 Mayuri Kashinath Shewale1‡
Mayuri Kashinath Shewale1‡ Jaromír Bláha1‡
Jaromír Bláha1‡ Jiří Synek1‡
Jiří Synek1‡ Martin Schebeck2‡
Martin Schebeck2‡ Martin N. Andersson3†‡
Martin N. Andersson3†‡ Dineshkumar Kandasamy3†‡
Dineshkumar Kandasamy3†‡ Anna Jirošová1*†‡
Anna Jirošová1*†‡- 1Faculty of Forestry and Wood Sciences, Czech University of Life Sciences Prague, Prague, Czechia
- 2Department of Ecosystem Management, Climate and Biodiversity, Institute of Forest Entomology, BOKU University, Vienna, Austria
- 3Department of Biology, Lund University, Lund, Sweden
Introduction: Bark beetles spend most of their lives under the bark of trees, with some species being economically significant pests that cause widespread tree mortality. Their behavior is primarily driven by olfactory signals, with aggregation pheromones playing a prominent role alongside volatiles from hosts, non-host trees, and associated microbes. These signals are detected by olfactory sensory neurons (OSNs) housed in hair-like sensilla on the antennae. In this study, we focused on two Ips species with distinct host preferences: Ips acuminatus, which infests pine species, and Ips cembrae, which primarily attacks European larch. To better understand species-specific adaptations and shared olfactory mechanisms, we compared their olfactory responses with those of Ips typographus, a major pest of Norway spruce. By investigating the frequency, specificity, and antennal distribution of various OSN classes, we aimed to uncover both conserved and different olfactory mechanisms across Ips species with different host associations.
Methods: We conducted single sensillum recordings (SSR) to examine OSN responses in the antennal olfactory sensilla of I. acuminatus and I. cembrae. The responses were compared to existing data from I. typographus to identify potential species-specific adaptations and conserved olfactory mechanisms. A panel of 57 ecologically relevant odorants was tested, comprising interspecific and intraspecific pheromones, along with compounds associated with host- and non-host trees, as well as symbiotic fungi.
Results and discussion: Based on their response profiles, we identified nineteen OSN classes in both I. acuminatus and I. cembrae. A few selected OSN classes were further analyzed using dose–response tests to assess their specificity and sensitivity. Three OSN classes in I. acuminatus and four in I. cembrae were specific to host-related compounds. Two OSN classes responded to non-host volatiles, while one OSN class exhibited strong responses to microbial volatiles in both species. Several OSN classes specific to pheromone compounds, non-host, and microbial volatiles showed similar response profiles in both I. acuminatus and I. cembrae as well as in OSN classes previously reported in I. typographus, potentially reflecting close phylogenetic relationships and shared ecological traits among these species.
1 Introduction
Abiotic disturbances in forests are becoming more frequent and severe due to climate change, leading to increased bark beetle infestations (Jaime et al., 2024). Rising temperatures and prolonged droughts can indirectly influence disturbances such as wildfires or windthrow, creating more favorable conditions for bark beetles (Allen et al., 2015; Jakoby et al., 2019; Senf et al., 2018). Warmer climates accelerate beetle development and reproduction, resulting in outbreaks with significant ecological and economic impacts (Biedermann et al., 2019; Dobor et al., 2020; Sommerfeld et al., 2021; Hlásny et al., 2021). The increasing frequency of droughts and extreme weather events will further impact forests, making them more vulnerable to infestations (Netherer et al., 2024; Wermelinger, 2004). Bark beetles (Coleoptera: Curculionidae: Scolytinae) comprise more than 6,000 species worldwide (Hulcr et al., 2015; Knížek and Beaver, 2007), including some of the most destructive conifer pests, particularly in the northern hemisphere. Bark beetles spend most of their life cycle under the tree bark, where many species feed and develop in the phloem. Many bark beetle species vector symbiotic fungi (particularly ophiostomatoid fungi), which further impair tree defenses and contribute to tree decline and potential mortality (Krokene, 2015).
Bark beetles rely on a diverse array of semiochemicals to coordinate their host selection, mass attack, and regulation of colonization density (Byers, 2007). The colonization process typically begins with the detection of host tree volatiles, which help beetles identify and locate suitable coniferous hosts (Jirošová et al., 2022a; Moliterno et al., 2023). In contrast, non-host volatiles (NHVs) emitted by deciduous trees act as repellents, helping beetles avoid unsuitable hosts (Zhang and Schlyter, 2004). Once a host is selected, pioneer beetles release aggregation pheromones, which attract conspecifics and facilitate coordinated mass attacks, a crucial strategy for overcoming tree defenses (Christiansen and Bakke, 1988; Wood, 1982; Raffa et al., 2016; Keeling et al., 2021). As colonization progresses, bark beetles also respond to volatiles produced by their symbiotic ophiostomatoid fungi, which can influence both aggregation behavior and host suitability assessment (Jirošová et al., 2022b; Kandasamy et al., 2019, 2023). To avoid overcrowding and resource depletion, beetles release anti-aggregation pheromones at later stages, which regulate colonization density and promote dispersal to uncolonized trees (Frühbrodt et al., 2024). Beyond intraspecific signaling, bark beetles are also capable of detecting volatiles emitted by other bark beetle species and associated fungi, suggesting a broader role for interspecific chemical communication in mediating competition and spatial distribution (Andersson et al., 2009; Schiebe et al., 2019; Yuvaraj et al., 2024; Zhao et al., 2019; Kandasamy et al., 2019, 2023).
The pine bark beetle, Ips acuminatus, and the larch bark beetle, Ips cembrae, are both ecologically significant species of coniferous forests in Europe (Papek et al., 2024; Postner, 1974). Ips acuminatus mainly infests stressed Scots pine (Pinus sylvestris), with outbreaks increasing due to drought and warming (Liška et al., 2021; Wermelinger et al., 2008; Thabeet et al., 2009). It prefers the thin-barked upper trunk and crown, avoiding competition by I. sexdentatus (Pettersson, 2000; Pfeffer, 1955; Wood and Bright, 1992), and other bark beetle species such as Tomicus piniperda and Tomicus minor (Foit and Čermák, 2014; Hlávková and Doležal, 2022). Males release a pheromone blend of S-(−)-ipsenol, S-(+)-ipsdienol, and (4S)-cis-verbenol to attract conspecifics (Bakke, 1978; Francke et al., 1986), with mating occurring in a polygynous system (Kirkendall, 1989, 1990). It is associated with ophiostomatoid fungi, including blue-stain species that may support beetle development and survival (Francke-Grosmann, 1965; Villari et al., 2012; Papek et al., 2024).
Similarly, I. cembrae primarily infests European (Larix decidua) and Japanese larch (Larix kaempferi) but can also colonize other conifer species (Postner, 1974). While typically a secondary pest of weakened or felled trees, warming and drought can trigger outbreaks (Grodzki, 2008; EFSA on Plant Health et al., 2017). It colonizes the entire trunk (Pfeffer, 1955) and competes in the crown with bark beetles from several genera, such as Pityophthorus, Pityogenes, and Cryphalus (Postner, 1974). Males emit S-(−)-ipsenol, S-(+)-ipsdienol, and 3-methyl-3-buten-1-ol to initiate aggregation (Kohnle et al., 1988; Stoakley et al., 1978), followed by mating with 2–4 females in a chamber (Postner, 1974). Ips cembrae also vectors Endoconidiophora laricola, a pathogenic blue-stain fungus that contributes to tree mortality (Redfern et al., 1987; Kirisits, 2004; Jankowiak et al., 2007).
The primary olfactory organ of bark beetles are the club-shaped antennae (Payne et al., 1973), covered with multi-porous sensilla that house olfactory sensory neurons (OSNs) (Hallberg, 1982a). In Ips species, the flattened antennal club has the most olfactory sensilla concentrated on the anterior surface, organized into three distinct sensory bands labeled A, B, and C (Hallberg, 1982a; Shewale et al., 2023). Most bark beetle OSNs are narrowly tuned, responding strongly to a single or a few structurally similar compounds, while some exhibit broader tuning (Andersson et al., 2009; Kandasamy et al., 2019, 2023). The dendritic membrane of the OSNs contains chemoreceptor proteins, such as odorant receptors (ORs) (Clyne et al., 1999) and ionotropic receptors (IRs) (Benton et al., 2009), which translate odor information of the environment into electrical signals. These signals can be interpreted by the brain, potentially leading to behavioral responses (Andersson et al., 2015).
Early single sensillum recording (SSR) experiments in bark beetles investigated OSN responses to pheromones and some host volatiles in I. typographus (Tømmerås, 1985). Studies on olfactory detection of mainly pheromone compounds are also available for other Ips species, such as Ips pini, Ips paraconfusus, and Ips grandicollis (Ascoli-Christensen et al., 1993; Mustaparta et al., 1979; Mustaparta et al., 1980, 1977). Ips typographus, a major pest of Norway spruce (Picea abies), is the most well-studied Ips species in terms of peripheral odor detection, with extensive research reporting the antennal abundance of different OSN classes and the spatial distribution of OSNs tuned to pheromones, host volatiles, NHVs, and microbial volatiles (Andersson et al., 2009; Kandasamy et al., 2019, 2023; Raffa et al., 2016; Schiebe et al., 2019; Yuvaraj et al., 2024). The comprehensive OSN data from I. typographus allows for detailed comparison with OSN data from other congeneric species with different host preferences to better understand their olfactory detection mechanisms. Although semiochemicals are often classified as pheromones, allomones, or kairomones, these classes might be unclear in this study, given their overlapping ecological activities. Therefore, we classify compounds in this study based on their biosynthetic origin, i.e., beetle-produced, host-derived, or microbial, to maintain clarity and avoid confusion. The chemical communication mechanisms that underlie their behavior, including pheromone-mediated aggregation and detection of plant and microbial volatiles, are critical for understanding their success as pests. Addressing these knowledge gaps is especially important given the increasing risks posed by these species under changing environmental conditions.
This study aimed to functionally characterize the OSN classes in two Ips species with different host preferences, specifically I. acuminatus on pine and I. cembrae on larch. By comparing our findings with existing data for the spruce bark beetle, I. typographus, we investigated whether the OSN frequencies and response patterns vary between species, and which of these patterns are conserved across Ips species. Using SSR, we examined OSN responses to 57 ecologically relevant odorants, including pheromones and volatiles from host trees, non-host trees, and fungi. Gas chromatography-electroantennographic detection (GC-EAD) using essential oils was also performed to investigate whether the antennae of the studied species respond to host volatiles. This study advances our understanding of olfactory adaptations in Ips species, particularly in pheromone communication and host detection. Additionally, our findings provide new insights that could be useful for species-specific monitoring and pest management, essential for maintaining forest health under climate change.
2 Materials and methods
2.1 Bark beetle collection
Both bark beetle species were collected from forests near the village of Rouchovany in central Czech Republic (49.0704°N, 16.1076°E) during late spring 2024. Species identification was conducted directly in the field. Branches of P. sylvestris (DBH 2–10 cm) infested by I. acuminatus were collected, along with logs of L. decidua (DBH 20–50 cm) infested by I. cembrae. Infested logs were maintained in university rearing facilities (FFWS, CULS) within insect cages (60 × 60 × 110 cm) under controlled laboratory conditions (25°C during the day, 19°C at night, 60% RH, and a 16:8 light/dark photoperiod). Adult beetles began emerging three to four weeks after field collection. The emerged beetles were collected and sexed under a stereomicroscope based on external morphology, specifically by the shape of elytral spines (Pfeffer, 1955; Zhang and Niemeyer, 1992). Before use in experiments, adult beetles were individually stored in Falcon tubes lined with moist paper at 4°C for at least 1 week. Each adult beetle was used for ten screenings using single sensillum recordings, whereas each beetle was used only once for dose–response studies. To obtain enough beetles, another batch of I. acuminatus was collected in spring 2024 from naturally infested P. sylvestris in northeastern Austria (Schönberg am Kamp; 48.5185°N, 15.7322°E) due to unavailability at the original location. Colonized logs (60 cm in length) were transferred to incubators at BOKU University, Vienna, where they were maintained at 25° C with a 16:8 light/dark photoperiod and monitored daily for newly emerged beetles. The emerged beetles were sexed and then express-mailed to Lund University, Sweden, for subsequent SSR experiments.
2.2 Chemical stimuli
The odor panel included 57 ecologically relevant compounds, including beetle pheromones, host-, non-host-, and microbial-related volatiles (Supplementary Table 2). These compounds were selected based on previous studies on Ips species, including I. typographus (Andersson et al., 2009; Kandasamy et al., 2023). Stock odor solutions (10 μg/μL) were prepared in paraffin oil and further diluted for use in experiments. For stimulation, 10 μL of the solution was applied to a piece of filter paper placed inside glass Pasteur pipettes. Control stimuli consisted of paraffin oil alone. Pipettes were stored at −18°C between experiments and replaced frequently to minimize odor depletion (Andersson et al., 2012b). For GC-EAD experiments, essential oils of L. decidua and P. sylvestris were purchased from Oshadhi Ltd. (United Kingdom). Stock odor solutions (10 μg/μL) were prepared in hexane and further diluted for use. For GC-EAD experiments, 1 μL of the solution was directly injected into GC.
2.3 Single-sensillum recordings (SSR)
SSR was performed on live adult individuals of I. acuminatus and I. cembrae to investigate OSN response profiles using previously described procedures (Andersson et al., 2012a). Beetles were immobilized in a 200 μL pipette tip, leaving the antennae and head exposed. One antenna was carefully secured with dental wax onto a microscope slide, ensuring optimal positioning for electrode insertion and light penetration from below. Mounted antennae were observed using a light microscope (Nikon Eclipse E6000FN) at ×500 magnification. Electrophysiological recordings were conducted using tungsten microelectrodes that were electrolytically sharpened with 1M KNO₃. The reference electrode was inserted into a pre-made hole in the beetle’s pronotum, while the recording electrode was inserted at the base of an olfactory sensillum. The recording electrode was mounted on a Sensapex micromanipulator (uMp-3, Oulu, Finland) for precise positioning. Signals were amplified and digitized using an IDAC4 interface (Syntech), and real-time recordings were visualized in AutoSpike v. 3.9 (Syntech). A continuous stream of charcoal-filtered and humidified air (1.2 L/min) was directed onto the antenna via a 6 mm inner diameter glass tube positioned 15 mm from the antenna. Odor stimuli were delivered as 0.5 s puffs (0.3 L/min) using a stimulus controller (CS-02, Syntech), allowing the odorant to mix into the continuous airflow and reach the antenna. Odor pipettes for screening experiments were used for a maximum of two consecutive days or ten stimulations per compound. Dose–response pipettes were freshly prepared daily and used for a maximum of two stimulations. To characterize OSN response profiles, a high-dose stimulus (10 μg; 10 μL of a 1 μg/μL solution) was used for initial screening. Odor compounds were tested in random order, and OSNs were allowed to regain basal spontaneous activity between stimulations. OSNs were classified based on their response profiles during the screening experiments. Additionally, five OSN classes for I. acuminatus and three OSN classes for I. cembrae were selected for dose–response tests. Compounds were tested in increasing concentrations (10 pg to 10 μg) to minimize sensory adaptation, starting with the least active compound identified during the screening phase.
2.4 Data analysis
Neuronal responses were quantified offline in AutoSpike v3.9 by calculating spike frequencies during the first 0.5 s of the odor response and subtracting the pre-stimulation activity. Responses to the paraffin oil control were also subtracted. At the screening dose, responses below 20 Hz were considered biologically non-significant. Excitatory responses were categorized as intermediate responses (40–60 Hz) and strong responses (>80 Hz). Poor-quality recordings or incompletely screened neurons were excluded. All data graphs and heatmaps were generated using GraphPad Prism (version 10.1.2, GraphPad Software, San Diego, CA, USA). The venn diagram was created using InteractiVenn (Heberle et al., 2015).
