 Xizhe Zhang
Xizhe Zhang Yanhui Ye†
Yanhui Ye†- Resources and Environment College, Xizang Agriculture and Animal Husbandry University, Nyingchi, Tibet, China
Introduction: As an important global ecological security barrier, the Qinghai-Tibet Plateau is a key region for exploring how global climate change affects the grassland ecosystem. Therefore, this study explored the mechanism of the impact of intensified atmospheric Phosphorus (P) deposition on the alpine grassland plant communities in Sejila Mountain, Nyingchi, Tibet.
Methods: A field experiment with four different gradients of phosphate fertilizer application (0, 50, 75, and 100 kg hm−2 a−1) was designed. The variation patterns of plant morphology, biomass, nutrient content, and stoichiometric ratio characteristics in response to P were systematically analyzed.
Results: P addition significantly affected the total community biomass and root-shoot ratio, showing a trend of initial increase followed by decline (P < 0.05). Additionally, P addition significantly influenced the plant traits (average height, total coverage, and abundance) and aboveground biomass of plant community (Poaceae, Cyperaceae, and Forbs). The plant traits and aboveground biomass of Poaceae and Forbs increased significantly with increasing of P addition levels. At a P addition of 100 kg hm−2 a−1, a decline in these parameters was observed. The total coverage and aboveground biomass of Cyperaceae plants showed a significant downward trend. P addition had limited effects on plant carbon (C) and nitrogen (N) contents and their respective stoichiometric ratios. However, it significantly increased P contents in both aboveground and belowground plant parts (P < 0.05), consequently reducing the C:P and N:P stoichiometric ratios in plants. This effectively enhanced plant P use efficiency.
Discussion: This study highlights the significant role of P addition in shaping the plant community structure and nutrient cycling of alpine grasslands. However, excessive P addition may exacerbate ecological competition among plants, potentially leading to nutritional imbalances and soil environmental degradation.
1 Introduction
Phosphorus (P) deposition encompasses contributions from both natural and anthropogenic sources. As a critical nutrient for plant growth, P plays an indispensable role in ecosystem functioning by influencing plant productivity, carbon sequestration, and nutrient cycling through atmospheric deposition (Dissanayaka et al., 2020). Atmospheric P deposition has gained increasing attention due to its dual natural and human-driven origins, including weathering processes, volcanic activities, agricultural runoff, dust transport, and emissions from combustion sources (Pan et al., 2021). In recent decades, global climate change and intensified human activities have substantially increased P deposition, particularly in regions such as Asia and Europe (Penuelas et al., 2020). This trend is primarily attributed to agricultural fertilization practices, windborne dust from arid and semi-arid areas, and industrial emissions. While P deposition enhances nutrient availability in terrestrial ecosystems, it is also associated with a suite of environmental challenges. Alpine grassland ecosystems, particularly those on the Qinghai-Tibet Plateau, are highly sensitive to changes in nutrient availability due to their nutrient-limited conditions and harsh environmental constraints. These ecosystems provide vital ecosystem services, such as climate regulation, water conservation, soil fertility maintenance, and support for pastoral livelihoods (Yang et al., 2020). The addition of P promotes the increase in aboveground biomass of alpine grasslands, drives the proliferation of bacteria and fungi, enhances their activity levels, and further regulates the functional traits of plants, thereby propelling changes in the overall ecological balance and ecological functions of alpine grasslands (Sun J. et al., 2022). Plant biomass is a key metric for assessing grassland ecosystem health and restoration, as it reflects the system’s capacity for carbon storage and primary production. Biomass production in grasslands is intricately linked to the availability and stoichiometry of carbon (C), nitrogen (N), and P. Among these elements, P is often the most limiting nutrient in alpine ecosystems due to its low bioavailability and slow turnover rates (Mesquita et al., 2020).
P addition can profoundly affect the structure and functioning of alpine grassland ecosystems (Sun Y. et al., 2022). Experimental studies have shown that P inputs promote plant biomass accumulation, enhance vegetation cover, and increase primary productivity (Mao et al., 2021). However, these benefits are accompanied by significant alterations in plant community composition. P addition tends to favor species with higher P demands, such as grasses and forbs, while suppressing species less dependent on P, such as sedges. This shift in community structure can lead to a decline in plant diversity, particularly in nutrient-poor environments where competition for P is a key driver of species coexistence. Over the long term, P addition may result in soil P accumulation, altering the chemical and biological properties of soils and complicating the long-term trajectory of plant biomass dynamics. P enrichment can increase soil microbial activity and alter the composition of microbial communities, which play a critical role in nutrient cycling. Enhanced microbial activity may accelerate the mineralization of organic matter, thereby influencing the availability of other essential nutrients such as nitrogen. Moreover, the interaction between P and nitrogen availability can lead to imbalances in nutrient ratios, potentially reducing the efficiency of nutrient uptake and utilization by plants.
In addition to its effects on plant biomass, P addition significantly influences the stoichiometry of C, N, and P within alpine grassland (Sun Y. et al., 2022). Changes in stoichiometric ratios are particularly important as they regulate key ecological processes, including nutrient cycling, plant-microbe interactions, and ecosystem productivity. P addition exerts profound effects on the contents of C, N, and P in alpine grasslands. It can alter the structure of soil bacterial communities, influence nitrogen forms in the soil, and affect plant nitrogen uptake (Zhang Z. et al., 2024). Additionally, P addition regulates plant growth and microbial activity in soils, indirectly impacting soil carbon content. In alpine grasslands, P addition enhances plant P uptake, influencing the forms of P in the soil. More importantly, P addition significantly affects the stoichiometric ratios of plants, soil, and microbial biomass, notably reducing C:P and N:P ratios, while its effects on the C:N ratio are relatively limited (Sun Y. et al., 2022). Such imbalances in stoichiometric ratios could have potential negative consequences for the functionality of grassland ecosystems.
The alpine grasslands of Sejila Mountain, located in Nyingchi, southeastern Tibet, are representative of high-altitude ecosystems characterized by cool winters, mild summers, and distinct wet and dry seasons. These grasslands are nutrie nt-limited and highly sensitive to external nutrient inputs, making them an ideal system for studying the effects of P deposition. Most of the current studies on the impact of P addition on alpine grasslands are short-term experiments. In this study, we conducted a long-term P addition experiment to study its impacts on the plant traits, biomass, nutrient contents (C, N, and P), and stoichiometric characteristics of the alpine grasslands in Sejila Mountain. The main objectives are as follows: (1) Analyze the responses of different plant functional groups to P addition; (2) Reveal the impacts of P addition on the plant communities in alpine grasslands; (3) Explore the relationships among plant traits, biomass, and nutrient contents under P addition. This is of great significance for clarifying the potential mechanisms of the changes in plant communities and the dynamics of nutrient cycling in alpine grasslands under P deposition, and provides a scientific basis for the protection and restoration of alpine grasslands.
2 Materials and methods
2.1 Study area
The experiment was conducted on the alpine grasslands of the western slope of Sejila Mountain in Nyingchi City, Tibet Autonomous Region (29°37′30″N, 94°37′13″E). The site exhibits an elevation of 4,400 m and an average slope of 21.4° (Figure 1). This plot is located in the humid mountain warm temperate zone and semi-humid mountain temperate zone. The annual average temperature is −0.73°C, and the annual average precipitation is 1,134.1 mm. Precipitation is mainly concentrated from April to October. The frost - free period is 180 days, and the average relative humidity ranges from 60% to 80%. The primary vegetation types include Poaceae, Cyperaceae, and Forbs. The soil types in the area are dominated by mountain brown soils and acidic brown soils. Basic soil parameters are presented in Table 1.
