Abstract
Forest management in Central Europe is affected by numerous biotic and abiotic factors, and wildlife damage to forest stands is one of the major ones. While damage to young forest stands is predominantly attributed to wild ungulates, the potential negative impact of the European hare (Lepus europaeus) on tree plantations has been largely overlooked. Therefore, this study aimed to quantify hare damage in comparison with ungulates, determine which developmental stages of trees are most attractive to hares, compare browsing preferences among key commercial species, and assess the effectiveness of commonly used protection measures. We evaluated 209 calamity clearcuts with 75,912 seedlings to determine the proportion and structure of browsing damage. Research revealed a substantial proportional damage caused by the European hare, accounting for 10.12%, while wild ungulates were responsible for 7.11% of browsing damage. The species distribution played a crucial role in the damage rate. Hares caused the greatest browsing damage on silver birch (Betula pendula—37%), while ungulates preferred Scots pine (Pinus sylvestris—29%). Moreover, browsing intensity caused by hares was seasonally affected with a significantly higher damage ratio in autumn (<14%) compared to the spring season (<2%) when clearcuts offer enough alternative fodder opportunities. Common protection measures, such as coating and fencing, proved largely ineffective against hare browsing. Surprisingly, sycamore maple (Acer pseudoplatanus) and European beech (Fagus sylvatica) were more heavily damaged by hares when coated than unprotected seedlings. The results showed that the European hare plays a crucial role in clear-cut reforestation, highlighting the need for enhanced focus on protective measures, including population management and the use of specialized fencing made of dense wire mesh compared to the usual protection against wild ungulates.
Introduction
European forests are currently facing a wide range of large and small-scale disturbances, including biotic and abiotic factors, which affect forest biodiversity, structure, stability, and species composition (Nagel et al., 2017; Brázdil et al., 2018). The negative factors affecting forest ecosystems are being intensified by ongoing climate change. They impact different tree species in different ways and generally destabilize forest stands (Allen et al., 2010; Lindner et al., 2014; Hlásny et al., 2017; Vacek et al., 2022, 2023). Forest management must respond to these changes by appropriate management measures, primarily by changing the tree species structure (Vacek et al., 2023). The traditional conifer monoculture plantations typical of Central Europe are now being replaced by broadleaved tree species predominately planted in mixtures that show greater resistance to climatic extremes (Seliger et al., 2023). Moreover, these mixed-species stands provide other benefits such as water regime improvement, amelioration effects, and increased and stable radial increment (González de Andrés et al., 2017; Pretzsch et al., 2020; Steckel et al., 2020; Vacek et al., 2021).
On the one hand, these newly established resilient forest ecosystems consisting of mixtures of broadleaved and coniferous species are much more resilient to ongoing climate change (Zamora-Pereira et al., 2021), on the other hand, may be endangered by other factors. In this context, the damage caused by wildlife is one of the most serious aspects affecting young forest stands with increasing intensity throughout Europe (Hardalau et al., 2024). In this case, wild ungulates such as red deer or roe deer, and also introduced ones (sika or fallow deer), are highlighted as species with frequent negative impacts on young forest stands (Gerhardt et al., 2013). This is especially true for newly afforested extended clearcuts which arose over the last few years in Central Europe due to the unprecedented bark beetle calamity (Schwegmann et al., 2023). Browsing and bark damage by wild ungulates can locally represent a considerable constraint on the successful regeneration of forest stands. Nevertheless, a high proportion of afforested clearcuts within forest ecosystems may also provide suitable biotopes for the European hare (Lepus europaeus Pallas)—another wildlife species that was heretofore only tenuously connected to the damage to young forest stands (Jansson and Pehrson, 2007).
From a historical perspective, the European hare abundance was at its highest during the 1970s. In general, the habitat type that is primarily preferred by hares is rich fragmented farmland with high biodiversity (Marboutin and Aebischer, 1996; Smith et al., 2005). In the suitable landscape consisting of a mixed, arable agriculture environment, the hare densities can reach up to 134 ind./100 ha (Krivopalova et al., 2024), or even more. However, hares can also inhabit forest habitats where the population densities are lower compared to open fields (Gryz and Krauze-Gryz, 2022). Moreover, forest habitats affected by disturbances may offer more open areas, and the lack of continuous forest cover over larger expanses could create potentially suitable conditions for hares.
From the point of view of their diet, hares prefer weeds or grasses, depending on what the habitat offers (Schai-Braun et al., 2015). If the European hare does not have access to sufficiently energy-rich food, it increases its overall food intake (Hackländer et al., 2002). The proportion of grasses in their diet decreases between summer and autumn, when forbs begin to dominate. Another shift occurs in winter with the onset of snow cover. During this period, the proportion of shrubs in the diet increases significantly (43% compared to a maximum of 3% in snow-free months) (Green et al., 2013). According to Hjältén et al. (2004), deciduous trees and shrubs are significantly more palatable to hares than conifers. Their diet predominantly includes goat willow (Salix caprea L.), common aspen (Populus tremula L.), and European blueberry (Vaccinium myrtillus L.). The ability to utilize woody plant shoots as a nutrient source enables hares to survive during periods when higher-quality food is scarce (Hulbert et al., 2001). When energy-rich food sources are limited, hares also include less nutritious species in their diet, and overall diet diversity decreases as food availability declines (Rödel et al., 2004).
Based on their feeding behavior, it is evident that the European hare may represent a significant threat to the regeneration and survival of young forest plantations. While the negative impact of wild ungulate populations on forest regeneration has been extensively studied and is frequently mentioned (Ramirez et al., 2018; Champagne et al., 2021; Unkule et al., 2022; Bernard et al., 2024; Brabec et al., 2024) the role of the hare has often been overlooked, or the damage it causes is not distinguished from that caused by other wildlife species.
Therefore, we tested the following research hypotheses:
-
Browsing damage by the European hare represents a higher proportion of total seedling damage compared to ungulates.
-
The susceptibility of seedlings to hare browsing decreases with advancing developmental stage, as hares preferentially target younger individuals with thinner shoots.
-
Broadleaved tree species are more heavily browsed by hares than coniferous species.
-
Common protective measures (repellent coatings and conventional fencing) are ineffective against hare browsing.
Materials and methods
Area of interest
The research was conducted in the Drahanská vrchovina (Drahany Highlands) in southwestern Czech Republic (Figure 1; N 49°22.81753′, E 16°48.70538′ – center of the study area). Mid altitudes (500–700 m a. s. l.) are characterized by extensive forest cover (53%). Climatically, the area belongs to the warm-summer humid continental climate (Dfb) according to the Köppen-Geiger classification (Peel et al., 2007). The average annual temperature ranges from 6.2 to 7.9 °C, and the annual precipitation ranges from 600 to 800 mm. The average number of days with snow cover is 70 (Český hydrometeorologický ústav—Czech Hydrometeorological Institute). The forest typology indicates the prevalence of the nutrient-rich series (Series mesotrophicum) in this area, particularly the nutrient-medium category (Categoria oligo-mesotrophica). Nutrient-rich series are well-developed, fresh soils with fast decomposition, dominated by mesophytes, and characterized by high forest productivity and brush hazard. The second most represented is the acidic series (Series acidophilum), specifically the acidic category (Categoria acidophila). This series develops on mineral-poor, well-aerated soils with poor decomposition, low water-holding capacity due to rapid drainage, and low sorption and cation exchange capacity (Viewegh et al., 2003). The vegetation cover consists mainly of monospecific Norway spruce [Picea abies (L.) Karst.] forests and mixed stands of spruce, European beech (Fagus sylvatica L.), sycamore maple (Acer pseudoplatanus L.) and Scots pine (Pinus sylvestris L.), which have often replaced the original beech-dominated forests, mostly by artificial planting.
Figure 1
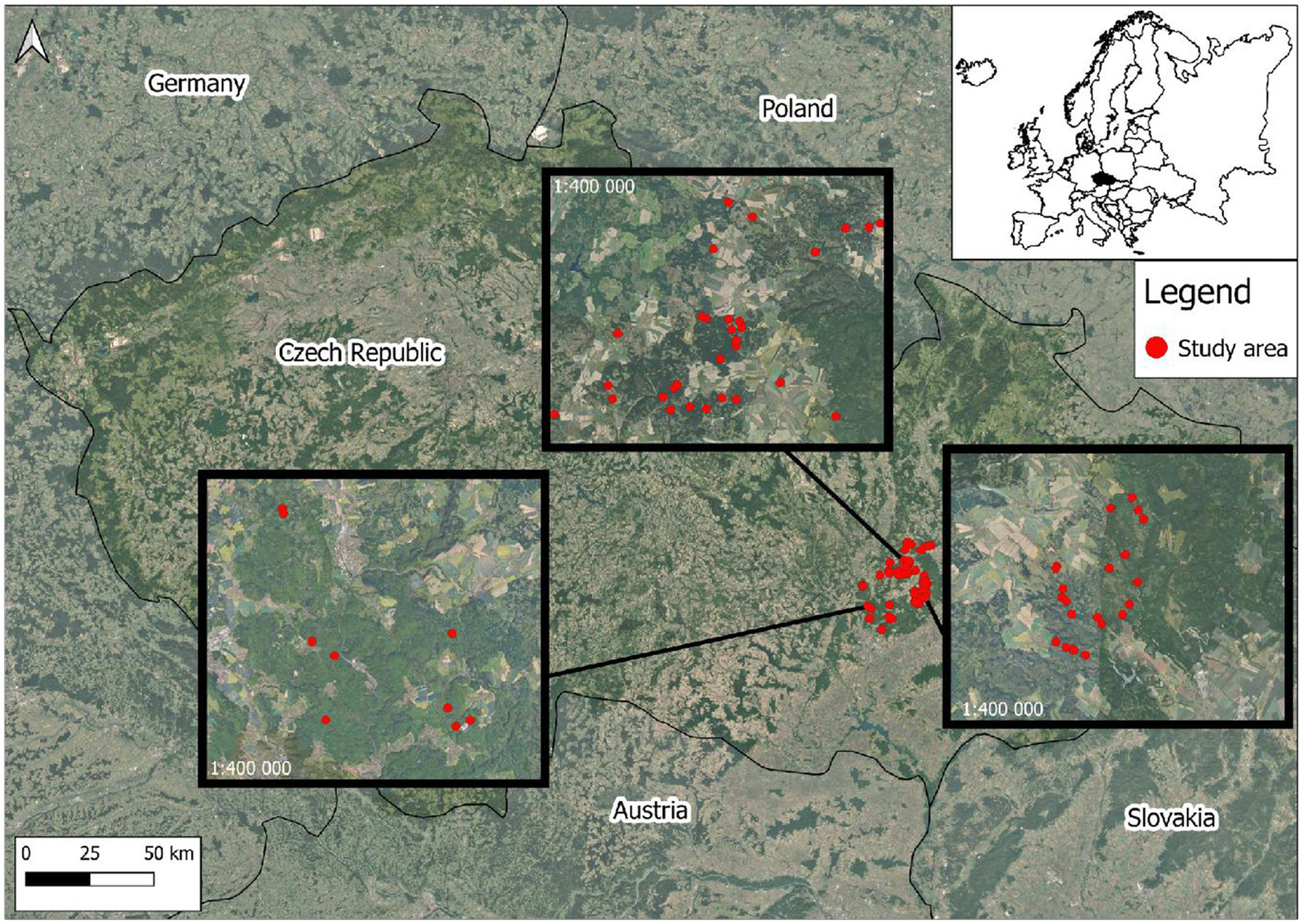
The area of interest—Drahanská vrchovina—with the main locations of the monitored plots (red dots).
