 Beatriz Fernandes1,2*
Beatriz Fernandes1,2* Rute Duarte1,2
Rute Duarte1,2 Cristiana Paiva1,3
Cristiana Paiva1,3 João Oliveira-Pacheco1
João Oliveira-Pacheco1 Anabela Cachada1,3
Anabela Cachada1,3 Ruth Pereira1,2
Ruth Pereira1,2- 1GreenUPorto/INOV4Agro, Vairão, Portugal
- 2Departamento de Biologia da Faculdade de Ciências da Universidade do Porto, Porto, Portugal
- 3Departamento de Geociências, Ambiente e Ordenamento do Território da Faculdade de Ciências da Universidade do Porto, Porto, Portugal
The use of plant protection products is often related to environmental contamination, including copper-based products as the Bordeaux Mixture (BM) and synthetic fungicides like folpet (active substance), both of which face use restriction from the European Union due to their toxicity. As alternatives, plant-based products such as E. globulus extracts (EE) or essential oils (EO) are considered more sustainable and environmentally friendly products with known biocidal properties, but their impacts on non-target species remain poorly known. These products might be effective not only on existing pathogenic species that infect grapevines, but also on other potential pathogens, emerging in a context of climate change. This study aimed to evaluate the fungicidal effects of E. globulus EE and EO, alone or in combination with BM and folpet through growth inhibition assays using target (Botrytis cinerea), potentially target (Chaetomium elatum) and non-target (Lentinus sajor caju and Phanerochaete chrysosporium) fungal species, under in vitro conditions. Different exposure methods were also assessed. Results showed that EE alone inhibited the growth of the fungi species, though sensitivity varied with the application methods. The combination of EE with BM was only effective when incorporated into the growth medium, becoming a promising alternative to reduce BM use, however, the same cannot be said for the mixture of EE and folpet. E. globulus EO effectively inhibited the target fungi growth, still, the non-target species were also affected, highlighting that a careful evaluation of plant-based products is of utmost importance, considering the effects that they have on non-target organisms of the ecosystem.
1 Introduction
Fungicides are commonly applied in viticulture to treat common diseases such as downy mildew and grey mold, caused by Plasmopara viticola (Koledenkova et al., 2022) and B. cinerea, respectively (Jiang et al., 2023). Copper-based fungicides are the most applied plant protection products (PPP) to treat downy mildew, however, its constant use over the years, especially in organic viticulture where no alternatives exist, has led to an accumulation of Cu in soils, adversely affecting the non-target organisms of the ecosystem (Tamm et al., 2022). This lead to Cu-based fungicides to be included in the list of candidates for substitution in the European Union and the European Commission emitted a regulation to limit Cu use to a maximum of 28 kg per ha over a period of 7 years (European Commission, 2018). Regarding synthetic fungicides, that have been used to control, not only downy mildew, but also grey mold caused by B. cinerea, in vineyards under integrated production mode (IPM), the active ingredient (a.i.) folpet, a member of the class of phthalimides (National Center for Biotechnology Information, 2024), is one of the most used a.i., approved in almost all European Member states (IUPAC, 2024). Even though this fungicide has low persistence in soil, it is known to be toxic towards earthworms and adverse effects were also reported in some non-target aquatic organisms (Acar et al., 2023; Adams et al., 2021; EFSA, 2023). Taking this into consideration, under the pressure of climate changes that is contributing for high pressure of infections with known pathogens but also for the appearance of new potential pathogenic fungi and diseases (Kumar and Mukhopadhyay, 2024), finding appropriate solutions to replace both copper and conventional synthetic PPP is a pressing matter.
A promising alternative that is becoming the focus of research for replacing the use of synthetic PPP are the plant-based products (Alengebawy et al., 2021; Wightwick et al., 2013). These products are often described as being eco-friendly, environmentally safe, not as prone to develop resistance, generally safe for humans and other non-target organisms, and target specific, allowing a more precise control of pests and diseases (Cordeau et al., 2016; Gupta et al., 2023; Zhao et al., 2017). However, there are few studies addressing the toxicity of these plant-based products to non-target organisms (Machado et al., 2023). Notwithstanding, the ability of these products to be used as alternative PPP is possible due to the composition of plants in polysaccharide, protein, alkaloids, anthocyanins, carotenoids, flavonoids, phenolic acids and other phytochemicals with bioactivity (Zhao et al., 2017). In the specific case of compounds such as monoterpenes, sesquiterpenes, phenols, alkaloids, flavonoids, and tannins, they are part of the essential oils found in the leaves of plants. These are responsible for the allelopathy, a chemical interaction between living organisms where plants can interfere with the establishment and growth of other species by releasing inhibitory molecules into the surrounding environment (Kong et al., 2019; Macías et al., 2019).
One of the most known species with this capacity is Eucalyptus globulus, a species belonging to the Eucalyptus genus, which is an economically important hardwood tree originated from southeast Australia, disseminated at temperate regions around the world (Águas et al., 2014; Queirós et al., 2020).
The antifungal potential of Eucalyptus oils (Ayed et al., 2023; Pedrotti et al., 2019), and, in the specific case of E. globulus, their essential oils were effective against Alternaria solani Sorauer that causes early blight in tomato (Tomazoni et al., 2017), B. cinerea and Colletotrichum acutatum the most important fungal root diseases in grapes (Pedrotti et al., 2019) and P. viticola (Fialho et al., 2017). While most studies focus on the use of essential oils, and even though they are effective antifungals, they remain expensive products that needs specialized equipment and expertise for their extraction and purification. Thus, aqueous extracts such as the one proposed by Pinto et al. (2021) may become important from the farmers perspective as it could be effective but less expensive and easily obtained and prepared. Some studies were already conducted on the efficacy of E. globulus and other Eucalyptus species aqueous extracts on fungal growth of C. gloeosporioides, A. alternata, A. mali, Diplodia. seriata and Botryosphaeriadothidea responsible for diseases in apple orchards (Ahmad et al., 2020; Hajji-Hedfi et al., 2024), Fusarium oxysporum the causal agent of Bayoud disease on date palm (Bouhlali et al., 2020) and Penicillium expansum, P. italicum, Aspergillus niger, P. digitatum, B. cinerea, and Rhizopus stolonifer (Yao et al., 2021) species that cause severe post-harvest diseases.
In addition to antifungal activity of plant-based products for target species with agronomic importance, it is important to understand how its application affects the non-target organisms of the ecosystem. This is the case of Lentinus sajor caju and Phanerochaete chrysosporium, two white rot fungi naturally present in the ecosystem that have key roles in the cycling of carbon and nitrogen, organic matter decomposition and biotic interactions (Bing-Ru et al., 2006; Galindo et al., 2013). Nonetheless, little is known of how these species behave in contact with E. globulus aqueous extracts/essential oils.
Therefore, this study aimed to evaluate the efficacy of both E. globulus aqueous extract and essential oil on the mycelial growth of two non-target (L. sajor caju and P. chrysosporium), one target (B. cinerea) and one potential target fungal species isolated from a vineyard, to substitute or reduce the use of traditional fungicides. Furthermore, as part of a strategy to at least reduce the amount of Cu and folpet applied in vineyards, E. globulus aqueous extract was also tested in combination with those PPP.
2 Materials and methods
2.1 Test species
Two Basidiomycota (L. sajor caju (Fries) Fries and P. chrysosporium (Burdsall 38,388)) and two Ascomycota (B. cinerea and C. elatum) species of the families Leotiomycetes and Sordariomycetes, respectively, were used in this study. The two Basidiomycota fungi and B. cinerea were gently obtained from two collections from the University of Aveiro, but presently maintained at GreenUPorto of the Faculty of Sciences of the University of Porto. For each species, individual cultures are kept, in the dark, at 4 °C in Malt Extract Agar (MEA) (Thermo Scientific™ Oxoid™ Malt Extract Agar, dehydrated). Before the beginning of the assays, new individual cultures were obtained where P. chrysosporium was cultured in MEA for 3 days at 25 °C and L. sajor caju and B. cinerea for 5 days at 28 °C. In the specific case of the fungi isolated from infected grapevines leaves, at Campus de Vairão vineyard of the Faculty of Sciences of the University of Porto (41.3500°N, 8.6833°W), it was cultured in Rose-Bengal Chloramphenicol Agar, a selective medium to supress the growth of bacteria and other microorganisms. After this first step, the mycelium was transferred to MEA and grown for 5 days at 25 °C.
2.2 DNA extraction and identification of the isolated fungal species
For the identification of the fungal species collected in the vineyard, DNA was extracted from mycelium of previously cultured fungus in Malt Extract Agar (MEA) for 5 days, using an automated nucleic acid extraction apparatus (MGISP-NE32, MGI tech GmbH©) with MagaBio Fungal Genome DNA Purification Kit (Bioer Technology©). DNA concentration and quality ratios were evaluated using a Thermo Scientific™ μDrop Duo plates in a Multiskan SkyHigh microplate reader. PCR amplification was performed with a NZY Taq II 2x Green Master Mix (with Taq polymerase, dNTPs, buffer and MgCl2), forward and reverse primers (ITS1 and ITS4) (Martin and Rygiewicz, 2005), ultra-pure H2O and the extracted DNA. The conditions for PCR amplification included an initial denaturation at 94 °C for 3 min, followed by denaturation at 94 °C for 21 s, annealing at 55 °C for 30 s, extension at 72 °C for 50 s, final extension at 72 °C for 5 min, during 35 cycles as described in the technical manual (NZYtech, 2024), using a T100 Thermal Cycler (Bio-Rad Laboratories©). Electrophoresis was performed in agarose gel with 1x TAE buffer and SYBR Safe dye (Thermo Fisher Scientific©) and the resulting gel was observed on ChemiDoc Imaging System (Biorad©). PCR products were purified using NZYGelpure kit from NZYtech© and sequenced at Eurofins Scientific. Sequences were checked for similarity using BLAST from NCBI. An additional set of primers were also used for multi-locus sequencing typing, namely, NL1 & NL4 for the D1/D2 domains of the 28S nrRNA gene region (LSU), gRPB1-A & fRPB1-C for partial fragments of the largest subunit of the RNA polymerase ІІ (rpb1) gene and RPB2AM-1bf & RPB2AM-7R for partial fragments of the rpb2 gene region (Wang et al., 2016). PCR conditions were similar to those previously described with the exception of the annealing temperature of the primers of rpb1 regions (51.5 °C) and the extension time (2 min for LSU and rpb2 and 30 s for rpb1). Sequences were deposited on GenBank with the accession numbers PV892941 (ITS), PV892836 (LSU), PV974811 (rpb1) and PV974810 (rpb2).
2.3 Preparation of E. globulus aqueous extract (EE)
The aqueous extract was obtained following the methodology of Pinto et al. (2021). Briefly, young leaves from new sprouts of E. globulus trees were collected and dried at 60 °C. Then, 250 g of dried leaves were fragmented into small pieces and underwent an extraction with 1 L of deionized water, at 70 °C for 30 min, with constant agitation. After this period, the leaves were separated from the liquid phase being this centrifuged at −4 °C and 150,00× g twice for 15 min. Finally, distilled water was added to the supernatant until the initial volume was achieved (concentration of 250 g dw L−1) and the obtained aqueous extract was stored at −80 °C until use.
