Abstract
Hardwood forests dominated by sugar maple (Acer saccharum) at its northern limit (Quebec) are threatened by rising regional temperatures and recurrent summer drought. In a rainfall exclusion experiment, we imposed artificial drought in six maple stands where American beech (Fagus grandifolia) saplings proliferated in the understory (with overstory beech presence) or were absent. Two paired plots were established in sites where beech proliferated vs. those that were beech-free, i.e., covered by rainfall excluders vs. left uncovered for summer 2021 and 2022. Responses to the four resulting treatments were subjected to Bayesian analysis. Dendroclimatological estimates of annual basal area increment (BAI) produced by three canopy maples within 12 plots, together with foliar nutrients of the same mature maple trees and soil nutrient pools (ion-exchange probes, with increasing burial duration), were compared across plots. Comparisons included ordination scores for foliage (DCA1, Detrended Correspondence Analysis) and Principal Component Analysis (PCA: PC1 and PC2) for soils. Soil water content (SWC) beneath excluders was 56% of adjacent rainfall-accessible plots; beneath understory beech saplings, this deficit negatively affected maple BAI, but only in 2022. Diagnosis and Recommendation Integrated System (DRIS) indices indicated foliar Mg insufficiency and Ca sufficiency. Mg insufficiency increased with rainfall exclusion, regardless of beech sapling presence. Rainfall exclusion reduced soil nutrient supply levels relative to controls; some supply rates (total pools, PC1, PC2, acidity) progressively increased over probe burial duration (2021: 14–56 days) in a diffusion-dependent manner under excluders, while reaching a plateau (equilibrium-dependent adsorption) with accessible rainfall. NO3-N progressively dominated beech-free soil inorganic-N pools over time, which diminished regardless of rainfall exclusion. Likewise, soil base cations (Ca + Mg + K) remained high in beech’s absence, and vice versa for acidity (Al + Fe + Mn). Modest rainfall manipulations decreased BAI, altered adsorption processes, and shifted soil pools from endo- to ectomycorrhizal dominance. Even short-term moisture deficiency in maple soils may incur further stress in already nutrient-stressed stands, initiating nutrient cycle uncoupling between foliar and soil pools.
1 Introduction
Sugar maple (Acer saccharum Marsh.) is subject to widespread “maple decline” (Boakye et al., 2023), where the vitality and persistence of this iconic long-lived and highly shade-tolerant tree across the northern hardwood forest are threatened by soil cation depletion and imbalance, Al toxicity, pest and pathogen outbreaks, and forestry operations (Duchesne et al., 2002; Phillips et al., 2013; Bal et al., 2015; Bishop et al., 2015). The effects of these stress factors are exacerbated by warmer seasonal temperatures, reduced snowpacks, and summer droughts (Moreau et al., 2019, 2020; Copenheaver et al., 2020; Stern et al., 2023), which are projected to increase in both severity and duration across the region (Boulanger et al., 2017; Coble et al., 2017; Moreau et al., 2022; Cholet et al., 2022). At the northernmost limit of its range (Quebec, Canada), sugar maple dominates broadleaf stands across three adjacent bioclimatic domains (Saucier et al., 1998). The forest canopy is co-dominated by Betula alleghaniensis Britt. in the sugar maple–yellow birch domain, by Tilia americana L. in the sugar maple–American basswood domain, and by Carya cordiformis [Wangenh.] K. Koch in the sugar maple–bitternut hickory domain. Unlike sugar maple, these three species have lower shade tolerances (Leak et al., 2014) and ectomycorrhizal (ECM) root systems, as does American beech (Fagus grandifolia Ehrh.).
American beech is likely to replace declining maple as the canopy dominant given that it exhibits shade tolerance equal to that of sugar maple, together with more flexible responses to environmental disturbances than the latter species (Bose et al., 2017a, 2017b; Boakye et al., 2023; Leduc et al., 2024), both in the canopy and understory. Indeed, beech is occurring more frequently in sugar maple stand understories, with various studies reporting very dense understory beech saplings (Duchesne and Ouimet, 2009; Nolet and Kneeshaw, 2018; Moreau et al., 2020). The understory proliferation of beech in sugar maple stands, particularly through root suckering, may be triggered by infection and eventual mortality of mature beech in the overstory due to the widespread occurrence of Beech Bark Disease (Roy and Nolet, 2018; Elenitsky et al., 2020; Kish et al., 2022). Consequences of sapling proliferation (not BBD-susceptible) have been associated with reduced diversity within forest stands (Cale et al., 2015), competition for available soil N (Simon, 2023), imbalances in other soil nutrients (Zarfos et al., 2024), and altered water fluxes (Rasoanaivo et al., 2024). In the presence of recurrent moisture deficits, these responses have widespread implications in terms of the recruitment and competitive relationships among northern hardwood species (Cleavitt et al., 2021; Henry and Walters, 2023), together with biogeochemical cycles and ecosystem services.
Drought has been proposed as an environmental driver that uncouples foliar nutrients from soil nutrient supplies and soil nutrient cycles (Evans and Burke, 2013; Schönbeck et al., 2020; Liu et al., 2023). Accordingly, we performed a two-year-long experiment in sugar-maple-dominated northern hardwood stands, where American beech was present in both the canopy and understory or absent from these strata. In turn, these conditions were crossed with the growing-season-long absence or presence of rainfall excluders to induce soil moisture deficiencies. Under these four treatment conditions, we employed dendroclimatology to determine whether the imposition of the drought treatment resulted in a decreased BAI (basal area increment) in the canopy maples. We further assessed foliar nutrient sufficiency or deficiency in canopy maples using diagnosis and recommendation integrated system (DRIS) and monitored soil nutrient supply rates across the four treatments. In the last task, we assumed that the mobility of nutrient ions in soil solution can be severely reduced in the presence of soil moisture deficits, decreasing to such an extent that drought-affected element concentrations, their stoichiometries, and stocks would markedly differ from those measured under moisture-sufficient conditions both in plants and the soil (Sardans and Peñuelas, 2012).
As Sardans and Peñuelas (2012) and others have noted in long-term studies of drought, its progression results in a decrease in fine root abundance and increasing stress in aboveground tissues that are subject to possible hydraulic failure, leaf drop, and diminished macronutrient contents, including N, P, K, Mg, and S (Sardans and Peñuelas, 2007; Brunner et al., 2015; Urbina et al., 2015; Grams et al., 2021). With drought-induced alteration of element ratios between soil exchange surfaces (including roots) and a diminished solution phase, and reduced uptake and translocation to stems and leaves, element stoichiometry is likely to change, particularly in the latter tissues, which also see a concomitant increase in C and Fe content. Urbina et al. (2015) have further suggested that plants growing within a species-diverse community setting would exhibit greater stoichiometric plasticity than those individuals grown in a monoculture. As a component of our correlation analyses, we posited that observed variation in canopy species composition among sites (weighted shade tolerance) would exert an effect, albeit a small one, on soil nutrient pool compositions in each growing season.
We had four goals. As briefly described above, our first objective was to determine whether we could effectively detect a drought signal of maple growth decline under experimental rainfall exclusion conditions that included enforced soil moisture deficits. Our second objective was to compare the response of foliar macronutrient status to rainfall exclusion in sugar maple stands with and without a dense beech sapling-dominated understory. We posited that the stress of experimentally imposed soil moisture deficits would worsen any existing nutrient insufficiencies in the leaves. Our third objective was to compare the response of soil supply rates and dynamics of various nutrients to rainfall exclusion in sugar maple stands with and without a dense beech-dominated understory using ion-exchange membrane technology. We predicted that drought imposed by rainfall excluders would interrupt the time course toward achieving equilibrium-dependent ion adsorption, which was monitored through sequential PRS retrievals after 14, 28, 42, and 56 days of burial in 2021. As a fourth objective, we generated correlations among above- and below-ground variables (foliage vs. soil) separately for the rainfall exclusion and rainfall-accessible treatments. We hypothesized that correlations among individual elements, element ensembles (total nutrient pools, dimensionally reduced pool compositions), and between foliage versus soil (plus other plot-level variables) would be weaker under drought conditions (rainfall exclusion = Yes) than under higher ambient moisture levels (rainfall exclusion = No). We further posited that the onset of soil moisture deficit would weaken correlations among soil nutrients, suggesting their subsequent decoupling.
2 Materials and methods
2.1 Study region
The study was conducted in the Kenauk Nature Reserve (Quebec, Canada), a private land area (265 km2) in the yellow birch–sugar maple bioclimatic domain (Saucier et al., 1998). Within the reserve, the late-successional and uneven-aged hardwood stands we selected have experienced few large-scale natural disturbances; others have been subjected to recent forestry operations (Roy et al., 2021a, 2021b). The regional climate is cold-temperate continental. The annual temperature is 5.3°C (running average, 1981–2010). Annual precipitation averages 1,204 mm (station 7035110, Notre-Dame-de-la-Paix, Environment & Climate Change Canada). Soils supporting these forests have developed from glacial till, with parent materials including gneiss, quartzite, and granite (Lajoie, 1967; Soil Classification Working Group, 1998). Soils are classified as Dystric Brunisols in the Canadian system (USDA: Typic Dystrochrepts) with moderate humus (Roy et al., 2021a, 2021b).
2.2 Experimental design
Six sugar maple-dominated sites were selected from among neighboring hardwood forest stands, which were undisturbed by human activities (Table 1). Two 20 m × 20 m plots were established on each site, in which individual trees > 9 cm DBH (diameter at breast height, 1.3 m) were identified to species and counted to estimate density (stems ha−1). Total canopy basal areas (BA, m2 ha−1) were recorded (Table 1), along with those of individual species, after being standardized to a per-hectare basis. Saplings (woody stems with DBH < 9 cm) in the understory were recorded by species in 100 m2 quadrats (Appendix 1). These data were combined to estimate total understory basal area (m2 ha−1). Our working sapling classification includes poles to emphasize the overwhelming understory dominance of American beech; Natural Resources Canada generally defines saplings as being 1–2 m tall, with 2–4 cm DBH, while excluding 5–9 cm DBH poles (Forestry Glossary).1
Table 1
| Site ID | Elev. (m) | Lat. | Long. | Proliferation | Exclusion | Total Sapl. BA (Beech) | Canopy BA (Beech) | Canopy SM BA | SM DBH Mean ± SD |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 239 | 45.76 | −74.90 | Yes | Yes | 3.28 (3.11) | 35.46 (7.52) | 20.43 | 29.83 (± 8.02) |
| 1 | 240 | 45.76 | −74.90 | Yes | No | 3.04 (2.25) | 36.44 (8.85) | 9.65 | 26.17 (± 5.69) |
| 2 | 258 | 45.74 | −74.93 | Yes | Yes | 3.23 (2.50) | 29.91 (0.73) | 27.16 | 29.10 (± 3.68) |
| 2 | 260 | 45.74 | −74.93 | Yes | No | 2.67 (2.38) | 41.22 (15.47) | 25.25 | 35.67 (± 4.45) |
| 3 | 239 | 45.74 | −74.95 | Yes | Yes | 3.10 (3.10) | 25.01 (3.40) | 18.83 | 30.17 (± 6.52) |
| 3 | 237 | 45.75 | −74.95 | Yes | No | 2.49 (2.46) | 31.03 (5.94) | 19.38 | 37.53 (± 4.02) |
| 5 | 258 | 45.75 | −74.95 | No | Yes | 0.93 (0.0) | 34.81 (0.0) | 26.37 | 29.03 (± 5.20) |
| 5 | 256 | 45.75 | −74.94 | No | No | 0.88 (0.0) | 44.08 (0.0) | 38.35 | 38.23 (± 6.56) |
| 6 | 284 | 45.76 | −74.94 | No | Yes | 0.37 (0.0) | 30.52 (0.0) | 30.25 | 35.90 (± 13.44) |
| 6 | 279 | 45.76 | −74.94 | No | No | 0.71 (0.0) | 32.61 (0.0) | 32.46 | 35.00 (± 11.73) |
| 7 | 264 | 45.76 | −74.94 | No | Yes | 0.82 (0.0) | 36.53 (0.0) | 33.40 | 32.77 (± 3.91) |
| 7 | 271 | 45.76 | −74.94 | No | No | 0.06 (0.02) | 31.34 (0.1) | 27.16 | 36.20 (± 1.95) |
Six sugar maple (SM) stands within Kenauk Nature Reserve (Quebec): three sites where American beech saplings had proliferated (Yes) vs. three beech-sapling-free sites (No).
