 Hong Ren
Hong Ren- Institute of Ecological Environment Planning and Technology, Taiyuan, Shanxi, China
This study evaluates the decadal population dynamics of vegetation restoration in a 2.8 hm² biodiversity-ecosystem functioning plot within Pingshuo open-pit mine’s reclamation area, comparing 2013 and 2023 survey data. This decade-long study focuses on survival dynamics, growth patterns, and interspecific interactions among four pioneer species: locust, oil pine, sea buckthorn, and Caragana microphylla. Through experimental plots with diverse planting configurations, mixed models revealed critical trade-offs between facilitation and competition. Oil pine and locust exhibited mutually beneficial interactions, achieving near-complete survival in paired configurations, while combinations involving sea buckthorn and caragana microphylla suffered marked competitive suppression. Monoculture outcomes varied. Locust thrived independently, whereas oil pine showed enhanced survival in mixed communities. Morphological traits displayed configuration-dependent plasticity, with locust-sea buckthorn combinations optimizing growth across species, and oil pine demonstrating superior basal development in isolation. Notably, shrub vulnerability to niche overlap contrasted with tree resilience, suggesting divergent adaptive strategies in moisture-limited environments. The study highlights the importance of species compatibility, functional trait complementarity, and phased planting approaches to balance competitive dynamics. Key findings propose the locust-sea buckthorn configuration as a strategic model for promoting multi-species growth, offering actionable insights for ecological restoration in degraded post-mining landscapes.
1 Introduction
While essential for resource extraction, open-pit mining causes profound disturbances to ecosystems, resulting in severe land degradation, loss of biodiversity, and habitat fragmentation. Ecological reclamation of post-mining landscapes has emerged as a critical strategy for restoring ecosystem functions and mitigating long-term environmental impacts. Among reclamation practices, vegetation restoration plays a key role in stabilizing soils, enhancing carbon sequestration, and promoting biodiversity recovery. However, the long-term success of such efforts depends on understanding population dynamics, species adaptability, and species interactions under different restoration models. Despite extensive research on short-term restoration outcomes, decadal-scale assessments of plant population dynamics remain scarce, limiting insights into the stability and sustainability of restored ecosystems.
In recent years, Bai et al. (2018) applied Biodiversity-Ecosystem Functioning (BEF) experimental plots to the ecological restoration process at the Pingshuo open-pit mine (Zhang and Zhang, 2003; Mark, 2013; Bai et al., 2018; Zhao et al., 2018). Vegetation restoration often takes 50-100 years to cover with ideal vegetation, while artificial vegetation restoration and reconstruction can significantly accelerate the succession process and promote environmental improvement (Haigh and Gentcheva-Kostadinova, 2002; Xiang, 2009; Yang et al., 2015; Han et al., 2018). In extremely degraded mining ecosystems, a vegetation restoration system that combines artificial intervention and natural succession has been established (Harris et al., 1996; Gao et al., 2001; Hao et al., 2003; Zhao, 2003; Wang et al., 2010). Over time, the populations under different vegetation compositions will show distinct population characteristics due to the interaction between artificially planted vegetation and the regional natural environment (Zhang, 2014; Xing, 2015; Zhao et al., 2015; Zhang et al., 2016a; Zhang et al., 2016b; Dong and Li, 2018).
A growing amount of literature highlights the importance of species selection and planting configurations for restoration success, particularly in degraded mining landscapes. Studies in semi-arid regions have shown that mixed-species planting models often outperform monocultures by enhancing niche complementarity, resource use efficiency, and resistance to environmental stressors (Zuppinger-Dingley et al., 2014; Dai et al., 2023). For example, Caragana microphylla is often prioritized in reclamation designs due to their ability to improve soil fertility and facilitate the establishment of companion species. Similarly, drought-tolerant pioneer trees are often selected for their rapid growth and soil-stabilizing capacity. However, the long-term viability of these configurations remains controversial, as interspecific competition, allelopathic effects, or mismatched ecological strategies may undermine early gains. Existing research has also highlighted trade-offs between survival rates and growth performance. For example, fast-growing species may exhibit high mortality under prolonged drought, whereas slower-growing conifers often exhibit greater resilience despite slower canopy development (Decarsin et al., 2024). These findings indicate the need for empirical assessments of the performance of different restoration models, ranging from single plantings to complex multi-species assemblages, on decadal time scales in challenging post-mining environments.
In this study, we focus on the main restoration configurations previously tested in semi-arid mining areas, including monocultures of locust, oil pine, sea buckthorn, and caragana microphylla, as well as mixed plantings. These configurations were chosen on the basis of their roles in early reclamation, namely, oil pine for its stress tolerance, caragana microphylla for soil enrichment, locust for rapid biomass accumulation, and sea buckthorn for erosion control. However, their long-term coexistence and interactions—particularly in terms of competition for light, water, and nutrients—remain poorly understood. By examining these models over a decade, we aim to unravel whether initial functional traits (e.g., nitrogen fixation, drought resistance) translate into sustained ecological benefits or lead to unintended outcomes such as dominance hierarchies or population decline.
This study investigates the decadal population dynamics of woody vegetation in a 2.8-hectare BEF plot within the Pingshuo reclamation area, comparing data from 2013 and 2023. Focusing on species composition, survival rates, and growth parameters across restoration models, we aim to elucidate (1) how plant communities evolve over a decade in a reclaimed mining landscape, (2) the performance of key species under varying planting configurations, and (3) the stability of interspecies relationships and ecosystem structure. By integrating population ecology principles with long-term field observations, this work advances the theoretical framework for ecological restoration in degraded mining systems while providing actionable insights for improving reclamation practices in semi-arid regions worldwide.
2 Materials and methods
2.1 Study area
The study area is located in the western mine dump of the Antaibao open-pit coal mine in Shuozhou City, Shanxi Province of China (112°10′58″-113°30′E, 39°23′-39°37′N). The region has a typical temperate semi-arid continental monsoon climate. The annual precipitation is 449.0 mm, with rainfall concentrated in July, August, and September, accounting for 75% of the annual precipitation. The annual evaporation ranges from 1786.6 to 2598.0 mm, with the maximum evaporation occurring in May, June, and July, exceeding four times the amount of precipitation. The ≥10°C annual accumulated temperature is between 2200 and 2500°C, with a daily temperature difference of 18-25°C. The annual temperature difference can even reach 61.8°C. The frost-free period lasts about 115 to 130 days. The average annual wind speed ranges from 2.5 to 4.2 m/s, with a maximum wind speed of 20 m/s and gusts reaching up to 24 m/s. The area experiences more than 35 days of strong winds (above level 8) annually. The regional zonal soil is chestnut soil and Castano-cinnamon soil. According to Chinese vegetation regionalization, the study area belongs to the mountain forest-steppe zone. The area is sparsely distributed with drought-tolerant plants such as ice grass and Stipa krylovii, with lower vegetation coverage.