2.4 Exposure of fungi to the EE and essential oil
Growth inhibition assays were performed following the methodology of Venâncio et al. (2017). Each fungus species was exposed to EE in Petri dishes (diameter of 90 mm) following two different methods. In the first method, the EE was incorporated in the MEA in the maximum concentration of 50% (v/v), which allowed the medium to solidify, in a final volume of 25 mL per plate. In the second method, the EE (non-diluted and diluted) was spread on the surface of the medium at concentrations of 100% and 50%, with a final volume of 500 µL per plate. As for the essential oil (Segredo da Planta™), different concentrations were incorporated into the medium (5, 2.5, 1.25, 0.63, 0.31, 0.16 μL mL−1) from a stock solution of 150 μL mL−1 prepared with Tween 20 (1:1), based on the previous study by Tomazoni et al. (2017). A control was also performed for both conditions with only distilled and sterilized water added to the MEA. When testing EO, a control with only Tween 20 was also prepared. Petri dishes were then inoculated with a single circular 7 mm mycelia plug removed from the edge of an actively growing colony for each fungus and incubated at the optimal temperature in the dark, at the temperatures reported in Section 2.1. Five replicates were performed for each treatment and the diameter of mycelial growth was recorded daily using a ruler. Each measurement was taken three times per plate, as also performed by Venâncio et al. (2017). The assay finished when all Petri dishes from the control with distilled water were completely filled with mycelium.
2.5 Exposure of fungi to Cu and folpet combined with EE
Two fungicides were tested in this study: Bordeaux Mixture (BM) using the commercial formulation Covicampo® Bordelés (containing 20% (w/w) of copper in the form of copper sulphate, provided by Vigo University) and the commercial formulation Folpetis WG (Selectis®) which contains 80% of folpet. These fungicides are already used in viticulture to control species as P. viticola and B. cinerea.
As described in the previous section, the combinations of EE and BM were tested by both methods: incorporation in the growth medium and surface application, with different concentrations used for each method. When incorporated, the corresponding weight of BM to obtain a concentration of 100, 80, 60% and 50% of the recommend dose (20 g L−1), was added to the MEA previously prepared, after cooling to 40 °C, and tested individually. BM in the different concentrations was also combined with EE diluted at 50%, according to Table 1. For this, MEA was first prepared with 50% of the total volume of water necessary, after cooling to 40 °C, BM in different concentrations was added (similarly to the individual testing), followed by the EE, in constant agitation. For surface application, the mixtures of EE plus BM and EE plus folpet were applied according to the concentrations shown in Table 1. In all the cases, a control using only distilled water was also prepared.
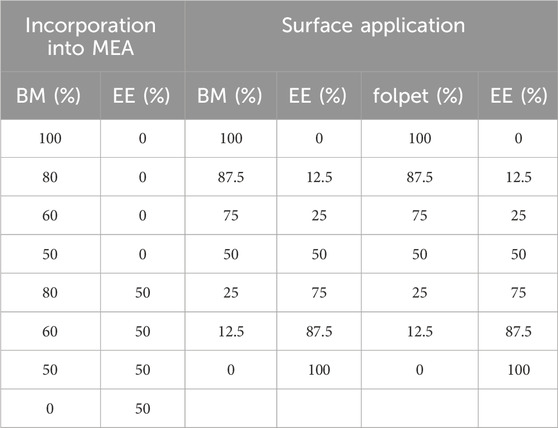
Table 1. Concentrations of Bordeaux Mixture (BM), E. globulus aqueous extract (EE) and folpet defined for the fungi growth inhibition assays according to different exposure methods. The percentages of BM and folpet are relative to the recommended dose of 20 g L−1 and 1.875 g L−1, respectively.
2.6 Data analysis
To analyse the effects of the different treatments on the four species tested, the software GraphPad Prism 8 (GraphPad Software Inc., San Diego, USA) was used. For each statistical test conducted, normality of the data and homoscedasticity of variances were assessed using Shapiro-Wilk and Levene’s tests, respectively, with a significance level of p ˂ 0.05. If data did not meet these assumptions, non-parametric (Kruskal-Wallis and Kolmogorov-Smirnov) tests were used instead. The effects of EE on the average maximum diameter achieved by the mycelium of the fungi at the end of the assay, applied to the surface or incorporated into the growth medium, were analysed using one-way ANOVA and unpaired t-tests, respectively. A comparison between all treatments was made in the case of surface application through the Tukey Test whereas for incorporation only a comparison between CTR and EE at 50% was made. For the effects of combinations of EE plus BM, whether incorporated or spread on the surface of the growth medium, on the average maximum diameter achieved at the end of the test, as well as folpet plus BM, one-way ANOVA was performed, followed by a multiple comparisons test (Tukey for EE plus BM and Dunn’s for folpet plus BM). For the effect of E. globulus EO at the last day of the assay, on the average maximum, a one-way ANOVA followed by a multiple comparison test (Tukey for L. sajor caju and B. cinerea and Dunn’s for P. chrysosporium and C. elatum) was performed whereas for the effects of EO throughout the fungi exposure, on the average daily diameter, a two-way repeated measures ANOVA was performed instead, using time and concentration as factors. Additionally, EC50 values, i.e., the concentration of EO that resulted in a 50% reduction effect on the fungal growth, as well as confidence intervals were estimated, using the software R (version 4.4.1. for Windows). For fitting the dose-response model and determining EC50, the drc package was used (Ritz et al., 2015). The drm() function was employed with a three-parameter Weibull type 1 model (W1.3) for L. sajor caju and B. cinerea, four-parameter log-logistic model (LL.4) for P. chrysosporium and three-parameter log-logistic model (LL.3) for C. elatum.
3 Results
3.1 Identification of the field isolated fungal species
DNA extraction and PCR amplification were conducted using 4 different sets of primers that codify different regions, namely, ITS1 and ITS4 for 5.8S ribosomal RNA gene, NL1 & NL4 for the D1/D2 domains of the 28S nrRNA gene region (LSU), gRPB1-A & fRPB1-C for partial fragments of the largest subunit of the RNA polymerase ІІ (rpb1) gene and RPB2AM-1bf & RPB2AM-7R for partial fragments of the rpb2 gene region. A BLAST search of the ITS region showed 98% coverage and 99.81% similarity with the reference species C. elatum (GenBank: MH871792.1). With LSU and rpb1 regions, an identification of C. elatum was made, showing a 100% coverage and 99.82% similarity with the reference strain CBS 126657 (GenBank: MH875647.1) for LSU and 98% coverage and 99.57% similarity with the reference strain CBS 374.66 (GenBank: KF001775.1) for rpb1. Rpb2 region showed 99% coverage and 100% similarity with the reference species C. elatum (GenBank: MG890098.1).
3.2 Effects of EE on non-target and target fungi
The analysis of fungal growth when exposed to EE spread on the surface of the MEA showed no significant effects on the non-target species L. sajor caju and P. chrysosporium nor on C. elatum (Table 2), at both concentrations tested (100% and 50%). However, for B. cinerea, a significant reduction in growth was observed at 100% of EE (H = 14.00, df = 2, p = 0.0010), applied on the surface of the agar, though growth of the mycelium was similar to the control when the EE was diluted to 50%. In contrast, when EE was incorporated into the MEA, it demonstrated enhanced efficacy, inhibiting the growth of the fungi C. elatum and both non-target species (for L. sajor caju: t (4) = 31.77, p < 0.0001; for P. chrysosporium: t (4) = 30.32, p < 0.0001), significantly in relation to the control (Table 2). These findings suggest that according to the exposure methodology, fungal response will vary.
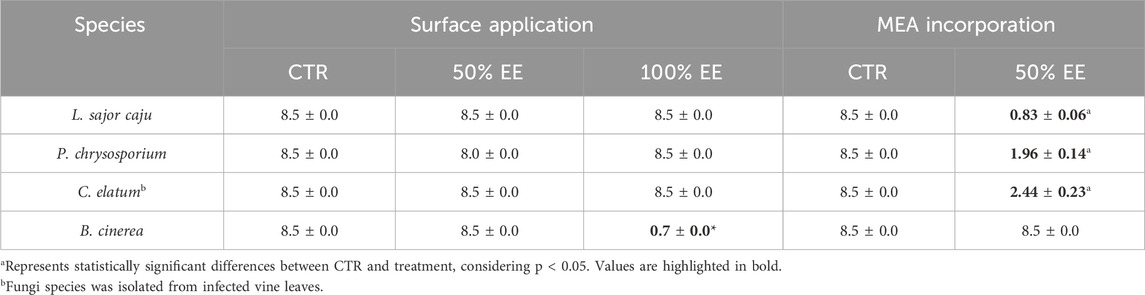
Table 2. Average maximum diameter (cm) of non-target (L. sajor caju and P. chrysosporium), one potentially target (C. elatum), and target (B. cinerea) species exposed to E. globulus aqueous extract (EE) by two different methods: surface application and incorporation into the growth medium (MEA), on the last day of the assay.
3.3 Effects of the combination of EE plus BM on non-target and target fungi
The effects of different combinations of BM with EE, applied to the surface of the MEA, on the four fungi species are shown in Figure 1. No statistically significant changes were recorded for L. sajor caju, P. chrysosporium or C. elatum, as their growth at the end of the assay was similar to that of the control. Thus, no standard deviation was recorded as the radial growth of the fungi achieved the limit of the plate at the same time as in the CTL, in all the replicates. On the other hand, for B. cinerea, significant growth inhibition was observed. The 100% EE treatment was the most effective, reducing growth by 88.5% compared to the control while also being statistically significantly different from the remaining treatments (F (7, 32) = 8.106, p < 0.0001). Moreover, it seems that the treatments with higher EE generally resulted in greater growth reduction when comparing with lower EE concentrations.