The total study area is 118,300 ha and, in terms of game management, is divided into 121 hunting grounds. The area has a high population density of roe deer (Capreolus capreolus L.). In hunting grounds where research plots are established (23 hunting grounds covering 34,800 ha), an average of 19.91 roe deer per 1,000 ha are hunted. The European rabbit (Oryctolagus cuniculus L.) is not found in the area. The European fallow deer (Dama dama L.) and red deer (Cervus elaphus L.) are rarely seen. The average fallow deer and red deer harvests are 1.52 individuals/1000 ha and 1.83 individuals/1000 ha, respectively. Hares are not harvested in these hunting grounds due to low hunting interest in forest areas where the ungulates are the main hunted species. The overview of game populations deliberately includes data on the harvest for the species concerned, which is considered the most reliable in hunting statistics (Kahlert et al., 2015). With all limitations acknowledged (e. g. Hunting effort or reporting of the data), these hunting bags primarily indicate the composition/ratio of the ungulate harvest and the dominant occurrence of roe deer in the study area. The location was purposely chosen with regard to the destruction of parent stands by the bark beetle disaster, and the subsequent availability of plantings up to three years old, and the presence of game, allowing for monitoring the impact of browsing on the survival and development of seedlings.
Data collection
For monitoring damage to seedlings caused by wildlife, forest plantations up to three years old were randomly selected in 2020 and 2021. Monitoring took place from April 1 to April 30 in both years which reflects the browsing after whole non-vegetation season. The browsing was not evaluated during the vegetation season due to fast regeneration of seedlings and impossible estimation of the cause of damage. A total of 209 plots with an area ranging from 0.1 ha to 10 ha (average 1.33 ha, SD 1.46 ha) were evaluated over the two years. The areas were selected after a systematic survey of established young stands (plantations). The size of each clearing was determined using GIS tools (ArcGIS 10.6). GPS coordinates were entered for each plot. The basic characteristics of the plantation were first recorded for each assessed forested area: tree species, planting age, and method of protection against game, if applicable. Subsequently, we made an inventory of growing seedlings and, for each hectare, even an incomplete one, we evaluated 250 seedlings. The evaluation was carried out on line transects systematically distributed throughout the clearing to spread observations across the stand and minimize potential local clustering effects, with two transect lines located in the central part of the clearing at a distance of ten meters from each other, with one line on either edge. Additional transects were incorporated into the remaining portion of the plot to ensure uniform coverage of the entire plantation. Twenty-five seedlings were evaluated in each transect.
The evaluation also included an analysis on the impact of protective measures against terminal browsing. Of the total number of monitored plots, 19 were fenced with 160 cm high, 2.0/2.8 mm forest knot mesh with 15 horizontal wires spaced 15 cm apart and denser mesh at the bottom to keep away hares (dimensions and number of meshes from the bottom: four meshes 5 × 15 cm, five meshes 10 × 15 cm, two meshes 15 × 15 cm, and three meshes 20 × 15 cm). The mesh was attached to wooden posts. Another 129 areas were treated with repellent coating—with Aversol (TORA, Czech Republic), Cervacol (AgroProtec, Czech Republic), and Morsuvin (Nera Agro, Czech Republic) commonly used in the area. The remaining 61 plots were untreated. All individuals observed were included in the total number evaluated, regardless of damage. In cases where terminal browsing damage was found, the height from the above-ground part to the terminal browsing spot (with an accuracy of ±0.1 cm), and the diameter of the root collar were measured using a digital caliper (±0.03 mm) on 15 randomly selected damaged seedlings. For comparison, the same parameters were also recorded for 30 randomly selected undamaged seedlings on the same plot. All data were entered in field forms and then digitized. The records were continuously checked for completeness and consistency. The resulting data were used to evaluate the extent of damage to seedlings and to statistically assess the influence of habitat factors.
Evaluation of browsing damage
The evaluation of damage to seedlings was based on differentiating two main groups according to who browsed the terminals—specifically, hares and cloven-hoofed game, predominantly roe deer in our case (according to the game hunting figures). The species responsible were determined by their dentition differences, which are reflected in the nature of the damage and the resulting wound on the seedling terminal (Figure 2). The European hare has sharp incisors in both the upper and lower jaws. The dental formula is I 2/1, C 0/0, P 3/2, M 3/3 (I = incisors, C = canines, P = premolars, M = molars; upper/lower jaw). As a result, the shoots, or in our case, primarily the terminals, are severed to create a clean, smooth cut on the stem, typically at a 45-degree angle. This type of browsing is usually found up to a height of 50 cm, which corresponds to the maximum reach of a hare standing upright on its hind legs, without snow cover. In contrast, cloven-hoofed animals do not have incisors in their upper jaw. The dental formula is I 0/3, C 0/1, P 3/3, M 3/3. When feeding, they tear off plant material between their lower incisors and hard palate, resulting in irregular, frayed damage to shoots (Pepper and Hedge, 1998). The edges are usually ragged, often with visible tissue disintegration (Vowles et al., 2016). The height of the damage commonly ranges between 30 and 150 cm because only artificial regeneration was evaluated. Natural regeneration was not included in the damage evaluation because the post-disturbance sites had been mostly artificially regenerated. During the evaluation, both the height of the damage and the nature of the cut were recorded at all plots, utilizing a magnifying glass if necessary. Based on these characteristics, it was possible to reliably identify the browser.
Figure 2
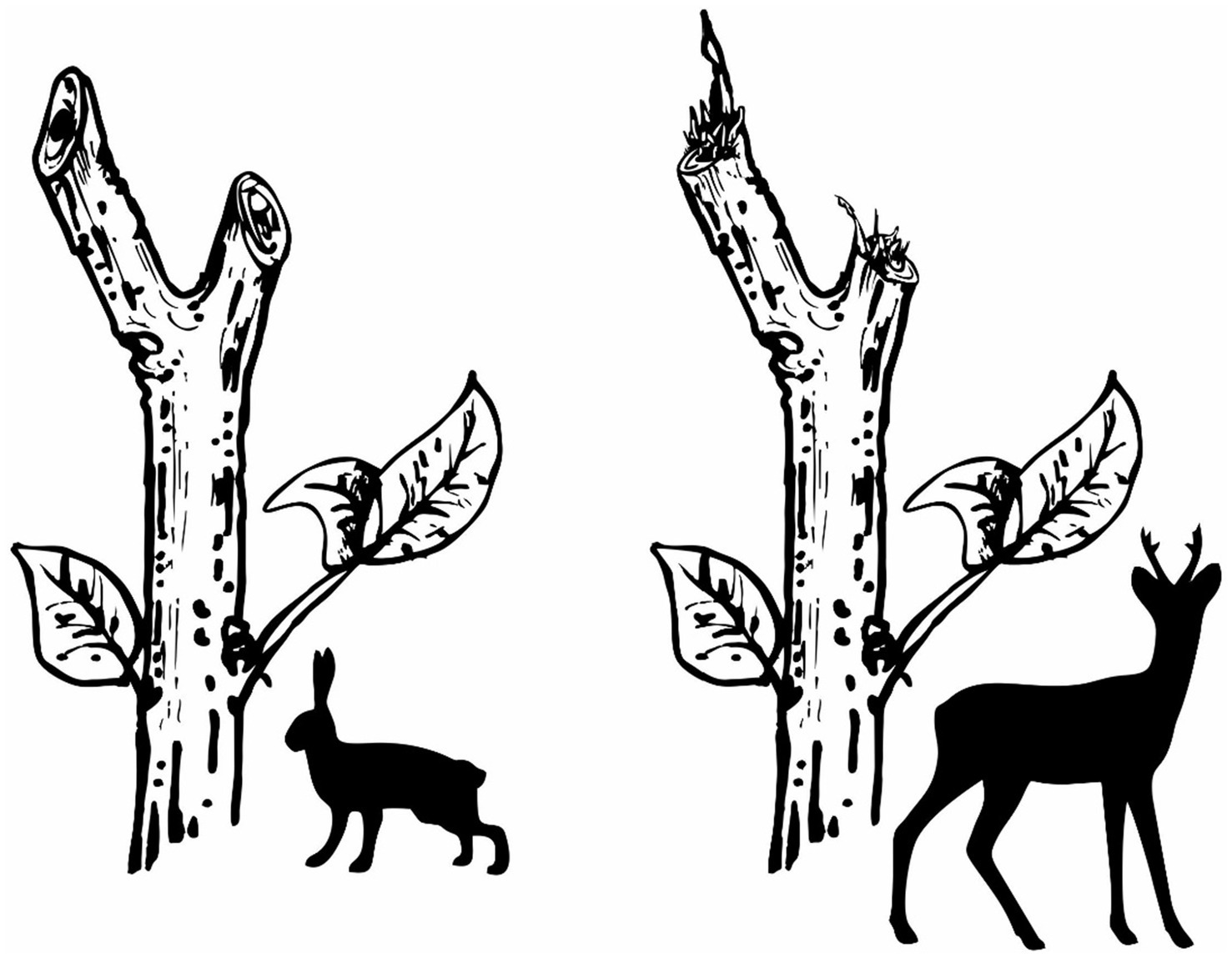
Browsing damage caused by the hare (smooth cut) and cloven-hoofed game (ragged, irregular).
Statistical assessment
As a first analysis, we have provided basic statistics about overall tree species composition and mean damage across all plots.
For testing for differences in number of damaged trees between hares and ungulates (marked “species” in model description), we analyzed the proportion of trees damaged using a binomial generalized linear mixed model (GLMM) with a logit link function. Species (hares/ungulates) was included as a fixed effect, and plot identity was included as a random intercept to account for non-independence of observations within plots. Each plot is considered observational unit in this analysis.
For evaluating the dependence of damage intensity on the plot area, linear regression was employed. However, due to a very low R2 value and overall bad fit, a spline curve is also presented to illustrate the character of the data. This analysis is performed separately for treated and untreated variants of seedlings after the first and second growing season.
For species with the most observations (beech and spruce), a relative count of damaged individuals based on the season of plantation is depicted. Separate GLMMs with binomial distribution were fitted for damages/ungulates. Model marginal means are depicted in respective plot along with its 95% confidence interval. Models in form (number of damaged trees, number of not damaged trees) ~ tree species: planting season (spring/autumn) + (1 | location) were used for subsequent comparisons separately for each tree species. Comparisons of marginal means were done using R package “emmeans” (Lenth, 2023) as in previous case. Only data after one vegetation season were used for construction of mentioned models to avoid counting damage over more years.
Regarding the choice of seedlings by their diameter (by hare only, not applicable for damage by ungulates), the relationship between seedling diameter (in position of damage caused by hare) and seedling height (the rest above damage, respectively) was evaluated by linear regression with random intercept, where location was considered random effect. For construction of GLMM, R packages “lme4” (Bates et al., 2015) and “lmerTest” (Kuznetsova et al., 2016) were used.
Tree species preference (in terms of damage) for both hare and ungulates was further examined. For the most prevalent species in this study (beech, spruce, pine, oak, and maple), separate (for hares/ungulates) generalized linear mixed effect models with binomial distribution were constructed for in following form (number of damaged trees, number of not damaged trees) ~ tree species + (1 | location). A statistical comparison of model predicted marginal means of damaged individuals’ ratio was performed using R package “emmeans.” To avoid counting damage from previous years, only data for trees after 1 vegetation season were used.
For testing for differences in damaged trees ratio between coated/non-coated trees by repellent, separate GLMMs for hares/ungulates were created. Models in form (number of damaged trees, number of not damaged trees) ~ tree species: coating (yes/no) + (1 | location) were used for subsequent comparisons separately for each tree species. Comparisons of marginal means were done using R package “emmeans” as in previous case. Data for trees after 1 vegetation season only were used for identical reasons as in cases above.
