 Khalid Awadh Al-Mutairi
Khalid Awadh Al-Mutairi- Department of Biology, Faculty of Science, University of Tabuk, Tabuk, Saudi Arabia
Climate change, hydrological alterations, and anthropogenic pressures are collectively driving rapid ecological shifts in Saudi Arabia’s arid regions, particularly in Tabuk and Khulais. This study evaluates the status and conservation strategies of floristic diversity in these regions through a bibliometric and ecological synthesis. Using bibliometric analysis of 102 selected publications from 1984 to 2025, major thematic clusters were identified, focusing on policy interventions, hydrological impacts, biodiversity loss, and vegetation dynamics. Among these, 20 studies specifically addressed the Tabuk region’s ecological challenges and conservation responses. The findings reveal that rising temperatures (0.70–1.30 °C) and drastic rainfall reduction (up to 85%) have intensified aridity, decreased soil moisture, and accelerated biodiversity degradation. Invasive species such as Prosopis juliflora exacerbate the situation by outcompeting native flora for scarce resources. While rocky ecosystems exhibit resilience due to microclimatic buffering, fragile valley ecosystems require urgent, targeted measures like artificial water input and soil stabilization. Hydrological projects, including dam construction, have yielded mixed outcomes—enhancing species richness in riparian zones while simultaneously disrupting natural seed dispersal and causing habitat fragmentation. Rare and endemic species such as Rheum palaestinum and Astragalus collenettiae serve as vital indicators of ecosystem health and genetic heritage, underscoring the need for species-specific conservation initiatives. The study recommends integrated strategies that combine habitat restoration, sustainable water management, and long-term biodiversity monitoring. Furthermore, ecotourism development and community participation are highlighted as valuable components in achieving conservation goals, aligning with Saudi Arabia’s Vision 2030 and national biodiversity targets. This work contributes a comprehensive framework for managing arid-region floristic diversity under changing climatic conditions. By bridging ecological research with socio-economic and policy dimensions, the study offers valuable insights for sustainable forest and landscape management in desert-margin ecosystems globally.
1 Introduction
Arid ecosystems, which are ecologically vulnerable, are some of the most vulnerable as a result of climate change and hydrological changes reshaping ecosystems worldwide. Minor temperature and precipitation changes can destabilize the already delicate balance between water availability and biodiversity, thereby generating habitat destruction, extinction of endemic plant species, and spread of invasive species (Cardinale et al., 2012; Maestre et al., 2012; Ali et al., 2016). These difficulties emphasize the requirement for more research on the effects of climate change and human activity on ecosystem processes (IPCC, 2014; Ghesquière et al., 2016; Liu et al., 2019). The extremely arid climate of Saudi Arabia helps one a lot to realize these ecological interruptions (Al-Masri and Alsharif, 2021).
The area is extreme in aridity and lack of water supply and also suffers from severe environmental stresses due to climate change. A few of the human-induced changes that continue to modify hydrological regimes include the construction of dams and land use change, which both influence soil characteristics and plant community composition (Al-Munqedhi et al., 2022; Alsherif and Almaghrabi, 2022). Other arid and semi-arid ecosystems, such as the Cape Floristic Region of South Africa, where species distribution changes due to climate change imperil floristic diversity, have extensively documented the worldwide implications of such changes (Alsherif and Almaghrabi, 2022; Anju and Kumar, 2024). Invasive species such as Prosopis juliflora take advantage of such environmental stresses, replacing natural vegetation and modifying ecological processes. These trends highlight the urgent need for anticipatory biodiversity monitoring and management interventions to pre-empt species decline and ecosystem degradation (Jewitt et al., 2017; Azeem et al., 2023; Bai et al., 2022). The decision to make the Tabuk and Khulais regions (Figure 1) the focus of this review is a product of their ecological vulnerability, rising development pressure, and relative underrepresentation in global floristic and conservation research. Found in northwestern and western Saudi Arabia, respectively, both regions exhibit unique biogeographic features, ranging from steep mountains and basaltic plateaus to ephemeral wadis and secluded valleys.
Figure 1. Location of Tabuk (in blue) and Khulais (in green) in Saudi Arabia arid ecosystem. Source: GoogleMap.
These micro-habitats harbor endemic and rare plant species like Rheum palaestinum and Astragalus collenettiae that serve as important indicators of ecosystem health and stability in hyper-arid conditions (Alsherif and Almaghrabi, 2022). Moreover, Tabuk is undergoing intensified urbanization and agricultural development as a component of the countrywide NEOM development project, while Khulais is dealing with hydrological disruption due to damming and groundwater extraction. These changes are compounded by climate projections of rising temperatures and further reductions in rainfall (Sillmann et al., 2013; Zittis et al., 2022; Lelieveld et al., 2016). This combination of ecological interest and anthropogenic pressure makes the two regions model case studies both for assessing the degree of floristic diversity and for forming locally relevant conservation strategies. Focusing on the two regions allows a targeted bibliometric and ecological synthesis that can extrapolate to broader desert conservation policy in Saudi Arabia and similar arid ecosystems worldwide. To identify the floristic plant diversity of Tabuk’s and Khulais’ desert ecosystems, recent studies provide valuable ecological and conservation data. The Tabuk region, in particular, shows considerable floristic diversity as a result of both natural edaphic factors and anthropogenic disturbances such as woodcutting, overgrazing, and urbanization (Al-Mutairi, 2017).
A complete survey of Tabuk area showed 163 plant species distributed across 41 families and 124 genera, with high diversity indices being measured in areas such as Sharma and Aldesah (Al-Mutairi, 2022). Indices such as species richness, evenness, Brillouin, Menhinick, Margalef, equitability, and Chao-1 estimates indicate the ecological richness of selected microhabitats. The diversity patterns have significant correlations with environmental parameters. Soil pH, electrical conductivity, texture, and nutrient content—particularly calcium, potassium, phosphorus, organic matter, bicarbonate, and sodium—were found to be key determinants of spatial plant distribution. Principal coordinate analyses also revealed high correlations of pH and potassium with the first axis, with organic matter and sodium associated with the second axis (Al-Mutairi, 2022). Given the rising anthropogenic pressures, Tabuk needs urgent conservation efforts to prevent irretrievable damages to the regional plant communities (Al-Mutairi, 2017). In comparison, the Khulais region faced significant climatic changes during the previous decade, as reflected in decreasing yearly rainfall and rising mean yearly temperatures (Alsherif and Almaghrabi, 2022). Different drought indices, including those of Lange, De Martonne, and Emberger, all show the region is facing heightened aridity.
