 Jorge Palmero-Barrachina1
Jorge Palmero-Barrachina1 Petr Blazek1
Petr Blazek1 Santiago Sabaté2,3*Teresa Sauras-Yera2Samuel Allasia-Grau2
Santiago Sabaté2,3*Teresa Sauras-Yera2Samuel Allasia-Grau2 Daniel Nadal-Sala2,3Sven Kallen4Tiago de Santana4
Daniel Nadal-Sala2,3Sven Kallen4Tiago de Santana4 Emil Cienciala1,5*
Emil Cienciala1,5*- 1IFER - Monitoring and Mapping Solutions, Jilove u Prahy, Czechia
- 2Department of Evolutionary Biology, Ecology and Environmental Sciences, Universitat de Barcelona, Barcelona, Spain
- 3CREAF (Centre de Recerca i Aplicacions Forestals), Barcelona, Spain
- 4Stichting Life Terra, Amsterdam, Netherlands
- 5IFER – Institute of Forest Ecosystem Research, Jilove u Prahy, Czechia
This study presents a comprehensive methodology for estimating potential biomass and carbon accumulation in European afforestation activities expected over a 40-year timespan, developed for the Life Terra project (LIFE19 CCM/NL/001200). We synthesized data from allometric equations, Yield tables, National Forest Inventories, and National Greenhouse Gas Inventory Reports across four European biogeographic regions: Alpine, Atlantic, Continental, and Mediterranean. While Life Terra encompasses six planting categories (ecological restoration, timber plantations, agroforestry/food forests, gardens, green infrastructure, and others), our analysis focused primarily on timber plantations due to data availability and reliability constraints. The study showed significant regional variations in planting density and growth patterns. Initial planting densities in timber plantations varied substantially across biogeographic regions (1,869–7,702 trees/ha), following exponential decline patterns over time. By year 40, individual tree biomass estimates ranged from 0.08 to 0.20 t/tree across regions and species types (conifers and broadleaves), with survival rates varying between 22.0 and 49.7%. This translated to stand-level biomass estimates of 54.7–232.6 t/ha at age 40 years. Our biomass estimates generally aligned with country-specific literature and IPCC default values, though showing considerable variation across sites, highlighting the importance of local conditions in tree growth and stand dynamics. The study provides a robust framework for assessing carbon sequestration potential in European afforestation projects, while acknowledging key uncertainties related to survival/mortality rates and climate change impacts. This methodology remains open to refinement through additional biomass equations and revised Yield tables. The future field validation studies should also include non-timber plantation categories that are not covered here.
1 Introduction
Afforestation and reforestation activities play a crucial role in enhancing greenness and biodiversity, as well as mitigating climate change by sequestering carbon through the growth of woody vegetation (Barry et al., 2014; Krause et al., 2017; Pérez-Silos et al., 2021). These activies enhance carbon fixation by expanding tree-covered area and promoting the growth of tree biomass over time (Masera et al., 2003; Wong and Dutschke, 2003; Vilén et al., 2016; Mo et al., 2023; Greenleaf, 2024).
For greenhouse gas emission inventories under UNFCCC, the mandatory ecosystem carbon pool changes that must be reported include living biomass, dead organic matter (litter and deadwood), and soil organic and inorganic layers (IPCC, 2006). Among these, living biomass estimation, together with soil C dynamic changes following afforestation, is the key and most dynamic component for assessing the effectiveness of tree planting efforts in carbon fixation. It involves quantifying the amount of living biomass, including both above-ground and below-ground components, within the afforested and/or reforested area. Above-ground biomass refers to the stems, branches and leaves of the trees and plants visible above the ground, while below-ground biomass encompasses the root system. Accurately estimating both components is essential for understanding the total carbon stored in both reforested and afforested ecosystems and its changes over time, directly linking to the amount of CO2 fixed from the atmosphere.
Life Terra1 is one of the largest recent tree planting projects based on the understanding that tree planting is a cost-effective nature-based solution for carbon capture. Since the Life Terra project has not been finalized yet, the exact number of trees and countries where it has helped with planting is still changing. However, with its ambitious goal to plant or facilitate the planting of millions of trees, Life Terra’s activities already span more than a dozen European countries across several biogeographic regions. Although the project duration is limited (2020–2025), it develops monitoring tools to enable long-term tracking of its afforestation efforts. Life Terra also promotes natural regeneration as a complementary strategy to enhance ecosystem resilience and biodiversity. As a part of this, a reliable projection of growth performance and survival rate are necessary to quantify expected biomass and carbon accumulation in trees in the mid-term.
To make the estimation feasible, the procedure must be tailored to the available input information, incorporating data on expected survival rate and leveraging locally available biomass equations and sampling programs, such as sample-based national forest inventories (NFIs). The estimation procedure should be designed to be applicable at both tree and stand level.
The aim of this contribution is twofold. First, to provide a transparent methodological description summarizing the estimation approaches and assumptions used by Life Terra to assess the expected biomass and carbon accumulation of its current planting activities over the next four decades (40 years since planting). Second, to discuss the estimation challenges (uncertainties) and offer related recommendations.
2 Methods
Estimation of the total living biomass of trees in the Life Terra timber plantations, expected to be reached at the age of 40 years, is specific to biogeographic regions, tree species, and/or tree species groups and types. The estimation uses local allometric equations and Yield tables to the greatest extent practicable, as described below.
2.1 Biogeographic regions
Each plantation is primarily classified by using biogeographic regions. Life Terra has primarily planted trees in the Atlantic, Continental, and Mediterranean regions, and to a lesser extent in the Alpine and Boreal regions (EEA, 2016). Representative local allometric equations and Yield tables were selected to best reflect the characteristics of each biogeographic region, except for the Boreal region, due to the (as of date) limited project exposure of Life Terra there.
2.2 Species groups
Life Terra has planted or facilitated the planting of over 550 tree and shrub species since the project’s inception. Species information is available in the Life Terra database. While the biomass estimation procedure requires species-specific data on allometry and growth performance, this information is generally not available for all species. Therefore, in addition to species-specific estimations, we grouped the species by functional type: broadleaves and conifers. However, species grouping could also be applied based on genus, as well as similarities in tree shape and wood density. This grouping approach (broadleaves and conifers) enables us to apply average biomass values from well-documented species to similar but data-deficient species within the same group, taking into account biogeographic region, country, and productivity class (site index), as detailed below.
Regarding shrubs, due to severely limited literature on their biomass estimates, Life Terra takes a conservative approach by excluding them from any CO2 accounting while emphasizing other environmental benefits.
2.3 Tree biomass functions (allometric equations)
Tree biomass functions are essential tools to estimate tree biomass based on measurable dimensions. A variety of published approaches are available for larger trees (diameter at breast height (DBH) > 5 cm, age >15 years) that estimate the dimensions of merchantable biomass (Zianis and Mencuccini, 2004; Pilli et al., 2006; Somogyi et al., 2007; Forrester et al., 2017; Vonderach et al., 2018). These approaches may either use tree or stand volume and convert it to biomass or apply specific biomass functions tailored for particular tree species. These functions typically rely on DBH and tree height, and often incorporate additional explanatory (input) variables. The basic biomass function form is:
where are the parameters of the equation, b is the biomass of tree component i (stem, branches, leaves, roots), and x1 and x2 are independent variables (most commonly DBH and tree height). The leaf component (leaves) is typically estimated for the winter stage, meaning that deciduous broadleaves are counted without leaves (which form a part of litter) while the biomass estimates for most conifers include needles. It is important to note that most of the data provided on tree biomass may include only above-ground biomass (AGB) components. Since Life Terra is focused on carbon and CO2 equivalents stored in the entire tree, it is necessary to estimate total tree biomass (TB), which also includes the below-ground component (BGB).
The set of applicable biomass functions used to estimate tree biomass for this project is summarized in Table 1. This information was primarily collected from two sources. First, we reviewed the most recent national submissions of greenhouse gas (GHG) inventories for the Land use, land-use change and forestry (LULUCF) sector under UNFCCC2 for the selected European countries. Secondly, we consulted the NFI reports. Complementarily, we used the other suitable published literature.
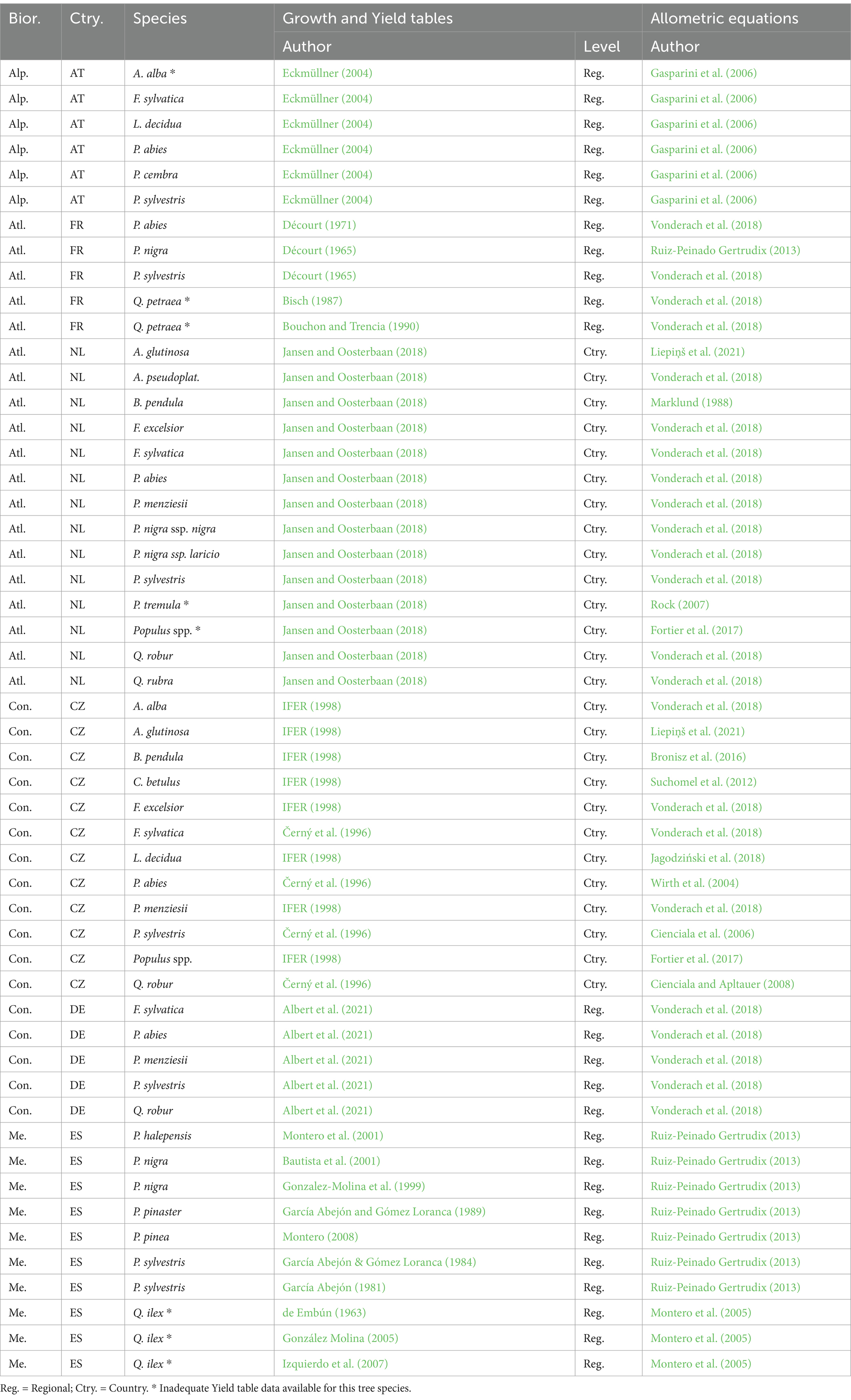
Table 1. The literature sources for growth and Yield tables and biomass equations by biogeographic region (Bior.), country (Ctry.), and species used in this study.
When the below-ground biomass component was absent in the allometric equations, appropriate expansion factors from Table 4.4 of the 2006 IPCC Guidelines for National Greenhouse Gas Inventories (NGHGI) (IPCC, 2006) were used.
2.4 Growth and Yield tables
Growth and Yield tables are forestry tools describing stand properties for individual species classified by site productivity. Yield tables usually require some estimate of stand age and thus cannot easily be applied to uneven-aged stands. Growth tables attempt to overcome this limitation by tabulating growth under various stand conditions (Vanclay, 1994). Since afforestation and reforestation activities typically result in even-aged stands, our approach is based solely on growth and Yield tables for ordinary managed even-aged stands (Table 1). They contain stand dimensions such as stand mean DBH, stand height and the number of trees per hectare for the expected lifespan of these stands. Therefore, the stand information associated with full stocking at the age of 40 years can be retrieved.
Site index is usually defined as the stand height that the dominant and codominant trees in fully stocked, even-aged stands attain at a given age. Yield tables are often classified by site index as a productivity indicator. An age of 100 years is often used to assess the site index (Mäkinen et al., 2017). For example, in a Norway spruce forest stand in the Czech Republic, a site index of 32 indicates that the dominant and codominant Norway spruce trees reach a height of 32 m at age 100 years. In total, 19 Yield tables (each for up to 15 tree species) from six different countries (Austria, Czech Republic, France, Germany, the Netherlands, and Spain) were used to estimate mean stand-specific dimensions (mean tree DBH and height) at age of 40 years for different site fertility levels (site index) (Table 1).
Since the specific fertility class of Life Terra plantations is mostly undetermined, the estimation procedure assumes that the likely productivity corresponds to the mean site index from the published Yield tables. This assumption is based on two main points: (i) there is scientific evidence supporting the hypothesis that tree growth performance is increasing in Europe in recent decades (Kahle, 2008; Cienciala et al., 2018; Pretzsch et al., 2020; Pretzsch and Hilmers, 2024), which has been influenced by environmental changes including nitrogen deposition and temperature shifts affecting the growing season. (ii) The concurrent increase in drought-related extremes and evidence of drought’s impacts on productivity–particularly in Mediterranean, but also temperate regions– may offset these productivity gains (Shestakova et al., 2019; Büntgen et al., 2021; Korosuo et al., 2023). Taking these two aspects into account, the mean site conditions from the Yield tables were used, with the average site index serving as the conservative default estimate. The references to the national and/or regional growth and Yield tables collected and used for this study are summarized in Table 1.
2.5 Estimation procedure for tree-level biomass
To estimate tree biomass in Life Terra plantations at an assumed stand age of 40 years, the following steps were performed (Figure 1):
1. Input Data: Stand dimensions (DBH and height for corresponding stand age) from country (or regional) Yield tables were used as input variables for species-specific allometric equations, covering the published range of site indices. It is important to note that we used the principal stand (main crop after thinning) from these Yield tables to also account for management interventions affecting stand density over time.
2. Biomass Calculation: The allometric equations from Step 1 provided outputs that were used to calculate the mean total dry weight biomass (t/tree) for different biogeographic regions, countries, tree species, and a productivity level that corresponds to a mean growth tree performance. The mean growth performance was determined using data from country-specific NFIs, whenever possible. This is the case for the main tree species of Czech Republic, Germany, Spain (just the province of Madrid), and France (Cienciala et al., 2022; Palmero-Barrachina et al., 2023) (Table 1). For all other cases, a mean site index of the specific range given Yield tables is used to characterize stand productivity. The corresponding stand dimensions and density associated with the selected site index were derived from Yield tables. Linear interpolation was applied to derive these stand dimensions and density when the NFI-derived mean growth performance (NFI-derived site index) fell between two site classes in the Yield tables.
3. Aggregation by categories: The biomass outputs from Step 2 were aggregated to derive representative tree biomass values for broader categories, i.e., functional type (broadleaved and coniferous) and biogeographic region.