2.2 Methods
2.2.1 Field study
The BEF plot at the western mine dump of the Antaibao open-pit coal mine was established in September 2013, with an average altitude of 1456 m, and the entire plot has a relatively flat terrain.
Before coal mining, the land use type in this area was barren land, with vegetation such as sandthorn, black locust, and reedmace. After reclamation, the platform area includes 5 cereal crops (CC) land plots, 15 herbaceous plants (HP) land plots, 12 arbor-shrubs (AS) land plots, and 4 wild grass (WG) land plots. In CC regions, the vegetation types are buckwheat and corn. In HP regions, the vegetation type is while purple medic. In AS regions, the vegetation types are oil pine, white poplar, black locust, jujube, and sandthorn. In WG regions, the vegetation types are feather grass and ice grass.
The plot follows the technical methods of the Center for Tropical Forest Science (CTFS), selecting two tree species (locust and oil pine) and two shrub species (sea buckthorn and caragana microphylla) as pioneer species for reclamation. As a native tree species, the oil pine has low moisture requirements, easily survives in harsh conditions, and is able to adapt to the environment of a drainage site. Locust is a tree with a strong root system that draws moisture from the soil without relying on external moisture. It is a light-loving, medium-sized, deciduous broadleaf pioneer tree species of the legume family that can participate in the transformation and migration of nitrogen in the soil, has a high seedling survival rate, and is often used for phytoremediation. Sea buckthorn is a deciduous shrub that has a distinct clonal character in its growth and reproduction. Dividing and natural thinning are often applied to regulate the population density of sea buckthorn and to increase the survival rate in order to achieve the best growth. As a leguminous shrub species, caragana microphylla is tolerant to drought and cold, tolerant to barrenness and strongly influenced by water quantity. It requires external precipitation for its growth, has a fairly high survival rate, and is an important pioneer species in semi-arid areas. Therefore, these four plants are easy to survive in harsh environments.
A total of 15 combinations of the four plant species were randomly generated, with three experimental replicates for each combination, resulting in 45 experimental plots (Figure 1; Table 1). The total plot area is 2.8 hm2, divided into 45 square plots, each measuring 25 m × 25 m. Each plot contains 12 rows × 12 columns, totaling 144 plants, with 1.5 m distance between plot edges and 2 m × 2 m spacing between plants.
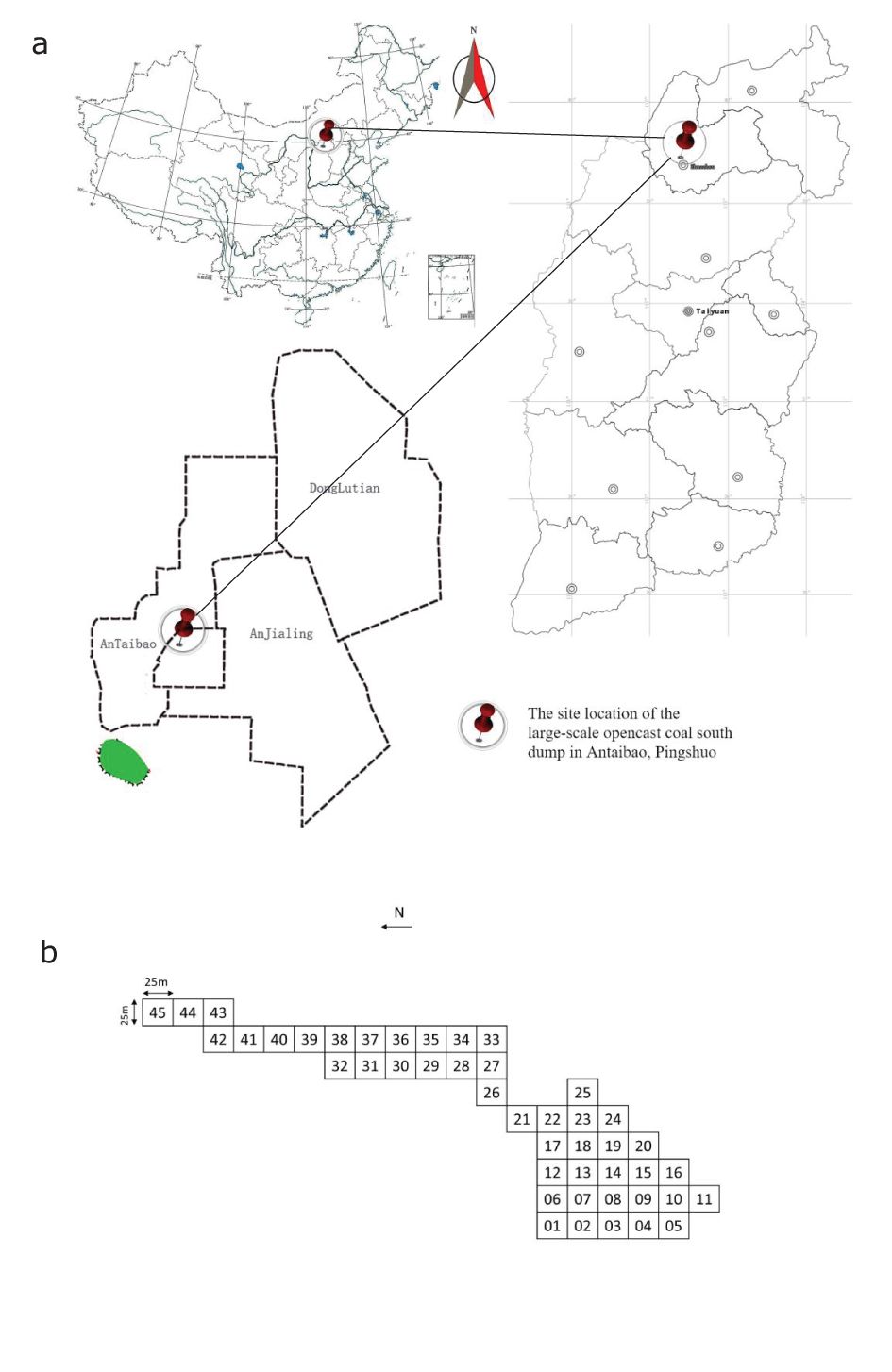
Figure 1. (a) Location of Antaibao open-pit coal mine; (b) Diagram of setting BEF sample site.