Total sapling basal area (BA), m2 ha−1 (Beech BA in parentheses), together with total canopy BA (Beech BA in parentheses) are shown. Plots within sites (20 m x 20 m) were paired: rainfall excluder-covered (Exclusion: Yes) vs. rainfall-accessible (Exclusion: No). SM DBH (cm) is mean ± SD of three canopy sugar maples in each plot.
Saplings: Our working classification includes poles (2–9 cm DBH); Natural Resources Canada (NRC) defines saplings as 1–2 m tall, with 2–4 cm DBH, explicitly excluding 5–9 cm DBH poles (Forestry Glossary, https://cfs.nrcan.gc.ca/terms/read/996). Canopy DBH (Diameter at breast height, cm) means and standard deviations (± SD) are based on three canopy sugar maples (DBH > 10 cm DBH) in each of 12 plots (n = 36). Sapling and over-story beech BA are positively correlated (r = 0.785, [0.294, 0.924], BF10 = 20.75). Ele., elevation above sea level; Lat., latitude; Long., longitude, west of the prime meridian.
American beech saplings dominated the understory of three stands (Proliferation = Yes: sites 1, 2, and 3) but were absent or very scarce in three others (Proliferation = No: sites 5 and 6; site 7 contains some beech). The six sites were subdivided into two plots (each 20 m × 20 m), pairing rainfall exclusion with rainfall-accessible soils to compare plots with enforced soil moisture deficiency (SWC, soil water content) with rainfall-accessible soils on each site. According to the experimental layout, 12 plots resulted from four treatments (Table 1), each with three replicates: no beech proliferation–no rainfall exclusion (No–No); no beech proliferation–rainfall exclusion (No–Yes); beech proliferation–no rainfall exclusion (Yes–No); and beech proliferation–rainfall exclusion (Yes–Yes). Rainfall excluders covered the designated plots at the beginning of two consecutive growing seasons. Rainfall was intercepted (> 75%) and diverted off the plot by 3.6 m × 3 m clear polyethylene tarps (Kleton, 8 mil-thick; Tenaquip Ltd., Senneville, QC). These were deployed in a grid pattern to form an impermeable tent suspended from the trees at ~1.5 m above the ground. In 2021, exclusion tents were set in place from early July to mid-September and subsequently removed before autumn litter fall. They were again erected from June to mid-September 2022.
The efficacy of rainfall excluders relative to uncovered plots was monitored through in situ data-logger recordings of summer variation (2021, 2022) in soil water content (SWC, % or cm3 cm−3) and temperature (0–10 cm). Placed near the plot centers, the loggers measured these properties (15-min intervals) within excluder-covered and rainfall-accessible plots with Decagon 5TM sensors (Decagon Devices, Pullman, WA). When the experiment ended, manual measurements integrated over the uppermost 12 cm of mineral soil were taken at 42 grid points superimposed on each plot using portable TDR (Time-Domain-Reflectometry). TDR was equipped with 12-cm-long waveguides (FieldScout TDR 100, Spectrum Technologies Inc., Aurora, IL). Bulk pH of the sandy loam soils was determined with a combination electrode on 7:1 (Fermentation-Humification [F-H] layer) and 2:1 (mineral) water-soil slurries (CPVQ, 1997). Results are reported in Table 2 and Appendix 2.
Table 2
| Site ID | Prolif. | Excl. | Foliar DCA1 | 2021 PC1 | 2021 PC2 | 2022 PC1 | 2022 PC2 | SWC % | F-H pH | Min. pH |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Yes | Yes | 7.667 | 0.360 | −1.924 | 0.403 | −0.897 | 12.6 | 4.45 | 3.90 |
| 1 | Yes | No | 8.333 | 0.267 | −1.315 | 0.411 | −2.070 | 22.81 | 3.93 | 3.99 |
| 2 | Yes | Yes | 6.333 | 0.484 | −1.542 | 0.725 | −1.388 | 9.71 | 4.47 | 4.35 |
| 2 | Yes | No | 6.667 | 1.685 | 0.530 | 2.113 | −0.702 | 14.20 | 4.64 | 4.15 |
| 3 | Yes | Yes | 14.000 | 0.812 | −0.429 | 1.569 | −1.419 | 9.40 | 5.01 | 4.42 |
| 3 | Yes | No | 4.333 | 1.856 | −0.074 | 0.419 | −1.190 | 11.26 | 4.44 | 4.15 |
| 5 | No | Yes | 26.33 | −0.1046 | −1.297 | −1.908 | 0.058 | 5.98 | 5.73 | 5.15 |
| 5 | No | No | 28.667 | −2.550 | 1.356 | −1.255 | 1.384 | 12.87 | 5.26 | 5.18 |
| 6 | No | Yes | 36.667 | −2.404 | 0.271 | −1.249 | 1.668 | 8.43 | 5.25 | 5.34 |
| 6 | No | No | 30.333 | −1.975 | 1.315 | 0.476 | 3.003 | 18.59 | 4.45 | 4.63 |
| 7 | No | Yes | 20.000 | −0.685 | −1.633 | −1.145 | −0.095 | 6.79 | 4.56 | 4.35 |
| 7 | No | No | 21.000 | −0.772 | 0.382 | −0.558 | 1.447 | 13.71 | 4.95 | 4.79 |
Detrended Correspondence Analysis (DCA) of sugar maple foliar nutrients and soil nutrient pool Principal Component Analysis (PCA) ordination scores for 2021 and 2022.
Soil Water Content (%) was measured on 1-09-2022 (Parsons et al., 2025), together with F-H and mineral soil pH.
Soil moisture content% (mean ± SD) in rainfall excluder plots is 8.82 ± 2.35% compared to 15.57 ± 4.30% in rainfall-accessible plots, according to manual TDR measurements (0–12 cm depth). Mean (± SD) Fermentation-Humus (F-H) layer and mineral soil pH values under beech sapling proliferation were about one unit lower than beech sapling-free plots, i.e., 4.1 ± 0.4 vs 5.0 ± 0.5 and 4.2 (± 0.2) vs 4.9 (± 0.4), respectively.
2.3 Basal area increment (BAI)
Three canopy sugar maples were sampled near the 12-plot centers to avoid possible edge effects and cored at breast height. From 12 to 16 September 2022, two increment cores were taken from opposite sides of each tree, together with its DBH (see Table 1, means ± SDs). Mounted cores were sanded flat prior to scanning (CDendro, optical resolution = 1,200 dpi). Annual radial increments between 1993 and 2022 were determined by measuring ring widths (± 0.001 mm) using CDendro (v. 7.5, Cybis Elektronik & Data AB, Saltsjöbaden, Sweden). Subsequent cross-dating ensures the determination of the exact year for each growth ring while verifying potential missing, false, or locally absent rings (Yamaguchi, 1991; Fritts, 2012).
Annual BAI (cm2 y−1) was estimated by subtracting the transverse-sectional area of the tree bole at year n-1 from that of year n. Annual BAI removed age-related growth trends while retaining suppression or release events that were incurred by potential disturbances, such as drought. To better control for possible biases generated by variation in inter-annual growth and diameter differences among individuals, we calculated the ratio between annual BAI estimated in 2021 and 2022, respectively 1 and 2 years after rainfall exclusion, divided by the average annual BAI estimated for 2010–2020.
2.4 Sugar maple foliar nutrition
To assess foliar nutrition and growth, leaves from the three aforementioned maple trees were sampled in each plot (N = 36). Fully expanded leaves were harvested from the trees on 25–29 July 2022, when tissue chemical compositions were stable. Two mid- to upper-crown branches (10+ m height, opposite directions) per tree were cut to obtain 10 healthy, completely formed leaves per branch. Harvested leaves were oven-dried (60°C for 48 h) prior to milling (Retsch MM 400 Vibro-Mill, Haan, Germany) to pass a 60-μm mesh. Between 1.7 and 1.8 g of ground leaf material per tree produced a compressed pellet (10 mm thickness, 13 mm diameter) that was suitable for X-ray spectroscopy (XRF). Sample pellets were prepared and measured by Laboratoire iForêt (Département Science et Technologie, Université TELUQ, Montreal, QC). Foliar elemental concentrations (% or g kg−1) of Nitrogen (N), P (Phosphorus), K (Potassium), Ca (Calcium), and Mg (Magnesium) were determined using a Vanta M series X-ray fluorescence spectrometer (XRF), which was equipped with an 8–50 kV rhodium tube and silica drift detector (Olympus, Tokyo, Japan). We have successfully applied this instrument to previous nutrient assessments in other forested sites of southern Quebec (Grimond et al., 2024).
2.5 Diagnosis and recommendation integrated system (DRIS)
We calculated DRIS (Diagnosis & Recommendation System; Beaufils, 1973; Walworth and Sumner, 1987) indices to understand overall plant nutritional status in fully expanded mid-summer leaves of sugar maple growing in rainfall-exclusion plots versus excluder-free controls, in the presence or absence of beech saplings. Nutrient ratios were calculated by this procedure and compared against external foliar standards for sugar maple (reference population; Appendix 3) to determine whether they were in excess or deficient (i.e., > 0 or < 0). Means (± SD) were estimated for the five previously described foliar nutrients, which had been determined by XRF. These estimates and their coefficients of variation (CV) were combined with reference data (Lozano and Huynh, 1989) for healthy sugar maple stands to calculate their respective DRIS indices.
Significant nutrient deficiency is indicated by a DRIS less than −15, while nutritional quality is deemed sufficient when the DRIS index for a given element is zero or greater (Masson et al., 2019). The nutrient disequilibrium index (NDI) is the summed absolute values of the nutritional DRIS indices (Walworth and Sumner, 1987). A total index of nutritional stress indicates the following: NDI < 60, balanced nutrient status; NDI 60–100, slight nutrient deficiency; NDI 100–200, moderate nutrient deficiency; and NDI > 200, major nutrient deficiency. Ideally, the NDI would approach zero when foliar nutrient contents are sufficient (Ouimet et al., 2013). In addition to Masson et al. (2019), the NDI has been used successfully by Moore and Ouimet (2006) to interpret a 10-year-long study of dolomite liming rate effects on sugar maple nutrition and growth in southern Quebec.
2.6 Soil nutrient supplies
Bioavailability of six major soil nutrients (NO3−, NH4+, P, K, Ca, Mg) was measured using Plant Root Simulator probes (PRS, Western Ag Innovations, Saskatoon, SK) that were buried and retrieved at regular intervals (Hangs et al., 2004). Cation or anion exchange membranes (adsorbing surface area: 17.5 cm2 probe−1) are encapsulated in thin plastic cases, facilitating easy insertion into mineral soil. Probes were deployed across Sites 1, 2, 3, 5, 6, and 7 to incubate in plots where rainfall exclusion was not implemented, paired with probes in plots where excluder tents were erected for the same duration. In each plot, four anion and four cation probes were buried to a depth of 5–10 cm for two burial periods (13 July to 6 September 2021; 28 June to 10 August 2022). Following the initial burial, a time course of probe retrievals was followed in 2021 at 14, 28, 42, and 56 days to determine whether ion adsorption remained diffusion-dependent or attained an adsorption–desorption equilibrium (Schoenau et al., 1993; Hangs et al., 2004). The duration of burial of 2022 PRS was limited to 43-day increases or decreases in supply rates and was not useful for following adsorption behavior.
Upon retrieval, the PRS were bulked into one composite sample per plot for each probe type. Probes were cleaned and stored in 4°C deionized water before complete analysis by the manufacturer (Western Ag Innovations). Probe membranes were then eluted with 0.5 M HCl. Extracted nitrate (NO3−) and ammonium (NH4+) were determined by flow-injection colorimetry (San++ Analyzer, Skalar Inc., The Netherlands). The remaining 13 nutrients were measured by inductively coupled plasma-optical emission spectrometry (Optima 8,300, Perkin-Elmer, Inc.). Nutrient supply rates per probe were calculated by company analysts from these concentrations (μg per 10 cm2 per 6 weeks).
We derived additional indices from supply rates across PRS retrieval dates in 2021 and 2022: nitrate-nitrogen (NO3-N) as a percentage of total inorganic-N (NH4-N + NO3-N) supply rates (nitrate%); aluminum (Al), iron (Fe), and manganese (Mn) as soil acidity indicators (summed Al + Fe + Mn supply rates); and base cation sums, i.e., Ca + Mg + K supply rates. For each incubation, have created 15-element compositions of N (NO3-N and NH4-N) and all mineral nutrients that were measured by ICP, which formed the vector [NH4|NO3|P|Ca|Mg|K|Fe|Mn|Zn|S|Al|Cu|B|Pb|Cd]. Copper (Cu), boron (B), lead (Pb), and cadmium (Cd) were often below detection limits, and observations were assigned zeros. Given their sparse non-zero responses, these data were not included in the 11-nutrient compositions, which were subjected to dimensional reduction using PCA ordination (see below).
