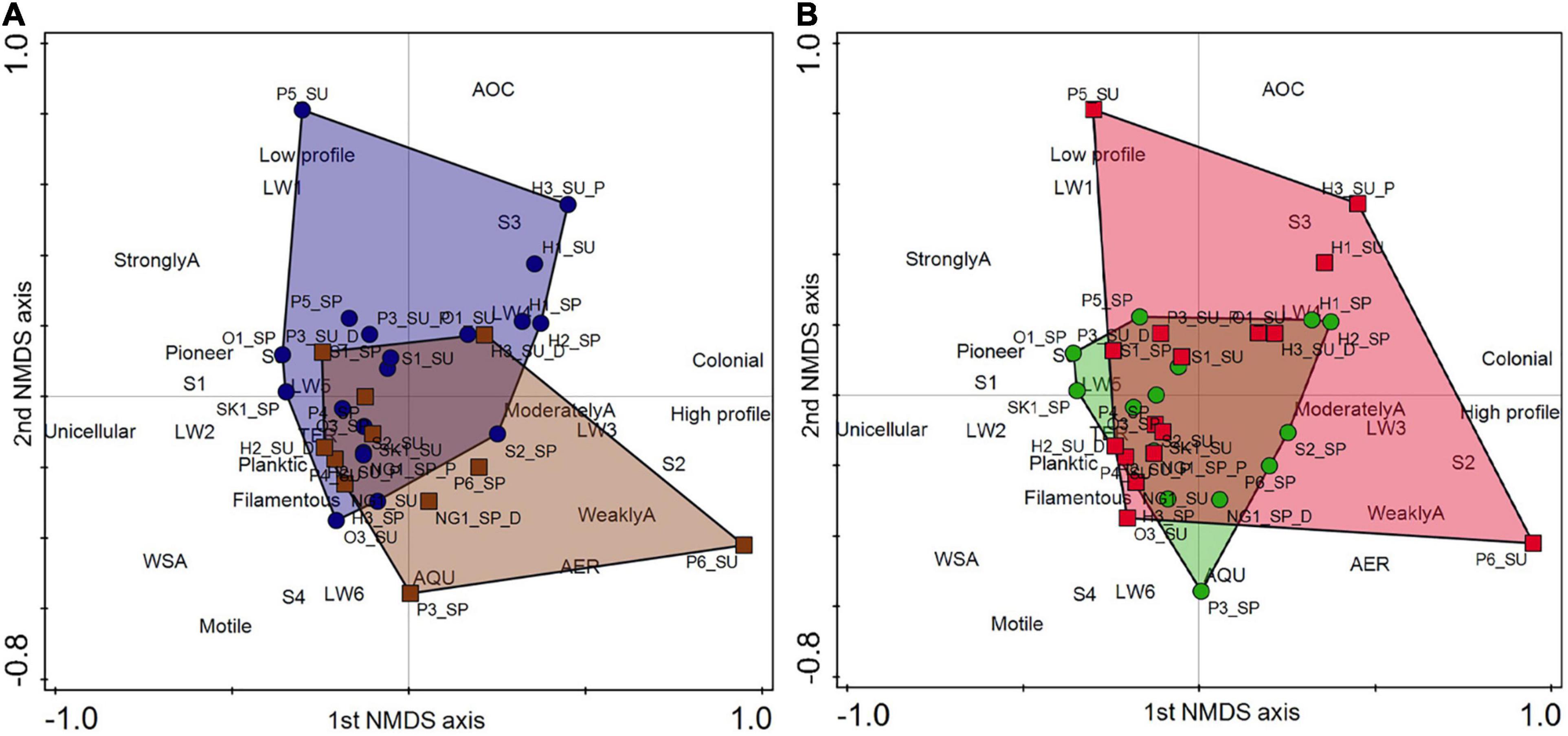
Figure 3. Results of the trait-based Non-metric Multi-dimensional Scaling (NMDS) analysis of the mean trait profiles of diatom assemblages. Trait categories and their abbreviations are found in Table 1. (A) Dry phase is marked with brown squares (their covered area is brown). Aquatic phase is marked with blue circles (their covered area is blue). (B) Spring period is marked with green circles (their covered area is green). Summer period is marked with red squares (their covered area is red).
Similarly to the taxonomy-based analysis, a less pronounced difference in trait composition was found between spring and summer (Figure 3B). All trait categories were found in both seasons, only their proportion was different. In summer, the following trait categories characterized the assemblages: high profile guild and/or S2 size and/or colonial category and/or low profile guild and/or LW1 category.
Biodiversity and Diatom Indices
Although both taxa and trait composition of benthic assemblages of the two phases clearly separated, hydrological regimes studied here did not result in significant differences either in the taxonomy-, or in the trait-based diversity metrics (p > 0.05; Figure 4). Despite the seasonal overlap in taxa and trait composition, the FDis was reduced significantly by summer (p = 0.012). The other diversity metrics, however, did not show seasonal differences (p > 0.05; Supplementary Figure 1).
