 Camille Auger1
Camille Auger1 Michel Labrecque
Michel Labrecque Joan Laur
Joan Laur- 1Institut de recherche en biologie végétale, Université de Montréal, Montréal, QC, Canada
- 2Montréal Botanical Garden, Montréal, QC, Canada
As waste from engineered landfills decomposes, it produces nitrogen (N)-laden leachate that cannot be directly released into the environment. Plants such as willows have the potential to phytofilter this polluted water but must be able to tolerate large loads of contaminants and flooding conditions. To date, however, it is mainly exotic species that have been used in for the treatment of leachates in a pilot project conducted on a technical landfill. It would be useful to compare the effectiveness of native species from eastern Canada with that of the Salix miyabeana ‘SX64’, a willow cultivar used in the pilot project. Three willows indigenous to Canada: S. amygdaloides, S. bebbiana and S. nigra, were tested alongside S. miyabeana. A mesocosm experiment was conducted under semi-controlled conditions over six weeks to document the impact of various nitrogen overfertilization and flooding treatments on plant development, and to test the plants’ tolerance to these constraints to evaluate their suitability for large-scale vegetative filters. Overall, growth and biomass production of S. nigra and S. amygdaloides were not affected by the treatments. Furthermore, S. nigra was ten times more efficient than the cultivar of S. miyabeana in terms of decontamination capacity. While still in the juvenile phase, S. nigra plants removed the equivalent of 240m3 per hectare of the N-contaminated water initially applied (60 kg of N), under both permanent and cyclic flooding. These results suggest that native species could be given greater in future projects.
Highlights
1. Intermittent leachate application had more beneficial effects on willow growth than conventional fertigation.
2. Salix miyabeana ‘SX64’ and Canadian indigenous willows adjusted to nitrogen overfertilization and flooding while removing nitrogen from leachate, without experiencing a reduction in biomass or growth.
3. Salix nigra demonstrated several adaptive features to flooding, such as the development of adventitious and upward growing roots and lenticels. It stood out for the amount of nitrogen stored in its leaf biomass, significantly higher than that of the other species tested.
1 Introduction
Each year, five million tons of residual material are sent to engineered landfills in Québec, Canada (Recyc-Québec, 2018). At these sites, rainwater percolating through waste layers becomes loaded with contaminants and forms what is known as leachate, a liquid containing large amounts of dissolved organic and inorganic compounds, salts, and heavy metals (Kjeldsen et al., 2002). The decomposition of the organic matter in wastewater leads to the release of large number of compounds including nitrogen (Burton and Watson-Craik, 1998), which is essential for plant growth but potentially leading to nutrient imbalances and negative environmental consequences when supplied in excessive quantities (Barak et al., 1997; Song et al., 2016; Hao et al., 2020; Xu et al., 2022).
Rather than employing cost-prohibitive physicochemical processes, an alternative way to treat leachate is to use phytofiltration plantations (Pilon-Smits, 2005). Unlike other rather slow-acting phytotechnologies like phytoremediation, phytofiltration is quite immediately tangible, an efficiency that goes a long way towards the wider deployment of nature-based solutions. The crops chosen for this purpose must be particularly nutrient-demanding and able to grow well on waterlogged and contaminated substrates (Justin et al., 2010). This is the case for several poplar or willow species and cultivars (Kuzovkina et al., 2008; Jerbi et al., 2023a; Muklada et al., 2022), including those developed in breeding programs for energy crops. While not originally developed for any phytotechnologies, they are characterized by faster growth rates, high nitrogen uptake capacity and the potential to stimulate soil microbiota activity (Aronsson and Perttu, 2001; Newton et al., 2022). In species tolerant to semi-permanent flooding, such properties are also associated with others of interest since leaf nitrogen concentration and specific leaf area (SLA) increase to promote photosynthesis and gas exchanges (Rodriguez et al., 2018; Mozo et al., 2024). Furthermore, several types of adaptations have been observed in flood-tolerant willows, including enlarged lenticels, floating or adventitious roots, all of which facilitate gas exchanges otherwise limited by flooding (Kuzovkina et al., 2004; Pezeshki et al., 1996; Jackson and Attwood, 1996; Li et al., 2006; Wang et al., 2016; Fujita et al., 2021). For these reasons, short-rotation coppice (SRC) of Salix miyabeana SX series were previously chosen for an integrative experimental project at pilot scale (Benoist et al., 2023) conducted on a technical landfill site Sainte-Sophie (Québec, Canada), of which the present study is a part. But while those cultivars show strong tolerance to contaminants (Frédette et al., 2019; Grenier et al., 2015) and have proven their relevance in water phytoremediation projects (Guidi Nissim et al., 2014; Frédette et al., 2019; Lévesque et al., 2017; Jerbi et al., 2023b), they originate from exotic provenance. Their use as a monoculture to treat the nitrogen-rich leachate from an inoperative engineered landfill fails to promote local biodiversity and to ensure ecosystem resilience to pests, disease, and environmental changes (Licinio et al., 2023; Massenet et al., 2021). To address this shortcoming, a composite design including native willow species can be envisioned. Indeed, an interesting research hypothesis could be that local willow species may also tolerate these growing conditions and could be an effective alternative that supports both local biodiversity and infrastructure resilience. To this end, the present study was undertaken to compare S. miyabeana ‘SX64’ performance to that of three indigenous willow species from Canada (S. amygdaloides, S. bebbiana, and S. nigra) under controlled conditions mimicking those found in the landfill site, i.e., an especially severe nitrogen overfertilization combined with flooding. Although a variety of contaminants can be found in landfill leachate, the release of nitrogen to the environment represents the greatest challenge to landfill management (Kjeldsen et al., 2002). Yield, plant nutritional status, physiological and morphological parameters were thoroughly investigated to efficiently determine whether native species could match or exceed the performance of the selected cultivar in terms of growth, biomass production, and nitrogen removal efficiency, while also offering added ecological benefits to filtering plantations.