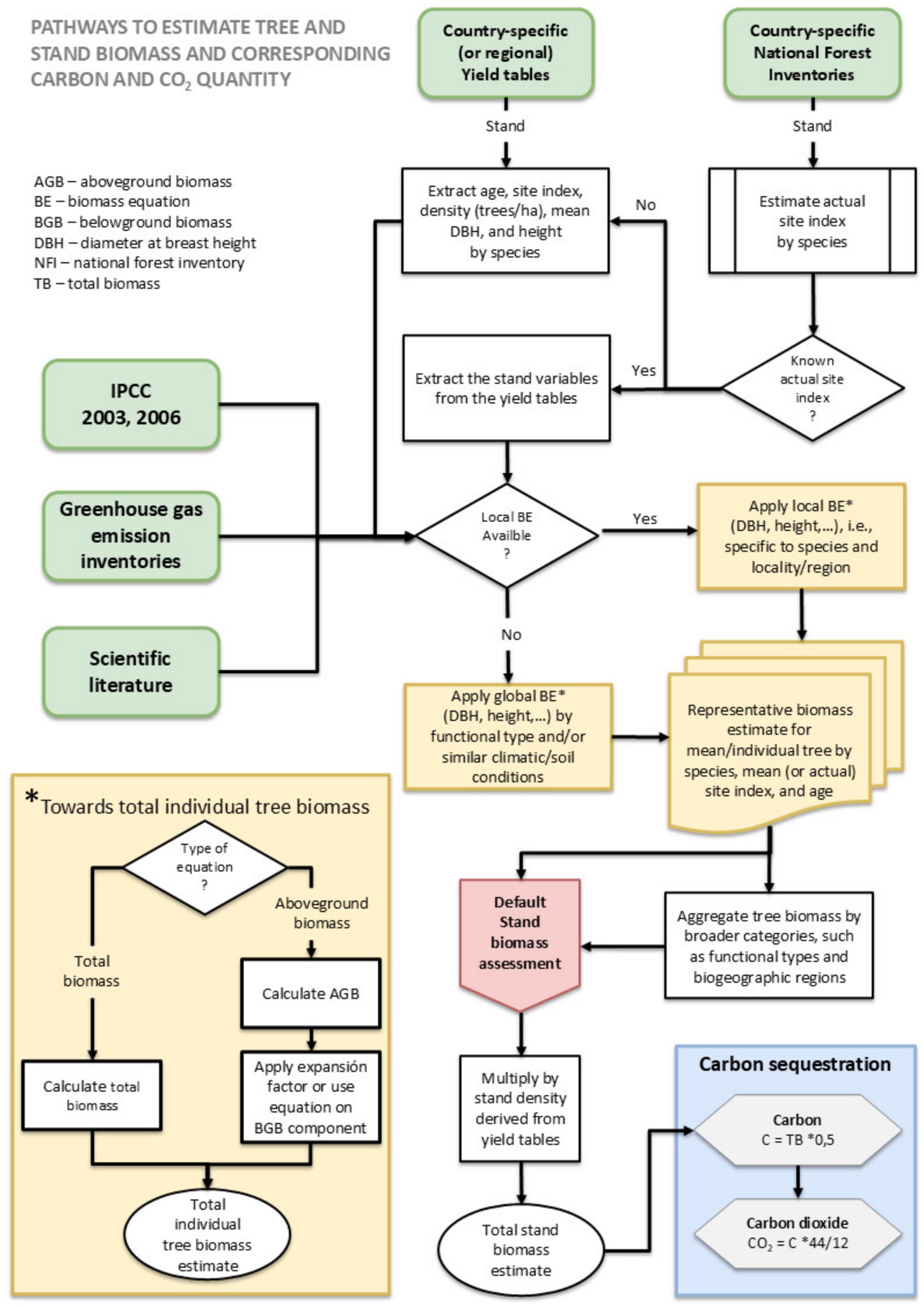
Figure 1. individual steps to estimate tree and stand biomass and corresponding carbon and CO2 quantity.
2.6 From tree level to plantation level
The Life Terra database includes the number of trees planted at each site, along with details about the species, planting date, planting objective, and location. It is important to stress that we do not use any allometry applicable to young trees to estimate stand biomass for current (young) plantation. Instead, we only use planting density as the initial information for survival rate considerations (described below) during the assumed stand development. This is because the estimation applicable to young trees remains generally highly uncertain and occasionally published literature on young tree allometry (e.g., Annighöfer et al., 2016; Pajtík et al., 2022) should be applied with care, specifically with respect to planting density. Therefore, for scaling biomass estimates from tree level to the stand level, the tree count at planting for each site is used together with survival rate considerations for the assumed stand level projection.
2.7 Mortality and survival considerations
To estimate biomass at a stand age of 40 years, it is essential to account for tree mortality and management interventions that may reduce the number of trees at a site over time (Bugmann et al., 2019). Our methodological framework uses the principal stand from the Yield tables (main crop after thinning) for average site conditions. Survival rate is determined as the proportion of standing living trees at a given age, e.g., at 40 years of age, relative to the original stand density at planting. It should be noted that mortality is a complementary fraction to survival (Mortality (%) = 100 – Survival (%)), and these terms are interchangeable. Since we are focusing on standing living trees, we decided to use the term survival thereon (Calzada and Millán, 2004; Pausas et al., 2004).
Timber plantations aim to grow trees for wood production. Life Terra forbids production cycles below 40 years as stated in its agreements with landowners, while allowing sustainable management practices such as regular thinning. Life Terra also prohibits the use of invasive species and monocultures, promoting biodiversity and ecological sustainability. To assess the survival rate in timber plantations, we derived stand density models (age-related decline) by functional type and biogeographic region through analysis of the applicable Yield tables. Most commonly, stand tree density declines in an exponential manner, which can be described by a function
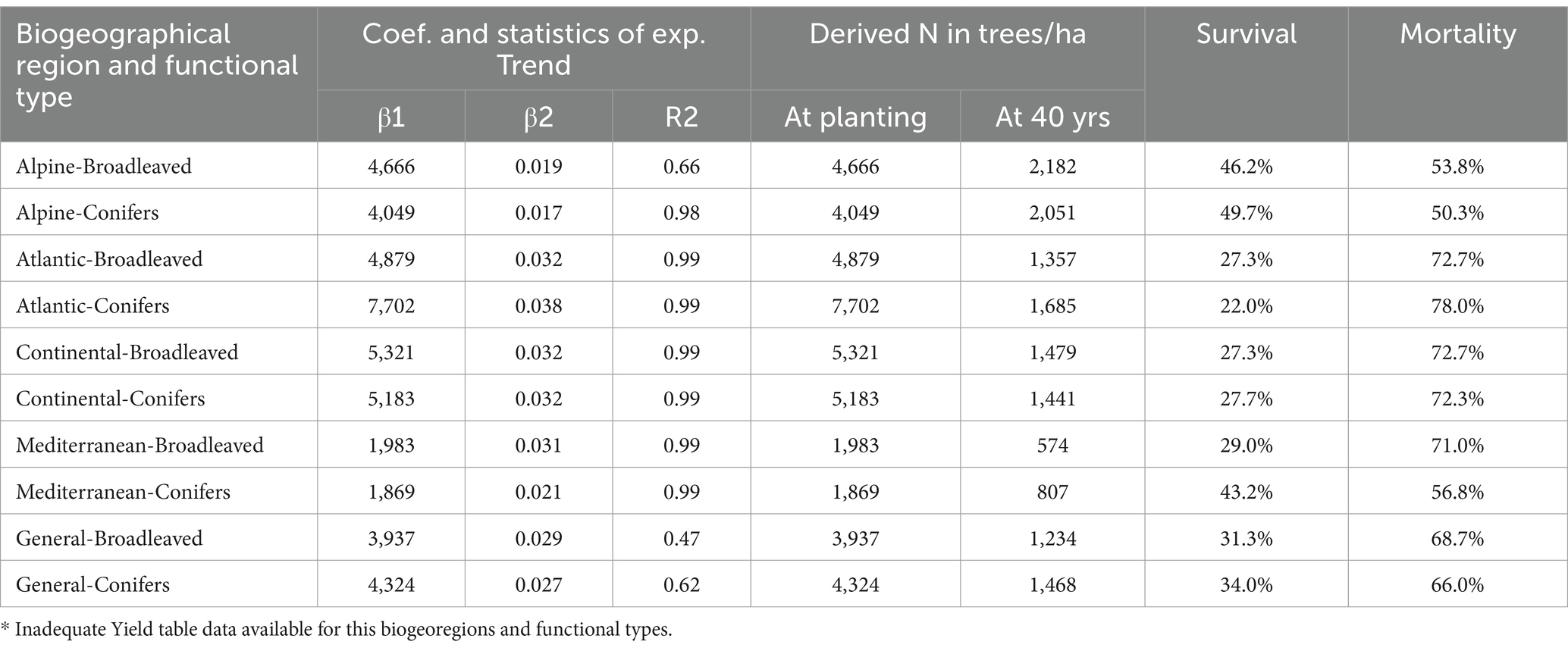
Table 2. Survival rate and mortality (natural and or due to management interventions) in timber plantations by biogeographic region, functional type (broadleaves and conifers) incl. surviving trees per ha at age 40 years compared to initial planting density.
Two additional functions, having the same form as described above, named “General broadleaves” and “General conifers” (continuous and dashed black lines in Figure 2), were derived from data across biogeographic regions, countries, tree species, functional types, and mean site indices. These two additional functions are showing the average age-related decline for broadleaves and conifers, regardless of the biogeographic region.
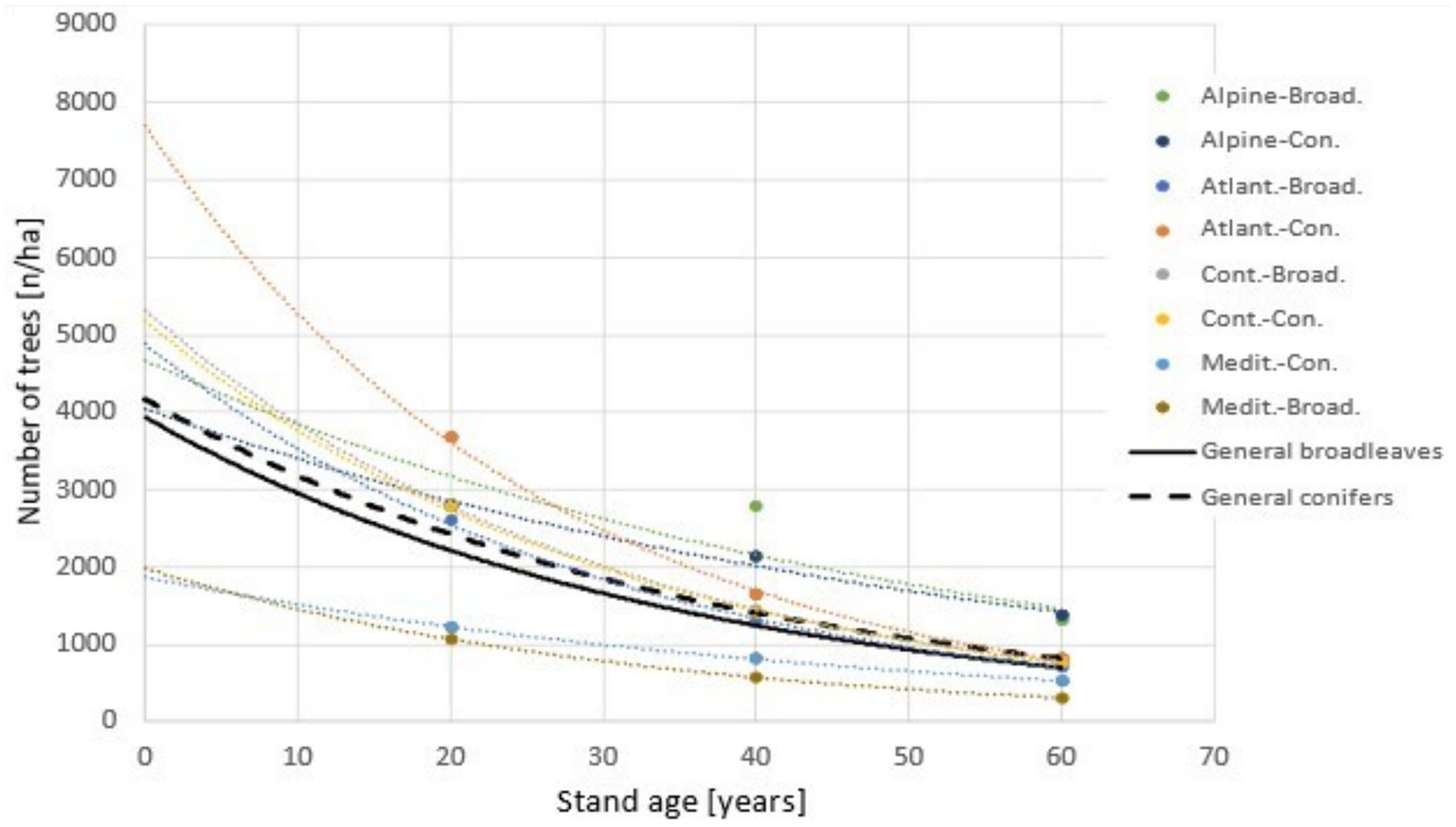
Figure 2. Estimated trends in the mean number of trees/ha over time by biogeographic regions and functional types, including an exponential smoother to highlight the trend in data.
2.8 From dry weight biomass to C, and from C to CO2 equivalent
To quantify the amount of carbon (C) in biomass, a factor of 0.5 is applied, reflecting that approximately 50% of dry weight biomass consists of carbon (IPCC, 2006). While this proportion varies slightly between species (see, e.g., Thomas and Martin, 2012), it serves as a standard estimate. The CO2 equivalent is calculated using the molar weight ratio of CO2 to C, i.e., 44/12.
3 Results
The tree level estimates by biogeographic region, country, species, functional type and reference site index (SI) are summarized in Table 3 and the aggregated assessment shown in Figure 3.
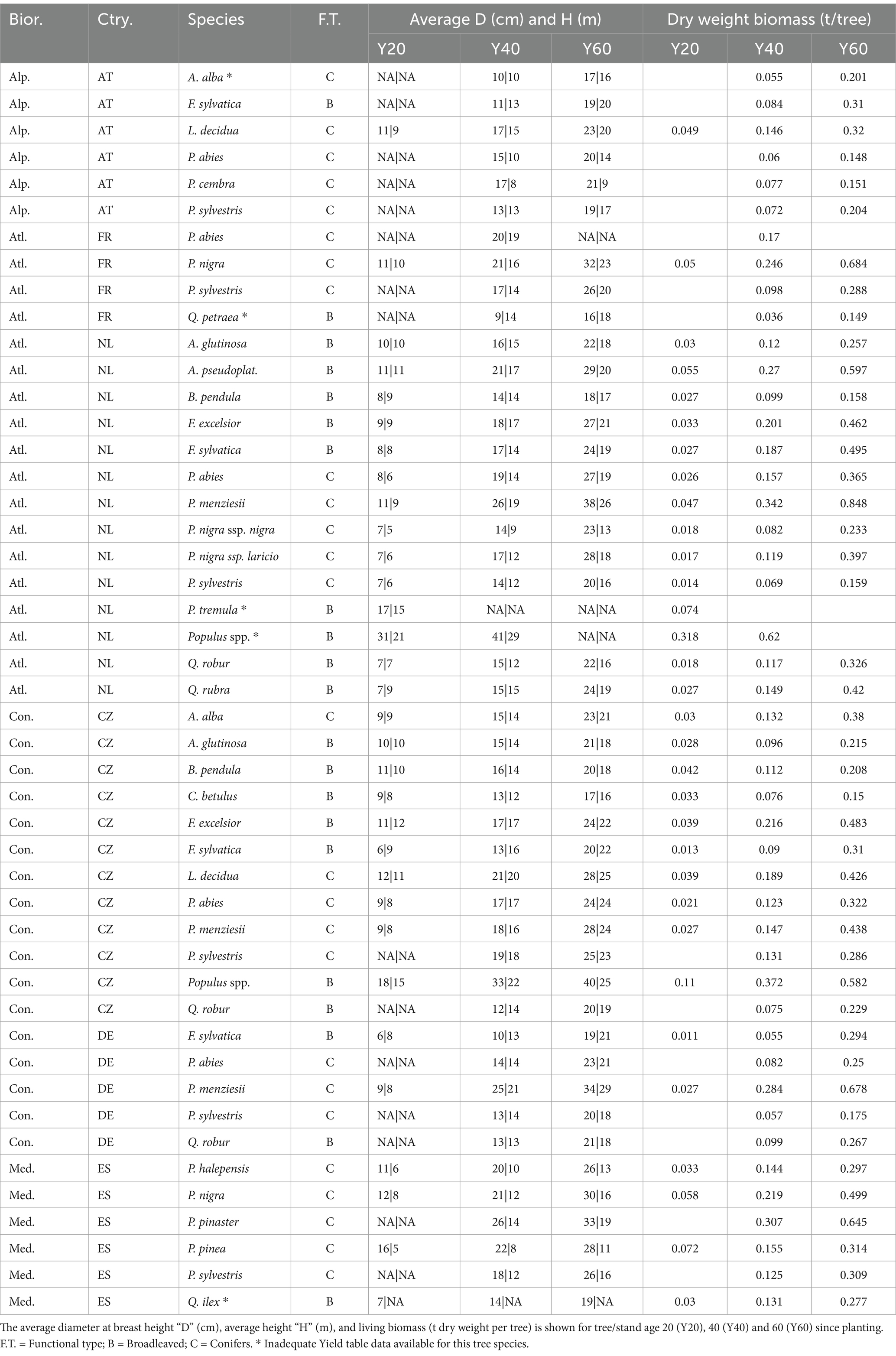
Table 3. Tree level estimates by biogeographic region, country, species, and functional type (Type).