For comparison of the relative number of damaged seedlings inside and outside fenced enclosures, GLMM with binomial distribution in following form: (number of damaged individuals, number of not damaged individuals) ~ tree species: fencing (yes/no) + (1 | location), similarly as in previous cases. Also, comparison of damaged individuals ratio inside/outside fenced enclosure was performed using R package “emmeans” as in previous cases. The results are presented in form of bar plot.
All statistical computations were made in R software (R Core Team, 2021) and plots were created in its package “ggplot2” (Wilkinson, 2011). Mixed effect models were computed using R package “lme4.” An alpha level of 0.05 was used in all statistical analyses.
Results
Extent of damage caused by hares compared to ungulates
The impact of wildlife on the artificial regeneration of forest tree species was evaluated in 75,912 seedlings. The highest proportion of browsing damage was found in European beech (Fagus sylvatica L.—50.3% of all seedlings assessed), followed by Norway spruce (Picea abies [L.] Karst.—18.6%), pedunculate oak (Quercus robur L.—17.3%), Scots pine (Pinus sylvestris L.—4.84%), Norway maple (Acer platanoides L.—3.05%), sycamore maple (Acer pseudoplatanus L.—1.78%), small-leaved lime (Tilia cordata Mill.—1.55%), and European hornbeam (Carpinus betulus L.—1.36%), with other tree species having minimal representation (<1%). A total of 7,685 seedlings (10.12%, SD 17%) were damaged by hares, while ungulates damaged 5,401 seedlings (7.11%, SD 15%).
The results of comparison of damage intensity between hares and ungulates showed significant differences (p < 0.001) in damage odds ratio (1.57 for hares), meaning that the tree is 1.57 likely to be damaged by hare than by ungulate according to constructed GLMM model. Overall numbers of damaged trees for hares/ungulates are presented above.
The relationship between the size of the research plots (in ha) and the proportion of damaged seedlings (in %) from four scenarios according to treatment (coating versus no treatment) and monitoring period (one versus two growing seasons) is shown in Figure 3. In none of the cases was a statistically significant dependence confirmed (all models show very low R2 values and insignificant regression coefficients – not treated/ after 1 veg. Season: R2 = 0.04, p = 0.21, treated/ after 1 veg. Season: R2 = 0.04, p = 0.09, not treated/ after 2 veg. Seasons: R2 = 0.01, p = 0.56, treated/ after 2 veg. Seasons: R2 = 0.001, p = 0.78). The data shows only a weak, non-linear indication of higher damage on smaller plots (primarily those around 1 ha), particularly on the untreated ones after the first growing season. However, this “peak” is significantly influenced by several extreme values and cannot be interpreted as a robust trend. The variance of values is considerable, especially for smaller plots, where both very low and extremely high damage can occur. Contrastingly, for larger plots, the damage is mostly low, but exhibits no clear trend. For the treated plots, damage is lower overall, but the relationship with the plot size is also inconsistent. A similar development was also observed in the evaluated damage caused by cervids. No correlation was confirmed between clearing size and browsing level; considerable variation exists across seasons and treatment regimes.
Figure 3
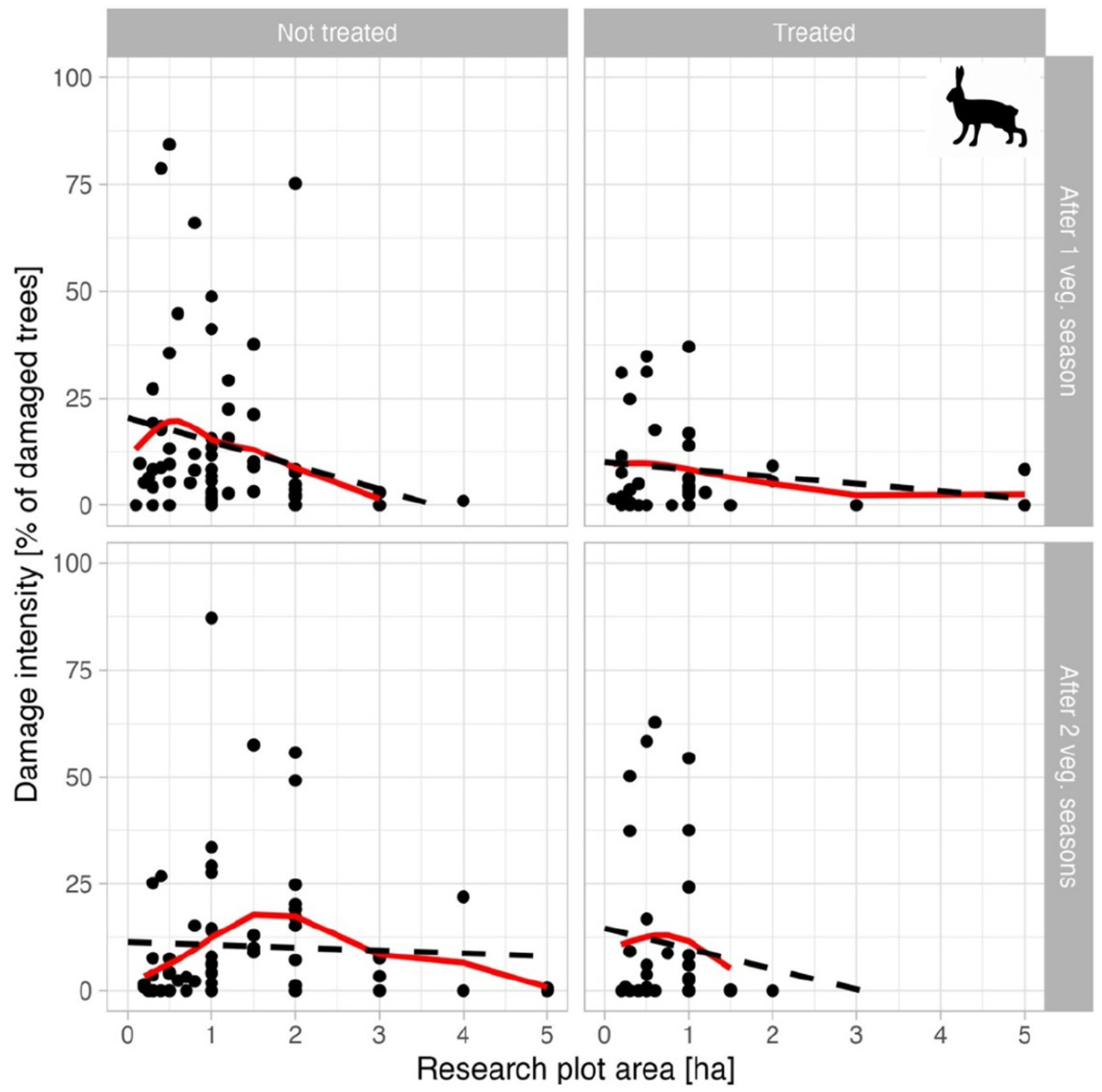
The effect of plot size and treatment on the intensity of hare browsing. To maintain the proportions of the individual graphs, the figure does not show three outliers (not shown in plot, but included in analysis) for the 10-ha plot, namely 0.8% in the upper left graph and 0.4 and 20.4% in the lower left graph. A red line in each subplot depicts spline line (smoothing parameter of 0.7 was used), black dashed line depict linear regression line.
The model predicted (marginal means) relative proportion of damaged seedlings of the most common commercial tree species, i.e., beech and spruce, depending on the season they were planted (spring versus autumn) and game species (hare versus cloven-hoofed game) is shown in Figure 4. In all combinations, autumn plantings were more susceptible to damage than spring ones. For hare-induced damages, marginally insignificant results are observed for both tree species (p = 0.07 for beech, 0.08 for spruce). In autumn, marginal means reached 9.5% for beech and 13.9% for spruce, in spring, marginal values reached values around 1% in both cases. For cloven-hoofed game-induced damage, no significant differences were found – for spring, marginal means reached around 2% in both cases, for autumn, marginal means reached 5.2% for beech and 7.8% for spruce.
Figure 4
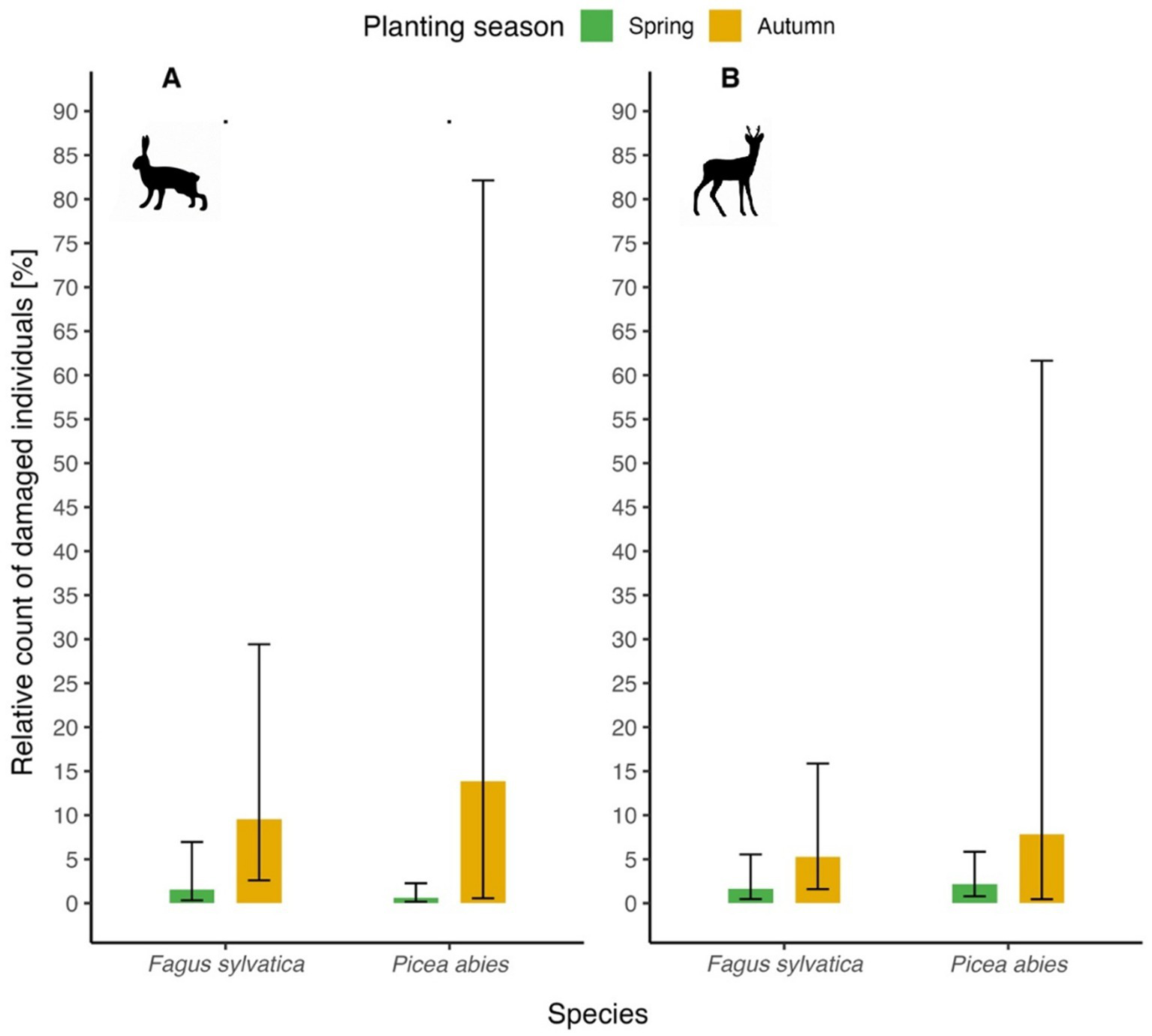
Relative count of damaged beech and spruce seedlings by planting season (spring versus autumn). (A) hare-induced damage; (B) cloven-hoofed game-induced damage.