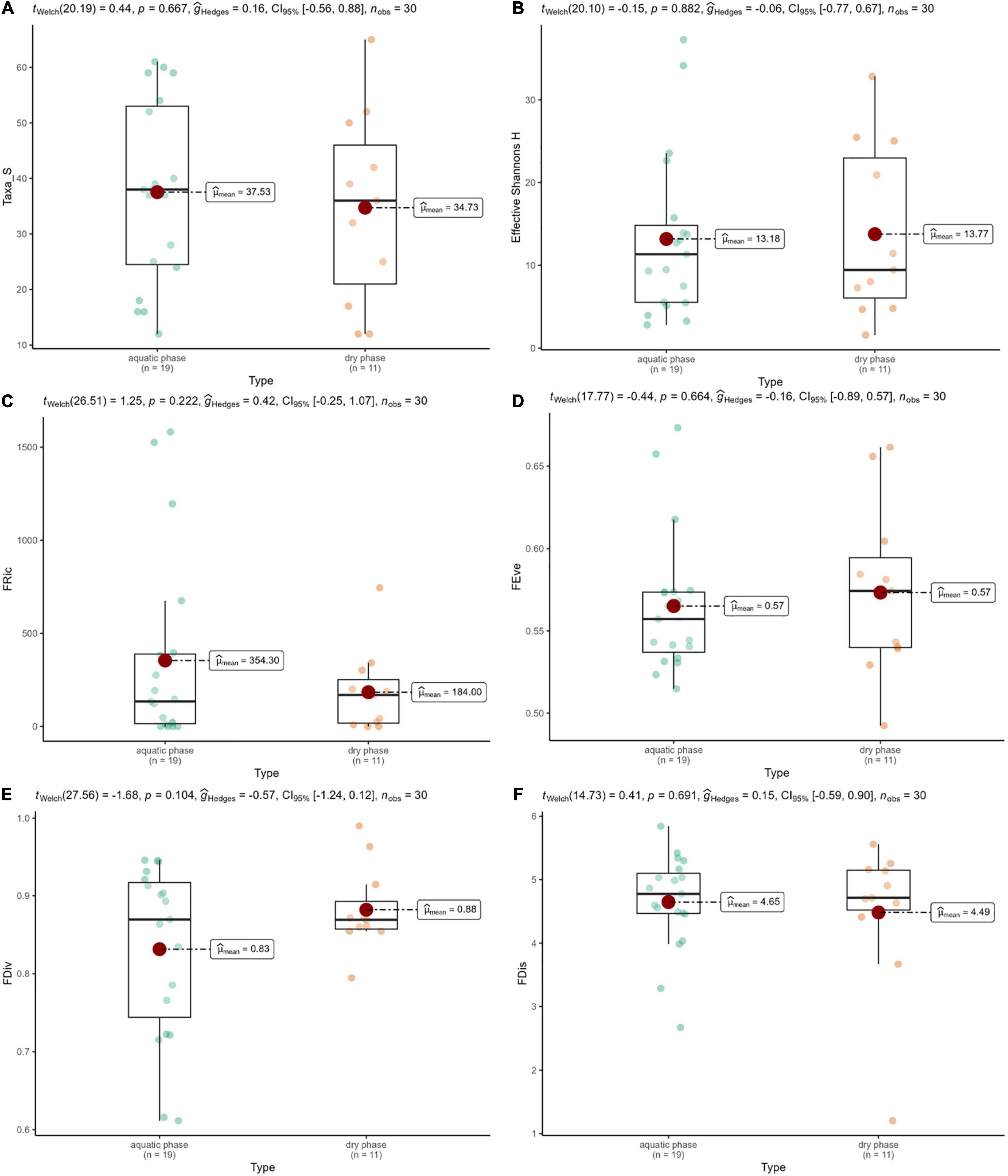
Figure 4. Taxa_S (A), effective Shannon’s H (B), functional richness (C), functional evenness (D), functional divergence (E), and functional dispersion (F) in the dry and aquatic phases. Results of ANOVA tests: p > 0.05 in all cases.
In the case of TID index, we found strong positive correlation between diatom-based ecological quality and hydrological regime: it was significantly lower in streams with dry streambed than in aquatic phase (p = 0.0122; Figure 5B). Overall, neither hydrological regime nor seasonal changes resulted in significant differences in the other indices (p > 0.05; Figure 5 and Supplementary Figure 2).
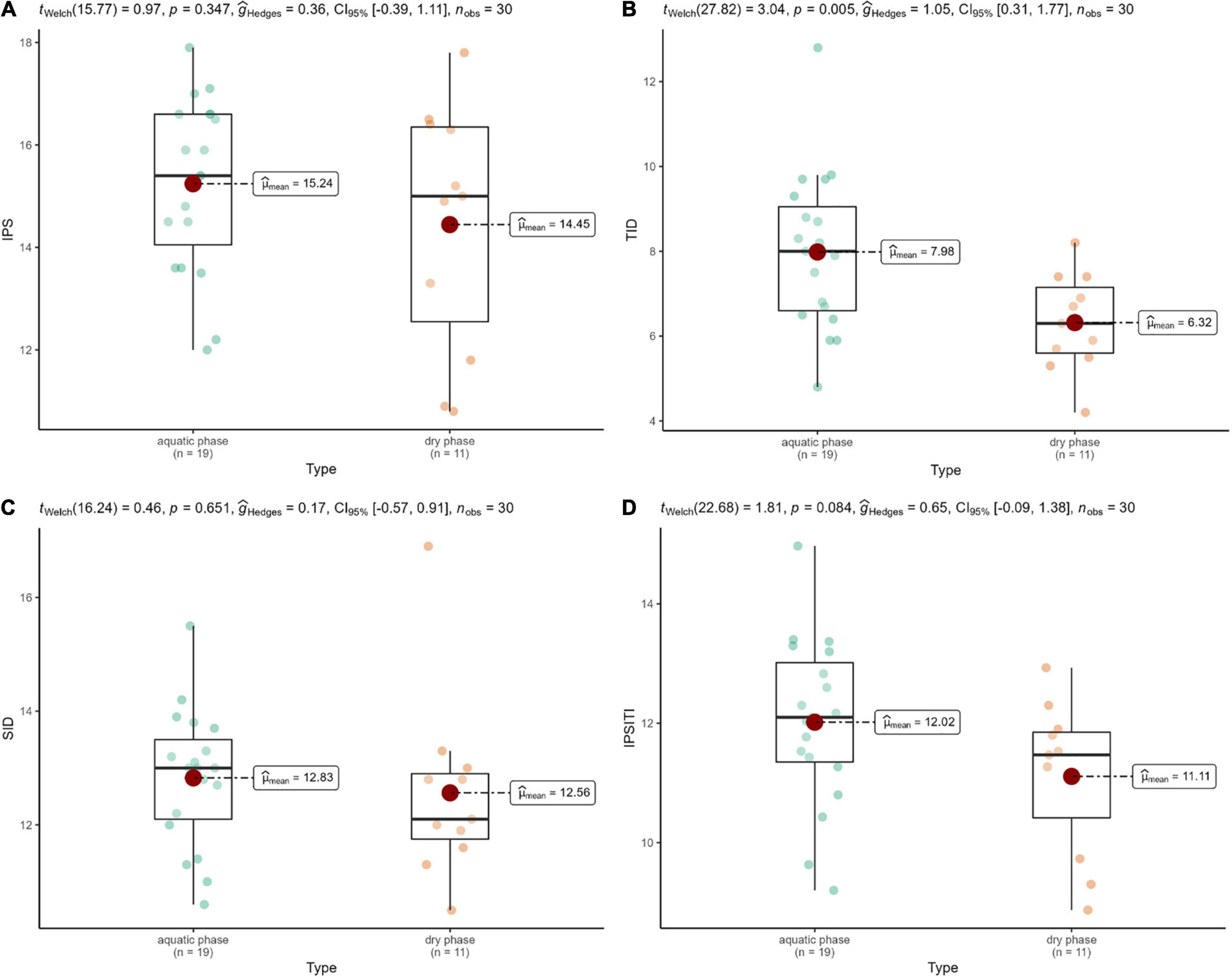
Figure 5. IPS (A), TID (B), SID (C), and IPSITI (D) diatom indices in the dry and aquatic phases. Results of ANOVA tests: p = 0.005 in the case of TID, p > 0.05 in the other cases.
Discussion
Compositional Changes
Intermittent small streams provide a variety of services to human communities and they contribute to the integrity of river networks (see more in Magand et al., 2020). The assemblages living here are special compared to permanent flowing waters, since lentic and terrestrial species appear regularly beside lotic taxa during intermittence (Stubbington et al., 2017). Additionally, intermittence requires special adaptation strategies, thus, functional composition of diatom assemblages continuously changes during drying up of streams (Sabater et al., 2017).
Here, we hypothesized that there would be clear compositional differences between streams with dry streambed (dry phase) and streams in aquatic phase both in terms of taxonomical and trait levels. We also assumed that indicator taxa could be identified in both phases. The results supported our hypotheses. We found clear, hydrological regime induced compositional differences both at taxonomy and trait levels and indicative species could be recorded too. Periphytic algae, including diatoms sensitively indicated the intermittence independently of the ecoregion (Novais et al., 2014, 2020; B-Béres et al., 2016, 2019; Falasco et al., 2016a; Várbíró et al., 2020; Lukács et al., 2021).