2.7 Statistical analyses
Prior to the application of Bayesian methods, dimensional reduction was performed on foliar and soil nutrient ensembles using PAST version 4.2 A detrended form of Correspondence Analysis (DCA) was selected as a robust alternative to Compositional Data Analysis (Greenacre, 2021). We assumed that foliar measurements formed sets of bounded variables, each summing to a constant value, where the resulting compositions would not necessarily occupy Euclidean space. In contrast, principal component analysis (PCA) was applied to soil nutrient pool ensembles to alleviate “arching” that is frequently incurred in CA through the distortion of unimodal response curves in ordination space,3 which was not eliminated with DCA. The unrotated two-dimensional PCA solutions are based on their correlation matrices. Both DCA and PCA were performed in PAST, and the resulting ordination scores were used in ANOVA and correlation analyses.
Bayesian generalized linear mixed-effect models (GLMM) were fitted to responses in our regressions. These were performed in R v.4.0.1 (R Core Team, 2021), using package BRMS 2.16.1 (Bürkner, 2017). To minimize the influence of prior assumptions regarding our results, we assigned uninformative flat priors to model parameters. We ran four Markov chain Monte Carlo (MCMC) chains, each comprised of 2,500 warm-up iterations (discarded) and 2,500 sampling iterations. Convergence among chains was assured by checking parameters, i.e., Rhat < 1.01 (Vehtari et al., 2021). We did not conduct traditional significance tests on individual univariate responses. Rather, GLMM-estimated means and slopes were reported, together with their 95% credible intervals.
Modeling of BAI ratios between treatment years and the long-term average employed GLMM with a Gamma distribution. This model considered basal area increments as a function of rainfall exclusion, beech sapling proliferation, and their interaction. Ratios ≥ 1.0 indicate comparable or greater growth increments over the intervention (consecutive growing season rainfall exclusion in 2021 and 2022) relative to previous reference years. Ratios < 1.0 would indicate a growth decrement (slowed growth under rainfall excluders). We accounted for random effects by considering individual trees as nested within sites. Regarding leaf nutrient models, we utilized GLMMs with a Beta distribution to analyze leaf nutrients. These models incorporated the interaction and main effects of rainfall exclusion and beech proliferation. Individual trees were nested within sites and incorporated as random effects. Soil nutrient models focused on the six most abundant ions (NO3-N, P, K, Mg, Ca, NH4-N), employing similar GLMMs with a Gamma distribution. These models were constructed with soil nutrients as a function of interaction and main effects of rainfall exclusion and beech proliferation. Year was a random effect; plots nested within-sites were additional random effects.
Bayesian ANOVA focused on foliar DRIS indices and was further applied to soil nutrient responses for 2021 and 2022. We estimated 95% credible intervals using naive priors, together with estimates of Bayes Factors (BF; Johnson et al., 2023), by implementing models in JASP (JASP Team 2020, Version 0.13; [Computer software]). Clear differences among means were determined according to estimated credible intervals and Bayes Factors (BF), which lent support either to the null model (BF01) or to the best alternative (BF10). BF10 values < 1.0 support the null, while values > 1.0 would support the alternative, weakly so at BF10 = 3.0, moderately so at BF10 = 5.0, and strongly so at BF10 = 10.0 (Johnson et al., 2023).
Soil nutrient supply estimates were obtained from PRS measurements made with increasing durations of burial and probe removals. Deployment and retrieval of PRS probes were more extensive in the former (year 2021: 14, 28, 42, 56 days) than in the latter (year 2022: 13, 43 days) growing season. Following burial, we assumed that ion adsorption onto the PRS membranes was initially diffusion-dependent. Ion concentrations would progressively increase (or decrease) under this constraint until equilibrium-dependent exchange was achieved and concentrations plateaued (Schoenau et al., 1993). Under moisture-sufficient conditions, we expected that equilibrium-dependent exchange would occur as indicated by trends in mean nutrient concentrations, while increasing or decreasing diffusion-dependent trends would prevail under soil moisture deficiency.
Effectiveness of PRS burials was assessed against the main effects of incubation (burial) duration and its two- and three-way interactions with the other fixed factors (Proliferation, Exclusion). Separate Bayesian ANOVAs were conducted for 2021 and 2022, given the inclusion of responses for nitrate% (percent of total inorganic-N pool), P, base cation sums, acidity, and the broader suite of nutrients as PCA scores. We hypothesized that variation around mean nutrient estimates would increase with increasing moisture variability, as indicated by percentage coefficients of variation (CV%). When ranked across treatments, these responses were subjected to concordance analysis (Legendre, 2005). Kendall’s Coefficient of Concordance was calculated as W, where W = 1 for perfect agreement among the ranks, and W = 0 for no agreement.
Understanding whether foliar nutrient concentrations and their respective soil nutrient supplies were uncoupled by the imposition of soil moisture deficits required that foliage and soil be compared on a common basis, i.e., the plot level. We summarized elemental means (± SD) or those of the ordination scores for each of the 12 plots and subjected these data to Bayesian analysis. Coefficients of Variation (CV) were calculated from means and SDs (data not shown). We posited that leaf and soil compartments would be “uncoupled” in the presence of soil moisture deficits, as indicated by very weak Pearson correlations among variables (anecdotal evidence: BF10 ≤ 3.0) and 95% credible intervals including zero; separate correlations were run for the subsets of data where rainfall excluders were installed (Exclusion = Yes) and where excluders were absent (Exclusion = No).
The choice of indices to be correlated was important in determining potential nutrient stresses or limitations due to drought (Bal et al., 2015): First, we compared the entire foliar five-element ensemble and its proportions to determine if it tracked against the soil nutrient ensemble (separate PCA scores for 2021 and 2022). Second, we examined correlations between DRIS indices (N, P, K, Ca, Mg) and their corresponding soil concentrations in each year, together with nitrate%, base cations, inorganic-N, and acidity. Third, we extended correlations of foliar and soil nutrients beyond soil moisture and pH to attributes of the forest plots. These included sugar maple DBH, basal area estimates, and percentages of overstory contributions (a crude BA-weighted shade-tolerance index), mineral and F-H layer pH, and understory beech and sugar maple BA, which were recorded in initial and revised stand inventories (Appendix 1).
3 Results
3.1 Maple BAI decreases after 2 years of summer SWC deficit under beech sapling proliferation
In 2021, rainfall exclusion did not affect the ratio of annual BAI to long-term average basal area increment. Its credible intervals all included zero (Table 3; Figure 1a). In 2022, rainfall exclusion exerted a positive effect on the ratio of annual BAI to long-term average BAI. This response was offset by a greater negative Proliferation*Exclusion interaction, i.e., the moisture deficit under dense understory beech saplings resulted in decreased sugar maple BAI (Table 3; Figure 1b). Tent deployment over two consecutive growing seasons (2021, 2022) produced clear differences not only in manually measured soil water content (SWC) but also in seasonal moisture data obtained from in situ loggers in 2021 and 2022 (Appendix 2). The intensive manual instrument measurements on 1 September 2022 (see Table 4) indicated that rainfall exclusion reduced soil moisture (mean = 0.09 cm3 cm−3 or 8.8%) to 56% of the volume reported in excluder-free plots (mean = 0.15 cm3 cm−3 or 15.6%).
Table 3
| Model parameters | 2021 | 2022 |
|---|---|---|
| Intercept | 0.07 (−0.54, 0.68) | 0.0 (−0.40, 0.41) |
| Rainfall exclusion yes | 0.25 (−0.20, 0.72) | 0.52 (0.17, 0.87) |
| Beech yes | 0.03 (−0.88, 1.00) | 0.06 (−0.58, 0.63) |
| Rain exclusion yes: Beech yes | −0.38 (−1.07, 0.27) | −0.59 (−1.09, −0.10) |
Posterior estimates and their 95% credible intervals of GLMM model theta parameters for rainfall exclusion and beech proliferation effects on basal area increment (BAI) ratios for 2021 and 2022 relative to the long-term average for 2010–2020.
Credible intervals for estimates that are highlighted in bold exclude zero.
Figure 1

Ratio of basal area increments (BAIs) between (a) 2021 and (b) 2022 and long-term averages (2010–2020; 2015–2020) across rainfall-exclusion and beech-proliferation experimental plots, predicted by GLMM regression. Stars represent estimated values, closed circles represent observed values, and error bars indicate 95% credible intervals.
Table 4
| Beech | Excl. | Mean NDI | SD NDI | Lower credible | Upper credible | Nutrient stress |
|---|---|---|---|---|---|---|
| Yes | Yes | 182.35 | 234.65 | 1.98 | 362.72 | Moderate deficiency |
| Yes | No | 260.81 | 373.13 | −26.01 | 547.62 | Major deficiency |
| No | Yes | 472.74 | 510.27 | 80.51 | 861.97 | Major deficiency |
| No | No | 627.25 | 574.58 | 185.60 | 1068.904 | Major deficiency |
Mean (± SD) Nutritional Deficiency Indices (NDI) for summed absolute DRIS values of N, P, K, Mg, and Ca in leaves of 36 sugar maples located in rainfall exclusion (Yes) vs. rainfall-accessible (No) plots, crossed with beech proliferation (Yes) vs. beech-free (No) conditions.
The 95% credible intervals are estimated from DRIS nutrient responses to four treatments (fixed effects).
The 95% credible limits around the estimates shown in boldface type exclude zero.
3.2 Sugar maple foliar nutrient insufficiencies may not be exacerbated by rainfall exclusion
Prior to applying DRIS, foliar nutrients displayed varying degrees of insufficiency relative to their reference values. Descriptive statistics for macronutrient concentrations of mid-canopy leaves from 36 maple trees are summarized in Appendix 3. Accordingly, foliage is severely Mg-deficient; 100% of the trees are below its deficiency threshold. In contrast, only leaves from 11% of the trees were Ca-deficient, with Ca concentrations averaging 60% higher than the element’s deficiency threshold. These two extremes bracket deficiency estimates for P, K, and N based on their respective thresholds.
Mid-summer foliar concentrations (% or g kg−1) of N, P, Mg, and K apparently were not affected by rainfall exclusion. Moreover, similar concentrations were observed in plots with and without beech sapling proliferation (Figure 2; Table 5). Leaf Ca responded differently; foliar Ca levels were, respectively, 79 and 73% higher on rainfall-accessible and rainfall-exclusion plots that were beech-free, relative to those plots where beech saplings proliferated (Figure 2). Only Ca and N were above their sufficiency thresholds across the four treatments, while P and Mg responses consistently remained below threshold values (Appendix 3). For K, only foliar responses for the maples in beech-free plots under rainfall excluders were above their sufficiency threshold.
Figure 2

Leaf nutrients (%) across rainfall-exclusion and beech-proliferation treatments, predicted by GLMM regression. Stars represent estimated values, closed circles represent observed values, and error bars indicate 95% credible intervals. Foliar concentrations (%) are shown for (a) iron, (b) phosphorus, (c) calcium, (d) magnesium, and (e) potassium.
Table 5
| Parameters | Nitrogen | Phosphorus | Calcium | Magnesium | Potassium |
|---|---|---|---|---|---|
| Intercept | 0.53 (0.42, 0.64) | −2.26 (−2.38, −2.14) | 0.15 (−0.08, 0.38) | −3.59 (−4.63, −2.67) | −0.44 (−0.63, −0.26) |
| Rainfall exclusion = yes | 0.07 (−0.062, 0.20) | 0.05 (−0.06, 0.20) | 0.09 (−0.09, 0.27) | −0.17 (−1.01, 0.65) | 0.15 (−0.01, 0.30) |
| Proliferation = yes | 0.04 (−0.12, 0.20) | 0.01 (−0.17, 0.18) | −0.59 (−0.89, −0.26) | 0.36 (−0.89, 1.53) | −0.06 (−0.35, 0.22) |
| Rainfall exclusion = yes: Proliferation = yes | −0.05 (−0.23, 0.13) | 0.06 (−0.04, 0.16) | 0.05 (−0.220, 0.32) | 0.19 (−0.96, 1.36) | −0.16 (−0.38, 0.06) |
| R 2 | 0.37 (0.07, 0.80) | 0.49 (0.23, 0.74) | 0.81 (0.61, 0.96) | 0.56 (0.13, 0.95) | 0.57 (0.23, 0.88) |
Posterior estimates and their 95% credible intervals of GLMM model theta parameters for the effects of rainfall exclusion and beech proliferation on sugar maple leaf nutrients (as percentages).