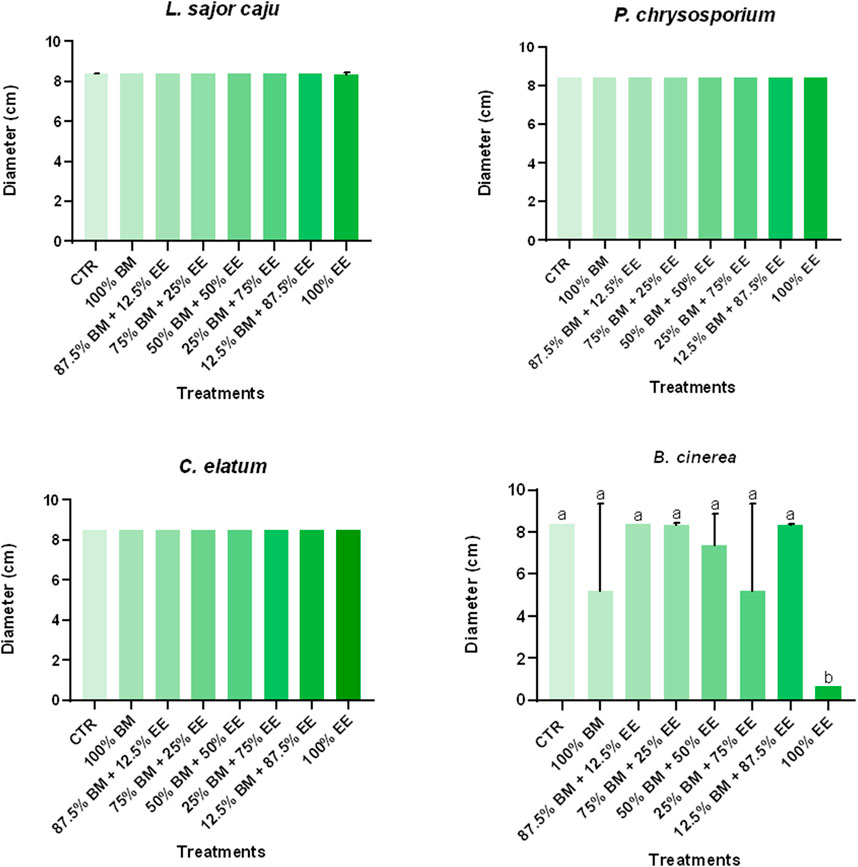
Figure 1. Average diameter of non-target (L. sajor caju and P. chrysosporium), one potentially target (C. elatum), and target (B. cinerea) fungi species achieved on the last day of the assay under exposure to a mixture of EE and BM in different concentrations, spread on the surface of the growth medium (MEA). Different letters represent statistically significant differences between treatments (Tukey test: p < 0.05). The maximum diameter that can be achieved by the mycelium when no growth inhibition is registered is 8.5 cm.
On the other hand, the incorporation of BM, both individually and in combination with EE, into the MEA (Figure 2) had measurable effects on the growth of all tested species. For L. sajor caju, significant differences between the treatments were found (F (8, 36) = 28.72, p < 0.0001). For BM exposure, only the 50% BM significantly reduced growth, compared to the control. Similar effects were observed in treatments containing 80% BM + 50% EE, 50% BM + 50% EE and 50% EE. Overall, the combination of BM with EE seemed to be more detrimental for this fungus. The same tendency was also observed for the other non-target fungus, P. chrysosporium. In all the treatments, a statistically significant reduction of growth was registered in relation to the control (F (8, 36) = 2,728, p < 0.0001). Additionally, the treatments with the combination of BM + EE as well as EE alone, proved to be the most efficient in inhibiting the growth of this species. When comparing the sensitivity of the two non-target species, P. chrysosporium was more affected, with a greater growth reduction across all treatments than L. sajor caju, at the end of the assay.
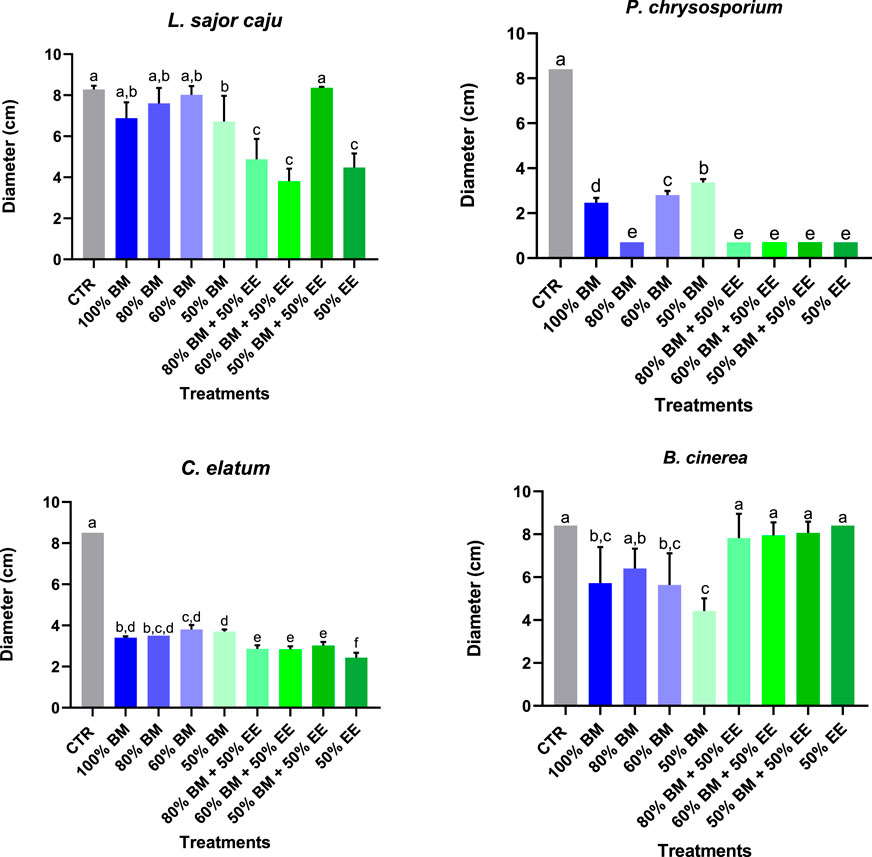
Figure 2. Average diameter of non-target (L. sajor caju and P. chrysosporium), one potentially target (C. elatum), and target (B. cinerea) fungi species achieved on the last day of the assay, under exposure to a mixture of EE and BM in different concentrations, incorporated on the growth medium (MEA). Different letters represent statistically significant differences between treatments (Tukey test: p < 0.05).
Similarly to P. chrysosporium, the growth of C. elatum was significantly affected by all treatments (F (8, 36) = 726.6, p < 0.0001), with the combination of BM and EE, as well as the 50% EE, being slightly more detrimental to this species than BM alone. In the specific case of the target species B. cinerea, no statistically significant changes were observed in treatments combining BM and EE, however, BM at concentrations of 100%, 60% and 50% significantly reduced its radial growth (F (8, 36) = 11.80, p < 0.0001). Additionally, it is also possible to notice that, in terms of species sensitivity, the growth of C. elatum was more affected in all treatments comparing with B. cinerea.
3.4 Effects of the combination of folpet and EE on non-target and target fungi
The results of the application of different combinations of the fungicide folpet and EE on the surface of the MEA are shown in Figure 3. Particularly on this assay, only the recognized target species was considered as this fungicide is recommended to be used against B. cinerea. For L. sajor caju (H (7) = 31.26, p < 0.0001), the highest concentrations of folpet (100% and 87.5% FP + 12.5% EE) significantly inhibited the growth of the fungi, with inhibition rates of 51% and 48% respectively. However, as the concentration of this fungicide decreases and EE increases, the growth reaches control levels, with no significant changes. Similarly, P chrysosporium (H (7) = 32.93, p < 0.0001) also had a significant decrease in growth in 100% FP of 7.1% in relation to the control, however, in the remaining treatments no significant changes were registered. In addition, P. chrysosporium seemed to be less affected by these treatments compared to L. sajor caju. This result agrees with previous assays in what regards no effect of EE, when applied to the surface of the MEA. The target species, B. cinerea (H (7) = 29.04, p = 0.0001), had similar results to L. sajor caju, as the growth was significantly affected at the highest folpet concentration, having an inhibition of 57.6% compared to the control, followed by an increase as the concentration of folpet decreases and EE increases. Somewhat surprisingly, was the no effect observed on the growth of this target species, when exposed to 100% EE, contrary to what was observed on the previous two assays, where this concentration was tested against B. cinerea.
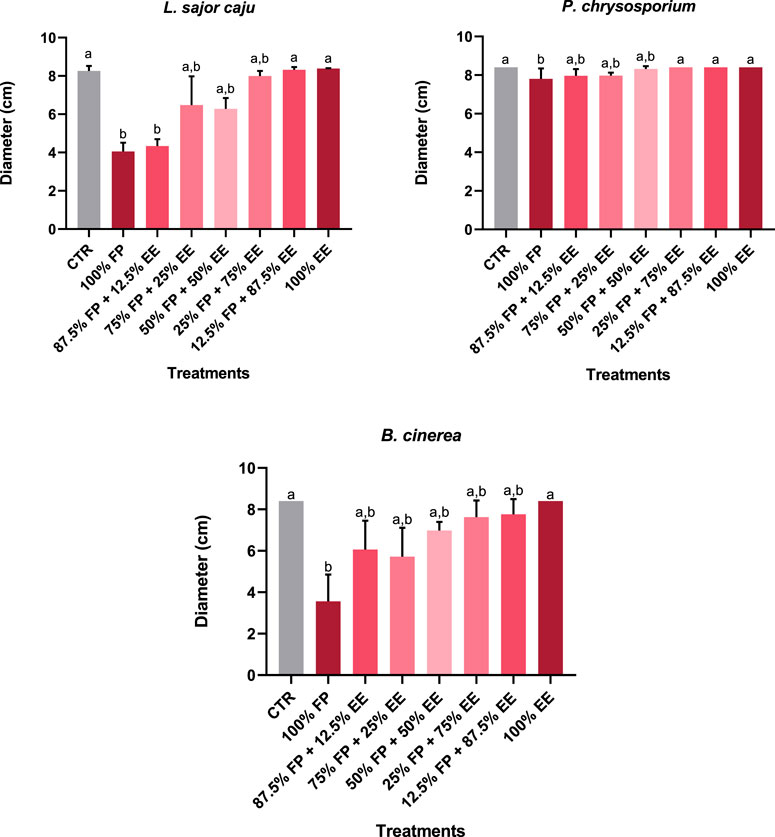
Figure 3. Average diameter of non-target (L. sajor caju and P. chrysosporium), target (B. cinerea) fungi species achieved on the last day of the assay, under exposure to a mixture of EE and folpet in different concentrations, spread on the surface of the growth medium (MEA). Different letters represent statistically significant differences between treatments (Dunn’s test: p < 0.05).
3.5 Effects of EO on non-target and target fungi
The efficacy of an E. globulus commercial essential oil (EO) on fungal growth was also evaluated in this study. A two-way ANOVA was performed to understand the effect of EO concentration and time on fungi growth. For all the species tested, statistically significant interaction was observed between these factors (Table 4). Simple main effects analysis showed that both EO concentration and time had a statistically significant effect on all species growth (Table 3).
