Abstract
Herbivore browsing can influence the structure, composition, and processes of an ecosystem, and its effects can cascade to other functional components, especially when coupled with other disturbances. Ungulate browsing is not random, as it can be influenced by various biophysical factors. This study assessed overall herbivore browse utilisation in the Kazuma Forest Reserve (KFR) in the Chobe District of Northern Botswana at two scales of aggregation: vegetation class and study area. Vegetation of the KFR can be divided into three broad classes: shrubland vegetation, grassland vegetation, and woodland vegetation from which 23 sampling plots were selected. In each sampling plot, browsed and unbrowsed terminal shoots of all woody species were counted, from which browse utilisation and selection were determined. In total, 46 woody species were recorded in the KFR. The Sorenson Similarity Index reveals a weak overlap in species composition across the three vegetation classes of the KFR. Overall proportionate browse utilisation by the herbivore community of the KFR was influenced by vegetation type, whereas selection was not. Utilisation of most woody species was low, and few species were preferentially browsed by the KFR herbivore community. Both woody species utilisation and selection were not consistent across the two scales of aggregation. Woodland vegetation, on nutrient-poor Kalahari Sands, was associated with the least and highest proportion of browsed and unbrowsed woody species, respectively. However, even low browsing pressure can effect notable changes in ecosystem structure, composition, and hence function. The results of this study did not find any effect of predation risk on browse utilisation or selection across the three vegetation classes of the KFR. In conclusion, this study has laid a benchmark for future monitoring of browsing pressure and changes in wood species community structure in the KFR. It recommends further research to assess the effect of environmental and biological factors on browse utilisation and selection in the KFR.
1 Introduction
The structure of ecological systems is shaped by extant biophysical components and their interactions, and among such is herbivory. Large herbivores are vectors of important processes that can influence the structure, composition, and function of an ecosystem (Frerker et al., 2013; Sankaran et al., 2013; Pekin et al., 2014; Skarpe et al., 2014; Möst et al., 2015; Machar et al., 2018; Bödeker et al., 2021). Coupled with other factors, e.g., disturbances, herbivory can promote feedback loops (Makhabu and Skarpe, 2006; Hasler and Senn, 2012; Möst et al., 2015) and exert top-down controls in semi-arid savannas (Sankaran et al., 2013). In the absence of disturbances, browsing pressure can regulate plant diversity by reducing and regulating the abundance of dominant species and excluding lesser competitors (Pekin et al., 2014) and can drive short- and long-term changes in land cover (Pekin et al., 2014; Fox et al., 2017; Sianga et al., 2017). Therefore, as a function of its intensity, herbivory can cause changes in the relative biomass and abundance of plant species (Owen-Smith and Chafota, 2012; Sankaran et al., 2013; Pekin et al., 2014; Möst et al., 2015; Machar et al., 2018).
Ultimately, herbivory can induce changes in ecosystem processes (Frerker et al., 2013; Möst et al., 2015) and successional pathways (Frerker et al., 2013; de Boer et al., 2015; Möst et al., 2015). Its effects can cascade to other functional components of an ecosystem through the competitive release of understory species (de Boer et al., 2015), its influence on predation risk (Valeix et al., 2012), and shifts in grazer–browser ratios (Rutina et al., 2005; Makhabu et al., 2006; de Boer et al., 2015) and can promote the expansion of invasive species (Machar et al., 2018). It is therefore one of the key drivers of the structure, composition, and function of savanna ecosystems (Mosugelo et al., 2002; Frerker et al., 2013; Sankaran et al., 2013; Pekin et al., 2014; Skarpe et al., 2014; Möst et al., 2015; Machar et al., 2018). Consequently, browsing pressure could have an impact on the integrity of ecosystems (Möst et al., 2015) and hence their ability to deliver ecosystem goods and services (Fox et al., 2017; Mojeremane et al., 2021).
All herbivores can be selective in their food habits (Owen-Smith and Chafota, 2012). Herbivore browsing is influenced by multiple biophysical factors: local ecological conditions (Frerker et al., 2013), top-down regulation of browsers (Valeix et al., 2012; Frerker et al., 2013), physicochemical protection mechanisms and palatability of plants (Makhabu and Skarpe, 2006; Möst et al., 2015; Hejcmanová et al., 2019), community structure and composition (Owen-Smith and Chafota, 2012; Frerker et al., 2013), availability and proximity to surface water, edaphic environment, and soil nutrients (Owen-Smith and Chafota, 2012), and anthropogenic influences (Fox et al., 2017). Climate change will compound the influence of most of these factors (Fox et al., 2017). Therefore, herbivore browsing and its effect on local ecology will vary at multiple scales consistent with the variability of its underlying drivers and their interactions.
Northern Botswana possesses extensive woodlands dominated by Zambezi teak Baikiaea plurijuga and mopane Colophospermum mopane (Fox et al., 2017). However, the conversion of woodland canopy forest to open savanna has been observed, as in some other high wildlife density areas of northern Botswana; Moremi Game Reserve and the Okavango Delta (Ben-Shahar, 1996); and Chobe National Park (Mosugelo et al., 2002; Rutina et al., 2005; Makhabu et al., 2006; Fox et al., 2017). The Kazuma Forest Reserve (KFR) has realised a 65% conversion from canopy forest into open savanna, and shrubland had become the dominant land cover by 2013 (Fox et al., 2017). The observed vegetation changes in northern Botswana are mainly ascribed to the felling of mature canopy trees by the African elephant (Loxodonta africana) and intensive seedling predation by medium-sized ungulates (Ben-Shahar, 1993; Mosugelo et al., 2002; Rutina et al., 2005; Makhabu et al., 2006; Skarpe et al., 2014). However, the large landscape land cover change analysis by Fox et al. (2017) was not conclusive on the relationship between elephant density and woodland loss. This suggests that the observed changes in the Chobe District and KFR were compounded by other determining factors, e.g., fire (Fox et al., 2017) and possibly herbivory.
The primary purpose of protected areas is to secure the conservation of natural ecological assemblages and processes (Möst et al., 2015). Understanding the effect of browsing pressure on the vegetation community is necessary to guide the management of forests and accompanying biodiversity (Sankaran et al., 2013; Pekin et al., 2014). Given its potential impact on vegetation, browsing pressure is of immediate concern to forest management, and hence, its monitoring and consequent changes in community structure are critical (Frerker et al., 2013; Bödeker et al., 2021). Regular and routine browsing inventories serve as a monitoring and planning tool for adaptive management (Bödeker et al., 2021). Although there has been some research on herbivory in the Chobe District, most (e.g., Rutina et al., 2005; Makhabu and Skarpe, 2006; Makhabu et al., 2006; Owen-Smith and Chafota, 2012) focussed on a single herbivore and specifically the African elephant. Based on the above, this study aimed to determine overall herbivore browsing pressure in the KFR. Specific objectives were to: (a) determine browse utilisation of extant woody species, (b) assess browse selection or avoidance of extant species, and (c) assess the influence of vegetation type on browse utilisation and selection across the three different vegetation types of the KFR. Furthermore, consistent with Frerker et al. (2013) and Mosbacher and Williams (2009), the results of this study will serve as a baseline for future monitoring of both browsing pressure and its potential effect on the KFR vegetation community. This study is not aware of any previous research that assessed overall browsing pressure that can also act as a benchmark for future browse monitoring in the KFR or the Chobe District.
2 Materials and methods
2.1 Ethical note
As required by the Laws of Botswana, this study was authorised by the Government of Botswana [Permit No. ENT 8/36/4 XXXX (29)]. It was approved and funded by the Research Technology Development and Transfer Committee of the Botswana University of Agriculture and Natural Resources. The Department of Wildlife and National Parks provided security for the research team from dangerous animals during field data collection. No trees or wildlife species were removed nor destroyed during field data collection.