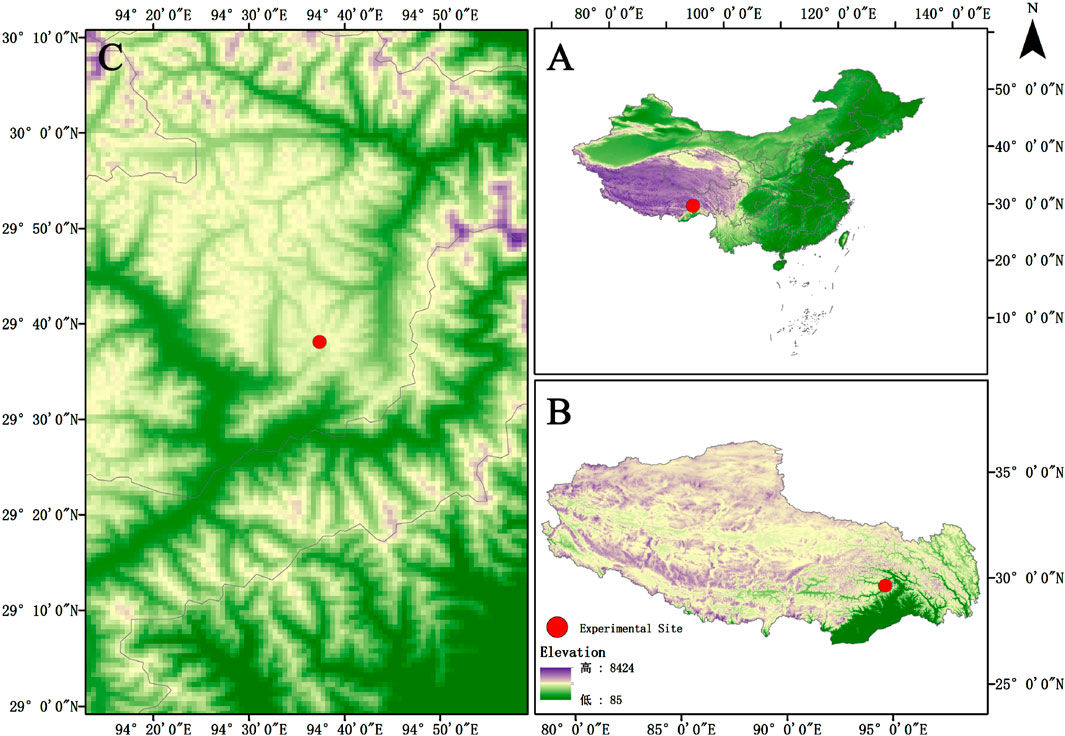
Figure 1. Geographical Location Map of the Research Area. (A) map of China; (B) map of Xizang; (C) Specific location of research site.
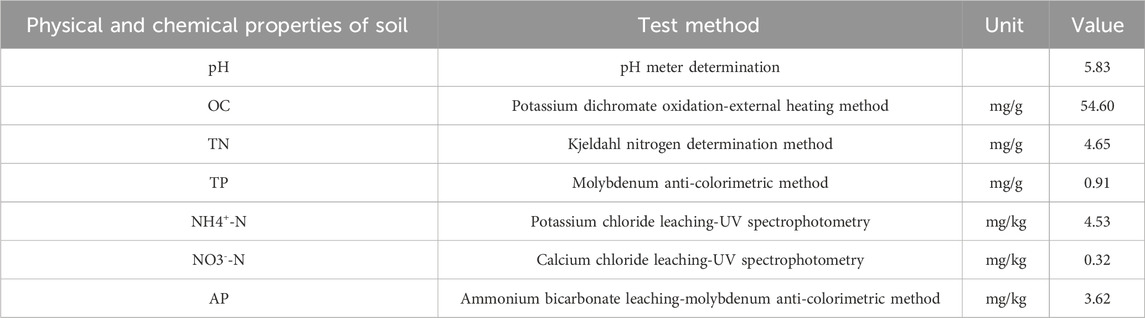
Table 1. Soil parameters and measurement methods of the test site.
2.2 Experimental design
The experiment employed a field P fertilizer application approach with controlled treatments. P addition is drawn from the P addition treatment test standard (50 kg hm−2 a−1) carried out by Zhai Jiaying in 2009 for alpine grasslands. Considering that the atmospheric P sedimentation has shown an increasing trend year by year over time, high concentration addition tests have been carried out based on the above P addition standards. Three replicated blocks of equal area were established, with a 5 m wide buffer zone between blocks. Each block contained four treatment groups, corresponding to P application rates of 0, 50, 75, and 100 kg hm−2 a−1, designated as CK, P50, P75, and P100, respectively. Each treatment group consisted of a plot measuring 3 m × 3 m, with a 1 m wide buffer zone between plots. A total of 12 plots were established. Translated from what was initiated in 2019, at the beginning of August each year, dissolve calcium superphosphate [Ca(H2PO2)] in water at the specified concentration, and evenly sprinkle it within the sample plot with the help of a watering can. The amount of water applied each time is roughly equivalent to a natural precipitation of 2.0 mm. In the control sample plot, the same amount of water is applied. After evaluation, the amount of water addition during fertilization will not have an impact on the ecosystem processes.
2.3 Measurement of plant traits and biomass
In the middle of August 2024, the number of species in each experimental area was recorded in detail. The heights and quantities of plants in each functional group were measured through the direct measurement method and the quadrat method, and the coverage rate was measured by using the line transect method. Randomly select three small plots of 30 cm × 30 cm. Plants within the subplots were clipped at ground level according to their functional groups, and subsequently oven-dried at 105°C for 30 min to deactivate enzymes, followed by drying at 80°C to constant weight. The aboveground biomass of each functional group was then determined by weighing. Plant roots were collected from the soil layer of 0–20 cm in depth using a soil auger with a diameter of 5 cm (Contains 95% of underground plant biomass). For each small sample plot, the auger was used to drill three times in an S-shape pattern, and the root samples obtained from the drilling were then mixed. Subsequently, the root samples were rinsed thoroughly with deionized water, and stones and humus mixed in them were carefully picked out. The treated root samples were placed in an oven at 80°C and dried to a constant weight before being weighed, so as to calculate the belowground biomass per unit area. The root-shoot ratio of the community was calculated by dividing the belowground biomass of the community by the aboveground biomass of the community.
2.4 Determination of the nutrient contents of C, N, and P in plants
The collected aboveground and belowground plant samples were ground into powder using a mechanical grinder. Organic carbon content was measured using the potassium dichromate oxidation method, total nitrogen was determined using the Kjeldahl method, and total P was quantified using the molybdenum-antimony anti-colorimetric method. By dividing the molar content of any two elements in the plant by each other, calculated the plants C:N, C:P, N:P.
2.5 Data analyses
Data processing was performed using Microsoft Excel 2019. Statistical analysis was carried out with IBM SPSS Statistics 20. One - way analysis of variance (ANOVA) was used to evaluate the responses of plant biomass, plant traits of different functional groups, and nutrient contents under different P addition gradients. Multiple comparisons were conducted using the Least Significant Difference (LSD) test, with the significance level set at P < 0.05. The figures were plotted using Origin 2021 software, and the results were presented as Mean ± Standard Error (Mean ± SE). The Spearman correlation analysis method was employed to investigate the relationships among plant biomass, plant traits, and nutrient contents in alpine grasslands under P addition.