In this study, mainly colonial taxa belonging to high profile guild dominated the dry phase (but see Meridion circulare – low profile guild taxa). Current evidences about the dominance of high profile guild and colonial forms in intermittent streams are inconsistent among studies (Falasco et al., 2016a; Novais et al., 2020; Várbíró et al., 2020). These traits can support survival in permanent waters, where obviously the environmental conditions, especially the water regime, are more stable than in intermittent streams (Várbíró et al., 2020). Novais et al. (2020) also found, that the relative abundances of the high profile guild are higher during aquatic (flow) phase than in dry phase.
In contrast to high profile guild, colonial lifestyle, especially in case of those colonial taxa which form mucous tube can be advantageous in intermittent streams (Falasco et al., 2016a) and during dry phase (Sabater et al., 2017). In our study, mucilage-forming taxa were not dominant during the dry phase, only non-tube forming colonial taxa such as Diadesmis contenta. Although Frustulia vulgaris, which forms a mucous tube, was one of the indicator species of dry phase, this species was never dominant in samples.
Beside the above mentioned taxa, aerophilous and/or motile species were revealed as indicative species in dry phase. Appearance of aerophilous taxa in intermittent streams is a well-known phenomenon (Novais et al., 2014; Falasco et al., 2016a; Sabater et al., 2016, 2017; B-Béres et al., 2019). Similar to high profile guild and colonial taxa, it is not easy to ascribe the members of motile guild into a water regime phase or type, i.e., they are diverse in their habitat preferences with respect to water phase. Motile taxa can be considered as physical disturbance indicators, since their dominance within assemblages usually increases with siltation and in depositional habitats (Falasco et al., 2016b; Novais et al., 2020), which are common during intermittence. Additionally, motile taxa are able to migrate from the sediment surface to the deeper, wet layers during drying. After rewetting, they return to the surface and colonize it relatively fast (McKew et al., 2011). Our previous studies, however, pointed out that intermittence is a strong selective pressure for motile taxa independently from the ecoregions (Várbíró et al., 2020) and this guild is characteristic for permanent streams in the Pannonian Ecoregion (B-Béres et al., 2019). Here, indicator species of dry phase belonging to motile guild were small sized and aerophilous or subaerial taxa. These characteristics together and their additive ecological benefits made survival possible for diatoms bearing these traits.