2.2 Study area
The Kazuma Forest Reserve is approximately 194 km2 (Fox et al., 2017), the second smallest and one of the six protected areas dedicated to forest conservation in Botswana. Protected areas, i.e., forest reserves (FRs) and Chobe National Park (CNP), constitute approximately 58% of the total land area of the Chobe District (Fox et al., 2017). The KFR abuts Kazuma Pan National Park of Zimbabwe (Figure 1). Consequently, it provides ecological connectivity between the two major conservation landscapes of Chobe (Botswana) and Hwange–Matetsi (Zimbabwe). All six formally declared FRs of Botswana are in the Chobe District and are devoid of human settlements. However, where they are near settlements, as it is the case with KFR, livestock grazing is permitted during the day. Its southern boundary is marked by the wildlife-proof fenced Pandamatenga Commercial Arable Farms, whereas the western boundary is shared with Controlled Hunting Area No. 5, where there is a growing subsistence pastoral farming.
Figure 1
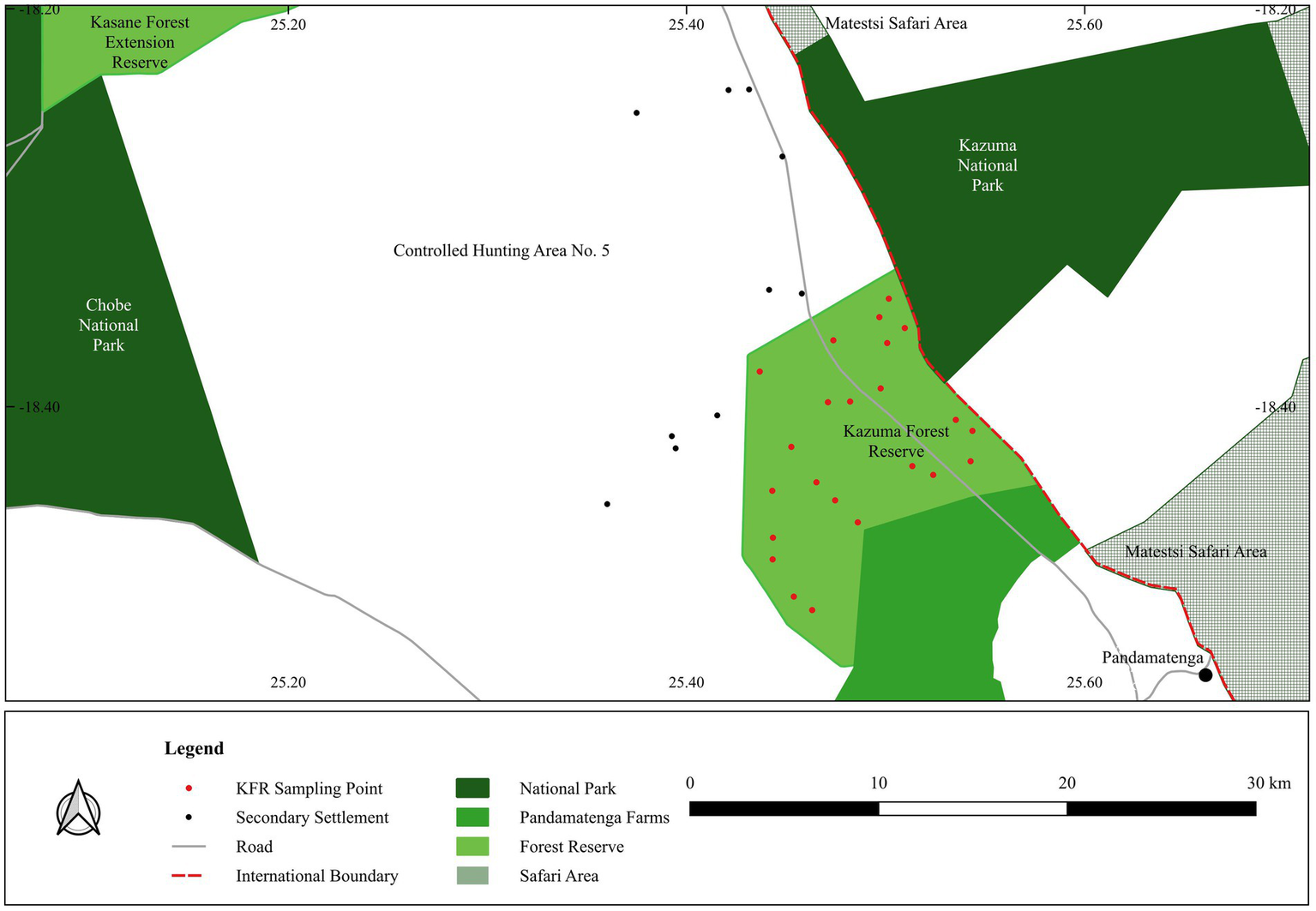
Map of the Kazuma Forest Reserve in relation to major land units in the republics of Botswana and Zimbabwe. Secondary settlements are also depicted (subsistence livestock farming settlements).
The climate of the area is described as semi-arid or subtropical, with distinct hot summer and cool winter seasons (Mmolotsi et al., 2012). Rainfall is highly variable, with nearly the entire annual rainfall budget concentrated during the November–March wet season, followed by a general absence of precipitation from April to October (Fox et al., 2017).
Three vegetation types can be discerned in the KFR. The Baikiaea plurijuga dominated woodland vegetation on nutrient-poor Kalahari Sands with low water retention capacity, open grassland in the northern fringes of the KFR associated with nutrient-rich black vertisols and hydrological features (i.e., natural water pans), and shrubland vegetation also on Kalahari Sands. Due to high humus content, vertisols are characterised by high nutrient content, high water retention capacity, and low percolation rates. The woodland vegetation is the tallest canopy vegetation dominated by Baikiaea plurijuga and other tall woody species. However, also on Kalahari Sands, the shrubland vegetation is characterised by woody shrub trees. These vegetation types, particularly the woodland and shrubland savanna, are not distinct but are rather a mosaic of fine-scale patches. High fire frequency and density are also characteristics of the KFR (Fox et al., 2017).
Typical of northern Botswana, high wildlife diversity is characteristic; mega-herbivores, e.g., African elephant and giraffe (Giraffa camelopardalis), large herbivores (e.g., Cape buffalo (Syncerus caffer), zebra (Equus quagga), eland (Tragelaphus oryx), and kudu (Tragelaphus strepsiceros)), medium-to-small ungulates (e.g., impala (Aepyceros melampus), steenbok (Raphicerus campestris), and duiker (Sylvicapra grimmia)), and large carnivores (e.g., African lion (Panthera leo melanochaita), spotted hyaena (Crocuta crocuta), and leopard (Panthera purdas)).
2.3 Data collection
In total, 23 20 m-by-20 m plots were selected in the representative vegetation types of the KFR. Attempts were made to randomise the distribution of the sampling plots. However, consideration for vehicular access and the safety of the research team against the presence of dangerous wildlife did influence the final distribution of sampling plots (Figure 1). Each sampling plot was georeferenced at its centre, measured and marked prior to sampling. At each sampling plot, the vegetation type was assigned based on physiognomic features. Within each sampling plot, all woody species were identified to the lowest taxonomic level. Data were collected during the wet season, November to December 2021.