These climatic shifts have had a profound effect on the floristic structure of Khulais. A recent study revealed a considerable reduction in the frequency of most plant species—particularly in agricultural areas and valley communities—compared to relatively stable populations in rocky and mountainous communities (Alsherif and Almaghrabi, 2022).
Both regions demonstrate that conservation must be spatially appropriate, ecologically grounded, and sensitive to climate projections. This dual regional focus establishes a sound foundation for developing targeted conservation strategies that further Saudi Arabia’s Vision 2030 goals while safeguarding endemic and native plant biodiversity in desert ecosystems.
A hotspot of biological diversity in Saudi Arabia’s desert environment, Tabuk is a territory with a variety of ecosystems including desert plains, wadis, and coastal environments. Supportive of some 1,800 plant species which have been recorded (Kürschner and Neef, 2011; Fakhry and El-Kenany, 2019), the area is home to endemic and endangered plant species such as A. collenettiae and R. palaestinum which are extremely susceptible to threats from human activities and environmental factors. Maintained biodiversity and ecosystem processes rely on intensified conservation efforts owing to the ecological significance and vulnerability of the region. Considering Tabuk specifically, this review article attempts to integrate the body of evidence regarding the ecological effects of hydrological manipulations and climate change on vegetation diversity in Saudi Arabia’s hyper-arid regions. This article aims to tackle large areas of research gap, such as the lack of long-term biodiversity monitoring and adaptive management of native and alien species—using bibliometric analysis and based on evidence from the literature.
2 Methodology
A Systematic Literature Review (SLR) approach based on the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guideline (Moher et al., 2009; Figure 2) was employed in this study to assess the scientific literature on the effects of climate change on floristic composition and plant diversity. The review aimed at uncovering dominant research themes, identify temporal patterns, and investigate knowledge gaps regarding the impact of climate change on plant populations worldwide.
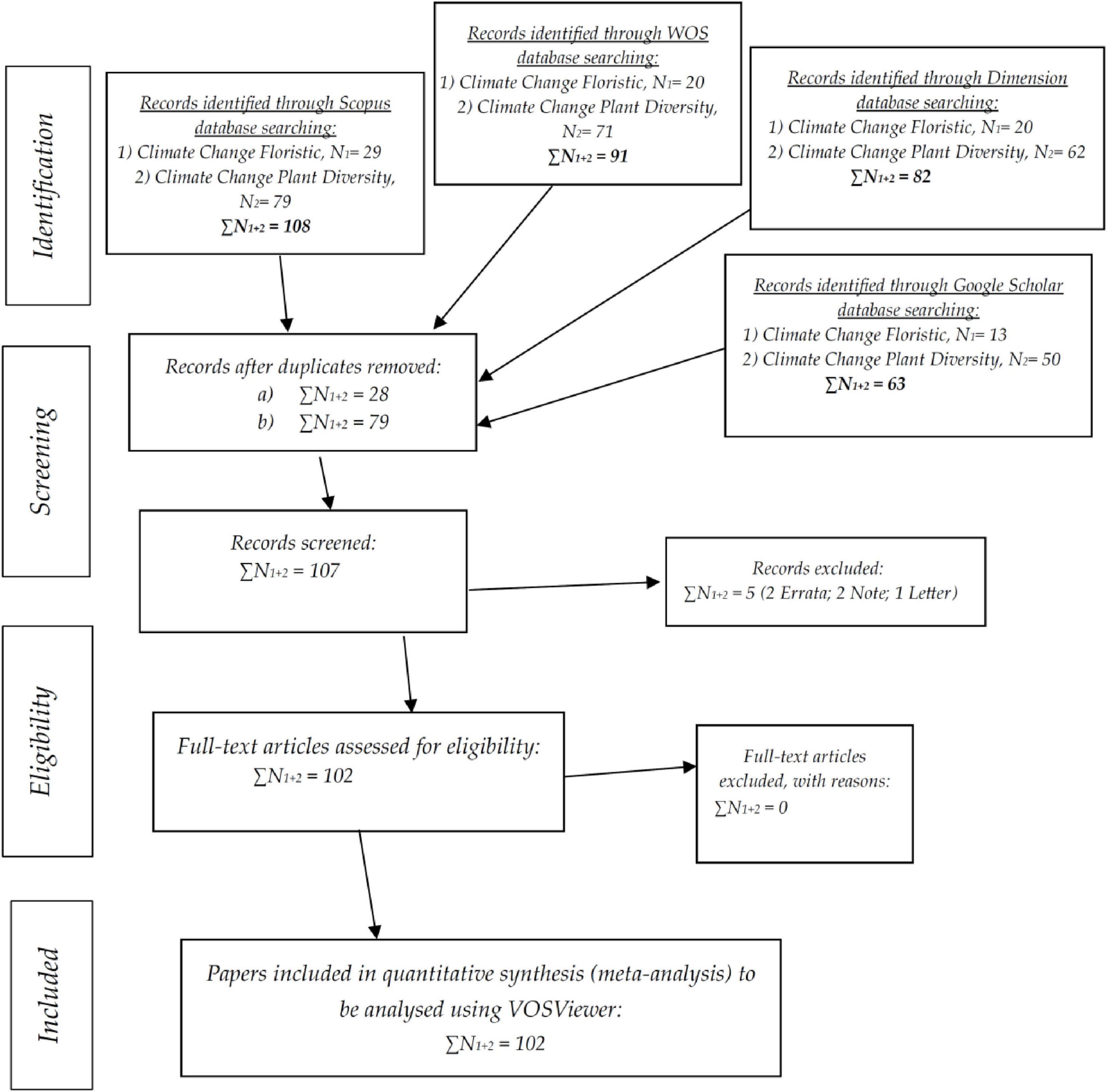
Figure 2. Flowchart of the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) [adapted from Moher et al. (2009)], used in the present study.
To determine the inclusivity and strength of the pool of literature, a multi-databases searching strategy was employed. The search operations were conducted between 1 and 3 March 2025 across four leading research platforms: Scopus, Web of Science (WOS), Dimensions, and Google Scholar. Two keyword searches were employed, “Climate Change Floristic” and “Climate Change Plant Diversity.” Boolean operators and field specifications were adjusted minimally to fit the format and constraints of each respective database. Filters were set to include only English language peer-review articles and review papers published between 1984 and 2025. Scopus search yielded 108 papers (29 in “Floristic” and 79 in “Plant Diversity”), while WOS yielded 91 papers (20 and 71, respectively). Dimensions gave 82 papers (20 and 62, respectively), and Google Scholar yielded an additional 63 (13 and 50, respectively) to arrive at the number of initial records of 344.