A further indicator species was the low profile and colonial Meridion circulare during dry phase, which was often dominant in streams with dry streambed (max. 36%). This species is usually found in small streams in winter and spring (Stenger-Kovács et al., 2013). In this study, M. circulare was more abundant in spring than in summer. But this species, as a member of the low profile guild, tolerates the physical disturbances which can be characteristic during intermittence. We believe these two characters (favor low temperature and tolerate disturbances) lead to the high number of M. circulare in streams with dry streambed in spring. In summer, its relative abundance decreased in the dry phase too, pointing out a more significant temperature-dependent than intermittence-dependent appearance of this taxon.
In this study, streams in aquatic phase were dominated mostly by pioneer category (e.g., Achnanthidium minutissimum and Amphora pediculus) and/or low profile guild, small and medium sized, unicellular taxa (e.g., Planothidium lanceolatum, Reimeria sinuata, and R. uniseriata) belonging usually to WSA or AOC spreading trait groups. It is a well-known phenomenon that physical disturbance is indicated by small sized, unicellular, low profile taxa (Berthon et al., 2011; B-Béres et al., 2019; Novais et al., 2020). In intermittent streams right after the rewetting of the streambed, these traits, i.e., small size, low profile guild, unicellular and/or pioneer, support the fast spreading of species and colonization of habitats. In our study, the relative lack of aquatic macrovegetation can enhance the chance for spreading of species bearing these traits.
Moisture preference should be an important trait in intermittence studies, since it clearly refers to the tolerance of water shortage. However, Novais et al. (2020) pointed out that most taxa they found in Portuguese streams do not have clear moisture preference. Here, more than 71% of the taxa related to WSA or AOC moisture categories meaning that these taxa are able to tolerate aerophilic conditions for a while. These taxa were characteristic in aquatic phase, while true aerophilic taxa preferred dry phase, as we mentioned above.
Mostly unicellular, stalk-forming Gomphonema species were indicative of aquatic phase. Long peduncle i.e., stalk provides an opportunity for taxa to live in the upper layers of biofilm. This life-form subserves an efficient access to light and nutrients but it exposes taxa to high flow disturbances (Passy, 2007). Here, most of the studied streams run through forests. Thus, there is no significant anthropogenic impact on them and their nutrient content is relatively low. In addition, flow conditions in the aquatic phase are comparatively stable. These circumstances, as we mentioned above, favor stalk-forming taxa.
Biodiversity Changes
We hypothesized that biodiversity would be reduced in the dry phase. The results, however, did not confirm our assumption. Neither taxonomical nor functional diversity metrics changed significantly due to drying. These results suggest that taxonomical and functional redundancies were able to compensate for biodiversity loss. Although these results are in accordance with observations of Novais et al. (2020), they contradict the very recent results of studies by Crabot et al. (2021) and Tornés et al. (2021). Crabot et al. (2021) and Tornés et al. (2021) highlighted that functional redundancy decreases with flow intermittence. But this decrease is time-dependent, the longer the dry period the smaller the protective effect of the functional redundancy on assemblages. In our study, the duration of the dry period of streams was very variable (from 7 to 266 days), however, in most cases, it was lower than 50 days, which is lower than the threshold of duration resulting in diversity reduction in the paper of Tornés et al. (2021). They found a significant decrease in biodiversity only after 50 days. Crabot et al. (2021) also noted that any 10% increase in flow intermittence results in a 5% decrease in functional redundancy. In our case, the annual flow intermittence does not reach the 10% in most cases.
Independently of the water regime, a slight decrease in FDis was revealed in summer. The FDis is strongly connected to the dominance of the trait values (Laliberté and Legendre, 2010). Because other metrics like FDiv and FRich did not change significantly, decrease in FDis did not reveal a shift of assemblages’ composition to extreme traits, and traits did not disappear, only their proportions changed. This was probably due to the biotic interactions that might be stronger during summer. Since proportion of other algal groups, e.g., green algae, cyanobacteria, usually increases in summer (Biggs and Stokseth, 1996; Suren et al., 2003) resulting in an enhanced resource competition and thus, a biotic pressure on diatoms.
Changes in Diatom Indices
Almost 10,000 watercourses are registered in Hungary (Stubbington et al., 2018), but less than 10% of them are monitored regularly2. This means that there is no information about the risk of vulnerability or the risk of intermittence as well as the ecological quality for more than 9000 small Hungarian watercourses. These ecosystems, however, are strongly endangered both by anthropogenic and natural impacts including climatic extremes. To the best of our knowledge, this paper is the first where diatom-based ecological quality of very small intermittent hilly streams were studied in the Pannonian Ecoregion. Since there is no diatom index for evaluating the status of these streams, altogether four diatom indices regularly used in WFD-based biomonitoring in Hungary (Várbíró et al., 2012) were analyzed here. Based on former studies by Falasco et al. (2016b) and by Novais et al. (2020), only slight differences in diatom indices were hypothesized between aquatic and dry phases. The results supported our hypothesis, since drying up of streams had significant negative influence only on TID, while water regime had no impact on the other three diatom indices. The stability of the studied indices points out their applicability in ecological status assessment of intermittent streams. However, it has to be highlighted that much more sampling sites including permanent ones as well as differently loaded sites should be involved in the analyses. Our study is only the first step in this way, but the results are encouraging. These diatom indices were developed mostly for assessing anthropogenic impact on permanent waters with larger catchment areas than the waterbodies studied here. Nevertheless, our results suggest that these indices are stable enough to assess the status of very small permanent streams even in dry phase. We think that the higher values of TID in the aquatic phase were attributable to the higher proportion of small, pioneer taxa considered as indicators of good or excellent waters. These taxa, as we mentioned above, are good colonizers and tolerate physical disturbances caused by rewetting. Analyzing this diatom index together with the others can be a good way to indicate both nutrient supply and climate-induced influences on benthic diatom assemblages in small intermittent streams in the Pannonian Ecoregion.