2.5 Gas chromatography coupled electroantennographic (GC-EAD) experiments
For GC-EAD, the head of the beetle with antennae was prepared and connected between glass microelectrodes filled with Ringer’s solution (Olsson and Hansson, 2013). Signals from the antenna were recorded using a Universal probe (Syntech) and integrated with IDAC 2 (Syntech). The results were processed using the software GcEad v. 4.6.1 (Syntech). At least five recordings were made for each sample, and a volatile was considered active if at least two antennal responses were recorded in I. acuminatus and I. cembrae. For the experiments, an Agilent 7890B GC was used, equipped with an HP-5 column (Agilent Technologies, Inc), 30 m in length, 0.32 mm in diameter, and with a film thickness of 0.25 μm, ending in a splitter. From the splitter, 5 m of the same column was led to the FID detector and 1 m of the column on the antenna. At the column outlet, the chemical components were mixed with humidified air at a flow rate of 2 L/min and blown onto the prepared antenna. Samples were injected splitless, and the carrier gas for the GC was helium with a constant column flow rate of 3 mL/min. The inlet temperature was set to 250°C, the initial oven temperature was set to 40°C for 1 min, then increased by 10°C/min to 100°C, held for 0.5 min, then increased by 20°C/min to 150°C, and then increased by 40°C/min to a final temperature of 300°C, with 3 min hold. The FID temperature was set to 300°C.
3 Results
3.1 General classification of OSN types
The responses of OSNs in I. acuminatus (IAc) and I. cembrae (IC) were examined using single sensillum recordings (SSR) from antennal olfactory sensilla. Most sensilla housed two OSNs, distinguishable by their spike amplitudes (A neuron: large amplitude cell; B neuron: small amplitude cell). Some sensilla appeared to house a single OSN, while a few seemed to house three. However, the presence of three neurons was rare and sometimes difficult to confirm due to suboptimal signal quality. The OSNs frequently responded to multiple compounds, but the primary compounds elicited the strongest responses, frequently exceeding 80 Hz. The compounds that triggered weaker secondary responses were often structurally similar to the primary compounds. The OSN response activity generally followed a phasic-tonic pattern with a sharp initial rise in firing rate, followed by a gradual decline to baseline levels (Supplementary Figure 1). However, most responses were rather tonic, with increased firing well beyond stimulus offset, whereas some neurons responded in a phasic manner, with responses quickly returning to baseline activity. The highest response frequencies reached 150 Hz in I. acuminatus and 200 Hz in I. cembrae. Notably, the compounds that elicited the strongest responses at the high screening dose (10 μg on the filter paper) were also associated with the lowest detection thresholds.
A screening experiment using 57 ecologically relevant odorants at 10 μg revealed that 69 out of 82 contacted sensilla (~84%) in I. acuminatus (males, n = 17; females, n = 23) and 62 out of 85 sensilla (~73%) in I. cembrae (males, n = 28 and females, n = 18), responded to at least one compound. The remaining sensilla (12 in I. acuminatus and 23 in I. cembrae) did not respond to any of the tested compounds. Additionally, a small number of sensilla (three in I. acuminatus and two in I. cembrae) were excluded due to poor recording quality or signal loss during the experiment, which prevented OSN classification. OSNs responding strongly (>80 Hz) to at least one compound were categorized into OSN classes based on their response profiles (Figures 1, 2). In contrast, OSNs with weak to intermediate responses (20–80 Hz) were not assigned to any OSN class because their primary compounds were likely missing from the test odor panel (Supplementary Table 1 shows detailed OSN responses).
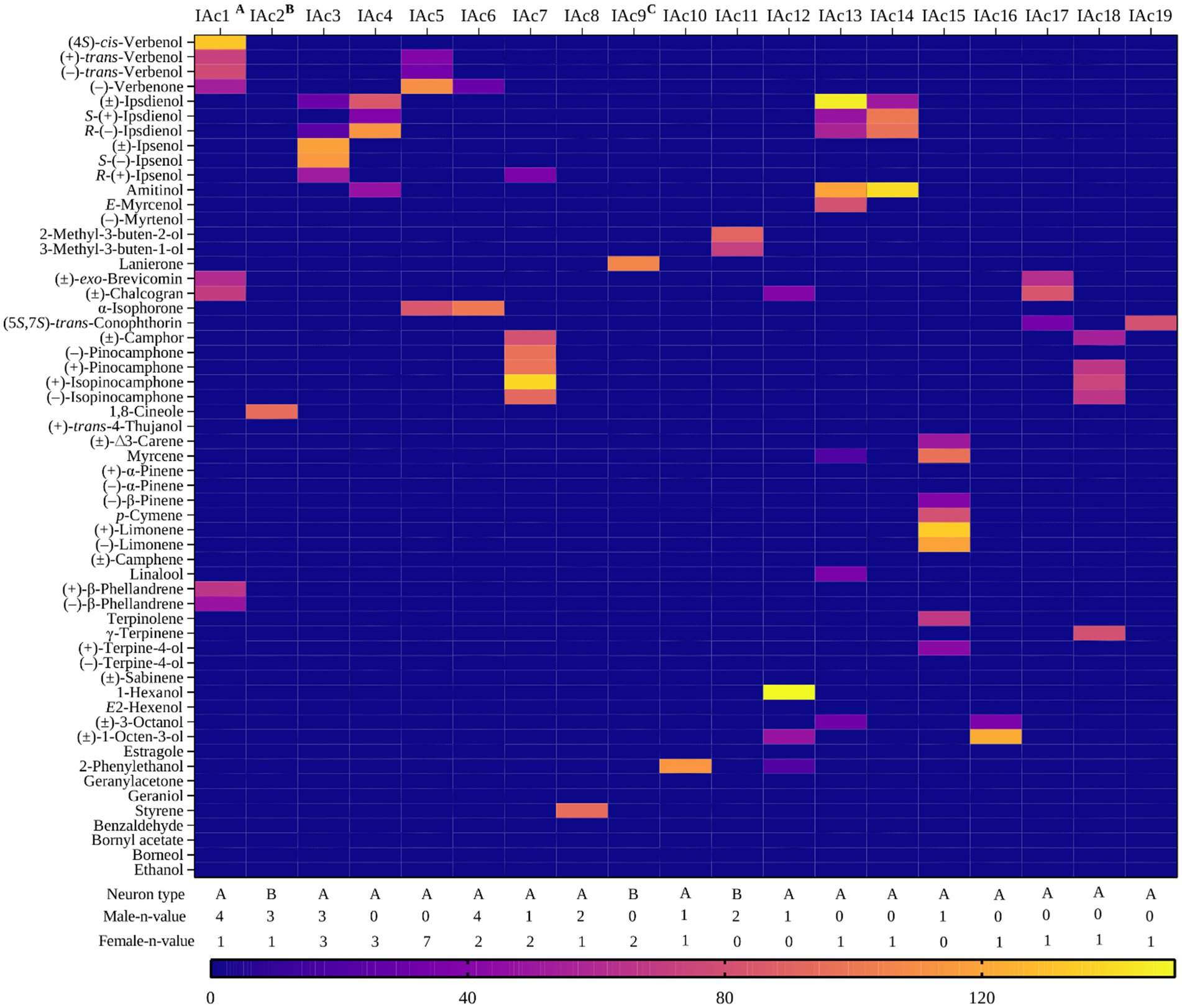
Figure 1. Heat map representing response profiles of identified olfactory sensory neurons classes responding strongly (>80 Hz) to one or more compounds at a 10 μg stimulus dose in Ips acuminatus. The stimulus eliciting the primary response in the 19 OSN classes are as follows: IAc1: (4S)-cis-verbenol, IAc2: 1,8-cineole, IAc3: (±)-ipsenol, IAc4: R-(−)-ipsdienol, IAc5: (−)-verbenone, IAc6: α-isophorone, IAc7: (+)-isopinocamphone, IAc8: styrene, IAc9: lanierone, IAc10: 2-phenylethanol, IAc11: 2-methyl-3-buten-2-ol, IAc12: 1-hexanol, IAc13: (±)-ipsdienol, IAc14: amitinol, IAc15: (−)-limonene, IAc16: 1-octen-3-ol, IAc17: (±)-chalcogran, IAc18: γ-terpinene, and IAc19: (5S,7S)-trans-conophthorin. A and B illustrate the co-localization of IAc1 with IAc2, while C shows the co-localization of IAc9 with IAc4 and IAc5.
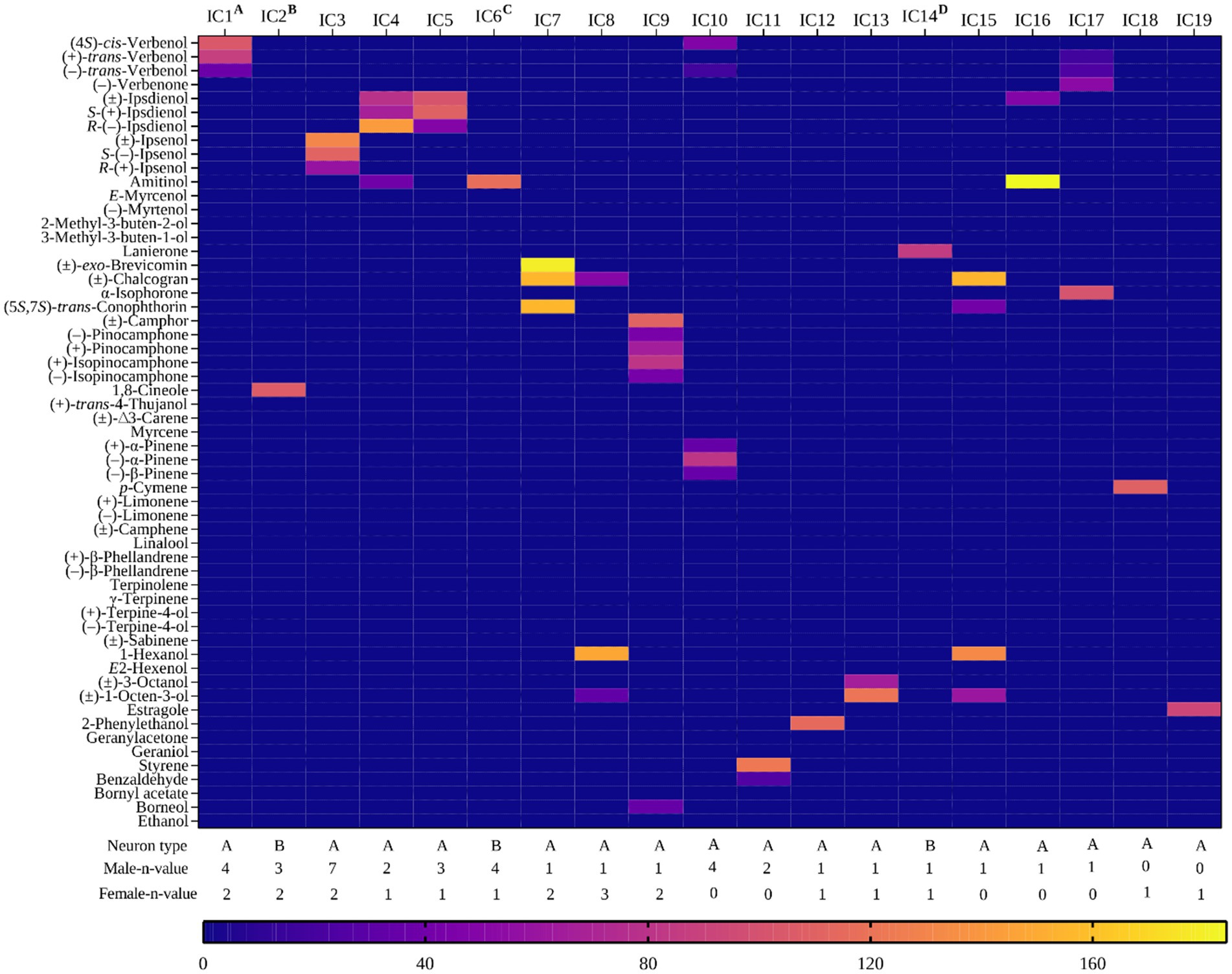
Figure 2. Heat map showing response profiles of identified OSN classes responding strongly (>80 Hz) to one or more compounds at the 10 μg stimulus dose in Ips cembrae. The stimulus eliciting the primary response in the 19 OSN classes are as follows: IC1: (4S)-cis-verbenol, IC2: 1,8-cineole, IC3: (±)-ipsenol, IC4: R-(−)-ipsdienol, IC5:S-(+)-ipsdienol, IC6: amitinol (B neuron), IC7: (±)-exo-brevicomin, IC8: 1-hexanol, IC9: (±)-camphor, IC10: (−)-α-pinene, IC11: styrene, IC12: 2-phenylethanol, IC13: 1-octen-3-ol, IC14: lanierone, IC15: (±)-chalcogran, IC16: amitinol (A neuron), IC17: α-isophorone, IC18: p-cymene, and IC19: estragole. A indicates IC1 co-localized with IC2, B illustrates IC2 co-localized with IC1 and IC10, C represents IC5 co-localized with IC4 and IC10, and D highlights IC14 co-localized with IC8.
3.2 Olfactory sensory neuron responses in I. acuminatus
3.2.1 OSNs responding to aggregation pheromone components in I. acuminatus
Three OSN classes in I. acuminatus were strongly activated by its aggregation pheromone components, with distinct ligand specificities and dose-dependent responses. The IAc1 class (I. acuminatus OSN class 1) responded strongly (>80 Hz) to (4S)-cis-verbenol (n = 5), which is the major aggregation component in this species. Weaker secondary responses (<60 Hz) were elicited by structurally similar compounds, including (+)-trans-verbenol, (−)-trans-verbenol, (−)-verbenone, and chalcogran. These A neuron OSNs were co-localized with a B neuron OSN class (IAc2), which strongly responded (>80 Hz) to 1,8-cineole, a host tree defense compound (Figure 3A). Sensilla housing IAc1 neurons were predominantly located on the distal antennal club in the sensory band C (Figure 4A).