2 Materials and methods
2.1 Experimental design and treatments
The experiment was conducted at the Phytozone, a plant growth research facility at the Montréal Botanical Garden (45°33′43.2″N; 73°34′18.0″W), in a polyethylene grow tunnel. Dormant cuttings (20-cm) of four willow species, S. miyabeana ‘SX64’, obtained from Ramo Inc., a Quebec willow producer, as well as three Canadian indigenous species, S. amygdaloides, S. bebbiana and S. nigra, kindly provided by Verbinnen’s Nursery Ltd. (Ontario, Canada), were planted in plastic bags filled with a mix (1:1) of mature compost and BM6 all-purpose high porosity potting substrate (80% peat moss, 20% perlite; Berger, Quebec, Canada) to allow them to root. After 35 days, 120 willows (30 of each species) were transplanted into 12 L pots filled with high quality plantation soil (pH: 6.4, organic matter: 5.8%, CEC: 23 meq.100 g-1, P: 326 kg ha-1, K: 224 kg ha-1, Ca: 3,849 kg ha-1, Mg: 478 kg ha-1, Al: 579 ppm, Mn: 16.1 ppm, Cu: 3.68 ppm, Zn: 13.25 ppm, B: 1.11 ppm, Fe: 190 ppm; Pépinière Mucci, Quebec, Canada). A bag of green tea (Tetley) and a bag of rooibos tea (Twinings) were weighed and buried in each of the pots at plantation to assess soil microbial activity following the protocol for the tea bag index (TBI) method (Keuskamp et al., 2013).
Willows were exposed to five treatments: (1) adequate watering and fertilization regime (control), (2) nitrogen overfertilization, (3) flooding, (4) combination of flooding and nitrogen overfertilization, and (5) a cyclic regime. A complete randomized block design including the five treatments and the four willow species was set up and replicated in six blocks for a total of 120 pots (5 × 4 × 6 experimental units). Treatments began on 18 June 2020, a week after transplantation, and lasted for 6 weeks.
For the flooding treatments, the pots were placed in pairs in larger containers (40 × 60 × 36 cm) which were filled up to 10 cm below the soil surface with either tap water (flooding treatment) or a fertilizing solution (flooding and nitrogen overfertilization treatment). Each willow receiving the nitrogen overfertilization treatment was exposed to a dose equivalent to 800 kg of N.hectare-1.season−1 in the form of an ammonium sulphate (NH4)2 SO4) solution (250 mg of N.l-1) without any other form of fertilization. ‘Cyclic treatment’ was applied over a 2-week cycle. During the first week, the willows were flooded and fertilized with nitrogen following the same procedure as for the ‘flooding and nitrogen overfertilization’ treatment. The second week, the liquid was emptied from the containers and no nitrogen was applied. The removed water was stored in a cold room and reused to refill the containers the following week.
2.2 Data collection
Plant height increment was measured weekly throughout the experiment. The presence or absence of upward growing roots, adventitious roots and hypertrophied lenticels, which are morphological characteristics associated to flooding (Shuwen et al., 2006), was noted during the last week of the experiment while teabags, roots, stems and leaves were harvested.
Total leaf area was measured using a LI-3100C Area Meter. Biomass weight was measured after the entire root systems and aerial parts of all 120 plants were washed and oven-dried (70 °C, 72 h) (Hamilton Cabinet Dryer). One leaf per plant was weighed individually before being dried to determine the fresh weight. This leaf was then reweighed and dried to determine the dry mass and thus calculate the leaf water content (WC). For nutrient content analysis, sub-samples were sent to a certified laboratory (Agroenvirolab, LaPocatière, Québec, Canada, accredited by CEAEQ and ISO-CEI 17025). The foliar content of total P, K, Ca, Mg, and S was analyzed by ICP-OES (Carter and Gregorich, 2007). Total nitrogen was determined by combustion analysis (CHNOS Elemental Analyzer Vario Micro Select; Elementar Analyse systeme GmbH, Hanau, Germany). Samples of the water contained in the tanks (leachate) were taken at the very end of the experiment and sent to an accredited laboratory (H2Lab, Quebec, Canada) to measure total nitrogen. An approximation of the total amount of foliar nitrogen removal (kg.ha-1) envisioned for the four willow species according to the different treatments was calculated by multiplying the leaf dry weight by the percentage of leaf N. We then cautiously multiplied the result by three, since the experiment lasted only 6 weeks (whereas full growing season lasts >6 months in Québec, eastern Canada) and by 16,000, the usual number of trees planted per hectare in conventional SRC willow plantation. Such a calculation, although imperfect, is intended as a proxy for the first-year potential of cuttings at maximum growth rate (4 months of spring and summer) and therefore when fertigation could be first planned in situ. The potential for total above-ground filtration and root nitrogen sequestration were not considered in this study since the stem and root nitrogen contents were not measured.
2.3 Data analysis
Statistical analyses were performed using R version 3.6.3. The significance level of the analyses was set at α = 0.05. Linear mixed models (nlme library) were built for each of the components under study. The “block” variable was used as a random intercept in all models. The “species” and “treatment” variables were used as fixed variables in all models. The significance of the interaction was verified using ANOVA followed by a Tukey’s test. For all tests, differences were considered to be significant at P ≤ 0.05.
3 Results
3.1 Plant growth and biomass production differ between species and nitrogen fertigation regimes
Throughout the experiment, plant growth patterns were stable without reaching a plateau regardless of willow species (Figure 1A) but ‘nitrogen overfertilization’ quickly induced the greatest growth with Salix miyabeana trees reaching almost 2 m after only 6-week (dark blue lines, p < 0.001). When treatment included ‘flooding’ (different shade of green), the height of S. miyabeana, S. amygdaloides and S. nigra with or without nitrogen application did not significantly differ from ‘control’ (purple lines) whether overfertilization occurred. The growth of S. bebbiana was much more severely impacted by inundation with its height being 50% lower (i.e., < 0.4 m; p < 0.05) than that of ‘control’. Individuals of all species in the cyclic treatment (lightest green lines) had equal or greater growth than individuals in other treatments with flooding.