3 Results
3.1 Effect of P addition on various plant functional groups
3.1.1 Effects of P addition on the Poaceae
The traits of the plant and aboveground biomass of the Poaceae showed a trend of increasing first and then decreasing under P treatment (Figure 2), and the highest value was reached under P75 treatment, with each plant trait increasing by 34.20%, 62.03% and 450.00% compared with CK, respectively, and the increase effect of average height and abundance of the Poaceae plants was more significant (P < 0.05); the aboveground biomass increased significantly by 140.35% compared with the control (P < 0.05); but under P100 treatment, each plant trait and aboveground biomass began to show a downward trend, indicating that excessive P addition has an adverse effect on the traits of the Poaceae and aboveground biomass. The addition of P addition not significantly affect the C content of Poaceae, and the N content showed a trend of decrease first and then increase (Figure 3). With the increase of P addition level, the P content of Poaceae plants increased significantly (P < 0.05); under P100 treatment, the P content of Poaceae plants increased by 122.10% compared with the control group. In the stoichiometric ratio characteristics, C:N showed a trend of increasing first and then falling; both C:P and N:P showed a significant trend of downward, which were 109.37 and 4.58 under P100 treatment.
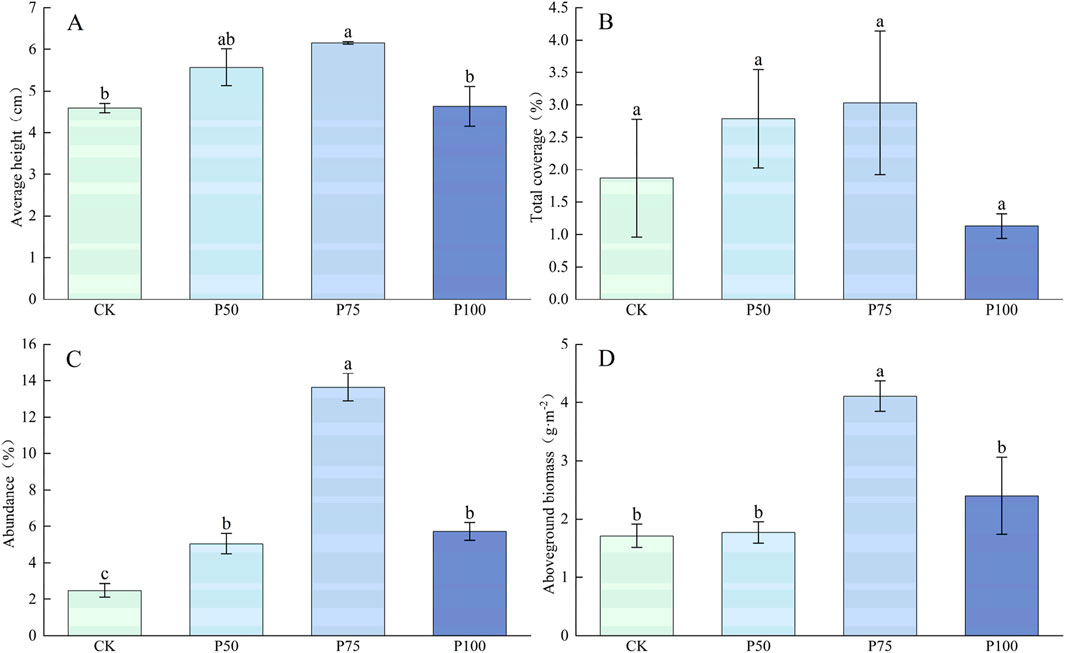
Figure 2. Changes in the traits and aboveground biomass of the Poaceae under P addition. (A) Average height, (B) Total coverage, (C) Abundance, (D) Aboveground biomass. Lower case letters in the graphs indicate significant differences mong the different level of P addition (P < 0.05, LSD test).
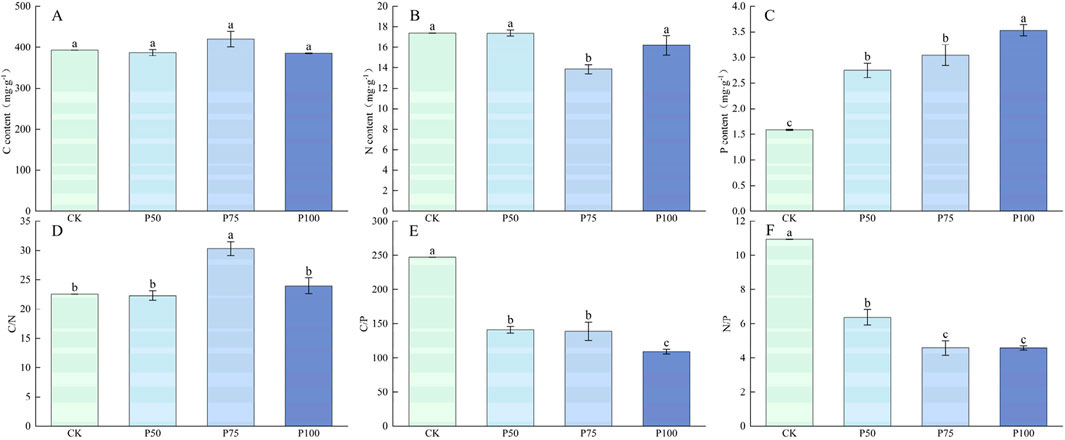
Figure 3. Changes in the nutrient content and stoichiometric ratio characteristics of the Poaceae under P addition. (A) C content, (B) N content, (C) P content, (D) C/N, (E) C/P, (F) N/P. Lower case letters in the graphs indicate significant differences mong the different level of P addition (P < 0.05, LSD test).
3.1.2 Effects of P addition on the Cyperaceae
In this study, the mean height and abundance of Cyperaceae plants showed an upward trend with the increase in P addition, but the difference was not significant, indicating that P addition had no significant effect on the mean height and abundance of Cyperaceae (Figure 4). P addition resulted in a significant downward trend in the total coverage and aboveground biomass of Cyperaceae (P < 0.05). The total coverage and aboveground biomass were the lowest under P100 treatment, with a decrease of 62.85% compared with CK treatment, and aboveground biomass decreased to 8.83 g m−2. P addition had no significant effect on C and N content in Cyperaceae (Figure 5). However, P addition significantly affected the P content of Cyperaceae (P < 0.05). With the increase in P addition level, the P content of Cyperaceae significantly increased, and under P100 treatment, the P content of Cyperaceae increased by 140.52% compared with the control group. P addition had no significant effect on C:N in Cyperaceae, but C:P and N:P showed a significant downward trend with the increase in P addition concentration (P < 0.05), which decreased to 110.01 and 6.12 under P100 treatment.
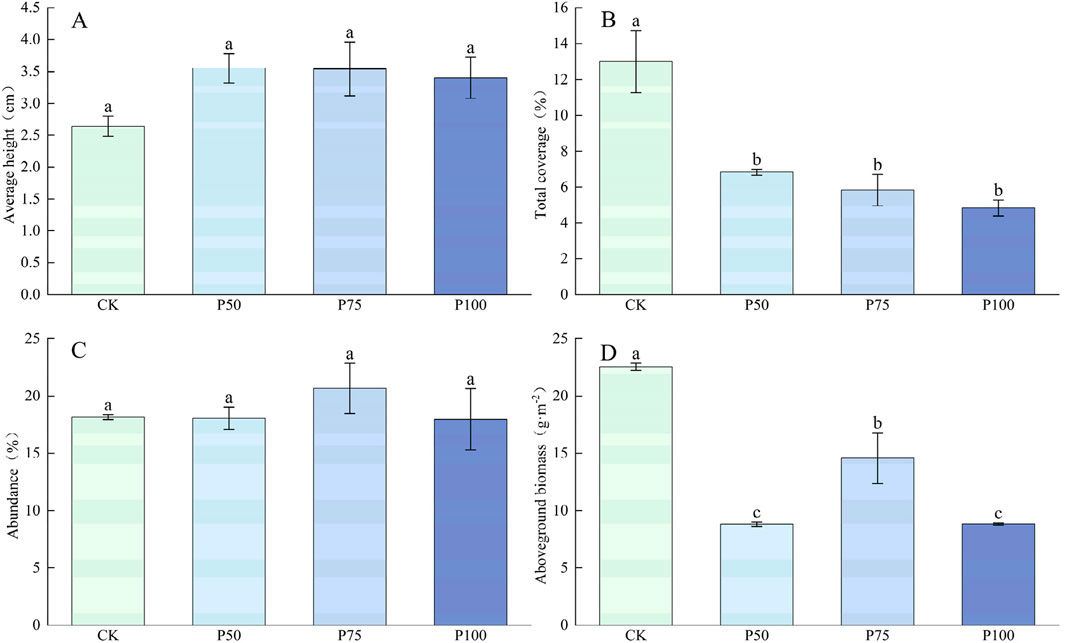
Figure 4. Changes in the traits and aboveground biomass of the Cyperaceae under P addition. (A) Average height, (B) Total coverage, (C) Abundance, (D) Aboveground biomass. Lower case letters in the graphs indicate significant differences mong the different level of P addition (P < 0.05, LSD test).