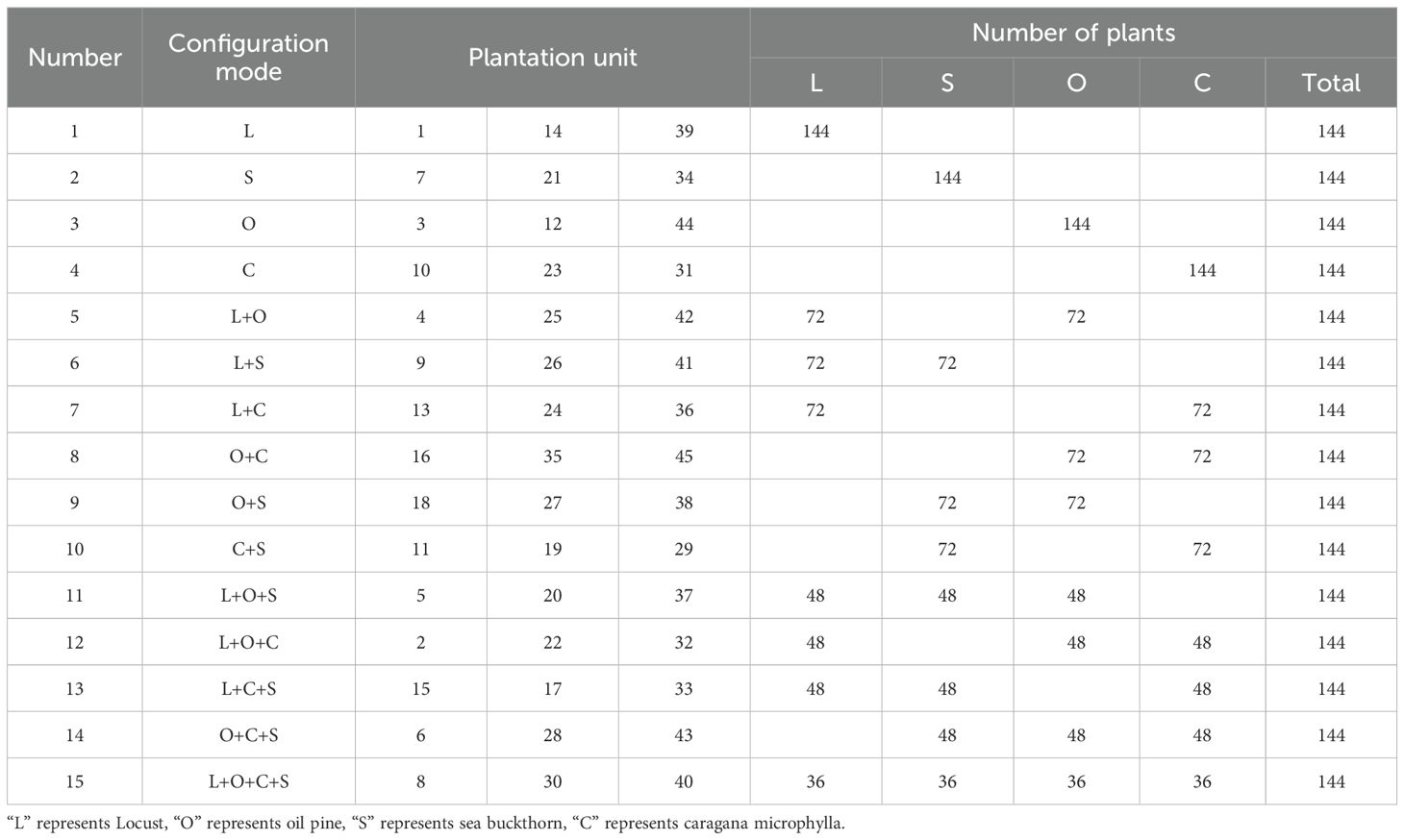
Table 1. Basic overview of BEF sample site (“L” represents Locust, “O” represents oil pine, “S” represents sea buckthorn, “C” represents caragana microphylla).
Data collection for the plot was based on the individual plots as the survey units. In July 2022, all tree and shrub species in the plot were identified, and measurements including species names, diameter at breast height (DBH), height, coordinates, and growth status were recorded.
2.2.2 Data processing
The data used in this study were collected in July 2022 and 2013, respectively. In order to study the dynamics of forest communities, researchers have developed the concept of mortality rate. Individuals were designated as “mortality” if they were found dead or not found during the second sample plot survey. The species mortality rate (M) was subsequently calculated as outlined by Condit et al. (1999). The dynamics of four pioneer species (oil pine, locust, sea buckthorn, and caragana microphylla) under different vegetation restoration models were analyzed, including average crown diameter (Fx), average height (Hx), average base diameter (Rx), average diameter at breast height (Dx), and population characteristics through Excel and SPSS.
The mortality rate (M), Fx, Hx, Rx, Dx were calculated as follows:
Where: N0 is the number of individuals in a population at the first survey, St is the number of individuals in that population that survived the second survey, t is the interval between the two surveys, and i represents the total number of surviving individuals in a given vegetation model.
3 Results
In 2013, 1620 plants of each of the four species were planted, totaling 6480 plants. After 10 years, 4263 plants survived, with an overall survival rate of 65.79%. The annual mortality rate was 8.38%. The survival rates for the four populations were: oil pine (91.98%) > black locust (76.60%) > locust (56.11%) > sea buckthorn (38.46%). A total of 169 invasive trees and shrubs were found in the plot over the 10 years, including 73 elm trees, 91 poplars, and 5 willows. The tree density in the plot decreased from 2314 individuals per hectare in 2013 to 1583 individuals per hectare in 2022.
Over the 10 years, mortality was observed in all 45 plots. The lowest mortality occurred in the oil pine monoculture model, where only 6 out of 144 oil pines died, with an annual mortality rate of 0.85%. The highest mortality occurred in the sea buckthorn monoculture model, where 98 out of 144 sea buckthorns died, with an annual mortality rate of 22.82%.
As shown in Figure 2, the survival rate of the oil pine monoculture model was higher compared to other combinations in the 15 vegetation combination models. The three experimental plots with oil pine had annual survival rates of 91%. The survival rates of three vegetation combination models were below 50%, namely the sea buckthorn monoculture (39%), the combination of sea buckthorn and caragana microphylla (37%), and the combination of sea buckthorn, caragana microphylla, and locust (47%). Other vegetation combinations had survival rates ranging from 60% to 70%.