Tree species attractiveness by seedling size
Typically, hares damage seedlings of low to mid-height, while in taller individuals, they select parts of a specific diameter to browse (Figure 5). Regression analysis with location as a random effect (random intercept) confirmed a marginally statistically significant (p = 0.048) but a very slight relationship between these two variables (y = 2.61–0.005x) with random effect of location varying between −1.54 and 2.83. Marginal R2 reached only value of 0.004, with random effects included (conditional R2 = 0.73), showing that random effect (intercept) of location “described” most of variability. The diameter of the seedling at the site of damage was chiefly between 2 and 3 mm (overall mean 2.43 mm, SD 0.85), with values only rarely approaching 4 mm, even in seedlings taller than 60 cm. The slope coefficient of the regression line (−0.005) shows that for every 10 cm of shoot height, the diameter at the point of browsing decreases by less than 0.1 mm on average (which is practically negligible value). It can therefore be concluded that hares prefer thinner shoots.
Figure 5
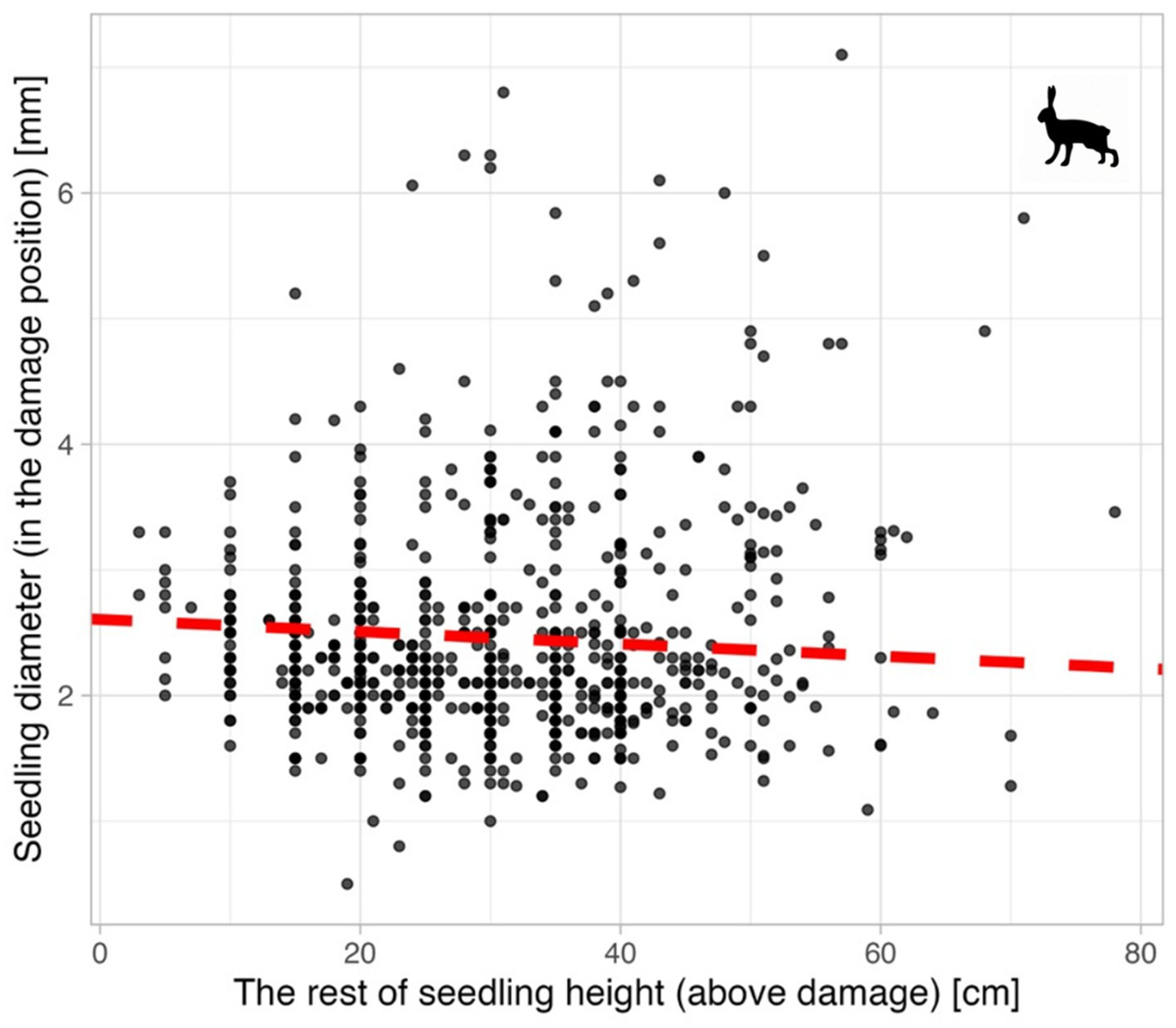
The relationship between the above-ground part and diameter of the seedling at the hare-browsed spot. Linear regression line depict model intercept and fixed effect slope.
Species preference and seedling protection
In terms of species preference, the animals mainly damaged native species such as birch, Norway maple, sycamore maple, and alder. Hares grazed most on the seedlings of deciduous trees, such as birch (overall damage of 37%), sycamore maple (22%), beech (15%), and Norway spruce (11%), while other tree species were damaged by less than 10%. Cloven-hoofed animals primarily grazed on terminal shoots of pine (29%), alder (21%), birch (20%), and sycamore (18%). For damage by hares, used GLMM with binomial distribution does not show any significant differences between model predicted marginal means (which take into account location variability in species distribution using location as a random model effect), which varied between 0.2% of trees damaged for oak and 10.4% of trees damaged for maple (beech – 3.8%, pine – 8.0%, spruce – 0.8%; Figure 6A). In case of damage by wild ungulates, model showed significant difference of pine (marginal mean = 57.65%) compared to other tree species (which values varied between 1.2% for maple and 3.1% for oak, beech – 2.7%, spruce – 2.2%), showing its significant preference (Figure 6B).
Figure 6
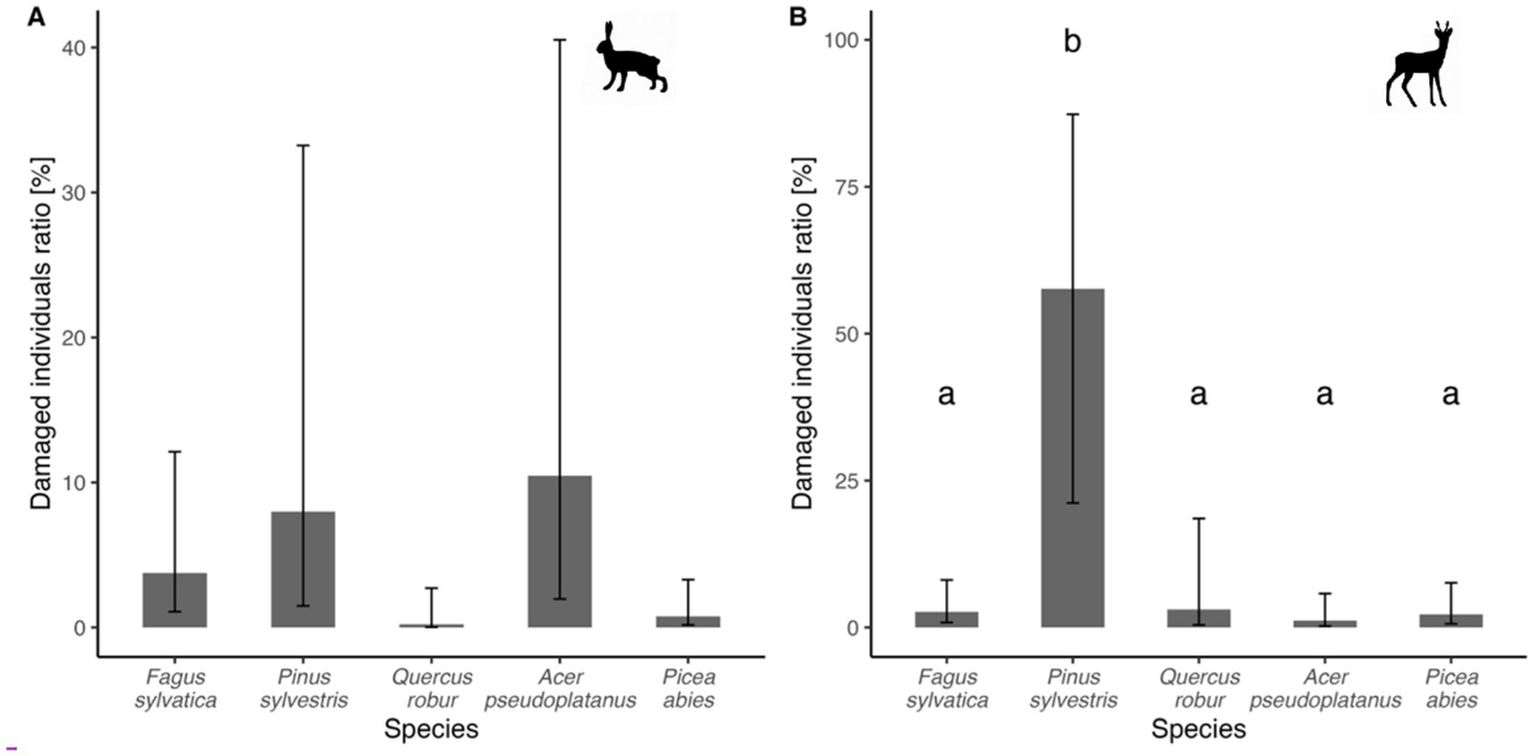
Comparison of marginal means of damaged individuals ratio for selected tree species for damages by hares (A) and ungulates (B). Different indices above subplot B show statistically significant differences (variants with different index above respective bar depict significant difference and vice versa). Error bars depict 95% confidence interval for respective marginal mean.
In the case of wild ungulates, the repellent coating did not prove to be an effective tool for protecting seedlings of almost any tree species (p ˃ 0.05), except for pine (p = 0.04, model marginal mean for protected individuals = 18.9%, for unprotected individuals = 57.8%). In case of maple, marginal insignificance was found (p = 0.09, model marginal mean for protected individuals = 1.2%, unprotected individuals = 28.6%). Other values of model marginal means varied around 2% (Figure 7).
