 Shiv Paul1,2
Shiv Paul1,2 Khilendra Singh Kanwal1*
Khilendra Singh Kanwal1* Anil Kumar1
Anil Kumar1 Sher Singh Samant1,2
Sher Singh Samant1,2 Indra Dutt Bhatt1
Indra Dutt Bhatt1 Rakesh Chandra Sundriyal1,3
Rakesh Chandra Sundriyal1,3 Swaran Lata2
Swaran Lata2- 1G.B. Pant National Institute of Himalayan Environment, Kosi-Katarmal, Almora, Uttarakhand, India
- 2ICFRE-Himalayan Forest Research Institute, Shimla, India
- 3Department of Forestry & Natural Resources, Hemvati Nandan Bahuguna Garhwal University, Srinagar, India
The populations of globally significant species in the Himalaya are invariably shrinking due to multiple forms of human manipulation. Over the decades, various species’ natural habitats have been influenced by changing climate patterns, and the trends are consistently increasing, which is an issue of concern. The Hippophae rhamnoides ssp. turkestanica Rousi is a critical species native to the Himalaya, belonging to the Elaeagnaceae family. It is widely known for its significant ecological, economic, and social benefits. There is a great need to conserve this species in the cold desert region to improve the livelihood and socioeconomic status of tribal communities. Therefore, the current study aims to assess the population status and predict highly suitable areas for Trans-Himalaya species under changing climatic conditions. The machine learning algorithm showed that Bio_6 (minimum temperature of the coldest month), elevation, and slope were the best suitable variables for the habitat prediction along with the CMIP6 project’s MIROC6 and CMCC-ESM2 climate change models to identify the potential distribution area of the species for the future under the SSP245 (middle of the way) and SSP585 (fossil-fueled development) scenarios, respectively. The result found that a 2.20% (4,028 km2) area is highly suitable for the species’ occurrence. The potential suitability areas of the species are predicted to expand along the boundary of the present distribution areas at the pace of climatic change. The current study findings will help to determine the species resource reserve in the Trans-Himalaya. The predicted distribution maps of the species would help policymakers and decision-makers design appropriate management and sustainable utilization approaches in the near future.
1 Introduction
The population and habitat of medicinal plants in the Himalaya are consistently decreasing due to various anthropic factors. Plants have been used for various therapeutic purposes since ancient times (Grover et al., 2002). Indigenous peoples worldwide have a long history of using medicinal plants and their derived products for daily use. In the current scenario, natural supplements of plants or their substitutes as synthetic medicines have acquired great attention among the people (Wyk and Wink, 2018). India is one of the largest exporters of unprocessed herbal drugs worldwide. The Himalayan region is known for its vast reserves of medicinal plants due to their promising conditions (Kumar et al., 2024). However, in the present scenario, the demand for transporting raw materials has accelerated, and people are doing it for business. As a result, illegal logging and unscientific harvesting of medicinal plants are increasing in the wild (Badola and Aitken, 2003). People harvest the plants on a large scale from the wild rather than cultivating them (Samant et al., 1998; Kumar et al., 2021). Therefore, the most significant species are fighting for survival and have reached the verge of extinction. Hence, monitoring the status and habitat of the important plants is very crucial for maintaining the hot spot legacy of the Himalayan region (Kumar et al., 2023). The existence and habitat of medicinal plants encounter severe challenges, which need to be conserved, and machine learning techniques known as ecological niche modeling would be significant tools for mapping the niche suitability of the species (Figure 1) (Kumar, 2023). In this context, species distribution modeling of the Hippophae rhamnoides ssp. turkestanica was performed using a machine learning algorithm. This species is native to the Himalaya region and belongs to the Elaeagnaceae family. It is widely known as a lifeline for tribal communities, following various usages, such as medicinal, commercial, and livelihood purposes (Figure 2).
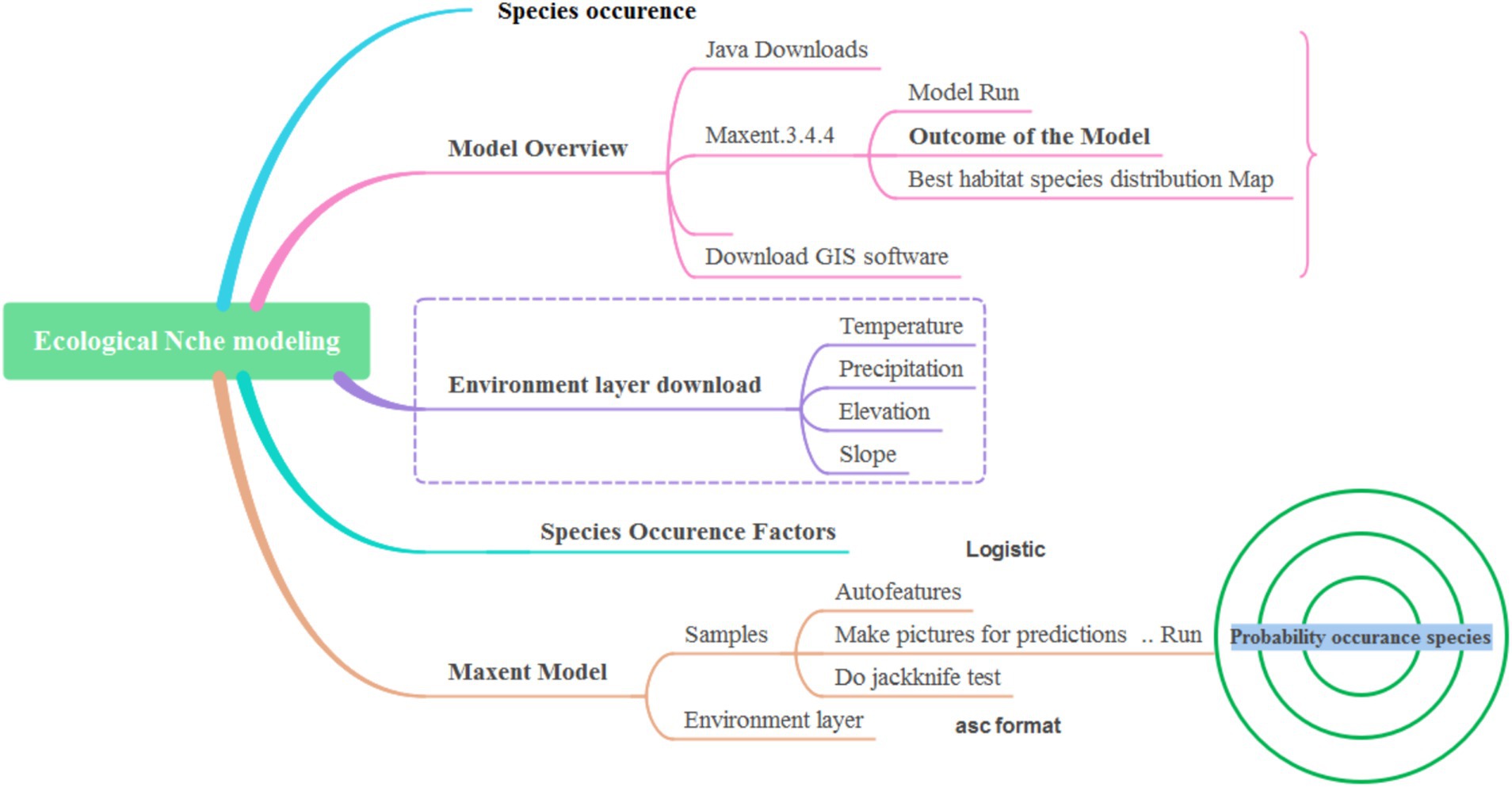
Figure 1. Overview of species distribution patterns through the ecological niche model.

Figure 2. Seabuckthorn plant with fruits and the marginal agricultural land habitat of the species.
In addition, H. rhamnoides ssp. turkestanica is commonly known as “Seabuckthorn” an ancient, multipurpose shrub with exceptional tolerance to varied temperature conditions and considerable genetic variety. The species is distributed in Afghanistan, Mongolia, Tadzhikistan, China, Tibet, Trans, and the North Western Himalayan Indian Himalayan Region (IHR) between an altitudinal range of 2,500–4,100 m amsl. Fruits are particularly significant as a rich source of vitamins A, B1, B2, B5, C, E, K, P, and others. Various researchers have documented that its ecological, agricultural, nutritional, medicinal, fuel, fodder, ornamental qualities, and antioxidant properties are well known to cure multiple diseases (Singh et al., 2003; Chaurasia et al., 2008; Singh, 2008; Paul, 2021). Food items made from fruit processing, such as syrup, carbonated juice, beer, sweet wine, tea, jam, and jelly, are in high demand both domestically and abroad (Singh et al., 2012). Besides, these plants have aphrodisiac properties and are also used for treating lung infection (Kala, 2005). Previous studies showed that H. rhamnoides ssp. turkestanica is vulnerable in the Cold Desert area of the IHR (Ved et al., 2003; Kala, 2005; Singh et al., 2022).
The earlier study also revealed that the consequential species of the Himalayan region are facing several obstacles due to the changing environmental conditions of the mountain region, and these obstacles need to be addressed promptly; otherwise, the population of these species will become extinct if proper action is not taken earnestly (Kattel, 2022). ENM, also known as habitat distribution, species distribution, and climate envelope modeling, are alternative names for comparable correlative or mechanistic modeling in which field observations have been linked to environmental predictor factors based on statistically or theoretically determined response surfaces (Guisan and Thuiller, 2005; Sillero, 2011) (Figure 1). A prevalent paradigm in broad-scale ecology and biogeography uses correlational modeling techniques with known species occurrences (Barve et al., 2011). For ENM, many algorithms are available, and these can also be differentiated by the requirements for data input (Qiao et al., 2019). Among these algorithms, the maximum entropy (MaxEnt) algorithm is one of the best suitable models for habitat niche modeling; it considers presence-only data, and with very low sample sizes, it typically induces good results: 13 records for generalist species and three for specialist species (van Proosdij et al., 2016; Ray et al., 2018; Sillero et al., 2021). Therefore, the MaxEnt algorithm for identifying suitable areas has been performed in the current study.