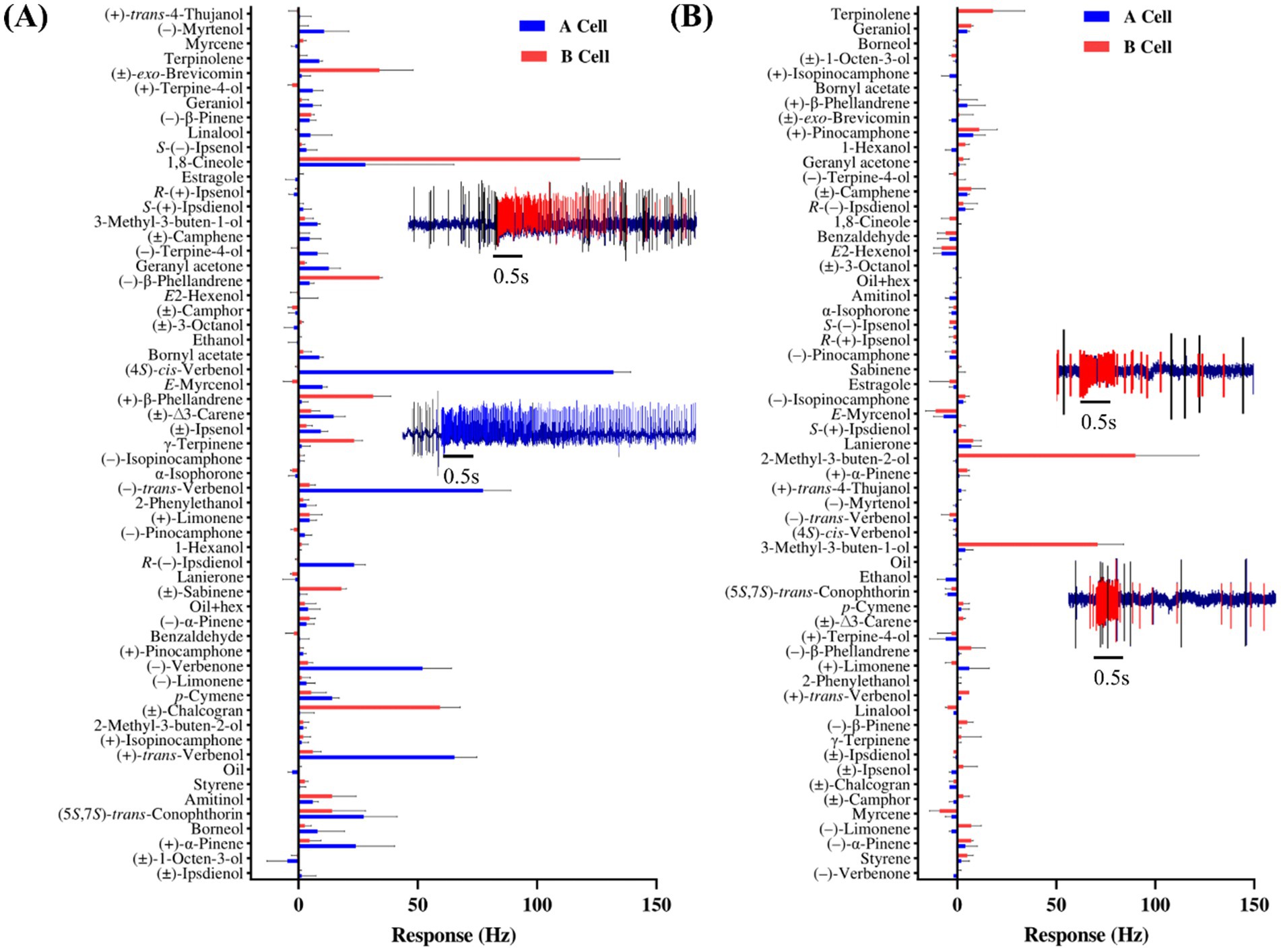
Figure 3. Single sensillum recordings from Ips acuminatus reveal co-localization of OSN classes with different spike amplitudes and response profiles. (A) The response pattern of the large-spiking A neuron with primary response to (4S)-cis-verbenol, co-localized with a small-spiking B neuron responding primarily to 1,8-cineole (n = 3 sensilla). Response frequencies (Hz) to the 10 μg compound dose are shown as mean ± the standard error of the mean (SEM). Representative action potential traces display the A neuron responses to (4S)-cis-verbenol and the B neuron responses to 1,8-cineole. (B) Responses of B neurons to both 2-methyl-3-buten-2-ol and 3-methyl-3-buten-1-ol (10 μg), co-localized with an unresponsive A neuron in the same sensillum (n = 3 sensilla). Representative action potential traces show the B neuron responses to these compounds.
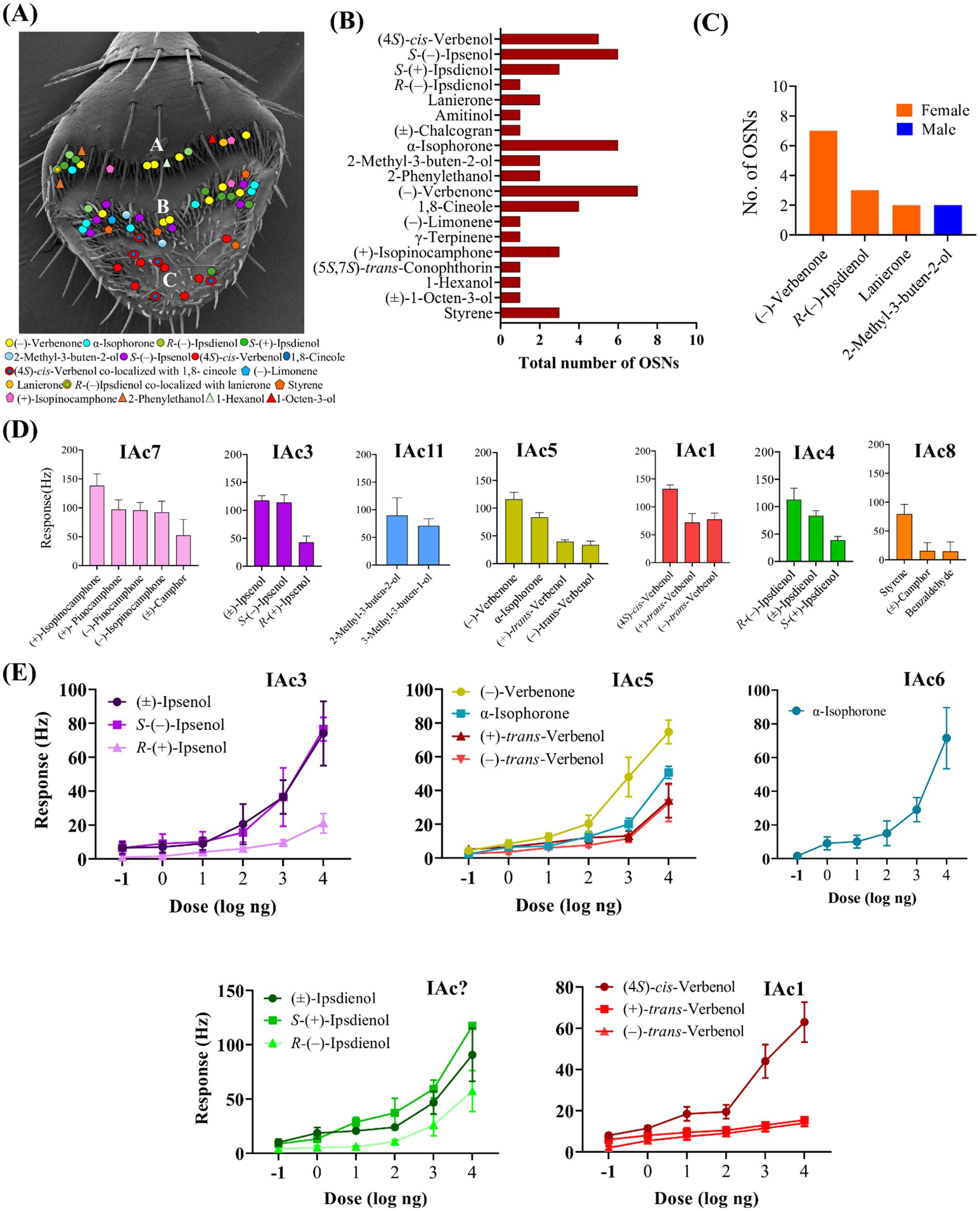
Figure 4. (A) Distribution of identified olfactory sensory neuron classes in Ips acuminatus on the antennal surface with three sensory bands A, B, and C. (B) Total number of each of the 19 OSN classes identified in I. acuminatus, with primary responses to compounds from different ecological origins. (C) Total number of OSN that were only found in one of the sexes of I. acuminatus. (D) Mean response (Hz) of the different OSN classes, including their secondary responses; from left to right: OSN classes IAc7: (+)-isopinocamphone (n = 3), IAc3: S-(−)-ipsenol (n = 6), IAc11: 2-methyl-3-buten-2-ol (n = 3), IAc5: (−)- verbenone (n = 6), IAc1: (4S)-cis-verbenol (n = 3), IAc4: R-(−)-ipsdienol (n = 3), and IAc8: styrene (n = 3). Error bars indicate standard error of the mean (SEM). (E) Dose–response curves of five OSN classes in I. acuminatus; IAc3 class: S-(−)-ipsenol (n = 4); IAc5 class: (−)-verbenone (n = 6); IAc? class: S-(+)-ipsdienol (n = 3)[the IAc? OSN class was observed only in dose–response tests and not while screening]; IAc1 class: (4S)-cis-verbenol (n = 4), and IAc6 class: α-isophorone (n = 4). Mean response values are shown, with error bars indicating the standard error of the mean (SEM).
The IAc3 class responded primarily (>80 Hz) to S-(−)-ipsenol (n = 6), which is the naturally occurring enantiomer of ipsenol. These A neurons showed weak secondary responses (<60 Hz) to R-(+)-ipsenol and ipsdienol enantiomers. Dose–response tests confirmed the high specificity for S-(−)-ipsenol, with a response threshold as low as 100 pg (Figure 4E). The OSNs of this class were mainly distributed in the sensory band B (Figure 4A). The IAc4 class also exhibited enantiomer-specific detection, responding strongly (>80 Hz) to R-(−)-ipsdienol (n = 3). These A neurons were co-localized with lanierone-responsive B neurons (IAc9 class, described below). Weaker secondary responses were observed for the S- (+)-enantiomer, the racemic mixture, and amitinol. This class was distributed on the sensory bands A and B on the antennal surface (Figure 4A) and was observed only in females. In contrast to the screening experiments, dose–response studies demonstrated the strongest response to the other enantiomer, to S-(+)-ipsdienol, with minimal responses to secondary compounds at lower concentrations (Figure 4E). Interestingly, during initial screening experiments, we did not identify any sensilla that strongly responded to S-(+)-ipsdienol. This discrepancy between the screening and dose–response data suggests that two distinct OSN classes likely exist in I. acuminatus, each specifically tuned to either S-(+)-ipsdienol or R-(−)-ipsdienol.
Additionally, OSN class IAc13, (n = 1, A neuron) responded strongly (>80 Hz) to racemic ipsdienol, with intermediate secondary responses to amitinol, E-myrcenol and ipsdienol enantiomers. Another OSN class, IAc14 (n = 1, A neuron), responded most strongly to amitinol with weaker secondary responses to racemic ipsdienol and its enantiomers (Figure 1; Supplementary Table 1).
3.2.2 OSNs responding to other beetle-produced compounds in I. acuminatus
Five OSN classes were tuned to additional beetle-produced compounds. The IAc5 class responded strongly (>80 Hz) to (−)-verbenone (n = 7) and exhibited weaker secondary responses to α-isophorone, (+)-trans-verbenol, and (−)-trans-verbenol. These A neurons, primarily distributed across the sensory bands A and B, mostly in the middle region of the antennal club, displayed dose-dependent responses with a response threshold at ~1 ng (Figure 4E). This was the most abundant OSN class in this species (Figure 4B) and was found exclusively in females (n = 7). The IAc6 class showed high specificity (>80 Hz) to α-isophorone (n = 6). These A neurons did not respond to verbenone or any other compounds from the odor panel, and dose–response tests revealed an exceptionally low response threshold at 10 pg, indicating high sensitivity (Figure 4E). This OSN class was distributed mainly in the sensory band B on the antennal club (Figure 4A). Additional pheromone-responsive OSN classes included IAc9, a B neuron class strongly responding (>80 Hz) to lanierone (n = 2, both females), with one B neuron co-localized with an R-(−)-ipsdienol-responsive A neuron and the other B neuron with a non-responsive A neuron. The IAc10 class, also an A neuron, responded strongly (>80 Hz) to 2-phenylethanol (n = 2). Another OSN class, IAc11, a B neuron, strongly responded to 2-methyl-3-buten-2-ol (n = 2, both males) with secondary responses to 3-methyl-3-buten-1-ol (Figure 4C). These OSNs were co-localized with non-responsive A neurons (Figure 4D). Most of these pheromone-sensitive OSN classes were distributed in all three sensory bands on the antennal surface (Figure 4A). Another A neuron class, IAc17, showed a strong response (>80 Hz) to chalcogran followed by intermediate secondary responses (>50 Hz) to (±)-exo-brevicomin and weaker responses (<40 Hz) to (5S,7S)-trans-conophthorin (Figure 4D).
3.2.3 OSN classes responding to host, non-host, and microbial volatiles in I. acuminatus
We observed three OSN classes with strong primary responses (>80 Hz) to host volatiles. OSN class IAc2 was a B neuron, which showed strong responses to only 1,8-cineole. OSN class IAc15 was an A neuron, which responded strongly (>80 Hz) to (−)-limonene and (+)-limonene followed by intermediate (>50 Hz) secondary responses to myrcene, p-cymene, terpinolene, and ∆-3-carene, and weak responses (<40 Hz) to (+)-terpine-4-ol and (−)-β-pinene (Figure 1). OSN class (IAc18) was an A neuron, which primarily responded strongly (>80 Hz) to γ-terpinene and secondarily to (+)-isopinocamphone, (−)-isopinocamphone, (+)-pinocamphone, and racemic camphor (Figure 4D). Most of the OSN classes responding to host volatiles were distributed on the sensory band B (Figure 4A). Notably, pheromone-sensitive and host-specific OSN classes were generally not spatially segregated across the antennal surface, with the exception of (4S)-cis-verbenol-responsive neurons, which were exclusively localized within sensory band C (Figure 4A).
Three OSN classes, all A neurons, exhibited strong responses (>80 Hz) to non-host volatiles. OSN class IAc12 responded strongly to 1-hexanol. Secondary responses (<50 Hz) were observed for other compounds, such as racemic 1-octen-3-ol and chalcogran. OSN class IAc16 displayed strong responses to racemic 1-octen-3-ol and weak responses to racemic 3-octanol (Figure 1). Another OSN class, IAc19, responded strongly (>80 Hz) to the bicyclic ketal (5S,7S)-trans-conophthorin (n = 1). Two OSN classes exhibited strong responses (>80 Hz) to microbial volatiles (Figure 4B). The IAc7 class was an A neuron, which responded strongly to (+)-isopinocamphone (n = 3). Secondary responses (60–80 Hz) were observed to related oxygenated monoterpenes from trees and microbes, including (−)-isopinocamphone, (+)-pinocamphone, (−)-pinocamphone, and racemic camphor. These responses suggest broad tuning to structurally similar oxygenated host monoterpenes (Figure 4D). The IAc8 class was an A neuron specific to styrene, with secondary responses to benzaldehyde and racemic camphor (n = 3).
3.3 Olfactory sensory neuron responses in I. cembrae
3.3.1 OSN classes responding to aggregation pheromone components of I. cembrae
We identified two OSN classes specific to the aggregation pheromone components of I. cembrae. OSN class IC1 was an A neuron (n = 6), which responded primarily (>80 Hz) to (4S)-cis-verbenol with dose-dependent responses and a response threshold of 100 pg. Weaker secondary responses (40–60 Hz) to (+)-trans-verbenol and (−)-trans-verbenol were observed (Figure 5E). Five out of six of these (4S)-cis-verbenol-responsive OSNs were co-localized with IC2, which was a B neuron responding to 1,8-cineole (Figure 6A). The distribution was mostly in the distal antennal club region on sensory band C, while few were located on sensory band B (Figure 5A). (4S)-cis-Verbenol is a male-produced aggregation pheromone component in both I. typographus and I. acuminatus but not reported as an aggregation pheromone component in I. cembrae.