Figure 7
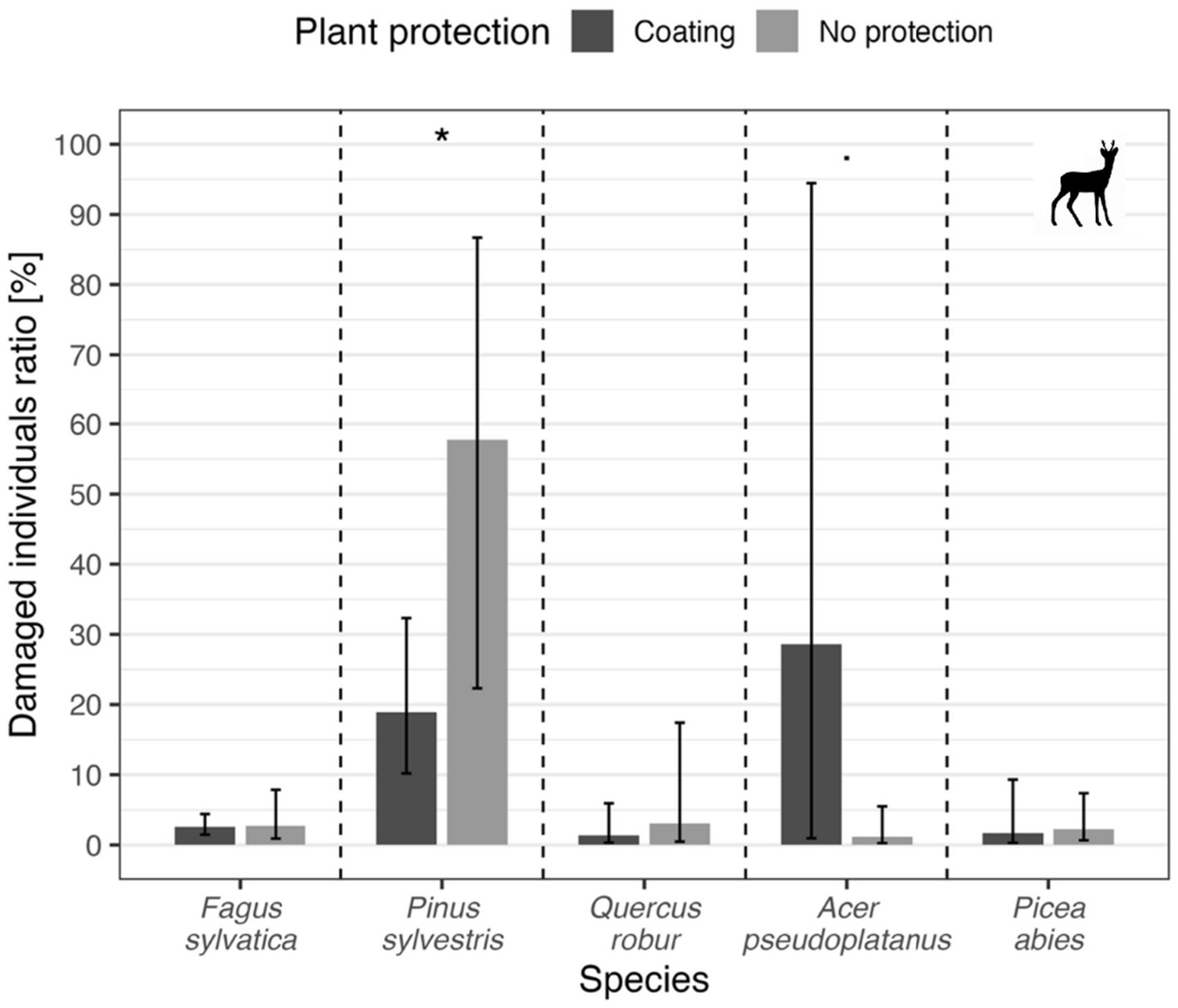
Proportion of cloven-hoofed game-induced damage by tree species and protection. Bars show GLMM predicted marginal mean value for each variant, error bars depict a 95% confidence interval for each ratio. Symbols above each species bar pair depict statistical significance for the comparison of particular species (asterisk stand for p-value between 0.01 and 0.05, dot stands for marginal insignificance, i.e., p-value between 0.05 and 0.1).
Similarly, the coating did not repel hares effectively (p > 0.05) also except for pine (p < 0.001), where significant difference between coated and non-coated trees was found (model marginal mean for protected individuals = 0.3%, unprotected individuals = 7.9%). Specifically, in the case of European hare, the highest proportion of damaged seedlings was recorded in sycamore maple, oddly, in the variant with a protective coating—an overall average of 45% (marginal mean also 45%), while in untreated seedlings, the overall proportion was only 17% (marginal mean = 10%, see Figure 8).
Figure 8
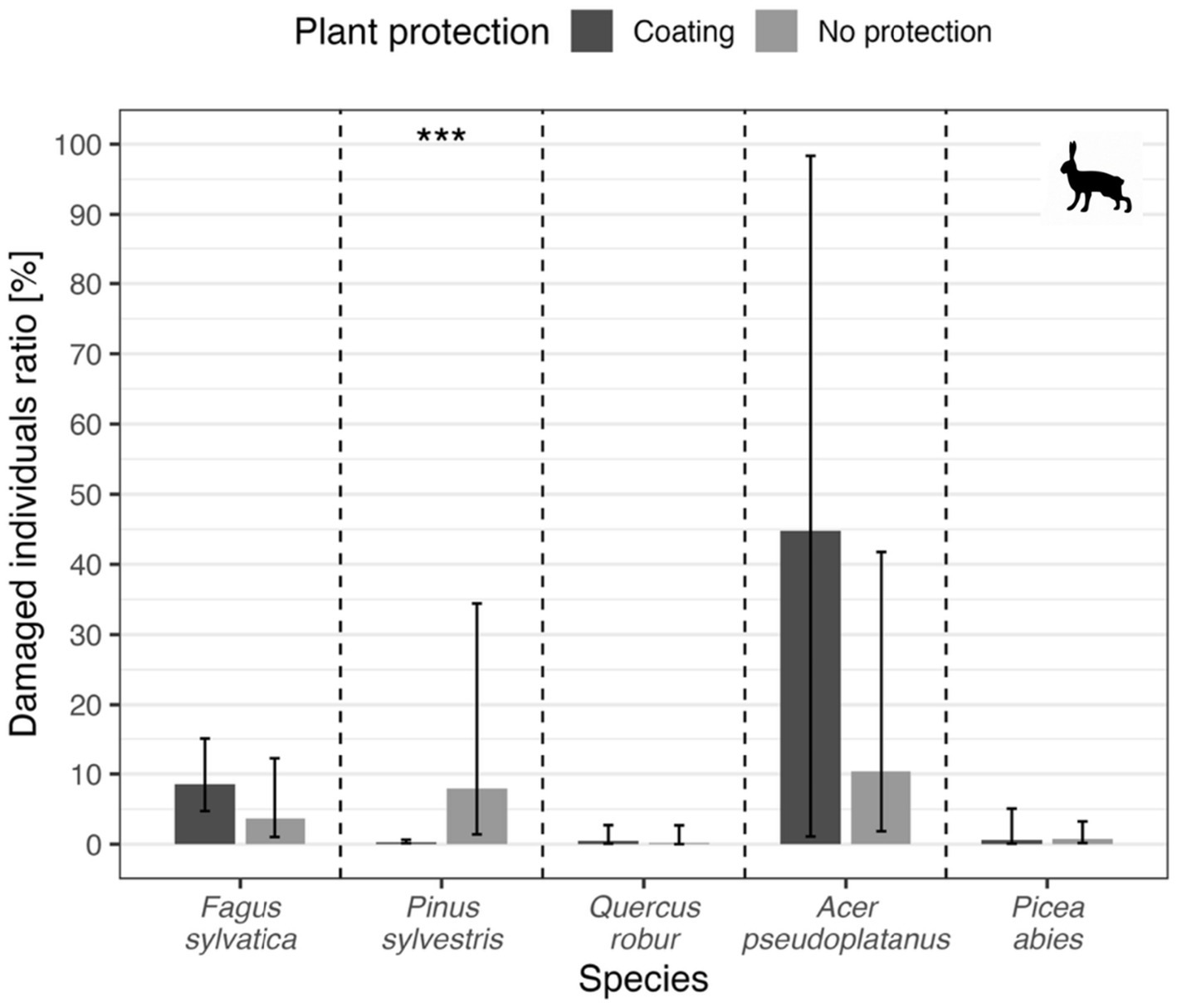
Proportion of hare-induced damage by tree species and protection. Bars show GLMM predicted marginal mean value for each variant, error bars depict a 95% confidence interval for each ratio. Symbols above each species bar pair depict statistical significance for the comparison of particular species (***p < 0.001).
The difference was not statistically significant though (p = 0.41), probably due to high variability, but still, opposite to the assumption of forest protection and coating application objectives. Similarly, in beech seedlings, which showed second highest difference ratio where coated individuals showed higher damage ratio than non-coated, the overall proportion of damage in coated individuals was 14% (marginal mean = 8.6%), however, in untreated individuals, it was approximately 9% (marginal mean = 3.7%), which again represented a statistically insignificant difference (p = 0.22), probably due to the same reason as in previous case. The opposite, but expected, positive effect of the coating was found in pine, where the overall difference between the variants was minimal—8% in coated and 10% in untreated seedlings, but, probably due to species clustering among plots, marginal means showed highly significant difference (0.3% for coated individuals vs. 7.9% for non-coated individuals, p < 0.001). In spruce, overall damage values were very low in both variants (approximately 2%), and the difference was not statistically significant (p > 0.05). The effectiveness of the coating in protecting against European hare browsing varied significantly between tree species, while it led to lower damage in some species (e.g., pine), in others (especially sycamore maple), a higher proportion of damaged individuals was recorded in the protected variant.
Figure 9 shows the percentage of damaged seedlings for the three main tree species—beech, oak, and spruce—and compares two planting variants: fenced and unfenced plots. In the case of hares, no significant differences were found, but in the case of beech, marginal insignificance was found (p = 0.06), where the most profound difference was observed – model marginal mean = 1.1% for trees in fenced enclosure, 8.0% for trees without fence protection. Other variants varied around 1%.
Figure 9
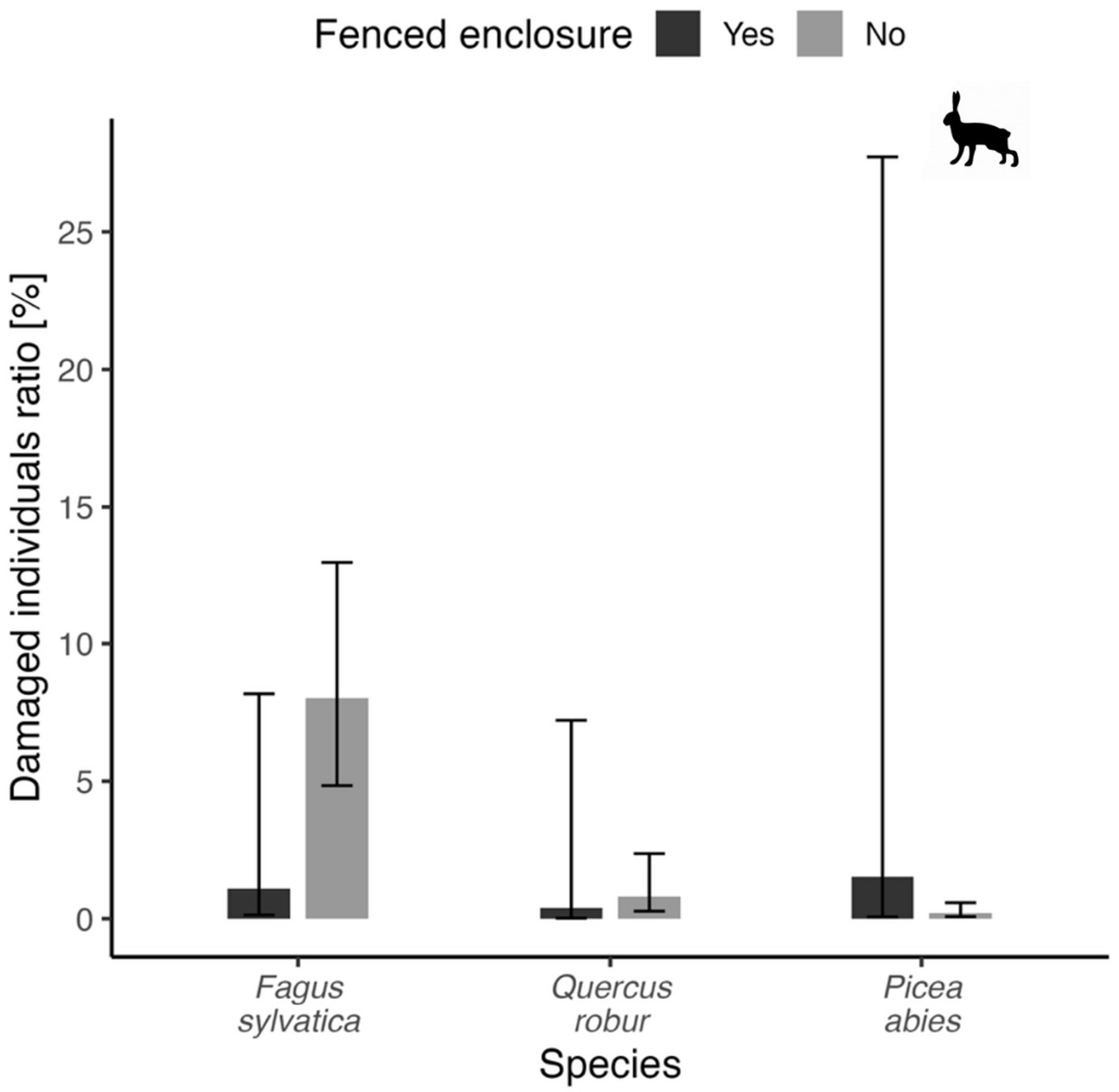
Proportion of damaged seedlings (%) in three tree species from fenced and unfenced plots for damages by hare. Bars show marginal mean values, error bars depict 95% confidence interval for each depicted variant.