Climate change in the 20th century has already directly impacted the geographical distribution of many species (Beaumont et al., 2008). In such a way, the trends of species shifting along with climate change are continuing and increasing, and in the 21st century, it has reached the threshold capacity (Chapin et al., 2004). However, species respond for survival and adapt to biotic and physical surroundings under changing climatic conditions (Byrne et al., 2008; Stigall, 2008; Crawford, 2008; Tattersall et al., 2012). These days, rapid changes in climate, habitat, and the species’ adaptably responses to environmental factors have reached beyond their capacity to survive; consequently, the population has reached its marginal scale (Thompson et al., 2009; Roy et al., 2024). Therefore, ENM is often used to estimate the possible impact of climate change on species ranges. Therefore, we gathered Coupled Model Intercomparison Project Phase 6 (CMIP6) future climate data under two Shared Socioeconomic Pathways (SSPs) from the www.worldclim.org website from 2081 to 2100. Shared Socio-economic Pathways (SSPs) are a set of concepts for how society will progress in the 21st century and how the global environment will change. Every SSP includes quantitative information derived from integrated assessment, demographic, and socioeconomic models to estimate future socioeconomic growth (O’Neill et al., 2016; Kishore et al., 2024). SSP245 is the medium pathway of future greenhouse gas emissions, an update of scenario RCP4.5, with an additional radiative forcing of 4.5 W/m2 by the year 2100. This scenario is predicated on the assumption that climate protection measures are being implemented. The SSP585 scenario represents the highest end of the range of future paths and updates the RCP8.5 pathway with an increased radiative forcing of 8.5 W/m2 by the year 2100. This scenario shows a development path dominated by fossil fuels, which is linked to high levels of greenhouse gas emissions (Riahi et al., 2017; Böttinger and Kasang, 2021). However, Lal and Samant (2017) and He et al. (2023) have tried to estimate the ENM of H. rhamnoides ssp. turkestanica in the cold desert region, but the future habitat suitability across all the Trans-Himalayan still needs to be explored. For this instance, the ecological niche modeling of H. rhamnoides ssp. turkestanica was performed. The tribal people of the cold desert region of the Trans-Himalayan region widely use this species for multiple purposes. It is essential to comprehend this species’ status adaptation and future distribution range by the pace of changing environmental conditions across the Himalaya. It would help to conserve the status of this species. In the literature, multiple research papers have been published on species distribution modeling. However, no frequent study and conservation status have been documented in H. rhamnoides ssp. turkestanica across Trans-Himalayan region (Tamchos and Dorjey, 2024). Therefore, predicting species distributions under historic and changing climatic conditions by ecological niche modeling (ENM), and identifying the current population status of the species, is necessary for species conservation, restoration of habitats, and anticipation of future consequences. The study will aid in understanding population status, environmental factors conducive to the species’ presence, and area change (reduction/expansion) as climatic conditions change.
2 Materials and methodology
2.1 Population assessment
The present research on the quantitative assessment of the populations of H. rhamnoides ssp. turkestanica was carried out in the Changthang landscape (Changthang Cold Desert Wildlife Sanctuary, Rong Valley, and Gya-Meru region), Ladakh, India. The topography of the landscape is formed of deep gorges and vast plateaus. The climate of Changthang is poor and unpredictable. The summers are warm but short, and thunderstorms can occur at any time of year, often with hail. The winters are cold and Arctic-like, despite the latitude, due to the high elevation (Dolkar et al., 2018) (Figure 3).
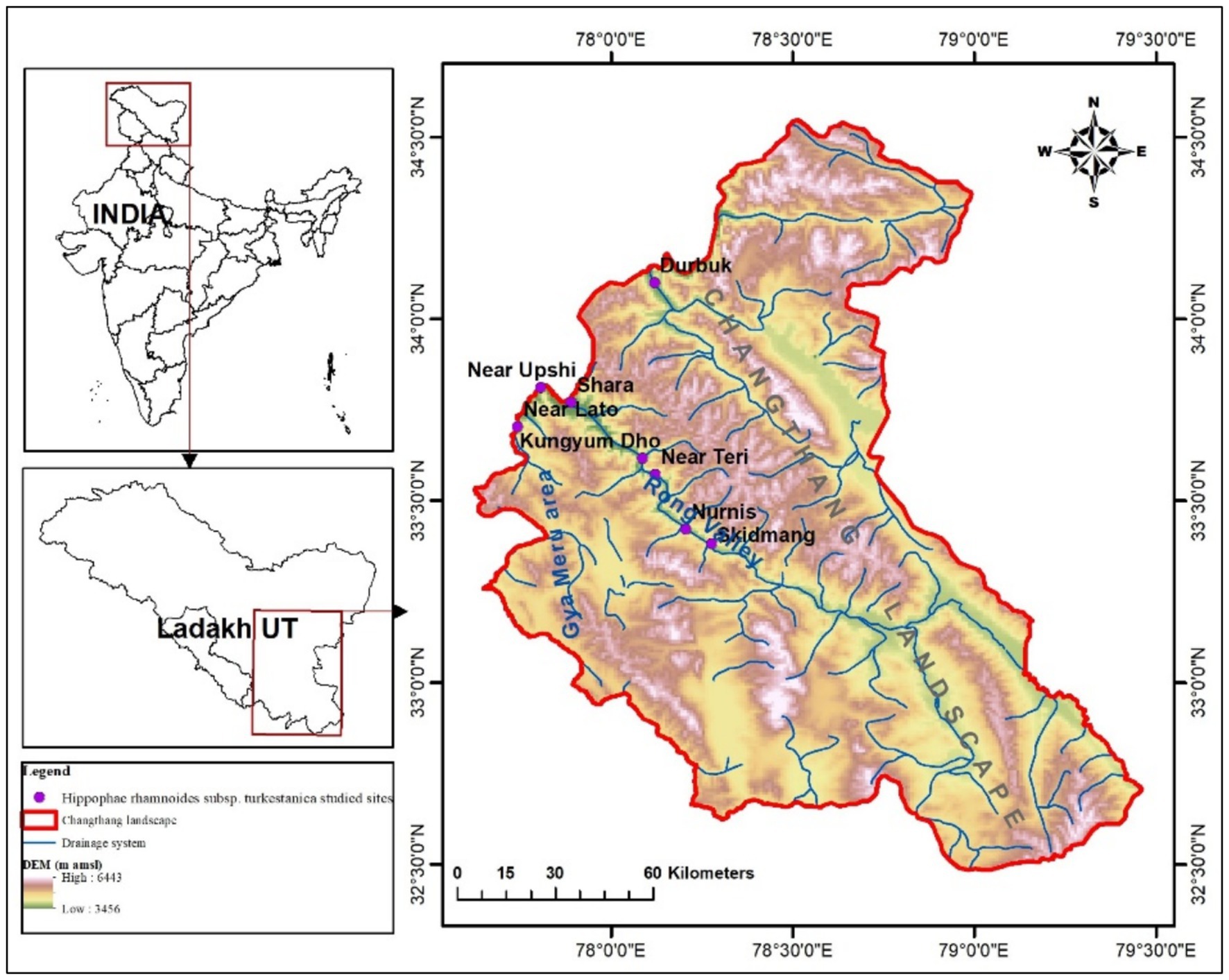
Figure 3. Map shows studied populations of H. rhamnoides ssp. turkestanica in Changthang.
2.2 Sampling methods
The habitats/sites supporting H. rhamnoides ssp. turkestanica across the Changthang landscape was selected to assess populations during 2019–2020. Habitats were identified based on physical features. The geo-reference of each population was obtained with the help of a hand-held Global Positioning System (Garmin Montana 650). For assessing the vegetation in each site and habitat, a 20 × 20 m plot was selected, and 10 quadrats of 5 × 5 m for shrubs and 20 quadrats of 1 × 1 m for herbs were randomly laid. Standard ecological methods were followed to collect and analyze the data (Curtis and McIntosh, 1950; Mueller-Dombois et al., 1974; Rana et al., 2011; Arya and Samant, 2016; Sharma and Samant, 2019). Plants were identified with the help of local floras (Chaurasia et al., 2008; Dvorský et al., 2018). In each quadrat, the number of individuals of each species was noted. Species richness (the number of species present in any specific area), density, frequency, and relative density of species have been calculated by following (Dhar et al., 1997; Joshi and Samant, 2004; Shukla et al., 2021).
2.3 Ecological niche modeling
For the species distribution modeling, the entire natural habitat of the species was evaluated. The present study has tried to cover the Trans-Himalayan region. During field data species assessment, most of the locations and habitats of H. rhamnoides ssp. turkestanica was present in cold desert regions such as Ladakh Union Territory, Lahaul and Spiti district, and Poh tehsil of Kinnaur district in Himachal Pradesh, which covers an area of ⁓1,54,000 km2, respectively. The specific secondary distribution habitat and location of H. rhamnoides ssp. turkestanica have been collected from two important sources (GBIF, 2024) and iNaturalist1 data in the GeoCAT web portal.2 Other sources in scientific academia, such as Google Scholar and published and unpublished data records, were also considered in the current study.
2.4 Species occurrence data
A total of 130 (08 primary and 122 secondary) occurrence points of H. rhamnoides ssp. turkestanica were gathered from primary and secondary sources (GBIF, 2024; Singh et al., 2011). Spatial bias arises because occurrence records are usually gathered in easily accessible, roadside, and well-liked study locations. We understand that sampling biases might cause data to be misinterpreted. Therefore, to eliminate sampling bias and remove duplicate points, the Geodesic Point Decimate tool in QGIS Desktop 3.28.1 was used to spatially thin the 130 occurrence points. Only 88 data points were left after spatial thinning at a 5 km distance, and these 88 points were finally used for niche modeling. The gathered occurrence points were converted into a Keyhole Markup Language Zipped (KMZ) file using ArcGIS 9.2 and uploaded into Google Earth, and illogical points were eliminated.
2.5 Environmental variables
The Digital Elevation Model (DEM) and the present (19 bioclimatic variables) and future climate data at the resolution of 30 arc-seconds (~1 km2) were obtained from the www.Worldclim.org website (Fick and Hijmans, 2017). For future climate data, we used two global climate models (GCMs), i.e., Model for Interdisciplinary Research on Climate-Sixth version (MIROC6) and Centro Euro-Mediterraneo sui Cambiamenti Climatici-Earth System Model Version 2 (CMCC-ESM2), under two SSPs, i.e., fossil-fueled development (SSP585) and optimistic (SSP245) climate scenarios, for the 2081–2,100 period. Earlier studies have demonstrated that these GCMs are most frequently applied in South Asia and exhibit the best performance in the Indian monsoon region (Bisht et al., 2016; Sun et al., 2021; Vinod and Agilan, 2024).
Data on slope and aspect were obtained from DEM using ArcGIS 9.2. Soil data (30 arc-seconds) were obtained from the FAO soil portal,3 Global Human Footprint (Geographic) v2 (1995–2004) (hfootp_geo) from the websites of Socioeconomic Data and Applications Center4 and World-Direct Normal Irradiation (DNI) GIS Data from the ENERGYDATA.INFO website,5 and Global land cover (300 m) from the European Space Agency GlobCover Portal (Arino et al., 2012).
Using the nearest neighbor interpolation approach in ArcGIS 9.2, land cover, soil, human footprint, and DNI data were resampled to the bioclimatic variables’ resolution (30 arc-seconds). Finally, all these variables have been clipped to the Trans-Himalayan region’s shapefile and converted to MaxEnt-compatible ^ASCII format using ArcGIS 9.2. Under climate change, no gap vegetation projections were available, and we considered the vegetation to be unchanged over this period. Because topographical data (slope, aspect, and elevation) were considered stationary, the same data were utilized in future datasets (Paul and Samant, 2023; Paul et al., 2023).