Following the identification stage, all the articles identified were exported into a citation manager, and duplicates were removed in a systematic manner. A total of 107 unique records remained following deduplication. The records were next screened for assessment of relevance by title and abstract. Five records were eliminated since they were not in the full-text mode, made up of two errata, two notes, and one letter, leaving a dataset of 102 papers which were available for more in-depth review.
All articles contained within the data set satisfied the inclusion criteria of being systematic reviews or empirical studies that had explicitly examined the impact of climate change on plant diversity or floristic composition, especially in arid and semi-arid habitats. The excluded materials were largely non-academic works such as opinion articles, brief communications, and editorial remarks that were not empirically based.
There is a total of 102 candidate articles was subsequently exported in CSV format (Supplementary Table 1) for bibliometric analysis using VOSviewer (version 1.6.20), created by Van Eck and Waltman (2010, 2014, 2017). The metadata of every article including titles, abstracts, authors, affiliations, journal titles, keywords, and publication years were harvested. Keyword co-occurrence analysis was subsequently performed using full counting to ensure balanced keyword representation across the corpus. Analysis employed a threshold of five occurrences in order to introduce keywords to the final mapping procedure.
Bibliometric analysis produced network visualizations which revealed general areas of research focus and clusters in climate change and plant diversity. Overlap visualizations also facilitated examination of trends in time, whereby chosen topics of research are proved to evolve over time. This integrative and structured approach provided a good foundation for later interpretation of research directions, ecological outcomes, and policy implications, particularly regarding climate change impacts on Saudi Arabian arid regions and other dryland areas’ biodiversity.
3 Results
3.1 Literature on “floristic” and “climate change plant diversity”
Each of the various clusters shown in Figure 3 is a product of climate change studies’ bibliometric network analysis, thereby displaying a focal point topic. “Climate change” is the most connected node, thereby highlighting its centripetal role across multiple science disciplines. Four major groups resulted, each characterized by differing research topics: the red group (plant dynamics and model development), blue group (abiotic processes and grasslands), purple group (phylogeny, genetics, and conservation), and green group (ecosystems and biodiversity conservation). Collaboration between these groups, combining biology, ecology, environmental science, and conservation planning, is indicative of the interdisciplinary nature of climate change research.
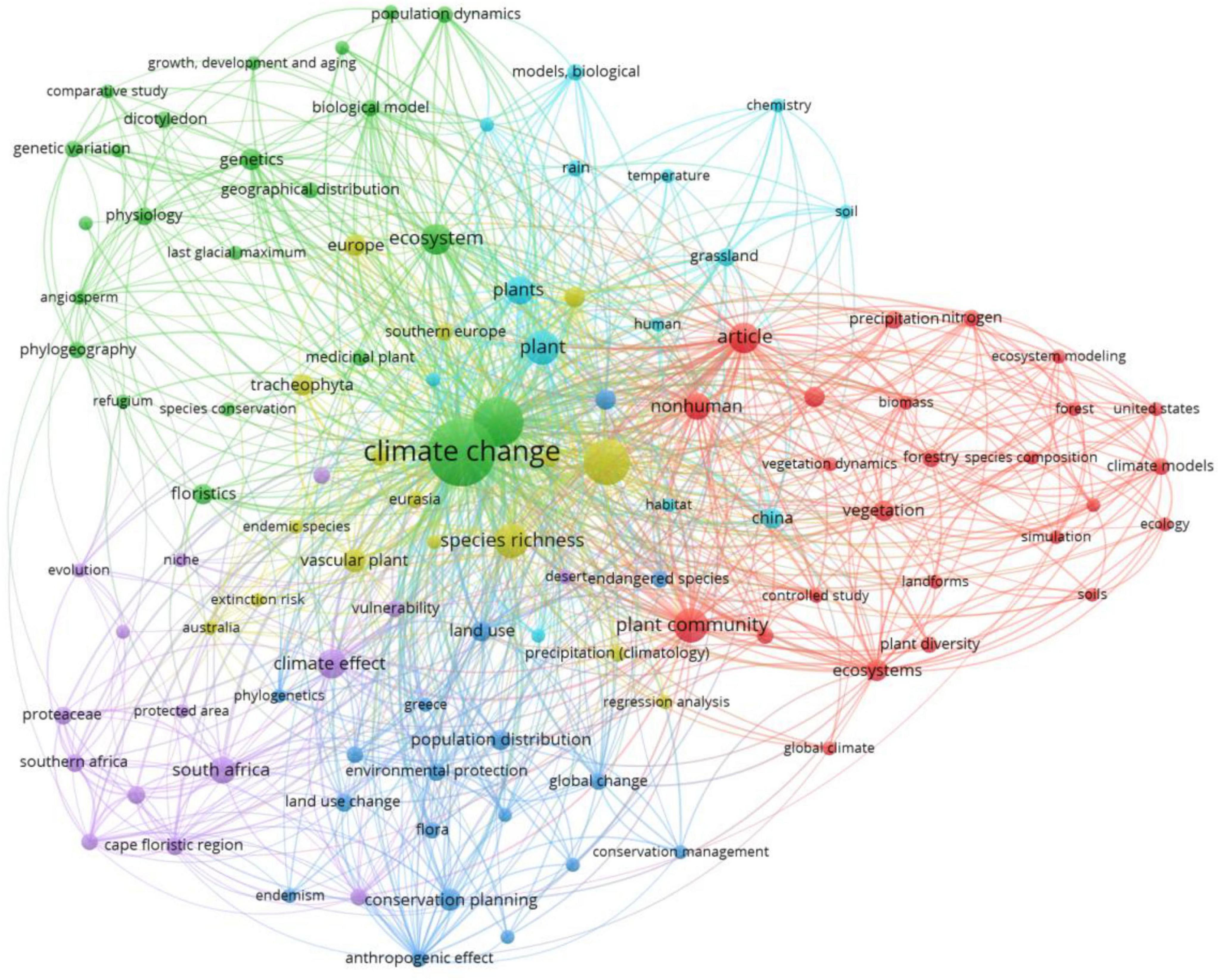
Figure 3. Visualization of similarity using VOSviewer based on Scopus database using the keyword “climate change floristic” retrieved 28 articles (1981–2024), while with the keyword “climate change plant diversity” yielded 78 articles (2000–2025), searched on 2 March 2025.