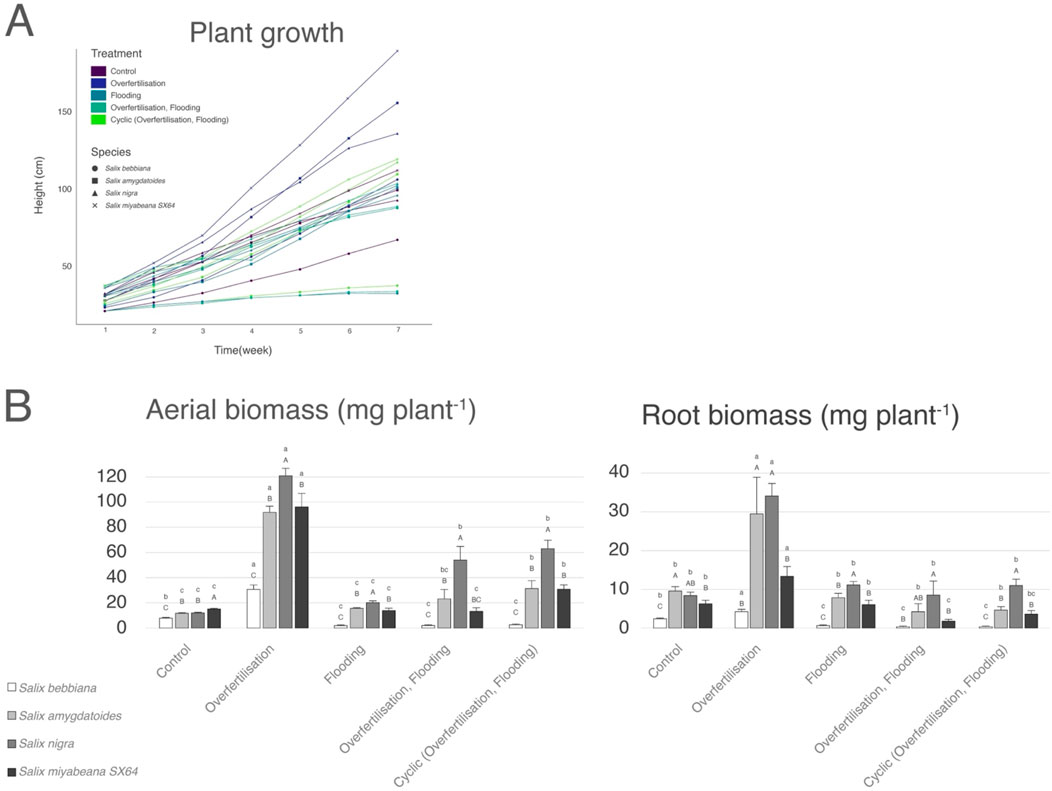
Figure 1. Willow growth pattern and overall biomass production in response to different fertigation conditions. (A) Height of cuttings of four willow species was monitored over a 6-week long experiment. Plant height is expressed in cm. (B) Average aerial and root biomass (in mg) of the four willow species at the end of the experiment. Values are mean + - SD. Different lower-case letters indicate significant difference between treatments for the investigated species (p < 0.05). Different capital letters indicate significant difference between species in response to a specific treatment (p < 0.05).
Total dry biomass results differed from those reported above for growth increment. Under all treatment conditions except control, S. nigra performed much better than other species and S. bebbiana was the least productive (Figure 1B); p < 0.05, different capital letters indicate significant difference between species exposed to the same treatment). All trees benefited from nitrogen overfertilization. ‘Flooding + N’ treatment’ conditions were also beneficial to S. nigra, S. miyabeana and S. amygdaloides but the root biomass of S. miyabeana was particularly impacted by fertigation including the ‘cyclic’ regime that may better simulate an operational vegetative filter in which treatment could be intermittent (Figure 1B, left panel).
3.2 Nitrogen phytofiltration conditions alter willows in a species-dependent manner
In addition to plant growth parameters mentioned above, a few morphometrics, anatomical, and nutritional variables were studied to evaluate the potential of the different willow species to adapt to the artificially created environment of a filtering plantation.
In comparison to ‘control’, root-to-shoot ratio decreased in the presence of excessive nitrogen for all plant species, although the ‘cyclic’ treatment slightly alleviated this tendency for S. amygdaloides and S. nigra (Figure 2A). Regarding more specific physiological manifestations of plasticity at the leaf level, no comparisons were made between species but their respective response profiles to the different fertigation treatments were recorded. Neither the results for total leaf area nor those for leaf water content were like those of biomass (Figure 2B) and the specific behavior of S. nigra was highlighted. For total leaf area (left panel), a maximum was observed in the ‘overfertilization’ conditions for all species. While S. miyabeana and S. amygdaloides exhibited rather similar patterns (i.e., more leaves in ‘nitrogen overfertilisation’ condition but not when flooded), S. nigra leaf area remained high in the mesocosms that copied vegetative filters: ‘flooding + overfertilization’ and ‘cyclic’ treatments. On the contrary, both leaf area and leaf water content parameters were indicative of S. bebbiana impairment when flooded (leaf water content was as low as 44% in cyclic conditions) while, for the later, that of S. nigra remained invariable between treatments including ‘control’ (see Figure 2B right panel, RWC remained slightly above 70% in all conditions while S. miyabeana and S. amygdaloides was much more plastic in presence of ‘nitrogen’ but not when flooded). Leaf nutrient content was also impacted by the different treatments in a species-dependant manner (Table 1). While foliar potassium and magnesium contents were not altered for any species, the nutrient status of the plants was logically modified by the different treatments. While potassium and magnesium content varied very little regardless treatment or plant species, phosphorus and calcium monitoring more specifically revealed some disorders, especially for the leaves of S. bebbiana and S. miyabeana, which were respectively much more affected by flooding and/or the combination of flooding and nitrogen overfertigation. Calcium content dropped by about half in ‘flooding’ and ‘overfertilization + flooding’ for S. bebbiana and that number was even worst for S. miyabeana under ‘overfertilization + flooding’. On the other hand, the data collected for S. amygdaloides and S. nigra shows much greater stability, with values even much closer to the ones found in control condition for the cyclic treatment.
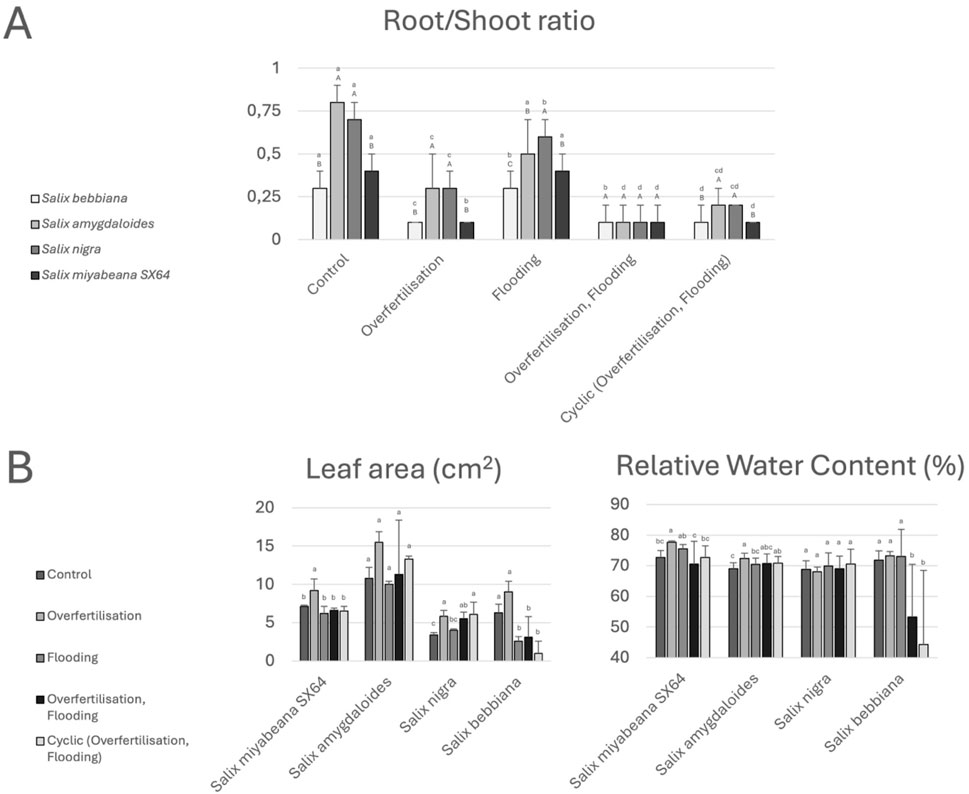
Figure 2. Morphometrics parameters of willow affected by different fertigation regimes. (A) Root to Shoot biomass ratio of the four willow species at the end of the experiment. (B) Leaf-related parameters (total leaf in cm2 and leaf relative water content in %). Values are mean + - SD. Different lower-case letters indicate significant difference between treatments for the investigated species (p < 0.05).