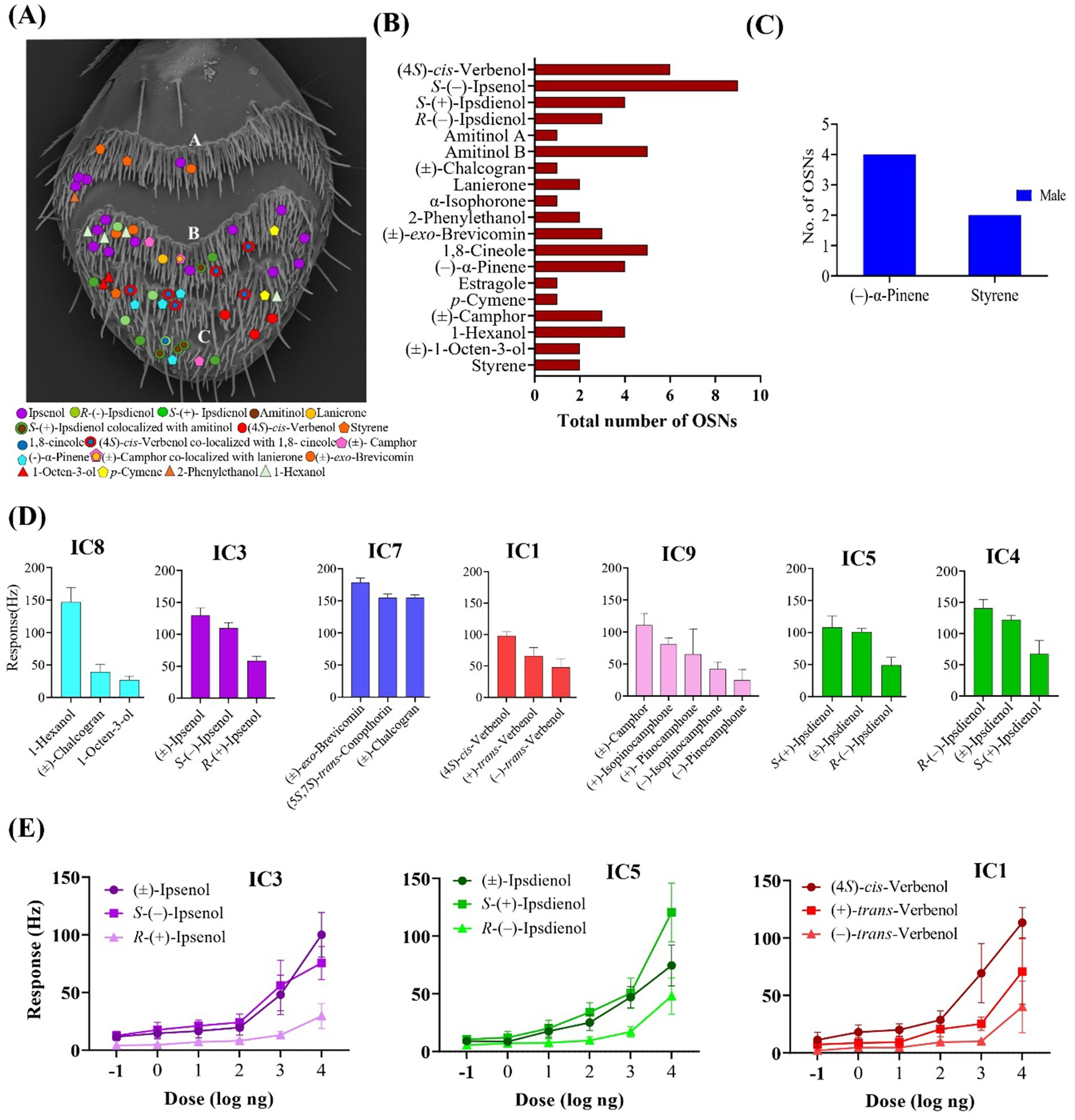
Figure 5. (A) Distribution of identified olfactory sensory neuron classes in the three sensory bands (A, B, C) on the antennal surface of Ips cembrae. (B) Total number of each of the 19 OSN classes identified in I. cembrae, with primary responses to compounds from different ecological origins (C) Total number of OSNs that were only found in males of I. cembrae. (D) Mean responses (Hz) of different OSN classes, including their secondary responses; from left to right: OSN classes IC8: 1-hexanol (n = 4), IC3: S-(−)-ipsenol (n = 9), IAc7: (±)-exo-brevicomin (n = 3), IC1: (4S)-cis-verbenol (n = 4), IC5: S-(+)-ipsdienol (n = 4), and IC4: R-(−)-ipsdienol (n = 3). Error bars indicate the standard error of the mean (SEM). (E) Dose–response curves of three pheromone-specific OSN classes in I. cembrae; IC3 class: S-(−)-ipsenol (n = 4); IC5 class: S-(+)-ipsdienol (n = 4), and IC1 class: (4S)-cis-verbenol (n = 3). Mean response values are shown, with error bars indicating the standard error of the mean (SEM).
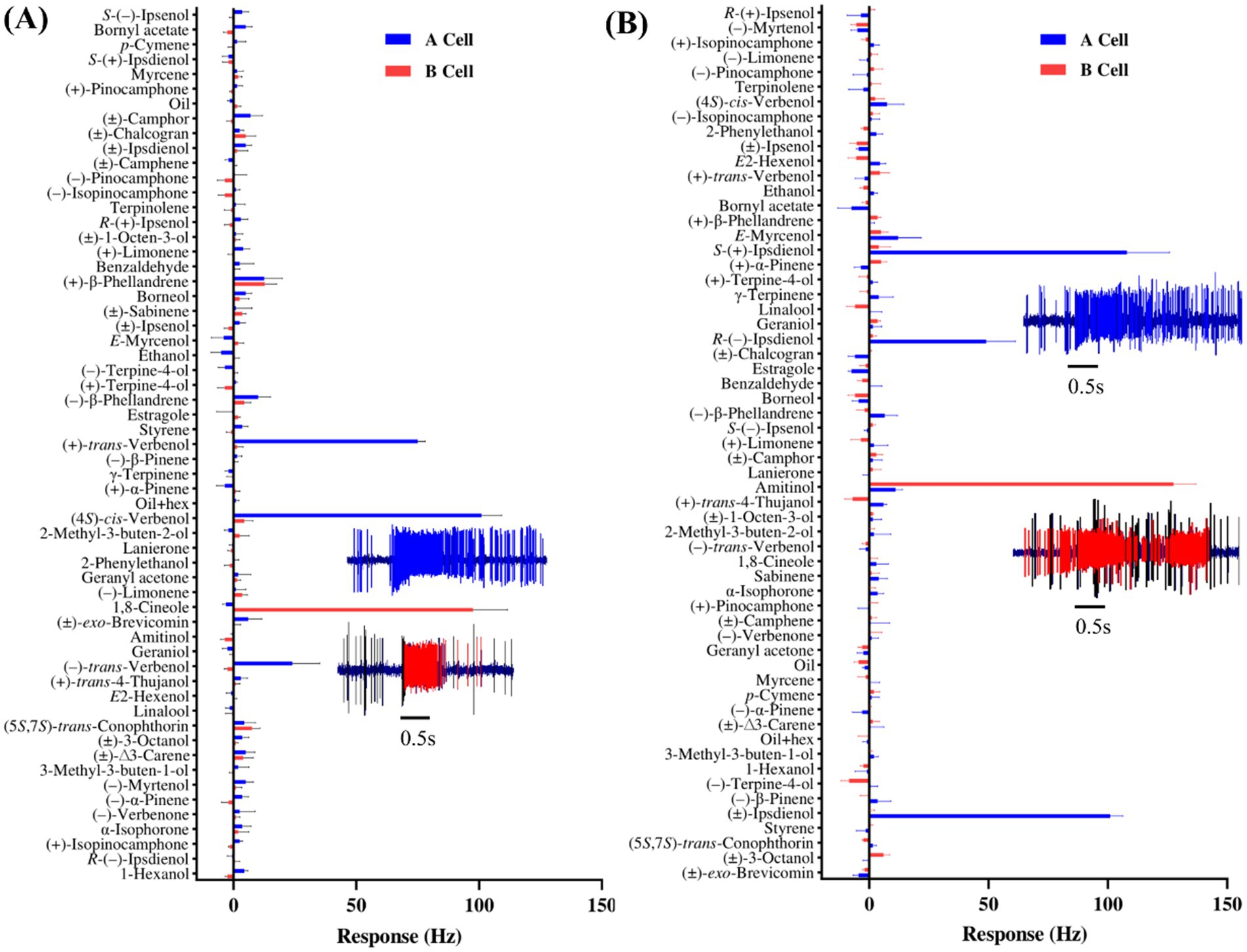
Figure 6. Single sensillum recordings from Ips cembrae, demonstrating co-localization of large spiking (A cell) and small-spiking (B cell) olfactory sensory neurons (OSNs) with responses to various pheromone compounds, volatiles from host- and non-host plants, as well as fungal symbionts (10 μg stimulus dose). (A) The response of A neurons primarily to (4S)-cis-verbenol, co-localized with B neurons that respond to 1,8-cineole (n = 3 sensilla). The representative action potential traces show A neuron response to (4S)-cis-verbenol and B neuron response to 1,8-cineole. (B) The response profile of A neurons responding to S-(+)-ipsdienol, which are co-localized with B neurons responsive to amitinol (n = 3). Representative action potential traces showing A neuron response to S-(+)-ipsdienol and B neuron response to amitinol. Response frequencies (Hz) are displayed as mean ± standard error of the mean (SEM).
The OSN class IC3 (A neuron) was the most frequently observed class (found in ~14% of the responding sensilla), responding strongly (>80 Hz) to racemic ipsenol and S-(−)-ipsenol (n = 9) (Figure 5B). Ipsenol is the major component of the aggregation pheromone in I. cembrae. The response threshold of these OSNs was around 100 pg (Figure 5E). R-(+)-ipsenol elicited minimal responses in this OSN class, consistent with its absence in the natural pheromone blend (Stoakley et al., 1978). This OSN class was uniformly distributed across sensory bands A and B (Figure 5A). The OSN class IC4, which was an A neuron, responded strongly to R-(−)-ipsdienol (n = 3) and weaker to the S-enantiomer and the racemic mixture (Figure 5D). However, we did not find this OSN class during dose–response tests. Additionally, OSN class IC5 (n = 4), also an A neuron, responded specifically to S-(+)-ipsdienol, with a response threshold of 1 ng (Figure 5E). Another OSN class, IC6, was a B neuron responding strongly to amitinol (n = 5); this class was always co-localized with OSN class IC5 (Figure 6B). The S-(+)-ipsdienol-specific OSN class and R-(−)-ipsdienol-specific OSN class were distributed across the sensory band C on the antennal club surface and rarely on sensory band B (Figure 5A). It is noteworthy that during our screening, we did not detect neurons that responded to 3-methyl-3-buten-1-ol, which is a pheromone component of I. cembrae.
3.3.2 OSNs responding to other beetle-produced compounds in I. cembrae
We observed four additional OSN classes strongly responding to different beetle-produced compounds. The OSN class IC7 (A neuron) strongly responded (>80 Hz) to (±)-exo-brevicomin (n = 3), with secondary responses to chalcogran and (5S,7S)-trans-conophthorin (Figure 5D). Another OSN class, IC14, a B neuron, responded strongly to lanierone (n = 2). These neurons were co-localized with OSN class IC9 or a non-responsive A neuron. Additionally, OSN class IC12 class (A neuron) responded strongly (>80 Hz) and specifically to 2-phenylethanol (n = 2) with no secondary responses. OSN class IC15 was an A neuron primarily tuned to chalcogran with secondary responses to 1-hexanol, 1-octen-3-ol, and (5S,7S)-trans-conophthorin (n = 1). OSN class IC16 (A neuron) displayed strong responses (>80 Hz) to amitinol with secondary responses to racemic ipsdienol (n = 1). Lastly, OSN class IC17 was an A neuron, which responded strongly to α-isophorone followed by secondary responses to (−)-verbenone, (+)-trans-verbenol, and (−)-trans-verbenol (n = 1). These OSN classes were mostly found in the distal region of the antennal surface (Figure 5A).
3.3.3 OSN classes responding to host, non-host, and microbial volatiles in I. cembrae
Five OSN classes were specifically tuned to host volatiles, including OSN class IC2, a B neuron specific for 1,8-cineole. The OSN class IC9 showed strong responses (>80 Hz) to camphor (n = 3), with strong secondary responses to (+)-isopinocamphone (>80 Hz) and weaker secondary responses to other related oxygenated monoterpenes, such as (−)-isopinocamphone, (+)-pinocamphone, (−)-pinocamphone, and borneol (Figures 4, 5D). This IC9 A neuron was co-localized with B neurons responding specifically (>80 Hz) to lanierone (IAc14). Additionally, the OSN class IC10 was an A neuron specific for (−)-α-pinene (n = 4, all males). This class also showed weak to intermediate (20–40 Hz) secondary responses to (4S)-cis-verbenol, (+)-α-pinene, (−)-β-pinene, and (+)-trans-verbenol. Another OSN class, IC18 (A neuron), showed strong responses (>80 Hz) to p-cymene (n = 1). Lastly, OSN class IC19 strongly responded (>80 Hz) to estragole (n = 1). The OSN classes corresponding to host volatiles were distributed on the sensory bands B and C on the antennal surface (Figure 5A).
Two OSN classes responded to non-host volatiles. The OSN class IC8 (A neuron) responded primarily to the green leaf volatile 1-hexanol (n = 4). This class also showed weak to intermediate secondary responses (<50 Hz) to other compounds, including chalcogran, racemic 1-octen-3-ol, and 2-phenylethanol. The IC13 class was also an A neuron responding to racemic 1-octen-3-ol (n = 2), with weaker secondary responses to racemic 3-octanol. These OSN classes were observed on sensory bands B and C on the antennal surface (Figure 5A). Additionally, the OSN class IC11 (A neuron) responded to microbial volatile styrene (n = 2, both males) (Figure 4), with weak secondary responses (<40 Hz) to benzaldehyde.
3.4 Comparative analysis of OSN profiles and distribution among I. acuminatus and I. cembrae with previously characterized I. typographus
Our comparative analysis revealed that I. acuminatus, I. cembrae, and I. typographus show both conserved olfactory adaptations and species-specific differences (Table 1, Supplementary Figure 2). Of the 23 OSN classes identified in I. typographus (Andersson et al., 2009; Kandasamy et al., 2019, 2023; Yuvaraj et al., 2024) and the 19 OSN classes found in both I. cembrae and I. acuminatus, 11 were shared among all three species based on similarities in response profiles. These shared OSN classes were tuned to beetle-produced compounds, host or non-host tree, and microbial volatiles (See Table 1 for details on OSN classes). Four OSN classes, specific to 2-methyl-3-buten-2-ol, (−)-verbenone, α-isophorone, (+)-isopinocamphone, and (5S,7S)-trans-conophthorin, respectively, were found exclusively in I. typographus and I. acuminatus (Andersson et al., 2009; Kandasamy et al., 2019, 2023), while two OSN classes tuned to p-cymene and estragole were shared between I. typographus and I. cembrae (Andersson et al., 2009; Raffa et al., 2016). Additionally, OSN classes specific to racemic chalcogran and α-isophorone were shared between I. acuminatus and I. cembrae. Species-specific OSN differences were particularly evident in responses to host and fungal volatiles. I. cembrae had four unique OSN classes tuned to amitinol (B neuron), racemic camphor, racemic exo-brevicomin, and (−)-α-pinene, respectively, whereas I. acuminatus had two unique OSN classes tuned to (−)-limonene and γ-terpinene (Supplementary Figure 2).