3.1.1 The green cluster: ecosystems and biodiversity conservation
The green cluster, which encompasses significant terms like “ecosystem,” “species richness,” “vascular plants,” and “land use,” is concerned mainly with ecosystems and biodiversity conservation. This cluster reflects research that investigates the effect of land use change and climate on biodiversity and ecosystem services. Research work in this field is more directed toward the need for conservation planning in halting biodiversity losses and promoting sustainable ecosystem management. Interest is specific in human-induced problems like habitat fragmentation and forest loss, as well as species richness as a measure of ecosystem condition. This category provides informative data for decision makers to prioritize top preservation programs by pointing out hotspots for conservation.
3.1.2 The red cluster: plant dynamics and predictive modeling
The red cluster deals with plant dynamics and predictive modeling using terms like “plant community,” “vegetation,” “biomes,” and “ecosystem modeling.” The cluster includes studies that try to know how the plant communities respond to varying climatic conditions by using scenario-based simulations and vegetation models. Tends to use climate data, extrapolating future biome change and providing data on anticipated changes in the distribution and constitution of vegetation. Of particular note in areas where ecosystems are most vulnerable to the impacts of climate change, this cluster emphasizes the role of climate models in the development of mitigation and adaptation policy.
3.1.3 The blue cluster: abiotic processes and grassland studies
The blue cluster, with its identification of words like “temperature,” “soil,” and “grassland,” emphasizes abiotic processes and how these affect plant ecosystems. Research in this cluster assesses the effect of abiotic factors like temperature, precipitation, and soil quality on grassland ecosystems. Also considered is the role of biogeochemical processes during climate change in nutrient cycling and ecosystem resilience. Usually considered to be carbon sinks, grasslands are actually the very thing necessary for mitigating the effects of climate change, so this category ranks them strongly. Research in this category also highlights how abiotic and ecosystem function occur together.
3.1.4 The purple cluster: phylogeny, genetics, and conservation
The purple cluster, which contains words like “phylogeny,” “genetics,” and “conservation planning,” reflects all three: phylogeny, genetics, and conservation planning. This cluster analyzes the evolutionary and genetic mechanisms of plants’ responses to climate change. It emphasizes the need for phylogenetic study and genetic variation in understanding how plants respond to environmental stresses. It also applies this knowledge to practical conservation methods like maintaining genetic diversity in order to improve species resistance and creating reserves to preserve evolutionary history.
3.1.5 Interconnectivity between clusters
The network analysis reveals strong connectivity among the clusters, in this case between the purple and the green ones. The use of words like “species richness” and “biodiversity” suggests the ways in which empirical data and theoretical frameworks may be combined by linking observed research on biodiversity with predicted models on vegetation change. In accord, the overlap of the purple and the green clusters points to growing interest in using phylogenetic and genomic data to inform conservation efforts. The blue cluster dynamics with other clusters reflect the manner in which abiotic drivers contribute to and respond to shifts in ecosystem processes.
3.1.6 Geographic scope and regional studies
Phrases like “South Africa,” “southern Europe,” and “China” are used to denote the geographic scope of the majority of studies that often are focused on regions most exposed to climate change and areas of high biodiversity hotspots. Knowledge of the local impact of climate change and conservation planning suitable to the local environment is dependent on these sites. For instance, studies analyze the relationship between land-use change and climate change and their combined effect on ecosystem services in southern Europe and South Africa, two highly rated territories for their biodiversity hotspots.
3.1.7 Emerging research trends
The network captures emergent patterns in climate change science, particularly increasing focus on human impact, conservation planning, and genetics. The application of the terms “genetics” and “conservation planning” signals increasing interest in using genetic studies to enhance species resilience. By marking emerging research fronts like socio-ecological perspectives, periphery nodes in the network underscore the need for integrating human activity, global climate models, and conservation policy.
3.2 “Floristic Tabuk” literature
Table 1 shows a broad summary of the key findings of 20 floristic diversity studies of Saudi Arabia’s Tabuk desert. Key findings of each study noted include records of plant species, dominant families and their ecological importance (with six papers outside Tabuk Province Supplementary Table 2). Sampling dates have also been included to address the time factor. It also indicates trends in the studies, for example, the preponderance of families like Asteraceae and Fabaceae and the prevalence of therophytes. Diversity is observed in species, methodology, and geographic location. The merging of 20 floristic diversity, vegetation structure, and ecological processes studies in various regions of Saudi Arabia thus provides novel valuable information on the biodiversity of this desert landscape. The table also refers to knowledge deficiencies, such as long-term biodiversity monitoring, genetic studies, and soil-plant interaction. The ideas put forward for the future are emphasized with a great requirement for interdisciplinary studies, targeted conservation action, and integration of emerging technology like DNA barcoding and remote sensing. This well-organized review that gives valuable information about the region’s biodiversity and its environmental challenges allows for successful conservation and sustainable management approaches.