Herbivore foraging decisions follow a sequence of place, plant species, individual plants, and finally a plant part (Hasler and Senn, 2012; Owen-Smith and Chafota, 2012). Therefore, herbivores’ browsing can be assessed at these different scales of aggregation (Owen-Smith and Chafota, 2012). Herbivores usually browse the tips of branches, leaving conspicuous signs (Hasler and Senn, 2012; Frerker et al., 2013). This study focussed on two scales of aggregation; terminal branch ends as browsable units (Mosbacher and Williams, 2009; Frerker et al., 2013; Bödeker et al., 2021) and vegetation type.
Within each sampling plot, all woody trees were inspected for browse or bite marks, and browsed and unbrowsed terminal shoots were counted (Mosbacher and Williams, 2009; Frerker et al., 2013; Pekin et al., 2014; Möst et al., 2015; Bödeker et al., 2021). Browsing height range of herbivores is species-specific (Owen-Smith and Chafota, 2012; Frerker et al., 2013; Hejcmanová et al., 2019; Bödeker et al., 2021). Therefore, to meaningfully assess browsing pressure, it is critical that sampling accommodates the reach of most extant herbivores (Bödeker et al., 2021). Elephant, the dominant browser of the KFR, browsing occurs mainly within the 1–2.5 m height range (Owen-Smith and Chafota, 2012); therefore, its upper browsing height was considered the maximum sampling height that accommodated most extant herbivores. Hejcmanová et al. (2019) also used 2.5 m as the upper sampling limit in their study of Western Derby eland (Tragelaphus derbianus) feeding behaviour in Senegal. Data were collected for individual trees and later aggregated by species, sampling plot, vegetation type, and study area. The total number of browsable units was estimated by the sum of browsed and unbrowsed branches.
2.4 Data analysis
Data were processed using the Microsoft Excel spreadsheet and summarised by plant species at the plot level and later aggregated by vegetation type and the study area. Generalised linear models with Poisson distribution and logarithmic link function were used to test for the possible influence of vegetation type on the frequency distribution of recorded species (i.e., species richness) and selection across the three vegetation types. Generalised linear models with a binomial distribution, logit link function, and the total number of browsable units as weights were used to test for the potential influence of vegetation type on proportionate browse utilisation. The Sorenson Similarity Index (Ss) was used for pairwise comparison of species composition across the three vegetation types of the KFR, where a is the number of species common to the two vegetation types while b and c are the number of species in the vegetation types being compared. The Ss varies from 0 to 1 for no overlap to perfect overlap in species composition, respectively, and values in the range of 0.51 to 0.75 were considered to indicate high similarities in species composition between the vegetation types being compared (Derebe and Girma, 2020).
The analysis of browse utilisation and selection was done at the vegetation type and aggregated at the study area level. Browse utilisation was based on the percentage of browsed terminal branches and separated into three ordinal browse utilisation categories: high (> 30%), moderate (15–30%), and low (< 15%) utilisation (Pekin et al., 2014).
The acceptance of a food type is the proportionate frequency at which it is used when encountered (Owen-Smith and Chafota, 2012). Similar to Hejcmanová et al. (2019), the Jacobs Index of Selectivity (Si) was used to test for browse selection at vegetation type and KFR scales, where ci is the relative proportion of browsed terminal branches of the ith woody species and ai is the relative proportion of available (total number) browsable units of the ith woody species. Jacobs Index ranges between −1 and 1, representing complete rejection or acceptance, respectively, of a browsed species. Woody species with Si < −0.33 and Si > 0.33 were considered avoided and selected, respectively, whereas those within this range were considered to have been accepted (not avoided nor selected). Species that were not browsed (Si = −1) were considered to have been rejected by the browser community of KFR.
3 Results
3.1 General results
In total, 46 woody species in 17 families were recorded in the 23 surveyed plots across the three vegetation types of the KFR. Recorded woody species were dominated by Family Fabaceae (n = 15, 32.61%) and Family Combretaceae (n = 9, 19.57%), while other families comprised at most two species (4.35%). One woody species recorded in the woodland vegetation was not definitively identified. The three vegetation types of the KRF were sampled adequately and in proportion to their availability in the reserve (χ2 = 2.70, df = 2, p = 0.26). Although the woodland and grassland vegetation types were associated with the highest and lowest species richness, respectively (Table 1), the number of recorded woody species in the grassland (χ2 = 0.18, df = 1, p = 0.67) and shrubland (χ2 = 1.38, df = 1, p = 0.24) vegetation types were not significantly different from that in the woodland vegetation and there was no data overdispersion (deviance–degree of freedom ratio = 0.96). The Sorenson Similarity Indices reveal a weak overlap in species composition between the three vegetation types of the KFR (Table 2), with the most overlap of 0.40 observed between the shrubland and woodland vegetation types.
Table 1
| Vegetation type | Number of sampling plots | Species richness (percentage) | ||
|---|---|---|---|---|
| Browsed | Not browsed | Total | ||
| Shrubland | 9 | 24 (88.89) | 3 (11.11) | 27 (100) |
| Grassland | 4 | 20 (83.33) | 4 (16.67) | 24 (100) |
| Woodland | 10 | 27 (81.82) | 6 (18.18) | 33 (100) |
| KFR (All) | 23 | 37 (80.43) | 9 (19.57) | 46 (100) |
Frequency and percentage (in brackets) distribution of sampling plots and species richness of browsed and unbrowsed woody species in the Kazuma Forest Reserve.
Table 2
| Paired vegetation types | Number of shared species | Sorenson’s Similarity Index |
|---|---|---|
| Grassland–shrubland | 15 | 0,37 |
| Grassland–woodland | 13 | 0,31 |
| Shrubland–woodland | 20 | 0,40 |
Comparison of woody species composition across the three vegetation types of the Kazuma Forest Reserve using the Sorenson Similarity Index.
3.2 Browse utilisation
Table 3 indicates that the majority of woody species aggregated by study area (71.74%) and at vegetation type (62.96–81.82%) were utilised at low intensity. Only two species, Vangueria infausta in the shrubland vegetation, and Commiphora glandulosa in the grassland vegetation and at the study area level, were browsed at high intensity (Figure 2). The woodland vegetation and shrubland vegetation had the highest (81.82%) and lowest (62.96%) proportions, respectively, of woody species browsed at low intensity (Table 3).
Table 3
| Browse utilization | KFR | Vegetation type | ||
|---|---|---|---|---|
| Shrubland | Grassland | Woodland | ||
| High (>30%) | 1 (2.17) | 1 (3.71) | 1 (4.17) | 0 (0.0) |
| Moderate (15–30%) | 12 (26.09) | 9 (33.33) | 6 (25.00) | 6 (18.18) |
| Low (<15%) | 33 (71.74) | 17 (62.96) | 17 (70.83) | 27 (81.82) |
| Total | 46 (100) | 27 (100) | 24 (100) | 33 (100) |
Frequency distribution and percentage of woody species by browse utilisation (percentage browsed terminal shoots) and vegetation type in the Kazuma Forest Reserve (KFR).