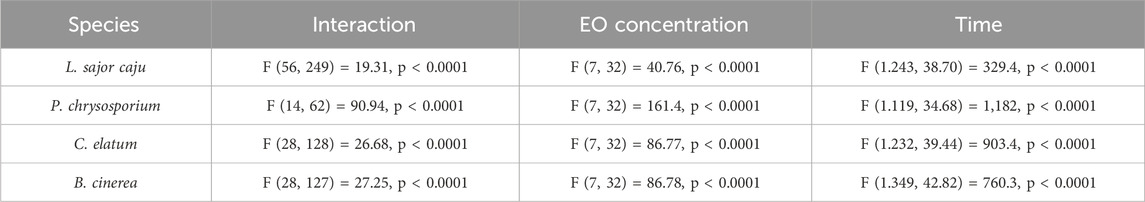
Table 3. Two-way ANOVA results for the effects of E. globulus essential oil (EO), time of exposure and the interaction between factors on the growth of two non-target (L. sajor caju and P. chrysosporium), one potentially target (C. elatum) and one target (B. cinerea) fungi species.
Looking in more detail at the growth of all fungi species exposed to different concentrations of EO, over time (Figure 4), and for L. sajor caju, there was a total growth inhibition (100%) or more prolonged lag-time, that achieved 7 days and the fungi did not recovered, displaying a radial growth inhibition of 92% and 76% at the end of the assay at 5.00 and 2.50 μL mL−1 EO, respectively (Figure 5). Notwithstanding, no significant differences were registered between days in these two concentrations. For the remaining EO concentrations, even though the growth rate was slow in the initial 4 days, the fungi was able to grow steadily, and no significant changes were registered at the end of the assay, comparing with the control. As for the Tween, it is visible that fungi growth decreased upon exposure, and significant effects in relation to the control were registered. However, after day 9 of the assay, these effects were no longer significant.
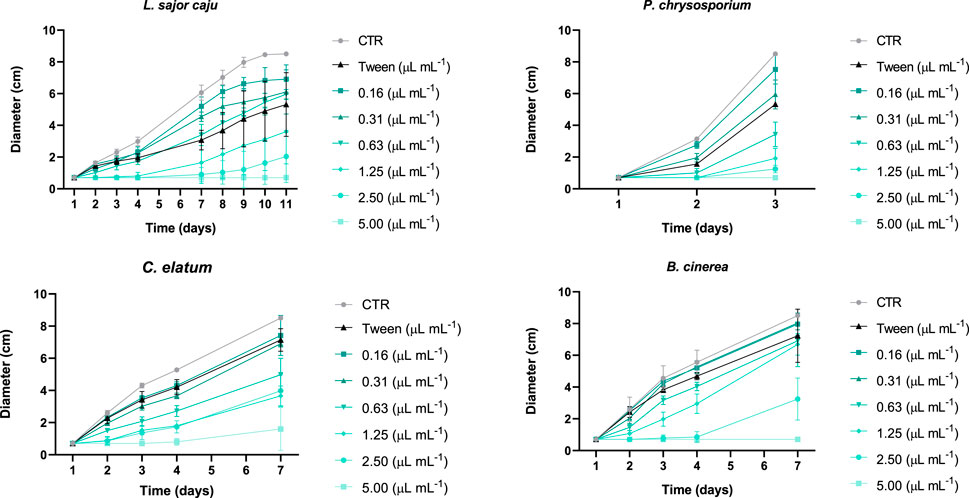
Figure 4. Effect of the application of E. globulus essential oil on the growth of different fungi species (non-target (L. sajor caju and P. chrysosporium), one potentially target (C. elatum) and one target (B. cinerea)) expressed as diameter (cm) over time (days). Error bars correspond to the standard deviation.
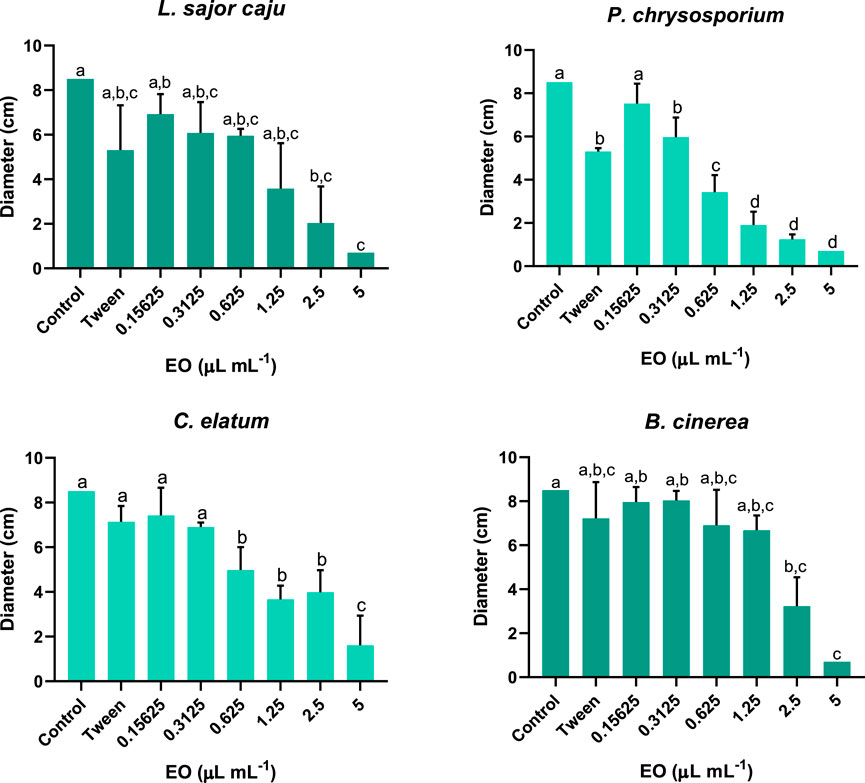
Figure 5. Average growth, expressed in centimetres, for four fungal species, L. sajor caju, P. chrysosporium, C. elatum and B. cinerea, exposed to different concentrations of E. globulus essential oil (EO), recorded at the last day of the assay. Error bars correspond to the standard deviation. Different letters represent statistically significant differences between treatments (Tukey and Dunn’s test: p < 0.05).
Regarding P. chrysosporium, the assay was much less extensive than that of the previous fungus, having a total duration of 3 days, as this species has a highest growth rate. At the control and at the lowest concentration of EO (0.16 μL mL−1) the fungus had a significantly different diameter when compared with the other treatments at day 2 (F (7, 32) = 170.0; p < 0.0001). When comparing the mean diameter of P. chrysosporium at the last day of the assay (Figure 5), except for the lowest EO concentration, all the other treatments were significantly different from the control, showing a growth decrease in a dose-response manner (F (7, 29) = 98.47; p < 0.0001). But clearly concentrations higher than (0.63 μL mL−1) caused a greater growth inhibition when compared with the lowest concentrations and the Tween (F (7, 29) = 98.47; p < 0.0001). Indeed, the exposure to Tween was detrimental to the growth of P. chrysosporium, showing a significant decrease of 37.6% at the end of the assay, compared to the control.
For C. elatum and the target species, the duration of the assay was the same, with the fungi in the control group occupying the entire Petri dish after 7 days of incubation. The exposure to EO showed significant effects on the growth of C. elatum having these occurred already at day 2 for the highest concentrations up to 0.63 μL mL−1 when compared to the CTL, being these differences maintained until the end of the assay, as can also be seen in Figure 5. Contrary to what was observed for the other three species, the highest EO concentration of 5.00 μL mL−1 did not totally inhibited the growth of C. elatum. Even though in the initial 4 days of the assay no growth was recorded, it seems that the fungus was able to tolerate the toxicity and recover its ability to grow. Nonetheless, EO concentrations above 0.63 μL mL−1 had the highest toxicity towards this species. Similarly, to B. cinerea, the treatment with Tween was not detrimental to this fungus, as, with the exception of day 4, no significant effects were recorded compared to the control, for the remaining days of the test.
Similarly, to L. sajor caju, B. cinerea growth was negatively affected by the two highest EO concentrations, reaching inhibitions of 62.1% at 2.50 μL mL−1 and 100% 5.00 μL mL−1, compared to the control, at the last day of the assay (Figure 5). These negative effects were immediately observed on the second day, the same also occurring for 0.63 and 1.25 μL mL−1. As for Tween treatment, B. cinerea growth was not significantly affected with this exposure when compared to the control, throughout the duration of the assay.
In terms of species sensitivity, from Figure 5, that better shows the effect on the fungi growth at the end of the assay, and from the EC50 values presented in Table 4 it is possible to conclude that, P. chrysosporium was the most sensitive one (0.454 μL mL−1), followed by L. sajor caju (0.825 μL mL−1), C. elatum (1.20 μL mL−1), and B. cinerea (2.19 μL mL−1). This demonstrates that non-target species were overall more sensitive to the Eucalyptus EO than the target ones.
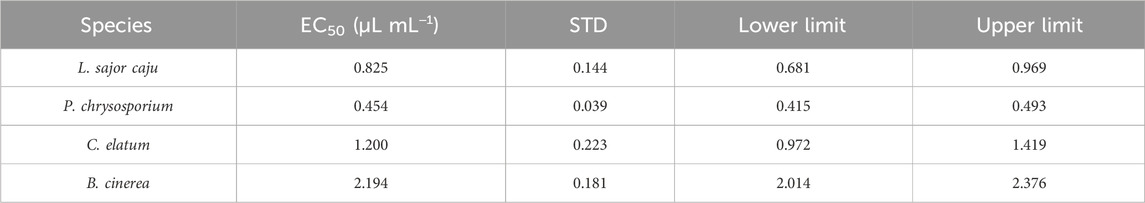
Table 4. EC50 values for the effect of E. globulus essential oil (EO) on the growth of two non-target (L. sajor caju and P. chrysosporium), one potential target (C. elatum) and the target (B. cinerea) fungi species. EC50 is the EO concentration (µL mL−1) that can inhibit in 50% the growth of the four species tested. Standard deviation (STD) and lower and upper limits are also presented.
4 Discussion
C. elatum, the species isolated at GreenUPorto vines, was already reported in Portugal in 2014 in olive trees (Global Biodiversity Information Facility, 2014), however, it is still a poorly known species and, to the best of our knowledge, there are no registered appearances in grapevines. Nevertheless, these species also deserve all attention, before becoming a problem once the conditions for their incidence and spread is achieved. Based on existing information about this species (herein discussed) it is assumed as a potential pathogenic/target species.