Autocorrelation between environmental variables can lead to collinearity, overfitting, and inaccurate interpretation of results (Amiri et al., 2020). Therefore, the correlation among variables was examined using the Band Collection Statistics tool in ArcGIS, and the variance inflation factor (VIF) was analyzed to see multicollinearity among variables. Thirteen environmental variables remained for modeling from the 26 selected variables after eliminating the variables with cross-correlation values larger than ± 0.80 (Rana et al., 2022) (Figures 4A–E). The finally selected variables were hfootp_geo, DNI, land cover, DEM, soil, slope, aspect, Bio_3 [isothermality (BIO2/BIO7) (×100)], Bio_6 (minimum temperature of coldest month), Bio_7 [temperature annual range (BIO5-BIO6)], Bio_8 (mean temperature of wettest quarter), Bio_15 [precipitation seasonality (coefficient of variation)], and Bio_19 (precipitation of coldest quarter).
2.6 Modeling parameters
To predict distribution maps, the most popular ENM tool is MaxEnt (Merow et al., 2013). In the present study, the MaxEnt version 3.4.4 k algorithm was used for predicting H. rhamnoides ssp. turkestanica distribution maps (Steven et al., 2022). To handle the model complexity, we applied 10 replicate effects, a cross-validated replicate run type, a maximum number of background points of 10,000, maximum iterations of 2,000, a default prevalence of 0.5, a Jackknife test to calculate the variable’s importance, and created response curves of each selected variable and threshold rule of 10 percentile training. To avoid model prediction overfitting, the regularization multiplier value of one was utilized by default. Ten percent of occurrence records were utilized for testing and 90% for model training (López-Martínez et al., 2016).
2.7 Model validation
MaxEnt’s output produced potential present and future distribution maps for H. rhamnoides ssp. turkestanica. The model’s accuracy was verified with the receiver operating characteristic (ROC) curve, True Skill Statistic (TSS), and area under curve (AUC) test values. The TSS was calculated by following Allouche et al. (2006). TSS values fall between −1 and +1, and values above 0.5 indicate acceptable model quality. Based on a 10-percentile training presence threshold value of 0.1046, we further divided distribution areas into four groups: unsuitable (0.00-threshold value), marginal (0.1046–0.50), moderate (0.50–0.75), and highly suitable (0.75–1) (Amiri et al., 2022; Paul and Samant, 2024). The shift in range and direction of species expansion and decrease have been calculated by subtracting the present predicted map from the maps of future distribution under various climate scenarios (Veera et al., 2019).
3 Results
3.1 Population assessment
A total of eight natural wild populations of the H. rhamnoides ssp. turkestanica has been studied in the riverine and dry slope (four sites, each) habitats, west (four sites), east, south, south-east, and north-west (one site, each) aspects, between 33°22.944’N and 34°05.998’N latitudes and 78°16.609′E and 77°44.618′E longitudes, altitude 3,464–4,014 m amsl, and slope varied from 5 to 500, in Changthang Landscape during 2019–2021. The population/habitat characteristics and geo-references of the sampled sites have been presented in Table 1.
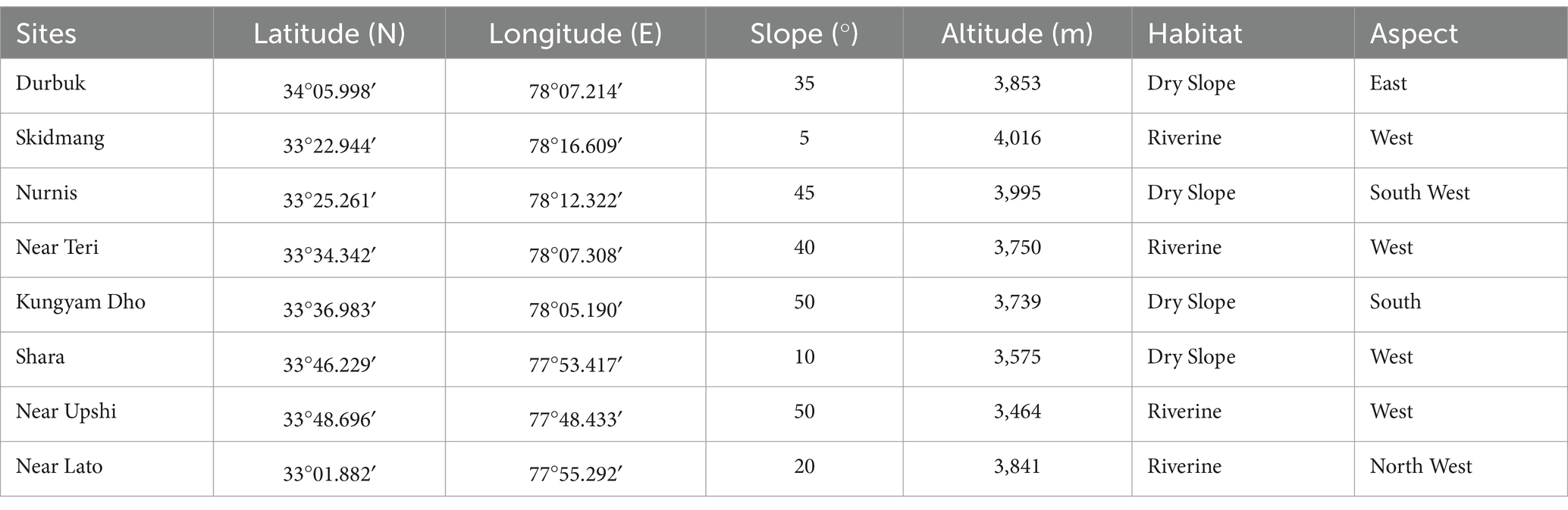
Table 1. Site characteristics and geo-references of the studied H. rhamnoides ssp. turkestanica populations in Changthang.
In the studied H. rhamnoides ssp. turkestanica populations, tree richness (number of species) ranged from 0 to 2; shrub richness ranged from 2 to 5; herbs richness ranged from 1 to 6; total tree density ranged from 0.90 to 2.50 Ind/100 m2; total shrubs density ranged from 30 to 61.2 Ind/100 m2; total herb density ranged from 2.25 to 16.50 Ind m−2; density of H. rhamnoides ssp. turkestanica ranged from 10.8 to 47.6 Ind/100 m2; relative density of H. rhamnoides ssp. turkestanica ranged from 27.55 to 93.45%; and frequency of H. rhamnoides ssp. turkestanica ranged from 50 to 70. Maximum density and abundance of H. rhamnoides ssp. turkestanica were recorded in the Skidmang population, and the maximum frequency and relative density were in the Shara population (Table 2). The studied sites did not contain any tree seedlings or saplings.
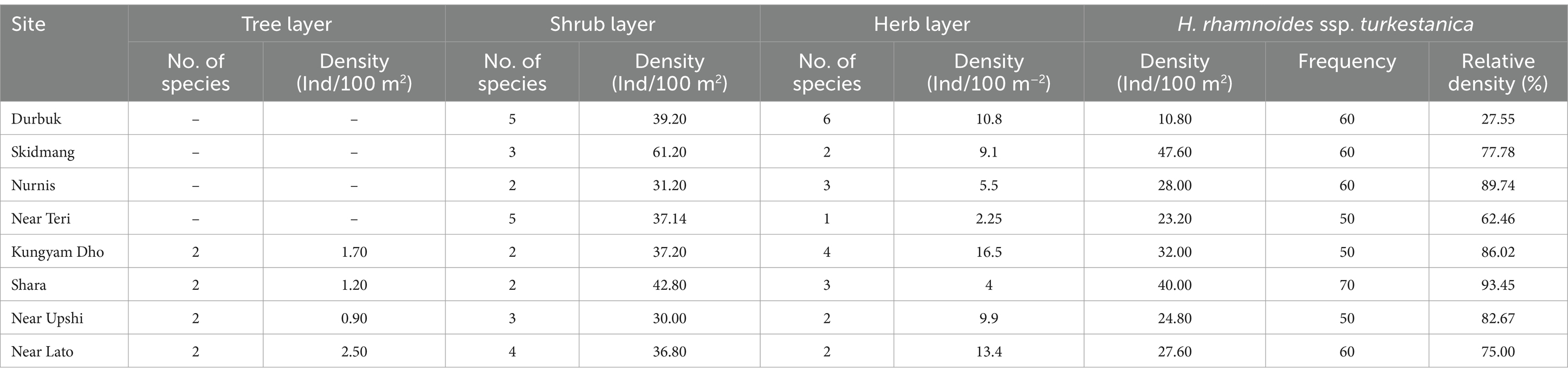
Table 2. Density, richness, and frequency of H. rhamnoides ssp. turkestanica in the studied populations.
3.2 Ecological niche modeling
3.2.1 Present distribution
The MaxEnt predicted present distribution model has AUC training and test values of 0.927 ± 0.005 and 0.895 ± 0.063, respectively (Table 3 and Figure 4A). Among the selected environmental variables, bio_6 (minimum temperature of the coldest month) has the highest relative contributions and importance to predict the MaxEnt model of the H. rhamnoides ssp. turkestanica (Table 3). Bio_6 is the environmental variable with the largest gain when utilized alone and slope significantly reduces the gain when it is removed, hence these two variables are looked to contain the most important information as per the Jackknife test (Figure 4B). The predicted response curves showed that regions with elevations between 2,300 and 3,600 m amsl and minimum temperatures of the coldest month between −16°C and −7°C are possibly favorable for the species distribution (Figures 4C,D).
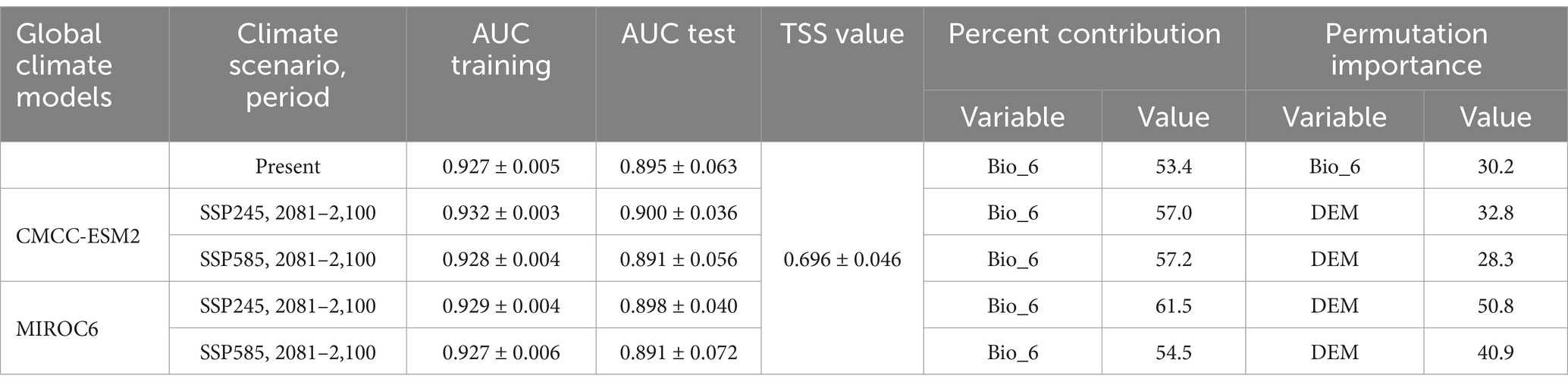
Table 3. H. rhamnoides ssp. turkestanica distribution models’ prediction accuracy.