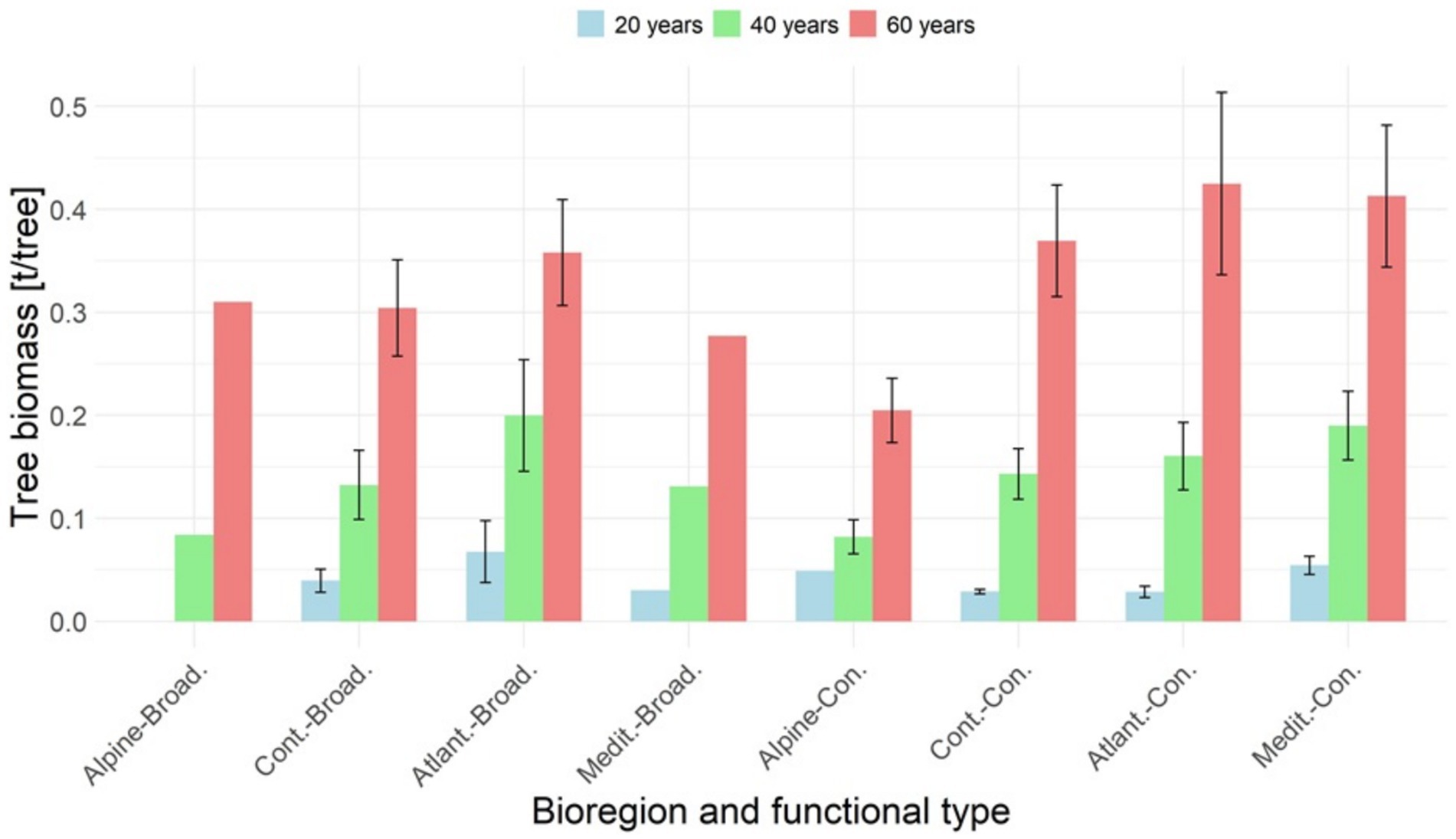
Figure 3. Living biomass in t/tree by biogeographic region and functional type, with broadleaves (Broad.) grouped on the left side of the graph, and conifers (Con.) on the right side of the graph. The bars indicate a mean value, with error bars showing the standard error if applicable.
3.1 Tree level biomass estimates
Figure 3 illustrates the average living biomass of individual trees at age 20, 40, and 60 years, categorized by biogeographic regions and functional types. Among these, the lowest tree-biomass is associated with the Alpine region, which aligns with stand densities (Figure 4, Table 4), but may also be influenced by the challenging environmental conditions at high altitudes. In these high-altitude areas, particularly for conifers, growth performance is limited by harsher environmental conditions, such as lower temperatures and poorer soils. The other species functional types and biogeographic regions (Continental, Atlantic, and Mediterranean) exhibit similar tree-biomass values.
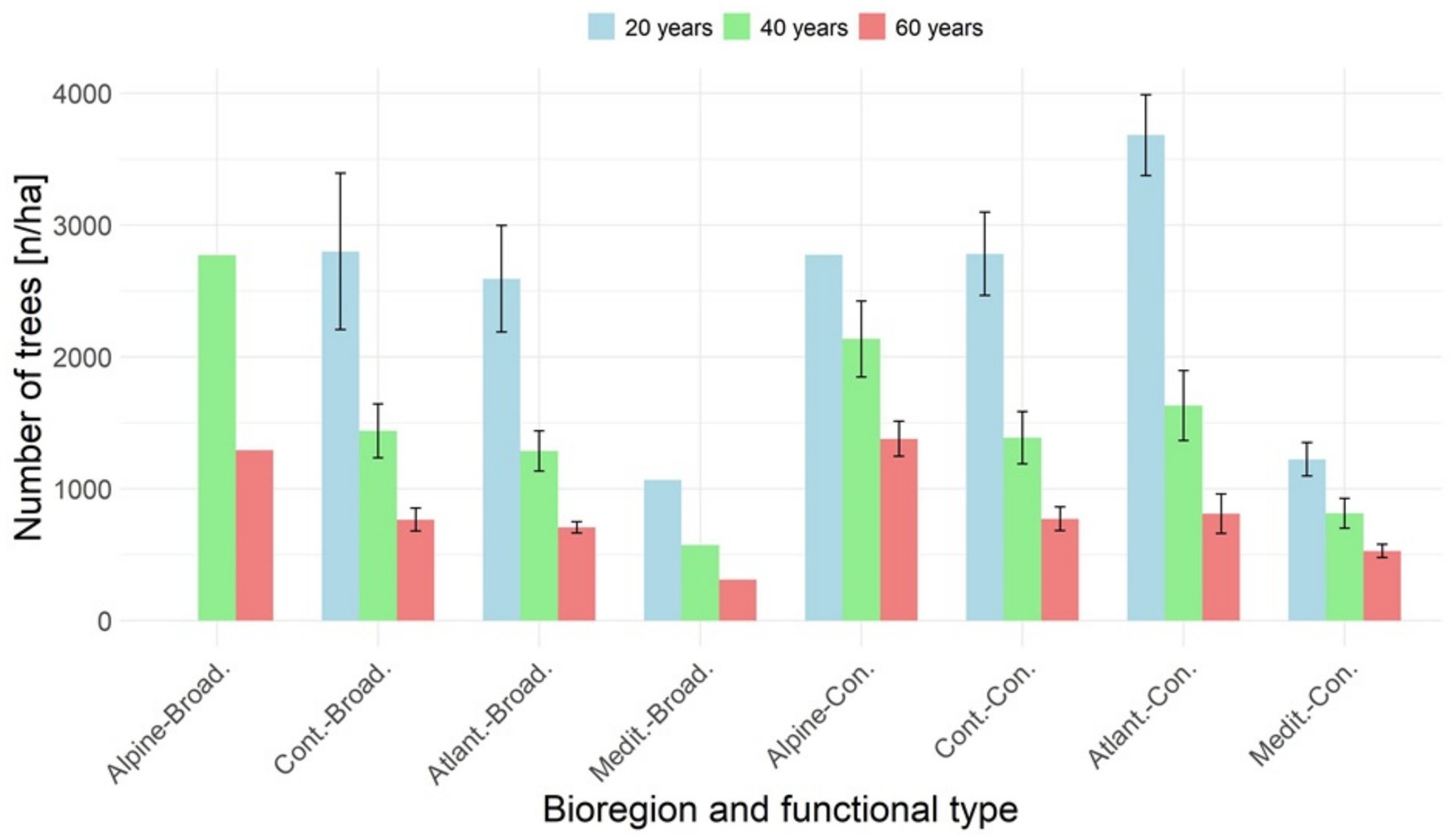
Figure 4. Assessed average number of trees per hectare for the age categories 20, 40 and 60 years, respectively, for the biogeographic regions and functional types.
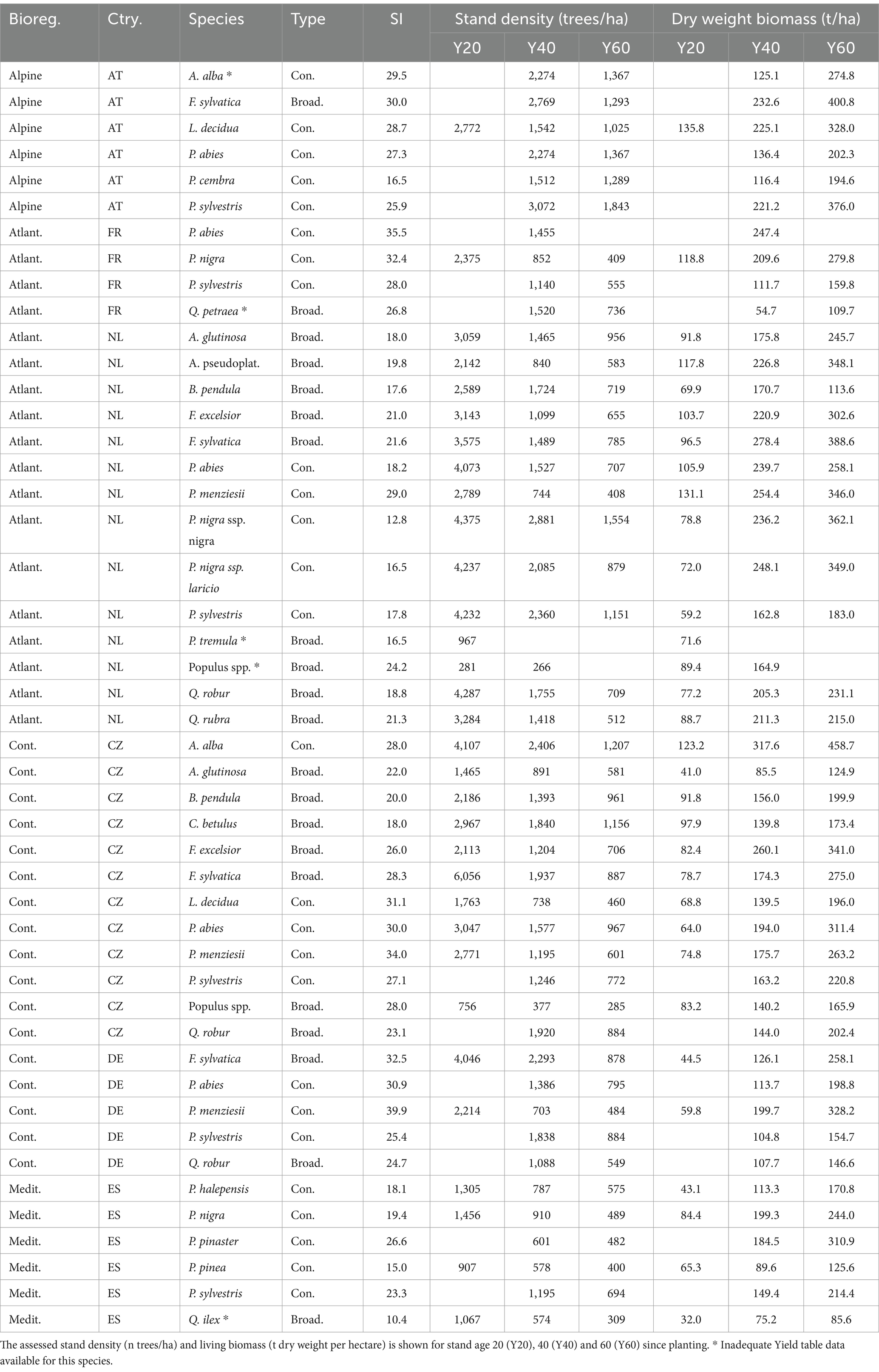
Table 4. Stand level estimates by biogeographic region, country, species, functional type (Type), and reference site index (SI).
By age 40, tree-biomass across biogeographic regions and functional types ranged on average from 0.08 to 0.20 t/tree, increasing to 0.20–0.42 t/tree by age 60. For a more comprehensive overview, Table 3 provides detailed country-specific biomass data by species.
3.2 Changes in stand density over time due to mortality and management interventions
For timber plantations, the age-related decline in mean tree density over time can be evaluated using Yield tables categorized by biogeographic regions, countries, tree species, forest group type, and mean site index at different ages. Figure 2 shows this age-related decline for managed even-aged stands by biogeographic regions and functional types, where the assessed number of trees/ha at specific age 20, 40, 60 years is estimated from the appropriate Yield tables (Table 1). As the trend is exponential, the function fitted to the processed data can be used to assess stand density along this age span, including the likely density at planting (Figure 2).
Stand densities for Alpine and Mediterranean broadleaves at 20 years of age were derived due to data unavailability. The ratio between the number of trees/ha at age 40 years and the estimated number of trees/ha at planting age represents the survival rate. Since this rate is derived from Yield tables, it accounts for both mortality and management interventions. Details of this assessment are provided in Table 2.
It is important to note that the survival rate based on Yield tables can only be applied to plantations that follow the expected planting density. For plantations where actual planting density is significantly lower (less than half the derived planting density), the survival rate correction is not applicable. The same happens when planting density is significantly higher (more than twice the derived planting density), since this could involve lower survival rate than expected.
3.3 Stand level biomass estimates
The stand level biomass estimates, applicable for timber plantations, are summarized by biogeographic regions and functional types in Figure 5 and Table 5.