The present study focused on the performance of E. globulus aqueous extract (EE) and essential oil (EO) on the growth of non-target, potentially target, and target fungi species, in a perspective to be used as fungicide or to reduce the use of active ingredients candidates for replacement, as is the case of copper (European Commission, 2018). In addition, different application methods were also tested as to understand the behaviour of the fungi under different types of exposure, considering that when fungicides are applied to treat fungal incidences, they mainly persist on the cuticle of leaves. As previously mentioned, the efficacy of extracts obtained from Eucalyptus leaves was already tested by several authors, against fungi species that are responsible for agronomic losses, either during production or in the post-harvest (Ahmad et al., 2020; Bouhlali et al., 2020; Hajji-Hedfi et al., 2024; Yao et al., 2021). In most of the studies already available, the preparation of the extracts was made using dried leaves, however, the state of development of the Eucalyptus trees when the leaves were collected is not mentioned. Indeed, young leaves as the ones collected for this study are visually different than the older ones and, as they are closer to the ground, they are more susceptible to herbivory thus it is possible that they have higher amounts of defensive specialized metabolites with biocidal properties (Pinto et al., 2021). Moreover, some of these extracts were prepared using ethanol, ethyl acetate and petroleum ether, making the extraction process more complicated and expensive in terms of future commercial use. Despite this variability on extracts preparation, these natural plant-based products are often seen as viable substitutes of fungicides due to their richness in volatile compounds, mainly from the monoterpene and sesquiterpene classes, and in polyphenols, such as ellagitannins, flavonols, and phenolic acids, being all these compounds reported as the ones responsible for the biocidal effect (Boulekbache-Makhlouf et al., 2013; Pinto et al., 2021; Pombal et al., 2014; Yong et al., 2019). Monoterpenes and sesquiterpenes are known for their lipophilic structures, which allows them to penetrate fungal cell membranes, disrupting its integrity, leading to leakage of essential ions and cellular content, ultimately leading to an inhibition of cellular respiration and energy production. Moreover, some terpenes are also capable of inhibiting fungal enzymes with key roles or even altering the fluidity of the cell membrane, thus impeding nutrient uptake (Câmara et al., 2024). As for polyphenols such as ellagitannins, flavonoids and phenolic acids, they have several modes of action in terms of antifungal activity. Ellagitannins, not only do they affect cell wall synthesis and permeability, but they can also deprive fungal cells of necessary nutrients and be responsible for oxidative stress and cellular damage. Generally, flavonoids can interfere with the cell wall but some can also inhibit fungal enzyme systems and disrupt DNA synthesis, successfully inhibiting fungal reproduction and produce reactive oxygen species (ROS), leading to cellular damage and death (Dias et al., 2021). Similarly to flavonoids, phenolic acids also affect cell wall synthesis and cell membrane integrity, interfering with intracellular signalling, and induce oxidative stress, damaging DNA, proteins and lipids, ultimately resulting in cell death (Hajji-Hedfi et al., 2024). Thus, in general, the biocidal effects and the mode of action are frequently described for individual components of the extracts rather than for the aqueous extracts on itself. Notwithstanding, in all studies testing aqueous extracts, the authors observed a decrease in mycelial growth, spore germination, and sporulation in a dose-dependent manner (depending on the endpoint measured) in fungi species such as A. mali, B. dothidea, D. seriata, F. oxysporum, C. gloeosporioides, A. alternata, A. niger, B. cinerea, P. digitatum, P. expansum, P. italicum and R. stolonifera (Ahmad et al., 2020; Bouhlali et al., 2020; Hajji-Hedfi et al., 2024; Yao et al., 2021). Additionally, only one application method (predominantly the method of medium incorporation) was tested.
In the present study, the exposure of L. sajor caju, P. chrysosporium and C. elatum to 100% and 50% of EE dispersed on the surface of the MEA showed no effects on the growth of these species. It cannot be neglected that in this methodology of exposure, the effective concentration of EE to which the fungi are exposed is lower than when EE is incorporated into the growth medium. Nevertheless, B. cinerea with a wide range of hosts, also known as a highly destructive pathogen, being ranked as the second species that causes more economic losses (Ullah et al., 2024 and references quoted herein), seemed to be sensitive to the 100% EE, with a remarkable inhibition of growth, at least in two out of three of the tests performed. However, when EE was incorporated into the growth medium an opposite trend was observed for B. cinerea, but not for the remaining species, being these significantly affected and showing a significant decrease in radial growth. In fact, all these fungi species are filamentous fungi which means that they grow through branching of hyphae that allows them to penetrate the culture medium in search of nutrients (Osma et al., 2011). Being the source of nutrient supply contaminated, it would be expected a decrease in growth. Nevertheless, B. cinerea was not affected by the incorporation of EE on MEA and, this can likely be explained by the complex multifunctional and multilayer mechanism of infection evolved by this species (Bi et al., 2023). In a former stage of infection B. cinerea combines the penetration of the host tissues with the surface development of hyphae. This surface development facilitates penetrating fungal cells to release cell death inducing proteins (CDIP) that will attack plant cells. Nevertheless, these proteins also penetrate fungal cells, as secondary effect. Still, the fungi is compensated by the development of plant necrotic tissues that will support the continuation of its infection and proliferation (Bi et al., 2023). This physiology of infection is likely the explanation for the contradicting responses of the tested fungi to the methodology of exposure and it should be considered whenever the sensitivity of fungi to different types of substances is to be analysed.
When the fungicidal effect of BM isolated and in combination with EE was tested, applied on the surface of the growth medium, none of the species was affected. B. cinerea was the only species that displayed a significant reduction in its radial growth once again when exposed to the non-diluted EE. On the other hand, when BM and EE were tested incorporated into the MEA (both isolated and combined), once again and consistently to what was observed in the first assay, B. cinerea was not the most sensitive species. BM alone was able to cause a significant inhibition in B. cinerea growth at concentrations of 100%, 60% and 50% of the recommended dose. However, BM alone only caused a maximum of 47.5% reduction in the growth of this fungi. The mixtures of BM and EE have not displayed any effect on the growth of this species. The limited fungicidal effect of BM on B. cinerea is in fact recognized. Usually, to treat the disease caused by this pathogenic fungi, synthetic fungicides such as hydroxyanilides, anilinopyrimidines, carboxamides, phenylpyrroles, and phtalamine are recommended (DRAPN, 2022). Nonetheless, these results suggest that copper-based fungicides could also have some protective effect on crops against this pathogen when they are applied to control P. viticola. A decrease in B. cinerea growth exposed to copper containing products was also observed by several authors. In light of increasing fungicide resistance, recent studies conducted by Parada et al. (2024) and Malandrakis et al. (2020) evaluated the efficacy of copper nanoparticles (Cu-NPs) on supressing B. cinerea growth and both studies reported an inhibition at Cu-NPs concentrations of 20, 40 and 160 mg L−1 and 300 and 500 μg mL−1, respectively. Even though both studies utilized purchased Cu nanoparticles, the ones from Parada et al. (2024) are oxide nanoparticles with <50 nm particle size whereas in the study of Malandrakis et al. (2020), particle size was 25 nm. In another study by Rodríguez-Ramos et al. (2023) where the effects of copper oxychloride (Cu-Oxy) in B. cinerea were assessed, 2 g L−1 of both commercial Cu-Oxy and pure Cu-Oxy were able to inhibit the fungi growth. Additionally, when evaluating the prevalence, spatial distribution and potential fungicides of cranberry fruit rot pathogens, Wood et al. (2023) stated that copper based fungicides were highly effective in inhibiting the mycelial growth of all fruit rot pathogens. Nevertheless, it is important to highlight that those studies are analysing different chemical forms of copper, than the ones tested in this study, what makes it difficult to elaborate robust conclusions. Nevertheless, BM is one the most common Cu-based product used in viticulture. Even though Cu is essential for the growth of fungal species, when in excess, it could lead to an upregulation of laccases to alleviate Cu toxicity, by scavenging reactive oxygen species, acting as a defence mechanism against Cu (Buddhika et al., 2021; Janusz et al., 2020). This mechanism, within others, is probably highly efficient in B. cinerea, a species that is known for expressing substantial amounts of extracellular laccase using a wide range of substrates as phenols, aromatic and aliphatic amines, hetero- and alicyclic compounds (Slomczynski et al., 1995) explaining the lower efficiency of BM on controlling this pathogen.
As for the remaining species, both the non-target fungi species and C. elatum demonstrated once again to be sensitive to the EE extract, and it was also proven that the Eucalyptus extract enhanced the growth inhibitory effect of the BM. But even more interesting is that 50% EE had the highest inhibitory effect (71.3%) on C. elatum, when compared with the mixture, being the EE likely a potential good candidate for copper replacement or at least for reducing its use. However, there is still little knowledge on the role of C. elatum, which limits the interpretation of these results. Moreover, some studies highlight the ability of this species to act as an antagonist, preventing diseases through antibiosis (Dwibedi et al., 2023), while other studies highlighted their capacity to switch its role from opportunistic colonizer to pathogen depending on abiotic and biotic conditions (Violi et al., 2007). As for the non-target fungi (L. sajor caju and P. chrysosporium) these were also sensitive to EE and EE + BM mixtures, in particular P. chrysosporium which was the most sensitive species. This result is most likely related to the morphology and physiology of these species, being commonly called white rot fungi. Both L. sajor caju and P. chrysosporium are Basidiomycota that are able to degrade wood using enzymes secreted by the hyphae (Goodell et al., 2020). These enzymes, mainly oxidative, such as laccase, lignin peroxidase, manganese peroxidase, versatile peroxidase and the dye-decolorizing peroxidases as well as carbohydrate active enzymes are able to break down lignin, cellulose and even hemicellulose, leading to the enzymatic erosion of the cell wall layers, allowing the hyphae to penetrate the wood (Goodell et al., 2020; Tura et al., 2016).Through the elongation of the tips of the hyphae, the fungi starts to seek pathways within the structure of the wood in order to ramify and grow while also absorbing nutrients, serving the role of building components in the area where they are growing (Sydor et al., 2022). This is only possible due to the fact that fungi hyphae are thin and flexible allowing them to spread in different directions and penetrate different substrates (Aleklett et al., 2021; Money, 2007). Thus, cultivating these fungi species using growth mediums is a common and successful practice in laboratory (Geris et al., 2024). In the case of this study, it seems that as hyphae penetrated the contaminated growth medium, they were in direct contact with the treatments probably absorbing its contents, leading to toxicity, ultimately resulting in the inhibition of growth. Considering all that was discussed so far, it is necessary that both EE and EE + BM mixtures are tested for other non-target species, before recommending their application to combat the potential pathogenic species as C. elatum, as well as other pathogens.