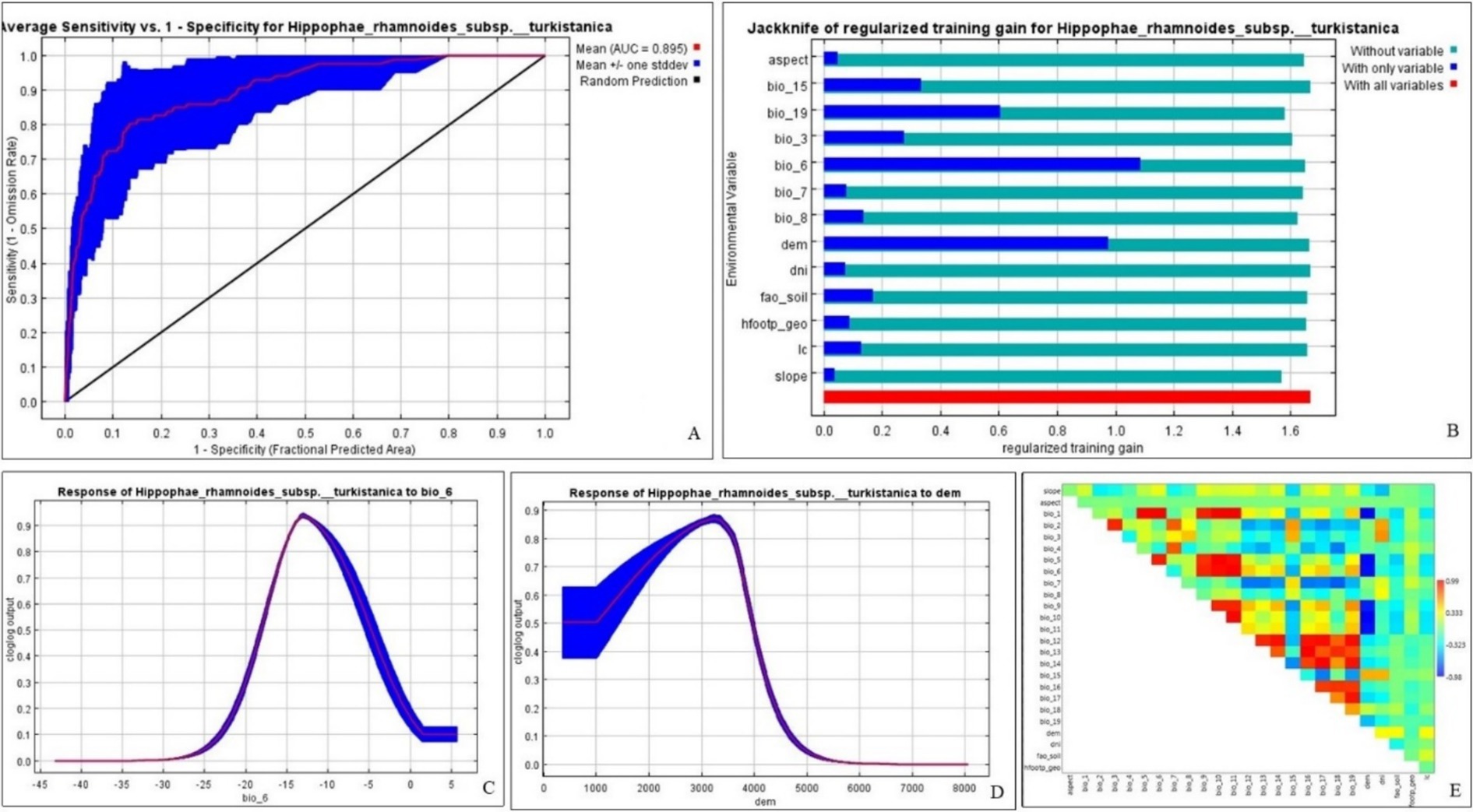
Figure 4. (A) ROC curve, (B) Jackknife evaluation outcomes, (C,D) key variables predicted response curves, and (E) matrix of correlations between variables.
The results of the MaxEnt model indicate that in the Trans-Himalayan region of India, the highly suitable class covers 4,028 km2 (2.20%) of the total geographical area for the occurrence of H. rhamnoides ssp. turkestanica, the moderate suitability class covers 5,464 km2 (2.98%), and the marginal suitability class covers 36,244 km2 (19.76%). In total, 45,736 km2 (24.93%) of the land is appropriate for the presence of H. rhamnoides ssp. turkestanica (Tables 4, 5; Figure 5). There are highly suitable regions in Pattan, Mayar, Tod, Spiti valleys, and Lower areas of Bhaga Valley in Lahaul and Spiti district and Chango, Nako, and Hango regions of Kinnaur district in Himachal Pradesh; Kidmang to Sanjak in Indus Valley, Shyok and Nubra valley, Panikhar, Sankoo, Kargil, and Dras regions, in Ladakh UT; and Gorikot, Shrigar, Thagas, Shimshal, and Karimabad regions in Pak occupied Kashmir within the altitudinal range of 2,400–3,600 m amsl on the floor and lower sides of the valleys (Figure 5).
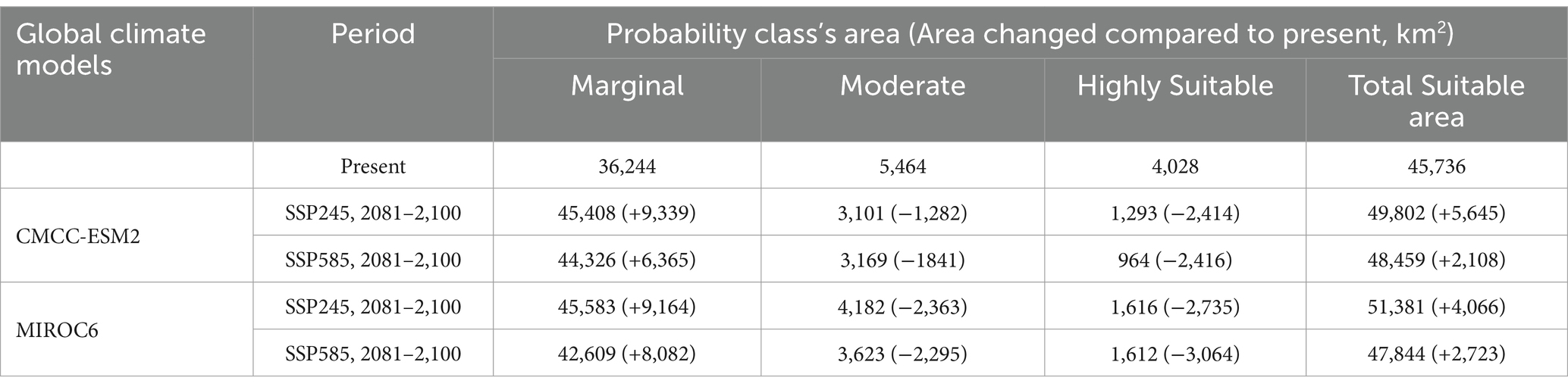
Table 4. Potential H. rhamnoides ssp. turkestanica suitability area predicted under climate change scenarios.
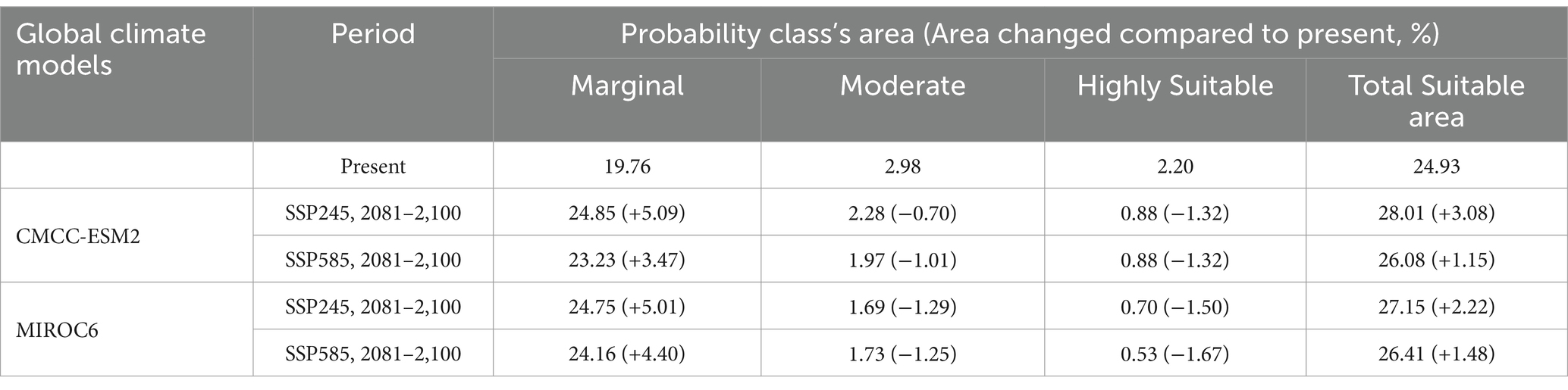
Table 5. Percentage changes in H. rhamnoides ssp. turkestanica suitability areas in different climate scenarios.