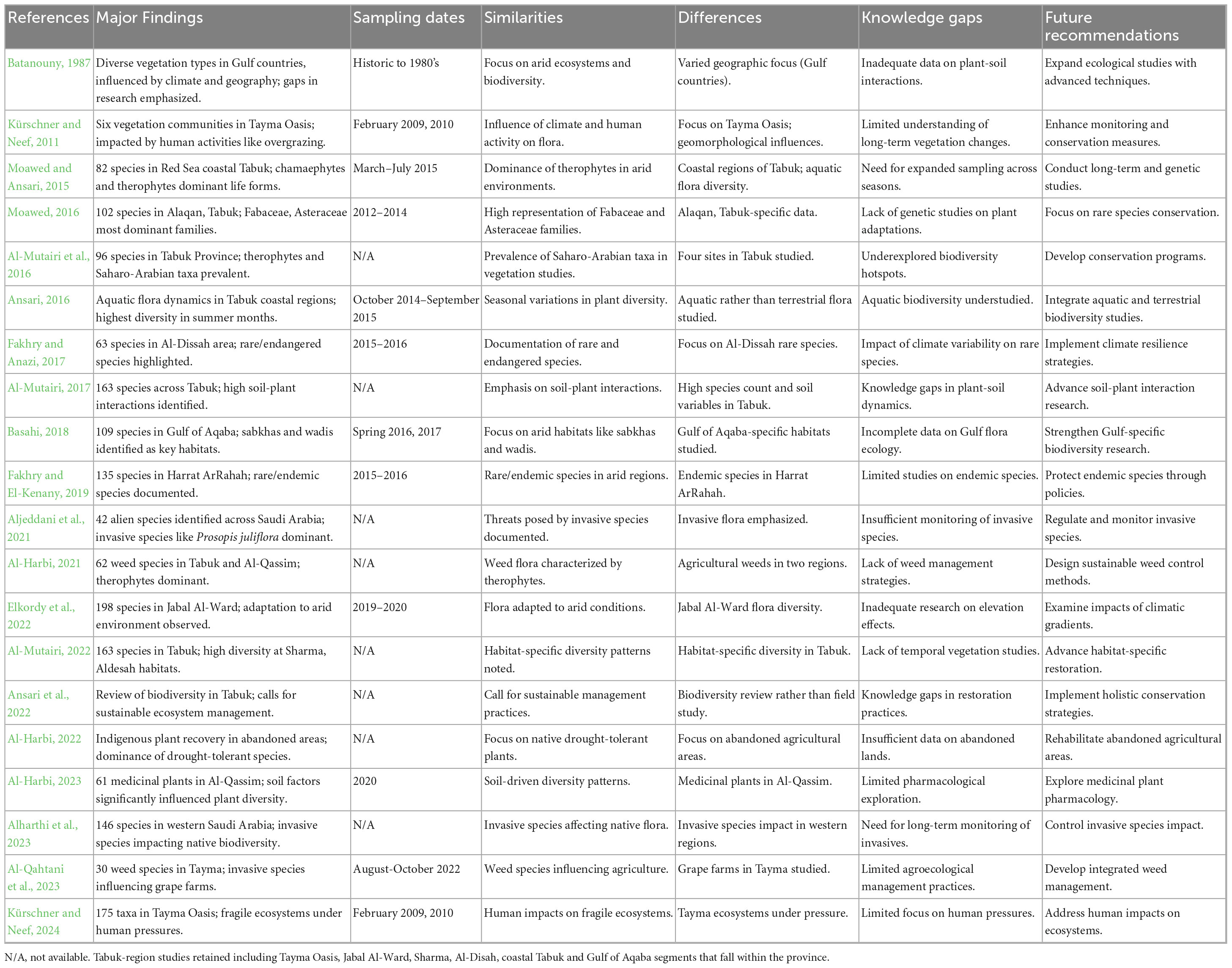
Table 1. Summary of major findings, sampling dates, similarities, differences, knowledge gaps, and future recommendations from 20 studies on floristic diversity in the Tabuk arid region, Saudi Arabia.
Based on the 26 (Table 1 and Supplementary Table 2) chosen papers, Figure 4 shows the occurrence of sampling dates in studies of floristic diversity in the Tabuk desert region of Saudi Arabia. Most of the research lacks specific sample date information, as represented by the “N/A” category with the most number of studies. Among studies with targeted sample periods, there were intensive sampling efforts during certain years or seasons, such as February 2009 and 2010, March to July 2015, and spring 2016 and 2017. There are also studies that cover historical records from the 1980’s to more recent years such as August–October 2022 and 2019–20. There needs to be greater systematic and periodic sampling to continue to study floristic dynamics in the region since this distribution reflects the variation of study activity with time.
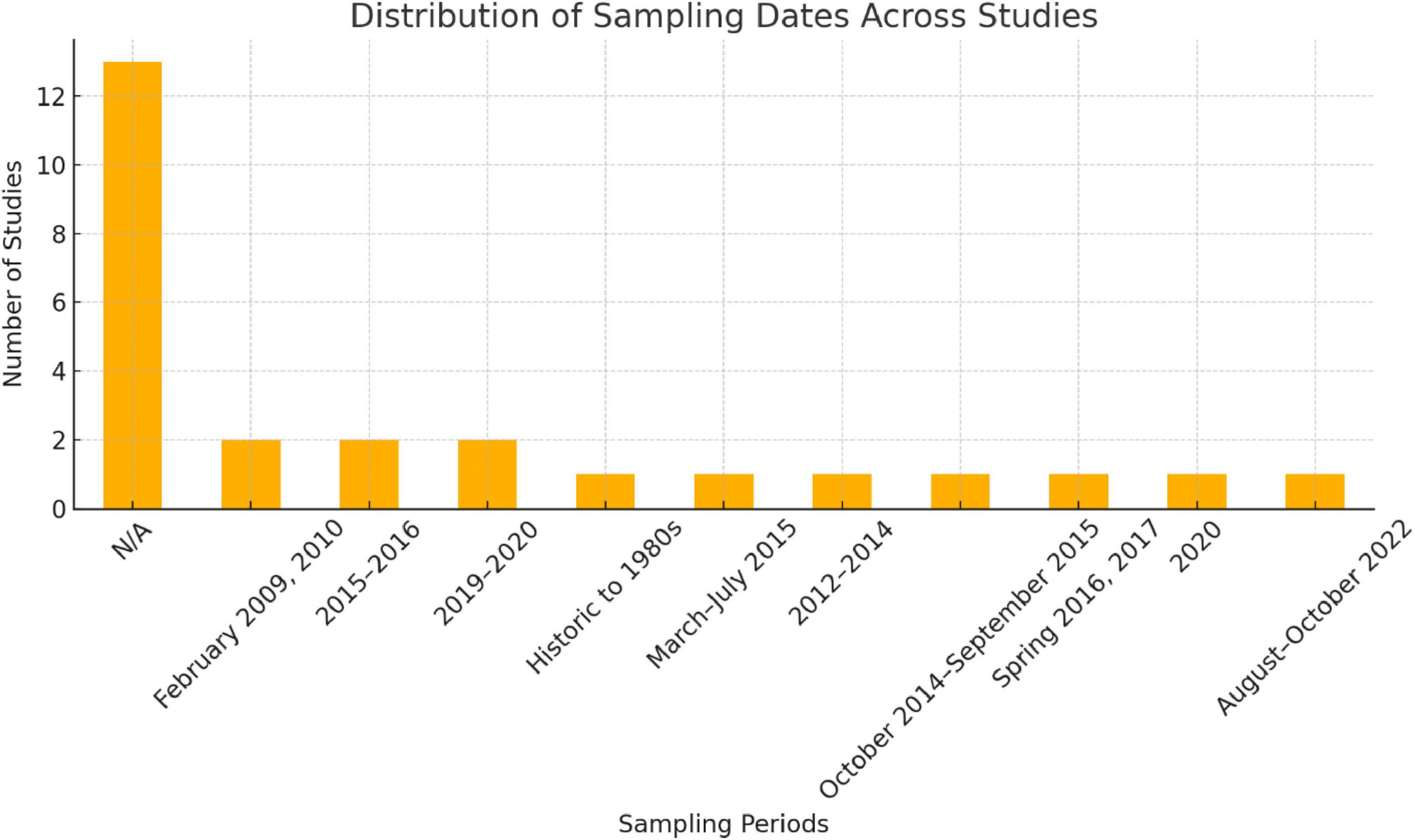
Figure 4. Distribution of sampling dates across studies on floristic diversity in the Tabuk arid region, Saudi Arabia.