Credible intervals for estimates in boldface type exclude zero. R2 is the unadjusted coefficient of determination.
Detrended correspondence analysis of the five foliar elements extracted two components, which are presented in Figures 3a,b. DCA1 yields ordination scores that can be tested against alternative Bayesian models including incubation, exclusion, and proliferation, with credible intervals excluding zero; ANOVA models for DCA2 scores had no support (BF10 ≤ 0.578). For foliar ordination scores (DCA1), moderate support is indicated only for proliferation (BF10 = 7.19; error % = 2.33). Leaf Ca appears to drive this separation, given its concentrations are strongly correlated with DCA1 scores (r = 0.932, [0.857, 0.964]). Thus, interpreting DCA1 would appear straightforward: foliar Ca increases moving along the axis of DCA1 from the origin toward the right, first through maples on plots containing beech saplings (Proliferation = Yes), then through beech-free plots (Proliferation = No). Over all treatments, DCA1 is inversely correlated only with N DRIS (r = −0.829, [−0.943, −0.3877]) but not with any of the other DRIS indices. When we considered the two levels of proliferation separately (Appendix 5), DCA1 was more strongly and inversely correlated with N DRIS under rainfall exclusion (r = −0.978, [−1.000, −0.457]), and more strongly correlated (r = −0.959, [−0.988, −0.320]) under rainfall-accessible conditions. Interpretation of DCA2 may not be as simple (Figure 3b), given poor ANOVA results. Yet, scores for DCA2 correlate moderately with foliar Mg (r = 0.685, [0.438, 0.818]). As depicted in Figure 3a, foliar (F) Mg varies among locations (Sites).
Figure 3

Two-dimensional ordination of foliar elements using detrended correspondence analysis. (a) DCA2 scores interpreted as variation among sites in terms of correlations with foliar Mg; (b) sugar maple foliar nutrient scores separated along DCA1 by the effect of beech sapling proliferation; (c) 2021 ordination plot of component scores along PC1 separated by the effect of beech sapling proliferation; (d) PCA scores for 2021 soil supply rates of 11 nutrients, indicated in the associated biplot; (e) 2022 ordination plot of component scores along PC2 separated by the effect of beech sapling proliferation; (f) PCA scores for 2022 soil supply rates of 11 nutrients, indicated in the associated biplot. The first three axes of the 2021 ordination explain 28.3, 24.7, and 12.4% of the variance, respectively. The corresponding three axes for PCA in 2022 explain 38.3, 28.3, and 11%, respectively. PC3 is not depicted for either ordination.
3.3 DRIS indices indicate severe Mg insufficiency and strong Ca sufficiency in beech-free plots
According to their respective DRIS indices (≥ zero), leaf N, P, and K were sufficient across the four treatments (Table 6). N-sufficiency estimates have credible intervals excluding zero, which overlap for higher sufficiency estimates of maple trees in treatments where beech saplings have proliferated. N-sufficiency is lower and similar (overlapping credible intervals) between treatments where beech is absent under canopy maples, despite weak support for Proliferation (BF10 = 2.38; error % = 0.91). No model support is evident for treatment differences in other DRIS indices. For beech-free, rainfall-accessible plots (Proliferation = No; Exclusion = Yes), credible intervals exclude zero (Table 6). Under this treatment combination, Ca is much more than sufficient (170.8), while Mg is very severely deficient (−309). Rainfall-accessible plots that are beech-free are comparably Ca-sufficient or Mg-deficient. When DRIS indices are ranked across the five elements, however, there is only moderate agreement in the ordering among the four treatments (W = 0.445).
Table 6
| Proliferation | Exclusion | Index N | Index P | Index K | Index Ca | Index Mg |
|---|---|---|---|---|---|---|
| Yes | Yes | 19.13 (13.89, 24.37) | 39.36 (−5.40, 84.12) | 22.73 (−18.37, 63.82) | 43.16 (−19.25, 105.56) | −124.37 (−270.59, 21.84) |
| Yes | No | 20.28 (15.36, 25.20) | 31.68 (−4.86, 68.22) | 19.23 (17.81, 56.28) | 14.22 (−6.48, 34.92) | −85.42 (−178.54, 7.70) |
| No | Yes | 6.93 (0.20, 13.66) | 59.65 (6.79, 112.52) | 71.95 (17.81, 126.09) | 170.83 (50.43, 291.23) | −309.36 (−533.31, −85.40) |
| No | No | 9.32 (2.71, 15.93) | 57.39 (−4.18, 118.97) | 43.33 (−3.98, 90.65) | 123.06 (29.05, 217.06) | −233.10 (−431.16, −35.04) |
Posterior estimates and their 95% credible intervals that were obtained from Bayesian ANOVA on DRIS indices for five foliar nutrients in response to fixed effects of beech proliferation (Proliferation) crossed with rainfall exclusion (Exclusion).
Site was considered a random effect in the models. Credible intervals for posterior estimates that are highlighted in boldface exclude zero.
NDI (Nutritional Deficiency Index) ranges more than three-fold across the four treatments (Table 4), from 182.4 (moderate deficiency) in plots where beech proliferated and are covered with excluders (Yes-Yes) to NDI = 200 + (major deficiency) in the three remaining treatments. The latter includes beech proliferation with no rainfall exclusion (Yes-No: 260.8, credible interval includes zero). In beech-free plots, nutrient deficiencies are very high, both in plots covered with excluders (No-Yes: 472.74) and those that are not covered (No-No: 627.25). Despite an increasing deficiency trend observed among the means, there are no differences among the four treatments (BF10 < 1.0).
3.4 Soil nutrient supplies may respond differently to drought depending upon beech proliferation
Regression analysis of the six major soil supply rates (μg 10 cm−2 6 weeks−1), combined across 2021 and 2022, suggests that P and NH4-N increase in the presence of beech, while NO3-N, Ca, and Mg decrease under the same treatment (Appendix 4). GLMM responses also depend upon the effects of rainfall exclusion, as indicated by their model summaries (Table 7). With credible intervals around parameter estimates for P that include zero, its supplies are low in beech-free plots yet increase by 40% where beech has proliferated. Under rainfall exclusion, P supplies are lower than in the rainfall-accessible plots but increase by more than 100% under beech saplings. Phosphate has low mobility in soil, made worse by soil moisture deficits and high spatial variability in the latter’s distribution.
Table 7
| Parameters | NO3-N | P | K | Mg | Ca | NH4-N |
|---|---|---|---|---|---|---|
| Intercept | 3.9 (2.26, 5.29) | 1.26 (−0.25, 2.62) | 4.39 (2.66, 5.71) | 5.01 (4.33, 5.67) | 7.04 (5.83, 8.34) | 1.08 (−1.73, 3.15) |
| Exclusion: yes | −1.36 (−2.37, −0.35) | −1.06 (−2.36, 0.24) | 0.48 (−0.19, 1.06) | −0.29 (−0.73, 0.14) | −0.48 (−1.28, 0.33) | −0.26 (−0.98, 0.42) |
| Prolif.: yes | −0.99 (−1.98, −0.06) | 0.25 (−0.94, 1.44) | 0.54 (−0.11, 1.17) | −0.34 (−0.72, 0.10) | −0.96 (−1.81, 0.11) | 1.58 (0.78, 2.40) |
| Exclusion: yes Prolif.: yes | 1.48 (0.11, 2.97) | 0.66 (−1.20, 2.45) | −0.91 (−1.73, −0.01) | 0.06 (−0.55, 0.65) | 0.12 (−1.04, 1.16) | −0.32 (−1.21, 0.67) |
Posterior estimates and their 95% credible intervals of model theta parameters for the effects of rainfall exclusion and beech proliferation on major soil nutrient pools.
Credible intervals around posterior estimates in boldface exclude zero.
Regardless of exclusion, supplies of NH4-N increase with beech sapling proliferation like P, as supported by 95% credible estimates (Table 7). In the presence of beech, decreases in Ca and Mg are more pronounced under rainfall-accessible conditions than under drought constraints, likely due to greater mobility of these elements. Yet, only their model intercepts provide credible estimates. More complex responses are exhibited by NO3-N, for example. Supplies are highest in rainfall-accessible plots and decline in the presence of beech and rainfall excluders. All parameter estimates for the NO3-N model are bracketed by credible intervals that exclude zero. Credible estimates for the model intercept and the proliferation*exclusion interaction are presented for K (Table 7) and, indeed, mirror the response of this element as depicted in Appendix 4. K increases in the presence of beech, then declines with beech proliferation under rainfall exclusion.
Temporal dynamics of the entire soil nutrient pool (supplies summed over each PRS incubation duration and treatment) were tested separately for 2021 and 2022. The duration of summer 2021 PRS burial was longer at 56 days (incubation: four probe retrievals) compared to 2022 (43 days, two retrievals). Focusing on longer-duration incubations under drought constraints, Exclusion + Incubation (BF10 = 21,194, error% = 1.89) is the overall best-supported model, yet it does not include beech sapling effects on total nutrient pools. Diverging patterns emerge when comparing supplies under rainfall exclusion vs. rainfall accessibility. In rainfall-accessible plots, nutrient supplies eventually reach a plateau (42–56 days) with increasing burial duration, while total nutrient adsorption remains diffusion-dependent under rainfall excluders, with lower means than in the first treatment. The 2021 responses are consistent with initial expectations for beech-free plots. Under beech proliferation (Figure 4), total soil pools in rainfall-accessible plots were about 26% lower than their beech-free counterparts.
Figure 4

Supply rates pooled over nutrients (AllSum) for each PRS retrieval (days) for 2021 and 2022, divided by the absence or presence of beech saplings (No vs. Yes). Error bars associated with the means here and in the following figures represent 95% credible intervals.
Short-term PRS incubations (2022) reveal that final pool sizes (43 days) differ in the absence vs. presence of beech saplings (Figure 4), but not with rainfall exclusion. Like 2021, candidate models (14 of 18) of 2022 responses are moderately to strongly supported, most notably Proliferation + Exclusion + Incubation + Proliferation*Exclusion + Exclusion*Incubation (BF10 = 401, error% = 4.43). Final total pools in beech-free, rainfall-accessible plots (No-No) are largest (mean 1,583 μg 10 cm−2 6 weeks−1 [589, 2,576]; Figure 4), while the smallest final pools (mean 549 μg 10 cm−2 6 weeks−1 [478, 619], Figure 4) were in plots where beech proliferated under excluders (Yes-Yes). Given limited numbers and durations of burials, we cannot ascertain whether final incubations have achieved equilibrium-dependent adsorption/desorption of the ion-exchange membranes.
Nutrients retained for 2021 ANOVAs exclude P, K, NO3-N, or S, as their responses exhibit weak support for Incubation (main effect BFincl ≤ 1.55) or its interactions. Given K’s weak support, we did not subject base cation sums (Ca + Mg + K) to more thorough analysis, despite strong support for Incubation provided by Mg and Ca (main effect BFincl: 52446 and 115.5, respectively). Other analyses including Incubation, which target individual nutrients or small groups, may include many competing candidates with credible intervals bracketing zero. The second grouping of nutrients among soil supplies that we consider is Al + Fe + Mn (acidity). In 2021, main effects of incubation on acidity within individual models are strongly supported for their components as follows: Al (BFincl = 1,548), Fe (BFincl = 495), and Mn (BFincl = 405). The strongest model among 13 candidates is Proliferation + Exclusion + Incubation + Proliferation*Incubation (BF10 = 505,038, error% = 2.36), which shows neither pronounced main effects of rainfall exclusion nor its interactions (Figure 5). Acidity supply rates are low, but these steadily increased with burial duration under beech sapling proliferation, while remaining constant and low under beech-free conditions.