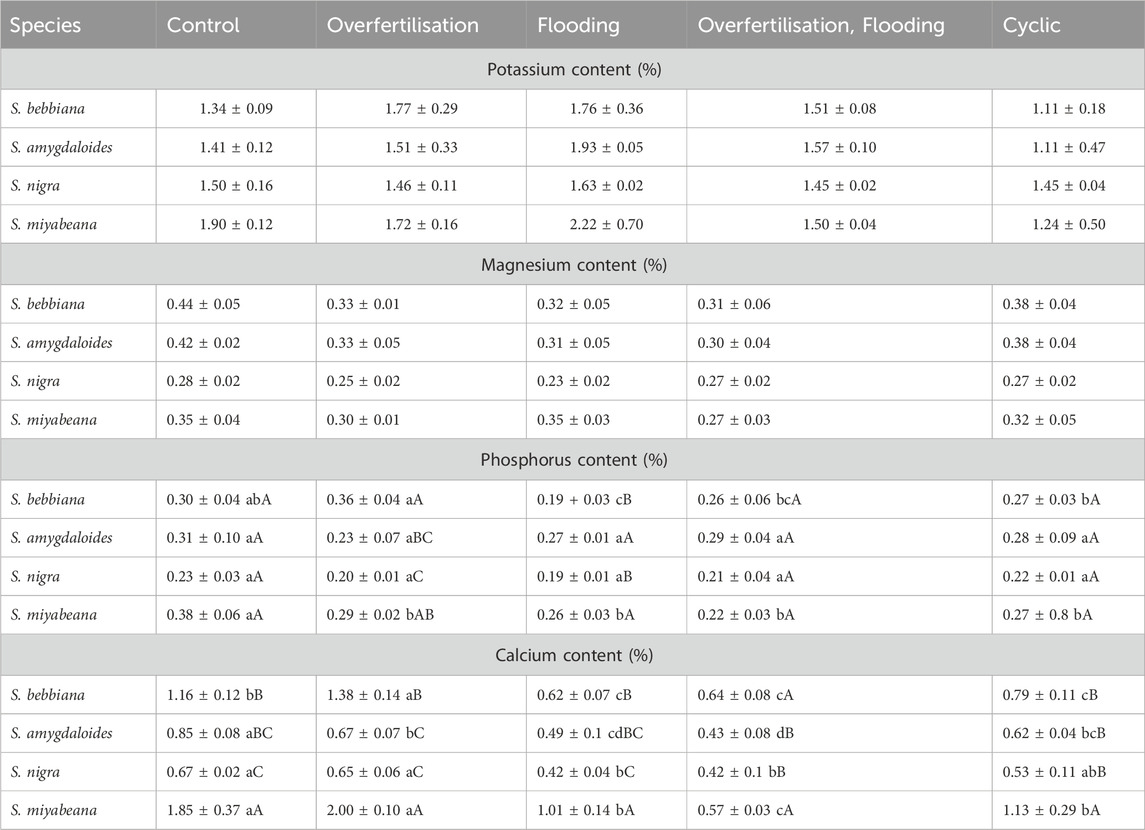
Table 1. Willow leaf nutrient content of the four willow species in response to different fertigation conditions. Potassium, magnesium, phosphorus and calcium leaf content (in percentage) of the four willow species and according to the different fertigation regimes. Values are mean + - SD. Different lower-case letters indicate significant difference between treatments for the investigated species (line; p < 0.05). Different capital letters indicate significant difference between species in response to a specific treatment (column; p < 0.05).
At the root and stem level (Figure 3), the presence of adventitious and upward growing roots as well as lenticels was recorded throughout the experiment. S. bebbiana was the species for which the least number of individuals exhibited all these characteristics (Figure 3, see lightest grey bars on left panel). In general, adventitious roots were the feature least observed (Figure 3 right panel, top block), while lenticels were the most common (Figure 3 right panel, bottom block), especially under flooded conditions (with or without overfertilization). Only some individuals of S. miyabeana and S. nigra developed adventitious roots in the cyclic treatment. Specimens of all species developed vertical roots, especially in ‘flooding’ and ‘cyclic’ conditions. Vertical roots were found in all individuals of S. amygdaloides in all treatments except ‘flooding + nitrogen’. Lenticels were almost systematically present on stems of S. miyabeana regardless of treatment conditions, but not in ‘control’ conditions for the other three species.
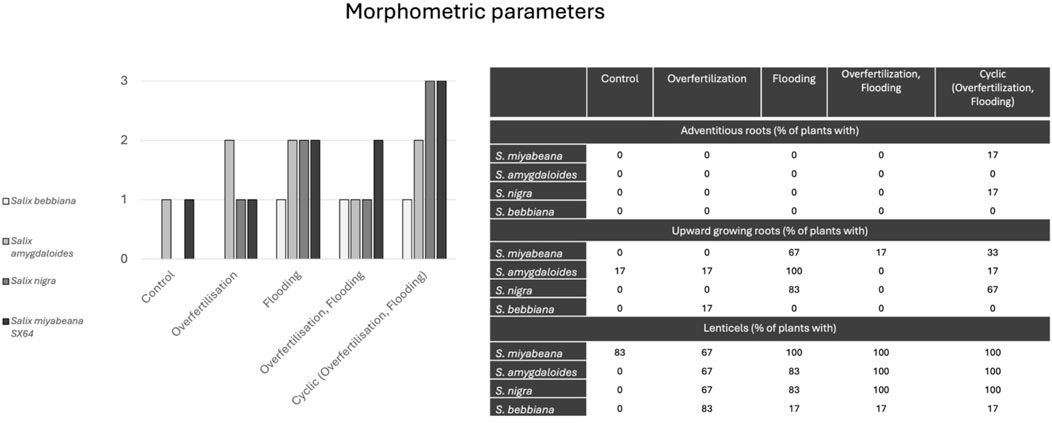
Figure 3. Anatomical parameters of willow affected by different fertigation regimes. Different morphological features such as adventitious roots, upright growing roots and lenticels were observed on roots and stems of willow exposed to different fertigation regimes on stems and roots according to the various treatments. Data summary on the left panel and observation details (%) on the right panel. No statistical test was performed.