3.2.1 Floristic diversity and dominant taxa
Collectively, the surveys yielded a total of almost 1,800 plant species and therefore the ecological diversity of the Tabuk desert region and its surroundings. Through the wide variety of life forms and phytogeographical relations of the species, they reflect the significance of the area in terms of biodiversity. These were established in various studies that the prevailing plant families like the Fabaceae, Poaceae, and Asteraceae possess a capacity to survive under arid conditions (Batanouny, 1987; Kürschner and Neef, 2011; Moawed and Ansari, 2015; Elkordy et al., 2022). Especially in deserts like those in Al-Qassim and Tabuk, therophytes and chamaephytes were the prevailing life forms (Al-Mutairi et al., 2016; Al-Harbi, 2023). Results explain how desert-adapted flora maintains ecological equilibrium and ecosystem functions like soil stabilization and carbon sequestration (Fakhry and Anazi, 2017; Alsobeai, 2024).
3.2.2 Spatial and temporal patterns
The study duration covered historical data like Batanouny (1987)’s and recent 2024 censuses (Youssef et al., 2024). The duration covered indicated the long-term processes of dynamics as well as the recent floristic dynamics of the Tabuk region. Research in such places as Tayma, Al-Fawzan Reserve, and Sharaan Nature Reserve recorded seasonality, where plant communities realized their highest diversity values during summer and spring (Kürschner and Neef, 2024; Youssef et al., 2024). Coastal locations like the Gulf of Aqaba, which had habitat-specific adaptation particularly in saline and wadi habitats, showed ecological resilience and complexity (Basahi, 2018; Aljedaani and Fakhry, 2023).
3.2.3 Ecological dynamics and human impact
Most cited as the main threats were urbanization, overgrazing, and invasive species; anthropogenic activities were cited as main stressors throughout the study. The invasive species P. juliflora were of most notable ecological effects, which included alteration of the soil’s properties to competitively outcompeting the native vegetation (Aljeddani et al., 2021; Alharthi et al., 2023). Tabuk and Rawdhat Al Masodi research indicates that chemical soil factors such as pH, potassium, and organic matter were reported to play a significant role in controlling the distribution of plants and composition of the community (Al-Mutairi, 2017; Al-Anazi and Al-Qahtani, 2018, 2019). By bringing into focus the intricate nexus between natural and human processes, our findings place the nexus in the limelight on the necessity for sustainable management practices (Al-Harbi, 2022).
3.2.4 Rare and endangered species
The literature reviewed revealed several rare and endangered species that are in need of urgent conservation. Two examples of the botanical rarity of the region include A. collenettiae, which occurs exclusively in Tabuk, and R. palaestinum, which is replicated near Harrat ArRahah (Fakhry and El-Kenany, 2019). Beyond acting as reservoirs of diversity, these species also serve as indicators of ecosystem health (Basahi, 2018; Al-Qahtani et al., 2023). Their vulnerability to habitat destruction and climate change especially in biodiversity hotspots such as Harrat ArRahah and Al-Fawzan Reserve suggests the importance of targeted conservation (Fakhry and El-Kenany, 2019).
3.2.5 Knowledge gaps and future directions
The research uncovered major knowledge gaps, including the importance of long-term biodiversity monitoring, genetic research to unravel mechanisms of plant adaptation, and more detailed research on the relationship between plants and soil (Moawed, 2016; Al-Mutairi, 2022). Emerging techniques like DNA barcoding and remote sensing were proposed to increase knowledge and documentation of plant diversity (Elkordy et al., 2022; Fakhry and Anazi, 2017). Maintenance of the ecosystem resilience depends on the management of invasive species and soil fertility improvement through managed restoration activities, as shown (Al-Harbi, 2022; Aljeddani et al., 2021). Future studies must therefore make it a priority to include multidisciplinary approaches if they are to support sustainable conservation policies and the Vision 2030 vision of Saudi Arabia (Ansari, 2016; Ansari et al., 2022). Taken together, these findings illustrate the ecological significance, problem, and biodiversity conservation potential of the Tabuk region through well-guided investigations and ventures. All references for the 20 articles have been provided to validate the results.
3.3 Khulais floristic diversity and recent change
The Khulais baseline of 251 accepted species and the decadal frequency shifts referenced in this study are derived from Alsherif and Almaghrabi’s survey (2011–2020) of the region’s flora (Alsherif and Almaghrabi, 2022). Their analysis, which coincided with declining precipitation and rising mean annual temperature, reported widespread reductions in species frequency with larger declines among agricultural weeds and valley taxa; rocky and mountainous taxa were comparatively stable. Only seven species increased in frequency, notably Prosopis juliflora and Trianthema portulacastrum.
To make our reuse of these published data auditable without duplicating the original inventory, we supply a single validation figure and concise summary checks. Figure 4 shows all sites referenced in our synthesis with habitat overlays and time windows, enabling visual verification of spatial coverage and habitat assignment. In our checks against the published list, all accepted binomials mapped unambiguously to families and life forms under the selected backbone; no changes were made to the original 251-species total. For transparency, we also report that the subset of species showing greater than ten percent frequency change (n = 50) falls primarily in agricultural and valley habitats, matching the original report’s pattern. These steps document the geo-referencing, habitat stratification, and taxonomic consistency of our analysis while respecting that the full inventory resides in Alsherif and Almaghrabi (2022).