Figure 5
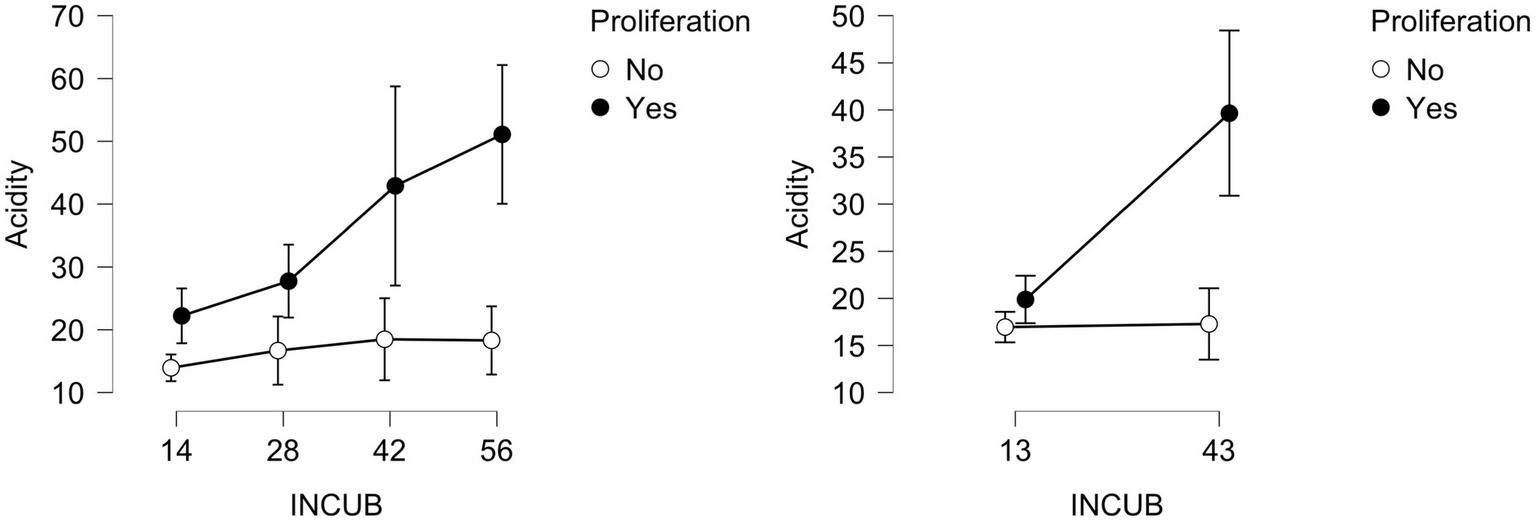
Acidity (Al + Fe + Mn) for each PRS retrieval (days) for 2021 and 2022, divided by the absence or presence of beech saplings (No vs. Yes).
In 2022, main effects and interaction of proliferation and incubation dominate acidity responses (BF10 = 4,626, error% = 2.94). Only Proliferation-Exclusion (Yes-Yes) incubations can be distinguished among the eight treatment means, given the otherwise frequent overlap of credible intervals among treatments: day 13, 19.40 [17.03, 21.77]; day 43, 42.23 [36.19, 48.28]. Day 43 PRS incubations produce trends similar to 2021: highest acidity under beech saplings and lowest acidity when beech-free (Figure 5). These trends are consistent with observations regarding bulk pH of mineral soil (0–15 cm) and Fermentation-Humification (F-H) layer of the 12 plots (Table 2). Similar trends are not reported for base cation pools. ANOVA of base cations (Ca + Mg + K) is not included, but their collective supply rates are inversely correlated with acidity (r = −0.336 [−0.511, −0.123]; BF10 = 13.4). When focusing on acidity vs. base cation responses under rainfall exclusion conditions, the correlation is −0.482 ([−0.719, −0.084]; BF10 = 3.7).
NH4-N pools are high at day 14 (2021) under beech saplings and progressively decrease with increasing burial duration. Fourteen of 18 candidate models are strongly supported (BF10: range = 138.2–83,901; error% = 0.52–14.82), with Proliferation + Exclusion + Incubation + Proliferation*Exclusion + Exclusion*Incubation having best support. Yet, of 16 treatment means estimated for NH4-N supplies, credible intervals for seven values include zero. Time-courses of NH4-N indicate a decline in 2021, very weakly so in beech-free plots compared to those where beech was profuse (Figure 6). Rank-ordering of NH4-N among the four treatments is fairly consistent over four PRS retrievals (Kendall’s W = 0.775).
Figure 6
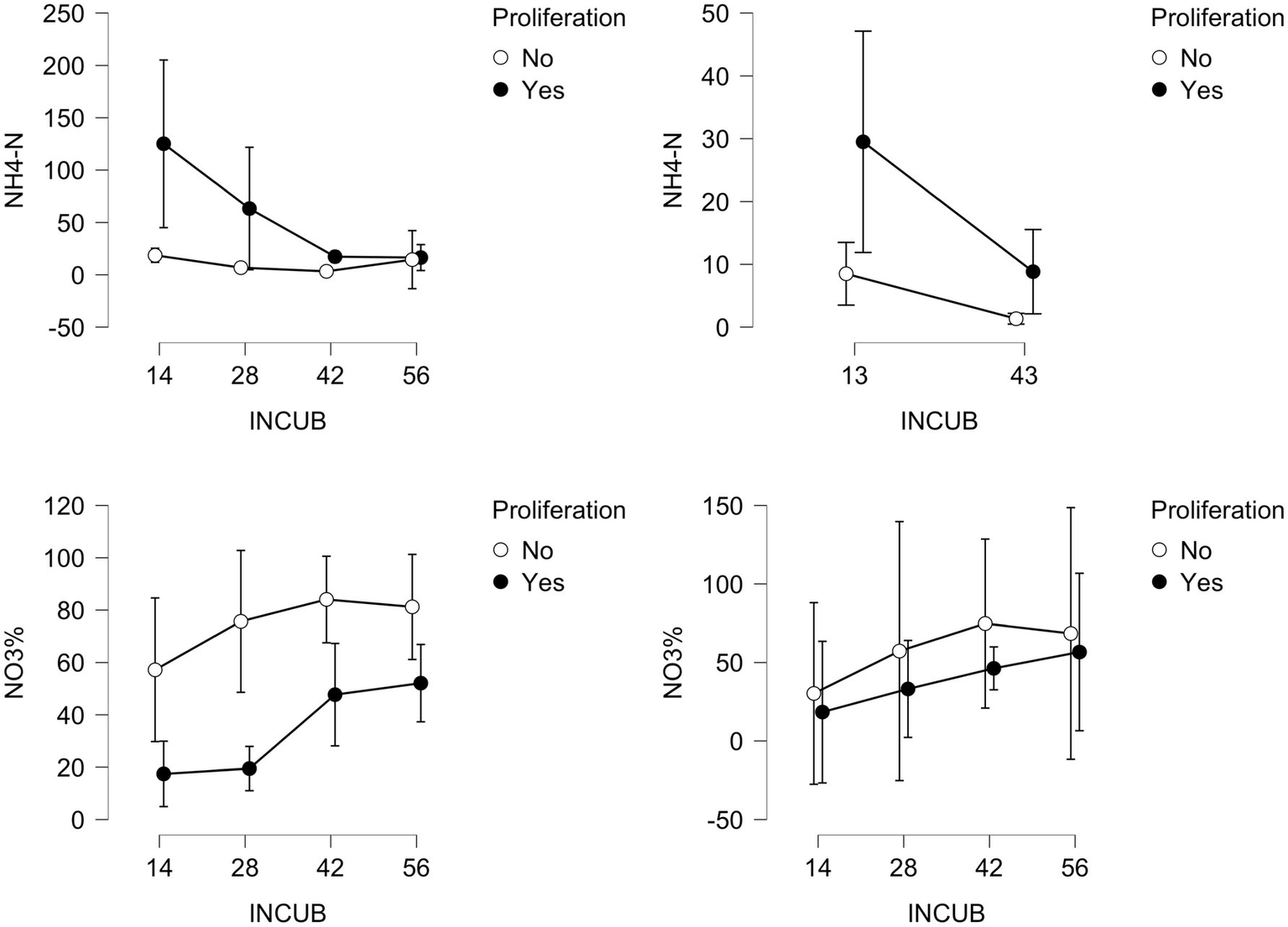
NH4-N supply rates for each PRS retrieval (days) for 2021 and 2022 (upper left panel and upper right panel, respectively), divided by the absence or presence of beech saplings (No vs. Yes). The lower panels depict nitrate % of inorganic-N pools in 2021, both in the absence and presence of rainfall excluders.
In 2022, the two NH4-N incubation durations ranked over four treatments are in perfect agreement (W = 1.00), not a robust result. Without enforced moisture deficits, one consistent feature of NH4-N responses that is shared between 2021 and 2022 is lower supply rates in beech-free compared to beech-proliferated plots (Figure 6). Another feature is the decreasing NH4-N pools over time, as well as those of total inorganic-N (not shown), which shows an increasing percentage of NO3-N (nitrate%) with time. In beech-free soils (Figure 6), nitrate constitutes 49% of inorganic-N pools at day 14, peaking at 84% by day 42, before falling to about 73% by day 56; under beech proliferation (Figure 6), nitrate progresses from 14 to 58% of inorganic-N over the entire burial duration in 2021. In 2022, final incubation pools of nitrate are 65 and 90% of inorganic-N in profuse-beech plots and beech-free plots, respectively.
Principal component analysis (PCA) extracted three axes from the 11 soil nutrient pools that explain 65.4 and 77.6% of the variance for the 2021 and 2022 datasets, respectively. Means of the PC1 and PC2 scores for the same components are summarized in Table 2 for further correlation analyses. In 2021, exchange membrane ion compositions steadily changed over the course of PRS burial in the presence of beech saplings but remained low and constant where beech was absent (Figure 7). Element pool composition progressively changed over the incubations before plateauing under the effects of rainfall exclusion.
Figure 7
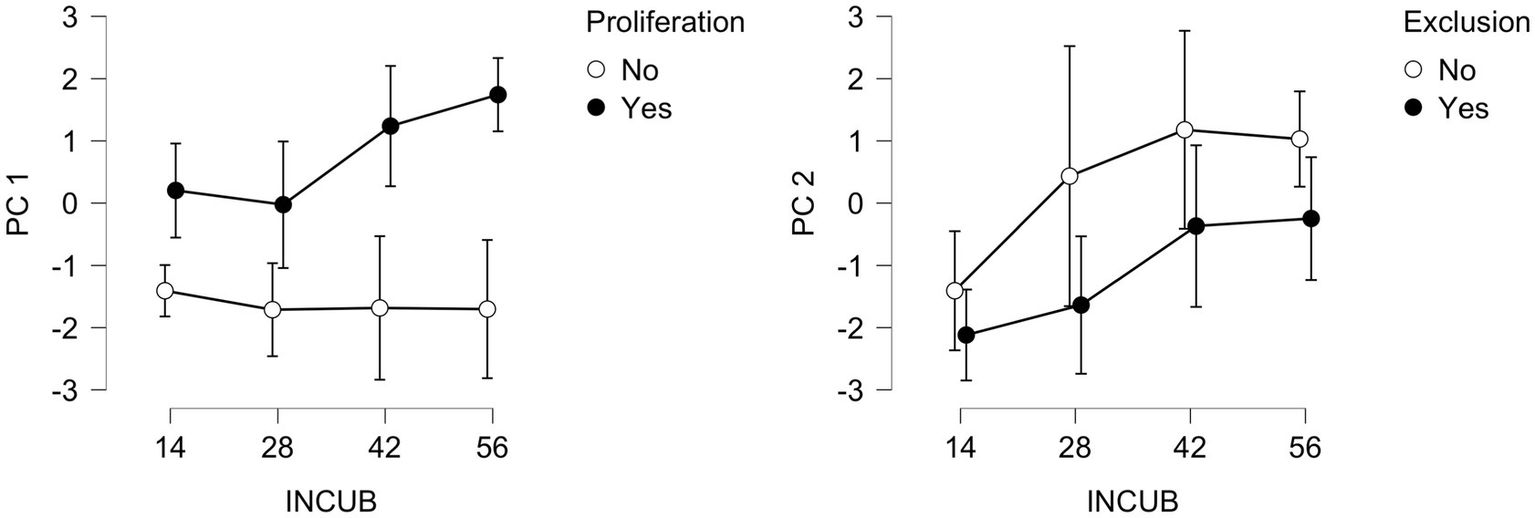
Principal component scores on PC1 and PC2 for each PRS retrieval (days) in 2021, divided by the absence or presence of beech saplings (No vs. Yes) or by exclusion (No vs. Yes). Apart from increased mean values over time, no other definitive trends are present in the 2022 data (not shown).