3.3 The filtering potential of local willow species
We investigated different components of the treatment potential of the envisioned vegetative filter which are summarized in Figure 4. We used the Tea Bag Index method to evaluate the carbon storage potential of willows (Figure 4A). While plant species did not significantly influence the stabilization factor, treatment did, with a very low level of stabilization under ‘control’ and ‘overfertilization’ regimes. A contrario, ‘flooding’ (with or without addition of nitrogen) showed the highest potential, and ‘cyclic’ conditions yielded intermediate values. For all species treated with an excess of nitrogen except S. bebbiana, N concentration in leaves was high, above 3% under such conditions (<2% under ‘control’ condition). Together with biomass data. These measurements enabled us to estimate the amount of nitrogen taken up by the willows (Figure 4B), i.e., >30 kg/ha for S. bebbiana, >75 kg/ha for S. miyabeana, S. amygdaloides and S. nigra under ‘N overfertilization’ conditions. The removal potential of S. nigra was comparable in all treatments that included nitrogen. No such level was obtained for the other species. For S. miyabeana, the data calculated for the ‘cyclic’ treatment was higher than for the other two treatments including flooding and ‘control’. For S. amygdaloides and S. bebbiana, no increase in nitrogen uptake under flooding with or without overfertilization was calculated.
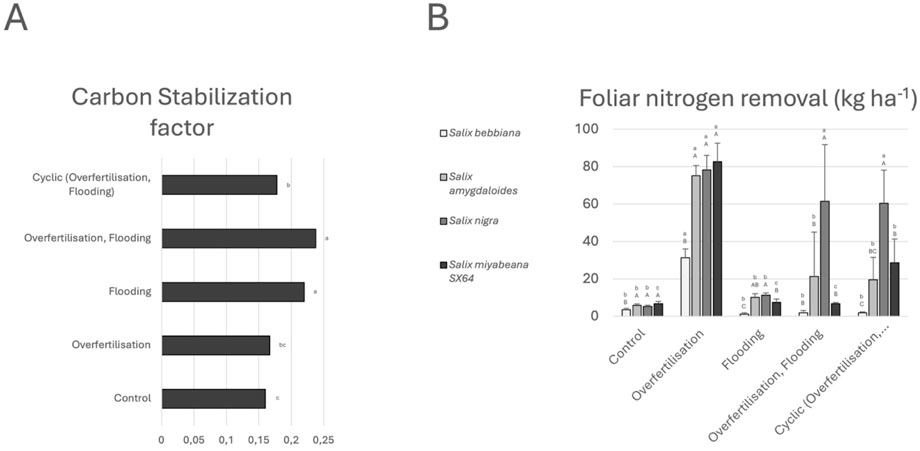
Figure 4. Treatment potential of willow plantations exposed to different fertigation regimes. (A) Rhizospheric carbon stabilisation factor according to the different fertigation regimes. No difference was measured depending on willow species, different lower-case letters indicate significant difference between fertigation regimes (p < 0.05). (B) Total amount of foliar nitrogen removal (kg ha-1) of the four willow species exposed to different fertigation regime. Values are mean + - SD. Different lower-case letters indicate significant difference between treatments for the investigated species (p < 0.05). Different capital letters indicate significant difference between species in response to a specific treatment (p < 0.05) performed.
4 Discussion
The promising results obtained during this experiment clearly indicate that integrating native plant species into vegetative filters established on landfill sites yields extensive benefits with no reduction in leachate treatment efficiency. While the implementation of such green infrastructure on landfill sites is already a good step forward, including adapted biodiversity elements and growing practices supporting the resilience of the entire ecosystem seems to be within reach. To do this, short experiments under controlled conditions accelerate innovation and adoption at the pace required by current ecological challenges and address the needs of the stakeholders involved (Guidi Nissim and Labrecque, 2021; Licinio et al., 2023; Nadeau et al., 2018a; Nadeau et al., 2018b; Markus-Michalczyk et al., 2020). Thus, the work presented here, which took place over a single growing season, established a protocol for species selection, and allows us to narrow down at least one new species that shows greater potential than Salix miyabeana for this context: S. nigra; and also, to outline a viable alternative irrigation protocol for phytofiltration: ‘cyclic’.
4.1 Impact of leachate irrigation on plant development
At first glance, all species benefited from nitrogen overfertilization with clear biomass gain, especially aboveground. Reduced root-to-shoot ratios were thus observed, which concurs with findings in the literature (Jerbi et al., 2015; Jerbi et al., 2020; Liu and Dickmann, 1992). In flooded conditions, however, opposite profiles were observed. Results for Salix bebbiana, a species that grows naturally in more xeric habitats (Isebrands and Richardson, 2013), contrast with those of the other tested species, showing both reduced growth and almost no adaptive traits such as upward growing roots, lenticels or longer stomata (Cameron et al., 2010; Jackson and Attwood, 1996; Jerbi et al., 2020; Kozlowski, 1997; Kuzovkina et al., 2004; Li et al., 2006; Pereira and Kozlowski, 1977; Wang et al., 2017). The biomass of more wetland-adapted willow species was only slightly affected, and apart from a nutrient imbalance at leaf level of S. miyabeana that could be indicative of stress response and alter long-term growth if no amendment is considered (Barak et al., 1997; Kozlowski, 1997), biomass distribution remained constant which is promising under the conditions herein reproduced.
A general pattern of response to leachate irrigation (the combined ‘flooding + overfertilization’ treatments) emerged for all three species adapted to riparian environments that clearly benefit from this treatment. Salix miyabeana, S. amygdaloides and S. nigra, performed equally well, or even better when irrigated, than in the ‘control’ conditions. These more flood-tolerant species had greater aboveground biomass production and a higher leaf surface area to maximize transpiration, thus resulting in increased foliar concentration of the applied nitrogen. Similar results were observed in Weih and Nordh (2002) when they tested different irrigation and fertilization regimes on fourteen willow species.