Figure 2
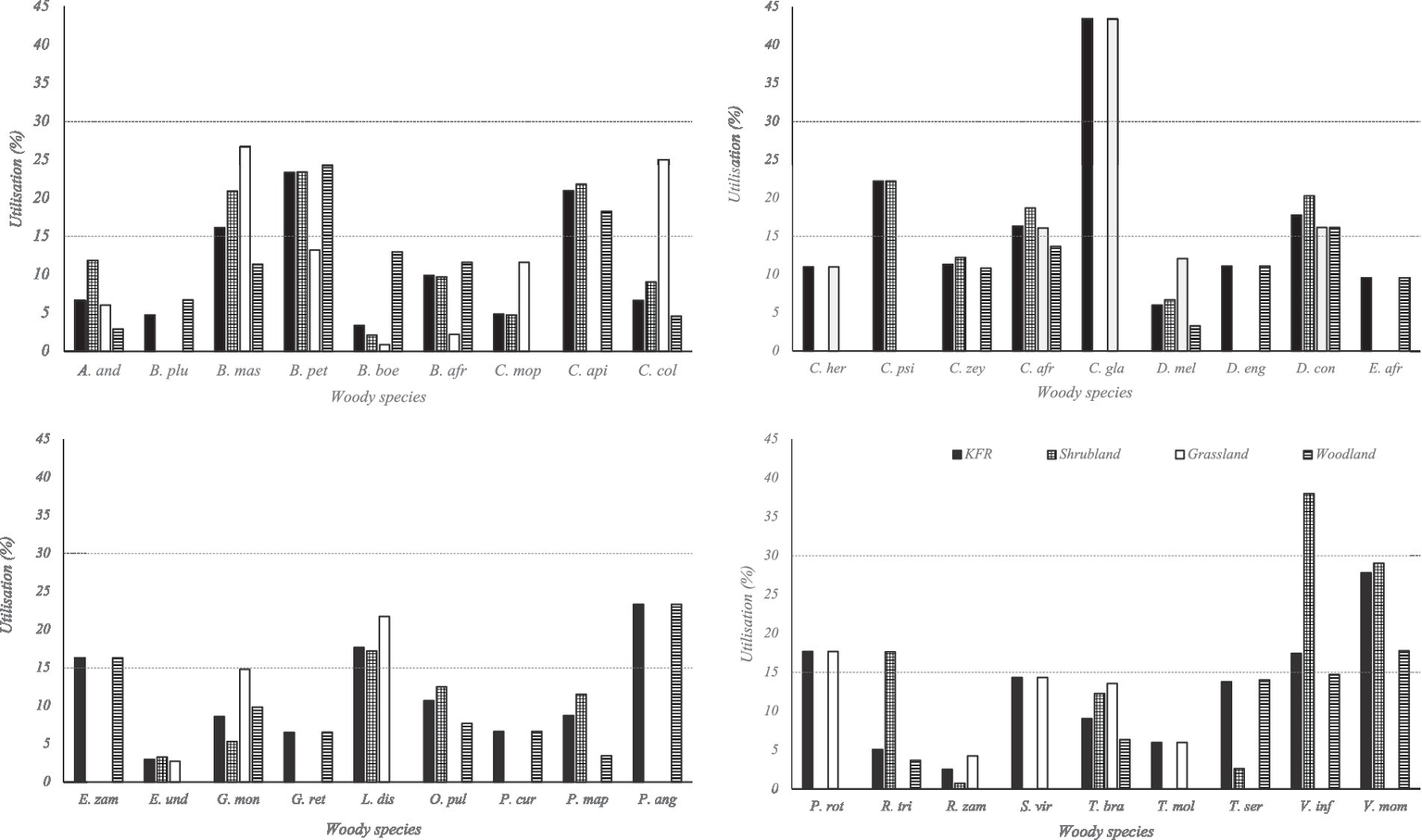
Graphical representation of woody species utilisation in the Kazuma Forest Reserve. The dotted horizontal lines indicate the 15 and 30% browse utilisation. A. and, Afzelia quanzensis; B. plu, Baikiaea plurijuga; B. mas, Baphia massaiensis; B. pet, Bauhinia petersiana; B. boe, Brachystegia boehmii; B. afr, Burkea africana; C. mop, Colophospermum mopane; C. api, Combretum apiculatum; C. col, Combretum collinum; C. her, Combretum hereroense; C. psi, Combretum psidioides; C. zey, Combretum zeyheri; C. afr, Commiphora africana; C. gla, Commiphora glandulosa; D. mel, Dalbergia melanoxylon; D. eng, Dialium englerianum; D. con, Diplorhynchus condylocarpon; E. afr, Erythrophleum africanum; E. zam, Erythroxylum zambesiacum; E. und, Euclea undulata; G. mon, Grewia monticola; G. ret, Grewia retinervis; L. dis, Lannea discolour; O. pul, Ochna pulchra; P. cur, Parinari curatellifolia; P. map, Pseudolachnostylis maprouneifolia; P. ang, Pterocarpus angolensis; P. rot, Pterocarpus rotundifolius; R. tri, Rhus trifoliate; R. zam, Rhizobium zambesiaca; S. vir, Securinega virosa; T. bra, Terminalia brachystemma; T. mol, Terminalia mollis; T. ser, Terminalia sericea; V. inf, Vangueria infausta; V. mom, Vitex mombassae.
Although species-specific browse utilisation varied with scales of aggregation, 19.57% (n = 9) of recorded woody species were not browsed (Afzelia quanzensis, Brachystegia spiciformis, Cassia abbreviata, Combretum imberbe, Crossopteryx febrifuga, Diospyros batocana, Hyphaene petersiana, Strychnos pungens, and Ximenia americana). Eight (n = 8, 17.39%) woody species (Amblygonocarpus andongensis, Baikiaea plurijuga, Brachystegia boehmii, Burkea africana, Dalbergia melanoxylon, Grewia monticola, Rhizobium zambesiaca, and Terminalia brachystemma) were consistently browsed at low intensity (< 15%) across all three vegetation types (Figure 2). Only one (n = 1, 2.17%) woody species (Diplorhynchus condylocarpon) was consistently used at high intensity (> 15%) across all three vegetation types of the KFR (Figure 2).
In addition, overall proportionate browse utilisation was significantly higher in the shrubland (β = 0.43, standard error = 0.06, p < 0.05) and woodland (β = 0.20, standard error = 0.05, p < 0.05) vegetation types than the grassland vegetation, with the odds increasing by 53.08 and 21.98%, respectively.
3.3 Browse selection
The frequency distribution of woody species by selection (i.e., number of species selected, accepted, avoided, and rejected) was not influenced by vegetation type (Poisson regression all p values >0.38). Table 4 indicates that the majority (n = 12–20, 50.00–60.61.6%) of recorded woody species were accepted by the KFR browser community at the vegetation type level. At the highest scale of aggregation, 56.52% (n = 26) of woody species were accepted. Fewer (n ≤ 4, 6.06–16.67%) woody species were selected in the three vegetation types and for the entire study area. Assuming that woody species that were not browsed (i.e., rejected) were also actively avoided, 33.33–37.04% of recorded woody species were avoided by the KFR browser community. At the highest scale of aggregation (i.e., the study area), nine (n = 9, 19.57%) woody species were rejected by the browser community of the KFR: Afzelia quanzensis, Brachystegia spiciformis, Cassia abbreviata, Combretum imberbe, Crossopteryx febrifuga, Diospyros batocana, Hyphaene petersiana, Strychnos pungens, and Ximenia americana (Table 5). Three (n = 3) to six (n = 6) woody species were rejected in the vegetation types they occurred. With the exception of the grassland vegetation, selected species constituted at most 7.41% (n = 2) of the recorded woody species. Only three species (n = 3, 6.52%), Bauhinia petersiana, Commiphora glandulosa, and Vitex mombassae, were selected at the highest level of aggression.
Table 4
| Selection | KFR | Vegetation type | ||
|---|---|---|---|---|
| Shrubland | Grassland | Woodland | ||
| Rejected | 9 (19.57) | 3 (11.11) | 4 (16.67) | 6 (18.18) |
| Avoided | 8 (17.39) | 7 (25.93) | 4 (16.67) | 5 (15.15) |
| Accepted | 26 (56.52) | 15 (55.56) | 12 (50.00) | 20 (60.61) |
| Selected | 3 (6.52) | 2 (7.41) | 4 (16.67) | 2 (6.06) |
| Total | 46 (100) | 27 (100) | 24 (100) | 33 (100) |
Frequency distribution and percentage of the number of woody species by selection and vegetation type in the Kazuma Forest Reserve (KFR).