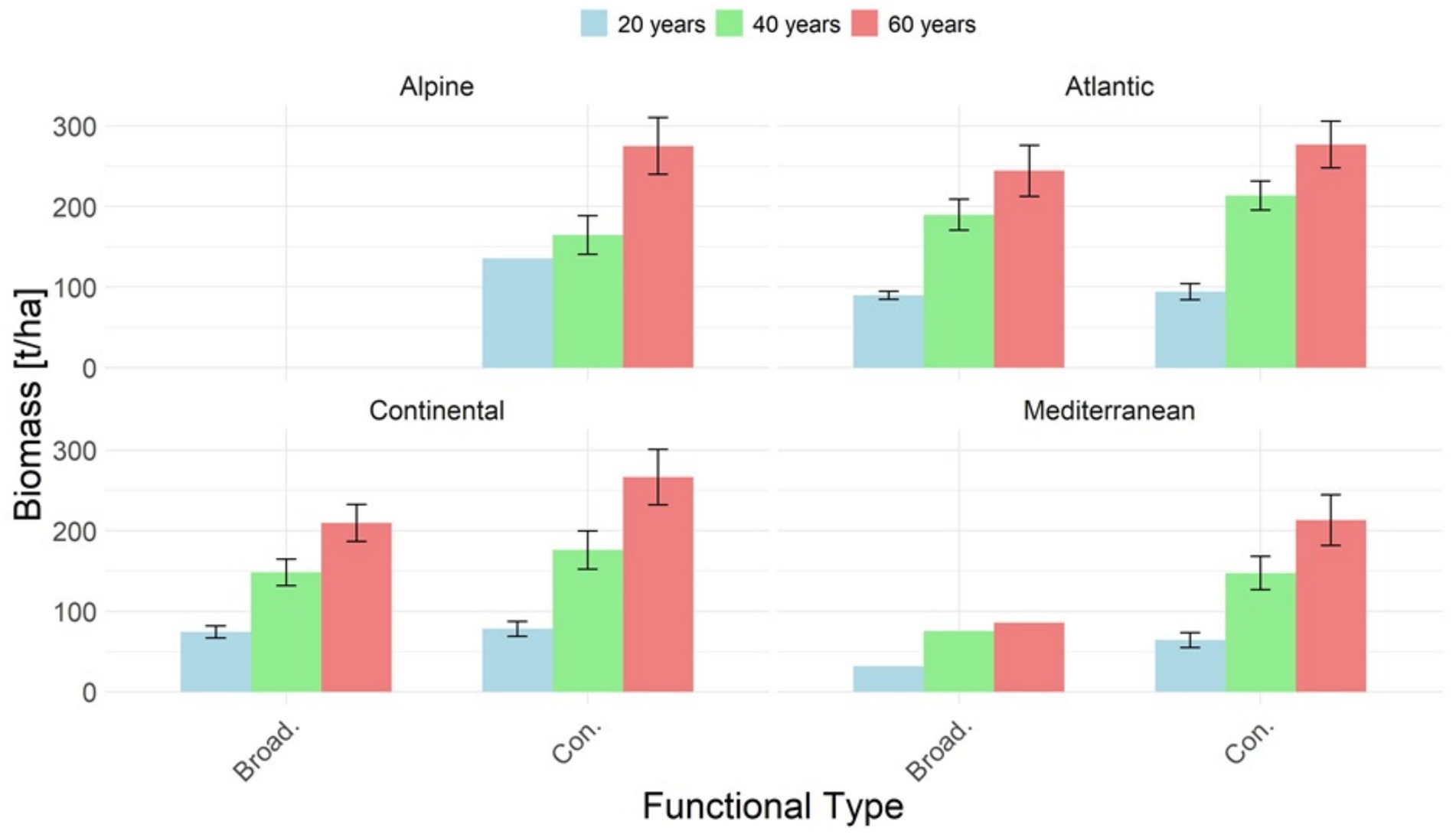
Figure 5. Living biomass in t/ha by biogeographic region and functional type. The bars indicate a mean value, with error bars showing the standard error if applicable.
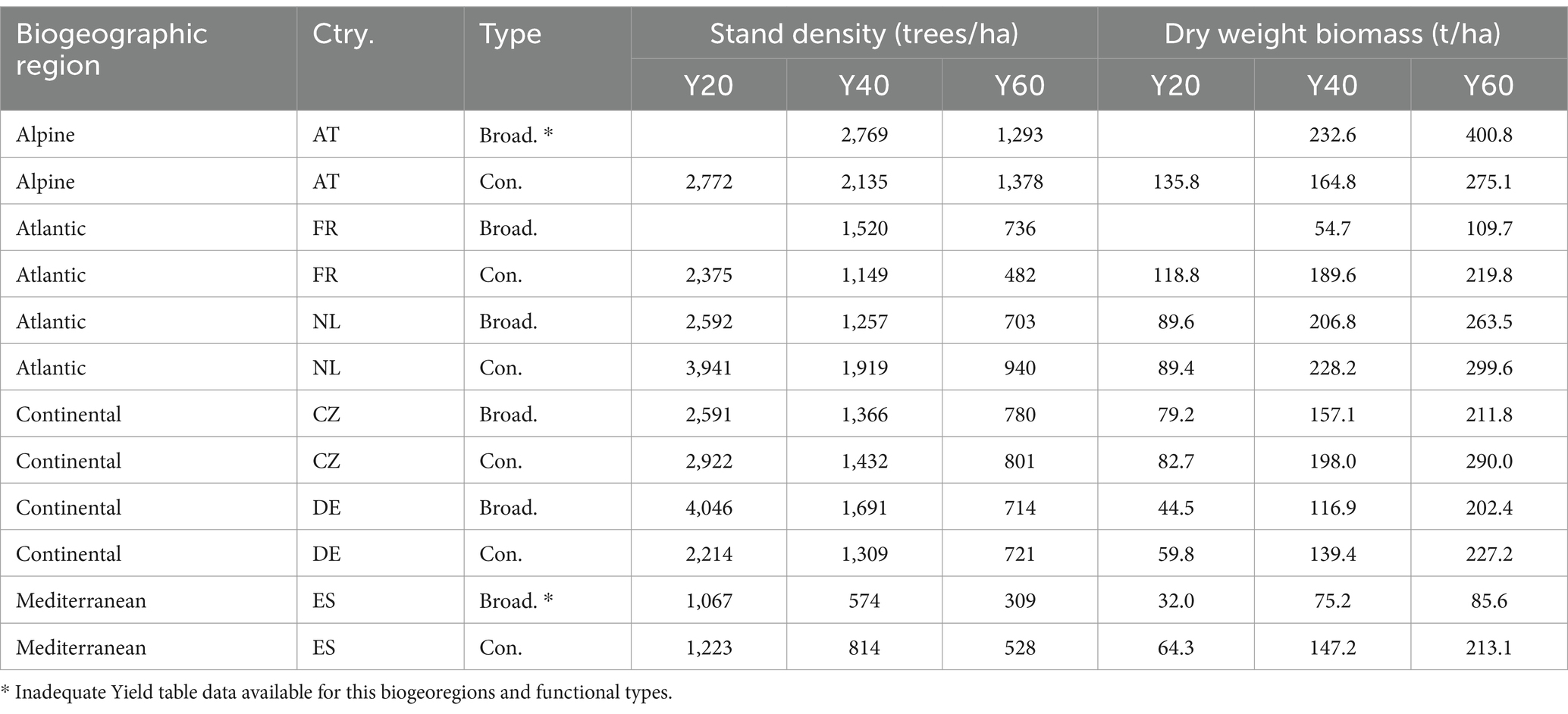
Table 5. Stand level estimates of living biomass (t dry weight per ha) by biogeographic region, country, functional type (broadleaves and conifers): underlying stand density (number of trees per ha) and dry weight biomass (t/ha), both for stand age 20 (Y20), 40 (Y40) and 60 (Y60) years.
Among all biogeographic region and functional types, the lowest stand-biomass is associated with the Mediterranean broadleaves, reaching 75.2 t/ha at age 40, and increasing to 85.6 t/ha at age 60 (Table 5). This lower biomass values in Mediterranean broadleaves are corresponding with stand density (Table 5, Figure 4).
While sufficient data were available for conifers in the Alpine region, the data for broadleaves in this region were too limited for meaningful comparison with other biogeographic regions. The limited data available were those of Fagus sylvatica in Austria (Tables 1, 5), which indicated a rapid biomass accumulation since early stand ages, reaching a biomass of 232.6 t/ha at 40 years and peaking at 400.8 t/ha at 60 years. This data should be verified and complemented with other sources, such as other Yield tables and/or country-specific NFIs.
Other functional types and biogeographic regions showed similar stand-biomass values, with the Atlantic region with somewhat larger biomass (Table 5, Figure 5). For a more complete overview of stand-biomass estimates, Table 4 provides the country-specific biomass data by species types.
4 Discussion
4.1 Main findings
This study provides a comprehensive assessment of biomass estimation for tree- and stand-level planting activities across different biogeographic regions in Europe (Alpine, Atlantic, Continental, and Mediterranean). We achieved biomass estimates for the mentioned biogeographic regions by using country and/or regional-specific data from Yield tables, NFIs, and NGHGIs. The following text provides a substantiation of the assessed biomass using the relevant literature.
4.2 Comparison with the IPCC, 2006
We compared our results with the prescribed IPCC default stand biomass values for forest ecosystems in comparable conditions as presented in Tables 4.7 (forests) and 4.8 (forest plantations) of Volume 4 of the 2006 IPCC Guidelines (IPCC, 2006). Since these data represent above-ground stand biomass, below-ground biomass fraction must also be added to these estimates, following Table 4.4 of Volume 4 (IPCC, 2006).
For the ecological zone “Temperate continental forest and mountain,” the IPCC (2006) gives an above-ground biomass for young forest ecosystems (≤20 years old) between 20 to 100 t/ha, which corresponds to a total stand biomass range between 28 to 140 t/ha. Next, specifically for plantations, the IPCC tables suggest a total biomass of 22 and 42 t/ha for broadleaves and conifers, respectively. These values are consistent with the estimates presented in our study, noting the fact that our estimates are applicable to age 20 years, not to an average biomass attributed to a range of the forest age class (1–20 years). Similarly, for medium and older stands attributed by IPCC by the age span of above 20 years, the derived IPCC default total biomass for coniferous and broadleaved plantation forests is 180–240 and 250 t/ha, respectively. This aligns well with the estimates for the Life Terra plantations expected at 60 years of age (Table 5).
For the Atlantic regions, the patterns of the assess biomass are similar, except of somewhat larger productivity for coniferous stands, which is about 300 t/ha according to the IPCC (2006) tables. This matches well the total biomass estimates for Atlantic biogeographic region for Life Terra coniferous plantations at the estimated age of 60 years (Table 5).
For the conditions of Mediterranean regions, the corresponding comparative IPCC biomass values are not available, and the region-specific comparison must rely on other evidence. Furthermore, biomass estimates for the Mediterranean region showed a similar range of values for conifer species as observed for the temperate region (Figure 5). On the contrary, the broadleaved plantations, represented only by even-aged stands of Quercus ilex, showed significatively lower biomass compared to other regions. This difference corresponds with stand density (Figure 4).
4.3 Comparison with other existing literature
4.3.1 Alpine biogeographic region
In our study, the Alps were considered a representative mountainous region for studying the Alpine biogeographic region. Since the early nineteenth-century, reforestation projects in the Alps were initiated, due to political changes, promoting most commonly naturally occurring conifer species including Picea abies and Larix decidua (Thom and Seidl, 2022).
These last-mentioned authors studied the ecosystem dynamic in Berchtesgaden National Park, southeastern Germany, by using permanent sample-plots, censused three times, i.e., in year 1984, 1996 and 2011. The species composition of this mentioned area at low elevations (< 800 m asl) is dominated by Fagus sylvatica. At the montane elevation belt, i.e., approximately from 800 to 1,400 meter above sea level, the species composition turns into a mixed forest of Picea abies, Fagus sylvatica and Abies alba. With increasing elevation, Picea abies gains dominance. Also, the subalpine elevation belt, i.e., from 1,400 meter above sea level to the timber (or tree) line, is composed by Picea abies, Larix decidua, and Pinus cembra. Although the Alpine region in our dataset (Table 1) contains mean tree and stand dimensions only from the Austrian Alps (Tyrol), the species composition fits with the one presented by Thom and Seidl (2022) for all the above-mentioned species. Therefore, it could be a good tool for estimating biomass in the Bavarian Alps as well.
Guidi et al. (2014) studied forest expansion on abandoned subalpine grasslands in Trentino (Southern Alps, Italy). Their study included two areas: an abandoned grassland now covered with 10-year-old Picea abies trees and shrubs (1,024 trees/ha), and an early-stage forest (age not specified) dominated by Picea abies (1,561 trees/ha). The 10-year-old stand had a mean DBH of 4.6 cm, and the estimated stand biomass was 6 t/ha, which corresponds with 0.006 t/tree. In the early-stage forest, the mean DBH was 11.0 cm, which corresponds with a total biomass of approximately 0.03 t/tree, based on the above-ground biomass equations used in their study and the IPCC, 2006 root-shoot ratio of 0.4 for conifer species with above-ground biomass < 50 t/ha in temperate mountain systems. In comparison with Guidi et al. (2014), our results for Picea abies indicate a total biomass of 0.06 t/tree for 40-year-old trees, corresponding to a mean DBH of 15 cm (Table 3).
We compared our results on broadleaves from the Alps with those reported by Smith et al. (2014), who developed biomass function for Betula pendula and Betula pubescens in Norway (Scandinavian Mountains). Their study provided data on stand density, mean tree dimensions, and biomass for these species, and they also compared their findings with the Norwegian NFI and datasets from Marklund (1987, 1988) and Bollandsås et al. (2009). The dataset used to develop the biomass functions of Smith et al. (2014) had a mean stand age of 50 years, a stand density of 1,449 trees/ha, mean DBH of 15.3 cm, a mean height of 12.0 meters, and above/ground biomass of 116.3 kg/tree. In comparison, the birch stands from Marklund (1987, 1988) had a mean age of 47 years (stand density not provided), a mean DBH of 12.7 cm and above-ground biomass 75 kg/tree. It is important to mention that all these mentioned datasets reflect typical forest conditions in the Scandinavian mountains, including a mix of natural regeneration and silvicultural treated stands, making them broadly applicable.
The total biomass for the species studied by the above-mentioned authors was derived using a root-to-shoot ratio of 0.24, as per the IPCC, 2006 table 4.4 (which applies to broadleaf species with above-ground biomass >150 t/ha in temperate mountain systems). This calculation resulted in a total biomass of 0.14 t/tree for Smith et al. (2014), and 0.09 t/tree for Marklund (1987, 1988). Our results are in close agreement with those of Marklund (1987, 1988) and Smith et al. (2014), with an average biomass of 0.084 t/tree for 40-year-old trees and a mean DBH of 11 cm (Table 3). Additionally, our stand density models (Table 2) suggest a stand density of 1,805 trees/ha for 50-year-old broadleaved species, which corresponds well with the mentioned studies.
4.3.2 Atlantic biogeographic region
In our study, the Atlantic part of France and the Netherlands were used as representative countries to estimate biomass at the Atlantic biogeographic region in Europe. According to Valade et al. (2018), 24% of the French forest is composed by Quercus robur (13.6%) and Q. petraea (10.4%). Including species such as Pinus sylvestris (5.9%), Pinus nigra (1.2%), and Picea abies (3.6%), our dataset (Table 1) is comprising the 34.7% of the species composition in French forests (including their Alpine, Continental and Mediterranean regions). Francini et al. (2024) used seven species groups to map the forest species area in the Netherlands combining Sentinel-2 harmonic predictors and national forest inventory data. All the species groups used by these mentioned authors are used in our dataset (Table 3).
Vos et al. (2023a, 2023b) studied forest biomass in the Netherlands, examining three key species: Fagus sylvatica, Pseudotsuga menziesii, and Pinus sylvestris. Their research documented stand characteristics including density, age, mean tree- and stand-dimensions, and biomass estimations.
For Fagus sylvatica stands at 46 years of age, they reported stand densities ranging from 1,100 to 840 trees/ha, with mean DBH ranging from 17 to 18 cm, yielding a total tree biomass up to 0.167 t/tree. Pseudotsuga menziesii stands aged between 59 and 66 years showed lower densities (127–240 trees/ha) but larger dimensions (mean DBH 44–52 cm), with biomass reaching over 1.103 t/tree at age 60 years. Pinus sylvestris stands aged 47–62 years exhibited intermediate densities (835–400 trees/ha) with mean DBH of 17–26 cm, and biomass values of 0.095 and 0.259 t/tree at 47 and 62 years, respectively.