When it comes to synthetic PPP application in viticulture, fungicides are also problematic in terms of ecosystems’ health, but its necessity is undeniable due to climate change and the increasing prevalence of fungal disease. This is the case of folpet, a fungicide that belongs to the group of phtalamids, that is commonly used in viticulture to control the growth of fungi species responsible for diseases such as grey mould, powdery mildew and black rot (DRAPN, 2022; Lamberth, 2019). Although this active substance, as well as its degradation products, have a very low or low persistence in soil, their residues were already detected in soil and water samples, and connected with (eco)toxic effects predicted for human health and aquatic organisms through modelling (Rosenbaum et al., 2008; Vallejo et al., 2019; Vázquez-Rowe et al., 2012). More concerning is the fact that some studies have also reported the presence of this fungicide in table grapes and wine, which could ultimately impact human health (Alonso González et al., 2022; Oulkar et al., 2019). In consequence, for example, in Portugal, its application was restricted for table grapes (DRAPN, 2022). To also explore the possible use of the Eucalyptus extract (EE) to replace or to reduce the dosages of this synthetic fungicide, mainly used to combat Botrytis, in the current study, Folpet alone and in combination with EE, were only spread on the surface of the growth medium, considering that B. cinerea has proven to be more sensitive to this exposure methodology. As expected, 100% of the recommended dose of folpet inhibited the growth of B. cinerea in 57.6% in relation to the control, and both non-target fungi were also significantly affected by this treatment. L. sajor caju proved to be as sensitive as the pathogenic species to this fungicide. The mixture with EE, was not as efficient as the folpet alone in the control of the B. cinerea, which is a concern related with the impact of this fungicide on relevant species of the ecosystems, and subsequently on their functions. As for the treatment with 100% EE and contrarily to what was observed in the previous assay, it did not inhibit the growth of B. cinerea, having recorded the same diameter as the control. This was an inconsistency found among these results that could indicate that the method used to prepare the EE might not allow the production of a product with consistent composition, due to differences in the plant material used, within other aspects. Particularly, the temperature used to prepare the extract, may account the for the volatilization of relevant compounds in terms of their allelopathy, introducing variability in the final composition of the extract. Something that needs to be addressed in future studies. It is also important to highlight the lack of studies on the effects of folpet on non-target fungi species, with key roles on the ecosystems, evidence brought by these species. Nevertheless, the effect of folpet on the growth of both white-rot fungi was not a surprise as this N-(trichloromethyl[thio]phtha-limide) fungicide has a broad-spectrum mode of action, by inhibiting microbial cells division (Zhong and Duncan, 2020).
Eucalyptus globulus essential oils proved to be effective in controlling the growth of the target species B. cinerea and of the potential target species C. elatum. On the other hand, the non-target species were also highly sensitive to the application of EO, representing a limitation to the use of this product as a fungicide, as it was observed for the synthetic fungicide. Indeed, white rot fungi such as L. sajor caju and P. chrysosporium, play a critical ecological role in decomposing lignin and recycling nutrients in agricultural soils (Bing-Ru et al., 2006; Galindo et al., 2013). By applying EO these processes could be disrupted, leading to reduced nutrient cycling, diminished soil fertility, and negative impacts on crop productivity. To mitigate these negative effects, strategies such as encapsulation or controlled-release formulations, localized application, synergistic combinations with other products to reduce the required concentration, and temporal targeting—applying treatments when non-target fungal species are less active—could be employed.
Regarding B. cinerea, the efficacy of E. globulus EO in controlling its growth was already evaluated in studies conducted by Fialho et al. (2017) and Diánez et al. (2018), where the authors stated that E. globulus EO was one of the most fungitoxic towards these species. EO from other plant species also proved to have antifungal activity, such as the ones from Cinnamomum zeylanicum, Origanum majorana, Melaleuca alternifolia, Mentha piperita, O. vulgare and Thymus vulgaris (Fialho et al., 2017; Rienth et al., 2019; Vigneron et al., 2023). In contrast, to the best of our knowledge, no studies were conducted so far on the effects of E. globulus EO on L. sajor caju and P. chrysosporium. However, there are studies on Trametes versicolor, also a white-rot fungi that belongs to Basidiomycota phylum. The study from Matan et al. (2009) demonstrated the antifungal activity of E. globulus EO incorporated in the growth medium in concentrations between 100 μL mL−1 and 800 μL mL−1, much higher than the ones used in this study, being the minimal inhibitory concentration of 600 μL mL−1 for this species. This effect was mainly attributed to the presence of eucalyptol, but the authors also indicated that a synergistic effect between minor constituents could also explain the inhibitory effect of this EO on T. versicolor growth. The fact that much higher concentrations of EO were needed to significantly reduce T. versicolor growth might suggest that EO chemical composition might also differ from the one used in the current study, although a remarkable difference in sensitivity between species might also result from different species physiology. One thing that cannot be neglected is the effect that Tween 20 (tested at the highest concentration of 5.00 μL mL−1) had on P. chrysosporium, as it was able to significantly reduce its growth. Thus, the effects that were observed for the remaining concentrations of EO might also have been caused by the presence of this surfactant and not to the EO alone. Future studies using Tween 20 alone as control for all EO concentrations should be considered to better understand this effect. Nonetheless, the use of Tween or other surfactants cannot be totally removed as they are essential to obtain an homogenous suspension and for EO to be able to penetrate the cells of the target organisms (Dudek-Wicher et al., 2021).
It is also important to highlight the appearance of C. elatum in vine plants, as up until this point, it had only been identified in olive trees in Portugal (Global Biodiversity Information Facility, 2014). Chaetomium elatum is a significant saprotrophic mold fungus of the genus Chaetomium, with occurrence in a wide variety of substrates. They are normally found in soil, air, plant, and cellulose debris and they are also capable of generating a broad spectrum of biologically active compounds of biotechnological interest (Dwibedi et al., 2023). However, in terms of pathogenicity, little is known regarding this species. In a study conducted by Violi et al. (2007), the authors established that under certain conditions, C. elatum functioned as a weak pathogen, being able to kill avocado plants. However, in the presence of the arbuscular mycorrhizal fungus Glomus intraradices, no negative impacts of C. elatum was detected. According to the authors, it seems that C. elatum has a multifunctional ability, as it can be both a rhizoplane and rhizosphere fungus, opportunistically colonizing plant roots and turning pathogenic only under extreme resource scarcity and intense intraspecific competition. In the specific case of vine plants, according to The American Phytopathological Society (APS), C. elatum is a disease of Vitis vinifera (Wilcox et al., 2015) inserted on the category of other berry rots and secondary invaders, raisin molds, and rots, however, the mode of infection of this species is not well-documented so far. Thus, attention should be given to this species as it could become a problem to grape production, especially in a context of climate change, where favourable conditions could arise for its incidence.
5 Conclusion
In summary, this study addressed the potential use of E. globulus aqueous extract (EE) and essential oils (EO) as sustainable and effective alternatives to the use of conventional plant protection products such as Bordeaux Mixture (BM) and folpet. EE demonstrated promising antifungal activity in this first in vitro screening, being its effectiveness influenced by the exposure method (incorporated or not in the culture medium). While B. cinerea growth was significantly affected when EE was spread on the surface of the growth medium (MEA), the same was not observed for the remaining species tested (C. elatum, L. sajor caju and P. chrysosporium) where the incorporation of EE into the MEA was more effective, being this difference likely related to the morphology, physiology, and mode of infection or spread of these species. This differentiated response depending on the method of exposure in the plate is something that needs to be considered in future studies that aim to assess the effect of chemical substances on fungi growth. When evaluating the possibility of reducing the amount of BM and folpet applied by combining it with EE, an enhanced efficacy of BM plus EE was recorded for C. elatum but not for B. cinerea, however, the same was not observed in the case of the mixture of folpet and EE. Even though this may demonstrate a potential use of EE to reduce PPP application, inconsistency of results regarding B. cinerea growth exposed to 100% EE on the surface of the MEA was also found. Thus, the preparation of these E. globulus aqueous extracts should be further investigated, adopting other methods such as the exchange of water for alcoholic solvents when preparing the extract, reducing the temperature used for the extraction process from E. globulus leaves or even adopting a closed system for these extractions. This optimization of extraction procedure is crucial for a further standardization of the methodology. Regarding EO, even though they were strongly effective in supressing the growth of B. cinerea and C. elatum, the non-target species were also very sensitive to these treatments, which poses a limitation to the use of these products as fungicides, before its impacts on other non-target species and ecosystem processes being assessed through field assays. The efficacy of EO in combination with these PPP should also be considered in the future to evaluate whether this mixture would be less detrimental to the non-target fungi species of the ecosystem.
Taking into consideration all of this information, the use of these plant-based products should be approached with caution, ensuring that their effects on ecologically important organisms with key functional roles are carefully evaluated beforehand. Furthermore, field studies are essential not only to determine the efficacy of potential biocides such as EOs, EE, and their mixtures with BM and folpet, but also to assess their impact on non-target species and ecosystem processes under natural conditions. This study shows that the replacement of copper-based products in viticulture is a highly demanding process, as it require in depth studies of the alternatives.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/supplementary material.
Author contributions
BF: Conceptualization, Formal Analysis, Methodology, Writing – original draft. RD: Formal Analysis, Investigation, Methodology, Writing – review and editing. CP: Investigation, Methodology, Writing – review and editing. JO-P: Investigation, Methodology, Writing – review and editing. AC: Conceptualization, Formal Analysis, Methodology, Supervision, Writing – review and editing. RP: Conceptualization, Formal Analysis, Methodology, Resources, Supervision, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by national funds through the FCT – Foundation for Science and Technology, I.P., under the project/support UID/05748/2025; BF (UI/BD/151040/2021), CP (2023.05266.BDANA) and RD (2024.01319.BD) acknowledge FCT for providing PhD scholarships.