Table 5
| Woody species | Jacob’s Index of Selectivity (Si) | |||
|---|---|---|---|---|
| KFR | Shrubland | Grassland | Woodland | |
| Afzelia quanzensis | −1.00 | −1.00 | ||
| Amblygonocarpus andongensis | −0.31 | −0.10 | −0.25 | −0.61 |
| Baikiaea plurijuga | −0.46 | −1.00 | −1.00 | −0.28 |
| Baphia massaiensis | 0.15 | 0.23 | 0.48 | −0.02 |
| Bauhinia petersiana | 0.37 | 0.30 | 0.16 | 0.42 |
| Brachystegia boehmii | −0.58 | −0.74 | −0.84 | 0.05 |
| Brachystegia spiciformis | −1.00 | −1.00 | −1.00 | |
| Burkea africana | −0.12 | −0.19 | −0.63 | −0.01 |
| Cassia abbreviate | −1.00 | −1.00 | ||
| Colophospermum mopane | −0.45 | −0.55 | 0.08 | |
| Combretum apiculatum | 0.26 | 0.21 | 0.22 | |
| Combretum collinum | −0.31 | −0.23 | 0.44 | −0.44 |
| Combretum hereroense | −0.06 | 0.06 | ||
| Combretum imberbe* | −1.00 | −1.00 | ||
| Combretum psidioides | 0.28 | 0.22 | ||
| Combretum zeyheri | −0.05 | −0.08 | −0.04 | |
| Commiphora africana* | 0.14 | 0.14 | 0.25 | 0.08 |
| Commiphora glandulosa* | 0.55 | 0.65 | ||
| Crossopteryx febrifuga* | −1.00 | −1.00 | ||
| Dalbergia melanoxylon* | −0.36 | −0.37 | 0.10 | −0.57 |
| Dialium englerianum | −0.06 | −0.03 | ||
| Diospyros batocana | −1.00 | −1.00 | ||
| Diplorhynchus condylocarpon* | 0.20 | 0.19 | 0.29 | 0.17 |
| Erythrophleum africanum | −0.13 | −0.10 | ||
| Erythroxylum zambesiacum | 0.13 | 0.16 | ||
| Euclea undulata | −0.62 | −0.63 | −0.57 | |
| Grewia monticola* | −0.19 | −0.47 | 0.20 | −0.09 |
| Grewia retinervis | −0.31 | −0.29 | ||
| Hyphaene petersiana* | −1.00 | −1.00 | ||
| Lannea discolor | 0.17 | 0.09 | 0.38 | |
| Ochna pulchra | −0.08 | −0.07 | −0.21 | |
| Parinari curatellifolia | −0.32 | −0.31 | ||
| Pseudolachnostylis maprouneifolia | −0.18 | −0.11 | −0.55 | |
| Pterocarpus angolensis | 0.30 | 0.34 | ||
| Pterocarpus rotundifolius* | 0.17 | 0.31 | ||
| Rhus trifoliate | −0.43 | 0.10 | −0.53 | |
| Rhizobium zambesiaca | −0.68 | −0.91 | −0.46 | −1.00 |
| Securinega virosa | 0.07 | 0.20 | ||
| Strychnos pungens* | −1.00 | −1.00 | ||
| Terminalia brachystemma | −0.16 | −0.08 | 0.16 | −0.30 |
| Terminalia mollis | −0.35 | −0.25 | ||
| Terminalia sericea | 0.05 | −0.69 | 0.10 | |
| Vangueria infausta | 0.17 | 0.45 | 0.12 | |
| Vitex mombassae | 0.39 | 0.35 | 0.20 | |
| Ximenia americana* | −1.00 | −1.00 | ||
| Unidentified | 0.04 | 0.07 | ||
Jacobs index of selectivity by woody species and vegetation type and aggregated at study area level in the Kazuma Forest Reserve.
Selectivity indices of rejected (Si = −1), avoided (Si < −0.33), and selected (Si > 0.33) are formatted in bold. Asterisks (*) denote spinescent woody species.
None of the woody species was consistently selected across all three vegetation types of the KFR or where they occurred in multiple vegetation types. Only one species (Rhizobium zambesiaca) was consistently rejected or avoided across all three vegetation types of the KFR. Brachystegia spiciformis and Euclea undulata were consistently rejected and avoided, respectively, in the two vegetation types they were recorded. Three species (Commiphora africana, Diplorhynchus condylocarpon, and Terminalia brachystemma) were consistently accepted across all three vegetation types of the KFR. Another three (Combretum apiculatum, Combretum zeyheri, and Ochna pulchra) were also consistently accepted, where they were recorded in more than one vegetation type. In some instances, browse selection varied between vegetation types and scales of aggregation. For example, Vangueria infausta was preferentially browsed only in the shrubland vegetation and Grewia monticola was avoided only in the same vegetation but browsed in proportion to its availability in the other vegetation types. Combretum collinum was selected and avoided in the grasslands and woodland, respectively, and accepted in the shrubland vegetation and at the highest scale of aggregation. In the exception of the woodland vegetation where it is dominant, Baikiaea plurijuga was rejected by the KFR browser in the other two vegetation types.
4 Discussion
The number of woody species (n = 46) recorded by this study was higher than approximately 27 species recorded in another area of the Chobe District (Owen-Smith and Chafota, 2012), located near and north of KFR. In the Chobe Riverfront, another area close to KFR, species richness was inversely related to nutrient quality (Aarrestad et al., 2014). Although not significantly different, woody species richness in the KFR was similarly highest and lowest in the nutrient-poor woodland vegetation and nutrient-rich grassland vegetation, respectively. Furthermore, although species richness was even, the Sorenson Similarity Index reveal notable dissimilarity (all Ss < 0.50) in woody species composition between the three vegetation types of the KFR, with only 28.26% (n = 13) of the recorded woody species being common to all vegetation types.
Herbivore browsing depends on their inherent preferences, which can be influenced by biophysical factors (Möst et al., 2015). Through their influence on vegetation, soils are the most important controller in the Chobe ecosystem (Skarpe et al., 2014). The grassland vegetation is associated with nutrient-rich vertisols with high water retention capacity, whereas the other two are on nutrient-poor Kalahari Sands with high percolation rates. This difference in the underlying substrate (i.e., nutrients and moisture) is likely the primary driver of the observed difference in woody species composition between the three vegetation types. In addition, although on the same substrate, the difference in physiognomy between the woodland (tall canopy stand) and shrubland (open short savanna) vegetation types could be the factor that differentiated species composition between the two vegetation types. Specific to the KFR (Fox et al., 2017) and other parts of the Chobe District (Neo, 2001; Mosugelo et al., 2002; Rutina et al., 2005; Makhabu et al., 2006), this difference in physiognomy is attributed to intensive browsing by elephants, seedling predation by smaller ungulates and fire. However, it should be noted that woody species richness was not significantly influenced by or different across the three vegetation types of the KFR.
The observed dissimilarity in woody species composition and the underlying substrate suggests that utilisation and selection of woody species by the KFR herbivore community is not likely to be consistent across the three vegetation types. The browser community of the KFR rejected (Table 1) 11–19.57% (n = 3–9) of extant woody species at all scales of aggregation, much comparable to six woody species rejected by elephants in another study site in the Chobe District (Owen-Smith and Chafota, 2012) even though this study covered the entire browser community of the KFR.