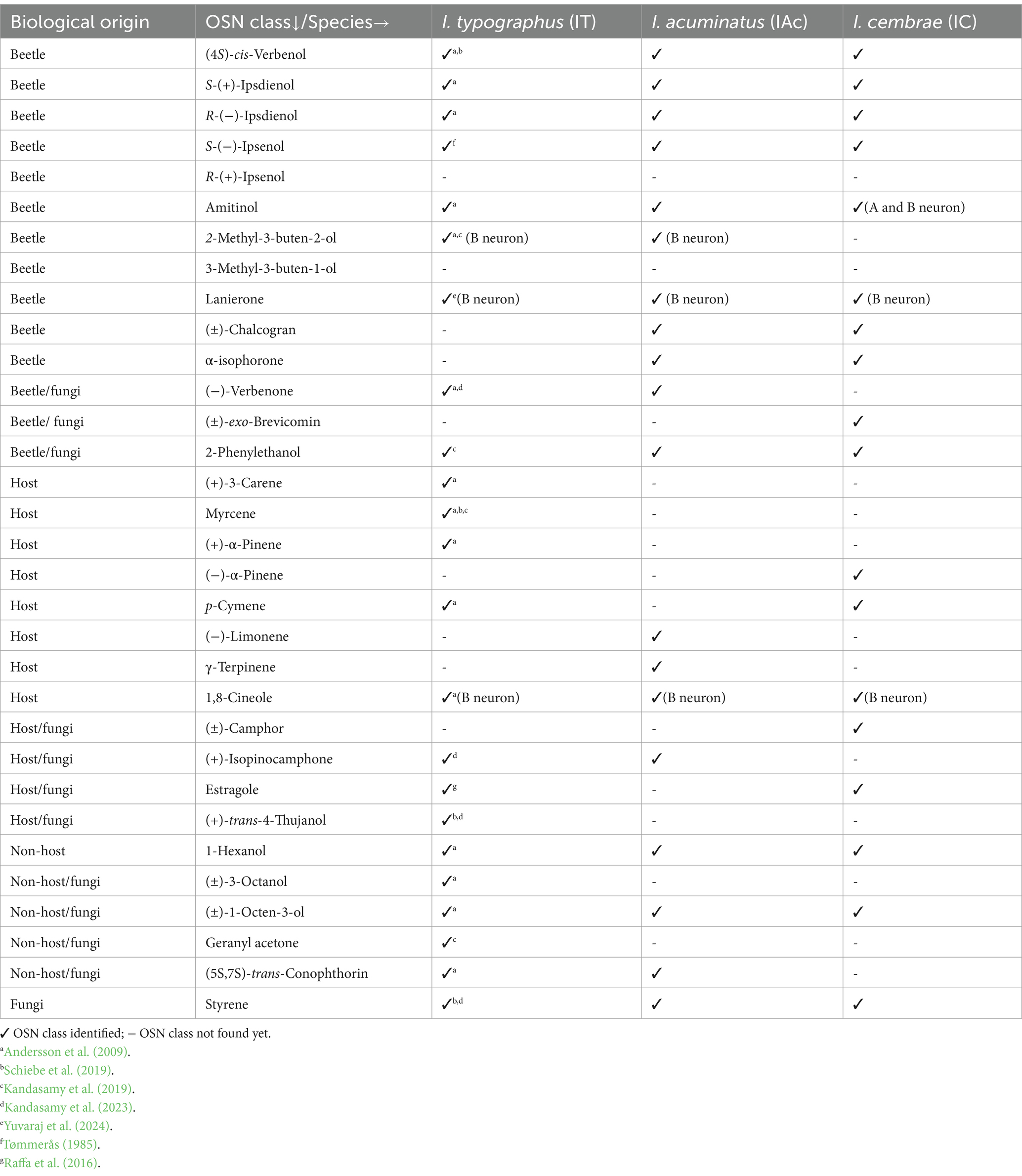
Table 1. Olfactory sensory neurons (OSNs) classified based on their response profiles at a 10 μg screening dose in Ips acuminatus and Ips cembrae and comparison to previously characterized OSN classes in Ips typographus.
3.5 Antennal responses of pine and larch essential oils in I. acuminatus and I. cembrae using GC-EAD
Since only a few OSN classes specific to host monoterpenes were identified in I. acuminatus (OSN classes IAc2, IAc15, and IAc18) and I. cembrae (OSN classes IC2 and IC9, IC18, and IC19), we further evaluated the antennal responses of both species to monoterpenes using GC-EAD analysis. We tested the antennae of I. acuminatus with pine essential oil and I. cembrae with larch essential oil, both with known chemical compositions, to assess their olfactory sensitivity to host-related compounds. GC-EAD analyses with pine essential oil revealed four potential chemical cues that elicited antennal responses from I. acuminatus, whereas I. cembrae responded to five potential cues in the larch essential oil (Figure 7). The EAD active compounds that elicited antennal response in I. acuminatus were identified as α-pinene, limonene, linalool, and isobornyl acetate (Figure 7A) while I. cembrae responded to β-pinene, p-cymene, linalool, terpinen-4-ol, and camphor (Figure 7B). Surprisingly, no EAD responses were observed in I. acuminatus to highly abundant pine host volatiles, such as 3-carene, terpinolene, and β-phellandrene. Similarly, I. cembrae showed no responses to key larch volatiles, including α-pinene, limonene, β-phellandrene, and myrcene.
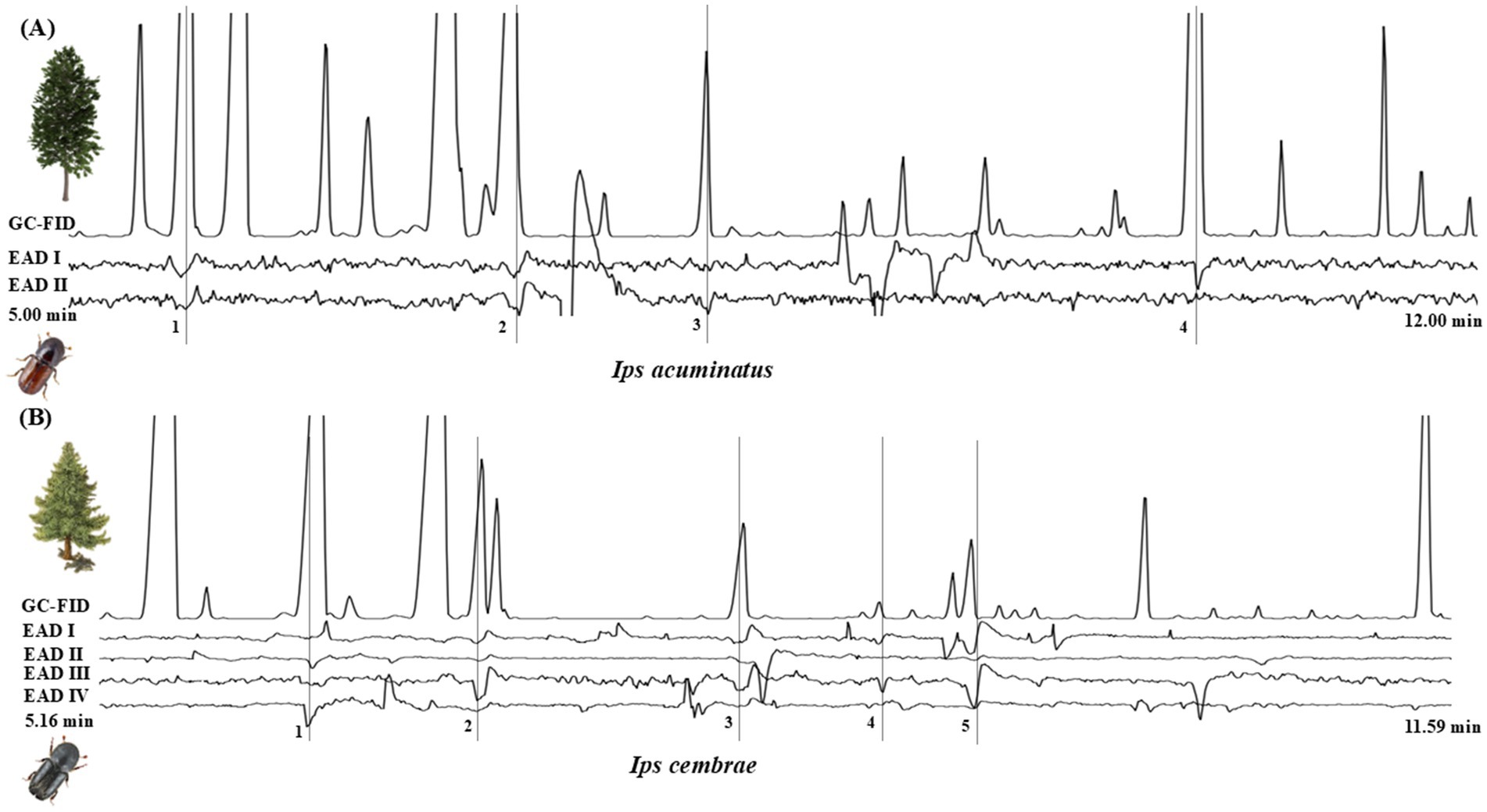
Figure 7. GC-EAD antennal responses at 10 ug of essential oil. (A) Ips acuminatus responses to pine essential oil, with peaks 1, 2, 3, and 4 corresponding to α-pinene, limonene, linalool, and isobornyl acetate, respectively (n = 2). (B) Responses of Ips cembrae to larch essential oil, with peaks 1, 2, 3, 4, and 5 corresponding to β-pinene, p-cymene, linalool, camphor, and terpinen-4-ol, respectively (n = 4).
4 Discussion
This study provides the first electrophysiological characterization of olfactory sensory neuron (OSN) responses in I. acuminatus and I. cembrae. By testing a comprehensive panel of ecologically relevant compounds, including pheromones, volatiles from the hosts and non-host trees, and associated microbes, we identified 19 OSN classes in both species. Most OSN classes exhibited narrow tuning, responding strongly to only one or a few structurally similar compounds, while fewer were broadly tuned. Furthermore, several of our dose–response tests in both species revealed greater OSN specificity at lower doses, consistent with findings in I. typographus, where OSNs exhibited high specificity to either pheromones or to compounds from the host or non-host trees and microbes (Andersson et al., 2009; Kandasamy et al., 2019, 2023).
Ips acuminatus and I. cembrae have several OSNs tuned to the enantiomers of ipsenol and ipsdienol, their key aggregation pheromone components (Francke and Vité, 1983; Renwick and Dickens, 1979). Interestingly, the ipsenol-responsive OSNs were highly specific, showing the strongest responses to the naturally occurring enantiomer, S-(−)-ipsenol, the main pheromone component in both species. This finding is consistent with previous studies in I. typographus and other Ips species such as I. pini and I. paraconfusus (Mustaparta et al., 1979; Mustaparta et al., 1980; Tømmerås, 1985). Also, the ipsdienol-responsive OSNs demonstrated enantiomer-specific tuning, suggesting two distinct OSN classes responding to R-(−)-ipsdienol and S-(+)-ipsdienol, respectively, in both species. In I. acuminatus, initial screenings identified only one OSN class primarily tuned to R-(−)-ipsdienol. However, dose–response tests revealed a stronger response to S-(+)-ipsdienol, suggesting the presence of distinct OSN classes tuned to each ipsdienol enantiomer. In I. cembrae, screenings identified distinct OSN classes specifically tuned to each ipsdienol enantiomer. However, dose–response testing was conducted only for S-(+)-ipsdienol, which exhibited high sensitivity. These results align with earlier reports of two enantiomer-specific ipsdienol-responsive OSN classes in I. typographus and other Ips species (Mustaparta et al., 1980; Tømmerås, 1985). Behavioral studies in I. acuminatus suggest that S-(+)-ipsdienol and S-(−)-ipsenol function as attractants, while R-(−)-ipsdienol likely serves as an attraction inhibitor in field (Bakke, 1978; Kohnle et al., 1986). (4S)-cis-Verbenol elicited strong responses in an OSN class in both I. acuminatus and I. cembrae. While this compound serves as a key aggregation pheromone component in I. acuminatus, it does not play a similar role in I. cembrae, despite the detection of trace amounts in this species (Kohnle et al., 1988). Interestingly, the presence of (4S)-cis-verbenol disrupts I. cembrae aggregation in field studies, possibly serving as an interspecific signal from I. typographus, which relies on this compound as a key aggregation pheromone (Schlyter et al., 1989).
A few examples of OSN co-localization were observed in both I. acuminatus and I. cembrae. The co-localization patterns of ipsdienol-responsive OSNs differed between the two species. In I. cembrae, S-(+)-ipsdienol-responsive A neurons were co-localized with amitinol-responsive B neurons. In contrast, in I. acuminatus, R-(−)-ipsdienol-responsive A neurons were co-localized with lanierone-responsive B neurons, corresponding to the observations in I. typographus (Yuvaraj et al., 2024). Additionally, in both species, (4S)-cis-verbenol- and 1,8-cineole-responsive OSNs were co-localized within the same sensilla, consistent with the co-localization pattern previously reported in I. typographus (Andersson et al., 2009). In I. typographus, such co-localization is thought to enhance the ability to differentiate odors based on spatial and temporal cues. It may also improve sensitivity to ecologically relevant odor blends by detecting specific ratio differences and regulating olfactory signaling at the peripheral level (Andersson et al., 2010; Baker et al., 1998; Binyameen et al., 2014; Bruce et al., 2005). Similar mechanisms may play a role in I. acuminatus and I. cembrae. Although the OSN responses in our study were characterized using pure compounds, it is well established that most insect semiochemicals function as multicomponent blends, with specific behavioral roles determined by the precise ratio and combination of constituents (Silverstein and Young, 1976). Such blends can activate distinct combinations of neurons depending on the ecological context, ultimately shaping behavioral outcomes.
Additionally, several OSN classes in I. cembrae and I. acuminatus responded to pheromone components produced by other Ips species, suggesting conserved detection mechanisms (Andersson et al., 2009; Tømmerås, 1985; Yuvaraj et al., 2024). Amitinol is not an aggregation pheromone component in either I. acuminatus (Bakke, 1978; Francke et al., 1986) or I. cembrae (Kohnle et al., 1986; Stoakley et al., 1978), despite one earlier report mentioning its presence in I. cembrae (Kohnle et al., 1986). Amitinol-responsive OSNs were identified in both species, exhibiting strong secondary responses to racemic ipsdienol, likely due to structural similarity and the presence of trace amounts of amitinol in the ipsdienol stimulus, corresponding to OSNs observed in I. typographus (Andersson et al., 2012b; Andersson et al., 2009). Interestingly, field studies indicate that amitinol enhances aggregation in I. cembrae but reduces attraction in I. acuminatus, potentially mediating interspecific interactions (Francke et al., 1986; Kohnle et al., 1986). Lanierone-responsive OSNs were identified in both species studied here, but the frequency of these OSNs was low. This is in absolute contrast to I. typographus, in which lanierone-specific OSNs represent the most abundant of all OSN classes (Yuvaraj et al., 2024). In I. acuminatus, OSN B neurons responsive to 2-methyl-3-buten-2-ol occurred at relatively low abundance, similar to observations in I. typographus (Kandasamy et al., 2019, 2023). Although this compound serves as an aggregation pheromone component in I. typographus (Lanne et al., 1989), its ecological role in I. acuminatus remains uncertain.
The OSN class responsive to (−)-verbenone, a known bark beetle anti-attractant (Frühbrodt et al., 2024), was the most abundant in I. acuminatus. Notably, we observed this OSN class only in female I. acuminatus and not at all in I. cembrae. Ips typographus possesses fewer (−)-verbenone-responsive OSNs than I. acuminatus; however, it exhibits strong behavioral avoidance in both sexes in laboratory assays (Yuvaraj et al., 2024) and in field studies (Schlyter et al., 1989). In I. typographus, (−)-verbenone-responsive neurons also show secondary responses to α- and β-isophorone, of which the latter compound was reported from hindguts of mated females (Birgersson et al., 1984). However, dose–response tests indicate that this OSN class is sensitive to α-isophorone (Kandasamy et al., 2023). In contrast, I. acuminatus possesses two distinct OSN classes: one specifically tuned to (−)-verbenone and another primarily responding to α-isophorone, with weak secondary responses to (−)-verbenone. Similarly, in I. cembrae, an OSN class specific to α-isophorone was also observed, exhibiting secondary responses not only to (−)-verbenone but also to trans-verbenol enantiomers, corresponding to an OSN class previously observed in I. typographus (Kandasamy et al., 2023). The behavioral role of α- and β-isophorone remains unclear.
Ips acuminatus and I. cembrae exhibited OSN responses to pheromones produced by non-Ips bark beetles, differing from OSN responses in I. typographus. Both species had an OSN class highly responsive to chalcogran, a pheromone of many Pityogenes species (Francke, 1977; Francke et al., 1995). Additionally, I. cembrae possessed a separate OSN class tuned to (±)-exo-brevicomin, which is a pheromone of Dendroctonus species and is also produced by a beetle symbiotic fungus (Zhao et al., 2019). This broader heterospecific pheromone detection in I. cembrae may reflect its ability to colonize different hosts, including Abies, Picea, and Pinus trees and frequent interactions with bark beetles from other genera such as Pityophthorus, Pityogenes, and Cryphalus (Postner, 1974; Pfeffer, 1955). In contrast, I. typographus OSNs primarily responsive to the non-host volatile (5S,7S)-trans-conophthorin exhibit strong secondary responses to both chalcogran and exo-brevicomin, likely due to structural similarities (Andersson et al., 2009).