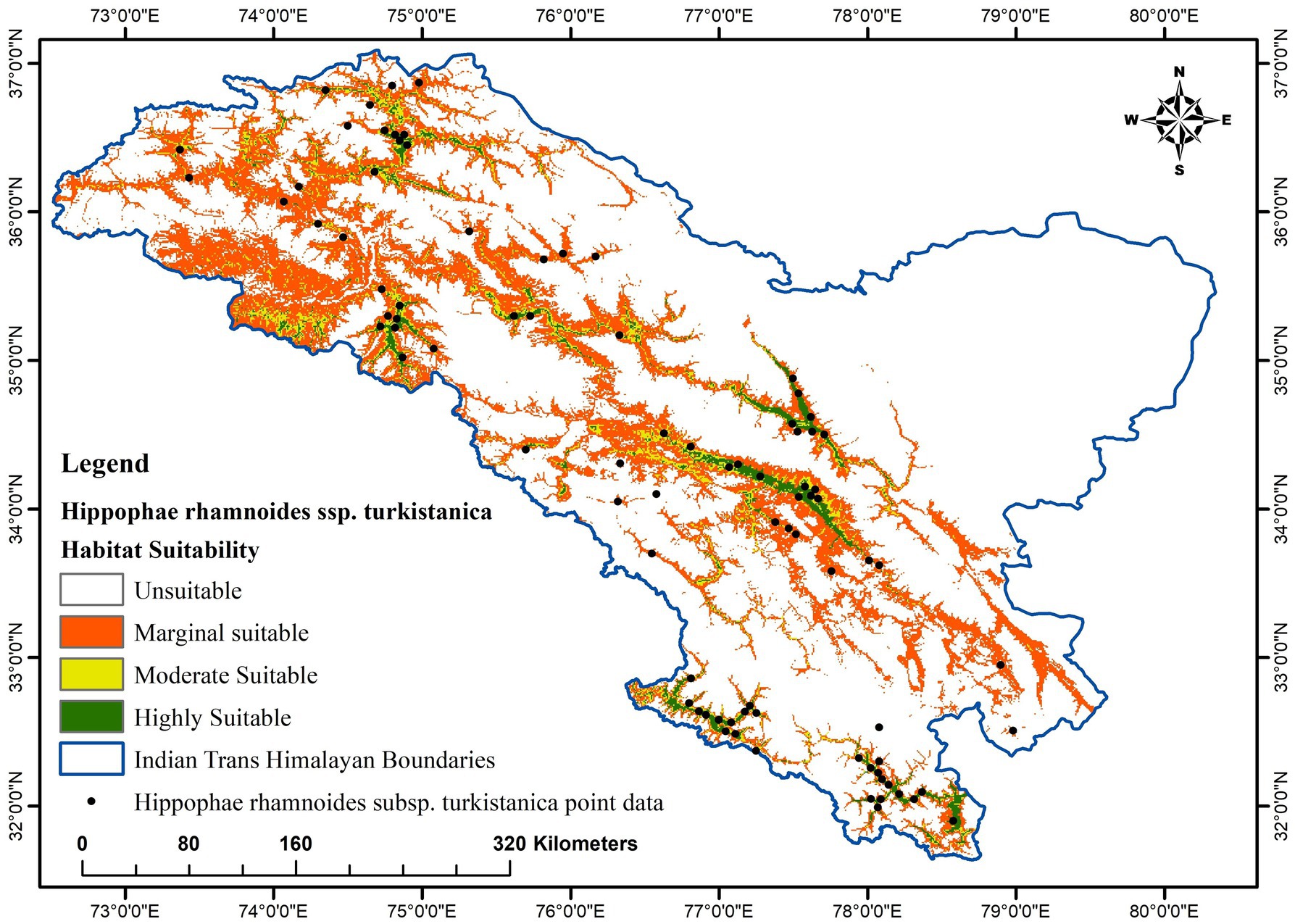
Figure 5. Distribution of H. rhamnoides ssp. turkestanica and potential suitability map in the Trans-Himalaya.
3.2.2 Future distribution
The TSS, AUC test, and training values for both GCMs (MIROC6 and CMCC-ESM2) were satisfactory for SSP245 and 585 climatic scenarios for the 2081–2,100 periods (Table 3). Based on the climate scenarios SSP 245 and SSP 585, the suitable area of H. rhamnoides ssp. turkestanica under the CMCC-ESM2 model was 49,802 km2 (28.01%) and 48,459 km2 (26.08%), respectively. Under MIROC6, it was 51,381 km2 (27.15%) and 47,844 km2 (26.41%) for SSP 245 and 585 climate scenarios, respectively (Tables 4, 5). The highly and moderately suitable areas under changing climatic conditions have been predicted to decrease in comparison to the predicted current distribution. However, the marginal and overall total suitability areas have been predicted to increase. In comparison to the current predicted model, the CMCC-ESM2 and MIROC6 models have predicted the suitability area for H. rhamnoides ssp. turkestanica will expand by 3.08 and 2.22%, respectively, for the SSP245 climate scenario in the 2081–2100 period. Similarly, under the SSP 585 scenario, the CMCC-ESM2 and MIROC6 models suggest that the total suitable area will expand by 1.15 and 1.48%, respectively (Tables 4, 5, and Figure 6). The projected suitability zones in different classes are represented in Tables 4, 5, and Figure 6 for selected SSPs and GCMs (Figure 7).
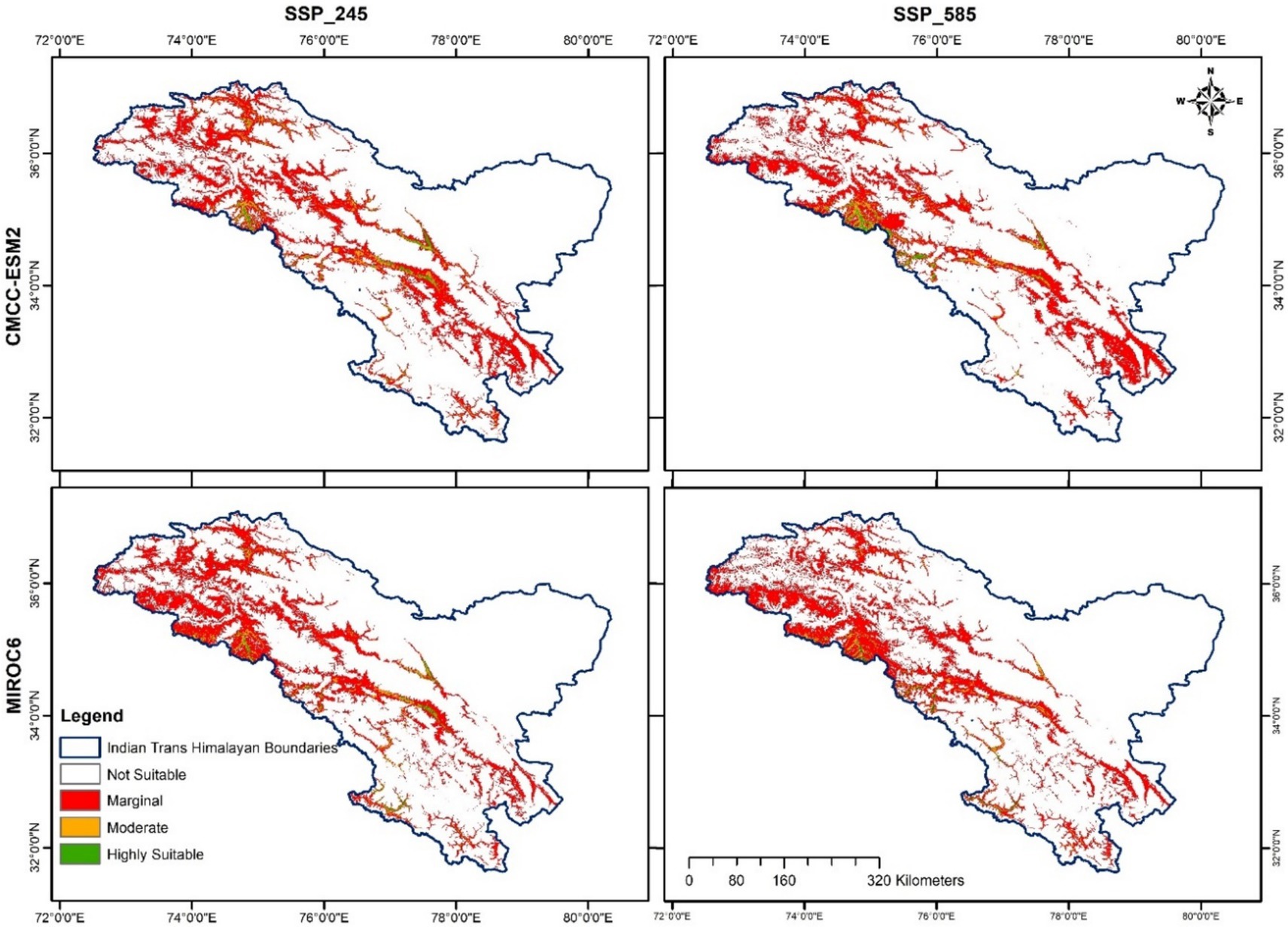
Figure 6. Future H. rhamnoides ssp. turkestanica distribution under climate change scenarios.
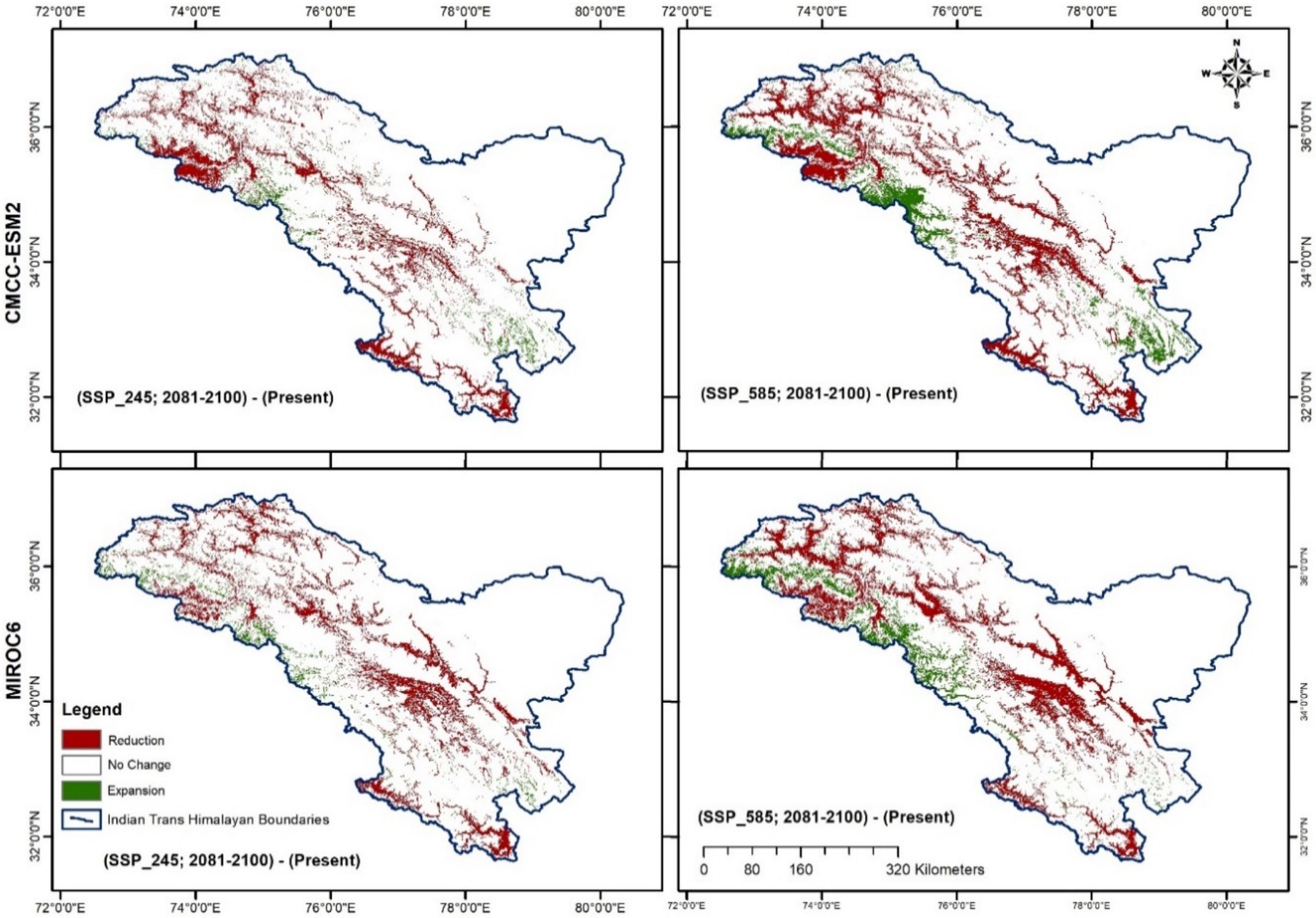
Figure 7. Effects of climate change on the spatial distribution of H. rhamnoides ssp. turkestanica are derived by subtracting future projections from the present predicted model.