Ordination biplots (Figures 3d,f) indicate that trace elements and several macronutrients (P, K, S) tend to cluster closely together, while NO3-N and NH4-N remain isolated. In 2021, six nutrients loaded onto PC1 (28.3% variance explained). In 2022, seven nutrients loaded onto PC1 (38.3% variance explained). According to their loadings (not shown), five elements (K, Fe, Mn, Zn, Al) were associated with PC1 in both years, while four elements (NO3-N, Mg, P, S) were associated with PC2 in both years (respectively 24.7 and 28.3% of variance explained), information that we can subsequently use to interpret gradients along the axes (Appendix 5). According to the ordination scores, soil nutrient ensembles in both years vary considerably in terms of their compositions. Yet, they clearly separate along PC1 in 2021 (Figure 3c), according to proliferation and the correlation with acidity (r = 0.845, [0.736, 0.903]). In 2022, scores separate along PC2 with respect to beech sapling proliferation (Figure 3e). In terms of trends over time, means and credible intervals for PC1 and PC2 exhibit progressively increasing values from day 14 to day 56 of the PRS incubations (Figure 7). For PC2, ordination scores are strongly correlated with P (r = 0.532, [0.302, 0.688]) and S (r = 0.878, [0.789, 0.924]). Separation of 2022 scores is not as clear-cut, but PC1 is strongly correlated with acidity (r = 0.888, [0.720, 0.948]) and its constituents, together with Zn (r = 0.812, [0.567, 0.909]) and S (r = 0.828, [0.598, 0.917]). In 2022, PC2 is strongly correlated with NO3-N (r = 0.878, [0.789, 0.924]), Ca (r = 0.964, [0.902, 0.984]), and Mg (r = 0.837, [0.616, 0.922]).
In examining the consistency of ranked CVs (not shown) for SWC and PC1 and PC2, agreement between moisture variability among plots and PC1 for 2021 and 2022 is moderate (W = 0.514). While agreement is lower, it is still moderately consistent (W = 0.428) for ranked CVs of SWC and PC2 in 2021 and 2022. Components are based on 11 nutrients (except Trace) with different mobilities, but the trends observed in these latter CV results are consistent with our hypothesis. Where beech is profuse and under rainfall exclusion (Yes-Yes), overall supply rates in their plots have more variable responses; beech-free plots under rainfall exclusion (No-Yes) are the least variable. Variation associated with rainfall exclusion (high CVs) vs. rainfall accessibility (low CVs) is very consistent among treatments across sites (Very good agreement: W = 0.822). When variation (CVs) in the 11 individual nutrients (plus SWC CVs) is ranked among the four treatments and their consistency assessed, there is only fair agreement among the treatments in both 2021 (W = 0.257; No-No < No-Yes = Yes-No <Yes-Yes) and 2022 (W = 0.392; No-Yes < No-No < Yes-No < Yes-Yes).
3.5 Uncoupling of foliar and soil nutrient pools may be related to soil moisture deficiency
Among the correlations performed between foliar and soil ordination scores, along with other variables (Appendix 5), we obtained 26 estimates with support at BF10 ≥ 5 under rainfall excluders (Exclusion: Yes) and 36 estimates with support in the absence of excluders (Exclusion: No). Given the small sample sizes for each estimate, we imposed a more stringent criterion of BF10 ≥ 10, reducing credible correlations to 13 and 15 under excluders vs. rainfall-accessible conditions, respectively. Under excluders, soil moisture deficits and spatial variability of SWC (see previous section), the lowest correlation relative to the revised criterion is r = 0.942 for soil PC1 (2022) vs. beech sapling basal area. This correlation suggests that variation in beech sapling density strongly controls the composition of the soil nutrient supply ensemble. The highest correlations were exhibited by the relationships Ca_2021 vs. Ca_2022 (r = 0.991), and Ca-DRIS vs. sugar maple DBH (r = 0.990). In the absence of rainfall excluders, the lowest and highest correlations were r = 0.942 for Ca_2021 vs. soil nitrate percentage in 2021, and r = 0.987 (Appendix 5). The lower and upper limits of their absolute values bracket eight strongly supported correlations between the above- and below-ground variables for the two levels of rainfall exclusion. Six and seven strong correlations were observed between two soil nutrient variables in the rainfall-exclusion plots and rainfall-accessible plots, respectively.
4 Discussion
4.1 Sugar maple BAI decreases with soil moisture deficits where understory beech proliferates
We investigated the effects of two consecutive growing seasons of enforced soil-moisture deficit using rainfall excluders in maple-dominated northern hardwood forests within southern Quebec (Canada). Excluder-covered plots (Exclusion: Yes) and excluder-free controls (Exclusion: No) were assigned to paired plots within maple stands with a dense beech sapling understory (Proliferation: Yes) vs. those that were relatively beech-sapling free (Proliferation: No). It was under these four treatment combinations that we assessed sugar maple BAI, foliar nutrition, and soil nutrient supplies.
Two consecutive growing seasons of experimental drought elicited a small positive effect of rainfall exclusion on the ratio of annual BAI to the long-term average BAI, but this was offset by beech sapling proliferation, which incurred a net negative BAI ratio response. Successful early detection of a maple growth decrement could be attributed not only to the methods of analysis that we used but also to the cumulative effects of moisture deficits and pre-existing foliar nutrient stress across all sites (see below). Our earlier prediction that proliferation and its interaction with rainfall exclusion would exert no effect on BAI was also refuted. Indeed, the responses we measured suggest that combinations of beech proliferation and rainfall exclusion treatments could be ordered from smallest to greatest decreases in mean BAI ratios as Yes-Yes ≤ No-No < Yes-No < No-Yes.
In 2022, the basal area increment ratio of sugar maple was highest in the absence of beech sapling proliferation (No), in the presence of rainfall exclusion (Yes), and lowest in the presence of both beech (Yes) and rainfall exclusion (Yes). There may be a simple explanation for the observed ordering, rather than the expected one where moisture stress effects should take precedence: Yes-Yes < No-Yes < Yes-No < No-No. A concurrent greenhouse study (Collin et al., 2025) found that maple seedling performance improved when beech litter mulch over mineral soil mitigated drought effects, i.e., soil moisture was three-fold higher under beech litter compared to in its absence. Unlike mature sugar maple or its saplings, which may resist moisture deficits through drought-deciduous responses, including leaf abscission and stomatal closure (Pallardy and Rhoads, 1993; Filewod, 2011), its seedling stage is very sensitive to soil moisture deficiencies given the lack of an extensive root system.
A lack of agreement between the two aforementioned rankings may underscore certain shortcomings or complexities in the experimental layout. For one, the selection of sites that would isolate the effects of understory beech from those of canopy beeches is a concern. Selecting sugar maple stands that contain only canopy beeches or only beech saplings may be extremely challenging since highly shade-tolerant beech seedlings can recruit under canopy beech through seed masting (short-distance dispersal) and long-distance animal dispersal (Tubbs and Houston, 1990), while root suckering can potentially be initiated under mature beeches with the onset of beech bark disease and individual tree decline and death. Second, we did not include plot sapling densities or record their spatial distributions, nor did we note pathogen/pest damage that would indicate the origins of these recruits (Reed et al., 2022), nor did we organize saplings into narrower size classes (i.e., 2–4 cm, 5–9 cm). A third concern is the complexity that natural stand dynamics can impose upon certain treatment responses or vice versa, particularly in our plots containing overstory and understory beech. The presence of shade-intolerant species in the overstory of these plots, such as red maple (A. rubrum), black cherry (Prunus serotina), and big-tooth aspen (Populus grandidentata), is likely the result of previous canopy gap disturbance, a condition that is not evident in plots that are beech sapling-free and that may encompass both exclusion treatments (Appendix 1).
We did not assess possible beech bark disease (BBD) damage, but the presence of several late-successional, shade-intolerant species in the canopy may represent the intersection of several powerful but conflicting drivers. A “perfect storm” of conditions likely emerges from an unusual convergence of (a) pathogen-driven overstory beech decline, (b) climate change-induced drought stress affecting maple vigor, (c) the state of stand base saturation and base cation availability, and (d) what we might refer to as co-opting of available soil moisture by dense mono-specific understory communities, all of which can conspire to hasten gap formation (Lawrence et al., 2018; Tourville et al., 2023; Rasoanaivo et al., 2024; Zarfos et al., 2024).
4.2 Rainfall exclusion does not necessarily exacerbate pre-existing foliar deficiencies
Of the 36 trees from which leaves were sampled, 100% had Mg concentrations below the reference value, while foliar deficiencies for P, K, N, and Ca strongly declined (Appendix 3). Despite apparent deficiencies, only leaf N was considered sufficient according to its DRIS index across all four treatments, barely so in plots where beech had not proliferated (DRIS N < 0). Of 20 DRIS indices had been calculated for the five foliar nutrients across the four treatments, only half of these estimates excluded zero from their credible estimates. What emerges from this incomplete evidence is that foliar Ca remains more than sufficient and that severity of Mg deficiency is further exacerbated by rainfall exclusion, even when beech saplings are not present, thereby confirming our initial prediction of drought stress increasing insufficiency.
The imposition of about 90 days of drought stress did not decrease the actual concentrations of the individual foliar nutrients. Our results contrast with those of Urbina et al. (2015), who observed decreases in K, N, P, Mg, and S, following 2, 6, and 12 weeks of consecutive drought. The contexts of our respective experiments differ: one species versus plant functional groups; two consecutive growing seasons of drought stress (undefined in terms of water potential) vs. consecutive years of severe drought; and DRIS analyses vs. stoichiometric shifts. Collectively, maples in the four treatments could be ordered according to their NDI in terms of increasing foliar deficiency as follows: Yes-Yes < Yes-No < No-Yes < No-No. In what be interpreted as a counter-intuitive response, foliage was rated most severely deficient in plots that were not subjected to rainfall exclusion, which contained no beech (No-No), indicating maple growth that is already experiencing nutrient stress. The least (moderately) deficient maple foliage was sampled from plots at the other treatment extreme, i.e., seasonal rainfall exclusion in plots containing beech saplings (Yes-Yes).
4.3 Rainfall exclusion diminishes many soil nutrient supply rates with increasing PRS incubation
In quantifying the effect of rainfall exclusion and beech proliferation on soil nutrients (Objective 3), we found that the GLMM of six major nutrient pools indicated that most means decreased with rainfall exclusion (except for K) in the absence of beech saplings. This response confirms preliminary expectations. Yet, some nutrients had higher mean concentrations in the presence of beech, i.e., NH4-N, P, and K, particularly where rainfall excluders were absent. Means subsequently decreased with seasonal rainfall exclusion, based on pooled 2021 and 2022 PRS probe data.
The greatest variation in nutrient supply rates (i.e., largest credible intervals) was exhibited by NO3-N, Ca, and Mg in uncovered, beech-sapling-free plots. The converse was true in the remaining treatment combinations for these nutrients (smaller credible intervals). Beech proliferation exerted opposite effects (large credible intervals) on P, K, and NH4-N, regardless of rainfall exclusion conditions. Sustained pool sizes may result from the accumulation of low mobility charged species such as PO4−3 and NH4+, which was greatly exacerbated by reduced soil water content (Gessler et al., 2017; Schönbeck et al., 2020). Furthermore, pooling variables aided in our understanding of nutrient ensemble dynamics during PRS incubations, particularly in relation to PRS burial duration.
Given the large overlapping credible intervals, we must view these and other mean responses with caution. For example, the summing of nutrient pools encompasses elements with very different mobilities in the soil, which vary strongly under different soil moisture conditions. Installed PRS probes were not surrounded by root-exclusion collars; therefore, we cannot discount ion competition between the latter and the probes. Nutrient supply rates were collectively smaller in the plots where beech proliferated than in those that were beech-free. Regardless of beech presence or absence, PRS probes apparently exhibited diffusion-dependent adsorption over increasing 2021 burial durations (linear increases or decreases), while only increasing element supplies exhibited equilibrium-dependent behavior, i.e., achieved a plateau by the end of the incubations when beech was absent and rainfall was freely available. In 2022, PRS had low and high supply rates at the beginning and end of the incubation period. Inorganic-N supplies declined linearly over time; by the end of the incubations, NO3-N dominated this pool, but only in the absence of beech. Decreased nitrate in the presence of beech could suggest a decrease in endomycorrhizal dominance of the root systems.
We confirmed that beech is associated with decreased available base cations in the PRS probe assays. These relationships could result from strong competition for nutrients and water, yet may simply reflect the preferential growth and development of beech seedlings or suckers in micro-sites that are unfavorable to sugar maple establishment. Effects of beech allelopathy and competition through rapid suckering likely contribute to the suppression of other understory vegetation (Tubbs and Houston, 1990; Boakye et al., 2023). The responses are consistent with literature reports (e.g., Dijkstra et al., 2001; Collin et al., 2017). Depletion of base cations, by whatever means, would result in the mobilization of Al + Fe + Mn. Indeed, 2021 incubations indicated that sequential removals of probes in the excluder-free plots that contained no beech had base cation supplies (not shown) that were consistently two- to three-fold higher than in plots where beech was present. Limited PRS results for 2022 confirm that exchangeable base cation supplies were 66% lower under beech proliferation in 43-day incubations conducted in excluder-free plots while decreasing by 53% in plots containing excluders.