Major monoterpene hydrocarbons such as α-pinene, β-pinene, limonene, and myrcene are key volatiles in coniferous trees, the primary hosts of Ips bark beetles (Wajs et al., 2007). However, OSN classes responding to monoterpenes were relatively rare in both species. In I. acuminatus, one OSN class responded mostly to (−)-limonene, with secondary responses to myrcene, p-cymene, terpinolene, and β-pinene. Another distinct OSN class was tuned specifically to γ-terpinene. Notably, we did not identify OSN classes for key pine volatiles such as α-pinene, 3-carene, β-pinene, and myrcene. In contrast, I. cembrae had an OSN class that responded primarily to (−)-α-pinene, with weak secondary responses to β-pinene. Another OSN class was specifically tuned to p-cymene. However, no OSN class was identified for major larch volatiles such as β-pinene, 3-carene, limonene, and myrcene. Given the suggested role of monoterpenes in bark beetle behavior (Erbilgin et al., 2007), we conducted GC-EAD analyses using pine and larch essential oils. Ips acuminatus antennae exhibited weak responses to α-pinene, limonene, linalool, and isobornyl acetate, whereas I. cembrae responded to β-pinene, p-cymene, linalool, and terpinen-4-ol. The absence or inconsistency of responses to major host volatiles, combined with the finding that I. acuminatus does not exhibit attraction to host trees in field studies (Brattli et al., 1998), suggests that these compounds may not play a primary role in host tree attraction for these species. Volatile compounds produced in minor amounts by Norway spruce, such as 1,8-cineole (Jirošová et al., 2022a; Schiebe et al., 2019) and estragole (Moliterno et al., 2023; Joseph et al., 2001), elicit antennal responses in I. typographus and function as anti-attractants. In this study, we identified OSNs specifically responsive to 1,8-cineole in both I. acuminatus and I. cembrae, while estragole-responsive OSNs were observed only in I. cembrae. Given that 1,8-cineole has been previously linked to conifer resistance against I. typographus attack (Schiebe et al., 2012), its detection by OSNs in the two species examined here suggests a similar ecological role in their host interactions.
Low-abundance oxygenated host monoterpenes, whose concentrations increase in stressed or fungus-infected conifers, likely play a crucial role in beetle discrimination of suitable hosts (Lehmanski et al., 2023). Although present only in trace amounts, these metabolites of monoterpene hydrocarbons can be produced via microbial activity or the tree’s own metabolism and may significantly influence bark beetle host selection and colonization strategies (Moliterno et al., 2023; Kandasamy et al., 2023). In our study, I. acuminatus exhibited strong OSN responses primarily to (+)-isopinocamphone and secondarily to structurally similar pinocamphone and camphor, closely resembling the OSN responses described in I. typographus (Kandasamy et al., 2019, 2023). In contrast, I. cembrae had OSNs primarily responsive to racemic camphor, with secondary responses to pinocamphone and isopinocamphone enantiomers. Isopinocamphone, an oxygenated metabolite of pinene (the main component of pine resin), and camphor, an oxidized metabolite of borneol from larch-derived bornyl acetate, are produced by beetle-symbiotic fungi (Kandasamy et al., 2023), which can also be associated with stressed host trees (Schiebe et al., 2019).
Ips acuminatus and I. cembrae vector different ophiostomatoid fungi (Papek et al., 2024; Jankowiak et al., 2007), whose volatile profiles have not yet been characterized but are likely to differ. Both species exhibited strong OSN responses to fungal volatiles (2-phenylethanol, styrene, 1-octen-3-ol), while (5S,7S)-trans-conophthorin-responsive OSNs were detected only in I. acuminatus. These OSN classes have previously been identified in I. typographus (Andersson et al., 2009; Kandasamy et al., 2019, 2023). trans-Conophthorin has been shown to disrupt aggregation pheromone activity in conifer-infesting bark beetles, including I. typographus, in field studies (Huber et al., 2000; Zhang et al., 2002; Zhang and Schlyter, 2004). These compounds, along with oxygenated host monoterpenes (Kandasamy et al., 2023) and (±)-exo-brevicomin (Zhao et al., 2019), are also produced by fungi. They likely indicate fungus-colonized or weakened host trees, potentially guiding beetles towards suitable hosts. Additionally, fungi may produce volatiles that elicit a positive response from beetles, as they potentially act as nutritional resources for bark beetles (Kandasamy et al., 2019, 2023). However, further research is needed to clarify their precise ecological roles and the mechanisms by which beetles interpret these chemical cues. Both species exhibited OSN responses to NHVs, helping conifer-feeding bark beetles avoid unsuitable angiosperm trees. OSNs in I. acuminatus and I. cembrae responded selectively to 1-hexanol, a known anti-attractant emitted by green leaves of non-host trees (Schlyter et al., 1989, 2000; Zhang et al., 1999).
Some OSN classes were only found in one of the sexes of I. acuminatus and I. cembrae, which may suggest differences in olfactory-driven behaviors between males and females. In I. acuminatus, OSN classes for (−)-verbenone, R-(−)-ipsdienol, and lanierone were observed in females, whereas 2-methyl-buten-2-ol OSNs were found in males. These female-biased responses may be associated with the species’ polygynous mating system, where males form large harems (2–12 females per male), and pseudogamous females breed independently (Kirkendall, 1989, 1990). Thus, these olfactory cues may help females to avoid overcrowded trees and reduce interspecific competition with other conifer bark beetle species (Papek et al., 2024). In I. cembrae, OSN classes specific to (−)-α-pinene and styrene were found in males, suggesting a role in host location. However, further recordings from additional sensilla and behavioral experiments are needed to determine whether these OSN classes are sex-specific, sex-biased, or simply missed during sampling.
The antennal distribution of OSNs varies among species. Ipsenol-responsive OSNs in I. cembrae were located mainly in sensory bands A and B, similar to I. typographus (Andersson et al., 2009), but restricted to band B in I. acuminatus, unless we failed to find them in band A. Conversely, ipsdienol-responsive OSNs occurred predominantly in bands B and C in I. cembrae, while in I. acuminatus they were distributed across bands A and B, resembling the distribution reported in I. typographus (Andersson et al., 2009). OSNs responding to (4S)-cis-verbenol are predominantly located in band C in all three species. These differences in OSN distribution may reflect species-specific olfactory adaptations related to their pheromone detection, distinct host selection, and chemical communication within their distinct ecological contexts. While our study was confined to OSNs housed in the antennae, it is noteworthy to point out that Ips beetles also possess chemosensory sensilla on the maxillary palps (Hallberg, 1982b; Hallberg et al., 2003), potentially capable of detecting less volatile or contact-mediated compounds, a subject of interest, which needs further investigation.
The olfactory responses we observed in I. acuminatus and I. cembrae are consistent with broader insect patterns, where selective olfactory systems are shaped by evolutionary pressures. OSNs are frequently specifically tuned to ecologically important stimuli also in non-beetle species, such as moths and Drosophila, whereas other neurons may be more broadly tuned (Hallem et al., 2004; de Bruyne and Baker, 2008; Andersson et al., 2015). Additionally, other congeneric species often share several conserved OSN classes, and display a few species-specific ones. This has been shown, for example, in beetles from other families, such as clover seed weevils (Apionidae) in the Protapion genus (P. fulvipes and P. trifolii) and scarab beetles (Scarabaeidae) in the Pachnoda genus (P. interrupta and P. marginata) (Bengtsson et al., 2011; Andersson et al., 2012a; Carrasco et al., 2019). At a molecular level, 12 odorant receptors (ORs) have been functionally characterized in I. typographus (Hou et al., 2021; Roberts et al., 2021, 2022; Yuvaraj et al., 2021, 2024; Biswas et al., 2024), with responses resembling several of the OSN responses observed in this study. While many OSN classes identified here exhibit response patterns similar to I. typographus, it remains unknown whether conserved ORs are responsible. Given that I. typographus and I. duplicatus share numerous conserved OR orthologs (Johny et al., 2024), similar conservation is likely in I. acuminatus and I. cembrae. Further OR characterization and comparative genomic analyses across Ips species could provide deeper insights into OSN specificity and pheromone detection mechanisms.
Overall, our findings provide valuable insights for improving bark beetle management by refining pheromone-based strategies. Although the pheromone-baited “trap and kill” approach has shown limited success due to spillover infestations and low overall efficacy (Jakuš et al., 2003), pheromone traps remain useful for monitoring beetle activity. Cross-attraction among Ips species has been observed (Byers, 1989; Etxebeste et al., 2012), emphasizing the need for species-specific approaches. Several anti-aggregation compounds show potential for spruce protection, including verbenone (Frühbrodt et al., 2024), spruce volatiles like trans-4-thujanol and 1,8-cineole (Andersson et al., 2010; Jirošová et al., 2022a), and others such as hexanol, 1-octen-3-ol, and trans-conophthorin (Schiebe et al., 2011; Zhang and Schlyter, 2004; Unelius et al., 2014). These have been repeatedly tested in various combinations against I. typographus (Schiebe et al., 2011; Zhang and Schlyter, 2004; Unelius et al., 2014; Jakuš et al., 2024), though they also suffer from spillover effects due to their repellent nature (Jakuš et al., 2003; Schiebe et al., 2011). Push-pull strategies, which combine anti-attractants with pheromone traps (Jakuš et al., 2022) or baited trap trees (Lindmark et al., 2022), offer a potential improvement. However, their effectiveness decreases under high beetle population density and severe tree stress (Deganutti et al., 2024). The use of anti-attractants for I. acuminatus and I. cembrae remains untested (Frühbrodt et al., 2024), highlighting the need for further behavioral studies with compound combinations designed according to this study to evaluate their field efficacy.
5 Conclusion
This is the first electrophysiological study to functionally characterize OSNs in I. acuminatus and I. cembrae, identifying 19 OSN classes in each species. These OSNs exhibited distinct tuning to aggregation pheromones, host monoterpenes, NHVs, and fungal-derived odors, highlighting their crucial role in bark beetle ecology. Comparative analysis with I. typographus revealed both conserved and species-specific OSN response patterns. While certain OSN profiles were shared across Ips species, suggesting common olfactory strategies for aggregation and host detection, species-specific differences likely reflect adaptations to their respective host tree preferences. The detection of heterospecific pheromones, along with fungal volatiles, further supports the role of multiple chemical cues in species coexistence and host colonization. Although OSN response profiles were generally similar between sexes, further research is needed to determine whether subtle differences influence mate and host selection behaviors. From an applied perspective, our findings support the use of specific compositions of ipsenol and ipsdienol mixtures, including their enantiomeric ratios, in combination with other detected compounds for species-specific Ips beetles monitoring and pest management. Integrating NHVs and host volatiles into conifer tree protection strategies could enhance its efficiency. Furthermore, future studies should explore a broader range of volatile compounds to identify additional OSN classes and incorporate molecular analyses of olfactory receptor function to refine our understanding of olfactory coding mechanisms in bark beetles. A deeper disentangling of these mechanisms could enable targeted interventions by disrupting the detection of key compounds by beetles at the gene level. This study lays the groundwork for further exploration of bark beetle olfactory systems, offering insights into ecological interactions and improved pest management strategies.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because In compliance with local legislation and institutional standards, the animal study did not require ethical review or approval. We have performed all beetle experiments that comply with the ARRIVE guidelines and were carried out in accordance with (Scientific Procedures) Act, 1986 and associated guidelines, EU Directive 2010/63/EU for animal experiments.
Author contributions
MKS: Data curation, Formal analysis, Investigation, Validation, Visualization, Writing – original draft, Writing – review & editing. JB: Data curation, Investigation, Writing – original draft. JS: Data curation, Writing – review & editing. MS: Methodology, Writing – review & editing. MA: Conceptualization, Methodology, Supervision, Validation, Writing – review & editing. DK: Conceptualization, Methodology, Supervision, Validation, Writing – review & editing. AJ: Conceptualization, Methodology, Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. MKS and AJ were funded by the funding agency, the Czech Science Foundation GACR 23- 07916s, Czech Republic. Research funding Internal Grant Commission at the Faculty of Forestry and Wood Sciences, Czech University of Life Sciences Prague, Czech Republic; [MKS, IGA: A_08_24] and ERASMUS Exchange Program for funding research internship at Lund University, Sweden. Publication fee funding: Faculty of Forestry and Wood Sciences, Czech University of Life Sciences, Prague. MS was financially supported by the Austrian Federal Ministry of Agriculture, Forestry, Regions and Water Management (‘Waldfonds’ projects ‘NewIPS’, grant no. 101686, and ‘IpsEMAN’, grant no. 101687). DK was funded by the Swedish Research Council VR (grant no: 2023-05256) and MA was funded by grants from the Swedish Research Council VR (grant no. 2022-03597), Formas (grant no. 2022-00902), and the Foundation in Memory of Oscar and Lili Lamm.
Acknowledgments
We thank Eva Papek (BOKU University, Vienna, Austria) for providing the beetles used in this study for the SSR experiments. We also thank Erling Jirle (Lund University, Sweden) for his valuable technical support. We acknowledge Venkatesh Pal Mahadevan and the Department of Forest Protection and Entomology, CZU Prague. We are grateful to Jakub Dušek for providing scanning electron micrographs of I. acuminatus and I. cembrae.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that Gen AI was used in the creation of this manuscript. The English language was edited using ScholarAI (2025), ScholarAI: AI-powered research assistant. OpenAI. Available at: https://notilo.ai.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2025.1588866/full#supplementary-material
SUPPLEMENTARY FIGURE 1 | Different olfactory sensory neurons (OSNs) showing unique response patterns when stimulated with 10 µg of each compound. The excitatory responses were predominantly phasic-tonic, although the extent of tonic firing varied among neurons. Typically, two OSNs (A,B neurons), with different spike amplitude were observed within the same sensillum. Panel (A) illustrates the responses of OSNs in I. acuminatus to α-isophorone, lanierone, (±)-chalcogran, 1-octen-3-ol, 2-phenylethanol, and (+)-isopinocamphone, whereas panel (B) shows the responses of OSNs in I. cembrae to α-isophorone, 2-octen-3-ol, (+)-isopinocamphone, and (±)-chalcogran.
SUPPLEMENTARY FIGURE 2 | Venn diagram illustrating the overlap in detected OSN classes among the three Ips species (I. typographus, I. acuminatus, and I. cembrae). The blue circle represents I. typographus, the orange circle represents I. acuminatus, and the green circle represents I. cembrae. Overlapping regions indicate OSN classes shared between two or more species, while unique sections represent species-specific OSN classes. The numbers within each region correspond to the number of OSN classes detected in that category. Data for I. typographus OSNs was sourced from Andersson et al. (2009); Schiebe et al. (2012); Kandasamy et al. (2019, 2023).