Conclusion
Here, we examined the influence of flow intermittence on benthic diatom communities in very small hilly streams (catchment area <10 km2) in the Pannonian Ecoregion. While the results revealed strong water regime dependent differences in taxa and trait composition of assemblages, no seasonal separation was found. However, our findings have highlighted that short-term drying up of streams might not necessarily cause biodiversity loss. It seems that taxonomical and functional redundancy could prevail. In contrast, decrease in functional dispersion in summer suggests proportional changes in trait values. But it has to be stressed that traits did not disappear from the system, and assemblages were not pushed to dominance of extreme traits. In this study, we first analyzed the usability of diatom-based quality indices in the assessment of drying up of small streams in this region. We showed that these metrics are useful tools for ecological evaluation of streams even in the dry phase. As we noted, these streams are strongly endangered to become episodic. However, it is not a simple shift from permanence to intermittency, the lengthening of the dry-phase in the future might result in irreversible compositional changes and significant diversity loss, threating the entire aquatic ecosystem not just benthic algae. Thus, any knowledge, which supports their ecological role in this region, can help to save them and their valuable microflora.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author Contributions
VB-B: conceptualization, data curation, project administration, supervision, and roles/writing – original draft. ZK, GV, GM, and VB-B: formal analysis and visualization. VB-B, PB, and ZC: funding acquisition. ZK, VB-B, IB, PB, and BP: investigation. VB-B, IB, and GV: methodology. PB, GB, ZC, and VB-B: resources. VB-B, PB, ZC, BP, GB, GV, and IB: writing – review and editing. All authors contributed to the article and approved the submitted version.
Funding
This project was financially supported by the KEHOP-1.1.0-15-2016-0002 project and by the National Research, Development and Innovation Office – NKFIH FK 132 142 (VB-B) and FK 135 136 (PB) grants, by the János Bolyai Research Scholarship of the Hungarian Academy of Sciences BO-00458-20-8 (VB-B); BO-00106-21-8 (PB) and by the ÚNKP-20-5 (VB-B) and ÚNKP-21-5 (VB-B and PB) New National Excellence Program of the Ministry for Innovation and Technology from the Source of the National Research, Development and Innovation Fund. ZC and BP were supported by the DRYvER project (http://www.dryver.eu/), which has received funding from the European Union’s Horizon 2020 Research and Innovation Programme under grant agreement No. 869226.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2022.834548/full#supplementary-material
Footnotes
References
Ács, F., Breuer, H., and Skarbit, N. (2015). Climate of Hungary in the twentieth century according to Feddema. Theor. Appl. Climatol. 119, 161–169. doi: 10.1007/s00704-014-1103-5
Acuña, V., Hunter, M., and Ruhi, A. (2017). Managing temporary streams and rivers as unique rather than second-class ecosystems. Biol. Conserv. 211, 12–19. doi: 10.1016/j.biocon.2016.12.025
Barthès, A., Leflaive, J., Coulon, S., Peres, F., Rols, J. L., and Ten-Hage, L. (2015). Impact of drought on diatom communities and the consequences for the use of diatom index values in the River Maureillas (Pyrénées-Orientales, France). River Res. Appl. 31, 993–1002. doi: 10.1002/rra.2793
B-Béres, V., Lukács, Á, Török, P., Kókai, Zs, Novák, Z., T-Krasznai, E., et al. (2016). Combined eco-morphological functional groups are reliable indicators of colonization processes of benthic diatom assemblages in a lowland stream. Ecol. Indic. 64, 31–38. doi: 10.1016/j.ecolind.2015.12.031
B-Béres, V., Tóthmérész, B., Bácsi, I., Borics, G., Abonyi, A., Tapolczai, K., et al. (2019). Autumn drought drives functional diversity of benthic diatom assemblages of continental streams. Adv. Water Resour. 126, 129–136. doi: 10.1016/j.advwatres.2019.02.010
Beck, J., and Schwanghart, W. (2010). Comparing measures of species diversity from incomplete inventories: an update. Methods Ecol. Evol. 1, 38–44. doi: 10.1111/j.2041-210X.2009.00003.x
Berthon, V., Bouchez, A., and Rimet, F. (2011). Using diatom life forms and ecological guilds to assess organic pollution and trophic level in rivers: a case study of rivers in south-eastern France. Hydrobiologia 673, 259–271. doi: 10.1007/s10750-011-0786-1
Bey, M. Y., and Ector, L. (2013). Atlas des Diatomées des Cours d’eau de la Région Rhône-Alpes. Available online at: http://www.auvergne-rhone-alpes.developpement-durable.gouv.fr/atlas-des-diatomees-a3480.html (accessed November 11, 2015).
Biggs, B. J. F., and Stokseth, S. (1996). Hydraulic habitat suitability for periphyton in rivers. Regul. Rivers Res. Manage. 12, 251–261. doi: 10.1002/(SICI)1099-1646(199603)12:2/3<251
Coste, M. (1982). Étude des Méthodes Biologiques D’appréciation Quantitative de la Qualité des Eaux. Lyon: CEMAGREF Division Qualité des Eaux, Agence de L’eau Rhone-Méditerranée-Corse.