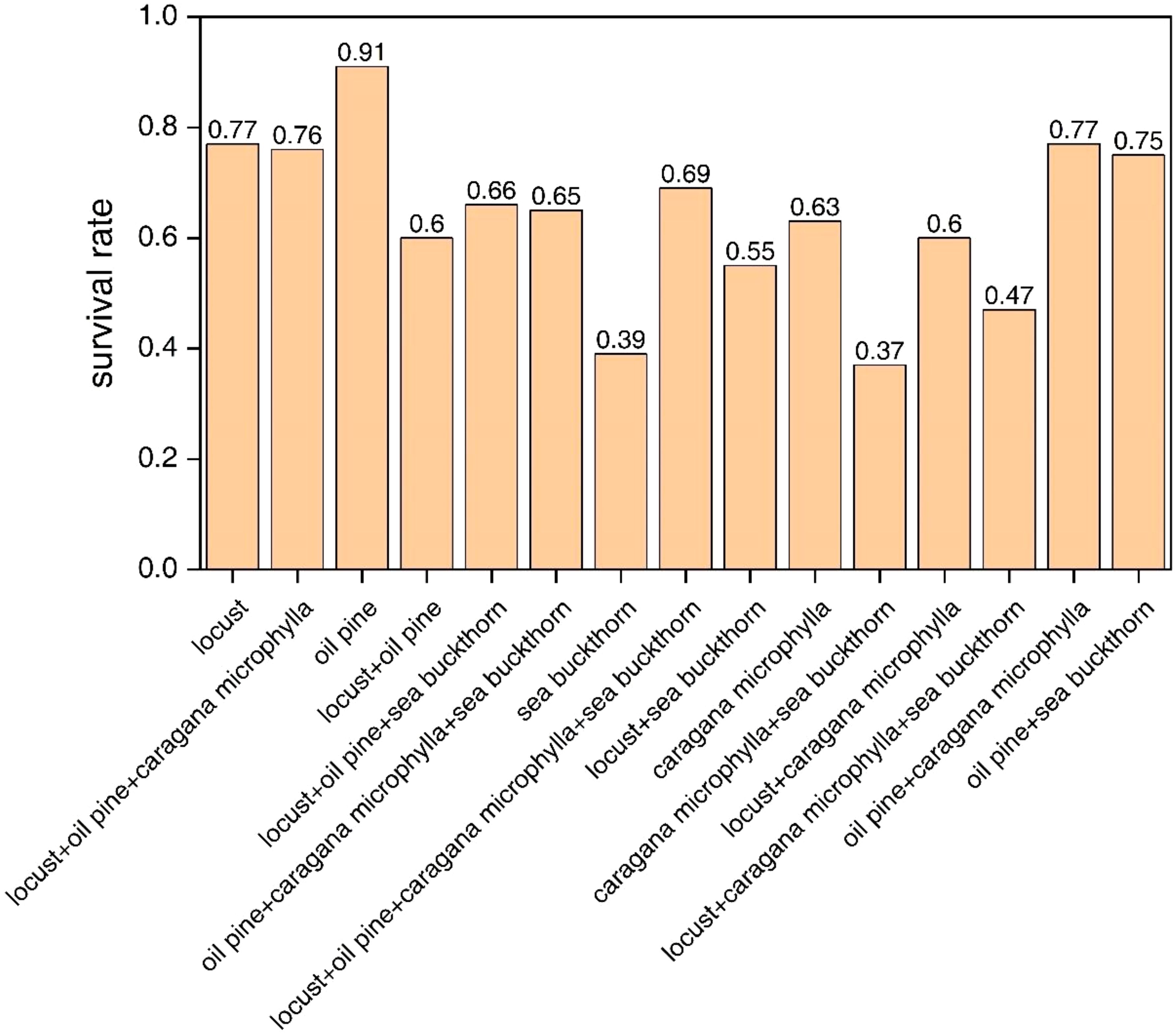
Figure 2. Overall survival of different vegetation restoration models.
3.1 Population characteristics of oil pine, locust, sea buckthorn, and caragana microphylla under different vegetation restoration models
In 2013, 1620 individuals of oil pine, locust, sea buckthorn, and caragana microphylla were planted in each model. After 10 years, the populations survived as follows: 1241 (76.60%) for locust, 1490 (91.98%) for oil pine, 909 (56.11%) for caragana microphylla, and 623 (38.46%) for sea buckthorn (Table 2).

Table 2. Number (plants) and percentage (%) of established pioneer species populations surviving under different vegetation configuration patterns (“L” represents Locust, “O” represents oil pine, “S” represents sea buckthorn, “C” represents caragana microphylla).
Oil pine demonstrated the highest overall survival rate (91.98%), peaking at 99.07% in a two-species model with locust and declining to 84.72% in a three-species model with caragana microphylla and sea buckthorn. Its annual mortality was lowest (0.20%) in the locust-oil pine combination and highest (3.32%) in the oil pine-caragana microphylla-sea buckthorn model. Oil pine monoculture survival (90.97%) was markedly lower than its optimal mixed model.
Locust exhibited the second-highest overall survival rate (76.60%), with its maximum survival (83.33%) achieved in a mixed model combining locust, oil pine, and caragana microphylla, while its lowest survival (70.37%) occurred in a two-species model with caragana microphylla. Notably, locust monoculture (77.08%) outperformed some mixed models. Annual mortality for locust ranged from 3.65% (lowest, in the three-species mixed model) to 7.16% (highest, in a locust-sea buckthorn combination).
Caragana microphylla, with the second-lowest overall survival (56.11%), showed the highest survival (72.22%) in a four-species model (locust, oil pine, caragana, sea buckthorn) and the lowest (42.59%) in a three-species model with locust and sea buckthorn. Its annual mortality extremes spanned 6.51% (four-species model) to 17.07% (two-species model with sea buckthorn).
Sea buckthorn, the least resilient species (overall survival: 38.46%), performed best in combination with oil pine (60.19%) and worst in a three-species model with locust and caragana microphylla (18.06%). Its mortality ranged from 10.16% (oil pine-sea buckthorn model) to 34.24% (three-species stress-sensitive model).
In conclusion, mixed models enhanced survival for dominant species but intensified stress for vulnerable species like sea buckthorn. Monoculture outcomes were inconsistent: locust thrived alone, while oil pine significantly benefited from interspecific interactions. These results underscore the importance of species-specific compatibility in designing effective vegetation restoration strategies.
3.2 Morphological characteristics of locust, oil pine, caragana microphylla, and sea buckthorn populations under different vegetation restoration models
The locust population exhibited distinct morphological variations across vegetation restoration models. As shown in Figure 3a, crown diameter was maximized in the mixed model of locust and sea buckthorn (15.80), while the smallest crown diameter occurred in the locust-oil pine combination (10.37), with an average of 12.11 across all models. For basal diameter, the highest value (5.420) was observed in the locust-sea buckthorn mixed model, whereas the lowest (4.170) appeared in the locust-oil pine-sea buckthorn combination, averaging 4.59 overall (Figure 3b). In terms of height, the tallest locust trees (4.37) developed in the locust-sea buckthorn mixed model, contrasting with the shortest (3.82) in the locust-oil pine-caragana microphylla combination, yielding an average height of 4.12 (Figure 3c). Notably, monoculture (locust alone) produced intermediate values for all traits (crown: 11.53; basal diameter: 4.941; height: 4.16), but consistently underperformed compared to the optimal mixed model (locust-sea buckthorn), which dominated in crown diameter, basal diameter, and height. These results highlight interspecific interactions, particularly with sea buckthorn, significantly enhanced locust’s morphological development, while combinations involving oil pine generally suppressed growth.