Our findings both align with and differ from these Dutch studies in notable ways. For Fagus sylvatica, we estimated slightly higher individual tree biomass (0.187 t/tree at age 40)—approximately 10% above their values – despite our younger stand age. Conversely, our biomass estimation for both conifer species were lower: Pinus sylvestris showed 0.069 and 0.159 t/tree at age 40 and 60, respectively, while P. menziesii reached 0.85 t/tree at age 60 (Table 3). These differences in conifer biomass can be attributed to variations in stand density, age, and local site conditions (Table 4).
Our stand density models (Table 2) showed strong agreement with Vos et al. (2023a, 2023b) for broadleaved species, predicting 1,120 trees/ha at age 46. Although our models predicted higher densities for coniferous stands, this discrepancy likely reflects differences in species composition. In particular, our dataset includes species such as Picea abies, which typically forms denser stands compared to the conifer species studied by the mentioned authors, such as P. sylvestris and P. menziesii, which generally have lower stand densities Vos et al. (2023a, 2023b).
Peichl et al. (2012) studied the above- and below-ground biomass, carbon and nitrogen allocation in afforested grassland in southwest Ireland. The study was conducted 5 years after planting. The stand density was 3,242 trees/ha at the moment of measurement, consisting of 80% Fraxinus excelsior and 20% Alnus glutinosa. By using the equations and mean dimensions reported by these authors, we derived tree biomass for Ash and Alder trees, 0.7 and 1.18 kg/tree, respectively. These correspond with 2.5 t/ha, from which 1.8 t/ha for Ash and 0.7 t/ha for Alder.
The biomass of Peichl et al. (2012) cannot be directly compared with our results due to differences in age. Obviously, due to this difference in age, their results on biomass are lower than ours (Tables 3, 4). Regarding our stand density models (Table 2), broadleaved stands at the Atlantic biogeographic region could reach an amount of 4,158 trees/ha at age 5 years, which is somewhat higher than observed by Peichl et al. (2012). However, our models are based on more genus such as Fagus, Acer, Betula, and Populus (Table 4).
Van Damme et al. (2022) compared biomass in Scots pine stands for two forest management strategies in Brasschaat, Antwerp, in the Campine region of Belgium: (i) thinning and group planting of Quercus robur and (ii) clear cut, followed by replanting of young oak. The study focused on the first 15-year-period after the intervention. These authors did not find significant differences between the two types of forest management. Their study included results on stand density, age, mean dimensions and biomass. These authors reported that a 14-year-old Quercus robur stand exhibited a stand density of 1,225 trees/ha, and a mean diameter at breast height of 14 cm. According to the biomass equations used in this study, these trees are associated with a individual tree biomass of 0.08 t/tree (without leaves). This number corresponds with 103.01 t/ha.
Our results on biomass and stand density (Tables 3, 4) are lower than observed by Van Damme et al. (2022). This discrepancy corresponds with differences in DBH at a young age. Our analysis indicates that the diameter at breast height of Quercus robur at age 20, for both Atlantic and Continental biogeographic regions, is commonly around ≤8 cm (Černý et al., 1996; Jansen and Oosterbaan, 2018).
Dubois et al. (2021) studied natural-regenerated pure Betula pendula stands in Western Europe (temperate oceanic bioclimatic zone). Their study included DBH and stand age for different experimental management interventions aimed at producing large-sized logs. Our DBH results for Betula pendula in the Atlantic biogeographic region at age 20 fall within the lower range of values reported by these authors at 20-year-age class (from 11 to 20 years). Older stands reported by these authors (i.e., 40-year and 60-year-age classes) showed higher DBH values than those estimated in our study (Table 3). This discrepancy could be attributed to differences in site fertility, stand density, and management practices, as we consider regular management interventions, whereas their study focused on experimental interventions to increase productivity.
Wellock et al. (2011a, 2011b) studied forest plantations across various sites in Ireland, reporting mean tree and stand characteristics such as DBH and age. Most of the afforestation stands included in their study were monocultures—primarily coniferous plantations—although some mixed-species stands were also included. We estimated biomass from their results using above-ground biomass equations (i.e., Cienciala et al. (2006) for Pinus spp., Fiedler (1986) for Picea spp., Bunce (1968) for Fraxinus and Acer spp., Pretzsch (2000) for Fagus spp., and Jagodziński et al. (2018) for Larix spp.) and suitable IPCC, 2006 coefficients to derive the below-ground biomass component.
We compared our results to those reported by Wellock et al. (2011a, 2011b), excluding mixed stands due to the absence of species-specific input data. For Fraxinus excelsior, they studied only one 17-year-old stand, with a central DBH value (from the range reported) corresponding to a total biomass of 0.037 t/tree. For Fagus sylvatica, a single 42-year-old stand was reported, with a central DBH of 17 cm and a total biomass of 0.172 t/tree. For Acer pseudoplatanus, only one 20-year-old stand was studied, with a central DBH of 5.1 cm (including trees from 3.1 cm DBH), corresponding to 0.005 t/tree. Our results are in close agreement with those for Fraxinus and Fagus spp. (Table 3). The slightly higher values observed for Acer spp. likely reflect the inclusion of smaller trees (<7 cm DBH) in the dataset, as well as differences in site conditions.
For Picea abies, three stands were studied by Wellock et al. (2011a, 2011b). Among these, the 41-year-old stand was the most similar to our results (Table 3), showing a central DBH of 17.1 cm and a total biomass of 0.152 t/tree. For Picea sitchensis, seven stands in the 20-year-age class (11 to 20 years) were studied by these authors. These stands can be classified in four site conditions: (i) stands reaching a central DBH of 5.1 cm (total biomass ≤ 0.001 t/tree), (ii) stands reaching a central DBH of 10.6 cm (total biomass = 0.037 t/tree), (iii) stands reaching a central DBH of 17.1 cm (total biomass = 0.152 t/tree), and (iv) stands reaching a central DBH of 25.1 cm (0.356 t/tree). Our results for Picea spp. at age 20 in the Atlantic region (Table 3) fell between the medium and lowest fertility classes reported by Wellock et al. (2011a, 2011b).
For the Picea stichensis 40-year-age class (31 to 40 years), Wellock et al. (2011a, 2011b) reported three different fertility classes: DBH 10.6 cm (total biomass = 0.037 t/tree), DBH 17.1 cm (total biomass = 0.152 t/tree), and DBH 35.1 cm (total biomass = 0.709 t/tree). Our results for Picea spp. in the Atlantic region at age 40 (Table 3) fell slightly above the medium fertility class from the range reported by these authors.
For Pinus contorta, Wellock et al. (2011a, 2011b) studied one 20-year-old stand. This stand had a central DBH of 10.1 cm (total biomass = 0.035 t/tree), which is higher than our estimates for Pinus spp. in the Atlantic region (Table 3). This discrepancy may be attributed to differences in species, site conditions, and stand density. Wellock et al. (2011a, 2011b) also studied two Pinus contorta stands belonging to the 40-year-age class. These stands, together with the only one Pinus sylvestris stand studied by these authors, had a central DBH of 35.1 cm (total biomass = 0.649 t/tree), which is much higher than our estimates for Pinus spp. in the Atlantic region (Table 3). This discrepancy could also be related to differences in site fertility and stand density.
For Larix kaempferi, Wellock et al. (2011a, 2011b) studied three different stands. Among these, the most similar to our results was 22-year-old stand with a central DBH of 10.6 cm (total biomass = 0.046 t/tree). The genus Larix is missing in our results for the Atlantic region, however other conifers studied for this region, such as Pinus nigra in France and Pseudotsuga menziesii in the Netherlands, had a similar biomass and DBH values (Table 3).
4.3.3 Continental biogeographic region
In our study, Czech Republic and Germany were considered representative countries for assessing biomass within the continental biogeographic region of Europe. The main trees species of Czech Republic are: Picea abies, Fagus sylvatica, Quercus robur, and Pinus sylvestris (Černý et al., 1996). However, our dataset includes a broader range of species, comprising nearly all species listed in relevant national reports such as the Green reports of the Czech Ministry of Agriculture. According to the most recent German NFI, Quercus and Fagus genus together account for 28.1% of Germany’s forest area. When adding, Picea (20.9%), Pseudotsuga (2.4%) and Pinus (21.8%), these genera collectively represent up to 73.3% of Germany’s forested area (Bundeswaldinventur Ergebnisdatenbank, n.d.). Therefore, our dataset provides a robust basis for assessing biomass in both the Czech Republic and Germany (Tables 3, 4).
Cukor et al. (2022) investigated the characteristics of 14-year-old forest stands stablished on agricultural lands in the Czech Republic. Their results included the stand density at planting and after 14 years, mean tree dimensions, wood volume and total biomass. For broadleaved species, the mean stand density at age 14 was 4,305 trees/ha, with Fagus showing the highest density (5,280 trees/ha) and Quercus the lowest (2,940 trees/ha). The mean survival rate for broadleaved species was 59.5%, varying from 79.3% for Tilia to 29.4% for Quercus. Accumulated biomass over a 14-year period averaged 61.25 t/ha, ranging from 14.8 t/ha for Fagus to 104.3 t/ha for Acer. When calculated per tree, the average biomass was 0.015 t/tree, with Fagus showing the lowest (0.003 t/tree) and Acer the highest (0.025 t/tree).
For conifers, represented by Picea, the stand density was 4,220 trees/ha at age 14, surprisingly higher than the initial planting density of 4,000 trees/ha, making survival assessment impossible. The species achieved a biomass of 72.3 t/ha (0.017 t/tree).
Our stand density models (Table 2) projected a 63.9% survival rate for continental broadleaved species at age 14, slightly lower than Cukor et al.’s (2022) observations. Mortality comparisons for conifers were not possible due to the unexpected increase in Picea density over time in their study.
Our 20-year-old Fagus sylvatica trees showed higher values (0.013 t/tree) compared to the 14-year-old Fagus reported by Cukor et al.’s (2022), while remaining within their reported biomass range. For Quercus, biomass estimation at age 20 was not possible due to lack of data (Černý et al., 1996; Albert et al., 2021). While Cukor et al. (2022) included Tilia and Acer genera in their study, these species were not present in our dataset (Table 3). The mean biomass at age 20 for our broadleaved species in this region (excluding Populus cul.) was 0.028 t/tree, exceeding Cukor et al.’s (2022) observations - a difference that could be attributed to differences in age and site productivity. Among conifer species, our 20-year-old Picea abies exhibited slightly higher biomass (0.021 t/tree) than their 14-year-old stands, as expected given the difference in stand age.
Werner et al. (2024) observed that, in some 8-year-old German forest plantations originated in 2013, the total stand-biomass depended on the species composition, showing average biomass values of 33.7 t/ha (range 21.5–40.5 t/ha) for monoculture broadleaved-stands, and 48.8 t/ha (range 41.3–53.5 t/ha) for mixed broadleaved-stands. Furthermore, these authors reported survival rates over the 8-year period on these young plantations, showing an average survival of 77.3% in broadleaved monoculture stands, and 86.9% for mixed-broadleaved stands (with a maximum of 83.3% at the mixed Acer platanoides and Tilia cordata plantation).
A direct comparison of our biomass estimates with those of Werner et al. (2024) is not feasible due to differences in the age of the stands studied. Our study focuses on forest stands ranging from 20 to 60 years in age, while these authors centred on much younger, 8-year-old forest stands. However, using our stand density models to 8-year-old broadleaved stands revealed a similar survival rate (77.4%).
Jagodziński et al. (2018) studied Larix decidua forest stands in Western and Southern Poland. These authors studied 12 stands in total, one from each 10-year-age class. Their results included stand age, DBH, heigh, stand density, and biomass. Our biomass estimates are lower (about 13%) than those reported by these authors at 40-year-age class (34 years). For the 20-year and 60-year age classes, this difference increases, reaching 43 and 49%, respectively, with our results being lower than those reported by these authors. These discrepancies correspond to differences in site fertility, tree-dimensions, and stand density (Table 3).
4.3.4 Mediterranean biogeographic region
In our study, Spain is considered a representative country to study the Mediterranean biogeographic region in Europe. Agroforest systems, such as Dehesa, are the forest-formation-type that covers most of the wooded-forest-area in this country (15%). This forest-formation-type is usually composed of Quercus spp., such as Q. ilex and Q. suber, Olea europaea and Fraxinus spp. Quercus ilex forest stands are the second largest forest-formation-type (14%), followed by Pinus halepensis stands (11%) (Bravo et al., 2017; Yearbook of Forest Statistics 2021, 2023; Bosques españoles y su evolución, n.d.). According to these last-mentioned authors, by adding Pinus sylvestris (5%), Pinus pinaster (4%), Pinus pinea (2%) and Pinus nigra (4%) we could assume that our dataset (Table 1) is covering 40% (up to 55% if agroforest systems are included) of the total wooded-forest area.
Q. ilex is primarily managed as coppice in Spain, making coppice management highly relevant in the Mediterranean biogeographic region (Ibàñez et al., 1999; González Molina, 2005; de Rigo and Caudullo, 2016). However, coppice rotation periods are typically under 30 years –shorter than the rotations permitted in Life Terra plantations. Yield tables for this species often reflect unmanaged or uneven-aged stands that are not applicable for Life Terra (Espelta et al., 2009; Centre de la Propietat Forestal, 2012; Berta et al., 2019). Since afforestation and reforestation activities typically result in even-aged stands, analyzing uneven-aged stands is irrelevant to our focus. Therefore, biomass estimation for this tree species was based solely on Yield tables for ordinary managed even-aged stands (Table 1) and should be interpreted with care.
Palacios-Rodríguez et al. (2022) studied a 26-year-old afforested cropland in Cádiz, southern Spain, planted with Ceratonia siliqua (330 trees/ha at planting). They reported an average dry biomass of 0.046 t/tree (mean DBH = 12.8 cm) and a remaining stand density of 293 trees/ha. This plantation was pruned but never harvested neither fertilised. In comparison, our results for Mediterranean broadleaves, represented by Quercus ilex, showed a total biomass of 0.03 t/tree (mean DBH = 7 cm) and a stand density of 1,067 trees/ha at age 20 (Tables 3, 4). The differences in tree dimensions can be attributed to variations in age, species, site productivity, and stand density.
Calama et al. (2024) investigated young afforestations in Spain using data from permanent sample-plots. Their analysis included stand age, stand density, mean tree dimensions, and total biomass estimates. For Q. ilex stands with an average age of 14 years (ranging from 1 to 30 years), they reported a total biomass of 7,383 kg/ha and a stand density of 756 trees/ha. This corresponds to approximately 0.01 t of biomass per tree. Our results for Q. ilex at age 20 showed higher biomass values (0.03 t/tree) compared to those reported by the authors. This difference could be attributed to differences in age and site productivity.
Other broadleaved species studied by Calama et al. (2024), all from Quercus genus at age 13–15 years, exhibited similar individual tree biomass values to Quercus ilex. However, stand densities varied significantly, ranging from 549 trees/ha for Q. suber to 1,367 trees/ha for Q. robur. These stand density values are consistent with our density models for broadleaved mediterranean species, which predict a stand density of 1,285 trees/ha at age 14 years (Table 2).
Calama et al. (2024) also analyzed conifer species from the Pinus genus, with stand age averaging around 20 years (ranging from 1 to 44 years). By deriving mean individual tree biomass values as we did above, the resulting biomass were approximately 0.04 t/tree for P. pinaster, 0.03 t/tree for P. sylvestris, 0.02 t/tree for P. halepensis, 0.02 t/tree for P. pinea, and 0.05 t/tree for P. nigra. These estimates align closely with our biomass results at the individual tree level (Table 3), with P. pinea being the notable exception–our results indicate higher biomass values (0.07 t/tree at age 20, for a stand density of 907 trees/ha). This discrepancy in biomass may be attributed to differences in stand characteristics, as Calama et al. studied younger trees (15 years) in denser stands (up to 2,375 trees/ha), which typically results in smaller individual tree biomass.