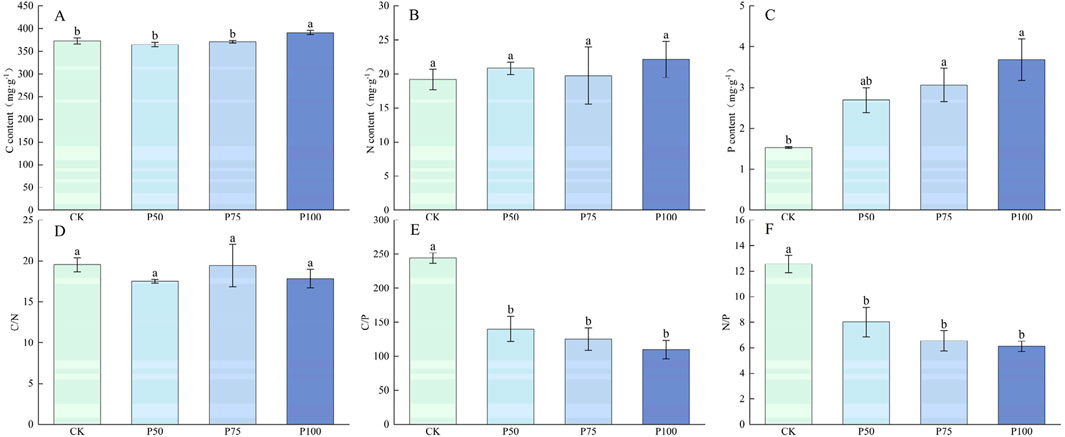
Figure 5. Changes in the nutrient content and stoichiometric ratio characteristics of the Cyperaceae under P addition. (A) C content, (B) N content, (C) P content, (D) C/N, (E) C/P, (F) N/P. Lower case letters in the graphs indicate significant differences mong the different level of P addition (P < 0.05, LSD test).
3.1.3 Effects of P addition on the Forbs
The plant traits and aboveground biomass of Forbs showed a trend of increasing first and then decreasing with P addition, and the changes were significant (P < 0.05, Figure 6), and reached the maximum value under P75 treatment. Compared with CK treatment, the average height, total coverage and abundance of Forbs increased by 73.86%, 51.55%, and 42.04%, and the aboveground biomass increased to 105.23 g m−2. In the case of P addition, the C content of Forbs showed a tendency to decrease first and then increase (Figure 7), with no significant effect on the N content, which significantly affected the P content of Forbs (P < 0.05). Under P100 treatment, the P content of Forbs increased by 117.50% compared with the control group. The C:N of Forbs did not change significantly with the increase of P addition level, but both the C:P and N:P ratios decreased significantly with the increase of P addition level (P < 0.05), with the lowest under P100 treatment, at 82.40 and 5.39.
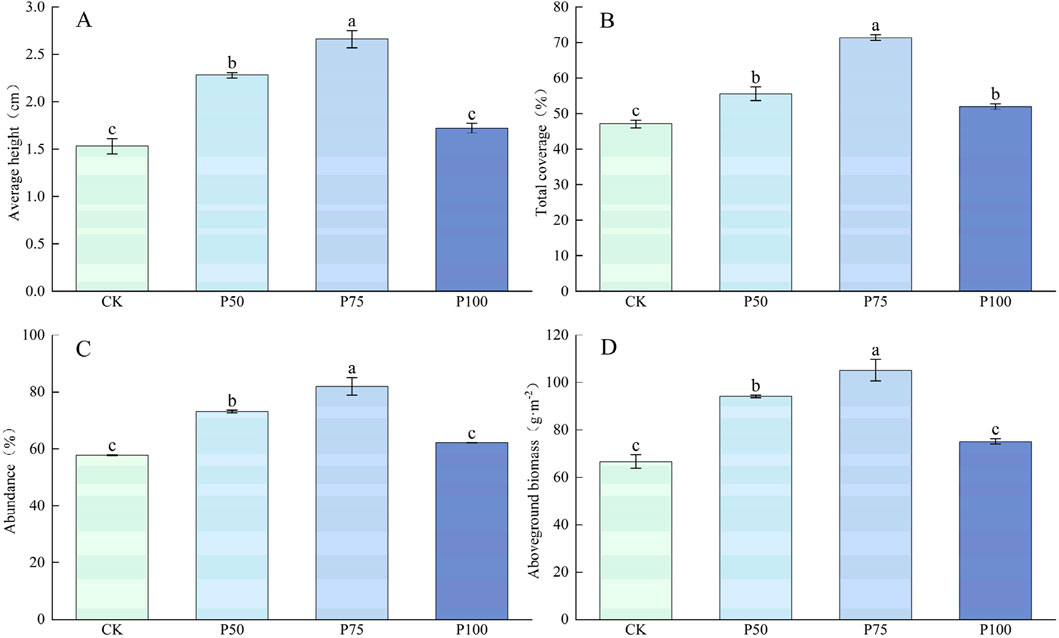
Figure 6. Changes in the traits and aboveground biomass of the Forbs under P addition. (A) Average height, (B) Total coverage, (C) Abundance, (D) Aboveground biomass. Lower case letters in the graphs indicate significant differences mong the different level of P addition (P < 0.05, LSD test).
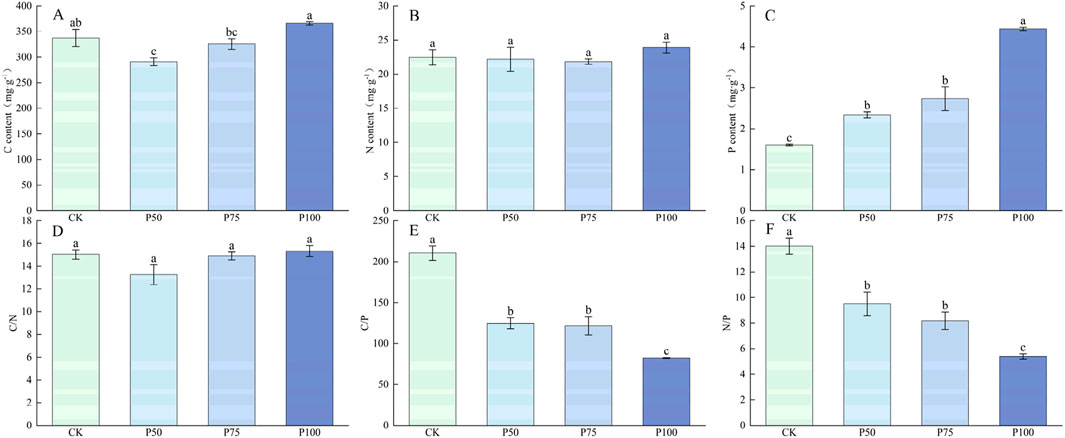
Figure 7. Changes in the nutrient content and stoichiometric ratio characteristics of the Forbs under P addition. (A) C content, (B) N content, (C) P content, (D) C/N, (E) C/P, (F) N/P. Lower case letters in the graphs indicate significant differences mong the different level of P addition (P < 0.05, LSD test).