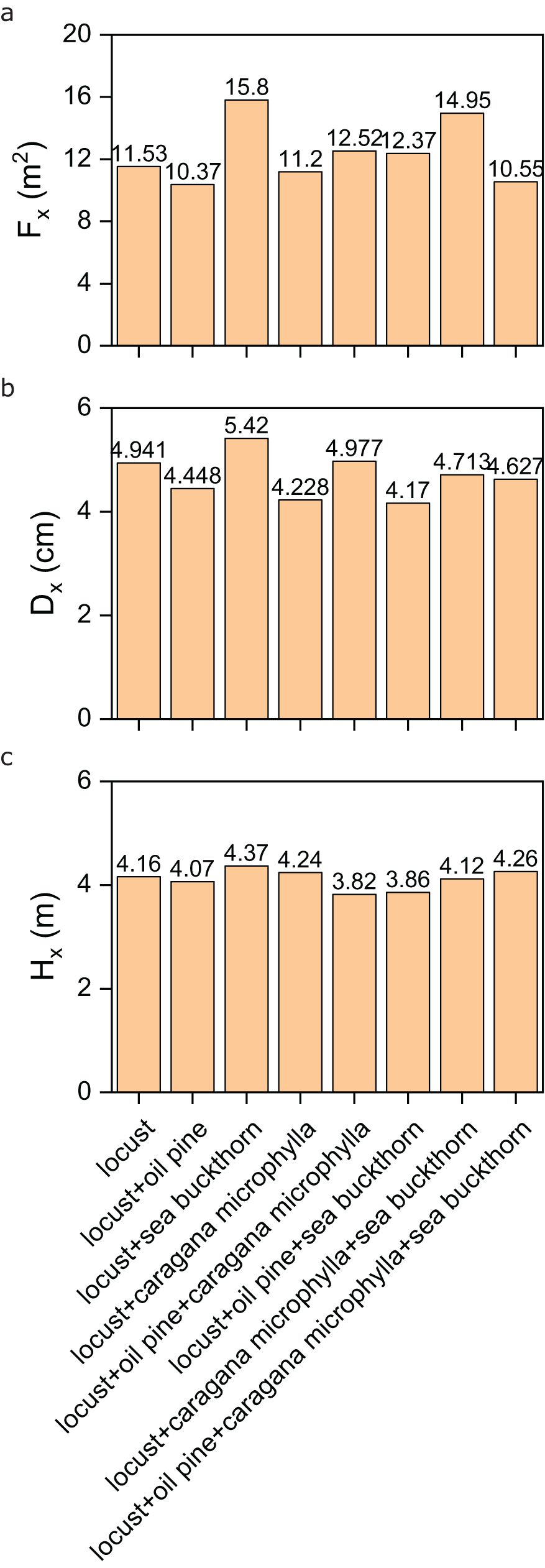
Figure 3. (a) The crown diameter of the locust population in different vegetation configuration models; (b) the base diameter of the locust population in different vegetation configuration models; (c) the height of the locust population in different vegetation configuration models.
The morphological traits of oil pine varied significantly across vegetation restoration models. As shown in Figure 4a, oil pine achieved its largest crown diameter (1.18) in the mixed model with locust and caragana microphylla, while its smallest crown diameter (0.97) occurred in monoculture. For basal diameter, monoculture (3.529) outperformed all mixed models, whereas the lowest value (1.546) was observed in the locust-oil pine-sea buckthorn combination (Figure 4b). Oil pine’s maximum height (1.42) occurred in two mixed models (locust-oil pine-caragana microphylla and locust-oil pine-caragana microphylla-sea buckthorn), contrasting with its minimum height (1.21) in the oil pine-caragana microphylla combination (Figure 4c).
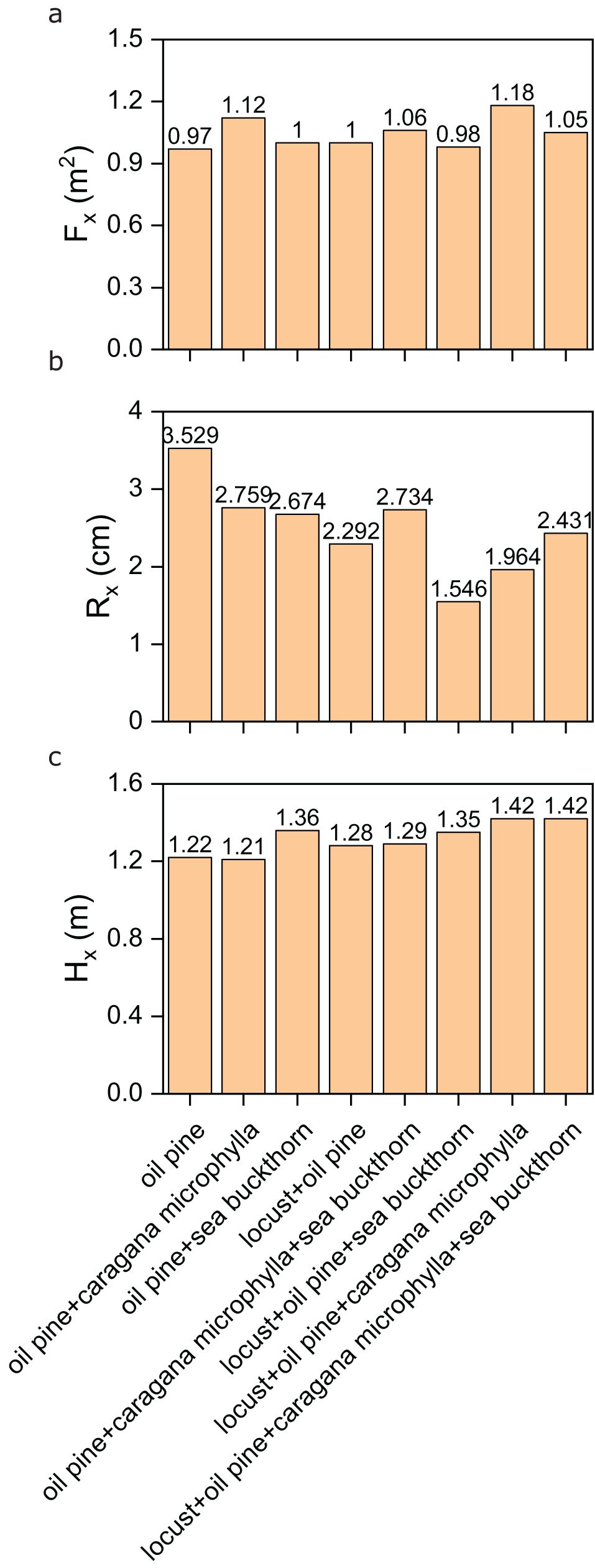
Figure 4. (a) The crown diameter of the oil pine population in different vegetation configuration models; (b) the base diameter of the oil pine population in different vegetation configuration models; (c) the height of the oil pine population in different vegetation configuration models.