Crabot, J., Mondy, C. P., Usseglio-Polatera, P., Fritz, K. M., Wood, P. J., Greenwood, M. J., et al. (2021). A global perspective on the functional responses of stream communities to flow intermittence. Ecography 44, 1511–1523. doi: 10.1111/ecog.05697
De Cáceres, M., and Legendre, P. (2009). Associations between species and groups of sites: indices and statistical inference. Ecology 90, 3566–3574. doi: 10.1890/08-1823.1
Díaz, S., Fargione, J., Chapin, F. S. III, and Tilman, D. (2006). Biodiversity loss threatens human well-being. PLoS Biol. 4:e277. doi: 10.1371/journal.pbio.0040277
Dudgeon, D., Arthington, A. H., Gessner, M. O., Kawabata, Z., Knowler, D. J., and Lévêque, C. (2006). Freshwater biodiversity: importance, threats, status and conservation challenges. Biol. Rev. 81, 163–182. doi: 10.1017/S1464793105006950
Dufrêne, M., and Legendre, P. (1997). Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecol. Monogr. 67, 345–366. doi: 10.2307/2963459
EC (2000). Directive 2000/60/EC of the European Parliament and of the Council of 23rd October 2000 Establishing a Framework for Community Action in the Field of Water Policy. Official Journal of the European Communities, 22 December, L 327/1. Brussels: European Commission.
EN 13946 (2014). Water Quality. Guidance Standard for the Routine Sampling and Pretreatment of Benthic Diatoms from Rivers. Brussels: European commission for standardization.
EN 14407 (2014). Water Quality. Guidance Standard for the Identification, Enumeration and Interpretation of Benthic Diatom Samples From Running Waters. Brussels: European commission for standardization.
Falasco, E., Piano, E., and Bona, F. (2016a). Diatom flora in Mediterranean streams: flow intermittency threatens endangered species. Biodivers. Conserv. 25, 2965–2986. doi: 10.1007/s10531-016-1213-8
Falasco, E., Piano, E., and Bona, F. (2016b). Suggestions for diatom-based monitoring in intermittent streams. Knowl. Manag. Aquat. Ecosyst. 417:38. doi: 10.1051/kmae/2016025
Hammer, Ø, Harper, D. A. T., and Ryan, P. D. (2001). PAST: paleontological statistics software package for education and data analysis. Palaeontol. Electron. 4:9.
He, F., Zarfl, C., Bremerich, V., David, J. N. W., Hogan, Z., Kalinkat, G., et al. (2019). The global decline of freshwater megafauna. Glob. Change Biol. 25, 3883–3892. doi: 10.1111/gcb.14753
IPCC (2021). Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press.
Kireta, A. R., Reavie, E. D., Sgro, G. V., Angradi, T. R., Bolgrien, D. W., Hill, B. H., et al. (2012). Planktonic and periphytic diatoms as indicators of stress on great rivers of the United States: testing water quality and disturbance models. Ecol. Indic. 13, 222–231. doi: 10.1016/j.ecolind.2011.06.006
Krammer, K., and Lange-Bertalot, H. (1997a). “Bacillariophyceae 1., Naviculaceae,” in Süsswasserflora Von Mitteleuropa, eds H. Gerloff, J. H. Heynig, and D. Mollenhauer (Heidelberg: Elsevier).
Krammer, K., and Lange-Bertalot, H. (1997b). “Bacillariophyceae 2., Bacillariaceae, Epithemiaceae, Surirellaceae,” in Süsswasserflora Von Mitteleuropa, eds H. Gerloff, J. H. Heynig, and D. Mollenhauer (Heidelberg: Elsevier).
Krammer, K., and Lange-Bertalot, H. (2004a). “Bacillariophyceae 3., Centrales, Fragilariaceae, Eunotiaceae,” in Süsswasserflora Von Mitteleuropa, eds H. Gerloff, J. H. Heynig, and D. Mollenhauer (Heidelberg: Spektrum Akademischer Verlag).
Krammer, K., and Lange-Bertalot, H. (2004b). “Bacillariophyceae 4., Achnanthaceae. kritische erganzungen zu Achnanthes s. l., Navicula s. str., Gomphonema. gesamt literaturverzeichnis teil 1–4,” in Süsswasserflora Von Mitteleuropa, eds H. Gerloff, J. H. Heynig, and D. Mollenhauer (Heidelberg: Spektrum Akademischer Verlag).
Laliberté, E., and Legendre, P. (2010). A distance-based framework for measuring functional diversity from multiple traits. Ecology 91, 299–305. doi: 10.1890/08-2244.1
Laliberté, E., Legendre, P., and Shipley, B. (2014). FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology. R Package Version 1.0-12. Vienna: R Foundation for Statistical Computing.
Lecointe, C., Coste, M., and Prygiel, J. (2003). Omnidia 3.2 Diatom Index Software including diatom database with taxonomic names, reference and codes of 11643 diatom taxa. Hydrobiologia 269, 509–513.