3.2 Effects of P addition on plant communities
P addition significantly affected the plant community biomass and root crown ratio in the alpine grasslands of Sejila Mountain (P < 0.05, Figure 8). The aboveground biomass, belowground biomass, total biomass, and root crown ratios of plant communities showed a consistent response pattern characterized by increased first and then decreased. Under P50 treatment, the belowground biomass, total biomass and root crown ratios of the plant community reached peaks, which were 1,464.12 g m−2, 1,568.80 m−2 and 15.16, which increased by 132.34%, 117.60% and 118.13% respectively compared with the control group; under P75 treatment, the above-ground biomass of the plant community reached the highest value of 123.91 g m−2. This trend may be due to the fact that excessive P addition changes soil conditions, thus exerting an inhibitory effect on plant growth.
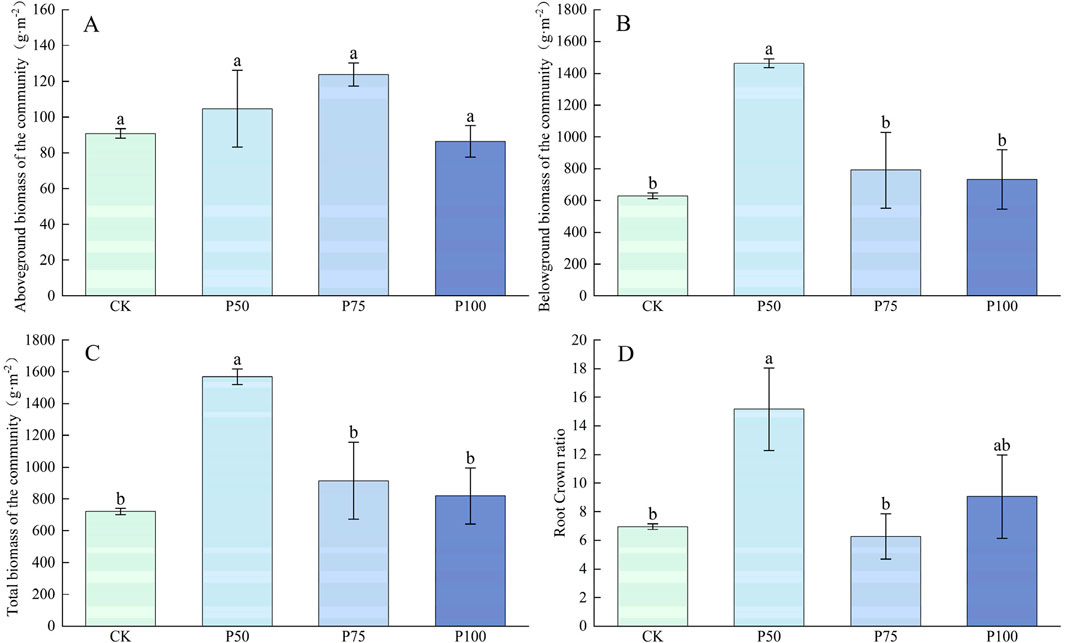
Figure 8. Changes in plant community biomass and root crown ratios by addition of P. (A) Aboveground biomass of the community, (B) Belowground biomass of the community, (C) Total biomass of the community, (D) Root crown ratio. Lower case letters in the graphs indicate significant differences mong the different level of P addition (P < 0.05, LSD test).
In the alpine grasslands plant community of Sejila Mountain, P addition had no significant effect on the C and N content of the aboveground and belowground plant parts (Figure 9). P addition significantly affected the P content of the aboveground and belowground parts of the plant community (P < 0.05). With the increase in the level of P addition, the P content in the plant community increased significantly, and under the P100 treatment, the P content of the aboveground and belowground parts of the plant community increased to 3.88 mg g−1 and 2.00 mg g−1. C:N in the aboveground and belowground parts of the plant community did not change significantly under the gradual increase in P addition levels. However, C:P and N:P showed a significant downward trend. Under P100 treatment, the C:P of the aboveground and belowground parts was 98.30 and 181.32, a decrease of 58.03% and 60.14% compared with the control treatment; N:P was 5.35 and 5.56, a decrease of 57.34% and 81.60% compared with the control treatment.
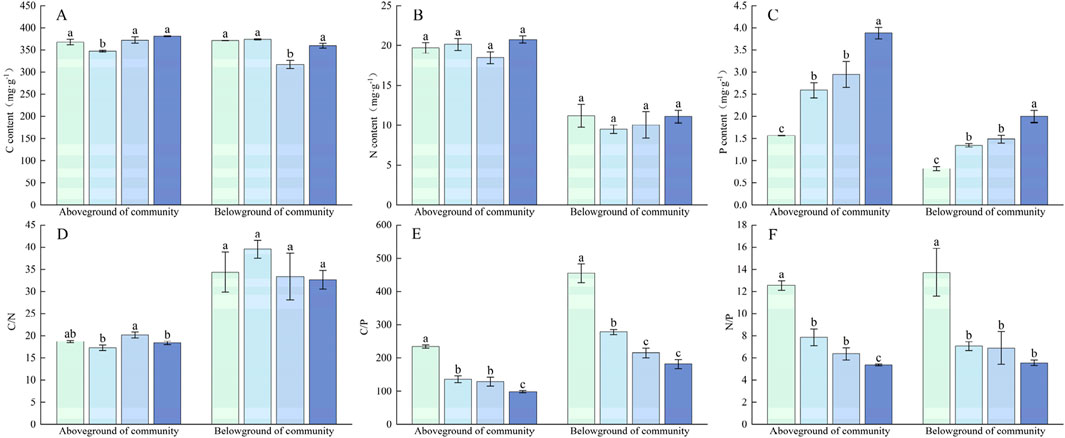
Figure 9. Changes in nutrient content and stoichiometric characteristics of P application on plant communities. (A) C content, (B) N content, (C) P content, (D) C/N, (E) C/P, (F) N/P. Lower case letters in the graphs indicate significant differences mong the different level of P addition (P < 0.05, LSD test).
3.3 Characteristics correlation analysis of P addition with plant traits, biomass, and nutrient contents
Spearman correlation analysis was conducted on plant traits, biomass and community nutrient contents in the alpine grasslands of Sejila Mountain under different gradients of P addition (Figure 10). The results showed that there were mostly positive correlations among the average height, total coverage and abundance within the Poaceae, Cyperaceae and Forbs respectively, indicating that these traits within the same group would affect each other and change jointly. The aboveground biomass of each plant group was significantly correlated with some of its own traits. The aboveground biomass of Poaceae had an extremely significant positive correlation with abundance (P < 0.001). The aboveground biomass of Forbs had extremely significant positive correlations with all its traits (P < 0.001). Notably, the aboveground biomass of Cyperaceae was negatively correlated with its average height, but positively correlated with total coverage and abundance. Meanwhile, there were also certain correlations among the aboveground biomasses of different plant groups, suggesting that they would influence the accumulation of biomass in the community. Community biomass was positively correlated with the aboveground biomasses of Poaceae and Forbs, but negatively correlated with that of Cyperaceae. The C, N and P contents of the community were not only correlated with each other, but also significantly related to the traits and biomass of plant groups. Specifically, the C and N contents had extremely significant negative correlations with the total coverage of Poaceae, there was an extremely significant positive correlation between C and N contents, and the P content had an extremely significant negative correlation with the total coverage of Cyperaceae. This indicates that nutrient contents can affect plant growth, and conversely, the growth status of plants can also have an impact on the nutrient cycling of the community.