Forage quality, among others, is controlled by the underlying soil substrates (Skarpe et al., 2014; Derebe and Girma, 2020). Therefore, relative to the nutrient-rich grassland vegetation, proportionate browse utilisation per plot was significantly higher in the woodland and shrubland vegetation types, both on nutrient-poor Kalahari Sands with low water retention capacity. This is consistent with Derebe and Girma (2020), who reported species richness in the Bohor reedbuck (Redunca redunca) diet to be inversely related to the quality of available forage. In addition, consistent with Mosbacher and Williams (2009), browse utilisation was significantly lower in the grassland vegetation, where species richness was the lowest. Collectively, these suggest that where nutritive value is low, herbivores increase the quantity of browse intake (use) and diversity of species consumed to optimise nutrient intake. However, when browse selection is considered, the effect of species richness on browsing is not evident as the proportion of selected woody species was highest and lowest in the grassland and woodland vegetation types, where species richness was lowest and highest, respectively. Moreover, when selected and accepted species in these two vegetation types are pooled, the combined proportion is comparable, ranging between 63 and 66.6%.
In addition, acceptance of browse may be inversely related to relative plant abundance or availability of alternative browse (Frerker et al., 2013). Results of this study are somewhat consistent with this assertion as the proportion of browsed woody species was lowest in the most species-rich woodland vegetation. In addition, results indicate that browse selection was not influenced by vegetation type. However, the differences in woody species composition between the three vegetation types of the KFR could be confounding. This suggests that where browsers are presented with fewer plant species, a higher proportion will be browsed. Acknowledging that feeding deterrents, such as thorns, can dissuade browsing (Ndlovu and Mundy, 2008; Hejcmanová et al., 2019; Wigley et al., 2019; Mramba, 2021), the majority (53.84%, n = 7) of spinescent woody species (e.g., Crossopteryx febrifuga, Hyphaene petersiana, Strychnos pungens, and Ximenia americana) were avoided or rejected by the browser community of the KFR. However, some spinescent species were browsed commensurate to their availability (e.g., Commiphora africana and Diplorhynchus condylocarpon) and even selected (Commiphora glandulosa) in some vegetation types. Dalbergia melanoxylon depicted a mixed pattern where it was accepted only in the grassland vegetation and avoided in others.
Some woody species of the KFR that were avoided and rejected by the browser community of the KFR include Brachystegia spiciformis, Euclea undulata and Rhizobium zambesiaca. In the northeastern Chobe, elephants avoided or rejected Baikiaea plurijuga and Ochna pulchra (Owen-Smith and Chafota, 2012). However, in the KFR, though rejected in the shrubland and grassland vegetation types, Baikiaea plurijuga was accepted in the woodland vegetation. This is consistent with the notion of wider browse diversity and increased browse intake to optimise required nutrients where nutritive value is low, i.e., the woodland vegetation is on nutrient-poor Kalahari Sands. In the KFR, Ochna pulchra was accepted in all the vegetation types it was recorded. Although Mramba (2021) found a comparable diversity of browsed tree species between two sites, in this study, woody species-specific utilisation and selection were not consistent across the three vegetation types and scales of aggregation. This is consistent with the additive effect of a wider browser community and differences in soil nutritive content.
Consistent with other studies in similar arid savanna environments (e.g., Owen-Smith and Chafota, 2012) and elsewhere (e.g., white-tailed deer Odocoileus virginianus in Allegheny, USA (Mosbacher and Williams, 2009)), utilisation of majority (62.96–81.82%) of woody species in the KFR was low (<15%) at all scales of aggregation. Goheen et al. (2007) also reported comparable and low (10–20%) browse utilisation in Laikipia District of Kenya. Consistent with Hejcmanová et al. (2019), the browser community of KFR selected very few woody species (< 16.5%) and avoided or rejected up to 37% of extant woody species at all scales of aggregation. This is comparatively lower than in another study site in the Chobe District, where elephants either neglected or rejected approximately 66% of woody species that were frequently encountered, including some very common species (Owen-Smith and Chafota, 2012). Once again, the lower proportion of pooled rejected and avoided woody species reinforces the additive effect of a wider browser community in the KFR.
Although browse utilisation was predominantly low and very few woody species were selected, the effect of browsing on ecosystem structure and processes in the KFR cannot be discounted. This is because due to species specificity of the effects of browsing, preferential browsing can induce directional shifts in woody species composition (Sankaran et al., 2013) and could eliminate favoured species (Owen-Smith and Chafota, 2012). In addition, even low browsing intensity can suppress dominant vegetation species (Sankaran et al., 2013; Pekin et al., 2014). According to recent population surveys by Chase et al. (2018), with the exception of the sable (Hippotragus niger) that decreased significantly, most herbivore populations of northern Botswana, including the African elephant, did not significantly change between the years 2014 and 2018. As the KFR is part of the northern Botswana wildlife range, it can be reasonably inferred that recent past browsing pressure could be comparable to the results of this study. Therefore, considering that notable vegetation changes and transition from the forest canopy to open savanna have already occurred in the KFR (Fox et al., 2017), results of this study are consistent with the notion that even at low browsing pressure shifts in vegetation structure are possible, especially where coupled with other disturbances (Pekin et al., 2014; Fox et al., 2017), e.g., fire that is known to regularly affect the KFR (Fox et al., 2017). However, acknowledging the difference in scale (i.e., area) and the characteristic high spatial heterogeneity of semi-arid African savannas, it can never be definitive that recent browser population trends and, hence, browsing pressure in the KFR were consistent with the large landscape.
Browse use and selection are directly related to palatability and the quality of browse (Hasler and Senn, 2012; Frerker et al., 2013; Skarpe et al., 2014) that are, in turn, a direct function of nutrient content (Skarpe et al., 2014; Derebe and Girma, 2020). Therefore, the physical environment can influence browsing pressure as mediated by underlying soil fertility and other soil properties (Skarpe et al., 2014; Derebe and Girma, 2020). Nutrient-poor sandy soils tend to be dominated by nutrient-poor vegetation (Owen-Smith and Chafota, 2012; Skarpe et al., 2014; Derebe and Girma, 2020). Consistent with the underlying soils’ nutrient effect on browse use, in the KFR overall proportionate browse use was significantly higher in vegetation types on nutrient-poor Kalahari Sands, which is consistent with an increase in browse utilisation to optimise nutrient intake. Even though the woodland and grassland vegetation types were associated with the highest and lowest total number of avoided and rejected woody species, the influence of vegetation type on woody species selection was not statistically significant.
In addition, browse utilisation is influenced by the chemical traits of browse (Hejcmanová et al., 2019). It is inversely related to lignin concentrations, condensed tannins, secondary metabolites, and toxic defensive compounds (Ndlovu and Mundy, 2008; Frerker et al., 2013; Owen-Smith and Chafota, 2012; Hejcmanová et al., 2019) and, directly related to higher crude protein and moisture contents (Makhabu and Skarpe, 2006; Derebe and Girma, 2020). Vegetation growing on nutrient-deficient soils tend to show a greater prevalence of chemical defences (Owen-Smith and Chafota, 2012; Wigley et al., 2019; Derebe and Girma, 2020) and hence are avoided by herbivores (Hejcmanová et al., 2019; Derebe and Girma, 2020). However, in the KFR the proportion of avoided and rejected woody species was comparable across the three vegetation types, ranging between 33.33 and 37.04%, but the nutrient-rich grassland vegetation was associated with the highest number of selected woody species. Leaves of most Combretaceae species are phenolic rich and hence were avoided by elephants but commonly eaten by ruminants (Owen-Smith and Chafota, 2012). In the KFR, the selection of Combretaceae species exhibits variability at species and vegetation type levels. Even though KFR hosts a wide diversity of browsers, Combretum imberbe was rejected, while Combretum collinum showed a mixed pattern as it was avoided and selected in the woodland vegetation and grassland vegetation, respectively. Terminalia sericea was avoided in the shrubland but taken commensurate to its availability in the woodland vegetation. This is consistent with site specificity (in this case vegetation type) of nutrient deficiencies and the fact that the nutritive value of foliage of the same plant species can vary across sites (Hejcmanová et al., 2019). Browse quality or chemistry was not measured as it was not part of the scope of this study; hence, the effect of browse quality or chemistry on utilisation and selection of woody species in the KFR cannot be speculated. However, this presents an opportunity for future research on browse utilisation in the KFR.