Pinus halepensis presented the lowest mean stand density among the species studied by Calama et al. (2024), with an average of 862 trees/ha and a range of 178–2,375 trees/ha. P. sylvestris showed the highest mean stand density, 1,443 trees/ha, ranging from 504 to 2,900 trees/ha. These mean stand density values are corresponding with our modelled estimates for Mediterranean conifers, which predict 1,228 trees/ha at age 20 years (Table 2).
Del Río et al. (2017) analyzed the effects of forest management on biomass in mediterranean pine forests. Their study, conducted on Pinus pinaster stands in Fuencaliente, Ciudad Real (Spain), provided stand level data on mean tree dimensions, stand density and above-ground biomass. Using their results and applying the below-ground biomass equations of (Ruiz-Peinado Gertrudix, 2013), we derived total tree biomass values ranging from 0.11 to 0.15 t/tree at age 33 and from 0.36 to 0.62 t/tree at age 59. Our biomass results for Pinus pinaster at age 60 years are around 5% above the range observed by these mentioned authors (Table 3).
The stand density of Pinus pinaster according to these last-mentioned authors ranged from 1,193 trees/ha to 1,570 trees/ha at age 33, and from 340 to 870 trees/ha at age 59. This range corresponds to the intensity of the management interventions. In comparison with (del Río et al., 2017), our stand density models are lower at age 33 (935 trees/ha) and aligned at age 59 years (541 trees/ha) (Table 2).
Hernández-Alonso et al. (2023) estimated carbon content in above- and below-ground tree biomass in mixed forest stands under Mediterranean conditions in Central Spain (Sierra de Francia-Quilamas and Sierra de Gredos) using sample plots. These mixed stands consisted of species from the genus Juniperus, Pinus, Arbutus, Castanea, Fraxinus, Ilex, and Quercus. Their results included stand density, age, and basal area, from which the mean diameter at breast height was derived. The estimated carbon content in the above- and below-ground tree components was 122.55 Mg C/ha, corresponding to an average biomass of 0.25 t/tree at age 51. For sample-plots with a mean age ≤60 years, the diameter at breast height ranged from 11.7 to 35.5 cm.
The findings of Hernández-Alonso et al. (2023) correspond with our results. The diameter at breast height and biomass ranges in our dataset correspond well with the measurements from their sample plots for stands aged ≤60 years (Table 3). Although our stand density models are derived from non-mixed forest stands, our results (Table 2) are consistent with the stand density data reported by these authors, especially for the stands located in Sierra de Gredos. This comparison is shown in (Figure 6) where the observed densities reported by these authors should fall between our conifer- and broadleaved-predicted densities. However, the slight systematic underestimation of stand density at increasing stand age suggests that our projections may be somewhat conservative for Mediterranean regions.
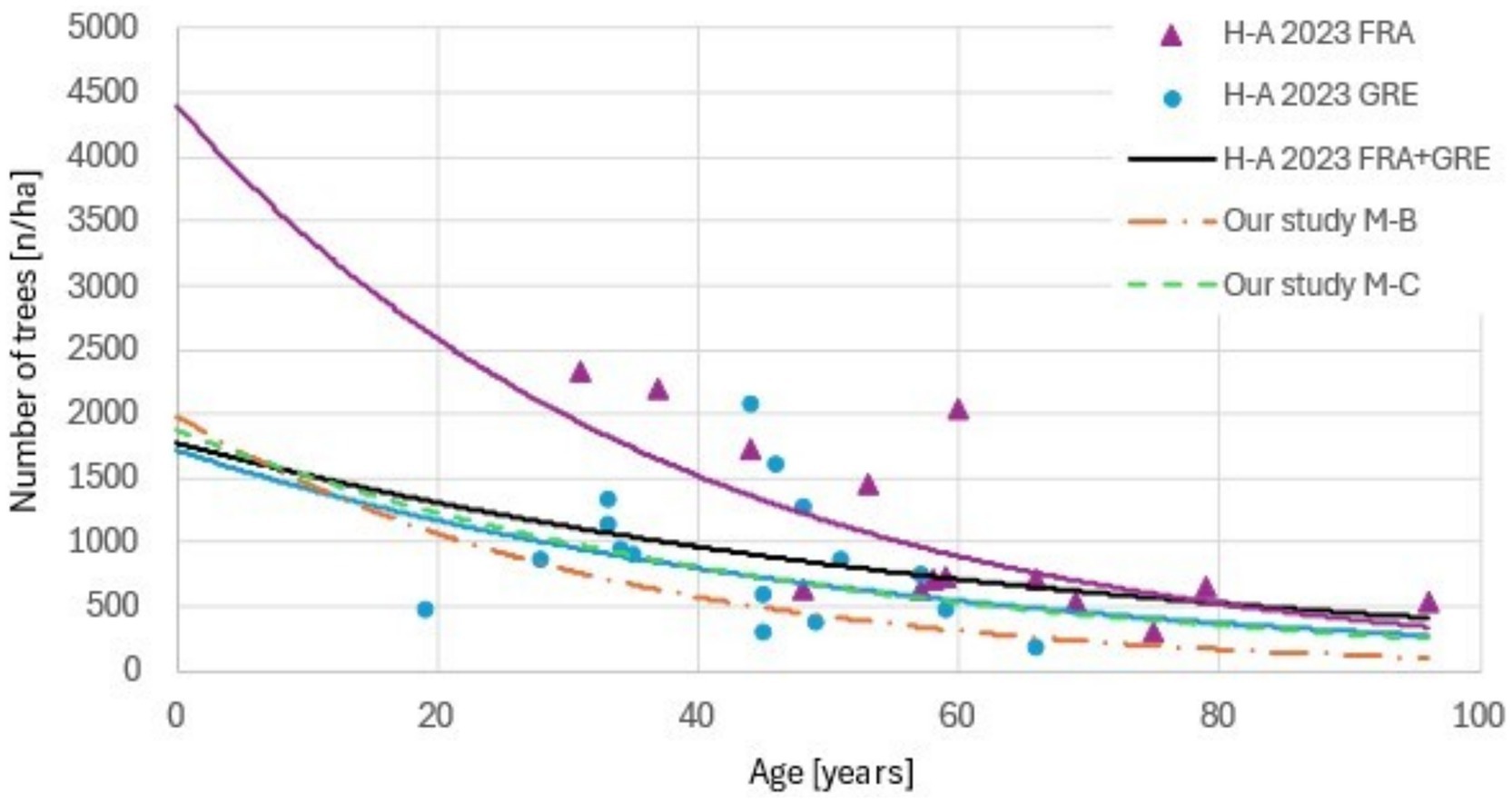
Figure 6. Comparison of our stand density models with stand density data extracted from Hernández-Alonso et al. (2023). “H-A 2023 FRA” and “H-A 2023 GRE” represent data from the Sierra de Francia-Quilamas and Sierra de Gredos localities, respectively. “M-B” and “M-C” denote Mediterranean broadleaves and Mediterranean conifers, respectively.
The higher stand densities reported by Hernández-Alonso et al. (2023) could be attributed to the relatively lower water deficit in their study areas compared to others of Spain. For instance, Gredos, with an annual rainfall of 500 mm according to the authors, would likely fall into the upper dry to sub-humid category, while their other study site would be classified as humid. Additionally, their species composition includes Juniperus, Arbutus, and Ilex, which typically develop as large shrubs or relatively small trees, potentially increasing stand density estimates.
Espelta et al. (2009) developed growth models for Q. ilex ssp. ballota and Q. ilex ssp. ilex in the Mediterranean region, focusing on Catalonia, Spain. Using data from the Spanish NFI cycles 2 and 3, they created models to describe DBH development over time across various stand densities, as illustrated in their Figure 12.5 (exact values could not be retrieved).
Our DBH projections at ages 40 and 60 years align with Espelta et al.’s (2009) findings, though trending slightly below their reported average values. While our DBH at age 20 is noticeably lower than their values, this discrepancy can be attributed to a methodological constraint in their study: their diameter growth curves begin at 7.5 cm (the minimum diameter threshold for the Spanish NFI data inclusion) for one-year-old plantations, which introduces an upward bias in the early-stage diameter estimates.
Centre de la Propietat Forestal (2012) studied management models for Q. ilex ssp. ballota and Q. ilex spp. ilex. Our results (DBH = 13.7 cm at age 40 years; Table 3) match their average site conditions for even-aged Q. ilex ssp. ballota stands, corresponding to their model “Qib02.” For even-aged Quercus ilex ssp. ilex, our results fall slightly below their average site conditions (between models “Qii06” and “Qii07”). However, the stand biomass values derived from the Centre de la Propietat Forestal (2012) data present challenges due to exceptionally high stand density at young stages (approximately 4,300 trees/ha at age 20). This resulted in unusually high estimated stand biomass values: 241.5 t/ha for 20-year-old even-aged Quercus ilex ssp. ballota stands and 207.3 t/ha for 20-year-old Q. ilex ssp. ilex stands of the same age.
While our individual tree estimates align with both Espelta et al. (2009) and Centre de la Propietat Forestal (2012), we excluded the stand-level biomass comparisons from our result due to the unrealistic stand density values and resulting stand biomass estimates in the reference data.
4.4 Implications
This study could be used as a template to assess biomass and carbon stock for afforested areas in Europe, which is an important metric for climate change mitigation strategies and policy planning, including initiatives like the Paris Agreement and LULUCF activities.
Since our methodology is based on Yield tables and NFIs, it is assuming full stocking. This offers an objective way for forest managers to make decisions about forest stand management (Burkhart et al., 2019). For example, for mean site productivity conditions (average site index) one could estimate actual stocking by comparing our results on Table 4 and the actual values observed at the field for a given stand. If the assessed actual stocking is below 80%, intensive thinning or clearing should be avoided to allow the forest to recover. Conversely, when the actual stocking exceeds 110%, managers might choose more intensive thinning or clearing to optimize stand health and productivity. These thresholds are general examples based on the Czech law (Forest Act, 1996), which includes the intensity of thinning depending on the species composition, stocking, and stand age.
Also, decisions regarding replanting can be guided using stand density models, as showed in Table 2. For instance, if we establish that stand density in stands younger than 20 years should not fall below 70% (with 100% representing the values derived from the models), one could decide objectively if it is needed to replant, and if so, how many trees per hectare are needed.
Therefore, our findings not only enhance biomass or carbon stock monitoring but also support sustainable forest management strategies. It is important to mention that these mentioned thresholds for thinning and replanting are just examples, and users (or future studies) should use country-specific thresholds.
4.5 Study limitations and recommendations
4.5.1 Associated challenges of estimates
While our study provides valuable insights into biomass estimation, its scope is limited by the specific planting objectives–it is primarily focused on timber plantations–, the geographic range, and the sample size of tree species analyzed. Nevertheless, our results align generally well with the country-specific literature reviewed, supporting their reliability and applicability. The observed variability in biomass across sites highlights the importance of site-specific factors in influencing growth and stand dynamics.
4.5.2 Shrubs and trees
While shrubs play a crucial role in enhancing biodiversity and preventing soil erosion, their contribution to biomass is relatively limited. That is why there is a lack of comprehensive data on the biomass of these species.
Hamelin et al. (2015) studied the above-ground biomass of buckthorns (Frangula alnus) under two different environmental conditions: in an open field and a plantation, located at Sainte-Catherine-de-Hatley, in Southeastern Québec, Canada. The authors developed allometric equations using age as the independent variable (ranging from 2 to 26 years) and total above-ground biomass as the dependent variable. The expansion factor from the table 4.4 of IPCC (2006), value 0.46 (representing “Domain = Temperate, other broadleaf ABG < 75 t/ha”) could be used to estimate the below-ground biomass component of this species. Applying this calculation results in a total biomass of 0.01 t/tree at age 20 years.
However, the biomass of this species should be used with care when aggregating species groups or types. If Frangula alnus is used to estimate the biomass of other species, it should be applied only to small broadleaved trees or large shrubs that reach a maximum height of 6–7 meters (non-merchantable trees), such as Crataegus monogyna. It should not be applied to small shrubs like Rosmarinus officinalis, as doing so could lead to an overestimation of the biomass.
Montero et al. (2020) developed models to predict variations in shrub-species dimensions, such as DBH and height, in Spain. These authors also parameterized biomass equations with canopy cover and mean height as independent variables. These equations may be applicable to the Mediterranean region when canopy cover and mean height are known. However, predicting canopy cover and survival rates for shrubs over a 40-year period involves considerable uncertainties. Due to these inherent uncertainties related to shrubs, the Life Terra project rigorously excludes these carbon and CO2 estimates from accounting while stressing other benefits of planting shrubs.
4.5.3 National Forest Inventories and National Greenhouse gas Inventories
Country-level site index is a suitable indicator for large-scale forest production assessments and intercomparison of growth performance among sites and regions. This assessment requires the analysis of National Sample-based Forest Inventories which are the major data source on European forests.
There is a wealth of published approaches for estimating the biomass of trees, especially for those with merchantable dimensions. Several countries of Europe use biomass functions to estimate the biomass of their forests and submit the results in their annual NGHGI. These NGHGI reports serve as a valuable resource for extracting relevant country-specific biomass equations, offering a practical tool to access the most applicable models. Given the complexity and effort involved in searching and analyzing the numerous published equations, these reports provide an efficient means of obtaining the necessary data for biomass estimation (Palmero-Barrachina et al., 2023).
4.5.4 Survival rate and management considerations
Since the estimation described in this text focuses on the biomass that trees would have at age 40 years, it is important to consider the uncertainty associated with the assumed survival rate, i.e., tree mortality and management interventions reducing the stand density over time.
In Life Terra’s timber plantations, it is assumed that the stand density at planting aligns with the values analyzed in the Yield table models. However, this assumption does not necessarily correspond to reality. While some timber plantations implement regular management interventions as prescribed in Yield tables, others do not. The correction of biomass estimates due to survival rate based on Yield tables can only be used for plantations that use the expected planting density (Figure 3 and Table 2). If the actual planting is significantly smaller (less than half of the derived planting density) or significantly higher (more than twice the derived planting density), the survival rate correction is not applicable (Cienciala et al., 2022). This issue is related to the use of the stocking mentioned in the implications section. Further research could focus on deriving specific survival rates for these cases.
Estimating tree survival is a complex process influenced by various abiotic (e.g., climate) and biotic (e.g., insect outbreaks, pests, diseases) factors, and their interaction with global warming (Seidl et al., 2017). Increased CO2 concentrations, temperature increases, altered precipitation patterns, and more frequent or intense disturbances—including droughts, beetle infestations, and wildfires—can significantly impact tree survival (Hartmann et al., 2022; Robbins et al., 2022; Korená Hillayová et al., 2023; Pretzsch and Grote, 2023). The result of the interaction of such factors is often difficult to anticipate. This makes the anticipated survival rates uncertain, as traditional models such as Yield tables are based on historical growth patterns and only project average stand characteristics over time, without accounting for the unpredictable nature of local disturbances and threats like droughts and beetle infestations. Finally, note that no projection may fully capture regional variations due to unexpected impacts related to global warming on tree survival (Hartmann et al., 2022).
4.5.5 Monitoring
Future research should focus on verifying and improving our findings through field measurement. One promising approach is to utilize sample-based forest inventories to monitor the dynamics of Life Terra plantations. These inventories would provide valuable, site-specific data on stand growth, biomass accumulation, and survival rates, allowing for the validation of our models and insights.
4.5.6 Applicability
Our methodology focuses on Life Terra timber plantations in four biogeographic regions (Alpine, Atlantic, Continental, and Mediterranean). Life Terra has also planted trees in the Boreal region and encompasses five additional planting categories beyond timber plantations: (i) ecological restoration projects, (ii) agroforestry systems, (iii) gardens, (iv) green infrastructure projects and (v) others. Therefore, it is important to discuss how our methodology can be applied to (a) other biogeographic regions and (b) non-timber plantations for which we currently have no information. As for (a) note that we have not elaborated examples from the boreal biogeographic region due to the (as of date) limited project exposure of Life Terra there. However, specifically Boreal region is fully suitable for biomass estimation following our methodology given the wealth of local allometric equations and growth and Yield tables available there. As for (b) Life Terra project works with other types of plantations as listed above for which literature is severely limited and trustable biomass estimation would require additional analytical effort and empirical evidence. Therefore, for these environments Life Terra adopted, e.g., a conservative survival rate assumption of 50% at age 40 (Calzada and Millán, 2004; Pausas et al., 2004), which should be further verified.