4.2 Phytotreatment with indigenous willow species
One of the objectives of the present study was to compare the tested species native to Canada to S. miyabeana ‘SX64’, which was expected to be the best performing species under leachate irrigation as it has been used frequently in nitrogen-rich water treatment with a high success rate (Guidi Nissim et al., 2014; Guidi Nissim et al., 2015; Jerbi et al., 2020; Lachapelle-T et al., 2019; Mirck and Volk, 2010; Sas et al., 2021). However, a similar or higher level of productivity was observed for well-adapted indigenous species in the present study.
In particular, S. nigra, recognized as a species well adapted to wetlands (Mosseler et al., 2017), displayed an interesting plasticity in physiological and morphological characteristics, as described above, and the most conservative isohydric behavior with no change in leaf water content (Sade et al., 2012). It was the most productive when nitrogen supply was maximized under flooded conditions. Furthermore, changes in its nutritional status also remained relatively buffered compared to others, although of course there was an overall increase in foliar N content, a good indicator of the plant’s ability to take up nitrogen from the substrate (Simon et al., 1990). The differences highlighted here show a clear distinction in response among willow species. S. nigra shows a wide range of adaptive traits that allow the conception of more versatile leachate phytofiltration components for engineered ecological landscaping.
4.3 Leachate application strategy and environmental applicability
The results from soil and leachate monitoring in this study highlight the need for a large-scale testing trial of these indigenous willow species using an intermittent ‘cyclic’ fertigation regime. In this experiment we tested two leachate application procedures and found that the ‘cyclic’ treatment was as promising as a conventional permanent fertigation regime in regard to nitrogen filtration potential. Of course, a significant amount of nitrogen-loaded water was leached at the end of the experiment (eq. > 200 kg ha-1 more in ‘flooding + overfertilization’ than in ‘flooding’ conditions), but plants were still in a juvenile phase with no mature root system to fully colonize the substrate and capture the available nutrient stock (Wang et al., 2017). Furthermore, the nitrogen concentration reported in leaves was indicative of an optimal nutrient supply (Kopinga and van den Burg, 1995) and consistent with the values reported in the literature (Aronsson and Perttu, 2001; Hofmann-Schielle et al., 1999), as was the projected nitrogen removal rate (Fabio and Smart, 2018), which could rank close to the >400 kg ha-1 estimates by Labrecque and Teodorescu (2001) when aboveground biomass is more realistically harvested at 2–3 years of age after being adequately fertilized.
5 Conclusion
Both Salix miyabeana ‘SX64’ and Canadian indigenous willows S. nigra and S. amygdaloides were able to adapt to nitrogen overfertilization and flooding while removing nitrogen from leachate, without experiencing any reduction in biomass or growth. Indeed, biomass of S. nigra even increased in the combined flooding and nitrogen treatments. That indigenous species of Canada also demonstrated several features adaptive to flooding, such as development of adventitious and upward growing roots, and lenticels. Regarding its phytoremediation capacities, it stood out for the amount of nitrogen stored in its leaf biomass, which was significantly higher than that of the other species tested. When comparing leachate application techniques, intermittent application had more beneficial effects on willow growth than conventional fertigation. Additional morphological adaptations were observed in this treatment. The total amount of nitrogen contained in the willow leaves and the amount of nitrogen remaining in the leachate at the end of the experiment did not differ between application methods, hence broadening the options for future use in engineered ecological landscaping. Pilot-scale experimentation would be necessary to confirm that these results persist over the long term.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
CA: Methodology, Conceptualization, Investigation, Formal Analysis, Writing – original draft, Visualization, Software, Data curation. ML: Resources, Writing – review and editing, Funding acquisition, Project administration, Formal Analysis, Methodology, Conceptualization, Validation, Supervision, Data curation. CS: Conceptualization, Investigation, Methodology, Formal Analysis, Software, Visualization, Data curation, Writing – original draft. JL: Formal Analysis, Supervision, Conceptualization, Software, Writing – review and editing, Methodology, Visualization, Investigation, Project administration, Validation, Data curation, Resources.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study received financial support from a NSERC Collaborative Research and Development Grant (RDCPJ-525307-18) and a provincial CRIBIQ Grant (2017-079-C35).
Acknowledgments
We would like to thank Waste Management Québec Inc. for its financial and technical support of this project. Thanks also to Yves Comeau and Karen Grislis for critical review of the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aronsson, P., and Perttu, K. (2001). Willow vegetation filters for wastewater treatment and soil remediation combined with biomass production. For. Chron. 77, 293–299. doi:10.5558/tfc77293-2
Barak, P., Jobe, B. O., Krueger, A. R., Peterson, L. A., and Laird, D. A. (1997). Effects of long-term soil acidification due to nitrogen fertilizer inputs in Wisconsin. Soil 197, 61–69. doi:10.1023/a:1004297607070
Benoist, P., Parrott, A., Lachapelle-T, X., Barbeau, L. C., Comeau, Y., Pitre, F. E., et al. (2023). Treatment of landfill leachate by short-rotation willow coppice plantations in a large-scale experiment in eastern Canada. Plants (Basel) 12 (2), 372. doi:10.3390/plants12020372
Burton, S. A. Q., and Watson-Craik, I. A. (1998). Ammonia and nitrogen fluxes in landfill sites: applicability to sustainable landfilling. Waste Manag. Res. 16, 41–53. doi:10.1177/0734242x9801600106
Cameron, K., Smart, L., Ballard, B., Volk, T., and Abrahamson, L. (2010). Salix miyabeana ‘SX64’, Willowpedia fact sheet. Cornell University.