No significant differences were found also for ungulates (p > 0.05 in all cases, Figure 10). Most visible difference was found in spruce – model marginal mean for trees inside fenced enclosure = 6.9%, for trees outside fenced enclosures = 0.9%. Contrary to original expectations, fenced enclosures did no prevent damages by ungulates.
Figure 10
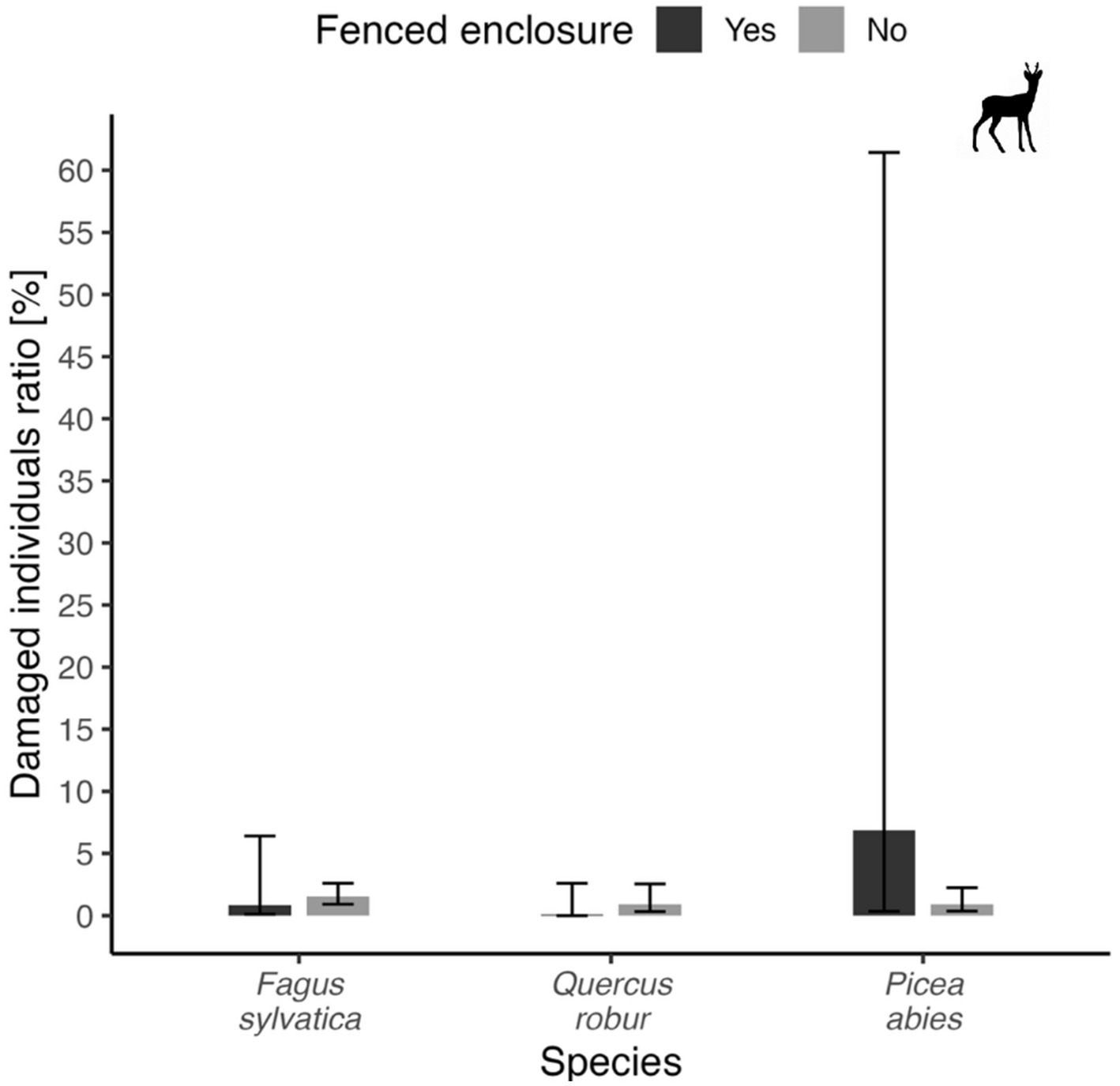
Proportion of damaged seedlings (%) in three tree species from fenced and unfenced plots for damages by ungulates. Bars show marginal mean values, error bars depict 95% confidence interval for each depicted variant.
Discussion
The results of the study clearly indicate that the European hare is a relevant, highly underestimated risk factor in forest regeneration, especially in post-calamity young deciduous tree plantations. The results of constructed GLMM model shows significantly, that trees were significantly more likely to be damaged by hares than by ungulates. The mean browsing damage to seedlings caused by hares was 10.1%, whereas browsing by wild ungulates averaged 7.1%. In general, the intensity of browsing varied significantly depending on the tree species. The highest damage was noted in birch, sycamore maple, and beech, while spruce and oak showed a lower proportion of damaged individuals. Similarly, in the Krkonoše Mountains in the Czech Republic, no more than 0.5% of sycamore maple seedlings were documented to survive due to browsing by hares (Vacek et al., 2018). These findings correspond to the selective feeding behavior of the hare, which prefers trees with soft bark and higher energy value (Hackländer et al., 2002; Schai-Braun et al., 2015). Studies report that hares consume dozens of plant species, but approximately 50% of their diet usually consists of only two or three preferred species (Puig et al., 2017). This selectivity—and high individual variability, seasonal changes, and local food availability—makes it challenging to predict browsing intensity. Nevertheless, it is possible to deduce a general rule that hares prefer energy-rich food and parts of plants with a higher fat content (Rödel et al., 2004; Katona et al., 2010) which is important to consider when planning the protection of plantings.
The ungulates, which may be mistakenly cited as the main limiting factor for the growth of regeneration and loss of species diversity in forest ecosystems (Schulze et al., 2014; Hardalau et al., 2024), caused 3% lower browsing damage in seedlings compared to hares. Wild ungulates primarily browsed terminal shoots of pine (29%)—contrary to most other references, where silver fir (Abies alba Mill.) (Bledý et al., 2024; Frei et al., 2025) and deciduous trees (Motta, 2003; Konôpka and Pajtík, 2015; Fuchs et al., 2024) are presented as suffering the most damage compared to Scots pine (Brichta et al., 2023). For example, a study of relict pine forests in the Czech Republic shows that only 1.6–12.2% of pine trees were damaged by browsing, while European beech was the most affected species, with 36.2–92.5% damage (Vacek et al., 2019). However, the results in our study may be due to the fact that Scots pine was not the main, but an allochthonous or admixed tree species. In general, as the proportion of a species decreases, its attractiveness increases significantly, regardless of the tree species (Brabec et al., 2024).
Regarding the hare-induced terminal browsing, it is also crucial to identify seasonality, which has been found to play a critical role in the risk of damage. Although the trend of a higher risk of browsing damage during the autumn compared to the spring was also observed in cloven-hoofed game, it was much more pronounced in European hares. The difference was found in Norway spruce, where the proportion of damaged individuals reached nearly 14% in the autumn, while in the spring, it did not exceed 2%. A similar trend is also evident in European beech. These values can be explained to some extent by the food preferences of hares, which mainly seek out weeds or grasses (Schai-Braun et al., 2015). The more varied species of grasses and herbs available in spring may be the primary source of food for hares. Contrastingly, in winter, it is often only dry bushgrass [Calamagrostis epigejos (L.) Roth] (Rebele and Lehmann, 2001) and other less suitable species that remain on post-calamity clearings, and for this reason, hares may prefer tree shoots in their diet. Therefore, protecting plantings through mowing weeds helps restore food sources for wild animals. However, the evaluation of the planting protection in terms of mowing and weed control by herbicide application was not the subject of this study.
Unquestionably, one of surprising results is the ambiguous effectiveness of repellent coatings. While a reduction in the browsing damage caused by wild ungulates was observed in Scots pine after the application of chemical protection, in as the case of sycamore maple, there was paradoxically found a marginal insignificantly opposite effect (p = 0.09) when the damage rate was incomparably higher for coated seedlings (1.2%) compared to untreated variant (28.6%). In the case of European hare, almost the same trend was found for sycamore maple with higher intensity of browsing on coated seedlings (45%) compared to the untreated ones (17%), however, despite the browsing rate of coated seedlings was almost 3 times higher, the differences were insignificant. There are several possible explanations for this finding. First, coated seedlings may have been more likely to be located in areas with higher expected game pressure, which may have influenced the results to some extent. Second, it is possible that hares browsed on parts of the seedlings below the coating level, especially in species with thin and attractive bark, such as sycamore maple. This phenomenon is consistent with the finding that hares adapt their behavior to food availability (Rödel et al., 2004; Mayer et al., 2018) and can specifically target unaffected parts of plants. These results indicate the need for a species-specific approach when choosing protective measures. Commonly used repellents may not be universally effective, and, without considering the target tree species or game species, their application may be ineffective or even counterproductive. Repellents appear to be an inappropriate measure for sycamore maple, while they may be somewhat effective for conifers. It should also be taken into account that hares browse close to the ground, and the application of a coating may not protect the most vulnerable parts of the seedling. In the case of cloven-hoofed game, seedlings with protective coating applied suffered higher damage rates than untreated individuals in most cases, except for pine, indicating the limited effectiveness of chemical protection even against this group of herbivores (Elmeros et al., 2011).
Contrarily, fencing proved to be the most effective method of protection against browsing, as it physically prevents access by both hares and wild ungulates. Despite the fencing, a small proportion of Norway spruce was browsed by both European hares and wild ungulates. However, this browsing most probably occurred because the fence had previously been damaged and the forage available inside was limited. Once wildlife discovered an opening, they consumed whatever vegetation was accessible, regardless of their usual food preferences, since spruce is otherwise not a preferred species. This measure confirms the conclusions of previous studies that recommend physical barriers as the most reliable and effective protection in areas with high game pressure (Gill, 1992; Redick and Jacobs, 2020). In many cases, mechanical protection of the most attractive tree species or extensive fencing of the restored forest part is practically the only possible way (Vacek et al., 2014). However, high economic costs and difficult implementation in large areas remain a disadvantage (Redick and Jacobs, 2020), requiring a combination of multiple strategies and careful planning that corresponds to local conditions. Additionally, current forestry practices in the context of climate change, such as creating forest gaps and clearcuts, can significantly increase the risk of damage to plantings by ungulates, as these open areas serve as attractive grazing habitats. Measures aimed at influencing the grazing behavior of game and dispersing its activity into the landscape could therefore be more effective than fencing and population control alone (Kuijper et al., 2009; Brabec et al., 2024).