Lukács, Á, Bácsi, I., Nemes-Kókai, Zs, Borics, G., Várbíró, G., T-Krasznai, E., et al. (2021). Strong influence of climatic extremes on diversity of benthic algae and cyanobacteria in a lowland intermittent stream. Ecohydrology 14:e2286. doi: 10.1002/eco.2286
Magand, C., Alves, M. H., Calleja, E., Datry, T., Dörflinger, G., England, J., et al. (2020). Intermittent Rivers and Ephemeral Streams: What Water Managers Need to Know. Technical Report – Cost ACTION CA 15113. Brussels: European Cooperation in Science and Technology, doi: 10.5281/zenodo.3888474
McCormick, P. V., and Cairns, J. (1994). Algae as indicators of environmental change. J. Appl. Phycol. 6, 509–526. doi: 10.1007/BF02182405
McKew, B. A., Taylor, J. D., McGenity, T. J., and Underwood, J. C. G. (2011). Resistance and resilience of benthic biofilm communities from a temperate saltmarsh to desiccation and rewetting. ISME J. 5, 30–41. doi: 10.1038/ismej.2010.91
Messager, M. L., Lehner, B., Cockburn, C., Lamouroux, N., Pella, H., Snelder, T., et al. (2021). Global prevalence of non-perennial rivers and streams. Nature 594, 391–397. doi: 10.1038/s41586-021-03565-52021
Mezősi, G. (2017). “Climate of hungary,” in The Physical Geography of Hungary. Geography of the Physical Environment, ed. G. Mezõsi (Cham: Springer), 101–119. doi: 10.1007/978-3-319-45183-1_2
Morris, E. K., Caruso, T., Buscot, F., Fischer, M., Hancock, C., Maier, T. S., et al. (2014). Choosing and using diversity indices: insights for ecological applications from the German Biodiversity Exploratories. Ecol. Evol. 4, 3514–3524. doi: 10.1002/ece3.1155
MSZ 12750-6:1971 (1971). Testing of SurfaceWaters. Determination of All Solved and Floating Matter Content (in Hungarian). Budapest: Magyar Szabványügyi Hivatal.
MSZ 1484-13:2009 (2009). Water Quality. Part 12: Determination of Nitrate and Nitrite. Content by Spectrophotometric Method. (in Hungarian). Budapest: Magyar Szabványügyi Hivatal.
MSZ 1484-15:2009 (2009). Water Quality. Part 15: Determination of Chloride Content by Argentometric Titration Method. (in Hungarian). Budapest: Magyar Szabványügyi Hivatal.
MSZ EN ISO 6878:2004 (2004). Water Quality. Determination of Phosphorus. Ammonium Molybdate Spectrometric Method. (in Hungarian). Budapest: Magyar Szabványügyi Hivatal.
MSZ ISO 7150-1:1992 (1992). Water Quality. Determination of Ammonium. Part 1: Manual Spectrophotometric Method. (in Hungarian). Budapest: Magyar Szabványügyi Hivatal.
Németh, J. (1998). A Biológiai Vízminõsítés Módszerei. Vízi Természet- és Környezetvédelem, Vol. 7. Budapest: Környezetgazdálkodási Intézet.
Novais, M. H., Morais, M. M., Rosado, J., Dias, L. S., Hoffmann, L., and Ector, L. (2014). Diatoms of temporary and permanent watercourses in Southern Europe (Portugal). River Res. Appl. 30, 1216–1232. doi: 10.1002/rra.2818
Novais, M. H., Morales, E. A., Penha, A. M., Potes, M., Bouchez, A., Barthes, A., et al. (2020). Benthic diatom community dynamics in Mediterranean intermittent streams: effects of water availability and their potential as indicators of dry-phase ecological status. Sci. Total Environ. 719:137462. doi: 10.1016/j.scitotenv.2020.137462
Pan, Y. D., Stevenson, R. J., Hill, B. H., Kaufmann, P. R., and Herlihy, A. T. (1999). Spatial patterns and ecological determinants of benthic algal assemblages in Mid-Atlantic streams, USA. J. Phycol. 35, 460–468. doi: 10.1046/j.1529-8817.1999.3530460.x
Passy, S. I. (2007). Diatom ecological guilds display distinct and predictable behavior along nutrient and disturbance gradients in running waters. Aquat. Bot. 86, 171–178. doi: 10.1016/j.aquabot.2006.09.018
Piano, E., Falasco, E., and Bona, F. (2017). How does water scarcity affect spatial and temporal patterns of diatom community assemblages in Mediterranean streams? Freshw. Biol. 62, 1276–1287. doi: 10.1111/fwb.12944
Potapova, M., and Hamilton, P. B. (2007). Morphological and ecological variation within the Achnanthidium minutissimum (Bacillariophyceae) species complex. J. Phycol. 43, 561–575. doi: 10.1111/j.1529-8817.2007.00332.x
R Core Team (2019). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Reid, A. J., Carlson, A. K., Creed, I. F., Eliason, E. J., Gell, P. A., Johnson, P. T. J., et al. (2019). Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 94, 849–873. doi: 10.1111/brv.12480
Rimet, F., and Bouchez, A. (2012). Life-forms, cell sizes and ecological guilds of diatoms in European rivers. Knowl. Manag. Aquat. Ecosyst. 406, 1283–1299. doi: 10.1051/kmae/2012018
Rott, E., Binder, N., Van Dam, H., Ortler, K., Pall, K., Pfister, P., et al. (1999). Indikationslisten für Aufwuchsalgen. Teil 2: Trophieindikation und Autökologische Anmerkungen. Wien: Bundesministerium für Land- und Forstwirtschaft.
Rott, E., Hofmann, G., Pall, K., Pfister, P., and Pipp, E. (1997). Indikationslisten für Aufwuchsalgen Teil 1: Saprobielle Indikation. Wien: Bundesministerium für Land- und Fortwirtschaft.