M. R. Carter, and E. G. Gregorich (2007). Soil sampling and methods of analysis. 2nd ed. (Boca Raton, FL: CRC Press).
Fabio, E. S., and Smart, L. B. (2018). Effects of nitrogen fertilization in shrub willow short rotation coppice production - a quantitative review. GCB Bioenergy 10, 548–564. doi:10.1111/gcbb.12507
Frédette, C., Comeau, Y., and Brisson, J. (2019). Ecophysiological responses of a willow cultivar (Salix miyabeana ‘SX67’) irrigated with treated wood leachate. Water, Air, and Soil Pollut. 230, 205. doi:10.1007/s11270-019-4244-4
Fujita, S., Noguchi, K., and Tange, T. (2021). Different waterlogging depths affect spatial distribution of fine root growth for Pinus thunbergii seedlings. Front. Plant Sci. 12, 614764. doi:10.3389/fpls.2021.614764
Grenier, V., Pitre, F., Guidi Nissim, W., and Labrecque, M. (2015). Genotypic differences explain most of the response of willow cultivars to petroleum-contaminated soil. Trees 29, 871–881. doi:10.1007/s00468-015-1168-5
Guidi Nissim, W., and Labrecque, M. (2021). Reclamation of urban brownfields through phytoremediation: implications for building sustainable and resilient towns. Urban For. and Urban Green. 65, 127364. doi:10.1016/j.ufug.2021.127364
Guidi Nissim, W., Voicu, A., and Labrecque, M. (2014). Willow short-rotation coppice for treatment of polluted groundwater. Ecol. Eng. 62, 102–114. doi:10.1016/j.ecoleng.2013.10.005
Guidi Nissim, W., Jerbi, A., Lafleur, B., Fluet, R., and Labrecque, M. (2015). Willows for the treatment of municipal wastewater: performance under different irrigation rates. Ecol. Eng. 81, 395–404. doi:10.1016/j.ecoleng.2015.04.067
Hao, T., Zhu, Q., Zeng, M., Shen, J., Shi, X., Liu, X., et al. (2020). Impacts of nitrogen fertilizer type and application rate on soil acidification rate under a wheat-maize double cropping system. J. Environ. Manage 270, 110888. doi:10.1016/j.jenvman.2020.110888
Hofmann-Schielle, C., Jug, A., Makeschin, F., and Rehfuess, K. E. (1999). Short-rotation plantations of balsam poplars, aspen and willows on former arable land in the Federal Republic of Germany. I. Site–growth relationships. For. Ecol. Manag. 121, 41–55. doi:10.1016/s0378-1127(98)00555-6
Isebrands, J. G., and Richardson, J. (2013). Poplars and willows: trees for society and the environment. Rome, Italy: CABI; FAO.
Jackson, M. B., and Attwood, P. A. (1996). Roots of willow (Salix viminalis L.) show marked tolerance to oxygen shortage in flooded soils and in solution culture. Plant Soil 187, 37–45. doi:10.1007/978-94-011-5696-7_4
Jerbi, A., Guidi Nissim, W., Fluet, R., and Labrecque, M. (2015). Willow root development and morphology changes under different irrigation and fertilization regimes in a vegetation filter. Bioenerg. Res. 8, 775–787. doi:10.1007/s12155-014-9550-5
Jerbi, A., Brereton, N. J. B., Sas, E., Amiot, S., Lachapelle, T. X., Comeau, Y., et al. (2020). High biomass yield increases in a primary effluent wastewater phytofiltration are associated to altered leaf morphology and stomatal size in Salix miyabeana. Sci. Total Environ. 738 (2020), 139728. doi:10.1016/j.scitotenv.2020.139728
Jerbi, A., Kalwahali-Muissa, M., Krygier, R., Johnston, C., Blank, M., Sarrazin, M., et al. (2023a). Comparative wood anatomy, composition and saccharification yields of wastewater irrigated willow cultivars at three plantations in Canada and Northern Ireland. Biomass Bioenergy 170, 106683. doi:10.1016/j.biombioe.2022.106683
Jerbi, A., Laur, J., Lajoie, K., Gallant, P. P., Barnabé, S., Pitre, F. E., et al. (2023b). Irrigation with primary wastewater alters wood anatomy and composition in willow Salix miyabeana SX67. Front. Plant Sci. 14, 1087035. doi:10.3389/fpls.2023.1087035
Justin, M. Z., Pajk, N., Zupanc, V., and Zupančič, M. (2010). Phytoremediation of landfill leachate and compost wastewater by irrigation of Populus and Salix: biomass and growth response. Waste Manag. 30 (6), 1032–1042. doi:10.1016/j.wasman.2010.02.013
Keuskamp, J. A., Dingemans, B. J. J., Lehtinen, T., Sarneel, J. M., and Hefting, M. M. (2013). Tea Bag Index: a novel approach to collect uniform decomposition data across ecosystems. Methods Ecol. Evol. 4, 1070–1075. doi:10.1111/2041-210x.12097
Kjeldsen, P., Barlaz, M. A., Rooker, A. P., Baun, A., Ledin, A., and Christensen, T. H. (2002). Present and long-term composition of MSW landfill leachate: a review. Crit. Rev. Environ. Sci. Technol. 32, 297–336. doi:10.1080/10643380290813462
Kopinga, J., and van den Burg, J. (1995). Using soil and foliar analysis to diagnose the nutritional status of urban trees. Arboriculture Urban Forestry. 21 17–25. doi:10.48044/jauf.1995.004
Kozlowski, T. T. (1997). Responses of woody plants to flooding and salinity. Tree Physiol. 17, 490. doi:10.1093/treephys/17.7.490
Kuzovkina, Y. A., Knee, M., and Quigley, M. F. (2004). Effects of soil compaction and flooding on the growth of 12 willow (Salix L.) species. J. Environmental Horticulture. 7. doi:10.24266/0738-2898-22.3.155
Kuzovkina, Y. A., Weih, M., Romero, M. A., Charles, J., Hust, S., McIvor, I., et al. (2008). “Salix: botany and global horticulture,” in Horticultural reviews. Editor J. Janick (Hoboken, NJ, USA: John Wiley and Sons, Inc.), 447–489.