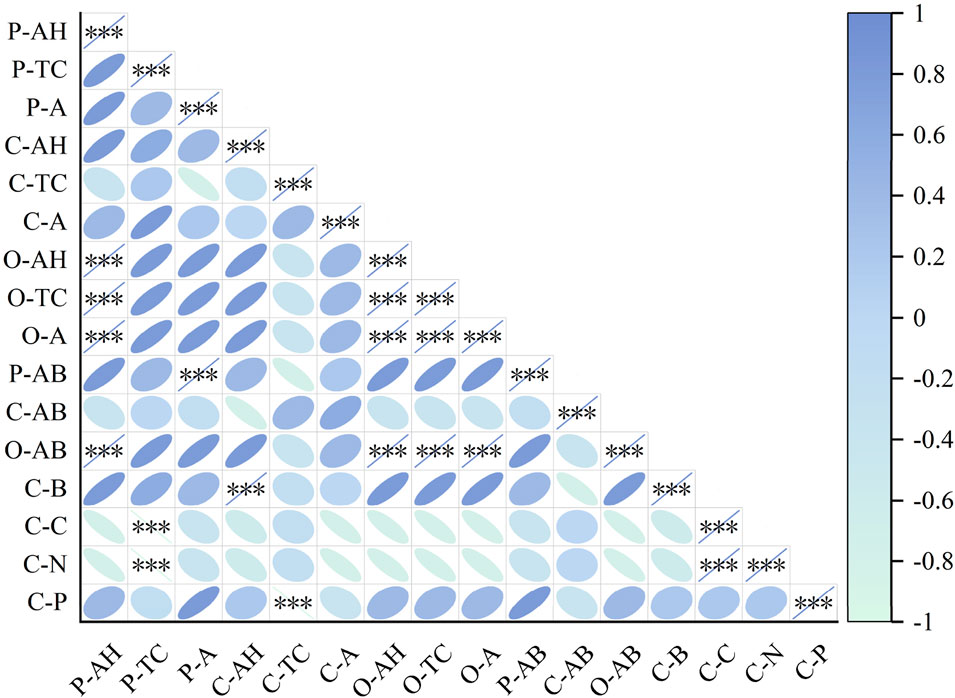
Figure 10. Spearman correlation heat map of plant traits, biomass and nutrient contents. P-AH, Average height of Poaceae; P-TC, Total coverage of Poaceae; P-A, Abundance of Poaceae; C-AH, Average height of Cyperaceae; C-TC, Total coverage of Cyperaceae; C-A, Abundance of Cyperaceae; O-AH, Average height of Forbs; O-TC, Total coverage of Forbs; O-A, Abundance of Forbs; P-AB, Aboveground biomass of Poaceae; C-AB, Aboveground biomass of Cyperaceae; O-AB, Aboveground biomass of Forbs; C-B, Community biomass; C-C, Community carbon content; C-N, Community nitrogen content; C-P, Community P content. *, ** and *** indicate significant correlations at the 0.05, 0.01 and 0.001 levels respectively.
4 Discussion
P is a component of important substances such as nucleic acids and phospholipids in plants, and is crucial for physiological processes such as energy metabolism and photosynthesis (Dissanayaka et al., 2020). Research has found that within the moderate range of P addition, grassland plants gain more abundant nutrients, enhance photosynthesis, and accelerate growth (Fan et al., 2021). In this study, it was found that moderate P supply can help improve material synthesis and accumulation in Poaceae and Forbs, thereby increasing height, coverage, and abundance. But when P addition exceeds a certain threshold, it can cause a series of problems. Excessive application of phosphate fertilizers leads to a surplus of P in the soil, increasing the risk of P loss (Zhang W. et al., 2020). These changes can interfere with the absorption of other nutrients by plants, affect their normal physiological functions, and thus inhibit plant growth. The P demand characteristics of Cyperaceae may be different from those of Poaceae and Forbs, and their growth may not be significantly regulated by the amount of P addition, so the average height and abundance changes are not significant. The decrease in total coverage may be due to P addition promoting the growth of other plants, intensifying interspecific competition, and putting Cyperaceae at a disadvantage in resource acquisition, resulting in a decrease in their coverage in the community (Yang et al., 2020).
Studies have shown demonstrated that P addition significantly increased the aboveground biomass of Poaceae in alpine grassland (Zhang W. et al., 2024). In this study, at a P application rate of 75 kg hm−2 a−1 (P75 treatment), the aboveground biomass of Poaceae and Forbs reached 4.11 g m−2 and 105.23 g m−2, respectively, representing increases of 140.35% and 58.20% compared to the control. However, this trend reversed at higher P levels. These results indicate that P addition initially promotes the growth of Poaceae and Forbs, but this stimulatory effect diminishes and may even turn inhibitory as P concentration continues to rise. Pedro et al. (2021) also supported the notion that P addition increases the aboveground biomass of non-nitrogen-fixing plants. However, when soil P concentration exceeds plant demand, a phenomenon of P saturation occurs. Excess P not only fails to further promote plant growth but may also intensify ecological competition among plant species (Kuang et al., 2021), leading to a decline in the aboveground biomass of Poaceae and Forbs. Similarly, observed in a multi-gradient nitrogen and P addition experiment in alpine grassland that the aboveground biomass of Cyperaceae significantly decreased with increasing P levels (Dong et al., 2022). This finding aligns with our study and may be attributed to Cyperaceae being more limited by nitrogen availability. In contrast, P addition promotes the growth of competing plant species, which intensify competition for resources such as light, water, and nutrients (Kuang et al., 2021), thereby suppressing the growth of Cyperaceae.
Plant biomass, as a core indicator of grassland ecosystem productivity, plays a crucial role in ecosystem stability and functionality (Gaowen et al., 2021). In a 5-year P addition experiment conducted in the alpine grasslands of Sejila Mountain, we observed a trend of initial increase followed by a decline in aboveground biomass, belowground biomass, total biomass, and root-to-shoot ratio of the plant community. Notably, the increase in belowground biomass was more pronounced than that in aboveground biomass under different P addition treatments, consistent with findings by Chen and Xiao (2023), which highlighted the stronger stimulatory effect of P addition on belowground biomass. P, as an essential nutrient for plant growth, can enhance plant growth, increase biomass, and optimize root-to-shoot ratio when applied in appropriate amounts (Lekberg et al., 2021). However, excessive P addition can induce adverse changes in the soil environment, such as salinity accumulation and pH imbalance (Yong et al., 2019), which disrupt soil microbial community structure and function, inhibiting the activity of certain microorganisms (Zhan et al., 2020). These environmental stresses eventually feedback onto plants, causing a decline in biomass and root-to-shoot ratio.
The contents of C, N, and P in plants are directly linked to primary productivity, species composition, and diversity in grassland ecosystems, playing a critical role in ecosystem function and regulation (De et al., 2019). Our results indicated that P addition had no significant effect on the C and N contents of the aboveground and belowground components of the plant community. Carbon, as a structural component in plants, is relatively stable and less influenced by external environmental factors (Sterner and Elser, 2017). Although P addition may alter soil microbial community structure, these changes did not significantly affect plant nitrogen content (Wang et al., 2020). However, the P content of the plant community significantly increased under P addition. The P contents of Poaceae, Cyperaceae, and Forbs responded similarly to P addition. Under the P100 treatment, the P contents of these functional groups increased by 122.1%, 140.52%, and 117.50%, respectively, compared to the control. This is likely because P addition elevated the concentration of available P (AP) in the soil (Cheng et al., 2020), providing a richer P source for plants and enhancing P uptake.