Our approach uses average site conditions to predict tree growth performance. It is possible that a significant part of the reforestation/afforestation activities would be carried out in unproductive or less productive sites. In such cases, users should use a site index lower than the average fertility class. However, using values of low site index also involves some uncertainties. Further studies could determine different values of site index to determine the entire range on fertility classes, for example: low-medium-high.
Our framework uses country-specific available information, including local allometric equations, Yield tables, NFIs, national submissions on GHG inventories for the LULUCF sector under UNFCCC. One potential enhancement to our methodology could involve creating categories based on forest types for example, using definition from the European Environment Agency (Barbati et al., 2007). This categorization could make our methodology more ecologically meaningful. Note however, that the accuracy and robustness of the estimates will primarily depend on local growth and Yield tables and suitable allometric equations.
5 Conclusion
This material presents the methodological approaches and assumptions used to assess tree and stand biomass in Life Terra plantation activities, with a particular focus on timber plantations. The assessment incorporates relevant literature available as of summer 2024. In the discussion section, we provide a detailed comparison of our biomass estimations with published studies of timber plantations across different biogeographic regions. While our biomass equations for individual trees are robust and reliable, estimates of anticipated survival rates and resulting stand densities require careful consideration. Although this study uses conservative survival rate assumptions, it cannot account for uncertainty related to future local random disturbances that may affect stand growth development at specific locations.
Given the dynamic nature of forest growth, ongoing monitoring will be essential to validate the projected biomass estimations against the actual growth patterns. This material is intended to remain open for further refinement and constructive comments with the general aim of continuously refining the assessment of the expected tree and stand biomass accumulation at age 40 since planting.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding authors.
Author contributions
JP-B: Data curation, Investigation, Visualization, Writing – original draft, Writing – review & editing, Methodology. PB: Writing – original draft, Writing – review & editing. SS: Investigation, Writing – original draft, Writing – review & editing. TS-Y: Investigation, Writing – original draft, Writing – review & editing. SA-G: Investigation, Writing – original draft, Writing – review & editing. DN-S: Investigation, Writing – original draft, Writing – review & editing. SK: Funding acquisition, Writing – original draft, Writing – review & editing. TS: Investigation, Project administration, Writing – original draft, Writing – review & editing. EC: Conceptualization, Data curation, Methodology, Project administration, Resources, Supervision, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was co-funded by Life Terra through the LIFE programme (LIFE19 CCM/NL/001200).
Acknowledgments
We thank our former colleague Jan Albert for initial draft and contribution to the concept of this paper.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
References
Albert, M., Nagel, J., Schmidt, M., Nagel, R.-V., and Spellmann, H. (2021). Eine neue Generation von Ertragstafeln für Eiche, Buche, Fichte, Douglasie und Kiefer : [Data set]. Zenodo. doi: 10.5281/zenodo.6343907
Annighöfer, P., Ameztegui, A., Ammer, C., Balandier, P., Bartsch, N., Bolte, A., et al. (2016). Species-specific and generic biomass equations for seedlings and saplings of European tree species. Eur. J. For. Res. 135, 313–329. doi: 10.1007/s10342-016-0937-z
Barbati, A., Corona, P., and Marchetti, M. (2007). European forest types — European Environment Agency. EEA technical report no 9/2006. Available online at: https://www.eea.europa.eu/en/analysis/publications/technical_report_2006_9 (Accessed April 19, 2025).
Barry, L. E., Yao, R. T., Harrison, D. R., Paragahawewa, U. H., and Pannell, D. J. (2014). Enhancing ecosystem services through afforestation: how policy can help. Land Use Policy 39, 135–145. doi: 10.1016/J.LANDUSEPOL.2014.03.012
Bautista, R., Alonso, A., Grau, J. M., and Gómez, J. A. (2001). Tablas de producción de selvicultura media para las masas de Pinus nigra Arn. de la Sierra de Cazorla, Segura y Las Villas. Congreso Forestal Español. Available online at: https://secforestales.org/publicaciones/index.php/congresos_forestales/article/view/15778 (Accessed January 29, 2025).
Berta, A., Levanič, T., Stojsavljević, D., and Kušan, V. (2019). Site index and volume growth percentage determination for privately owned uneven-aged stands of Quercus pubescens and Quercus ilex along the Croatian Adriatic coast. SEEFOR 10, 65–75. doi: 10.15177/SEEFOR.19-08
Bisch, J. L. (1987). “Un exemple de conversion d’une table de production en volume en tables de production en biomasse: le chêne dans le secteur ligérien” in Annales des sciences forestières (HAL open science: EDP Sciences), 243–258. Available online at: https://hal.science/hal-00882414v1
Bollandsås, O. M., Rekstad, I., Næsset, E., and RØsberg, I. (2009). Models for predicting above-ground biomass of Betula pubescens spp. czerepanóvii in mountain areas of southern Norway. Scand. J. For. Res. 24, 318–332. doi: 10.1080/02827580903117412
Bosques españoles y su evolución (n.d.). Available online at: https://www.miteco.gob.es/es/biodiversidad/temas/inventarios-nacionales/inventario-forestal-nacional/index.html (Accessed December 10, 2024).
Bouchon, J., and Trencia, J. (1990). Sylviculture et production du chêne. Rev. Forest. française 42, 246–253. doi: 10.4267/2042/26070
Bravo, F., Guijarro, M., Cámara, A., Díaz Balteiro, L., Fernández, P., Pajares, J., et al. (2017). La situación de los bosques en España-ISFE. Ed. Sociedad Española de Ciencias Forestales. 276.
Bronisz, K., Strub, M., Cieszewski, C., Bijak, S., Bronisz, A., Tomusiak, R., et al. (2016). Empirical equations for estimating aboveground biomass of Betula pendula growing on former farmland in Central Poland. Silva Fennica 50:1559. doi: 10.14214/SF.1559
Bugmann, H., Seidl, R., Hartig, F., Bohn, F., Una, J. B., Cailleret, M., et al. (2019). Tree mortality submodels drive simulated long-term forest dynamics: assessing 15 models from the stand to global scale. Ecosphere 10:e02616. doi: 10.1002/ECS2.2616
Bunce, R. G. H. (1968). Biomass and production of trees in a mixed deciduous woodland: I. Girth and height as parameters for the estimation of tree dry weight. J. Ecol. 56, 759–775. doi: 10.2307/2258105
Bundeswaldinventur Ergebnisdatenbank. (n.d.). Available online at: https://bwi.info/inhalt1.aspx (Accessed December 5, 2024).
Büntgen, U., Urban, O., Krusic, P. J., Rybníček, M., Kolář, T., Kyncl, T., et al. (2021). Recent European drought extremes beyond common era background variability. Nat. Geosci. 14, 190–196. doi: 10.1038/s41561-021-00698-0
Burkhart, H. E., and Avery, T. E. and Bullock, B. P. (2019). Forest measurements. Ed. Waveland Press. 434.
Calama, R., Madrigal, G., del Río, M., López-Senespleda, E., Pardos, M., Ruiz-Peinado, R., et al. (2024). Dynamic height growth equations and site index-based biomass models for young native species Afforestations in Spain. Forests 15:827. doi: 10.3390/F15050827/S1
Calzada, V. R. V., and Millán, J. A. A. (2004). Avances en el estudio de la gestión del monte mediterráneo. Mediterráneo: Fundación Centro de Estudios Ambientales del.
Centre de la Propietat Forestal (2012). Boscos d’alzina i carrasca. Tipologies i models de gestió. Centre de la Propietat Forestal. Available online at: https://cpf.gencat.cat/ca/detalls/Article/03_Boscos-dalzina-i-carrasca (Accessed February 17, 2025).
Černý, M., Pařez, J., and Malík, Z. (1996). Růstové a taxační tabulky hlavních dřevin Česk’ě republiky (smrk, borovice, buk, dub). Jílové u Prahy: IFER.
Cienciala, E., Palmero-Barrachina, J., Albert, J., Blazek, P., and de Santana, T. (2022). Guidance on biomass estimation at age 40 years since planting. Available online at: https://www.datocms-assets.com/50353/1677059743-guidance-on-biomass-estimation-at-age-40-years-since-planting-1.pdf (Accessed May 21, 2025).
Cienciala, E., Altman, J., Doležal, J., Kopáček, J., Štěpánek, P., Ståhl, G., et al. (2018). Increased spruce tree growth in Central Europe since 1960s. Sci. Total Environ. 620, 1637–1647. doi: 10.1016/j.scitotenv.2017.10.138
Cienciala, E., and Apltauer, J. (2008). Undefined (2008). Biomass functions applicable to oak trees grown in central-European forestry. J. For. Sci. 54, 109–120. doi: 10.17221/2906-JFS Z. E.-J. of F
Cienciala, E., Černý, M., Tatarinov, F., Apltauer, J., and Exnerová, Z. (2006). Biomass functions applicable to scots pine. Trees Struct. Funct. 20, 483–495. doi: 10.1007/S00468-006-0064-4
Cienciala, E., Palmero-Barrachina, J., Albert, J., Blažek, P., and Santana, T. (2022). Technical Annex of the Life Terra foundation. Available online at: https://www.datocms-assets.com/50353/1677059743-guidance-on-biomass-estimation-at-age-40-years-since-planting-1.pdf (Accessed December 14, 2024).
Cukor, J., Vacek, Z., Vacek, S., Linda, R., and Podrázský, V. (2022). Biomass productivity, forest stability, carbon balance, and soil transformation of agricultural land afforestation: a case study of suitability of native tree species in the submontane zone in Czechia. Catena 210:105893. doi: 10.1016/J.CATENA.2021.105893
de Embún, X. (1963). Diez temas sobre los arboles. Ministerio de Agricultura, Madrid: Servicio de Extensión Agraria.
de Rigo, D., and Caudullo, G. (2016). “Quercus ilex in Europe: distribution, habitat, usage and threats.,” in European atlas of Forest tree species, eds. J. San-Miguel-Ayanz, D. Rigode, G. Caudullo, T. Houston Durrant, and A. Mauri (European Comission: Publications Office of the European Union Luxembourg), 152–153. doi: 10.2760/776635
Décourt, N. (1965). Les tables de production pour le Pin sylvestre et le le Pin Laricio de Corse en Sologne. Rev. Forest. Française 15, 818–831. doi: 10.4267/2042/24705
Décourt, N. (1971). “Tables de production provisoires pour l’Épicéa commun dans le Nord-Est de la France” in Annales des sciences forestières. (Revue forestière française: EDP Sciences). 23, 411–418. doi: 10.4267/2042/20508
del Río, M., Barbeito, I., Bravo-Oviedo, A., Calama, R., Cañellas, I., Herrero, C., et al. (2017). Mediterranean pine forests: management effects on carbon stocks. Manag. Forest Ecosyst., 301–327. doi: 10.1007/978-3-319-28250-3_15
Dubois, H., Claessens, H., and Ligot, G. (2021). Towards Silviculture guidelines to produce large-sized silver birch (Betula pendula Roth) logs in Western Europe. Forests 12:599. doi: 10.3390/F12050599
Eckmüllner, O. (2004). Empfohlene Ertragstafeln für Nord-und Osttirol. Innsbruck: Amt der Tiroler Landesregierung.
EEA (2016). Biogeographical regions, Europe 2016, version 1. Available online at: https://sdi.eea.europa.eu/catalogue/inspire-pds/api/records/c6d27566-e699-4d58-a132-bbe3fe01491b (Accessed November 20, 2023).
Espelta, M. J., Grácia, M., Molowny-Horas, R., Ordoéz, J. L., Retana, J., Vayreda, J., et al. (2009). “Els alzinars - Manuals de gestió d’hàbitats” in Diputació de Barcelona. eds. M. Grácia and J. L. Ordoñez. Spain, 181.
Fiedler, F. (1986). Die Dendromasse eines hiebsreifen Fichtenbestandes. Beiträge für die Forstwirtschaft 20, 171–180.
Forest Act. Vyhláška o lesním hospodářském plánování (1996). Available online at: https://www.zakonyprolidi.cz/cs/1996-84/zneni-20240101 (Accessed April 21, 2025).
Forrester, D. I., Tachauer, I. H. H., Annighoefer, P., Barbeito, I., Pretzsch, H., Ruiz-Peinado, R., et al. (2017). Generalized biomass and leaf area allometric equations for European tree species incorporating stand structure, tree age and climate. For. Ecol. Manag. 396, 160–175. doi: 10.1016/J.FORECO.2017.04.011
Fortier, J., Truax, B., Gagnon, D., and Lambert, F. (2017). Allometric equations for estimating compartment biomass and stem volume in mature hybrid poplars: general or site-specific? Forests 8:309. doi: 10.3390/F8090309
Francini, S., Schelhaas, M. J., Vangi, E., Lerink, B. J., Nabuurs, G. J., McRoberts, R. E., et al. (2024). Forest species mapping and area proportion estimation combining Sentinel-2 harmonic predictors and national forest inventory data. Int. J. Appl. Earth Obs. Geoinf. 131:103935. doi: 10.1016/J.JAG.2024.103935
García Abejón, J. L. (1981). Tablas de producción de densidad variable para Pinus sylvestris L. en el Sistema Ibérico. INIA, Madrid: Serie Recursos Forestales.
García Abejón, J. L., and Gómez Loranca, J. A. (1984). Tablas de producción de densidad variable para “Pinus sylvestris” L. en el Sistema central. 36. Available online at: https://dialnet.unirioja.es/servlet/libro?codigo=125504 (Accessed January 29, 2025).
García Abejón, J. L., and Gómez Loranca, J. A. (1989). Tablas de producción de densidad variable para Pinus pinaster Ait. en el Sistema Central. INIA, Madrid: Serie Recursos Forestales.
Gasparini, P., Nocetti, M., Tabacchi, G., and Tosi, V. (2006). Biomass equations and data for forest stands and shrublands of the eastern Alps. Italy: USDA Forest Service - General Technical Report PNW.
González Molina, J. M. (2005). Introducción a la selvicultura general. León, Spain: Universidad de León, Secretariado de Publicaciones. 309.
Gonzalez-Molina, J., Meya, D., and Arrufat, D. (1999). Primeras tablas de selvicultura a la carta para masas regulares de “Pinus nigra Arn.” del Prepirineo catalán. Investig. Agraria. Sistemas Recursos Forest. 8, 49–62.
Greenleaf, M. (2024). Growing a better future. Environ. Soc. 15, 160–177. doi: 10.3167/ARES.2024.150108
Guidi, C., Vesterdal, L., Gianelle, D., and Rodeghiero, M. (2014). Changes in soil organic carbon and nitrogen following forest expansion on grassland in the southern Alps. For. Ecol. Manag. 328, 103–116. doi: 10.1016/J.FORECO.2014.05.025
Hamelin, C., Gagnon, D., and Truax, B. (2015). Aboveground biomass of glossy buckthorn is similar in open and understory environments but architectural strategy differs. Forests 6, 1083–1093. doi: 10.3390/F6041083
Hartmann, H., Bastos, A., Das, A. J., Esquivel-Muelbert, A., Hammond, W. M., Martínez-Vilalta, J., et al. (2022). Climate change risks to global Forest health: emergence of unexpected events of elevated tree mortality worldwide. Annu. Rev. Plant Biol. 73, 673–702. doi: 10.1146/ANNUREV-ARPLANT-102820-012804/1
Hernández-Alonso, H., Madrigal-González, J., Tornos-Estupiña, L., Santiago-Rodríguez, A., Alonso-Rojo, P., Morera-Beita, A., et al. (2023). Tree-size heterogeneity modulates the forest age-dependent carbon density in biomass and top soil stocks on Mediterranean woodlands. Plant Soil 486, 361–373. doi: 10.1007/S11104-023-05874-2
Ibàñez, J. J., Lledó, M. J., Sánchez, J. R., and Rodà, F. (1999). “Stand structure, aboveground biomass and production” in Ecology of Mediterranean evergreen oak forests. eds. F. Rodà, J. Retana, C. A. Gracia, and J. Bellot Springer, Berlin, Heidelberg: Ecological Studies. 137. doi: 10.1007/978-3-642-58618-7_3
IFER. (1998). Růstové tabulky dřevin České republiky. Jilove u Prahy, Czech republic. [Internal material, in Czech].