In terms of crown diameter, Caragana microphylla exhibited its largest crown diameter (4.82) in the oil pine-caragana microphylla mixed model, while the smallest (2.79) appeared in the four-species model (locust-oil pine-caragana microphylla-sea buckthorn) (Figure 5a). Its highest basal diameter (1.791) and tallest height (1.50) were recorded in monoculture and the oil pine-caragana microphylla-sea buckthorn model, respectively (Figures 5b, c). Conversely, the lowest basal diameter (1.408) and shortest height (1.30) occurred in mixed models involving locust and oil pine.
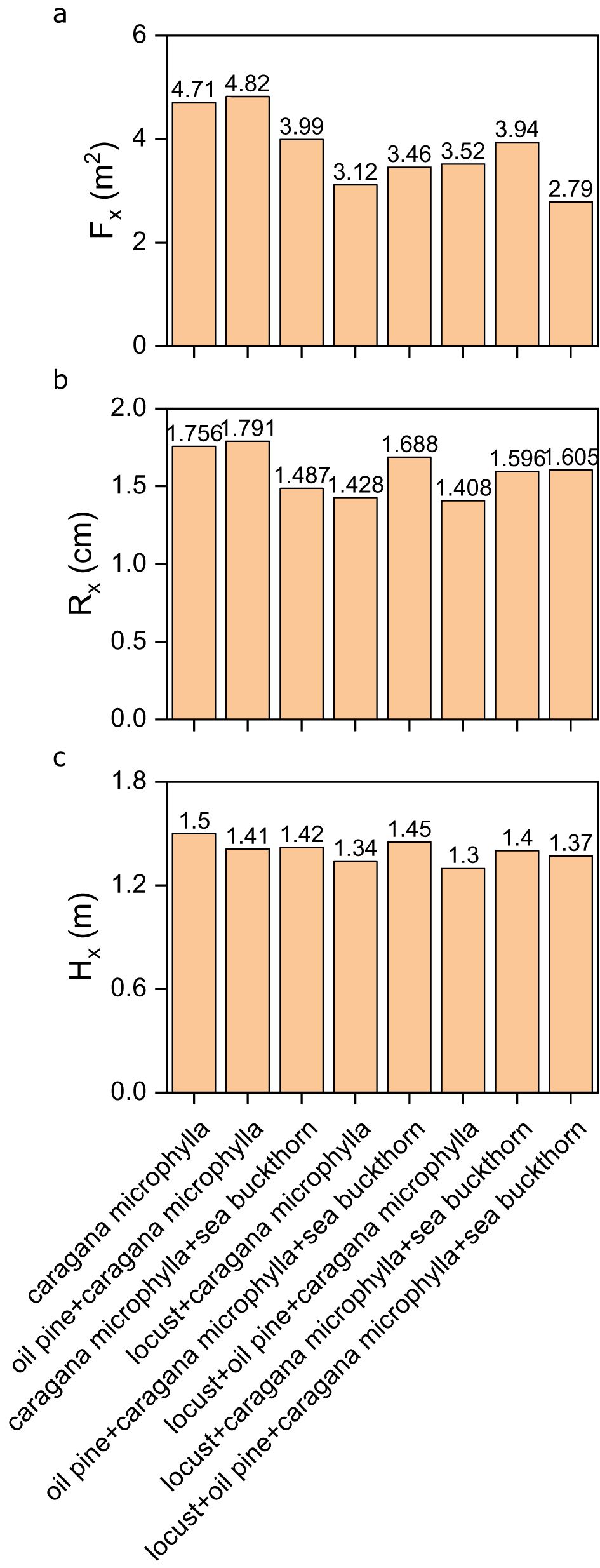
Figure 5. (a) The crown diameter of the caragana microphylla population in different vegetation configuration models; (b) the base diameter of the caragana microphylla population in different vegetation configuration models; (c) the height of the caragana microphylla population in different vegetation configuration models.
Sea buckthorn demonstrated extreme morphological responses. Its largest crown diameter (6.07) and tallest height (1.98) were observed in the locust-caragana microphylla-sea buckthorn model, whereas monoculture resulted in the smallest crown diameter (2.28) and shortest height (1.21) (Figures 6a, c). For basal diameter, the maximum (2.028) occurred in the locust-caragana microphylla-sea buckthorn combination, while the minimum (1.261) appeared in the four-species model (Figure 6b).
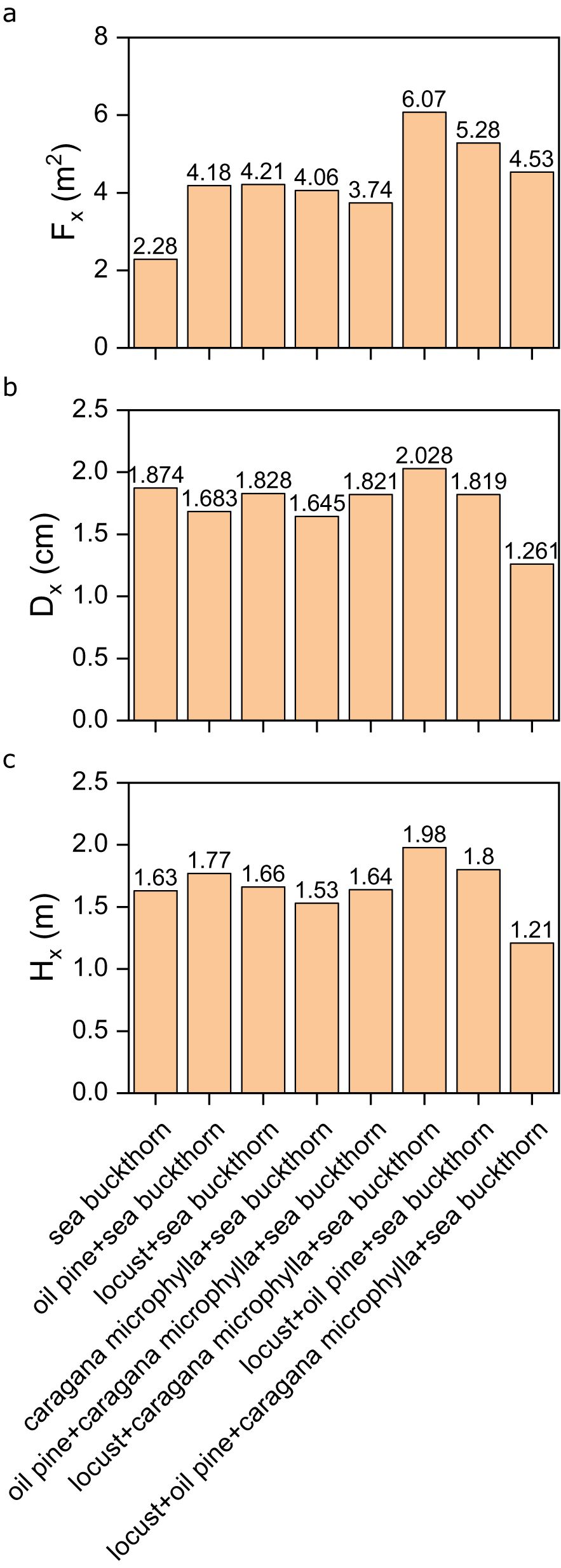
Figure 6. (a) The crown diameter of the sea buckthorn population in different vegetation configuration models; (b) the base diameter of the sea buckthorn population in different vegetation configuration models; (c) the height of the sea buckthorn population in different vegetation configuration models.