In other studies (e.g., Mosbacher and Williams, 2009; Owen-Smith and Chafota, 2012; Frerker et al., 2013), herbivores tended to select few plant species available to them. Similarly, the herbivore community of the KFR focussed (moderate-to-high utilisation) and preferentially browsed on relatively very few woody species constituting at most 16.7% (n = 4) in the grassland vegetation. On the other hand, a third (33–37%) of woody species were avoided or completely neglected by the KFR browser community at all scales of aggregation. This is considerably narrower than the two-thirds (66.6%) neglected and rejected woody species reported by Owen-Smith and Chafota (2012) for the African elephant also in the Chobe District just north of KFR. The disparity is certainly a result of the additive effect of other browser species in the KFR that were not considered by Owen-Smith and Chafota (2012).
Also, predation risk can influence the distribution of herbivores and hence their impact on forest structure (Theuerkauf and Rouys, 2008). Predation risk can alter herbivore habitat selection and foraging behaviour (Cain et al., 2024). Hunting efficiency of stalking carnivores (e.g., lion and leopard) requires a balance between cover and visibility (Van Orsdol, 1984; Cain et al., 2024). In the nearby Chobe Riverfront, this balance was optimum in the shrubland vegetation as it was preferred by lions, while the denser woodland vegetation was avoided (Neo, 2001; Andreassen et al., 2014). As browsing can be inversely related to predation risk (Frerker et al., 2013; Cain et al., 2024), it was expected that the shrubland vegetation would be associated with lower browsing pressure as a consequence of higher predation risk and the reverse true for the other denser vegetation types. On the contrary, the proportion (i.e., 37%) of woody species browsed at moderate and high utilisation was highest in the shrubland vegetation. In addition, overall proportionate browse utilisation was higher in the shrubland vegetation relative to the grassland vegetation where predation risk is expected to be comparatively lower due to low visibility as a result of tall dense grass. Even though indirect signs (i.e., tracks) and three lions were observed during field surveys, both browse utilisation and selection in the KFR were not consistent with the possible influence of predation risk, as driven by vegetation thickness/openness.
5 Conclusion
Unlike most studies cited herein that assessed the impact of a single or few herbivore species, this study looked at collective KFR herbivore community browsing and thus does not enable direct comparison with single-browser studies. Results of this study show that the browser community of the KFR rejected some woody species, light to moderate browsing of most and selected very few woody species. Similar to other landscapes, across all vegetation types, the herbivore community KFR selected or utilised at high intensity very few extant woody species. Though not definitive in this study, even at low browse utilisation and selection, herbivores can direct changes in a vegetation community (Sankaran et al., 2013; Pekin et al., 2014), and such changes have already been realised in the KFR (Fox et al., 2017). Consistent with Sankaran et al. (2013) and Mosbacher and Williams (2009), this study has laid a benchmark for future monitoring to assess the potential role of herbivory on changes in vegetation community structure of the KFR. Furthermore, this study recommends future research on environmental and biological correlates of browse utilisation and selection based on the sampling plots defined by this study and the development of a long-term monitoring programme for the Kazuma Forest Reserve.
Statements
Data availability statement
The raw data supporting the conclusions of this article will be made available on request by the corresponding author, without undue reservation.
Author contributions
GN-M: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. MR: Conceptualization, Investigation, Methodology, Writing – review & editing. KM: Data curation, Investigation, Methodology, Writing – review & editing. KO: Data curation, Investigation, Methodology, Writing – review & editing. TM: Data curation, Investigation, Methodology, Writing – review & editing. EK: Data curation, Investigation, Methodology, Writing – review & editing. RM: Conceptualization, Investigation, Methodology, Writing – review & editing. WM: Conceptualization, Investigation, Writing – review & editing. DT: Conceptualization, Investigation, Methodology, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. All funds and materials support for this study were provided by the Research Technology Development and Transfer Committee of the Botswana University of Agriculture and Natural Resources.
Acknowledgments
The authors would like to thank the Botswana University of Agriculture and Natural Resources for providing funds and logistical support and the Ministry of Environment, Natural Resources Conservation, and Tourism for permission to carry out the study. The Department of Wildlife and National Parks (Kasane office) is deeply appreciated for providing security against dangerous wildlife during field data collection.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial gains that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1
Aarrestad P. A. Hytteborn H. Masunga G. Skarpe C. (2014). “Vegetation; Between soils and herbivores” in Elephants and savanna woodand eocsystems;'A study from Chobe National Park, Botswana. eds. SkarpeC.DutoitJ. T.MoeS. R. (United kingdom: Wiley Blackwell & Zoological Society of London).
2
Andreassen H. P. Neo-Mahupeleng G. Flagstad O. Wegge P. (2014). “The Chobe Riverfornt lion population: a large predator as a responder to elephant-induced habitat heterogeneity” in Elephants and savanna woodland ecosystem: A study from Chobe National Park, Botswana. eds. SkarpeC.Du ToitJ. T.MoeS. R. (Hoboken, NJ: Wiley Blackwell & Zoological Society of London).
3
Ben-Shahar R. (1993). Patterns of elephant damage to vegetation in northern Botswana. Biol. Conserv.65, 249–256. doi: 10.1016/0006-3207(93)90057-8
4
Ben-Shahar R. (1996). Woodland dynamics under the influence of elephants and fire in northern Botswana. Vegetatio123, 153–163. doi: 10.1007/BF00118268
5
Bödeker K. Ammer C. Knoke T. Heurich M. (2021). Determining statistically robust changes in ungulate browsing pressure as a basis for adaptive wildlife management. Forests12:1030. doi: 10.3390/f12081030
6
Cain J. Kay J. Liley S. Gedir J. (2024). Mule deer (Odocoileus hemionus) resource selection: trade-offs between forage and predation risk. Front. Ecol. Evol.12:1121439. doi: 10.3389/fevo.2024.1121439
7
Chase M. Schlossberg S. Sutcliffer S. Seonyatseng E. (2018). Dry season aerial survey of elephants and wildlife in northern Botswana: July – October 2018. Elephant Without Borders/Department of Wildife and National Parks. Botswana. Available online at: https://elephantswithoutborders.org/site/wp-content/uploads/2018-Botswana-report-final-version-compressed-upload.pdf (Accessed December 9, 2024)
8
De Boer W. F. Oort J. Grover M. Peel M. (2015). Elephant-mediated habitat modifications and changes in herbivore species assemblages in Sabi sand, South Africa. Eur. J. Wildl. Res.61, 491–503. doi: 10.1007/s10344-015-0919-3
9
Derebe Y. Girma Z. (2020). Diet composition and preferences of Bohor reedbuck (Redunca redunca) in the compound of Alage college, central Rift Valley of Ethiopia. Ecol. Evol.10, 13370–13381. doi: 10.1002/ece3.6939
10
Fox J. T. Vandewalle M. E. Alexander K. A. (2017). Land cover change in northern Botswana: the influence of climate, fire, and elephants on semi-arid savanna woodlands. Land6:73. doi: 10.3390/land6040073
11
Frerker K. Sonnier G. Waller D. (2013). Browsing rates and ratios provide reliable indices of ungulate impacts on forest plant communities. For. Ecol. Manag.291, 55–64. doi: 10.1016/j.foreco.2012.11.041
12
Goheen J. Young T. Keesing F. Palmer T. (2007). Consequences of herbivory by native ungulates for the reproduction of a savanna tree. J. Ecol.95, 129–138. doi: 10.1111/j.1365-2745.2006.01196.x
13
Hasler H. Senn J. (2012). Ungulate browsing on European silver fir Abies alba: the role of occasions, food shortage and diet preferences. Wildl. Biol.18, 67–74. doi: 10.2981/09-013
14
Hejcmanová P. Miřejovská M. Homolka P. Hejcman M. (2019). Drivers of diet selection of critically endangered Western Derby eland during the food shortage period within conservation breeding in Senegal. Sci. Rep.9:8712. doi: 10.1038/s41598-019-45035-z
15
Machar I. Čermák P. Pechanec V. (2018). Ungulate browsing limits bird diversity of the central European hardwood floodplain forests. Forests9:373.