Sabater, S., Timoner, X., Bornette, G., De Wilde, M., Stromberg, J. C., and Stella, J. C. (2017). “The biota of intermittent rivers and ephemeral streams: algae and vascular plants,” in Intermittent Rivers and Ephemeral Streams, Ecology and Management, eds T. Datry, N. Bonada, and N. Boulton (Cambridge, MA: Academic Press), 189–216. doi: 10.1016/B978-0-12-803835-2.00016-4
Sabater, S., Timoner, X., Borrego, C., and Acuña, V. (2016). Stream biofilm responses to flow intermittency: from cells to ecosystems. Front. Environ. Sci. 4:14. doi: 10.3389/fenvs.2016.00014
Stenger-Kovács, Cs, and Lengyel, E. (2015). Taxonomical and distribution guide of diatoms in soda pans of Central Europe. Studia Bot. Hung. 46, 3–203. doi: 10.17110/StudBot.2015.46.Suppl.3
Stenger-Kovács, Cs, Körmendi, K., Lengyel, E., Abonyi, A., Hajnal, É, Szabó, B., et al. (2018). Expanding the trait-based concept of benthic diatoms: development of trait- and species-based indices for conductivity as the master variable of ecological status in continental saline lakes. Ecol. Indic. 95, 63–74. doi: 10.1016/j.ecolind.2018.07.026
Stenger-Kovács, Cs, Lengyel, E., Crossetti, L. O., Üveges, V., and Padisák, J. (2013). Diatom ecological guilds as indicators of temporally changing stressors and disturbances in the small Torna-stream, Hungary. Ecol. Indic. 24, 138–147. doi: 10.1016/j.ecolind.2012.06.003
Stuart-Smith, R. D., Bates, A. E., Lefcheck, J. S., Duffy, J. E., Baker, S. C., Thomson, R. J., et al. (2013). Integrating abundance and functional traits reveals new global hotspots of fish diversity. Nature 501, 539–542. doi: 10.1038/nature12529
Stubbington, R., Chadd, R., Cid, N., Csabai, Z., Milisa, M., Morais, M., et al. (2018). Biomonitoring of intermittent rivers and ephemeral streams in Europe: current practice and priorities to enhance ecological status assessments. Sci. Total Environ. 618, 1096–1113. doi: 10.1016/j.scitotenv.2017.09.137
Stubbington, R., England, J., Sefton, C., and Wood, P. J. (2017). Temporary streams in temperate zones: recognizing, monitoring and restoring transitional aquatic-terrestrial ecosystems. Wiley Interdiscip. Rev. Water 4, e1223. doi: 10.1002/wat2.1223
Stubbington, R., Paillex, A., England, A., Barthès, A., Bouchez, A., Rimet, F., et al. (2019). Comparison of biotic groups as dry- phase indicators of ecological quality in intermittent rivers and ephemeral streams. Ecol. Indic. 97, 165–174. doi: 10.1016/j.ecolind.2018.09.061
Suren, A. M., Biggs, B. J. F., Kilroy, C., and Bergey, L. (2003). Benthic community dynamics during summer low-flows in two rivers of contrasting enrichment 1. Periphyton. N. Z. J. Mar. Freshw. Res. 37, 53–70. doi: 10.1080/00288330.2003.9517146
ter Braak, C. J. F., and Šmilauer, P. (2002). CANOCO Reference Manual and CanoDraw for Win- dows User’s Guide: Software for Canonical Community Ordination (Version 4.5). Ithaca, NY: Microcomputer Power.
Thompson, L. M., Lynch, A. J., Beever, E. A., Engman, A. C., Falke, J. A., Jackson, S. T., et al. (2021). Responding to ecosystem transformation: resist, accept, or direct? Fisheries 46, 8–21. doi: 10.1002/fsh.10506
Tornés, E., Colls, M., Acuña, V., and Sabater, S. (2021). Duration of water flow interruption drives the structure and functional diversity of stream benthic diatoms. Sci. Total Environ. 770:144675. doi: 10.1016/j.scitotenv.2020.144675
van Dam, H., Mertens, A., and Sinkeldam, J. (1994). A coded checklist and ecological indicator values of freshwater diatoms from The Netherlands. Neth. J. Aquat. Ecol. 28, 117–133. doi: 10.1007/BF02334251
Várbíró, G., Borics, G., Csányi, B., Fehér, G., Grigorszky, I., Kiss, K. T., et al. (2012). Improvement of the ecological water qualification system of rivers based on the first results of the Hungarian phytobenthos surveillance monitoring. Hydrobiologia 695, 125–135. doi: 10.1007/s10750-012-1120-2
Várbíró, G., Borics, G., Novais, M. H., Morais, M. M., Rimet, F., Bouchez, A., et al. (2020). Environmental filtering and limiting similarity as main forces driving diatom community structure in Mediterranean and continental temporary and perennial streams. Sci. Total Environ. 741:140459. doi: 10.1016/j.scitotenv.2020.140459
Keywords: flow intermittence, small hilly streams, temperate region, water quality indices, aquatic ecosystem, compositional changes
Citation: B-Béres V, Kókai Z, Várbíró G, Mustazhapova G, Csabai Z, Pernecker B, Borics G, Bácsi I and Boda P (2022) Flow Intermittence Drives the Benthic Algal Composition, Biodiversity and Diatom-Based Quality of Small Hilly Streams in the Pannonian Ecoregion, Hungary. Front. Ecol. Evol. 10:834548. doi: 10.3389/fevo.2022.834548
Received: 13 December 2021; Accepted: 01 February 2022;
Published: 10 March 2022.
Edited by:
Chao Wang, Pearl River Fisheries Research Institute, Chinese Academy of Fishery Sciences, ChinaReviewed by:
Michael Alister Reid, University of New England, AustraliaPatrick Rioual, Institute of Geology and Geophysics (CAS), China
Copyright © 2022 B-Béres, Kókai, Várbíró, Mustazhapova, Csabai, Pernecker, Borics, Bácsi and Boda. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Viktória B-Béres, YmVyZXMudmlrdG9yaWFAZ21haWwuY29t