The C:N ratio is often regarded as an important indicator of nutrient balance in plants (Zhang J. et al., 2020). In this study, P addition had no significant effect on the C:N ratio in the aboveground and belowground components of the plant community. This suggests that P addition is not strongly correlated with nitrogen uptake or that plants can adjust nitrogen uptake and utilization to maintain a stable C:N balance (Sun Y. et al., 2022). In contrast, the C:P and N:P ratios of the plant community declined significantly with increasing P levels. This trend is attributed to enhanced P uptake efficiency, which increased P content in plants, while the absorption of carbon and nitrogen did not increase proportionally, thereby reducing C:P and N:P ratios (Mao et al., 2021; Sun Y. et al., 2022). In this study, Poaceae demonstrated stronger P uptake and utilization efficiency under P addition (Liu et al., 2020), resulting in faster growth and greater accumulation of nutrients. Conversely, Cyperaceae and Forbs showed weaker responses to P addition. The differential responses of plant species to P addition may be due to variations in their P uptake and utilization efficiency (Luor et al., 2022), as well as the influence of microbial communities altering the mineralization rate of organic P to affect P availability (Deforest and Moorhead, 2020). Despite these differences, the overall trend observed was a significant decline in C:P and N:P ratios with increasing P addition levels.
In the study of the alpine grasslands on Sejila Mountain, the correlations among plant traits, biomass, and community nutrient contents under different gradients of P addition exhibit unique patterns. Some scholars have found that within a certain range, P addition can promote the growth of Poaceae plants (Huang et al., 2018), and there is a positive correlation between their biomass and abundance, which is consistent with the extremely significant positive correlation between the aboveground biomass and abundance of Poaceae plants in this study. However, for Forbs, in this study, their aboveground biomass has an extremely significant positive correlation with all traits. This may be due to the unique environmental conditions of the alpine grasslands on Sejila Mountain, which make Forbs more sensitive to P addition and enable them to make more full use of P to promote their own growth. The results that the aboveground biomass of Cyperaceae is negatively correlated with the average height and positively correlated with the total coverage and abundance are relatively rare. It may be because P addition has changed the intra-specific or inter-specific competition relationships of Cyperaceae plants. Under high coverage and abundance, the competition among individuals intensifies, resulting in a decrease in the average height, but the overall biomass accumulation is more influenced by the total coverage and abundance. At the community level, in this study, the community biomass is positively correlated with Poaceae and Forbs, and negatively correlated with Cyperaceae. Studies on grasslands at different altitudes have shown that community biomass is affected by dominant plant groups (Chen et al., 2021). In the alpine grasslands on Sejila Mountain, Poaceae and Forbs play a positive and dominant role in the construction of community biomass, while Cyperaceae shows different correlations due to its own characteristics. In addition, the relationship between community nutrient contents and the traits and biomass of plant groups is consistent with previous studies on the interaction between nutrient cycling and plant growth, further emphasizing the importance of the complex feedback mechanism between the two in the alpine grassland ecosystem.
Given the complexity of P deposition, future research should focus on long-term monitoring and modeling to capture the spatiotemporal variability of P inputs. Additionally, integrated management strategies are needed to mitigate the environmental risks associated with excessive P deposition. Such strategies could include optimizing agricultural fertilization practices, controlling industrial emissions, and implementing land management practices that enhance the retention and utilization of P within ecosystems. By addressing these challenges, we can promote the sustainability and resilience of alpine grassland ecosystems in the face of global environmental change.
5 Conclusion
We conducted an in-depth analysis of a 5-year P addition experiment with varying gradients in the alpine grasslands of Sejila Mountain and drew the following key conclusions: P addition had significant and profound effects on plant traits, biomass, C, N, and P contents, as well as their stoichiometric ratios. These effects exhibited a high degree of consistency across plant communities and their constituent functional groups. Poaceae and Forbs’ overground biomass increase most under P75 treatment, but when P addition exceeds 100 kg hm−2 a−1, it will inhibit its growth, and P addition reduces the total coverage and biomass of the Cyperaceae. At the community level, however, P addition led to a notable increase in total biomass. Additionally, P addition substantially altered the elemental composition within plants, as evidenced by a significant increase in P content and a notable decrease in the C:P and N:P ratios. However, excessive P addition (P100) intensified ecological competition among plant species, posing potential threats to the functional stability and ecological balance of grassland ecosystems. Based on the above research results, in order to empirically test the impact of P addition on the ecosystem stability of the alpine grasslands in Sejila Mountain, it is very necessary to closely monitor the changes in plant community structure, and pay attention to the impact of precipitation on the effect of nutrient addition, and pay special attention to the dynamics of Cyperaceae plants. It is crucial to prevent ecological imbalances caused by fierce interspecies competition and achieve sustainable development of grassland ecosystems.
Data availability statement
The raw data supporting the conclusion of this article will be made available by the authors, without undue reservation.
Author contributions
XZ: Conceptualization, Methodology, Software, Formal analysis, Investigation, Data curation, Writing – original draft, Writing – review and editing, Visualization. YY: Conceptualization, Software, Investigation, Resources, Writing – review and editing, Visualization, Supervision, Project administration, Funding acquisition. JT: Conceptualization, Methodology, Validation, Formal analysis, Investigation, Data curation, Writing – original draft, Writing – review and editing, Visualization. ZN: Writing – original draft. ZC: Writing – original draft. JL: Writing – original draft. YH: Conceptualization, Methodology, Formal analysis, Investigation, Data curation, Writing – original draft, Writing – review and editing, Visualization.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by the National Natural Science Foundation of China (31860141; 31360119); Graduate Education Innovation Program of Tibet Agriculture and Animal Husbandry University (YJS2024-26; YJS2024-28; YJS2024-31); National College Student Innovation and Entrepreneurship Training Program (2024-02); Linzhi Apple Science and Technology Academy of Tibet (XY2024-03); Key Discipline Construction Project of Tibet Agriculture and Animal Husbandry University (XK2024-04); Xizang Agriculture and Animal Husbandry University Doctoral Program in Forestry (Phase I) funded by Grant 533325001.