4 Discussion
Medicinal plants have numerous health benefits for people, especially in areas where access to Western treatment is limited. Millions of individuals make their income as traditional healers, collectors, or marketers of medicinal plants. Both rural and urban residents rely heavily on the harvesting and trading medicinal plants as a source of revenue. Medicinal species are mostly harvested in an unsustainable manner, which could lead to the extinction of many populations (Applequist et al., 2020). The Indian Himalayan region is home to many priceless medicinal plants mentioned in Ayurvedic literature. This region’s diversity of medicinal plants is under a lot of stress from climate change, and it may eventually go extinct. The alpine ecosystems have changed due to climate change, and the current resources of medicinal plants have been negatively impacted by phenology pattern alterations, habitat fragmentation, invasion of new species, shifting range of distribution, and secondary metabolite changes (Kumar et al., 2012; Maikhuri et al., 2018).
The significant ecological, economic, and social advantages of a vulnerable plant H. rhamnoides ssp. turkestanica is well-known, and it has the potential to significantly improve the socioeconomic standing of the tribal communities and green the chilly desert region (Singh et al., 2011). In the current study, a total of eight wild populations of the species have been studied to check the population status, and the probable distribution under current and changing climatic conditions has been predicted. Salix alba L. and Populus balsamifera L. were associated tree species. Clematis tibetana Kuntze, Salix daphnoides Villo., Myricaria squamosa Desv., Rosa webbiana Wall. ex Royle, Festuca rubra L., Echinops cornigerus DC., Tanacetum gracile Hook.f. & Thomson, Physochlaina praealta (Decne.) Miers, and Lancea tibetica Hook.f. & Thomson were major associated shrub species. In the studied populations, the selected species have been reported from dry and riverine habitats. Habitat and aspect-wise, the maximum density of the selected species was recorded from the riverine habitat and the west aspect (Skidmang). The density of H. rhamnoides ssp. turkestanica has a significant negative correlation with slope (r = −0.620, n = 8, and p < 0.01) and latitude (r = −0.673, n = 8, and p < 0.01) (Figure 8). This indicates that the slope and latitude play an important role in species distribution.
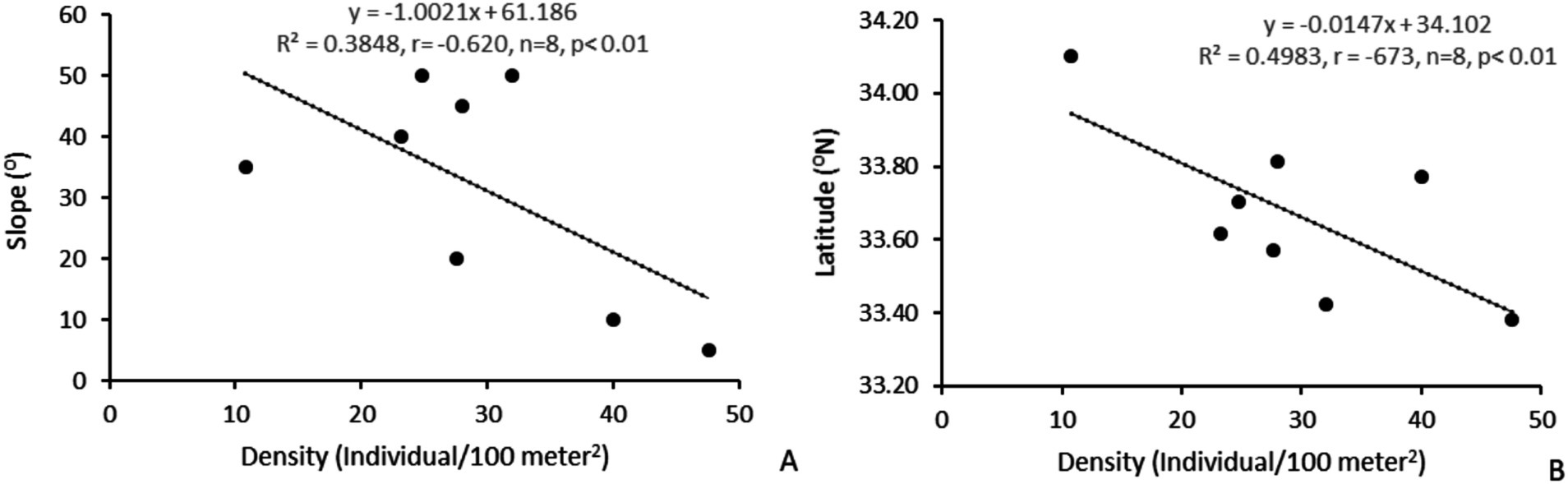
Figure 8. Correlation between (A) H. rhamnoides ssp. turkestanica density and slope and (B) density and latitude.
Predicted MaxEnt models are evaluated using ROC curves, AUC, and TSS values. AUC values are interpreted as follows: 1.0 indicates a perfect test, 0.9–0.99 is an excellent test, 0.8–0.89 is a good test, 0.7–0.79 is a fair test, and 0.51–0.69 is a poor test (Hanley and McNeil, 1982; Carter et al., 2016). AUC test values (0.891 ± 0.072 to 0.900 ± 0.036) and AUC training values (0.927 ± 0.006 to 0.932 ± 0.003) in the current study fell under excellent and good tests. The calculated average TSS value (0.696 ± 0.046) in the current ENM was above 0.5, indicating acceptable model quality (Allouche et al., 2006). The models were found suitable as the ROC curves were situated toward the left and top axes and far from the 1:1 line (Pearson, 2007) (Figure 4A).
Reduction of the high suitability area is seen when the current distribution map is subtracted from future distribution maps, but the overall total suitability area increases (Tables 4, 5; Figure 7). The increase in the total suitability area under optimistic (SSP245) climate scenarios was higher than fossil-fueled development (SSP585) scenario for the 2081–2100 period, a similar trend was also predicted for the H. rhamnoides ssp. turkestanica in China (He et al., 2023). As a result of global warming, there will be more arid and semiarid regions (Ramarao et al., 2019), which could potentially be the cause of the future growth of suitable habitats for H. rhamnoides ssp. turkestanica. In an era of rapid land-use transformations and increasing risk of climate change in the Trans-Himalayan highlands of Ladakh, 28 invasive wild-growing plants have already been reported. Roadsides and agricultural areas are among the sites that these invasive species most frequently occupy (Zargar et al., 2025). H. rhamnoides ssp. turkestanica is mostly found in the marginal agricultural land, and these invasive plants can affect the distribution of the species.
Based on MaxEnt’s prediction, the distribution of the species was most significantly influenced by the minimum temperature of the coldest month, suggesting that this species was the most temperature-sensitive (Figure 4B and Table 3). The minimum temperature of the coldest month (bio_6) was also identified as one of the major variables in a previous study that predicted the possible distribution map of H. rhamnoides ssp. turkestanica in the North Western Himalaya (Lal and Samant, 2017).
5 Conclusion and future considerations for the species conservation
The current study presents the population status and possible distribution map of H. rhamnoides ssp. turkestanica in the Trans-Himalaya, India, under present and changing climatic conditions. According to the study’s findings, the species’ distribution will be significantly impacted by the anticipated highly favorable locations. Policymakers and decision-makers can use the predicted distribution maps to create a suitable management plan for cultivating the species using various available propagation techniques to ensure the region’s economic growth with the active support of local communities and government organizations. Earlier studies have highlighted that the overall suitable area of the species would be expanded in the Trans-Himalaya due to changing climatic conditions, which need to be addressed for conservation purposes. The current study emphasizes the species-changing patterns and behavior of the habitat due to the pace of climate change’s impacts on the population of the significant plant species in the unexplored provinces of the IHR. These challenges need to be addressed by nature-based solutions. There is an urgent need to frame a region-based strategy according to the needs of the cold desert region to mitigate the adverse effects of climate change and preserve livelihood options in the IHR. However, to tackle these challenges, it is necessary to emphasize such a study through machine learning algorithms for habitat conservation at the individual species level as a cornerstone of climate resilience. Biodiversity credit and ecosystem restoration, with their associated species and biodiversity finance approach, should be introduced as law at the administrative level to prioritize biodiversity, including species conservation actions. By doing so, we can protect the hot spot legacy and integrity of the Himalaya and save the biodiversity for upcoming generations. However, the key roles and accountabilities of the people about the importance of the Himalaya for the existence of human civilization should also be enlightened to the people through awareness and education camps from door to door. In addition, proper coordination between local people, policy planners, and governments must be emphasized to address this risk. Efforts to build climate-resilient ecosystems can be sustained if all communities are in a single frame. Therefore, global and state delegate administrations should strengthen region-based mitigation strategies to address ecosystem vulnerability and habitat changes of the species across the IHR. It would be a significant way to handle the specific challenges of battling species vulnerability in the Himalaya.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
SP: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft. KK: Conceptualization, Data curation, Formal analysis, Investigation, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. AK: Data curation, Formal analysis, Writing – original draft. SS: Supervision, Writing – review & editing, Methodology, Conceptualization. IB: Project administration, Resources, Writing – review & editing. RS: Project administration, Resources, Supervision, Writing – review & editing. SL: Data curation, Formal analysis, Writing - review & editing.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Acknowledgments
The authors are thankful to the Director, G.B. Pant National Institute of Himalayan Environment, Kosi-Katarmal, Almora, Uttarakhand, and the Director, ICFRE-Himalayan Forest Research Institute, Conifer Campus, Panthaghati, Shimla, Himachal Pradesh, India, for facilities, encouragement, and inspiration. Help received from the Himachal Pradesh Forest Department, the Forest Department, Leh-Ladakh, and the UNDP-SECURE Himalaya team of the Wildlife Department, Leh-Ladakh has also been highly acknowledged.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
1. ^https://www.inaturalist.org/
2. ^https://geocat.iucnredlist.org/
3. ^https://www.fao.org/soils-portal/
References
Allouche, O., Tsoar, A., and Kadmon, R. (2006). Assessing the accuracy of species distribution models: prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 43, 1223–1232. doi: 10.1111/J.1365-2664.2006.01214.X
Amiri, M., Tarkesh, M., Jafari, R., and Jetschke, G. (2020). Bioclimatic variables from precipitation and temperature records vs. remote sensing-based bioclimatic variables: which side can perform better in species distribution modeling? Ecol. Inform. 57:101060. doi: 10.1016/j.ecoinf.2020.101060
Amiri, M., Tarkesh, M., and Shafiezadeh, M. (2022). Modelling the biological invasion of Prosopis juliflora using geostatistical-based bioclimatic variables under climate change in arid zones of southwestern Iran. J. Arid. Land 14, 203–224. doi: 10.1007/S40333-022-0004-1/
Applequist, W. L., Brinckmann, J. A., Cunningham, A. B., Hart, R. E., Heinrich, M., Katerere, D. R., et al. (2020). Erratum: Scientistsʼ warning on climate change and medicinal plants. Planta Med. 86:e1. doi: 10.1055/a-1113-1659
Arino, O., Ramos Perez, J. J., Kalogirou, V., Bontemps, S., Defourny, P., and Van Bogaert, E. (2012). Global land cover map for 2009 (GlobCover 2009). © Eur. Sp. agency Univ. Cathol. Louvain (UCL), PANGAEA. doi: 10.1594/PANGAEA.787668
Arya, S., and Samant, S. (2016). Habitat wise assessment of alpine vegetation and prioritization of communities for conservation in a part of Nanda Devi Biosphere Reserve of West Himalaya. Int. J. Life Sci. 5, 6–19. Available at: http://www.crdeepjournal.org/wp-content/uploads/2016/01/Vol-5-1-2-IJLS.pdf.