4 Discussion
4.1 Climate change impacts on floristic diversity in arid ecosystems
Figure 5 shows the synergistic effects of climate change on desert ecosystems from current bibliographic research. The bibliometric study illustrates the alteration of the climatic status of the Khulais area. Alsherif and Almaghrabi (2022) reported alarming climatic changes in Saudi Arabia’s hyper-arid ecosystems, including a reduction of annual rainfall by as much as 85%, and an increase in temperature of 0.70 °C–1.30 °C. This trend is consistent with broader trends across the semi-arid and arid Middle East and North Africa (MENA) region, indicating similar studies have reported record-low precipitation trends and persistent warming trends (Sillmann et al., 2013; Zittis et al., 2022; Lelieveld et al., 2016). These trends are reinforcing aridity and are further stressing already vulnerable ecosystems. The sudden drop in rainfall coupled with rising temperatures not just threatens native biodiversity but also intensifies water shortages, desertification, and conventional agriculture—making this a pressing agenda both for environmental sustainability and climate resilience in the region. These findings also validate the blue cluster of the bibliometric network, where the contributions made by abiotic factors such as soil quality, temperature, and water availability to ecosystem processes are featured. The green cluster illustrating habitats and species richness indicates the central role of water availability for biodiversity, corroborated by the lowered soil moisture in Khulais having inhibited native plant germination and seedling survival.
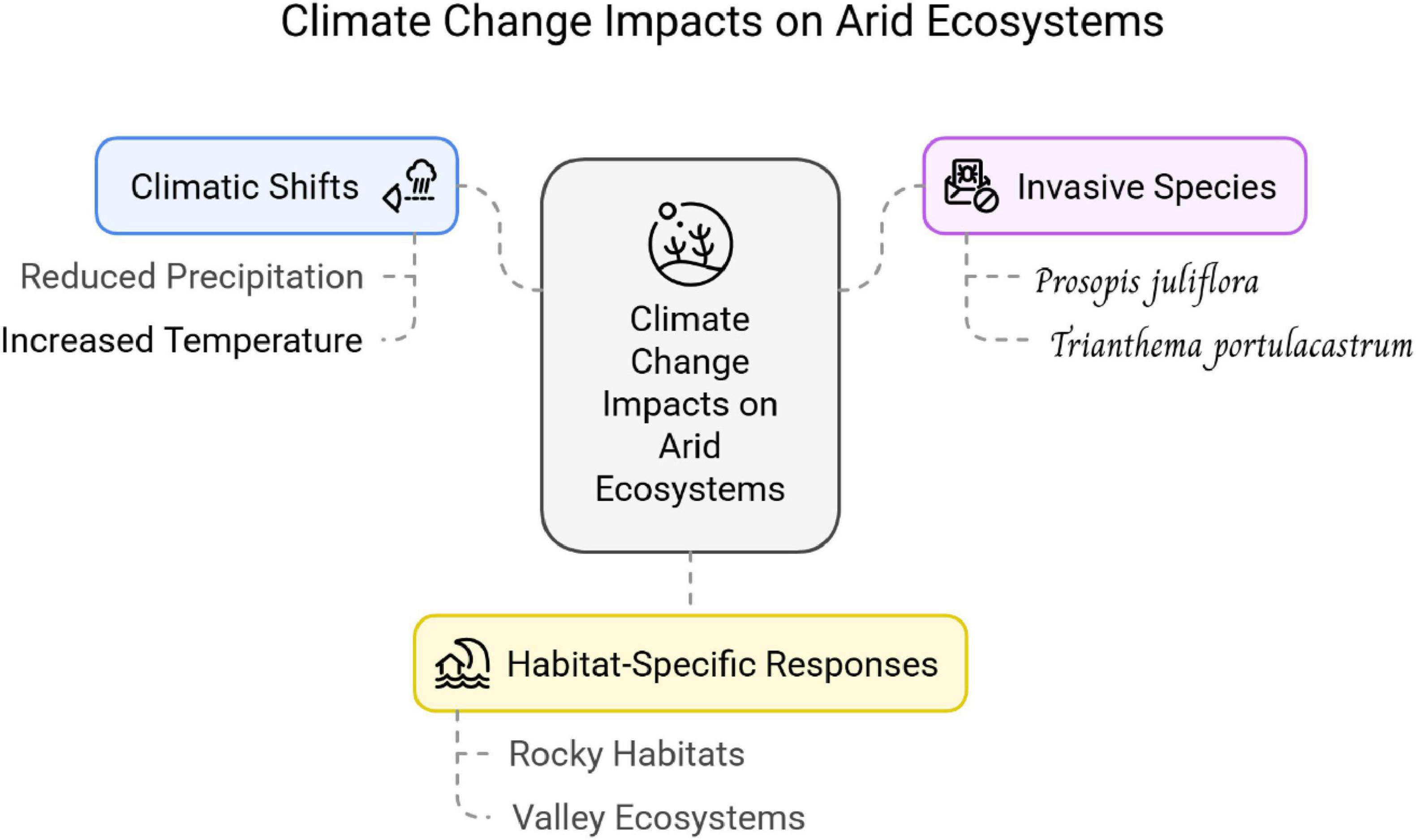
Figure 5. The overall climate change impacts on arid ecosystems based on present bibliographic analysis.
The global trends in the red cluster, which highlight vegetation shift and competition by invasive species, reflect the proliferation of invasive species like P. juliflora and T. portulacastrum in disturbed communities. Invasive species displace native vegetation through monopolization of limited resources, lowering diversity, and redefining ecological processes. The bibliometric synopsis indicates that invasive species are flourishing due to synergistic interactions between climate change and anthropogenic activities, thus highlighting the inter-linked nature of the two problems. Trends in biodiversity loss in regions such as the South West Australian Floristic Region (Yates et al., 2010) bear witness to the extent of such problems across the globe and the imperative of holistic management of dry ecosystems. The results thus serve to underscore even more the significance of habitat-specific adaptations in face of constraint from climate change.
Harsh environments are more robust than those of valleys due to their characteristic microclimatic conditions—e.g., reduced temperatures and restricted access to water—that assist in protecting against extreme aridity. The findings also endorse the purple cluster of the network, with emphasis on genetic and phylogenetic richness in enabling survival of the plants under environmental stress (Bakker et al., 2024; Belyazid et al., 2019). The rock ecosystem resilience developed requires concerted conservation efforts. While valley interventions are concerned about artificial fillings of water and stabilizing land, rock communities may be assisted by conserving microhabitats to facilitate typical plant communities (Schuchardt et al., 2023; Sekar et al., 2024).