References
Allen, C. D., Breshears, D. D., and McDowell, N. G. (2015). On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 6, 1–55. doi: 10.1890/ES15-00203.1
Andersson, M. N., Larsson, M. C., Blaženec, M., Jakuš, R., Zhang, Q.-H. H., and Schlyter, F. (2010). Peripheral modulation of pheromone response by an inhibitory host compound in a beetle. J. Exp. Biol. 213, 3332–3339. doi: 10.1242/jeb.044396
Andersson, M. N., Larsson, M. C., and Schlyter, F. (2009). Specificity and redundancy in the olfactory system of the bark beetle Ips typographus: single-cell responses to ecologically relevant odors. J. Insect Physiol. 55, 556–567. doi: 10.1016/j.jinsphys.2009.01.018
Andersson, M. N., Larsson, M. C., Svensson, G. P., Birgersson, G., Rundlöf, M., Lundin, O., et al. (2012a). Characterization of olfactory sensory neurons in the white clover seed weevil, Apion fulvipes (Coleoptera: Apionidae). J. Insect Physiol. 58, 1325–1333. doi: 10.1016/j.jinsphys.2012.07.006
Andersson, M. N., Löfstedt, C., and Newcomb, R. D. (2015). Insect olfaction and the evolution of receptor tuning. Front. Ecol. Evol. 3:53. doi: 10.3389/fevo.2015.00053
Andersson, M. N., Schlyter, F., Hill, S. R., and Dekker, T. (2012b). What reaches the antenna? How to calibrate odor flux and ligand-receptor affinities. Chem. Senses 37, 403–420. doi: 10.1093/chemse/bjs009
Ascoli-Christensen, A., Salom, S. M., and Payne, T. L. (1993). Olfactory receptor cell responses of Ips grandicollis (Eichhoff) (Coleoptera: Scolytidae) to intra- and interspecific behavioral chemicals. J. Chem. Ecol. 19, 699–712. doi: 10.1007/BF00985002
Baker, T. C., Fadamiro, H. Y., and Cosse, A. A. (1998). Moth uses fine tuning for odour resolution. Nature 393:530. doi: 10.1038/31131
Bakke, A. (1978). Aggregation pheromone components of the bark beetle Ips acuminatus. Oikos 31, 184–190. doi: 10.2307/3543561
Bengtsson, J. M., Khbaish, H., Reinecke, A., Wolde-Hawariat, Y., Negash, M., Seyoum, E., et al. (2011). Conserved, highly specialized olfactory receptor neurons for food compounds in two congeneric scarab beetles, Pachnoda interrupta and Pachnoda marginata. Chem. Senses 36, 499–513. doi: 10.1093/chemse/bjr002
Benton, R., Vannice, K. S., Gomez-Diaz, C., and Vosshall, L. B. (2009). Variant ionotropic glutamate receptors as chemosensory receptors in Drosophila. Cell 136, 149–162. doi: 10.1016/j.cell.2008.12.001
Biedermann, P. H., Müller, J., Grégoire, J. C., Gruppe, A., Hagge, J., Hammerbacher, A., et al. (2019). Bark beetle population dynamics in the Anthropocene: challenges and solutions. Trends Ecol. Evol. 34, 914–924. doi: 10.1016/j.tree.2019.06.002
Binyameen, M., Blaženec, M., Jakuš, R., Song, L., Jankuvová, J., Andersson, M. N., et al. (2014). Co-localization of insect olfactory sensory cells improves the discrimination of closely separated odor sources. Funct. Ecol. 28, 1216–1223. doi: 10.1111/1365-2435.12252
Birgersson, G., Schlyter, F., Löfqvist, J., and Bergström, G. (1984). Quantitative variation of pheromone components in the spruce bark beetle Ips typographus from different attack phases. J. Chem. Ecol. 10, 1029–1055. doi: 10.1007/BF00987511
Biswas, T., Sims, C., Yuvaraj, J. K., Roberts, R. E., Löfstedt, C., and Andersson, M. N. (2024). Functional characterization supports multiple evolutionary origins of pheromone receptors in bark beetles. Mol. Biol. Evol. 40, 1–15. doi: 10.1093/molbev/msad232
Brattli, J. G., Andersen, J., and Nilssen, A. C. (1998). Primary attraction and host tree selection in deciduous and conifer-living Coleoptera: Scolytidae, Curculionidae, Cerambycidae, and Lymexylidae. J. Appl. Entomol. 122, 345–352. doi: 10.1111/j.1439-0418.1998.tb01511.x
Bruce, T. J. A., Wadhams, L. J., and Woodcock, C. M. (2005). Insect host location: a volatile situation. Trends Plant Sci. 10, 269–274. doi: 10.1016/j.tplants.2005.04.003
Byers, J. A. (1989). Chemical ecology of bark beetles. Experientia 45, 271–283. doi: 10.1007/BF01951813
Byers, J. A. (2007). “Chemical ecology of bark beetles in a complex olfactory landscape” in Bark and Wood boring insects in living trees in Europe, a synthesis. eds. F. Lieutier, K. R. Day, A. Battisti, J. C. Grégoire, and H. F. Evans (Dordrecht: Springer), 89–134.
Carrasco, D., Nyabuga, F. N., Anderbrant, O., Svensson, G. P., Birgersson, G., Lankinen, Å., et al. (2019). Characterization of olfactory sensory neurons in the red clover seed weevil, Protapion trifolii (Coleoptera: Brentidae), and comparison to the closely related species P. fulvipes. J. Insect Physiol. 119:103948. doi: 10.1016/j.jinsphys.2019.103948
Christiansen, E., and Bakke, A. (1988). “The spruce bark beetle of Eurasia” in Dynamics of Forest insect populations. Population ecology. ed. A. A. Berryman (Boston, MA: Springer).
Clyne, P. J., Warr, C. G., Freeman, M. R., Lessing, D., Kim, J., and Carlson, J. R. (1999). A novel family of divergent seven-transmembrane proteins: candidate odorant receptors in Drosophila. Neuron 22, 327–338. doi: 10.1016/s0896-6273(00)81093-4
De Bruyne, M., and Baker, T. C. (2008). Odor detection in insects: volatile codes. J. Chem. Ecol. 34, 882–897. doi: 10.1007/s10886-008-9485-4
Deganutti, L., Biscontin, F., Bernardinelli, I., and Faccoli, M. (2024). The semiochemical push-and-pull technique can reduce bark beetle damage in disturbed Norway spruce forests affected by the Vaia storm. Agric. For. Entomol. 26, 115–125. doi: 10.1111/afe.12600
Dobor, L., Hlásny, T., and Zimová, S. (2020). Contrasting vulnerability of monospecific and species-diverse forests to wind and bark beetle disturbance: the role of management. Ecol. Evol. 10, 12233–12245. doi: 10.1002/ece3.6854
EFSA on Plant HealthJeger, M., Bragard, C., Caffier, D., Candresse, T., Chatzivassiliou, E., et al. (2017). Pest categorization of Ips cembrae. EFSA J. 15:e05039. doi: 10.2903/j.efsa.2017.5040
Erbilgin, N., Krokene, P., Kvamme, T., and Christiansen, E. (2007). A guest monoterpene influences Ips typographus (Coleoptera: Curculionidae, Scolytinae) responses to its aggregation pheromone. Agric. For. Entomol. 9, 135–140. doi: 10.1111/j.1461-9563.2007.00329.x
Etxebeste, I., Álvarez, G., Pérez, G., and Pajares, J. A. (2012). Field response of the six-toothed pine bark beetle, Ips sexdentatus (Col.: Curculionidae, Scolytinae), to pheromonal blend candidates. J. Appl. Entomol. 136, 431–444. doi: 10.1111/j.1439-0418.2011.01682.x
Foit, J., and Čermák, V. (2014). Colonization of disturbed scots pine trees by bark- and wood-boring beetles. Agric. For. Entomol. 16, 184–195. doi: 10.1111/afe.12048
Francke, W. (1977). 2-ethyl-l,6-dioxaspiro[4.4] nonane, principal aggregation pheromone of Pityogenes chalcographus (L.). Entomology 64, 590–591. doi: 10.1007/BF00450651
Francke, W., Bartels, J., Meyer, H., Schroder, F., Kohnle, U., Baader, E., et al. (1995). Semiochemicals from bark beetles: new results, remarks, and reflections. J. Chem. Ecol. 21, 1043–1063. doi: 10.1007/BF02033807
Francke, W., Pan, M.-L., Bartels, J., König, W. A., Vité, J. P., Krawielitzki, S., et al. (1986). The odour bouquet of three pine engraver beetles (Ips spp.). J. Appl. Entomol. 101, 453–461. doi: 10.1111/j.1439-0418.1986.tb00879.x
Francke, W., and Vité, J. P. (1983). Oxygenated terpenes in pheromone systems of bark beetles. Z. Angew. Entomol. 96, 146–156. doi: 10.1111/j.1439-0418.1983.tb03655.x
Francke-Grosmann, H. (1965). Ein Symbioseorgan bei dem Borkenkäfer Dendroctonus frontalis Zimm. (Coleoptera, Scolytidae). Naturwissenschaften 52:143. doi: 10.1007/BF00638532
Frühbrodt, T., Schebeck, M., Andersson, M. N., Holighaus, G., Kreuzwieser, J., Burzlaff, T., et al. (2024). Verbenone—the universal bark beetle repellent? Its origin, effects, and ecological roles. J. Pest. Sci. 97, 35–71. doi: 10.1007/s10340-023-01635-3
Grodzki, W. (2008). Ips cembrae Heer. (Col.: Curculionidae, Scolytinae) in young larch stands – a new problem in Poland. Forstschutz Aktuell 44, 1–42.
Hallberg, E. (1982a). Sensory organs in Ips typographus (Insecta: Coleoptera)—fine structure of antennal sensilla. Protoplasma 111, 206–214. doi: 10.1007/BF01281968
Hallberg, E. (1982b). Sensory organs in Ips typographus (Insecta: Coleoptera): fine structure of the sensilla of the maxillary and labial palps. Acta Zool. 63, 191–198. doi: 10.1111/j.1463-6395.1982.tb00778.x
Hallberg, E., Hansson, B., and Löfstedt, C. (2003). “Sensilla and proprioceptors” in Teilband/Part 36 Vol 2: Morphology, Physiology, and Development. ed. W. Kükenthal, (Berlin, Boston: De Gruyter) 267–288. doi: 10.1515/9783110893724.267
Hallem, E. A., Ho, M. G., and Carlson, J. R. (2004). The molecular basis of odor coding in the Drosophila antenna. Cell 117, 965–979. doi: 10.1016/j.cell.2004.05.012
Heberle, H., Meirelles, G. V., da Silva, F. R., Telles, G. P., and Minghim, R. (2015). InteractiVenn: a web-based tool for the analysis of sets through Venn diagrams. BMC Bioinformatics 16:169. doi: 10.1186/s12859-015-0611-3
Hlásny, T., Zimová, S., Merganičová, K., Štěpánek, P., Modlinger, R., and Turčáni, M. (2021). Devastating outbreak of bark beetles in the Czech Republic: drivers, impacts, and management implications. For. Ecol. Manag. 490:119075. doi: 10.1016/j.foreco.2021.119075
Hlávková, D., and Doležal, P. (2022). Cambioxylophagous pests of scots pine: ecological physiology of European populations—a review. Front. For. Glob. Change 5:806571. doi: 10.3389/ffgc.2022.806571
Hou, X. Q., Yuvaraj, J. K., Roberts, R. E., Zhang, D. D., Unelius, C. R., Löfstedt, C., et al. (2021). Functional evolution of a bark beetle odorant receptor clade detecting monoterpenoids of different ecological origins. Mol. Biol. Evol. 38, 4934–4947. doi: 10.1093/molbev/msab218
Huber, D. P. W., Gries, R., Borden, J. H., and Pierce, H. D. Jr. (2000). A survey of antennal responses by five species of coniferophagous bark beetles (Coleoptera: Scolytidae) to bark volatiles of six species of angiosperm trees. Chemoecology 10, 103–113. doi: 10.1007/PL00001811
Hulcr, J., Atkinson, T. H., Cognato, A. I., Jordal, B. H., and McKenna, D. D. (2015). “Morphology, taxonomy, and phylogenetics of bark beetles” in Bark beetles. eds. F. E. Vega and R. W. Hofstetter (London: Academic Press), 41–84. doi: 10.1016/B978-0-12-417156-5.00002-2
Jaime, L., Batllori, E., and Lloret, F. (2024). Bark beetle outbreaks in coniferous forests: a review of climate change effects. Eur. J. For. Res. 143, 1–17. doi: 10.1007/s10342-023-01623-3
Jakoby, O., Lischke, H., and Wermelinger, B. (2019). Climate change alters elevational phenology patterns of the European spruce bark beetle (Ips typographus). Glob. Chang. Biol. 25, 4048–4063. doi: 10.1111/gcb.14766
Jakuš, R., Modlinger, R., Kašpar, J., Majdak, A., Blaženec, M., Korolyova, N., et al. (2022). Testing the efficiency of the push-and-pull strategy during severe Ips typographus outbreak and extreme drought in Norway spruce stands. Forests 13:2175. doi: 10.3390/f13122175
Jakuš, R., Schlyter, F., Zhang, Q. H., Blaženec, M., Vaverčák, R., Grodzki, W., et al. (2003). Overview of development of an anti-attractant based technology for spruce protection against Ips typographus: from past failures to future success. Anz. Schädlingskd. J. Pest Sci. 76, 89–99. doi: 10.1046/j.1439-0280.2003.03020.x
Jakuš, R., Trubin, A., Singh, V. V., Zabihi, K., Jirošová, A., Modlinger, R., et al. (2024). Spruce protection against Ips typographus with anti-attractant blend of tree-based Semiochemicals: from small experimental plots to stand scales. Forests 15:356. doi: 10.3390/f15020356
Jankowiak, R., Rossa, R., and Mista, K. (2007). Survey of fungal species vectored by Ips cembrae to European larch trees in Raciborskie forests (Poland). Czech Mycol. 59, 227–239. doi: 10.33585/cmy.59209
Jirošová, A., Kalinová, B., Modlinger, R., Jakuš, R., Unelius, C. R., Blaženec, M., et al. (2022a). Anti-attractant activity of (+)-trans-4-thujanol for Eurasian spruce bark beetle Ips typographus: novel potency for females. Pest Manag. Sci. 78, 1992–1999. doi: 10.1002/ps.6819
Jirošová, A., Modlinger, R., Hradecký, J., Ramakrishnan, R., Beránková, K., and Kandasamy, D. (2022b). Ophiostomatoid fungi synergize attraction of the Eurasian spruce bark beetle Ips typographus to its aggregation pheromone in field traps. Front. Microbiol. 13:980251. doi: 10.3389/fmicb.2022.980251
Johny, J., Große-Wilde, E., Kalinová, B., and Roy, A. (2024). Antennal transcriptome screening and identification of chemosensory proteins in the double-spine European spruce bark beetle, Ips duplicatus (Coleoptera: Scolytinae). Int. J. Mol. Sci. 25:9513. doi: 10.3390/ijms25179513
Joseph, G., Kelsey, R. G., Peck, R. W., and Niwa, C. G. (2001). Response of some scolytids and their predators to ethanol and 4-allylanisole in pine forests of Central Oregon. J. Chem. Ecol. 27, 697–715. doi: 10.1023/A:1010345817756
Kandasamy, D., Gershenzon, J., Andersson, M. N., and Hammerbacher, A. (2019). Volatile organic compounds influence the interaction of the Eurasian spruce bark beetle (Ips typographus) with its fungal symbionts. ISME J. 13, 1788–1800. doi: 10.1038/s41396-019-0390-3
Kandasamy, D., Zaman, R., Nakamura, Y., Zhao, T., Hartmann, H., Andersson, M. N., et al. (2023). Conifer-killing bark beetles locate fungal symbionts by detecting volatile fungal metabolites of host tree resin monoterpenes. PLoS Biol. 21:e3001887. doi: 10.1371/journal.pbio.3001887
Keeling, C. I., Tittiger, C., MacLean, M., and Blomquist, G. J. (2021). “Pheromone production in bark beetles” in Insect pheromone biochemistry and molecular biology. eds. G. J. Blomquist and R. G. Vogt (Cambridge, MA: Academic Press) 2nd ed, 123–162. doi: 10.1016/B978-0-12-819628-1.00004-3
Kirisits, T. (2004). “The large larch bark beetle (Ips cembrae) as vector of blue-stain fungi” in Forstschutz Aktuell. (Wien, Austria: Bundesamt und Forschungszentrum für Wald (BFW), Institut für Forstschutz), 19–23.