Badola, H. K., and Aitken, S. (2003). The Himalayas of India: A treasury of medicinal plants under siege. Biodiversity 4, 3–13. doi: 10.1080/14888386.2003.9712694
Barve, N., Barve, V., Jiménez-Valverde, A., Lira-Noriega, A., Maher, S. P., Peterson, A. T., et al. (2011). The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecol. Modell. 222, 1810–1819. doi: 10.1016/j.ecolmodel.2011.02.011
Beaumont, L. J., Hughes, L., and Pitman, A. J. (2008). Why is the choice of future climate scenarios for species distribution modelling important? Ecol. Lett. 11, 1135–1146. doi: 10.1111/j.1461-0248.2008.01231.x
Bisht, V. K., Bhandari, A. K., Negi, B. S., Bhandari, A. K., and Kandari, L. S. (2016). Fritillaria roylei Hook. in Western Himalaya: Species Biology, Traditional Use, Chemical Constituents, Concern and Opportunity. Res. J. Med. Plant 10, 375–381. doi: 10.3923/rjmp.2016.375.381
Böttinger, M., and Kasang, D. (2021). The SSP Scenarios — English. Available online at: https://www.dkrz.de/en/communication/climate-simulations/cmip6-en/the-ssp-scenarios (Accessed November 18, 2024).
Byrne, M., Yeates, D. K., Joseph, L., Kearney, M., Bowler, J., Williams, M. A. J., et al. (2008). Birth of a biome: insights into the assembly and maintenance of the Australian arid zone biota. Mol. Ecol. 17, 4398–4417. doi: 10.1111/j.1365-294X.2008.03899.x
Carter, J. V., Pan, J., Rai, S. N., and Galandiuk, S. (2016). ROC-ing along: evaluation and interpretation of receiver operating characteristic curves. Surgery 159, 1638–1645. doi: 10.1016/j.surg.2015.12.029
Chaurasia, O. P., Khatoon, N., and Singh, S. B. (2008). Field guide floral diversity of Ladakh. WWF-India. Available online at: https://www.researchgate.net/profile/Arvind-Singh-21/post/Ladakh_Indian_Himalaya_plant_identification/attachment/59d63b2679197b8077998308/AS%3A409087150968834%401474545385175/download/1.pdf (Accessed October 29, 2024).
Chapin, F. S., Callaghan, T. V., Bergeron, Y., Fukuda, M., Johnstone, J. F., Juday, G., et al. (2004). Global Change and the Boreal Forest: Thresholds, Shifting States or Gradual Change? vol. 33. Available at: http://www.ambio.kva.se
Crawford, R. M. (2008). Plants at the Margin: Ecological Limits and Climate Change. Cambridge, UK: Cambridge University Press. Available at: https://books.google.co.in/books?hl=en&lr=&id=fvvIxkPjTFkC&oi=fnd&pg=PA5&dq=Crawford,+2008+&ots=Jt9aa6B9FG&sig=KaYfIpjVDhkNc0pIqRE-ES38jHA#v=onepage&q&f=false ().
Curtis, J. T., and McIntosh, R. P. (1950). The Interrelations of Certain Analytic and Synthetic Phytosociological Characters. Ecology 31, 434–455. doi: 10.2307/1931497
Dhar, U., Rawal, R. S., and Samant, S. S. (1997). Structural diversity and representativeness of forest vegetation in a protected area of Kumaun Himalaya, India: implications for conservation. Biodivers. Conserv. 6, 1045–1062. doi: 10.1023/A:1018375932740
Dolkar, T., Azim, H., and Sheikh, F. (2018). Ladakh: to study the pastoral Changpa nomadic life of Changthang region Jammu and Kashmir. Academia.edu. Available online at: https://www.academia.edu/download/96554588/1521136272_JK1505SS.pdf (Accessed October 29, 2024).
Dvorský, M., Klimeš, L., Doležal, J., Wild, J., and Dickoré, W. B. (2018). A field guide to the flora of Ladakh : Academia.
Fick, S. E., and Hijmans, R. J. (2017). WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315. doi: 10.1002/JOC.5086
Grover, J. K., Yadav, S., and Vats, V. (2002). Medicinal plants of India with anti-diabetic potential. J. Ethnopharmacol. 81, 81–100. doi: 10.1016/S0378-8741(02)00059-4
Guisan, A., and Thuiller, W. (2005). Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 8, 993–1009. doi: 10.1111/j.1461-0248.2005.00792.x
Hanley, J. A., and McNeil, B. J. (1982). The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology 143, 29–36. doi: 10.1148/radiology.143.1.7063747
He, X., Si, J., Zhu, L., Zhou, D., Zhao, C., Jia, B., et al. (2023). Modeling habitat suitability of Hippophae rhamnoides L. using MaxEnt under climate change in China: a case study of H. r. sinensis and H. r. turkestanica. Front. For. Glob. Chang. 5:1095784. doi: 10.3389/ffgc.2022.1095784
Joshi, H. C., and Samant, S. S. (2004). Assessment of forest vegetation and conservation priorities of communities in part of Nanda Devi biosphere reserve, west Himalaya. Part I. Int. J. Sustain. Dev. World Ecol. 11, 326–336. doi: 10.1080/13504500409469835
Kala, C. P. (2005). Indigenous uses, population density, and conservation of threatened medicinal plants in protected areas of the Indian Himalayas. Conserv. Biol. 19, 368–378. doi: 10.1111/j.1523-1739.2005.00602.x
Kattel, R. (2022). Dynamic capabilities of the public sector: Towards a new synthesis. Gower Street, London. Available at: https://www.ucl.ac.uk/bartlett/public-purpose/wp2022-07
Kishore, B. S. P. C., Kumar, A., and Saikia, P. (2024). Understanding the invasion potential of Chromolaena odorata and Lantana camara in the Western Ghats, India: an ecological niche modelling approach under current and future climatic scenarios. Ecol. Inform. 79:102425. doi: 10.1016/j.ecoinf.2023.102425
Kumar, A. (2023). Uncertainties and assumptions of bioclimatic envelope model while monitoring niche of significant species in the Indian Himalayan forest ecosystem. Hima Paryavaran 37, 54–56.
Kumar, A., Sathyakumar, S., Goraya, G. S., Gupta, A. K., Adhikari, B. S., and Rawat, G. S. (2021). Sustainable harvesting and cultivation protocols of threatened medicinal and aromatic plants of the Western Himalaya. Dehradun: Wildlife Institute of India.
Kumar, R. K., Sruajani, M. S., Arya, J. C., and Joshi, G. C. (2012). Impact of climate change on diversity of Himalayan medicinal plant: a threat to Ayurvedic system of medicine. Int. J. Res. Ayurveda Pharm. 3, 327–331.
Kumar, A., Verma, R. K., Kumar, P., and Dushyant, (2024). Indigenous Acquaintance and Perceptions of Local Inhabitants towards Biodiversity Conservation in Talra Wildlife Sanctuary, North Western Himalaya, India, 53–72 doi: 10.1007/978-3-031-51696-2_3
Kumar, V., Kumar, P., Bhargava, B., Sharma, R., Irfan, M., and Chandora, R. (2023). Transcriptomic and Metabolomic Reprogramming to Explore the High-Altitude Adaptation of Medicinal Plants: A Review. J. Plant Growth Regul. 42, 7315–7329. doi: 10.1007/s00344-023-11018-8
Lal, M., and Samant, S. S. (2017). Niche modelling-an approach to predict habitats for conservation of Hippophae species in trans and North Western Indian Himalaya. Int. J. Sci. Nat. 8, 437–446. Available online at: https://www.researchgate.net/publication/322518933 (Accessed June 21, 2023).
López-Martínez, V., Sánchez-Martínez, G., Jiménez-García, D., Pérez-De la, O. N. B., and Coleman, T. W. (2016). Environmental suitability for Agrilus auroguttatus (Coleoptera: Buprestidae) in Mexico using MaxEnt and database records of four Quercus (Fagaceae) species. Agric. For. Entomol. 18, 409–418. doi: 10.1111/afe.12174
Maikhuri, R. K., Phondani, P. C., Dhyani, D., Rawat, L. S., Jha, N. K., and Kandari, L. S. (2018). Assessment of climate change impacts and its implications on medicinal plants-based traditional healthcare system in central Himalaya, India. Iran. J. Sci. Technol. Trans. A Sci. 42, 1827–1835. doi: 10.1007/s40995-017-0354-2
Merow, C., Smith, M. J., and Silander, J. A. (2013). A practical guide to MaxEnt for modeling species’ distributions: what it does, and why inputs and settings matter. Ecography (Cop.). 36, 1058–1069. doi: 10.1111/j.1600-0587.2013.07872.x
Muller-Dombois, D. S., and Ellenberge, H. (1974). Aims and Methods of Vegetation Ecology. New York: John Willey and Sons.