4.4 Uncoupling of foliar and soil nutrients with rainfall exclusion
Concurrent monitoring over two consecutive summers indicated that plot soil moisture under rainfall excluders was about half the volumetric moisture content of soils in plots that were accessible to rainfall inputs. While these estimates are useful, neither soil bulk density nor stoniness in the six sites were determined to confirm the validity of the TDR estimates, nor was soil moisture linked to estimates of water potential for the soil or sugar maples. Direct estimates of plant-water relations, soil moisture storage capacity, soil saturation, and moisture retention were not made; these parameters are necessary for the construction of a stand-hydrologic model, but these aims are beyond the scope of the present study (e.g., Köcher et al., 2013; Yu et al., 2019; Harrison et al., 2020).
Drought is accompanied by decoupling among soil nutrient cycles (Dannenmann et al., 2016; Liu et al., 2023). Under our experimental drought regime, analyses reveal that among the 12 soil nutrients selected for the 2 years and correlated by excluder level (Yes vs. No), each resulted in 66 possible pairwise combinations. Five strongly supported correlations (BF10 ≥ 5) indicated sustained nutrient coupling under rainfall exclusion, while there were six such strong associations present when excluders were absent. These results suggest uncoupling of elements; under a less stringent criterion (BF10 > 3.0), 21 and 34 correlations emerged that were weakly to strongly correlated under the same treatments, respectively rainfall exclusion and rainfall-accessible conditions. Neither analysis, however, includes assessments of soil C. Moreover, the artificial drought conditions imposed on sugar maple in this study were very short-term, contrasting with many experimental durations (e.g., 11 years, Evans and Burke, 2013), which have been conducted in arid ecosystems (e.g., Liu et al., 2023; but see Guillén et al., 2022).
5 Conclusion
Sugar maple stands in the northern hardwood forest are predisposed to decline, as evidenced by foliar nutrient deficiency (high DRIS indices), together with extensive understory recruitment of American beech suckers and saplings. Consequently, both foliar element ensembles and soil nutrient compositions that we studied are modified by American beech sapling proliferation and by drought conditions induced by rainfall excluder installations: interaction is present between the two treatment factors. Manipulations of SWC over two consecutive summers were sufficient to decrease annual basal area increment (BAI) and further shift soil pools and processes from endomycorrhizal to ectomycorrhizal dominance, or to possible disruption and decoupling of soil nutrient cycles.
Given its endomycorrhizal rooting habit, sugar maple supplies many ecosystem services (ES) and economic benefits (Godman et al., 1990), which ECM species such as yellow birch, hickory, or beech especially cannot or do not necessarily provide, i.e., inorganic-N rather than an organic-N-based economy, more rapid nutrient cycling, and base cation-rich litter, together with providing high-value timber and syrup production (Midgley and Sims, 2020; Tourville et al., 2023). With increasing beech, the transition from a belowground community dominated by endomycorrhizae to one dominated by ectomycorrhizae reinforces conditions conducive to maple disappearance and its replacement by other canopy dominants that share similar life history characteristics (Phillips et al., 2013), including beech, and which exert their own effects on stand moisture dynamics (Harrison et al., 2020).
The loss of sugar maple is likely to be exacerbated by prolonged drought stress, whereas beech, as a competitor under conditions of moisture deficit, is more flexible in terms of its acquisition, allocation, and redistribution of available water stores among tissues and between size classes (Köcher et al., 2013; Rasoanaivo et al., 2024). In contrast, maple has demanding habitat (narrow) requirements to maintain growth performance and is particularly sensitive to environmental stresses, including nutrient imbalances, acidity, and soil moisture deficits (Bal et al., 2015); its gradual decline and replacement by other highly shade-tolerant species, particularly beech, has been documented in many studies. American beech establishment in maple stands through root suckering or mast-seeding of beech nuts and seedling recruitment would result in understory sapling proliferation. This process could exert profound effects on soil carbon storage and biogeochemical cycles of other nutrients, particularly in the face of prolonged drought, although to our knowledge, no study has yet addressed this question.
Statements
Data availability statement
The data supporting the conclusions of this article will be made available by the authors upon reasonable request.
Author contributions
WP: Formal analysis, Writing – original draft. CG: Conceptualization, Investigation, Methodology, Writing – review & editing. TS: Formal analysis, Writing – review & editing. AM: Conceptualization, Funding acquisition, Project administration, Supervision, Writing – review & editing. DR: Conceptualization, Funding acquisition, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was financially supported by the Ministère des Ressources Naturelles et des Forêts du Québec (MRNF: #3329–2019-142332178) and the Natural Sciences and Engineering Research Council of Canada (NSERC: ALLRP 558271–2020).
Acknowledgments
We thank Liane Nowell and the Kenauk Institute (Montebello, QC) for facilitating this research project and granting access to field sites. We also thank Simon Lebel Desrosiers (Laboratoire iForêt, TELUQ) for conducting XRF analyses, Benoit Gendreau-Berthiaume (ISFORT) for conducting the cross-dating of wood cores, and Pierrick Romain Arnault for updating the forest stand inventories. We thank the editor and the reviewers for their extensive comments ‘that greatly improved this paper.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2025.1518704/full#supplementary-material
Footnotes
1.^ https://cfs.nrcan.gc.ca/terms/read/996
References
1
Bal T. L. Storer A. J. Jurgensen M. F. Doskey P. V. Amacher M. C. (2015). Nutrient stress predisposes and contributes to sugar maple dieback across its northern range: a review. Forestry88, 64–83. doi: 10.1093/forestry/cpu051
2
Beaufils E. R. (1973). Diagnosis and recommendation integrated system (dris), a general scheme of experimentation and calibration based on principles developed from research in plant nutrition. Pietermaritzburg, South Africa: University of Natal.
3
Bishop D. A. Beier C. M. Pederson N. Lawrence G. B. Stella J. C. Sullivan T. J. (2015). Regional growth decline of sugar maple (Acer saccharum) and its potential causes. Ecosphere6, 1–14. doi: 10.1890/ES15-00260.1
4
Boakye E. A. Bergeron Y. Drobyshev I. Beekharry A. Voyer D. Achim A. et al . (2023). Recent decline in sugar maple (Acer saccharum marsh.) growth extends to the northern parts of its distribution range in eastern Canada. For. Ecol. Manag.545:121304. doi: 10.1016/j.foreco.2023.121304
5
Bose A. K. Weiskittel A. Wagner R. G. (2017a). Occurrence, pattern of change, and factors associated with American beech-dominance in stands of the northeastern USA forest. For. Ecol. Manag.392, 202–212. doi: 10.1016/j.foreco.2017.03.003
6
Bose A. K. Weiskittel A. Wagner R. G. (2017b). A three decade assessment of climate-associated changes in forest composition across the North-Eastern USA. J. Appl. Ecol.54, 1592–1604. doi: 10.1111/1365-2664.12917
7
Boulanger Y. Taylor A. R. Price D. T. Cyr D. McGarrigle E. Rammer W. (2017). Climate change impacts on forest landscapes along the Canadian southern boreal forest transition zone. Landsc. Ecol.32, 1415–1431. doi: 10.1007/s10980-016-0421-7
8
Brunner I. Herzog C. Dawes M. A. Arend M. Sperisen C. (2015). How tree roots respond to drought. Front. Plant Sci.6, 1–16. doi: 10.3389/fpls.2015.00547
9
Bürkner P.-C. (2017). Brms: an R package for Bayesian multilevel models using Stan. J. Stat. Softw.80, 1–28. doi: 10.18637/jss.v080.i01
10
Cale J. A. Teale S. A. Johnston M. T. Boyer G. L. Perri K. A. Castello J. D. (2015). New ecological and physiological dimensions of beech bark disease development in aftermath forests. For. Ecol. Manag.336, 99–108. doi: 10.1016/j.foreco.2014.10.019
11
Cholet C. Houle D. Sylvain J.-D. Doyon F. Maheu A. (2022). Climate change increases the severity and duration of soil water stress in the temperate forest of eastern North America. Front. For. Glob. Change5:879382. doi: 10.3389/ffgc.2022.879382
12
Cleavitt N. L. Battles J. J. Fahey T. J. van Doorn N. (2021). Disruption of the competitive balance between foundational tree species by chronic multiple interacting stressors in a northern deciduous temperate forest. J. Ecol.109, 2754–2768. doi: 10.1111/1365-2745.13687
13
Coble M. P. Vadeboncoeur M. A. Berry Z. C. Jennings K. A. McIntire C. D. Campbell J. L. et al . (2017). Are northeastern U.S. forests vulnerable to extreme drought?Ecol. Process.6:34. doi: 10.1186/s13717-017-0100-x
14
Collin A. Ghotsa Mekontchou C. Maheu A. Nolet P. Sotelo F. Rivest D. (2025). The presence of American beech litter can alter the growth response of sugar maple seedlings to drought. Ecosphere16:103. doi: 10.1002/ecs2.70103
15
Collin A. Messier C. Kembel S. W. Bélanger N. (2017). Low light availability associated with American beech is the main factor for reduced sugar maple seedling survival and growth rates in a hardwood forest of southern Quebec. Forests8:413. doi: 10.3390/f8110413
16
Copenheaver C. A. Shumaker K. L. Butcher B. Hahn G. E. Perkins L. J. Dukes C. J. et al . (2020). Dendroclimatology of sugar maple (Acer saccharum): climate-growth response in a late-successional species. Dendrochronologia63:125747. doi: 10.1016/j.dendro.2020.125747
17
CPVQ (1997). Méthodes d’analyse des sols, des fumiers et des tissus végétaux. Quebec: CPVQ.
18
Dannenmann M. Bimüller C. Gschwendtner S. Leberecht M. Tejedor J. Bilela S. et al . (2016). Climate change impairs nitrogen cycling in European beech forests. PLoS One11:e0158823. doi: 10.1371/journal.pone.0158823
19
Dijkstra F. A. Geibe C. Holmström S. Lundström U. S. van Breemen N. (2001). The effect of organic acids on base cation leaching from the forest floor under six north American tree species. Eur. J. Soil Sci.52, 205–214. doi: 10.1046/j.1365-2389.2001.00369.x
20
Duchesne L. Ouimet R. (2009). Present-day expansion of American beech in northeastern hardwood forests: does soil base status matter?Can. J. For. Res.39, 2273–2282. doi: 10.1139/X09-172
21
Duchesne L. Ouimet R. Houle D. (2002). Basal area growth of sugar maple in relation to acid deposition, stand health, and soil nutrients. J. Environ. Qual.31, 1676–1683. doi: 10.2134/jeq2002.1676
22
Elenitsky L. M. Walters M. B. Farinosi E. J. (2020). Tree regeneration structure following beech bark disease-motivated harvests: factors associated with patterns and management implications. Forests11:180. doi: 10.3390/f11020180
23
Evans S. E. Burke I. C. (2013). Carbon and nitrogen decoupling under an 11-year drought in the shortgrass steppe. Ecosystems16, 20–33. doi: 10.1007/s10021-012-9593-4
24
Filewod B. A. (2011). Effects of an early season heat wave on ecophysiological parameters related to productivity in sugar maple (Acer saccharum marsh.). Toronto, ON: University of Toronto.
25
Fritts H. C. (2012). Tree rings and climate. London: Academic Press.
26
Gessler A. Schaub M. McDowell N. G. (2017). The role of nutrients in drought-induced tree mortality and recovery. New Phytol.214, 513–520. doi: 10.1111/nph.14340
27
Godman R. M. Yawney H. W. Tubbs C. H. (1990). “Sugar Maple” in Silvics of North America: 2. Hardwoods, Agriculture Handbook 654. eds. BurnsR. M.HonkalaB. H. (Washington DC: USDA Forest Service).