In terms of food selection, hares typically prefer to nibble on thinner shoots of seedlings, with the diameter of damaged parts usually ranging between 2 and 3 mm. As the height of the seedling increases, the diameter of the browsed area increases only slightly, which confirms the weak but statistically significant relationship between the height of the seedling and the diameter of the damaged area. Similarly, Rao et al. (2003) emphasize the selective behavior of other species of hares (Lepus timidus L.) when browsing on young trees. Concerning the planting season, a significantly higher degree of damage was recorded in autumn plantings, regardless of tree or game species. This phenomenon can be explained by a lesser availability of other food during the autumn and winter months, which makes freshly planted seedlings more attractive (Reimoser and Gossow, 1996). On the other hand, the size of the plantation area had no statistically significant effect on the degree of damage. A review study by Smith et al. (2004) states that field size was negatively correlated with the number of the hares in half of the studies (neutral in the other studies), and that the relationship between the number of the hares and forests depends on the spatial scale. However, the higher variability in smaller areas from our study suggests that the degree of browsing is more influenced by local factors such as hare density, stand cover structure, and microclimatic conditions. This is consistent with the findings of Reichlin et al. (2006) and Voigt and Siebert (2019), who found hares prefer a mosaic landscape with sufficient cover in the peripheral parts of stands.
In relation to the formulated hypotheses, the results provide strong and coherent evidence supporting all of the stated assumptions. The first hypothesis, predicting that browsing damage by the European hare constitutes a higher proportion of total seedling damage than that caused by ungulates, was clearly confirmed. The second hypothesis was also supported, as the susceptibility of seedlings to hare browsing decreased with advancing developmental stage, reflecting the species’ preference for younger individuals with thinner shoots. The third hypothesis, which assumed that broadleaved tree species are more intensively browsed by hares than conifers, was likewise validated, with birch, sycamore maple, and beech being the most affected. Finally, the fourth hypothesis, suggesting the limited effectiveness of commonly applied protective measures such as repellents and fencing, was also confirmed. In particular, the ambiguous or even counterproductive effects of chemical repellents highlight the need for a more species-specific approach to protection. Overall, these findings underline the substantial and often underestimated influence of hares on forest regeneration and emphasize the necessity to develop more targeted and ecologically based protection strategies tailored to local conditions and species composition.
In this context, a limitation of the study is that the relationship between the degree of damage and microhabitat conditions has not been quantified in detail, which limits a more accurate interpretation of the influence of local factors such as stand cover structure or local hare density. Conversely, more than 200 sites and nearly 76,000 seedlings represent a robust sample for assessing general trends in selective browsing, not only by hares, but also by ungulates across different habitats, planting conditions, and set objectives. Here we should acknowledge that browsing is unquestionably influenced by the food supply across the evaluated plots and their surroundings (habitat context). In our study, the representation of tree species was not uniform; however, it reflected actual forestry practice and the afforestation of clearcuts after bark beetle calamity. For this reason, we do not attach excessive importance to minor differences in the preferences for individual tree species, which might vary in areas with a more diverse planting composition. Instead, we emphasize the general trends, on the basis of which hares must be regarded as a genuine threat to tree plantations under Central European conditions, a fact that has not yet been sufficiently highlighted.
Conclusion
The results of this extensive field study demonstrate that the rate of seedling damage caused by wildlife exhibits substantial species-specific variability and is influenced by multiple factors, including shoot height and diameter, planting season, and the application of protective measures. The most unexpected finding is the substantial extent of damage attributable to the European hare. The European hare prefers browsing on broadleaved tree species with thinner shoots, while ungulates tend to focus more on Scots pine. Autumn plantings were significantly more susceptible to damage than spring plantings, across both tree species and wildlife types. The analyses did not confirm a statistically significant effect of planting area size on the level of damage; however, the variance was high—particularly on smaller plots—suggesting a complex interplay of additional local factors. Repellent treatments yielded inconsistent results: while damage was reduced in some species (e.g., Scots pine), a negative effect was observed in others, particularly sycamore maple, where the treated seedlings experienced even higher levels of browsing. In the case of hare damage, repellents proved ineffective and, in some instances, increased the attractiveness of seedlings. Conversely, fencing was confirmed to be the most effective method for reducing browsing pressure. The annual browsing rate by ungulates and hares combined reached 17.2%, causing substantial damage to broadleaved species. Due to cumulative wildlife pressure, more susceptible species may eventually disappear from regeneration, whereas less affected species are able to outgrow the browsing pressure. For practical forestry and future research, it is recommended to abandon the blanket use of chemical repellents and instead implement targeted protection strategies tailored to individual tree species and specific game.
Statements
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
VS: Formal analysis, Resources, Data curation, Writing – original draft, Investigation, Validation, Conceptualization, Writing – review & editing, Methodology. JC: Formal analysis, Writing – review & editing, Methodology, Validation, Writing – original draft, Resources, Supervision, Funding acquisition, Conceptualization. RL: Methodology, Writing – review & editing, Formal analysis, Writing – original draft, Data curation. ZV: Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The research was funded by the Ministry of Agriculture of the Czech Republic, institutional support MZE-RO0123 and by Excelent Team 2025 project provided by the Faculty of Forestry and Wood Sciences, Czech University of Life Sciences in Prague.
Acknowledgments
We would like to thank Prof. Jiří Kamler, Ph.D., for his factual comments on the proposed methodological procedure. His suggestions contributed to the clarification of certain parts and helped to better formulate the key steps of the proposed methodology.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2025.1669069/full#supplementary-material
References
1
Allen C. D. Macalady A. K. Chenchouni H. Bachelet D. McDowell N. Vennetier M. et al . (2010). A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag.259, 660–684. doi: 10.1016/j.foreco.2009.09.001
2
Bates D. Mächler M. Bolker B. Walker S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw.67, 1–48. doi: 10.18637/jss.v067.i01
3
Bernard M. Barrere J. Morin X. Saïd S. Boulanger V. Granda E. et al . (2024). Elevation affects both the occurrence of ungulate browsing and its effect on tree seedling growth for four major tree species in European mountain forests. Ann. For. Sci.81:13. doi: 10.1186/s13595-024-01226-x
4
Bledý M. Vacek S. Brabec P. Vacek Z. Cukor J. Černý J. et al . (2024). Silver fir (Abies alba mill.): review of ecological insights, Forest management strategies, and climate change’s impact on European forests. Forests15:998. doi: 10.3390/f15060998
5
Brabec P. Cukor J. Vacek Z. Vacek S. Skoták V. Ševčík R. et al . (2024). Wildlife damage to forest stands in the context of climate change - a review of current knowledge in the Czech Republic. Central Eur. Forest. J.70, 207–221. doi: 10.2478/forj-2024-0016
6
Brázdil R. Stucki P. Szabó P. Řezníčková L. Dolák L. Dobrovolný P. et al . (2018). Windstorms and forest disturbances in the Czech lands: 1801–2015. Agric. For. Meteorol.250–251, 47–63. doi: 10.1016/j.agrformet.2017.11.036
7
Brichta J. Vacek S. Vacek Z. Cukor J. Mikeska M. Bílek L. et al . (2023). Importance and potential of scots pine (Pinus sylvestris L.) in 21st century. Central Eur. Forest. J.69, 3–20. doi: 10.2478/forj-2022-0020
8
Champagne E. Raymond P. Royo A. A. Speed J. D. M. Tremblay J. P. Côté S. D. (2021). A review of ungulate impacts on the success of climate-adapted Forest management strategies. Curr. For. Rep.7, 305–320. doi: 10.1007/s40725-021-00148-5
9
Elmeros M. Winbladh J. K. Andersen P. N. Madsen A. B. Christensen J. T. (2011). Effectiveness of odour repellents on red deer (Cervus elaphus) and roe deer (Capreolus capreolus): a field test. Eur. J. Wildl. Res.57, 1223–1226. doi: 10.1007/s10344-011-0517-y
10
Frei E. R. Conedera M. Bebi P. Zürcher S. Bareiss A. Ramstein L. et al . (2025). High potential but little success: ungulate browsing increasingly impairs silver fir regeneration in mountain forests in the southern Swiss Alps. Forestry98, 194–203. doi: 10.1093/forestry/cpae028
11
Fuchs Z. Vacek Z. Vacek S. Cukor J. Šimůnek V. Štefančík I. et al . (2024). European beech (Fagus sylvatica L.): a promising candidate for future forest ecosystems in Central Europe amid climate change. Central Eur. Forestry J.70, 62–76. doi: 10.2478/forj-2023-0020
12
Gerhardt P. Arnold J. M. Hackländer K. Hochbichler E. (2013). Determinants of deer impact in European forests - a systematic literature analysis. For. Ecol. Manag.310, 173–186. doi: 10.1016/j.foreco.2013.08.030
13
Gill R. M. A. (1992). A review of damage by mammals in north temperate forests: 1. Deer. Forestry65, 145–169. doi: 10.1093/forestry/65.2.145
14
González de Andrés E. Seely B. Blanco J. A. Imbert J. B. Lo Y. H. Castillo F. J. (2017). Increased complementarity in water‐limited environments in Scots pine and European beech mixtures under climate change. Ecohydrology, 10:e1810. doi: 10.1002/eco.1810
15
Green K. Davis N. E. Robinson W. A. McAuliffe J. Good R. B. (2013). Diet selection by European hares (Lepus europaeus) in the alpine zone of the Snowy Mountains. Australia. Eur. J. Wildl. Res.59, 693–703. doi: 10.1007/s10344-013-0723-x
16
Gryz J. Krauze-Gryz D. (2022). Why did Brown hare Lepus europaeus disappear from some areas in Central Poland?Diversity14:465. doi: 10.3390/d14060465
17
Hackländer K. Tataruch F. Ruf T. (2002). The effect of dietary fat content on lactation energetics in the European hare (Lepus europaeus). Physiol. Biochem. Zool.75, 19–28. doi: 10.1086/324770
18
Hardalau D. Codrean C. Iordache D. Fedorca M. Ionescu O. (2024). The expanding thread of ungulate browsing—a review of Forest ecosystem effects and management approaches in Europe. Forests15:1311. doi: 10.3390/f15081311
19
Hjältén J. Danell K. Ericson L. (2004). Hare and vole browsing preferences during winter. Acta Theriol.49, 53–62. doi: 10.1007/BF03192508
20
Hlásny T. Barka I. Kulla L. Bucha T. Sedmák R. Trombik J. (2017). Sustainable forest management in a mountain region in the Central Western Carpathians, northeastern Slovakia: the role of climate change. Reg. Environ. Chang.17, 65–77. doi: 10.1007/s10113-015-0894-y
21
Hulbert I. A. Iason G. R. Mayes R. W. (2001). The flexibility of an intermediate feeder: dietary selection by mountain hares measured using faecal n-alkanes. Oecologia129, 197–205. doi: 10.1007/s004420100725
22
Jansson G. Pehrson Å. (2007). The recent expansion of the brown hare (Lepus europaeus) in Sweden with possible implications to the mountain hare (L. timidus). Eur. J. Wildl. Res.53, 125–130. doi: 10.1007/s10344-007-0086-2
23
Kahlert J. Fox A. D. Heldbjerg H. Asferg T. Sunde P. (2015). Functional responses of human hunters to their prey-why harvest statistics may not always reflect changes in prey population abundance. Wildlife Biol.21, 294–302. doi: 10.2981/wlb.00106
24
Katona K. Biró Z. S. Szemethy L. Demes T. Nyeste M. (2010). Spatial, temporal and individual variability in the autumn diet of european hare (lepus europaeus) in Hungary. Acta Zool. Acad. Sci. Hung.56, 89–101.