IPCC (2006) in 2006 IPCC guidelines for National Greenhouse gas Inventories, prepared by the National Greenhouse gas Inventories Programme. eds. H. S. Eggleston, L. Buendia, K. Miwa, T. Ngara, and K. Tanabe. Hayama, Japan: Institute for Global Environmental Strategies.
Izquierdo, G. G., de Viñas, I. C. R., García, E. G., and González, G. M. (2007). “Modelos dinámicos dependientes e independientes de la edad: crecimiento en diámetro del" quercus ilex L." en Dehesas” in Cuadernos de la Sociedad Española de Ciencias Forestales. 23, 175–180.
Jagodziński, A. M., Dyderski, M. K., Gȩsikiewicz, K., and Horodecki, P. (2018). Tree- and stand-level biomass estimation in a Larix decidua mill. Chronosequence. Forests 9:587. doi: 10.3390/F9100587
Jansen, H. and Oosterbaan, A. (2018). Opbrengsttabellen. The Netherlands: Wageningen Academic. doi: 10.3920/978-90-8686-876-6
Kahle, H.-P. (2008). Causes and consequences of forest growth trends in Europe. Available online at: http://www.worldcat.org/title/causes-and-consequences-of-forest-growth-trends-in-europe/oclc/191846770 (Accessed June 20, 2017).
Korená Hillayová, M., Holécy, J., Korísteková, K., Bakšová, M., Ostrihoň, M., and Škvarenina, J. (2023). Ongoing climatic change increases the risk of wildfires. Case study: Carpathian spruce forests. J. Environ. Manag. 337:117620. doi: 10.1016/J.JENVMAN.2023.117620
Korosuo, A., Pilli, R., Abad Viñas, R., Blujdea, V. N. B., Colditz, R. R., Fiorese, G., et al. (2023). The role of forests in the EU climate policy: are we on the right track? Carbon Balance Manag. 18:15. doi: 10.1186/s13021-023-00234-0
Krause, A., Pughl, T. A. M., Bayer, A. D., Doelman, J. C., Humpenöder, F., Anthoni, P., et al. (2017). Global consequences of afforestation and bioenergy cultivation on ecosystem service indicators. Biogeosciences 14, 4829–4850. doi: 10.5194/BG-14-4829-2017
Liepiņš, J., Liepiņš, K., and Lazdiņš, A. (2021). Equations for estimating the above- and belowground biomass of grey alder (Alnus incana (L.) Moench.) and common alder (Alnus glutinosa L.) in Latvia. Scand. J. For. Res. 36, 389–400. doi: 10.1080/02827581.2021.1937696
Mäkinen, H., Yue, C., and Kohnle, U. (2017). Site index changes of scots pine, Norway spruce and larch stands in southern and Central Finland. Agric. For. Meteorol. 237-238, 95–104. doi: 10.1016/J.AGRFORMET.2017.01.017
Marklund, L. (1987). Biomass functions for Norway spruce (Picea abies (L.) karst.) in SwedenBiomassafunktioner för gran i Sverige. Available online at: https://search.worldcat.org/title/635932976 (Accessed December 13, 2024).
Marklund, L. G. (1988). Biomass functions for pine, spruce and birch in Sweden Rapport. Sweden: Sveriges Lantbruksuniversitet.
Masera, O. R., Garza-Caligaris, J. F., Kanninen, M., Karjalainen, T., Liski, J., Nabuurs, G. J., et al. (2003). Modeling carbon sequestration in afforestation, agroforestry and forest management projects: the CO2FIX V.2 approach. Ecol. Model. 164, 177–199. doi: 10.1016/S0304-3800(02)00419-2
Mo, L., Zohner, C. M., Reich, P. B., Liang, J., de Miguel, S., Nabuurs, G.-J., et al. (2023). Integrated global assessment of the natural forest carbon potential. Nature 624, 92–101. doi: 10.1038/s41586-023-06723-z
Montero, G. (2008). Selvicultura de Pinus pinea. Compendio de selvicultura aplicada en España. eds. R. Serrada, G. Montero, and J. A. Reque. Madrid: INIA-FUCOVASA. 431–470.
Montero, G., Cañellas, I., and Ruiz-Peinado, R. (2001). Growth and yield models for Pinus halepensis mill. For Syst 10, 179–201. doi: 10.5424/720
Montero, G., López-Leiva, C., Ruiz-Peinado, R., López-Senespleda, E., Onrubia, R., and Pasolodos, M. (2020). Producción de biomasa y fijación de carbono por los matorrales españoles y por el horizonte orgánico superficial de los suelos forestales: Ministerio de Agricultura. Madrid, Spain: Pesca Y Alimentación, Secretaría General Técnica.
Montero, G., Ruiz-peinado, R., and Muñoz, M. (2005). Producción de biomasa y fijación de CO2 por los bosques españoles. Monografías INIA 13,:274. Available online at: https://www.researchgate.net/publication/235639682_Produccion_de_Biomasa_y_Fijacion_de_CO2_Por_Los_Bosques_Espanoles (Accessed September 30, 2022).
Pajtík, J., Konôpka, B., and Šebeň, V. (2022). Allometric models for estimating aboveground biomass of young Norway spruce trees in the Western Carpathians, Slovakia. Cent. Eur. For. J. 68, 154–162. doi: 10.2478/forj-2022-0007
Palacios-Rodríguez, G., Quinto, L., Lara-Gómez, M. A., Pérez-Romero, J., Recio, J. M., Álvarez-Romero, M., et al. (2022). Carbon sequestration in carob (Ceratonia siliqua L.) plantations under the EU afforestation program in southern Spain using low-density aerial laser scanning (ALS) data. Forests 13:285. doi: 10.3390/F13020285/S1
Palmero-Barrachina, J., Blažek, P., and Cienciala, E. (2023). Supporting information from greenhouse gas submissions and sample-based forest inventories for biomass estimation of the Life Terra projects. Available online at: https://www.datocms-assets.com/50353/1676563069-suporting_information_on_ghgandnfi_ifer_f.pdf (Accessed December 14, 2024).
Pausas, J. G., Bladé, C., Valdecantos, A., Seva, J. P., Fuentes, D., Alloza, J. A., et al. (2004). Pines and oaks in the restoration of Mediterranean landscapes of Spain: new perspectives for an old practice - a review. Plant Ecol. 171, 209–220. doi: 10.1023/B:VEGE.0000029381.63336.20
Peichl, M., Leava, N. A., and Kiely, G. (2012). Above- and belowground ecosystem biomass, carbon and nitrogen allocation in recently afforested grassland and adjacent intensively managed grassland. Plant Soil 350, 281–296. doi: 10.1007/S11104-011-0905-9
Pérez-Silos, I., Álvarez-Martínez, J. M., and Barquín, J. (2021). Large-scale afforestation for ecosystem service provisioning: learning from the past to improve the future. Landsc. Ecol. 36, 3329–3343. doi: 10.1007/S10980-021-01306-7
Pilli, R., Anfodillo, T., and Carrer, M. (2006). Towards a functional and simplified allometry for estimating forest biomass. For. Ecol. Manag. 237, 583–593. doi: 10.1016/j.foreco.2006.10.004
Pretzsch, H. (2000). Die Regeln von reineke, yoda und das gesetz der räumlichen allometrie. Allgemeine Forst-und Jagdzeitung 171, 205–210. Available online at: https://www.cabidigitallibrary.org/doi/full/10.5555/20013005668
Pretzsch, H., and Grote, R. (2023). Tree mortality: revisited under changed climatic and Silvicultural conditions. Prog. Bot., 351–393. doi: 10.1007/124_2023_69
Pretzsch, H., and Hilmers, T. (2024). Structural diversity and carbon stock of forest stands: tradeoff as modified by silvicultural thinning. Eur. J. For. Res., 1–22. doi: 10.1007/S10342-024-01691-Z
Pretzsch, H., Steckel, M., Heym, M., Biber, P., Ammer, C., Ehbrecht, M., et al. (2020). Stand growth and structure of mixed-species and monospecific stands of scots pine (Pinus sylvestris L.) and oak (Q. robur L., Quercus petraea (Matt.) Liebl.) analysed along a productivity gradient through Europe. Eur. J. For. Res. 139, 349–367. doi: 10.1007/s10342-019-01233-y
Robbins, Z. J., Xu, C., Aukema, B. H., Buotte, P. C., Chitra-Tarak, R., Fettig, C. J., et al. (2022). Warming increased bark beetle-induced tree mortality by 30% during an extreme drought in California. Glob. Chang. Biol. 28, 509–523. doi: 10.1111/GCB.15927
Rock, J. (2007). Suitability of published biomass equations for aspen in Central Europe—results from a case study. Biomass Bioenergy 31, 299–307. doi: 10.1016/J.BIOMBIOE.2007.01.003
Ruiz-Peinado Gertrudix, R. (2013). Modelos para la estimación del carbono en la biomasa de los sistemas forestales: influencia de la selvicultura en los socks de carbono. Universidad de Valladolid. Instituto Universitario de Investigación en Gestión Forestal Sostenible. doi: 10.35376/10324/4437
Seidl, R., Thom, D., Kautz, M., Martin-Benito, D., Peltoniemi, M., Vacchiano, G., et al. (2017). Forest disturbances under climate change. Nat. Clim. Chang. 7, 395–402. doi: 10.1038/NCLIMATE3303
Shestakova, T. A., Voltas, J., Saurer, M., Berninger, F., Esper, J., Andreu-Hayles, L., et al. (2019). Spatio-temporal patterns of tree growth as related to carbon isotope fractionation in European forests under changing climate. Glob. Ecol. Biogeogr. 28, 1295–1309. doi: 10.1111/geb.12933
Smith, A., Granhus, A., Astrup, R., Bollandsås, O. M., and Petersson, H. (2014). Functions for estimating aboveground biomass of birch in Norway. Scand. J. For. Res. 29, 565–578. doi: 10.1080/02827581.2014.951389
Somogyi, Z., Cienciala, E., Mäkipää, R., Muukkonen, P., Lehtonen, A., and Weiss, P. (2007). Indirect methods of large-scale forest biomass estimation. Eur. J. For. Res. 126, 197–207. doi: 10.1007/s10342-006-0125-7
Suchomel, C., Pyttel, P., Becker, G., and Bauhus, J. (2012). Biomass equations for sessile oak (Quercus petraea (Matt.) Liebl.) and hornbeam (Carpinus betulus L.) in aged coppiced forests in Southwest Germany. Biomass Bioenergy 46, 722–730. doi: 10.1016/J.BIOMBIOE.2012.06.021
Thom, D., and Seidl, R. (2022). Accelerating Mountain Forest dynamics in the Alps. Ecosystems 25, 603–617. doi: 10.1007/S10021-021-00674-0
Thomas, S. C., and Martin, A. R. (2012). Carbon content of tree tissues: A synthesis. For. 3, 332–352. doi: 10.3390/f302033
Valade, A., Luyssaert, S., Vallet, P., Njakou Djomo, S., Jesus Van Der Kellen, I., and Bellassen, V. (2018). Carbon costs and benefits of France’s biomass energy production targets. Carbon Balance Manag. 13, 1–22. doi: 10.1186/S13021-018-0113-5
Van Damme, F., Mertens, H., Heinecke, T., Lefevre, L., De Meulder, T., Portillo-Estrada, M., et al. (2022, 2022). The impact of thinning and clear cut on the ecosystem carbon storage of scots pine stands under maritime influence in Flanders, Belgium. Forests 13:1679. doi: 10.3390/F13101679
Vanclay, J. K. (1994). Modelling forest growth and yield: applications to mixed tropical forests. Wallingford UK: Cab International.
Vilén, T., Cienciala, E., Schelhaas, M. J., Verkerk, P. J., Lindner, M., and Peltola, H. (2016). Increasing carbon sinks in European forests: effects of afforestation and changes in mean growing stock volume. Forestry 89, 82–90. doi: 10.1093/forestry/cpv034
Vonderach, C., Kändler, G., and Dormann, C. F. (2018). Consistent set of additive biomass functions for eight tree species in Germany fit by nonlinear seemingly unrelated regression. Ann. For. Sci. 75, 1–27. doi: 10.1007/S13595-018-0728-4/TABLES/30
Vos, M. A. E., de Boer, D., de Vries, W., den Ouden, J., and Sterck, F. J. (2023a). Aboveground carbon and nutrient distributions are hardly associated with canopy position for trees in temperate forests on poor and acidified sandy soils. For. Ecol. Manag. 529:120731. doi: 10.1016/J.FORECO.2022.120731
Vos, M. A. E., den Ouden, J., Hoosbeek, M., Valtera, M., de Vries, W., and Sterck, F. (2023b). The sustainability of timber and biomass harvest in perspective of forest nutrient uptake and nutrient stocks. For. Ecol. Manag. 530:120791. doi: 10.1016/J.FORECO.2023.120791
Wellock, M. L., LaPerle, C. M., and Kiely, G. (2011a). What is the impact of afforestation on the carbon stocks of Irish mineral soils? For. Ecol. Manag. 262, 1589–1596. doi: 10.1016/j.foreco.2011.07.007
Wellock, M. L., Reidy, B., Laperle, C. M., Bolger, T., and Kiely, G. (2011b). Soil organic carbon stocks of afforested peatlands in Ireland. Forestry 84, 441–451. doi: 10.1093/forestry/cpr046
Werner, R., Gasser, L. T., Steinparzer, M., Mayer, M., Ahmed, I. U., Sandén, H., et al. (2024). Early overyielding in a mixed deciduous forest is driven by both above- and below-ground species-specific acclimatization. Ann. Bot. 134, 1077–1096. doi: 10.1093/AOB/MCAE150
Wirth, C., Schumacher, J., and Schulze, E. D. (2004). Generic biomass functions for Norway spruce in Central Europe—a meta-analysis approach toward prediction and uncertainty estimation. Tree Physiol. 24, 121–139. doi: 10.1093/TREEPHYS/24.2.121
Wong, J., and Dutschke, M. (2003). Can permanence be insured? Technical and practical credits from afforestation and reforestation. Climate Policy. Hamburg Institute of International Economics (HWWA).
Yearbook of Forest Statistics 2021 (2023). Madrid: Ministry for Ecological Transition and the Demographic Challenge. Available online at: https://www.miteco.gob.es/es/ministerio/servicios/publicaciones.html (Accessed April 22, 2025).
Keywords: tree growth, tree survival/mortality, tree biomass, carbon sequestration, National Forest Inventories, growth and Yield tables, planting density
Citation: Palmero-Barrachina J, Blazek P, Sabaté S, Sauras-Yera T, Allasia-Grau S, Nadal-Sala D, Kallen S, de Santana T and Cienciala E (2025) Estimating medium-term (40 years) carbon uptake in living biomass from Life Terra’s afforestation and reforestation actions: challenges and recommendations. Front. For. Glob. Change. 8:1586743. doi: 10.3389/ffgc.2025.1586743
Edited by:
Ana Cristina Gonçalves, University of Evora, PortugalReviewed by:
Paula Soares, University of Lisbon, PortugalJokin Idoate Lacasia, Swiss Federal Institute for Forest, Snow and Landscape Research (WSL), Switzerland
Copyright © 2025 Palmero-Barrachina, Blazek, Sabaté, Sauras-Yera, Allasia-Grau, Nadal-Sala, Kallen, de Santana and Cienciala. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Emil Cienciala, ZW1pbC5jaWVuY2lhbGFAaWZlci5jeg==; Santiago Sabaté, c2FudGkuc2FiYXRlQHViLmVkdQ==