Acknowledgments
The authors want to acknowledge Artur Alves and Teresa Rocha-Santos from University of Aveiro for providing fungal cultures of L. sajor caju, P. chrysosporium and B. cinerea.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Acar, Ü., Erden, Y., Inanan, B. E., Kesbiç, O. S., Yılmaz, S., and Çelik, E. S. (2023). Evaluation of metabolic stress status in common carp (Cyprinus carpio) exposed to the fungicide folpet. Comp. Biochem. Physiol. Part C 263, 1–6. doi:10.1016/j.cbpc.2022.109494
Adams, E., Gerstle, V., and Brühl, C. A. (2021). Dermal fungicide exposure at realistic field rates induces lethal and sublethal effects on juvenile European common frogs (Rana temporaria). Environ. Toxicol. Chem. 40, 1289–1297. doi:10.1002/etc.4972
Águas, A., Ferreira, A., Maia, P., Fernandes, P. M., Roxo, L., Keizer, J., et al. (2014). Natural establishment of Eucalyptus globulus Labill. in burnt stands in Portugal. For. Ecol. Manage. 323, 47–56. doi:10.1016/j.foreco.2014.03.012
Ahmad, H., Venugopal, K., Rajagopal, K., De Britto, S., Nandini, B., Pushpalatha, H. G., et al. (2020). Green synthesis and characterization of zinc oxide nanoparticles using eucalyptus globules and their fungicidal ability against pathogenic fungi of apple orchards. Biomolecules 10, 425–13. doi:10.3390/biom10030425
Aleklett, K., Ohlsson, P., Bengtsson, M., and Hammer, E. C. (2021). Fungal foraging behaviour and hyphal space exploration in micro-structured soil chips. ISME J. 15, 1782–1793. doi:10.1038/s41396-020-00886-7
Alengebawy, A., Abdelkhalek, S. T., Qureshi, S. R., and Wang, M. Q. (2021). Heavy metals and pesticides toxicity in agricultural soil and plants: ecological risks and human health implications. Toxics 9, 42–34. doi:10.3390/toxics9030042
Alonso González, P., Parga Dans, E., Acosta Dacal, A. C., Zumbado Peña, M., and Pérez Luzardo, O. (2022). Differences in the levels of sulphites and pesticide residues in soils and wines and under organic and conventional production methods. J. Food Compos. Anal. 112, 104714. doi:10.1016/j.jfca.2022.104714
Anjos, A., Fernandes, P., Marques, C., Borralho, N., Valente, C., Correia, O., et al. (2021). Management and fire, a critical combination for Eucalyptus globulus dispersal. For. Ecol. Manage. 490, 119086–119089. doi:10.1016/j.foreco.2021.119086
Ayed, A., Polito, F., Mighri, H., Souihi, M., Caputo, L., Hamrouni, L., et al. (2023). Eucalyptus species and their antifungal and herbicidal activities. Plants 12, 1–21. doi:10.3390/plants12173068
Beaumelle, L., Tison, L., Eisenhauer, N., Hines, J., Malladi, S., Pelosi, C., et al. (2023). Pesticide effects on soil fauna communities—A meta-analysis. J. Appl. Ecol. 60, 1239–1253. doi:10.1111/1365-2664.14437
Bi, K., Liang, Y., Mengiste, T., and Sharon, A. (2023). Killing softly: a roadmap of botrytis cinerea pathogenicity. Trends Plant Sci. 28, 211–222. doi:10.1016/j.tplants.2022.08.024
Bing-Ru, L., Guo-Mei, J., Jian, C., and Gangi, W. (2006). A review of methods for studying microbial diversity in soils. Pedosphere 16, 18–24. doi:10.1016/s1002-0160(06)60021-0
Bouhlali, E. D. T., Derouich, M., Ben-Amar, H., Meziani, R., and Essarioui, A. (2020). Exploring the potential of using bioactive plant products in the management of Fusarium oxysporum f.sp. albedinis: the causal agent of bayoud disease on date palm (Phoenix dactylifera L.). Beni-Suef Univ. J. Basic Appl. Sci. 9, 46. doi:10.1186/s43088-020-00071-x
Boulekbache-Makhlouf, L., Meudec, E., Mazauric, J. P., Madani, K., and Cheynier, V. (2013). Qualitative and semi-quantitative analysis of phenolics in Eucalyptus globulus leaves by high-performance liquid chromatography coupled with diode array detection and electrospray ionisation mass spectrometry. Phytochem. Anal. 24, 162–170. doi:10.1002/pca.2396
Buddhika, U. V. A., Savocchia, S., and Steel, C. C. (2021). Copper induces transcription of BcLCC2 laccase gene in phytopathogenic fungus, botrytis cinerea. Mycology 12, 48–57. doi:10.1080/21501203.2020.1725677
Câmara, J. S., Perestrelo, R., Ferreira, R., Berenguer, C. V., Pereira, J. A. M., and Castilho, P. C. (2024). Plant-derived terpenoids: a plethora of bioactive compounds with several health functions and industrial applications — a comprehensive overview. Molecules 29, 3861–35. doi:10.3390/molecules29163861
Cordeau, S., Triolet, M., Wayman, S., Steinberg, C., and Guillemin, J. P. (2016). Bioherbicides: dead in the water? A review of the existing products for integrated weed management. Crop Prot. 87, 44–49. doi:10.1016/j.cropro.2016.04.016
Diánez, F., Santos, M., Parra, C., Navarro, M. J., Blanco, R., and Gea, F. J. (2018). Screening of antifungal activity of 12 essential oils against eight pathogenic fungi of vegetables and mushroom. Lett. Appl. Microbiol. 67 (4), 400–410. doi:10.1111/lam.13053
Dias, M. C., Pinto, D. C. G. A., and Silva, A. M. S. (2021). Plant flavonoids: chemical characteristics and biological activity. Molecules 26, 5377–16. doi:10.3390/molecules26175377
DRAPN (2022). Circular no 06/2022. Estação de Avisos Agrícolas do Entre Douro e Minho. Produtos fitossanitáios para a vinha 2022.
Dudek-Wicher, R., Paleczny, J., Kowalska-Krochmal, B., Szymczyk-Ziółkowska, P., Pachura, N., Szumny, A., et al. (2021). Activity of liquid and volatile fractions of essential oils against biofilm formed by selected reference strains on polystyrene and hydroxyapatite surfaces. Pathogens 10, 515–522. doi:10.3390/pathogens10050515
Dwibedi, V., Rath, S. K., Jain, S., Martínez-Argueta, N., Prakash, R., Saxena, S., et al. (2023). Key insights into secondary metabolites from various chaetomium species. Appl. Microbiol. Biotechnol. 107, 1077–1093. doi:10.1007/s00253-023-12365-y
EFSA Álvarez, F., Arena, M., Auteri, D., Leite, S. B., Binaglia, M., et al. (2023). Peer review of the pesticide risk assessment of the active substance folpet. EFSA J. 21, e08139. doi:10.2903/j.efsa.2023.8139
European Commission (2018). “Implementing regulation (EU) 2018/1981 of 13 December 2018 renewing the approval of the active substances copper compounds, as candidates for substitution,” in Accordance with regulation (EC) no 1107/2009 of the European parliament and of the council conce (Official Journal of the European Union). Brussels, Belgium: European Commission.
Fialho, R. D. O., Papa, M. D. F. S., Panosso, A. R., and Cassiolato, A. M. R. (2017). Fungitoxicity of essential oils on Plasmopara viticola, causal agent of grapevine downy mildew. Rev. Bras. Frutic. 39, 1–14. doi:10.1590/0100-29452017015
Galindo, T. P. S., Pereira, R., Freitas, A. C., Santos-Rocha, T. A. P., Rasteiro, M. G., Antunes, F., et al. (2013). Toxicity of organic and inorganic nanoparticles to four species of white-rot fungi. Sci. Total Environ. 458–460, 290–297. doi:10.1016/j.scitotenv.2013.04.019
Geris, R., Teles de Jesus, V. E., Ferreira da Silva, A., and Malta, M. (2024). Exploring culture media diversity to produce fungal secondary metabolites and cyborg cells. Chem. Biodivers. 21, e202302066. doi:10.1002/cbdv.202302066
Global Biodiversity Information Facility (2014). Occurences of Chaetomium elatum kunze. Available online at: https://www.gbif.org/occurrence/3346418269 (Accessed 18 February, 25).
Goodell, B., Winandy, J. E., and Morrell, J. J. (2020). Fungal degradation of wood: emerging data, new insights and changing perceptions. Coatings 10, 1210–1219. doi:10.3390/coatings10121210
Gupta, I., Singh, R., Muthusamy, S., Sharma, M., Grewal, K., Singh, H. P., et al. (2023). Plant essential oils as biopesticides: applications, mechanisms, innovations, and constraints. Plants 12, 2916–2929. doi:10.3390/plants12162916
Hajji-Hedfi, L., Rhouma, A., Hlaoua, W., Dmitry, K. E., Jaouadi, R., Zaouali, Y., et al. (2024). Phytochemical characterization of forest leaves extracts and application to control apple postharvest diseases. Sci. Rep. 14, 2014–2018. doi:10.1038/s41598-024-52474-w
IUPAC (2024). Folpet. “In: Dictionary of Toxicology. Springer, Singapore. 389. doi:10.1007/978-981-99-9283-6_991
Janusz, G., Pawlik, A., Świderska-Burek, U., Polak, J., Sulej, J., Jarosz-Wilkołazka, A., et al. (2020). Laccase properties, physiological functions, and evolution. Int. J. Mol. Sci. 21, 1–25. doi:10.3390/ijms21030966
Jiang, L., Qiu, Y., Dumlao, M. C., Donald, W. A., Steel, C. C., and Schmidtke, L. M. (2023). Detection and prediction of Botrytis cinerea infection levels in wine grapes using volatile analysis. Food Chem. 421, 136120. doi:10.1016/j.foodchem.2023.136120
Koledenkova, K., Esmaeel, Q., Jacquard, C., Nowak, J., Clément, C., and Ait Barka, E. (2022). Plasmopara viticola the causal agent of downy mildew of grapevine: from its taxonomy to disease management. Front. Microbiol. 13, 889472–29. doi:10.3389/fmicb.2022.889472
Kong, C. H., Xuan, T. D., Khanh, T. D., Tran, H. D., and Trung, N. T. (2019). Allelochemicals and signaling chemicals in plants. Molecules 24, 2737–19. doi:10.3390/molecules24152737
Kumar, D., and Mukhopadhyay, R. (2024). Climate change and plant pathogens: understanding dynamics, risks and mitigation strategies. Plant Pathol. 74, 59–68. doi:10.1111/ppa.14033
Lamberth, C. (2019). “Synthesis and applications of cyclic imides in agrochemistry,” in Imides - Medicinal, agricultural, synthetic applications and natural products chemistry. Editor F. A. Luzzio (Elsevier Inc.), 335–352.
Machado, S., Pereira, R., Marie, R., and Sousa, O. F. (2023). Nanobiopesticides: are they the future of phytosanitary treatments in modern agriculture? Sci. Total Environ. 896, 166401–166416. doi:10.1016/j.scitotenv.2023.166401
Macías, F. A., Mejías, F. J. R., and Molinillo, J. M. G. (2019). Recent advances in allelopathy for weed control: from knowledge to applications. Pest Manag. Sci. 75, 2413–2436. doi:10.1002/ps.5355
Malandrakis, A. A., Kavroulakis, N., and Chrysikopoulos, C. V. (2020). Synergy between Cu-NPs and fungicides against Botrytis cinerea. Sci. Total Environ. 703, 135557–11. doi:10.1016/j.scitotenv.2019.135557
Martin, K. J., and Rygiewicz, P. T. (2005). Fungal-specific PCR primers developed for analysis of the ITS region of environmental DNA extracts. BMC Microbiol. 5, 28–11. doi:10.1186/1471-2180-5-28
Matan, N., Woraprayote, W., Saengkrajang, W., Sirisombat, N., and Matan, N. (2009). Durability of rubberwood (Hevea brasiliensis) treated with peppermint oil, eucalyptus oil, and their main components. Int. Biodeterior. Biodegr. 63, 621–625. doi:10.1016/j.ibiod.2008.12.008
McGinley, J., Healy, M. G., Ryan, P. C., O’Driscoll, H., Mellander, P. E., Morrison, L., et al. (2023). Impact of historical legacy pesticides on achieving legislative goals in Europe. Sci. Total Environ. 873, 162312–162313. doi:10.1016/j.scitotenv.2023.162312
Money, N. P. (2007). “Biomechanics of invasive hyphal growth,” in The mycota - a comprehensive treatise on fungi as experimental systems for basic and applied research. Editors R. J. Howard, and N. A. R. Gow (Berlin Heidelberg: Springer).
Morin, S., and Artigas, J. (2023). Twenty years of research in ecosystem functions in aquatic microbial ecotoxicology. Environ. Toxicol. Chem. 42, 1867–1888. doi:10.1002/etc.5708
Motmainna, M., Juraimi, A. S., Ahmad-Hamdani, M. S., Hasan, M., Yeasmin, S., Anwar, M. P., et al. (2023). Allelopathic potential of tropical plants—A review. Agronomy 13, 2063–21. doi:10.3390/agronomy13082063
National Center for Biotechnology Information (2024). PubChem compound summary for cid 8607, folpet. Available online at: https://pubchem.ncbi.nlm.nih.gov/compound/Folpet (Accessed 29 October, 24).