Kirkendall, L. R. (1989). Within-harem competition among Ips females, an overlooked component of density-dependent larval mortality. Univ. Bergen 12, 477–487. doi: 10.1111/j.1600-0587.1989.tb00925.x
Kirkendall, L. R. (1990). Sperm is a limiting resource in the pseudogamous bark beetle Ips acuminatus (Scolytidae). Oikos 57:80. doi: 10.2307/3565740
Knížek, M., and Beaver, R. (2007). “Taxonomy and systematics of bark and ambrosia beetles” in Bark and Wood boring insects in living trees in Europe, a synthesis. eds. F. Lieutier, K. R. Day, A. Battisti, J. C. Grégoire, and H. F. Evans (Dordrecht: Springer), 141–176. doi: 10.1007/978-1-4020-2241-8_5
Kohnle, U., Kopp, S., and Francke, W. (1986). Inhibition of the attractant pheromone response in Ips acuminatus (Gyll.) by I. sexdentatus (Boerner) (Coleoptera, Scolytidae). J. Appl. Entomol. 101, 316–319. doi: 10.1111/j.1439-0418.1986.tb00864.x
Kohnle, U., Vité, J. P., Erbacher, C., Bartels, J., and Francke, W. (1988). Aggregation response of European engraver beetles of the genus Ips mediated by terpenoid pheromones. Entomol. Exp. Appl. 49, 43–53. doi: 10.1111/j.1570-7458.1988.tb02475.x
Krokene, P. (2015). “Conifer defense and resistance to bark beetles” in Bark beetles. eds. F. E. Vega and R. W. Hofstetter (London: Academic Press), 177–207. doi: 10.1016/B978-0-12-417156-5.00005-8
Lanne, B. S., Ivarsson, P., Johnsson, P., Bergström, G., and Wassgren, A. B. (1989). Biosynthesis of 2-methyl-3-buten-2-ol, a pheromone component of Ips typographus (Coleoptera: Scolytidae). Insect Biochem. 19, 163–167. doi: 10.1016/0020-1790(89)90087-5
Lehmanski, L. M., Kandasamy, D., Andersson, M. N., Netherer, S., Alves, E. G., Huang, J., et al. (2023). Addressing a century-old hypothesis–do pioneer beetles of Ips typographus use volatile cues to find suitable host trees? New Phytol. 238, 1762–1770. doi: 10.1111/nph.18865
Lindmark, M., Wallin, E. A., and Jonsson, B. G. (2022). Protecting forest edges using trap logs – limited effects of associated push-pull strategies targeting Ips typographus. For. Ecol. Manag. 505:119886. doi: 10.1016/j.foreco.2021.119886
Liška, J., Knížek, M., and Véle, A. (2021). Evaluation of insect pest occurrence in areas of calamitous mortality of scots pine. Cent. Eur. For. J. 67, 85–90. doi: 10.2478/forj-2021-0006
Moliterno, A. A. C., Jakuš, R., Modlinger, R., Unelius, C. R., Schlyter, F., and Jirošová, A. (2023). Field effects of oxygenated monoterpenes and estragole combined with pheromone on attraction of Ips typographus and its natural enemies. Front. For. Glob. Chang. 6:1292581. doi: 10.3389/ffgc.2023.1292581
Mustaparta, H., Angst, M. E., and Lanier, G. N. (1977). Responses of single receptor cells in the pine engraver beetle, Ips pini (SAY) (Coleoptera: Scolytidae) to its aggregation pheromone, ipsdienol, and the aggregation inhibitor, ipsenol. J. Comp. Physiol. A 121, 343–347. doi: 10.1007/BF00613013
Mustaparta, H., Angst, M. E., and Lanier, G. N. (1979). Specialization of olfactory cells to insect- and host-produced volatiles in the bark beetle Ips pini (say). J. Chem. Ecol. 5, 109–123. doi: 10.1007/BF00987692
Mustaparta, H., Angst, M. E., and Lanier, G. N. (1980). Receptor discrimination of enantiomers of the aggregation pheromone ipsdienol, in two species of Ips. J. Chem. Ecol. 6, 689–701. doi: 10.1007/BF00987679
Netherer, S., Lehmanski, L., Bachlehner, A., Rosner, S., Savi, T., Schmidt, A., et al. (2024). Drought increases Norway spruce susceptibility to the Eurasian spruce bark beetle and its associated fungi. New Phytol. 242, 1000–1017. doi: 10.1111/nph.19635
Olsson, S. B., and Hansson, B. S. (2013). “Electroantennogram and single sensillum recording in insect antennae” in Neuroethology of insect olfaction. eds. B. S. Hansson and T. A. Christensen (Cham: Springer), 157–177.
Papek, E., Ritzer, E., Biedermann, P. H., Cognato, A. I., Baier, P., Hoch, G., et al. (2024). The pine bark beetle Ips acuminatus: an ecological perspective on life-history traits promoting outbreaks. J. Pest. Sci. 97, 1093–1122. doi: 10.1007/s10340-024-01765-2
Payne, T. L., Moeck, H. A., Willson, C. D., Coulson, R. N., and Humphreys, W. J. (1973). Bark beetle olfaction—II. Antennal morphology of sixteen species of Scolytidae (Coleoptera). Int. J. Insect Morphol. Embryol. 2, 177–192. doi: 10.1016/0020-7322(73)90027-5
Pettersson, E. M. (2000). Vital volatiles: Host location in parasitic wasps attacking bark beetles. [Ph.D. dissertation]. [Göteborg: Chemical ecology, botanical institute, Univ.]. Available online at: http://hdl.handle.net/2077/13633 (Accessed January, 14,2025).
Pfeffer, A. (1955). Fauna ČSR. Svazek 6: Kůrovci-Scolytoidea. Praha: Brouci-Coleoptera. Nakladatelství Československé Akademie Věd.
Postner, M. (1974). “Ips cembrae” in Die Forstschädlinge Europas II. ed. B. Käfer (Hamburg: Paul Parey), 458–459.
Raffa, K. F., Andersson, M. N., and Schlyter, F. (2016). Host selection by bark beetles: playing the odds in a high-stakes game. Adv. Insect Physiol. 50, 1–74. doi: 10.1016/bs.aiip.2016.02.001
Redfern, D. B., Stoakley, J. T., Steele, H., and Minter, D. W. (1987). Dieback and death of larch caused by Ceratocystis laricicola sp. nov. following attack by Ips cembrae. Plant Pathol. 36, 467–480. doi: 10.1111/j.1365-3059.1987.tb02264.x
Renwick, J. A. A., and Dickens, J. C. (1979). Control of pheromone production in the bark beetle, Ips cembrae. Physiol. Entomol. 4, 377–381. doi: 10.1111/j.1365-3032.1979.tb00630.x
Roberts, R. E., Biswas, T., Yuvaraj, J. K., Grosse-Wilde, E., Powell, D., Hansson, B. S., et al. (2022). Odorant receptor orthologues in conifer-feeding beetles display conserved responses to ecologically relevant odors. Mol. Ecol. 31, 3693–3707. doi: 10.1111/mec.16494
Roberts, R. E., Yuvaraj, J. K., and Andersson, M. N. (2021). Codon optimization of insect odorant receptor genes may increase their stable expression for functional characterization in HEK293 cells. Front. Cell. Neurosci. 15:744401. doi: 10.3389/fncel.2021.744401
Schiebe, C., Blaženec, M., Jakuš, R., Unelius, C. R., and Schlyter, F. (2011). Semiochemical diversity diverts bark beetle attacks from Norway spruce edges. J. Appl. Entomol. 135, 726–737. doi: 10.1111/j.1439-0418.2011.01624.x
Schiebe, C., Hammerbacher, A., Birgersson, G., Witzell, J., Brodelius, P. E., Hansson, B. S., et al. (2012). Inducibility of chemical defenses in Norway spruce bark is correlated with unsuccessful mass attacks by the spruce bark beetle. Oecologia 170, 183–198. doi: 10.1007/s00442-012-2298-8
Schiebe, C., Unelius, C. R., Ganji, S., Binyameen, M., Birgersson, G., and Schlyter, F. (2019). Styrene, (+)-trans-(1R,4S,5S)-4-Thujanol and oxygenated monoterpenes related to host stress elicit strong electrophysiological responses in the bark beetle Ips typographus. J. Chem. Ecol. 45, 474–489. doi: 10.1007/s10886-019-01070-8
Schlyter, F., Birgersson, G., and Leufvén, A. (1989). Inhibition of attraction to aggregation pheromone by verbenone and ipsenol. J. Chem. Ecol. 15, 2263–2277. doi: 10.1007/BF01014114
Schlyter, F., Zhang, Q.-H., Anderson, P., Byers, J. A., Wadhams, L. J., Löfqvist, J., et al. (2000). Electrophysiological and behavioural responses of Tomicus piniperda and Tomicus minor (Coleoptera: Scolytidae) to non-host leaf and bark volatiles. Can. Entomol. 132, 965–981. doi: 10.4039/Ent132965-6
Senf, C., Dirk, P., Zhiqiang, Y., Sebald, J., Knorn, J., Neumann, M., et al. (2018). Temperate forests over the last three decades. Nat. Commun. 9:4978. doi: 10.1038/s41467-018-07539-6
Shewale, M. K., Nebesářová, J., Grosse-Wilde, E., and Kalinová, B. (2023). Microscopic morphology and distribution of the antennal sensilla in the double-spined bark beetle, Ips duplicatus (Coleoptera: Curculionidae). Microsc. Res. Tech. 86, 1610–1625. doi: 10.1002/jemt.24397
Silverstein, R. M., and Young, J. C. (1976). Insects generally use multicomponent pheromones. ACS Symp. Ser. 23, 1–13. doi: 10.1021/bk-1976-0023.ch001
Sommerfeld, A., Rammer, W., Heurich, M., Hilmers, T., Müller, J., and Seidl, R. (2021). Do bark beetle outbreaks amplify or dampen future bark beetle disturbances in Central Europe? J. Ecol. 109, 737–749. doi: 10.1111/1365-2745.13502
Stoakley, J. T., Bakke, A., Renwick, J. A. A., and Vité, J. P. (1978). The aggregation pheromone system of the larch bark beetle, Ips cembrae Heer. Z. Angew. Entomol. 86, 174–177. doi: 10.1111/j.1439-0418.1978.tb01925.x
Thabeet, A., Vennetier, M., Gadbin-Henry, C., Denelle, N., Roux, M., Caraglio, Y., et al. (2009). Response of Pinus sylvestris L. to recent climatic events in the French Mediterranean region. Trees 23, 843–853. doi: 10.1007/s00468-009-0326-z
Tømmerås, B. Å. (1985). Specialization of the olfactory receptor cells in the bark beetle Ips typographus and its predator Thanasimus formicarius to bark beetle pheromones and host tree volatiles. J. Comp. Physiol. A 157, 335–341. doi: 10.1007/BF00618123
Unelius, C. R., Schiebe, C., Bohman, B., Andersson, M. N., and Schlyter, F. (2014). Non-host volatile blend optimization for forest protection against the European spruce bark beetle, Ips typographus. PLoS One 9:85381. doi: 10.1371/journal.pone.0085381
Villari, C., Battisti, A., Chakraborty, S., Michelozzi, M., Bonello, P., and Faccoli, M. (2012). Nutritional and pathogenic fungi associated with the pine engraver beetle trigger comparable defenses in scots pine. Tree Physiol. 32, 867–879. doi: 10.1093/treephys/tps056
Wajs, A., Pranovich, A., Reunanen, M., Willför, S., and Holmbom, B. (2007). Headspace-SPME analysis of the sapwood and heartwood of Picea abies, Pinus sylvestris, and Larix decidua. J. Essent. Oil Res. 19, 125–133. doi: 10.1080/10412905.2007.9699244
Wermelinger, B. (2004). Ecology and management of the spruce bark beetle Ips typographus: a review of recent research. For. Ecol. Manag. 202, 67–82. doi: 10.1016/j.foreco.2004.07.018
Wermelinger, B., Rigling, A., Schneider Mathis, D., and Dobbertin, M. (2008). Assessing the role of bark- and wood-boring insects in the decline of scots pine (Pinus sylvestris) in the Swiss Rhone valley. Ecol. Entomol. 33, 239–249. doi: 10.1111/j.1365-2311.2007.00960.x
Wood, D. L. (1982). The role of pheromones, kairomones, and allomones in the host selection and colonization behavior of bark beetles. Annu. Rev. Entomol. 27, 411–446. doi: 10.1146/annurev.en.27.010182.002211
Wood, S. L., and Bright, D. E. Jr. (1992). Hosts of Scolytidae and Platypodidae. Great Basin Nat. Mem. 13:12.
Yuvaraj, J. K., Kandasamy, D., Roberts, R. E., Hansson, B. S., Gershenzon, J., and Andersson, M. N. (2024). Eurasian spruce bark beetle detects lanierone using a highly expressed specialist odorant receptor, present in several functional sensillum types. BMC Biol. 22:266. doi: 10.1186/s12915-024-02066-x
Yuvaraj, J. K., Roberts, R. E., Sonntag, Y., Hou, X. Q., Grosse-Wilde, E., Machara, A., et al. (2021). Putative ligand binding sites of two functionally characterized bark beetle odorant receptors. BMC Biol. 19, 16–21. doi: 10.1186/s12915-020-00946-6
Zhang, Q. H., and Niemeyer, H. (1992). Morphological characteristics for sexing living adults of Ips cembrae (Heer) (Col., Scolytidae). J. Appl. Entomol. 114, 403–409. doi: 10.1111/j.1439-0418.1992.tb01144.x
Zhang, Q. H., and Schlyter, F. (2004). Olfactory recognition and behavioral avoidance of angiosperm non-host volatiles by conifer-inhabiting bark beetles. Agric. For. Entomol. 6, 1–20. doi: 10.1111/j.1461-9555.2004.00202.x
Zhang, Q., Schlyter, F., and Anderson, P. (1999). Green leaf volatiles interrupt pheromone response of spruce bark beetle Ips typographus. J. Chem. Ecol. 25, 2847–2861. doi: 10.1023/A:1020816011131
Zhang, Q. H., Tolasch, T., Schlyter, F., and Francke, W. (2002). Enantiospecific antennal response of bark beetles to spiroacetal (E)-conophthorin. J. Chem. Ecol. 28, 1839–1852. doi: 10.1023/A:1020569303433
Keywords: electrophysiology, olfactory sensory neurons, bark beetles, aggregation pheromones, monoterpenes, semiochemicals
Citation: Shewale MK, Bláha J, Synek J, Schebeck M, Andersson MN, Kandasamy D and Jirošová A (2025) Comparative analysis of olfactory sensory neurons in two Ips species reveals conserved and species-specific olfactory adaptations. Front. For. Glob. Change. 8:1588866. doi: 10.3389/ffgc.2025.1588866
Edited by:
Milica Zlatkovic, University of Novi Sad, SerbiaReviewed by:
Paul Greer, University of Massachusetts Medical School, United StatesPatrick Christian Pageat, Institut de Recherche en Sémiochimie et Ethologie Appliquée (IRSEA), France
Jiaxing Fang, Chinese Academy of Forestry, China
Copyright © 2025 Shewale, Bláha, Synek, Schebeck, Andersson, Kandasamy and Jirošová. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Anna Jirošová, amlyb3NvdmFhQGZsZC5jenUuY3o=
‡ORCID: Mayuri Kashinath Shewale, orcid.org/0000-0001-8021-695X
Jaromír Blaha, orcid.org/0000-0002-5101-8302
Jiří Synek, orcid.org/0000-0003-1360-7680
Martin Schebeck, orcid.org/0000-0001-7376-4878
Martin N. Andersson, orcid.org/0000-0001-9807-8524
Dineshkumar Kandasamy, orcid.org/0000-0002-5450-3231
Anna Jirošová, orcid.org/0000-0002-5054-5821
†These authors share senior authorship