28
Grams T. E. E. Hesse B. D. Gebhardt T. Weikl F. Rötzer T. Kovacs B. et al . (2021). The Kroof experiment: realization and efficacy of a recurrent drought experiment plus recovery in a beech/spruce forest. Ecosphere12:e03399. doi: 10.1002/ecs2.3399
29
Greenacre M. (2021). Compositional data analysis. Annu. Rev. Stat. Appl.8, 271–299. doi: 10.1146/annurev-statistics-042720-124436
30
Grimond L. Rivest D. Bilodeau-Gauthier S. Khlifa R. Elferjani R. Bélanger N. (2024). Novel soil reconstruction leads to successful afforestation of a former asbestos mine in southern Quebec, Canada. New For.55, 477–503. doi: 10.1007/s11056-023-09989-3
31
Guillén L. A. Brzostek E. McNeil B. Raczka N. Casey B. Zegre N. (2022). Sap flow velocities of Acer saccharum and Quercus velutina during drought: insights and implications from a throughfall exclusion experiment in West Virginia, USA. Sci. Total Environ.850:158029. doi: 10.1016/j.scitotenv.2022.158029
32
Hangs R. D. Greer K. J. Sulewski C. A. (2004). The effect of interspecific competition on conifer seedling growth and nitrogen availability measured using ion-exchange membranes. Can. J. For. Res.34, 754–761. doi: 10.1139/x03-229
33
Harrison J. L. Reinmann A. B. Maloney A. S. Phillips P. Juice S. M. Webster A. J. et al . (2020). Transpiration of dominant tree species varies in response to projected changes in climate: implications for composition and water balance of temperate forest ecosystems. Ecosystems23, 1598–1613. doi: 10.1007/s10021-020-00490-y
34
Henry C. R. Walters M. B. (2023). Tree species size class patterns portend compositional shifts and low resilience in managed northern hardwood forests. Ecosphere14:e4621. doi: 10.1002/ecs2.4621
35
Johnson V. E. Pramanik S. Shudde R. (2023). Bayes factor functions for reporting outcomes of hypothesis tests. Proc. Natl. Acad. Sci. USA120:e2217331120. doi: 10.1073/pnas.2217331120
36
Kish R. James P. M. A. Mariani R. O. Schurman J. S. Thomas S. C. Young E. N. et al . (2022). Beech bark disease in an unmanaged temperate forest: patterns, predictors, and impacts on ecosystem function. Front. For. Glob. Change5:831663. doi: 10.3389/ffgc.2022.831663
37
Köcher P. Horna V. Leuschner C. (2013). Stem water storage in five coexisting temperate broad-leaved tree species: significance, temporal dynamics and dependence on tree functional traits. Tree Physiol.33, 817–832. doi: 10.1093/treephys/tpt055
38
Lajoie P. G. . (1967). Étude pédologique des comtés de Hull, Labelle et Papineau (Québec). Available online at: https://sis.agr.gc.ca/siscan/publications/surveys/pq/pq25/pq25_report.pdf (Accessed September 1, 2024).
39
Lawrence G. B. McDonnell T. C. Sullivan T. J. Dovciak M. Bailey S. W. Antidormi M. R. et al . (2018). Soil base saturation combines with beech bark disease to influence composition and structure of sugar maple-beech forests in an acid rain-impacted region. Ecosystems21, 795–810. doi: 10.1007/s10021-017-0186-0
40
Leak W. B. Yamasaki M. Holleran R. (2014). Silvicultural guide for northern hardwoods in the northeast. USDA Forest Service. Newtown Square, PA: Northern Research Station.
41
Leduc F. Chagnon C. Moreau G. Dumont S. St-Jean É. Achim A. (2024). American beech outgrows sugar maple at the sapling stage regardless of partial harvest intensity in northern hardwood forests. For. Ecol. Manag.553:121630. doi: 10.1016/j.foreco.2023.121630
42
Legendre P. (2005). Species associations: the Kendall coefficient of concordance revisited. J. Agr. Biol. Envir. St.10, 226–245. doi: 10.1198/108571105X46642
43
Liu C. Siri M. Ren C. Huang J. Feng C. Li K. (2023). Drought is threatening plant growth and soil nutrients of grassland ecosystems: a meta-analysis. Ecol. Evol.13:e1009. doi: 10.1002/ece3.10092
44
Lozano F. C. Huynh K. D. (1989). Foliar diagnosis of sugar maple decline by DRIS. Comm. Soil Sci. Plant Anal.20, 1895–1914. doi: 10.1080/00103628909368191
45
Masson E. Cogliastro A. Houle D. Rivest D. (2019). Variation in soil and foliar nutrition status along a forest edge–interior gradient in sugar maple forest fragments. Can. J. For. Res.49, 1463–1470. doi: 10.1139/cjfr-2019-0185
46
Midgley M. G. Sims R. S. (2020). Mycorrhizal association better predicts tree effects on soil than leaf habit. Front. For. Glob. Change3:74. doi: 10.3389/ffgc.2020.00074
47
Moore J.-D. Ouimet R. (2006). Ten-year effect of dolomitic lime on the nutrition, crown vigor, and growth of sugar maple. Can. J. For. Res.36, 1834–1841. doi: 10.1139/x06-081
48
Moreau G. Achim A. Pothier D. (2019). A dendrochronological reconstruction of sugar maple growth and mortality dynamics in partially cut northern hardwood forests. For. Ecol. Manag.437, 17–26. doi: 10.1016/j.foreco.2019.01.031
49
Moreau G. Achim A. Pothier D. (2020). An accumulation of climatic events has led to years of reduced growth for sugar maple in southern Quebec, Canada. Ecosphere11:e03183. doi: 10.1002/ecs2.3183
50
Moreau L. Thiffault E. Cyr D. Boulanger Y. Beauregard R. (2022). How can the forest sector mitigate climate change in a changing climate? Case studies of boreal and northern temperate forests in eastern Canada. For. Ecosyst.9:100026. doi: 10.1016/j.fecs.2022.100026
51
Nolet P. Kneeshaw D. (2018). Extreme events and subtle ecological effects: lessons from a long-term sugar maple–American beech comparison. Ecosphere9:e02336. doi: 10.1002/ecs2.2336
52
Ouimet R. Moore J. D. Duchesne L. (2013). Soil thresholds update for diagnosing foliar calcium, potassium, or phosphorus deficiency of sugar maple. Com. Soil Sci. Plant Anal.44, 2408–2427. doi: 10.1080/00103624.2013.803563
53
Pallardy S. G. Rhoads J. L. (1993). Morphological adaptations to drought in seedlings of deciduous angiosperms. Can. J. For. Res.23, 1766–1774. doi: 10.1139/x93-223
54
Phillips R. P. Brzostek E. Midgley M. G. (2013). The mycorrhizal-associated nutrient economy: a new framework for predicting carbon-nutrient couplings in temperate forests. New Phytol.199, 41–51. doi: 10.1111/nph.12221
55
Parsons W. F. J. Ghotsa Mekontchou C. Maheu A. Rivest D. (2025). Sugar maple litter decay rates are reduced more strongly by drought than by American beech proliferation in the understory. Ecology & Evolution, Art.ECE371416.
56
R Core Team (2021). R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing.
57
Rasoanaivo A. Ghotsa Mekontchou C. Rochon P. Nolet P. Maheu A. (2024). Increased tree water use with the development of a dense understory layer in a north American hardwood forest. Ecohydrology17:2648. doi: 10.1002/eco.2648
58
Reed S. E. Volk D. Martin D. K. H. Hausman C. E. Macy T. Tomon T. et al . (2022). The distribution of beech leaf disease and the causal agents of beech bark disease (Cryptoccocus fagisuga, Neonectria faginata, N. Ditissima) in forests surrounding Lake Erie and future implications. For. Ecol. Manag.503:119753. doi: 10.1016/j.foreco.2021.119753
59
Roy M.-E. Nolet P. (2018). Early-stage of invasion by beech bark disease does not necessarily trigger American beech root sucker establishment in hardwood stands. Biol. Invasions20, 3245–3254. doi: 10.1007/s10530-018-1771-7
60
Roy M.-E. Surget-Groba Y. Delagrange S. Rivest D. (2021a). Legacies of forest harvesting on soil properties along a chronosequence in a hardwood temperate forest. For. Ecol. Manag.496:119437. doi: 10.1016/j.foreco.2021.119437
61
Roy M.-E. Surget-Groba Y. Rivest D. (2021b). Legacies of forest harvesting on plant diversity and plant community composition in temperate deciduous forest. Appl. Veg. Sci.24:e12620. doi: 10.1111/avsc.12620
62
Sardans J. Peñuelas J. (2007). 2007 drought changes phosphorus and potassium accumulation patterns in an evergreen Mediterranean forest. Funct. Ecol.21, 191–201. doi: 10.1111/j.1365-2435.2007.01247.x
63
Sardans J. Peñuelas J. (2012). The role of plants in the effects of global change on nutrient availability and stoichiometry in the plant-soil system. Plant Physiol.160, 1741–1761. doi: 10.1104/pp.112.208785
64
Saucier J.-P. Bergeron J.-F. Grondin P. Robitaille A. (1998). Les regions écologiques du Québec méridional (3e version): un élément du système hiérarchique de classification écologique du territoire mis au point par le ministère des Ressources naturelles. Ste-Foy, Québec: Ministère des Ressources Naturelles du Québec.
65
Schoenau J. J. Qian P. Huang W. Z. (1993). Ion exchange resin membranes as plant root simulators. Proceedings, soils and crops workshop 1993, extension division. Saskatoon, SK: University of Saskatchewan, 392–400.
66
Schönbeck L. Gessler A. Schaub M. Rigling A. Hoch G. Kahmen A. et al . (2020). Soil nutrients and lowered source:sink ratio mitigate effects of mild but not of extreme drought in trees. Environ. Exp. Bot.169:103905. doi: 10.1016/j.envexpbot.2019.103905
67
Simon J. (2023). Relevance of organic vs. inorganic nitrogen in intra- and interspecific competition of seven central European tree species. Trees37, 1583–1591. doi: 10.1007/s00468-023-02418-0
68
Soil Classification Working Group (1998). The Canadian system of soil classification. Ottawa: Agriculture and Agri-Food Canada.
69
Stern R. L. Schaberg P. G. Rayback S. A. Hansen C. F. Murakami P. F. Hawley G. J. (2023). Growth trends and environmental drivers of major tree species of the northern hardwood forest of eastern North America. J. For. Res.34, 37–50. doi: 10.1007/s11676-022-01553-7
70
Tourville J. C. Zarfos M. R. Lawrence G. B. McDonnell T. C. Sullivan T. J. Dovčiak M. (2023). Soil biotic and abiotic thresholds in sugar maple and American beech seedling establishment in forests of the northeastern United States. Plant Soil491, 387–400. doi: 10.1007/s11104-023-06123-2
71
Tubbs C. H. Houston D. R. (1990). “American Beech” in Silvics of North America: 2. eds. BurnsR. M.HonkalaB. H. (Washington DC: Hardwoods, Agriculture Handbook 654, USDA Forest Service).
72
Urbina I. Sardans J. Beierkuhnlein C. Jentsch A. Backhaus S. Grant K. et al . (2015). Shifts in the elemental composition of plants during a very severe drought. Environ. Exp. Bot.111, 63–73. doi: 10.1016/j.envexpbot.2014.10.005
73
Vehtari A. Gelman A. Simpson D. Carpenter B. Bürkner C. P. . (2021). Rank-normalization, folding, and localization: an improved R-hat for assessing convergence of MCMC. (with Discussion). Bayesian Anal. 16, 667–718. doi: 10.1214/20-BA1221
74
Walworth J. L. Sumner M. E. (1987). “The diagnosis and recommendation integrated system (DRIS)” in Advances in soil science. ed. StewartB. A. (New York: Springer), 149–188.
75
Yamaguchi D. K. (1991). A simple method for cross-dating increment cores from living trees. Can. J. For. Res.21, 414–416. doi: 10.1139/x91-053
76
Yu S. Mo Q. Li Y. Li Y. Zou B. Xia H. et al . (2019). Changes in seasonal precipitation distribution but not annual amount affect litter decomposition in a secondary tropical forest. Ecol. Evol.9, 11344–11352. doi: 10.1002/ece3.5635
77
Zarfos M. R. Lawrence G. B. Beier C. M. Page B. M. McDonnell T. C. Sullivan T. J. et al . (2024). Environmental change drivers reduce sapling layer diversity in sugar maple-beech forests of eastern North America. Ecosystems27, 1021–1039. doi: 10.1007/s10021-024-00930-z
Summary
Keywords
Acer saccharum , basal area increment, DRIS, Fagus grandifolia , plant root simulators, rainfall exclusion, soil moisture deficit
Citation
Parsons WFJ, Ghotsa Mekontchou C, Shovon TA, Maheu A and Rivest D (2025) Do drought and understory beech interact to influence the nutrition and growth of sugar maple?. Front. For. Glob. Change 8:1518704. doi: 10.3389/ffgc.2025.1518704
Received
28 October 2024
Accepted
23 April 2025
Published
15 May 2025
Volume
8 - 2025
Edited by
Aziz Ebrahimi, Purdue University, United States
Reviewed by
Juan De Dios Franco-Navarro, Spanish National Research Council (CSIC), Spain
Martin Dovciak, SUNY College of Environmental Science and Forestry, United States
Updates
Copyright
© 2025 Parsons, Ghotsa Mekontchou, Shovon, Maheu and Rivest.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: David Rivest, david.rivest@uqo.ca
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.