Osma, J. F., Moilanen, U., Toca-Herrera, J. L., and Rodríguez-Couto, S. (2011). Morphology and laccase production of white-rot fungi grown on wheat bran flakes under semi-solid-state fermentation conditions. FEMS Microbiol. Lett. 318, 27–34. doi:10.1111/j.1574-6968.2011.02234.x
Oulkar, D. P., Shinde, R., Khan, Z., Organtini, K., Leonard, S., and Banerjee, K. (2019). Improved analysis of captan, tetrahydrophthalimide, captafol, folpet, phthalimide, and iprodione in fruits and vegetables by liquid chromatography tandem mass spectrometry. Food Chem. 301, 125216–125217. doi:10.1016/j.foodchem.2019.125216
Parada, J., Tortella, G., Seabra, A. B., Fincheira, P., and Rubilar, O. (2024). Potential antifungal effect of copper oxide nanoparticles combined with fungicides against botrytis cinerea and Fusarium oxysporum. Antibiotics 13, 215–11. doi:10.3390/antibiotics13030215
Pedrotti, C., Marcon, Â. R., Delamare, A. P. L., Echeverrigaray, S., da Silva Ribeiro, R. T., and Schwambach, J. (2019). Alternative control of grape rots by essential oils of two eucalyptus species. J. Sci. Food Agric. 99, 6552–6561. doi:10.1002/jsfa.9936
Pingali, P. L. (2012). Green revolution: impacts, limits, andthe path ahead. Proc. Natl. Acad. Sci. U. S. A. 109, 12302–12308. doi:10.1073/pnas.0912953109
Pinto, M., Soares, C., Martins, M., Sousa, B., Valente, I., Pereira, R., et al. (2021). Herbicidal effects and cellular targets of aqueous extracts from young eucalyptus globulus labill. Leaves. Plants 10, 1–18. doi:10.3390/plants10061159
Pombal, S., Rodilla, J., Gomes, A., Silva, L., and Rocha, P. (2014). Evaluation of the antibacterial activity of the essential oil and antioxidant activity of aqueous extracts of the Eucalyptus globulus L. leaves. Glob. Adv. Res. J. Agric. Sci. 3, 356–366.
Queirós, L., Deus, E., Silva, J. S., Vicente, J., Ortiz, L., Fernandes, P. M., et al. (2020). Assessing the drivers and the recruitment potential of Eucalyptus globulus in the iberian peninsula. For. Ecol. Manage. 466, 1–11. doi:10.1016/j.foreco.2020.118147
Rienth, M., Crovadore, J., Ghaffari, S., and Lefort, F. (2019). Oregano essential oil vapour prevents Plasmopara viticola infection in grapevine (Vitis Vinifera) and primes plant immunity mechanisms. PLoS One 14, e0222854–29. doi:10.1371/journal.pone.0222854
Ritz, C., Baty, F., Streibig, J. C., and Gerhard, D. (2015). Dose-response analysis using R. PLoS One 10, e0146021–13. doi:10.1371/journal.pone.0146021
Rodríguez-Ramos, F., Briones-Labarca, V., Plaza, V., and Castillo, L. (2023). Iron and copper on botrytis cinerea: new inputs in the cellular characterization of their inhibitory effect. PeerJ 11, 1–30. doi:10.7717/peerj.15994
Rosenbaum, R. K., Bachmann, T. M., Gold, L. S., Huijbregts, M. A. J., Jolliet, O., Juraske, R., et al. (2008). USEtox - the UNEP-SETAC toxicity model: recommended characterisation factors for human toxicity and freshwater ecotoxicity in life cycle impact assessment. Int. J. Life Cycle Assess. 13, 532–546. doi:10.1007/s11367-008-0038-4
Slomczynski, D., Nakas, J. P., and Tanenbaum, S. W. (1995). Production and characterization of laccase from botrytis cinerea 61-34. Appl. Environ. Microbiol. 61, 907–912. doi:10.1128/aem.61.3.907-912.1995
Sydor, M., Cofta, G., Doczekalska, B., and Bonenberg, A. (2022). Fungi in mycelium-based composites: usage and recommendations. Mater. (Basel) 15, 6283–34. doi:10.3390/ma15186283
Tamm, L., Thuerig, B., Apostolov, S., Blogg, H., Borgo, E., Corneo, P. E., et al. (2022). Use of copper-based fungicides in organic agriculture in twelve European countries. Agronomy 12, 673. doi:10.3390/agronomy12030673
Tomazoni, E. Z., Pauletti, G. F., da Silva Ribeiro, R. T., Moura, S., and Schwambach, J. (2017). In vitro and in vivo activity of essential oils extracted from Eucalyptus staigeriana, Eucalyptus globulus and Cinnamomum camphora against Alternaria solani sorauer causing early blight in tomato. Sci. Hortic. Amst. 223, 72–77. doi:10.1016/j.scienta.2017.04.033
Tudi, M., Ruan, H. D., Wang, L., Lyu, J., Sadler, R., Connell, D., et al. (2021). Agriculture development, pesticide application and its impact on the environment. Int. J. Environ. Res. Public Health 18, 1–23. doi:10.3390/ijerph18031112
Tura, D., Wasser, S. P., and Zmitrovich, I. V. (2016). “Wood-inhabiting fungi: applied aspects,” in Fungi: applications and management strategies (Florida, United States: Taylor & Francis Group, LLC), 1–48.
Ullah, I., Yuan, W., Khalil, H. B., Khan, M. R., Lak, F., Uzair, M., et al. (2024). Understanding botrytis Cinerea infection and gray mold management: a review paper on deciphering the rose ’ s thorn. Phytopathol. Res. 6, 42–17. doi:10.1186/s42483-024-00262-9
Vallejo, A., Millán, L., Abrego, Z., Sampedro, M. C., Sánchez-Ortega, A., Unceta, N., et al. (2019). Fungicide distribution in vitiviniculture ecosystems according to different application strategies to reduce environmental impact. Sci. Total Environ. 687, 319–329. doi:10.1016/j.scitotenv.2019.06.112
Vázquez-Rowe, I., Villanueva-Rey, P., Moreira, M. T., and Feijoo, G. (2012). Environmental analysis of ribeiro wine from a timeline perspective: harvest year matters when reporting environmental impacts. J. Environ. Manage. 98, 73–83. doi:10.1016/j.jenvman.2011.12.009
Venâncio, C., Pereira, R., Freitas, A. C., Rocha-Santos, T. A. P., da Costa, J. P., Duarte, A. C., et al. (2017). Salinity induced effects on the growth rates and mycelia composition of basidiomycete and zygomycete fungi. Environ. Pollut. 231, 1633–1641. doi:10.1016/j.envpol.2017.09.075
Vigneron, N., Grimplet, J., Remolif, E., and Rienth, M. (2023). Unravelling molecular mechanisms involved in resistance priming against downy mildew (Plasmopara viticola) in grapevine (Vitis vinifera L.). Sci. Rep. 13, 14664–13. doi:10.1038/s41598-023-41981-x
Violi, H. A., Menge, J. A., and Beaver, R. J. (2007). Chaetomium elatum (Kunze: chaetomiaceae) as a root-colonizing fungus in avocado: is it a mutualist, cheater, commensalistic associate, or pathogen? Am. J. Bot. 94, 690–700. doi:10.3732/ajb.94.4.690
Vurro, M., Miguel-Rojas, C., and Pérez-de-Luque, A. (2019). Safe nanotechnologies for increasing the effectiveness of environmentally friendly natural agrochemicals. Pest Manag. Sci. 75, 2403–2412. doi:10.1002/ps.5348
Wang, X. W., Lombard, L., Groenewald, J. Z., Li, J., Videira, S. I. R., Samson, R. A., et al. (2016). Phylogenetic reassessment of the Chaetomium globosum species complex. Persoonia Mol. Phylogeny Evol. Fungi 36, 83–133. doi:10.3767/003158516X689657
Wightwick, A. M., Salzman, S. A., Reichman, S. M., Allinson, G., and Menzies, N. W. (2013). Effects of copper fungicide residues on the microbial function of vineyard soils. Environ. Sci. Pollut. Res. 20, 1574–1585. doi:10.1007/s11356-012-1114-7
Wilcox, W. F., Gubler, W. D., and Uyemoto, J. K. (2015). Diseases of grape (Vitis vinifera L.). Available online at: https://www.apsnet.org/edcenter/resources/commonnames/Pages/Grape.aspx (Accessed 3 March, 25).
Wood, B., Mcbride, E., Nabetani, K., Grif, T., and Sabaratnam, S. (2023). Prevalence and spatial distribution of cranberry fruit rot pathogens in British Columbia, Canada and potential fungicides for fruit rot management. Front. Plant Sci. 14, 1274094–17. doi:10.3389/fpls.2023.1274094
Yao, S., Xu, J., Zhang, H., Gao, H., Shang, S., and Wang, D. (2021). in vitro activity of extracts of five medicinal plant species on plant pathogenic fungi. Front. Agric. Sci. Eng. 8, 635–644. doi:10.15302/J-FASE-2020343
Yong, W. T. L., Ades, P. K., Goodger, J. Q. D., Bossinger, G., Runa, F. A., Sandhu, K. S., et al. (2019). Using essential oil composition to discriminate between myrtle rust phenotypes in Eucalyptus globulus and Eucalyptus obliqua. Ind. Crops Prod. 140, 111595–111597. doi:10.1016/j.indcrop.2019.111595
Zhao, L., Feng, C., Wu, K., Chen, W., Chen, Y., Hao, X., et al. (2017). Advances and prospects in biogenic substances against plant virus: a review. Pestic. Biochem. Physiol. 135, 15–26. doi:10.1016/j.pestbp.2016.07.003
Keywords: ascomycota, basidiomycota, bordeaux mixture, folpet, growth inhibition assays, plant-based products
Citation: Fernandes B, Duarte R, Paiva C, Oliveira-Pacheco J, Cachada A and Pereira R (2025) Evaluation of alternatives to replace or reduce viticulture conventional fungicides: Eucalyptus globulus essential oil and aqueous extract. Front. Environ. Sci. 13:1675469. doi: 10.3389/fenvs.2025.1675469
Received: 30 July 2025; Accepted: 02 October 2025;
Published: 14 October 2025.
Edited by:
Olukayode Jegede, University of California, Davis, CA, United StatesReviewed by:
Feng Zhou, Henan Institute of Science and Technology, ChinaSivagnanapazham Kathiresan, Tamil Nadu Agricultural University, India
Copyright © 2025 Fernandes, Duarte, Paiva, Oliveira-Pacheco, Cachada and Pereira. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Beatriz Fernandes, dXAyMDE1MDMzNTJAZWR1LmZjLnVwLnB0