Key trends indicate that mixed models generally enhanced growth for caragana microphylla and sea buckthorn but suppressed oil pine’s basal diameter. Monoculture was advantageous only for oil pine’s basal diameter and caragana microphylla’s height, highlighting species-specific dependencies on interspecific interactions. The locust-sea buckthorn synergy consistently promoted morphological traits across multiple species, suggesting its potential as a strategic restoration configuration.
4 Discussion
The decade-long vegetation dynamics observed in this reclaimed mining area provide critical insights into the early-stage ecological recovery processes and the efficacy of artificial restoration strategies. The absence of major disturbances since the establishment of experimental plots in 2013 has allowed for the evaluation of species-specific interactions and community stability under controlled conditions. Notably, the dominance of artificially introduced pioneer species throughout the 10-year period highlights the slow pace of natural succession in such degraded ecosystems, contrasting sharply with undisturbed native ecosystems in the region, which typically exhibit faster species turnover and structural complexity development. This divergence underscores the inherent challenges of jump-starting ecological processes in heavily modified landscapes.
4.1 Interspecific facilitation and competition in mixed plantations
In the early stages of vegetation restoration, the effects of mutual influences among populations have already become apparent. The differential survival rates across planting configurations reveal nuanced interspecific relationships. The facilitative role of arborescent species (e.g., locust and oil pine) in enhancing shrub survival is particularly noteworthy. For example, caragana microphylla survival increased from 62.96% in monoculture to 72.22% when co-planted with sea buckthorn, oil pine, and locust. Similarly, oil pine achieved near-complete survival (99%) when paired with locust, suggesting strong complementary resource use strategies between these species. These findings align with the stress-gradient hypothesis, where facilitative interactions amongst plant species dominate under harsh conditions (Wang et al., 2019). The exceptional performance of locust+oil pine configurations (214/216 survival) may stem from complementary root architectures—locusts’ nitrogen-fixing capability potentially ameliorating soil conditions for the conifer (Zhang et al., 2020).
However, the reduced survival rate of sea buckthorn (38.66% to 30.56%) and caragana microphylla (62.96% to 42.59%) in their pairwise combination exemplifies competitive exclusion, likely driven by niche overlap in water and nutrient acquisition. This dichotomy between facilitation and competition emphasizes the need for species-specific compatibility assessments in restoration planning. Among the eight vegetation configuration models containing sea buckthorn, the combination of locust, caragana microphylla, and sea buckthorn configuration had the highest average crown diameter, base diameter, and height of the sea buckthorn population, with values of 6.07 m2, 2.028 cm, and 1.98 m, respectively. The observed morphological plasticity further suggests that certain combinations may optimize microhabitat conditions despite initial survival trade-offs.
The superior survival and growth of arborescent species (locust and oil pine) compared to shrubs (sea buckthorn and caragana microphylla) likely reflects adaptive advantages in this semi-arid continental monsoon climate. Deeper root systems in trees may access stable groundwater reserves, buffering against precipitation variability, while shrubs’ shallower roots make them more vulnerable to surface soil moisture fluctuations (Zhao et al., 2024). This pattern contrasts with natural succession trajectories in undisturbed regional ecosystems, where shrub dominance typically precedes tree establishment (Meng et al., 2023). The inversion here suggests artificial restoration may short-circuit natural successional sequences, potentially creating novel communities with uncertain long-term stability.
4.2 Implications for restoration practice for open-pit mine dump sites
In terms of population dynamics characteristics, tree-only, tree and tree, and shrub and tree vegetation configurations are superior to shrub-only and shrub and shrub configurations. In the early stages of vegetation restoration, locust is the fastest-growing plant species among the four pioneer species. Compared to locust and oil pine, the survival status of caragana microphylla and sea buckthorn is clearly disadvantaged. The sea buckthorn and caragana microphylla populations exhibit a reverse inhibitory effect on each other, while locust and oil pine populations exhibit a positive facilitative effect on each other. In terms of species selection framework, the success of locust and oil pine configurations demonstrates the value of combining deep-rooted trees with nitrogen fixers in moisture-limited environments. This aligns with global observations of improved restoration outcomes through functional trait complementarity (Zhang et al., 2025). In terms of temporal management strategies, the transient competitive suppression of shrubs by trees suggests phased planting approaches—establishing shrubs before introducing trees—may better mimic natural succession patterns. In terms of microhabitat engineering, the enhanced growth metrics in specific combinations (e.g., locust and sea buckthorn) indicate that certain species pairings can create favorable microenvironments, potentially accelerating soil development.
5 Conclusion
The study demonstrates that tree-dominated vegetation configurations, such as the combination of locust and oil pine, outperform shrub-based models in early-stage ecological restoration of open-pit coal mine spoil banks. Growth rates of locust were notably higher than those of shrubs like caragana microphylla and sea buckthorn. Additionally, interspecies positive interactions between locust and oil pine were observed, suggesting a beneficial synergy for vegetation restoration in arid regions. While mixed plantings demonstrate clear advantages over monocultures in survival rates, the artificial communities remain ecologically simplistic compared to natural benchmarks. Restoration strategies must balance short-term survival goals with longer-term objectives of functional and structural complexity. Future work should integrate assisted nucleation techniques with targeted native species introductions to bridge the gap between artificial restoration outcomes and natural ecosystem trajectories. The decadal data presented here provide a critical baseline for understanding multi-temporal scale recovery processes in post-mining landscapes.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Author contributions
HR: Data curation, Methodology, Supervision, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Bai W., Li S., and Di X. (2018). The interspecific association of dominant herbaceous plants under different vegetation types in the reclamation land of the Antaibao open-pit coal mine. J. Appl. Environ. Biol. 24, 140–145.