Labrecque, M., and Teodorescu, T. I. (2001). Influence of plantation site and wastewater sludge fertilization on the performance and foliar nutrient status of two willow species grown under SRIC in southern Quebec (Canada). For. Ecol. Manag. 150, 223–239. doi:10.1016/s0378-1127(00)00567-3
Lachapelle, T. X., Labrecque, M., and Comeau, Y. (2019). Treatment and valorization of a primary municipal wastewater by a short rotation willow coppice vegetation filter. Ecol. Eng. 130, 32–44. doi:10.1016/j.ecoleng.2019.02.003
Lévesque, S., Demers, E., Brisson, J., and Comeau, Y. (2017). Treatment of a mixed wood preservative leachate by a hybrid constructed wetland and a willow planted filter. Water Sci. Technol. 76, 164–171. doi:10.2166/wst.2017.197
Li, S., Reza Pezeshki, S., and Douglas, F. (2006). Shields, partial flooding enhances aeration in adventitious roots of black willow (Salix nigra) cuttings. J. Plant Physiology 163, 619–628. doi:10.1016/j.jplph.2005.06.010
Licinio, A., Laur, J., Pitre, F. E., and Labrecque, M. (2023). Willow and herbaceous species’ phytoremediation potential in Zn-contaminated farm field soil in eastern Québec, Canada: a greenhouse feasibility study. Plants 12 (1), 167. doi:10.3390/plants12010167
Liu, Z., and Dickmann, D. I. (1992). Responses of two hybrid Populus clones to flooding, drought, and nitrogen availability. I. Morphology and growth. Can. J. Bot. 70, 2265–2270. doi:10.1139/b92-281
Markus-Michalczyk, H., Crawford, M. L., and Baldwin, A. H. (2020). Salinity tolerance and occurrence of Salix nigra marshall (black willow) in tidal wetlands of chesapeake bay tributaries. Northeast. Nat. 27 (2), 229–241. doi:10.1656/045.027.0205
Massenet, A., Bonet, A., Laur, J., and Labrecque, M. (2021). Co-planting Brassica napus and Salix nigra as a phytomanagement alternative for copper contaminated soil. Chemosphere 279, 130517. doi:10.1016/j.chemosphere.2021.130517
Mirck, J., and Volk, T. A. (2010). Response of three shrub willow varieties (Salix spp.) to storm water treatments with different concentrations of salts. Bioresour. Technol. 101, 3484–3492. doi:10.1016/j.biortech.2009.12.128
Mosseler, A., Major, J. E., and Ostaff, D. (2017). Distribution of genetic variation in five coppice growth traits among natural populations of seven North American willow (Salix) species. Can. J. For. Res. 47 (1), 36–46. doi:10.1139/cjfr-2016-0307
Mozo, I., Monteoliva, S. E., Cerrillo, T., and Luquez, V. M. C. (2024). Flooding tolerance, biomass production, and leaf nitrogen assimilatory efficiency in 29 diverse willows (Salix spp.) genotypes during early growth. J. For. Res. 29 (6), 458–465. doi:10.1080/13416979.2024.2386704
Muklada, H., Fabio, E. S., and Smart, L. B. (2022). Growth, nitrogen uptake, and nutritional value of a diverse panel of shrub willow (Salix spp.) Genotypes in response to nitrogen fertilization. Agronomy 12 (11), 2678. doi:10.3390/agronomy12112678
Nadeau, M. N., Laur, J., and Khasa, D. P. (2018a). Mycorrhizae and rhizobacteria on precambrian rocky gold mine tailings: I. Mine-adapted symbionts promote white spruce health and growth. Front. Plant Sci. 9, 1267. doi:10.3389/fpls.2018.01267
Nadeau, M. N., Laur, J., and Khasa, D. P. (2018b). Mycorrhizae and rhizobacteria on precambrian rocky gold mine tailings: II. Mine-adapted symbionts alleviate soil element imbalance for a better nutritional status of white spruce seedlings. Front. Plant Sci. 9, 1268. doi:10.3389/fpls.2018.01268
Newton, K., Gonzalez, E., Pitre, F. E., and Brereton, N. J. B. (2022). Microbial community origin and fate through a rural wastewater treatment plant. Environ. Microbiol. 24, 2516–2542. doi:10.1111/1462-2920.16025
Pereira, J. S., and Kozlowski, T. T. (1977). Variations among woody angiosperms in response to flooding. Physiol. Plant 41, 184–192. doi:10.1111/j.1399-3054.1977.tb05555.x
Pezeshki, S. R., Pardue, J. H., and DeLaune, R. D. (1996). Leaf gas exchange and growth of flood-tolerant and flood-sensitive tree species under low soil redox conditions. Tree Physiol. 16, 453–458. doi:10.1093/treephys/16.4.453
Pilon-Smits, E. (2005). Phytoremediation. Plant Biol. 56, 15–39. doi:10.1146/annurev.arplant.56.032604.144214
Rodriguez, M. E., Doffo, G. N., Cerrillo, T., and Luquez, V. M. C. (2018). Acclimation of cuttings from different willow genotypes to flooding depth level. New For. 49, 415–427. doi:10.1007/s11056-018-9627-7
Sade, N., Gebremedhin, A., and Moshelion, M. (2012). Risk-taking plants: anisohydric behavior as a stress-resistance trait. Plant Signal. and Behav. 7, 767–770. doi:10.4161/psb.20505
Sas, E., Hennequin, L. M., Frémont, A., Jerbi, A., Legault, N., Lamontagne, J., et al. (2021). Biorefinery potential of sustainable municipal wastewater treatment using fast-growing willow. Sci. Total Environ. 792, 148146. doi:10.1016/j.scitotenv.2021.148146
Shuwen, L. I., Pezeshki, S., and Shields, J., F. (2006). Partial flooding enhances aeration in adventitious roots of black willow (Salix nigra) cuttings. J. plant physiology 163, 619–628. doi:10.1016/j.jplph.2005.06.010
Simon, M., Zsuffa, L., and Burgess, D. (1990). Variation in N, P, and K status and N efficiency in some North American willows. Can. J. For. Res. 20, 1888–1893. doi:10.1139/x90-253
Song, H., Che, Z., Cao, W., Huang, T., Wang, J., and Dong, Z. (2016). Changing roles of ammonia-oxidizing bacteria and archaea in a continuously acidifying soil caused by over-fertilization with nitrogen. Environ. Sci. Pollut. Res. 23, 11964–11974. doi:10.1007/s11356-016-6396-8
Wang, C., Li, C., Wei, H., Xie, Y., and Han, W. (2016). Effects of long-term periodic submergence on photosynthesis and growth of Taxodium distichum and Taxodium ascendens saplings in the hydro-fluctuation zone of the Three Gorges Reservoir of China. PLoS One 11 (9), e0162867. doi:10.1371/journal.pone.0162867
Wang, C., Xie, Y., He, Y., Li, X., Yang, W., and Li, C. (2017). Growth and physiological adaptation of Salix matsudana Koidz. to periodic submergence in the hydro-fluctuation zone of the Three Gorges dam reservoir of China, 13. 283. doi:10.3390/f8080283
Weih, M., and Nordh, N.-E. (2002). Characterising willows for biomass and phytoremediation: growth, nitrogen and water use of 14 willow clones under different irrigation and fertilisation regimes. Biomass Bioenergy 23 (6), 397–413. doi:10.1016/s0961-9534(02)00067-3
Keywords: contaminated water, flooding, landfill leachate, phytotechnology, Salix, ecophysiology
Citation: Auger C, Labrecque M, Susini C and Laur J (2025) The relevance of Eastern Canadian native willows as alternatives to Salix miyabeana in nitrogen leachate-treating vegetative filters. Front. Environ. Sci. 13:1599955. doi: 10.3389/fenvs.2025.1599955
Received: 25 March 2025; Accepted: 17 September 2025;
Published: 03 October 2025.
Edited by:
Aleksandra Wdowczyk, Wroclaw University of Environmental and Life Sciences, PolandReviewed by:
Aneta Spyra, University of Silesia in Katowice, PolandAnshu Gupta, University of Jammu, India
Yanjiao Gao, Liaoning University of Technology, China
Copyright © 2025 Auger, Labrecque, Susini and Laur. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Joan Laur, am9hbi5sYXVyQHVtb250cmVhbC5jYQ==