25
Konôpka B. Pajtík J. (2015). Why was browsing by red deer more frequent but represented less consumed mass in young maple than in ash trees?!J. For. Sci.61, 431–438. doi: 10.17221/70/2015-JFS
26
Krivopalova A. Mikula P. Cukor J. Ševčík R. Brynychová K. Šálek M. (2024). Adaptation of farmland mammalian specialist to urban life: escape behavior of European hare along the urban-rural gradient. Sci. Total Environ.951:175779. doi: 10.1016/j.scitotenv.2024.175779
27
Kuijper D. P. J. Cromsigt J. P. G. M. Churski M. Adam B. Jedrzejewska B. Jedrzejewski W. (2009). Do ungulates preferentially feed in forest gaps in European temperate forest?For. Ecol. Manag.258, 1528–1535. doi: 10.1016/j.foreco.2009.07.010
28
Kuznetsova A. Brockhoff P. B. Christensen R. H. Jensen S. P. (2016). Tests in linear mixed effects models. R Package Version2:33.
29
Lenth R. (2023). emmeans: estimated marginal means, aka least-squares Means_. R Package Version1:5.
30
Lindner M. Fitzgerald J. B. Zimmermann N. E. Reyer C. Delzon S. van der Maaten E. et al . (2014). Climate change and European forests: what do we know, what are the uncertainties, and what are the implications for forest management?J. Environ. Manag.146, 69–83. doi: 10.1016/j.jenvman.2014.07.030
31
Marboutin E. Aebischer N. J. (1996). Does harvesting arable crops influence the behaviour of the European hare Lepus europaeus?Wildlife Biol.2, 83–91. doi: 10.2981/wlb.1996.036
32
Mayer M. Ullmann W. Sunde P. Fischer C. Blaum N. (2018). Habitat selection by the European hare in arable landscapes: the importance of small-scale habitat structure for conservation. Ecol. Evol.8, 11619–11633. doi: 10.1002/ece3.4613
33
Motta R. (2003). Ungulate impact on rowan (Sorbus aucuparia L.) and Norway spruce (Picea abies (L.) karst.) height structure in mountain forests in the eastern Italian Alps. For. Ecol. Manag.181, 139–150. doi: 10.1016/S0378-1127(03)00128-2
34
Nagel T. A. Mikac S. Dolinar M. Klopcic M. Keren S. Svoboda M. et al . (2017). The natural disturbance regime in forests of the Dinaric Mountains: a synthesis of evidence. For. Ecol. Manag.388, 29–42. doi: 10.1016/j.foreco.2016.07.047
35
Peel M. C. Finlayson B. L. McMahon T. A. (2007). Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sc.11, 1633–1644. doi: 10.5194/hess-11-1633-2007
36
Pepper H. Hedge S. (1998). The prevention of mammal damage to trees in woodland. Edinburgh: Forestry Commission.
37
Pretzsch H. Steckel M. Heym M. Biber P. Ammer C. Ehbrecht M. et al . (2020). Stand growth and structure of mixed-species and monospecific stands of scots pine (Pinus sylvestris L.) and oak (Q. robur L., Quercus petraea (Matt.) Liebl.) analysed along a productivity gradient through Europe. Eur. J. For. Res.139, 349–367. doi: 10.1007/s10342-019-01233-y
38
Puig S. Rosi M. I. Videla F. Mendez E. (2017). Flexibility in the food selection by the European hare (Lepus europaeus) along the altitudinal gradient of the southern Andean Precordillera (Argentina). Mamm. Res.62, 75–87. doi: 10.1007/s13364-016-0288-7
39
R Core Team (2021). R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing.
40
Ramirez J. I. Jansen P. A. Poorter L. (2018). Effects of wild ungulates on the regeneration, structure and functioning of temperate forests: a semi-quantitative review. For. Ecol. Manag.424, 406–419. doi: 10.1016/j.foreco.2018.05.016
41
Rao S. J. Iason G. R. Hulbert I. A. R. Daniels M. J. Racey P. A. (2003). Tree browsing by mountain hares (Lepus timidus) in young scots pine (Pinus sylvestris) and birch (Betula pendula) woodland. For. Ecol. Manag.176, 459–471. doi: 10.1016/S0378-1127(02)00311-0
42
Rebele F. Lehmann C. (2001). Biological flora of central europe: Calamagrostis epigejos (L.) Roth. Flora196, 325–344. doi: 10.1016/S0367-2530(17)30069-5
43
Redick C. H. Jacobs D. F. (2020). Mitigation of deer herbivory in temperate hardwood forest regeneration: a meta-analysis of research literature. Forests11:1220. doi: 10.3390/f11111220
44
Reichlin T. Klansek E. Hackländer K. (2006). Diet selection by hares (Lepus europaeus) in arable land and its implications for habitat management. Eur. J. Wildl. Res.52, 109–118. doi: 10.1007/s10344-005-0013-3
45
Reimoser F. Gossow H. (1996). Impact of ungulates on forest vegetation and its dependence on the silvicultural system. For. Ecol. Manag.88, 107–119. doi: 10.1016/S0378-1127(96)03816-9
46
Rödel H. G. Völkl W. Kilias H. (2004). Winter browsing of brown hares: evidence for diet breadth expansion. Mamm. Biol.69, 410–419. doi: 10.1078/1616-5047-00163
47
Schai-Braun S. C. Reichlin T. S. Ruf T. Klansek E. Tataruch F. Arnold W. et al . (2015). The European hare (Lepus europaeus): a picky herbivore searching for plant parts rich in fat. PLoS One10:e0134278. doi: 10.1371/journal.pone.0134278
48
Schulze E. D. Bouriaud O. Wäldchen J. Eisenhauer N. Walentowski H. Seele C. et al . (2014). Ungulate browsing causes species loss in deciduous forests independent of community dynamics and silvicultural management in central and southeastern Europe. Ann. For. Res.57, 267–288. doi: 10.15287/afr.2014.273
49
Schwegmann S. Mörsdorf M. Bhardwaj M. Storch I. (2023). Effects of understory characteristics on browsing patterns of roe deer in central European mountain forests. Ecol. Evol.13:e10431. doi: 10.1002/ece3.10431
50
Seliger A. Ammer C. Kreft H. Zerbe S. (2023). Diversification of coniferous monocultures in the last 30 years and implications for forest restoration: a case study from temperate lower montane forests in Central Europe. Eur. J. For. Res.142, 1353–1368. doi: 10.1007/s10342-023-01595-4
51
Smith R. K. Jennings N. V. Harris S. (2005). A quantitative analysis of the abundance and demography of European hares Lepus europaeus in relation to habitat type, intensity of agriculture and climate.
52
Smith R. K. Jennings N. V. Robinson A. Harris S. (2004). Conservation of European hares Lepus europaeus in Britain: is increasing habitat heterogeneity in farmland the answer?J. Appl. Ecol.41, 1092–1102. doi: 10.1111/j.0021-8901.2004.00976.x
53
Steckel M. del Río M. Heym M. Aldea J. Bielak K. Brazaitis G. et al . (2020). Species mixing reduces drought susceptibility of scots pine (Pinus sylvestris L.) and oak (Quercus robur L., Quercus petraea (Matt.) Liebl.) – site water supply and fertility modify the mixing effect. For. Ecol. Manag.461:117908. doi: 10.1016/j.foreco.2020.117908
54
Unkule M. Piedallu C. Balandier P. Courbaud B. (2022). Climate and ungulate browsing impair regeneration dynamics in spruce-fir-beech forests in the French Alps. Ann. For. Sci.79:11. doi: 10.1186/s13595-022-01126-y
55
Vacek Z. Prokůpková A. Vacek S. Bulušek D. Šimůnek V. Hájek V. et al . (2021). Mixed vs. monospecific mountain forests in response to climate change: structural and growth perspectives of Norway spruce and European beech. For. Ecol. Manag.488:119019. doi: 10.1016/j.foreco.2021.119019
56
Vacek Z. Vacek S. Bílek L. Král J. Remeš J. Bulušek D. et al . (2014). Ungulate impact on natural regeneration in spruce-beech-fir stands in Černý důl nature reserve in the Orlické hory mountains, case study from central sudetes. Forests5, 2929–2946. doi: 10.3390/f5112929
57
Vacek S. Vacek Z. Bílek L. Remeš J. Hůnová I. Bulušek D. et al . (2019). Stand dynamics in natural scots pine forests as a model for adaptation management?Dendrobiology82, 24–42. doi: 10.12657/denbio.082.004
58
Vacek Z. Vacek S. Cukor J. (2023). European forests under global climate change: review of tree growth processes, crises and management strategies. J. Environ. Manag.332:117353. doi: 10.1016/j.jenvman.2023.117353
59
Vacek Z. Vacek S. Cukor J. Bulušek D. Slávik M. Lukáčik I. et al . (2022). Dendrochronological data from twelve countries proved definite growth response of black alder (Alnus glutinosa [L.] Gaertn.) to climate courses across its distribution range. Central Eur. Forest. J.68, 139–153. doi: 10.2478/forj-2022-0003
60
Vacek S. Vacek Z. Kalousková I. Cukor J. Bílek L. Moser W. K. et al . (2018). Sycamore maple (Acer pseudoplatanus L.) stands on former agricultural land in the sudetes – evaluation of ecological value and production potential. Dendrobiology79, 61–76. doi: 10.12657/denbio.079.006
61
Viewegh J. Kusbach A. Mikeska M. (2003). Czech forest ecosystem classification. J For Sci49, 74–82. doi: 10.17221/4682-jfs
62
Voigt U. Siebert U. (2019). Living on the edge - circadian habitat usage in pre-weaning European hares (Lepus europaeus) in an intensively used agricultural area. PLoS One14:e0222205. doi: 10.1371/journal.pone.0222205
63
Vowles T. Molau U. Lindstein L. Molau M. Björk R. G. (2016). The impact of shrub browsing by mountain hare and reindeer in subarctic Sweden. Plant Ecol Divers9, 421–428. doi: 10.1080/17550874.2016.1264017
64
Wilkinson L. (2011). ggplot2: elegant graphics for data analysis by WICKHAM. H. Biometrics67, 678–679. doi: 10.1111/j.1541-0420.2011.01616.x
65
Zamora-Pereira J. C. Yousefpour R. Cailleret M. Bugmann H. Hanewinkel M. (2021). Magnitude and timing of density reduction are key for the resilience to severe drought in conifer-broadleaf mixed forests in Central Europe. Ann. For. Sci.78:68. doi: 10.1007/s13595-021-01085-w
Summary
Keywords
browsing damage, Lepus europaeus , tree species attractiveness, post-calamity reforestation, wildlife damage, climate change adaptation
Citation
Skoták V, Cukor J, Linda R and Vacek Z (2025) The European hare as a hidden challenge for forest regeneration: common protection measures are ineffective. Front. For. Glob. Change 8:1669069. doi: 10.3389/ffgc.2025.1669069
Received
18 July 2025
Accepted
16 October 2025
Published
30 October 2025
Volume
8 - 2025
Edited by
Arshad Ali, Hebei University, China
Reviewed by
Yiannis G. Zevgolis, University of the Aegean, Greece
Anastazija Dimitrova, University of Molise, Italy
Updates
Copyright
© 2025 Skoták, Cukor, Linda and Vacek.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Vlastimil Skoták, vlastimil.skotak@gmail.com
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.