Condit R., Ashton P. S., Manokaran N., LaFrankie J. V., Hubbell S. P., and Foster R. B. (1999). Dynamics of the forest communities at Pasoh and Barro Colorado: Comparing two 50-ha plots. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences. 354, 1739–1748. doi: 10.1098/rstb.1999.0517
Dai J., Li Y., and Wang L. (2023). Mixed-species plantations alleviate deep soil water depletion and facilitate hydrological niche partitioning compared to pure plantations. For. Ecol. Manage. 539, 121017.
Decarsin R., Guillemot J., le Maire G., Blondeel H., Meredieu C., Achard E., et al. (2024). Tree drought-mortality risk depends more on intrinsic species resistance than on stand species diversity. Global Change Biol. 30, e17503.
Dong Q. and Li S. (2018). Ecological relationships of herbaceous plant communities under different vegetation types in the reclamation land of the Antaibao open-pit coal mine. Chin. Agric. Bull. 34, 95–100.
Gao G., Gao B., Zhou X., and Jin Y. (2001). Dynamic research on land reclamation in foreign industrial and mining areas. Soil Water Conserv. Res. 8, 98–103.
Haigh M. J. and Gentcheva-Kostadinova S. (2002). Ecological erosion control on coal-spoil banks: An evaluation. Ecol. Eng. 18, 371–377.
Han Y., Zhao W., Zhang Q., Wang Q., Shi N., Xiao N., et al. (2018). Restoration effect of different vegetation restoration models on abandoned mining sites. Soil Water Conserv. Res. 25, 120–125.
Hao R., Bai Z., Zhao J., Peng S., and Song Y. (2003). Vegetation dynamics in the reclamation process of large open-pit coal mine spoil heaps in the Loess Plateau. Acta Ecol. Sin. 23, 1470–1476.
Harris J. A., Birch P., and Palmer P. (1996). Land restoration and reclamation: Principles and practice, (Prentice Hall, Singapore).
Mark P. (2013). Experimental studies on biodiversity and ecosystem function. Biodiversity 21, 247–248.
Meng Y., Li S., Wang S., Meiners S., and Jiang L. (2023). Scale-dependent changes in ecosystem temporal stability over six decades of succession. Sci. Adv. 9 (40), eadi1279.
Wang F., Zhong T., Li S., Cheng Y., Tan H., and He X. (2010). Experimental study on vegetation optimization for ecological reclamation of spoil banks. Modern. Min. 26, 70–73.
Wang Y., Cadotte M. W., Chen Y., Fraser L. H., Zhang Y., Huang F., et al. (2019). Global evidence of positive biodiversity effects on spatial ecosystem stability in natural grasslands. Nat. Commun. 10, 3207.
Xiang Y. H., Li G. P., and Yu X. J. (2009). Ecological restoration and vegetation reconstruction techniques for waste dumps in modern open-pit mines. In Proceedings of the 2009 Annual Meeting and Academic Symposium of the Planning and Design Professional Committee, China Society of Soil and Water Conservation. 107–114.
Xing H. (2015). Correlation analysis of woody plant population dynamics and soil quality under different restoration models in the Antaibao open-pit coal mine. Master's thesis. (Shanxi University).
Yang Q., Zhao B., and Guo D. (2015). Research progress on vegetation restoration in northern China’s open-pit coal mining areas. Chin. J. Ecol. 34, 1152–1157.
Zhang Y. (2014). Analysis of the population dynamics of black locust under different vegetation configuration models in the Antaibao open-pit coal mine spoil bank (Shanxi University).
Zhang Q., Bi R., Wu Z., Song H., Bai Z., and Guo D. (2016a). Natural regeneration patterns and driving factors of species in the vegetation restoration area of the Pingshuo open-pit coal mine. Environ. Sustain. Dev. 41, 172–176.
Zhang Q., Bi R., Wu Z., Zhao B., Bai Z., and Guo D. (2016b). Spatiotemporal dynamics of natural regeneration of species in the vegetation restoration area of the Antaibao open-pit coal mine. Chin. J. Ecol. 35, 3223–3232.
Zhang X., Chen M., Shao T., Zhang G., Duan Q., Wang X., et al. (2025). Adaptation of plantations to drought events in arid and semi-arid regions: Evidence from tree resilience. For. Ecol. Manage. 578, 122437.
Zhang K., Cheng X., Dang H., Zhang Q., and Schrodt F. (2020). Biomass:N:K:Ca: Mg:P ratios in forest stands world-wide: Biogeographical variations and environmental controls. Global Ecol. Biogeogr. 29, 2176–2189.
Zhang Q. and Zhang D. (2003). Biodiversity and ecosystem function: Latest progress and trends. Biodiversity 11, 351–363.
Zhao P. (2003). Progress in physiological ecology research on vegetation restoration of degraded ecosystems. Chin. J. Appl. Ecol. 11, 2031–2036.
Zhao B., Guo D., and Bai Z. (2018). Community growth dynamics of the Robinia pseudoacacia × Pinus tabulaeformis reclamation model on the spoil heaps of open-pit coal mines in the Loess Plateau over 17–22 years. J. Agric. Environ. Sci. 37, 485–494.
Zhao Y., Wang L., Chun K. P., Ziegler A. D., and Evaristo J. (2024). Dynamic hydrological niche segregation: How plants compete for water in a semi-arid ecosystem. J. Hydrol. 630, 130677.
Zhao B., Wu Z., Xu Z., Yin Z., Wang Q., Wang X., et al. (2015). Species composition and spatial pattern changes of the Robinia pseudoacacia + Pinus tabulaeformis mixed forest at the reclamation site of an open-pit coal mine. Environ. Sustain. Dev. 40, 31–35.
Keywords: open-pit coal mine, vegetation restoration, biodiversity-ecosystem functioning (BEF), population dynamics, survival rate
Citation: Ren H (2025) Population dynamics of an experimental plot of biodiversity and ecosystem functioning in the Pingshuo open-pit mine. Front. Ecol. Evol. 13:1581588. doi: 10.3389/fevo.2025.1581588
Received: 22 February 2025; Accepted: 04 June 2025;
Published: 23 June 2025.
Edited by:
Darren Norris, Universidade Federal do Amapá, BrazilReviewed by:
Nayani Ilangakoon, University of Colorado Boulder, United StatesNebi Bilir, Isparta University of Applied Sciences, Türkiye
Copyright © 2025 Ren. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hong Ren, aGp0c3RjcmhAMTYzLmNvbQ==