O’Neill, B. C., Tebaldi, C., van Vuuren, D. P., Eyring, V., Friedlingstein, P., Hurtt, G., et al. (2016). The scenario model Intercomparison project (ScenarioMIP) for CMIP6. Geosci. Model Dev. 9, 3461–3482. doi: 10.5194/gmd-9-3461-2016
Paul, S. (2021). Diversity, distribution and conservation status of raw edible plant resources of the Madgram watershed, Lahaul Valley, Himachal Pradesh, India. J. Non-Timber For. Prod. 28, 150–156. doi: 10.54207/bsmps2000-2022-0w0dq1
Paul, S., Lata, S., and Barman, T. (2023). Habitat distribution modeling of the Pinus gerardiana under projected climate change in the North-Western Himalaya, India. Landsc. Ecol. Eng. 19, 647–660. doi: 10.1007/s11355-023-00570-w
Paul, S., and Samant, S. S. (2023). Population biology, ecological niche modelling of endangered and endemic Pittosporum eriocarpum Royle in Western Himalaya, India. J. Nat. Conserv. 72:126356. doi: 10.1016/J.JNC.2023.126356
Paul, S., and Samant, S. S. (2024). Population ecology and habitat suitability modelling of an endangered and endemic medicinal Meconopsis aculeata Royle under projected climate change in the Himalaya. Environ. Exp. Bot. 225:105837. doi: 10.1016/j.envexpbot.2024.105837
Pearson, R. G. (2007). Species’ distribution modeling for conservation educators and practitioners. Synth. Am. Museum Nat. Hist. 50, 54–89. Available online at: https://scholar.google.com/scholar?hl=en&as_sdt=0%2C5&q=Species’+distribution+modeling+for+conservation+educators+and+practitioners&btnG= (Accessed September 11, 2023).
Qiao, H., Feng, X., Escobar, L. E., Peterson, A. T., Soberón, J., Zhu, G., et al. (2019). An evaluation of transferability of ecological niche models. Ecography (Cop.). 42, 521–534. doi: 10.1111/ecog.03986
Ramarao, M. V. S., Sanjay, J., Krishnan, R., Mujumdar, M., Bazaz, A., and Revi, A. (2019). On observed aridity changes over the semiarid regions of India in a warming climate. Theor. Appl. Climatol. 136, 693–702. doi: 10.1007/s00704-018-2513-6
Rana, M. S., Samant, S. S., and Rawat, Y. S. (2011). Plant communities and factors responsible for vegetation pattern in an alpine area of the northwestern Himalaya. J. Mt. Sci. 8, 817–826. doi: 10.1007/s11629-011-2078-7
Rana, S. K., Rana, H. K., Stöcklin, J., Ranjitkar, S., Sun, H., and Song, B. (2022). Global warming pushes the distribution range of the two alpine ‘glasshouse’ Rheum species north-and upwards in the eastern Himalayas and the Hengduan Mountains. Front. Plant Sci. 13:925296. doi: 10.3389/fpls.2022.925296
Ray, D., Behera, M. D., and Jacob, J. (2018). Evaluating ecological niche models: a comparison between Maxent and GARP for predicting distribution of Hevea brasiliensis in India. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 88, 1337–1343. doi: 10.1007/s40011-017-0869-5
Riahi, K., van Vuuren, D. P., Kriegler, E., Edmonds, J., O’Neill, B. C., Fujimori, S., et al. (2017). The shared socioeconomic pathways and their energy, land use, and greenhouse gas emissions implications: an overview. Glob. Environ. Chang. 42, 153–168. doi: 10.1016/j.gloenvcha.2016.05.009
Roy, S., Wei, X., Weiskittel, A., Hayes, D. J., Nelson, P., and Contosta, A. R. (2024). Influence of climate zone shifts on forest ecosystems in northeastern United States and maritime Canada. Ecol. Indic. 160:111921. doi: 10.1016/j.ecolind.2024.111921
Samant, S. S., Dhar, U., and Palni, L. M. S. (1998). Medicinal plants of Indian Himalayan region. Nainital: Gyanodaya Prakashan.
Sharma, L., and Samant, S. S. (2019). Prioritization of habitats and communities for conservation in Cold Desert Biosphere Reserve, Trans Himalaya. India. Ecol. Res. 34, 509–523. doi: 10.1111/1440-1703.12010
Shukla, V., Singh, A., and Nautiyal, A. R. (2021). Population assessment and Phyto-chemical screening of Meconopsis aculeata Royle an endangered medicinal Plant of Western Himalaya. Springer Proc. Earth Environ. Sci., 429–443. doi: 10.1007/978-3-030-79065-3_33
Sillero, N. (2011). What does ecological modelling model? A proposed classification of ecological niche models based on their underlying methods. Ecol. Modell. 222, 1343–1346. doi: 10.1016/J.ECOLMODEL.2011.01.018
Sillero, N., Arenas-Castro, S., Enriquez-Urzelai, U., Vale, C. G., Sousa-Guedes, D., Martínez-Freiría, F., et al. (2021). Want to model a species niche? A step-by-step guideline on correlative ecological niche modelling. Ecol. Model. 456:109671. doi: 10.1016/j.ecolmodel.2021.109671
Stigall, A. L. (2008). Tracking Species in Space and Time: Assessing the Relationships Between Paleobiogeography, Paleoecology, and Macroevolution. Paleontol. Soc. Pap. 14, 233–248. doi: 10.1017/S1089332600001704
Sun, C., Zuo, J., Shi, X., Liu, X., and Liu, H. (2021). Diverse Inter-Annual Variations of Winter Siberian High and Link With Eurasian Snow in Observation and BCC-CSM2-MR Coupled Model Simulation. Front. Earth Sci. 9:761311. doi: 10.3389/FEART.2021.761311/BIBTEX
Singh, V. (2008). Seabuckthorn (Hippophae L.): A multipurpose wonder plant (Vol. III): Advances in research and development. Delhi: Daya Publishing House.
Singh, A., Butola, J. S., Samant, S. S., Sharma, P., Lal, M., and Marpa, S. (2012). Indigenous techniques of product development and economic potential of seabuckthorn: a case study of cold desert region of Himachal Pradesh, India. Proc. Natl. Acad. Sci. India Sect. B-Biol. Sci. 82, 391–398. doi: 10.1007/s40011-012-0042-0
Singh, V., Kallio, H., Sawhney, R. C., Gupta, R. K., Rongsen, L., Eliseev, I. P., et al. (2003). Seabuckthorn (Hippophae L.) a multipurpose wonder plant. Harvesting and processing technologies. Delhi: Indus Publishing Co.
Singh, A., Samant, S. S., Lal, M., Sharma, P., Butola, J. S., and Marpa, S. (2011). Assessment, mapping and harnessing economic potential of Hippophae species for the socio-economic development of tribal communities in Himachal Pradesh, India, in National Conference on Seabuckthorn: Emerging trends in R & D on Health Protection & Environmental Conservation (December 1–3, 2011), (CSK Himachal Pradesh agricultural university, Palampur-176062, Himachal Pradesh, India, 260–269.
Singh, A., Samant, S. S., Manohar, L., and Sharma, P. (2022). Conservation prioritization criteria to identify rarity of the plant species, habitats and communities in the Lahaul Valley, trans North-Western Himalaya, India. Arid Ecosyst. 12, 251–271. doi: 10.1134/S2079096122030131
Steven, J. P., Dudík, M., and Schapire, R. E. (2022). Maxent software for modeling species niches and distributions (Version 3.4.4). Available online at: https://biodiversityinformatics.amnh.org/open_source/maxent/ (Accessed June 21, 2023).
Tamchos, S., and Dorjey, K. (2024). Hippophae rhamnoides ssp. turkestanica: a potential species for sustainable propagation in the Cold Desert of Ladakh. Natl. Acad. Sci. Lett. 48, 89–94. doi: 10.1007/s40009-024-01431-2
Tattersall, G. J., Sinclair, B. J., Withers, P. C., Fields, P. A., Seebacher, F., Cooper, C. E., et al. (2012). Coping with thermal challenges: physiological adaptations to environmental temperatures. Compr. Physiol. 2, 2151–2202. doi: 10.1002/cphy.c110055
Thompson, D., Wallace, J., Jones, P. D., Jones, P. D., and Kennedy, J. J. (2009). Identifying signatures of natural climate variability in time series of global-mean surface temperature: Methodology and insights. J. Clim. 22, 6120–6141. doi: 10.1175/2009JCLI3089.1
van Proosdij, A. S. J., Sosef, M. S. M., Wieringa, J. J., and Raes, N. (2016). Minimum required number of specimen records to develop accurate species distribution models. Ecography (Cop.). 39, 542–552. doi: 10.1111/ecog.01509
Ved, D. K., Kinhal, G. A., Ravikumar, K., Prabhakaran, V., Ghate, U., Vijaya Shankar, R., et al. (2003). Conservation assessment and management prioritization for the medicinal plants of Jammu & Kashmir, Himachal Pradesh & Uttaranchal.
Veera, S. N. S., Panda, R. M., Behera, M. D., Goel, S., Roy, P. S., and Barik, S. K. (2019). Prediction of upslope movement of Rhododendron arboreum in Western Himalaya. Trop. Ecol. 60, 518–524. doi: 10.1007/s42965-020-00057-x
Vinod, D., and Agilan, V. (2024). Ranking of CMIP 6 climate models in simulating precipitation over India. Acta Geophys. doi: 10.1007/s11600-024-01313-7
Wyk, B.Van, and Wink, M. (2018). Medicinal plants of the world., Revised Edition. CAB International. Available at: https://books.google.co.in/books?hl=en&lr=&id=UAitDwAAQBAJ&oi=fnd&pg=PA3&dq=medicinal+plants+world&ots=gqAf04Rbnn&sig=ep5bvWWHOJ3r1zCi_9eL-M3Ll44
Keywords: distribution, MaxEnt, niche modelling, climate change, Trans-Himalaya, population assessment, environmental variables, model validation
Citation: Paul S, Kanwal KS, Kumar A, Samant SS, Bhatt ID, Sundriyal RC and Lata S (2025) Population status and impact of climate change on the distribution of vulnerable multipurpose plant Hippophae rhamnoides ssp. turkestanica for conservation in Trans-Himalaya, India. Front. For. Glob. Change. 8:1551024. doi: 10.3389/ffgc.2025.1551024
Edited by:
Jahangeer A. Bhat, Fiji National University, FijiReviewed by:
Aabid Hussain Mir, Government of Jammu & Kashmir, IndiaUmeshkumar L. Tiwari, Botanical Survey of India, India
Copyright © 2025 Paul, Kanwal, Kumar, Samant, Bhatt, Sundriyal and Lata. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Khilendra Singh Kanwal, a3NrYW53YWxAZ2JwaWhlZC5uaWMuaW4=