Acknowledgments
The author thanks Associate Professors YH for providing technical assistance.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Chen, C., and Xiao, W. (2023). The global positive effect of phosphorus addition on soil microbial biomass. Soil Biol. and Biochem. 176, 108882. doi:10.1016/j.soilbio.2022.108882
Chen, Z., Xiong, P., Zhou, J., Lai, S., Jian, C., Xu, W., et al. (2021). Effects of plant diversity on semiarid grassland stability depends on functional group composition and dynamics under N and P addition. Sci. Total Environ. 799, 149482. doi:10.1016/j.scitotenv.2021.149482
Cheng, H. Y., Yuan, M. S., Duan, Q. Y., Sun, R., Shen, Y., Yu, Q., et al. (2020). Influence of phosphorus fertilization patterns on the bacterial community in upland farmland. Industrial Crops and Prod. 155, 112761. doi:10.1016/j.indcrop.2020.112761
De, L. J. R., Jackson, B. G., Wilkinson, A., Pritchard, W. J., Oakley, S., Mason, K. E., et al. (2019). Relationships between plant traits, soil properties and carbon fluxes differ between monocultures and mixed communities in temperate grassland. J. Ecol. 107, 1704–1719. doi:10.1111/1365-2745.13160
Deforest, L. J., and Moorhead, L. D. (2020). Effects of elevated pH and phosphorus fertilizer on soil C, N and P enzyme stoichiometry in an acidic mixed mesophytic deciduous forest. Soil Biol. Biochem. 150, 107996. doi:10.1016/j.soilbio.2020.107996
Dissanayaka, S., Ghahremani, M., Siebers, M., Wasaki, J., and Plaxton, W. C. (2020). Recent insights into the metabolic adaptations of phosphorus deprived plants. J. Exp. Bot. 72 (2), 199–223. doi:10.1093/jxb/eraa482
Dong, J., Cui, X., Niu, H., Zhang, J., Zhu, C., Li, L., et al. (2022). Effects of nitrogen addition on plant properties and microbiomes under high phosphorus addition level in the alpine steppe. Front. Plant Sci. 13, 894365. doi:10.3389/fpls.2022.894365
Fan, L. L., Mekrovar, O., Li, Y. M., Li, K. H., Ma, X. X., and Mao, J. F. (2021). Effect of nutrient addition on the productivity and species richness of grassland along with an elevational gradient in Tajikistan. Front. Plant Sci. 12, 765077. doi:10.3389/fpls.2021.765077
Gaowen, Y., Masahiro, R., Julien, R., Hempel, S., and Rillig, M. C. (2021). Plant and soil biodiversity have non-substitutable stabilising effects on biomass production. Ecol. Lett. 24 (8), 1582–1593. doi:10.1111/ele.13769
Huang, J., Yu, H., Liu, J., Luo, C., Sun, Z., Ma, K., et al. (2018). Phosphorus addition changes belowground biomass and C:N:P stoichiometry of two desert steppe plants under simulated N deposition. Sci. Rep. 8 (1), 3400. doi:10.1038/s41598-018-21565-w
Kuang, X. L., Si, K. Y., Song, H. J., Peng, L., and Chen, A. (2021). Lime-phosphorus fertilizer efficiently reduces the Cd content of rice: physicochemical property and biological community structure in Cd-polluted paddy soil. Front. Microbiol. 12, 749946. doi:10.3389/fmicb.2021.749946
Lekberg, Y., Arnillas, C. A., Borer, E. T., Bullington, L. S., Fierer, N., Kennedy, P. G., et al. (2021). Nitrogen and phosphorus fertilization consistently favor pathogenic over mutualistic fungi in grassland soils. Nat. Commun. 12, 3484. doi:10.1038/s41467-021-23605-y
Liu, C. J., Gong, X. W., Dang, K., Li, J., Yang, P., Gao, X., et al. (2020). Linkages between nutrient ratio and the microbial community in rhizosphere soil following fertilizer management. Environ. Res. 184, 109261. doi:10.1016/j.envres.2020.109261
Luor, Y., Kuzyakov, Y., Zhu, B., Qiang, W., Zhang, Y., and Pang, X. (2022). Phosphorus addition decreases plant lignin but increases microbial necromass contribution to soil organic carbon in a subalpine forest. Glob. Change Biol. 28 (13), 4194–4210. doi:10.1111/gcb.16205
Mao, Q. G., Chen, H., Wang, C., Pang, Z., Mo, J., and Lu, X. (2021). Effect of long-term nitrogen and phosphorus additions on understory plant nutrients in a primary tropical forest. Forests 12 (6), 803. doi:10.3390/f12060803
Mesquita, C. P. B. D., Brigham, L. M., Sommers, P., Porazinska, D. L., Farrer, E. C., Darcy, J. L., et al. (2020). Evidence for phosphorus limitation in high-elevation unvegetated soils, Niwot Ridge, Colorado. Biogeochemistry 147 (7), 1–13. doi:10.1007/s10533-019-00624-y
Pan, Y., Liu, B., Cao, J., Liu, J., Tian, S., and Du, E. (2021). Enhanced atmospheric phosphorus deposition in Asia and Europe in the past two decades. Atmos. Ocean. Sci. Lett. 14 (05), 100051. doi:10.1016/j.aosl.2021.100051
Pedro, M. T., Suzanne, M. P., Selene, B., Chaneton, E. J., Firn, J., Risch, A. C., et al. (2021). Negative effects of nitrogen override positive effects of phosphorus on grassland legumes worldwide. Proc. Natl. Acad. Sci. U. S. A. 118 (28), 1–8. doi:10.1073/pnas.2023718118
Penuelas, J., Janssens, I. A., Ciais, P., Obersteiner, M., and Sardans, J. (2020). Anthropogenic global shifts in biospheric N and P concentrations and ratios and their impacts on biodiversity, ecosystem productivity, food security, and human health. Glob. Change Biol. 26 (4), 1962–1985. doi:10.1111/gcb.14981
Sterner, R. W., and Elser, J. J. (2017). Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere. Princeton University Press. Publication date: 2017-02-15. doi:10.1515/9781400885695
Sun, J., Wang, Y., Liu, S., Li, J., Zhou, H., Wu, G., et al. (2022). Editorial: patterns, functions, and processes of alpine grassland ecosystems under global change. Front. Plant Sci. 13, 1048031. doi:10.3389/fpls.2022.1048031
Sun, Y., Wang, C. T., Chen, X. L., Liu, S., Lu, X., Chen, H. Y. H., et al. (2022). Phosphorus additions imbalance terrestrial ecosystem C:N:P stoichiometry. Glob. Change Biol. 28 (24), 7353–7365. doi:10.1111/gcb.16417
Wang, S. H., Mori, T., Mo, J. M., and Zhang, W. (2020). The responses of carbon- and nitrogen-acquiring enzymes to nitrogen and phosphorus additions in two plantations in southern China. J. For. Res. 31 (04), 1319–1324. doi:10.1007/s11676-019-00905-0
Yang, J., Liu, Q. R., and Wang, X. T. (2020). Plant community and soil nutrient of alpine meadow in different degradation stages on the Tibetan plateau, China. J. Appl. Ecol. 31 (12), 4067–4072. doi:10.13287/j.1001-9332.202012.008
Yong, J., Zhu, W. X., Ru, Z. W., Li, H., and Zhang, Y. G. (2019). Effects of long-term fertilization and water addition on soil properties and plant community characteristics in a semiarid grassland. J. Appl. Ecol. 30 (7), 2470–2480. doi:10.13287/j.1001-9332.201907.034
Zhan, Y. N., Wang, Z., and Meng, Y. L. (2020). Biochar addition improves soil phosphorus availability: a meta-analysis. J. Appl. Ecol. 31 (4), 1185–1193. doi:10.13287/j.1001-9332.202004.024
Zhang, J., He, N., Liu, C., Xu, L., Chen, Z., Li, Y., et al. (2020). Variation and evolution of C:N ratio among different organs enable plants to adapt to N-limited environments. Glob. Change Biol. 26, 2534–2543. doi:10.1111/gcb.14973
Zhang, W., Gong, J., Zhang, S., Lambers, H., Dong, X., Hu, Y., et al. (2024). Soil phosphorus availability affects niche characteristics of dominant C3 perennial and sub-dominant C4 annual species in a typical temperate grassland of northern China. Plant Soil 504, 737–761. doi:10.1007/s11104-024-06655-1
Zhang, W., Zhang, Y., An, Y., and Chen, X. (2020). Phosphorus fractionation related to environmental risks resulting from intensive vegetable cropping and fertilization in a subtropical region. Environ. Pollut. 269, 116098. doi:10.1016/j.envpol.2020.116098
Keywords: phosphorus addition, alpine grassland, plant traits, nutrient content, stoichiometric ratio
Citation: Zhang X, Ye Y, Tao J, Niu Z, Cui Z, Li J and Han Y (2025) Responses of alpine grassland plant communities on Sejila Mountain in the Qinghai-Tibet Plateau to phosphorus addition. Front. Environ. Sci. 13:1601695. doi: 10.3389/fenvs.2025.1601695
Received: 28 March 2025; Accepted: 06 May 2025;
Published: 22 May 2025.
Edited by:
Tianjiao Feng, Beijing Forestry University, ChinaReviewed by:
Ling Han, Grassland Ecology, Nitrogen Cycle, Nitrogen Deposition, ChinaWang Hongyi, Heilongjiang Bayi Agricultural University, China
Ran Zhang, Beijing Forestry University, China
Copyright © 2025 Zhang, Ye, Tao, Niu, Cui, Li and Han. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yanying Han, MTA2NDMzNjAxNUBxcS5jb20=
†These authors share first authorship