16
Makhabu S. Skarpe C. (2006). Rebrowsing by elephants three years after simulated browsing on five woody plant species in northern Botswana. S. Afr. J. Wildl. Res.36, 99–102.
17
Makhabu S. Skarpe C. Hytteborn H. (2006). Elephant impact on shoot distribution on trees and rebrowsing by smaller browsers. Acta Oecol.30, 136–146. doi: 10.1016/j.actao.2006.02.005
18
Mmolotsi R. M. Obopile M. Kwerepe B. C. Sebolai S. Rampart M. P. Segwagwe A. T. et al . (2012). Studies on Mukwa (Pterocarpus angolensis, D. C.) dieback in Chobe Forest reserves in Botswana. J. Plant Stud.1, 154–157. doi: 10.5539/jps.v1n2p154
19
Mojeremane W. Makgobota K. Mathowa T. Obusitswe K. Rampart M. Teketay D. et al . (2021). Evaluation of dormancy-breaking treatments on seed germination of two leguminous tree species from Chobe District, Northern Botswana. Forestry Ideas27, 380–395.
20
Mosbacher E. Williams C. (2009). Browse preference and browsing intensity of white-tailed deer (Odocoileus virginianus) in Allegheny high plateau riparian forests, USA. Wildl. Biol. Pract.5, 11–21. doi: 10.2461/wbp.2009.5.4
21
Möst L. Hothorn T. Müller J. Heurich M. (2015). Creating a landscape of management: unintended effects on the variation of browsing pressure in a national park. Forest Ecol. Manag.338, 46–56. doi: 10.1016/j.foreco.2014.11.015
22
Mosugelo K. Moe S. Ringrose S. Nellemann C. (2002). Vegetation changes during a 36-year period in northern Chobe National Park, Botswana. Afr J Ecol40, 232–240. doi: 10.1046/j.1365-2028.2002.00361.x
23
Mramba R. (2021). Browsing behaviour of impala, Aepyceros melampus in two contrasting savannas. Global Ecol. Conserv.30:e01770. doi: 10.1016/j.gecco.2021.e01770
24
Ndlovu M. Mundy P. (2008). Browse preference of captive black rhinos at Chipangali wildlife orphanage, Zimbabwe. Pachyderm45, 41–46. doi: 10.69649/pachyderm.v45i.165
25
Neo G. (2001). Group dynamics, home ranges and habitat use by lions, Panthera leo, in Chobe National Park, northeastern Botswana. Ås: Norwegian University of Life Science.
26
Owen-Smith N. Chafota J. (2012). Selective feeding by a megaherbivore, the African elephant (Loxodonta africana). J. Mammal.93, 698–705. doi: 10.1644/11-MAMM-A-350.1
27
Pekin B. K. Wisdom M. J. Endress B. A. Naylor B. J. Parks C. G. (2014). Ungulate browsing maintains shrub diversity in the absence of episodic disturbance in seasonally-arid conifer Forest. PLoS One9:e86288. doi: 10.1371/journal.pone.0086288
28
Rutina L. P. Moe S. R. Swenson J. E. (2005). Elephant Loxodonta africana driven woodland conversion to shrubland improves dry-season browse availability for impalas Aepyceros melampus. Wildl. Biol.11:207–213. doi: 10.2981/0909-6396(2005)11[207:ELADWC]2.0.CO;2
29
Sankaran M. Augustine D. J. Ratnam J. (2013). Native ungulates of diverse body sizes collectively regulate long-term woody plant demography and structure of a semi-arid savanna. J. Ecol.101, 1389–1399. doi: 10.1111/1365-2745.12147
30
Sianga K. Fynn R. W. S. Bonyongo M. C. (2017). Seasonal habitat selection by African buffalo Syncerus caffer in the Savuti–Mababe–Linyanti ecosystem of northern Botswana. Koedoe.59:a1382. doi: 10.4102/koedoe.v59i2.1382
31
Skarpe C. Masunga G. Aarrestad R. A. Frost P. G. H. (2014). “Soils as a controller of and responder to elephant activity” in Elephant and savanna woodland ecosystem: A study from Chobe National Park, Botswana. eds. SkarpeC.DutoitJ. T.MoeS. (London: Wiley Blackwell & Zoological Society of London).
32
Theuerkauf J. Rouys S. (2008). Habitat selection by ungulates in relation to predation risk by wolves and humans in the Białowieża Forest, Poland. For. Ecol. Manag.256, 1325–1332. doi: 10.1016/j.foreco.2008.06.030
33
Valeix M. Hemson G. Loveridge A. Mills M. Macdonald D. (2012). Behavioural adjustments of a large carnivore to access secondary prey in human-dominated landscape. J. Appl. Ecol.49, 73–81. doi: 10.1111/j.1365-2664.2011.02099.x
34
Van Orsdol K. (1984). Foraging behaviour and hunting success of lions in queen Elizabeth National Park, Uganda. Afr. J. Ecol.22, 79–99. doi: 10.1111/j.1365-2028.1984.tb00682.x
35
Wigley B. J. Coetsee C. Augustine D. J. Ratnam J. Hattas D. Sankaran M. (2019). A thorny issue: Woody plant defence and growth in an east African savanna. J. Ecol.107, 1839–1851. doi: 10.1111/1365-2745.13140
Summary
Keywords
Chobe, woody species, Combretaceae, Fabaceae, Baikiaea plurijuga , woodland vegetation, grassland vegetation, shrubland vegetation
Citation
Neo-Mahupeleng G, Rampart M, Makgobota K, Obusitswe K, Makoi T, Kgosiesele E, Mmolotsi R, Mojeremane W and Teketay D (2025) Herbivore browse utilisation and selection in the Kazuma Forest Reserve, Northern Botswana. Front. For. Glob. Change 8:1500962. doi: 10.3389/ffgc.2025.1500962
Received
24 September 2024
Accepted
10 February 2025
Published
10 March 2025
Volume
8 - 2025
Edited by
Elisa Carrari, University of Florence, Italy
Reviewed by
Pascal Vittoz, Université de Lausanne, Switzerland
María José Corriale, University of Buenos Aires, Argentina
Keotshephile Kashe, University of Botswana, Botswana
Updates
Copyright
© 2025 Neo-Mahupeleng, Rampart, Makgobota, Obusitswe, Makoi, Kgosiesele, Mmolotsi, Mojeremane and Teketay.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gosiame Neo-Mahupeleng, gmahupeleng@buan.ac.bw
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.