The bibliometric evaluation justifies complicated conservation efforts by aligning with research from the South West Australian Floristic Region (Yates et al., 2010) and Romanian Carpathians (Stoica et al., 2017). Focusing on habitat adaptation strategies to enhance conservation effectiveness, this research offers viable methods for tackling the challenges posed by climate change in desert ecosystems.
4.2 Floristic diversity and planetary health in Saudi Arabia
Figure 6 illustrates the overall idea of Saudi Arabia’s floristic diversity and its functions according to the current bibliographic analysis. The floristic diversity of Saudi Arabia is comprised of a wide variety of plant species owing to the geographical and ecological distinctness of the country. The 27 studies reviewed have recorded over 1,800 species that belong to various families, life forms, and phytogeographical affinities. Some of the major families like Fabaceae, Poaceae, and Asteraceae are extremely adaptable to the prevalent dry and semi-arid conditions of the country (Batanouny, 1987; Kürschner and Neef, 2011; Moawed and Ansari, 2015). The upkeep of the ecological balance and wellbeing of the earth, especially in ecologically stressed areas, relies on ecosystem services such as soil stabilization, carbon sequestration, and provision of wildlife habitat, all thanks to these plants (Yang et al., 2021).
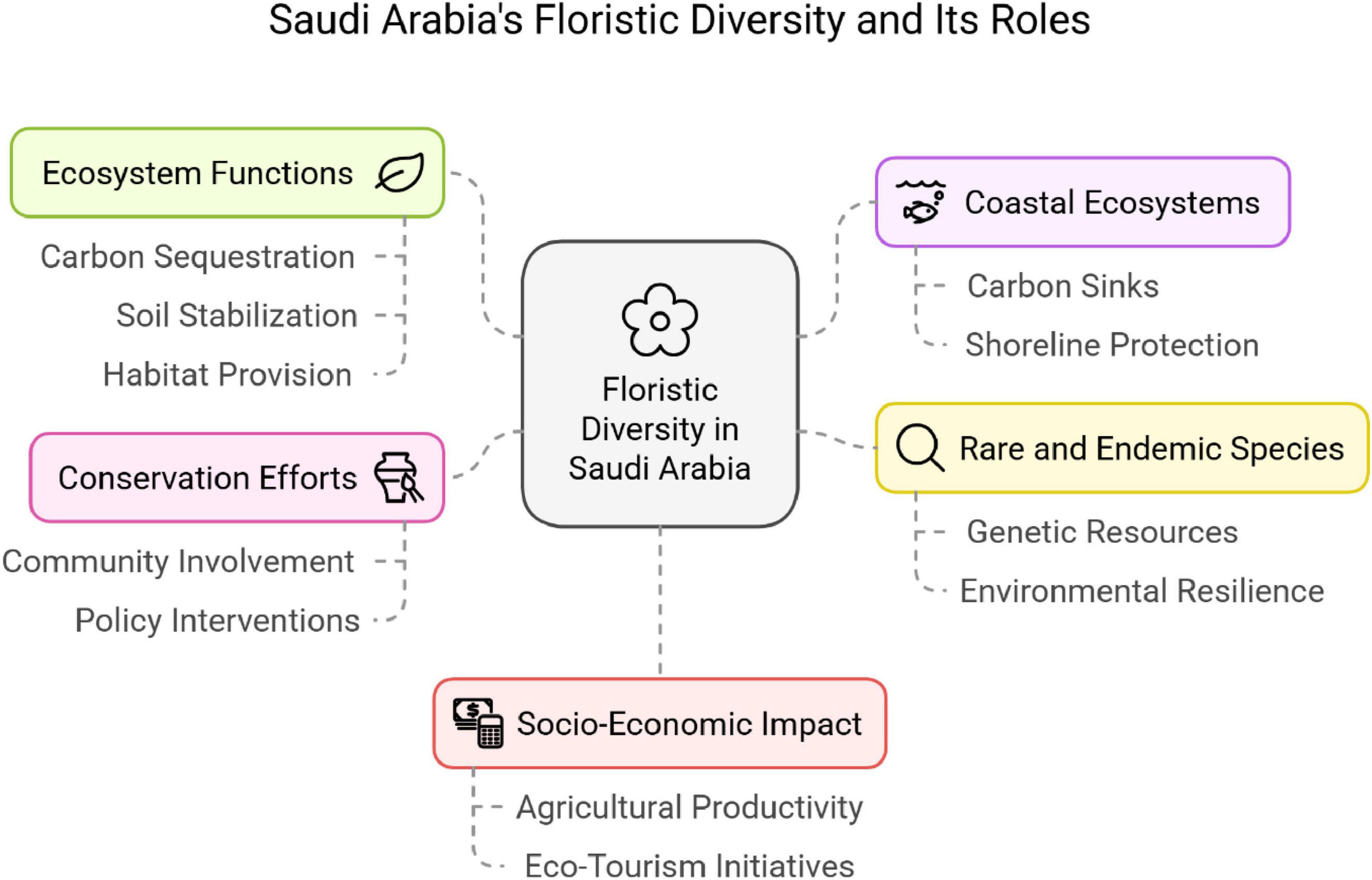
Figure 6. The overall concept of Saudi Arabia’s floristic diversity and its roles based on present bibliographic analysis.
Vegetation adaptation to extreme conditions, i.e., aridity and salt land, in areas such as Tabuk, Sharma, and Aldesah represents value to biodiversity to environmental sustainability. The coastal areas of the Red Sea and Aqaba Gulf, which are characterized by mangrove and halophyte covers, constitute an important ecological value. Coastal ecosystems are natural sinks of carbon because they fix atmospheric carbon dioxide, thereby mitigating the effects of climate change. They also save coasts from erosion and enable the stabilization of marine systems that are crucial for the sustainability of biodiversity and the economic viability of fisheries (Basahi, 2018; Moawed and Ansari, 2015). Habitats such as Harrat ArRahah and Al-Fawzan Reserve experience floristic diversity of significant magnitude, including endemic and rare species such as R. palaestinum and A. collenettiae.
In addition to being a vital ecological asset, these species also harbor valuable genetic resources for environmental adaptation in the future. They must be conserved so that ecosystem resilience and genetic diversity are maintained and these ecosystems can be used as natural buffers to environmental degradation (Alsobeai, 2024). Successful conservation involves bringing together ecological research, community involvement, and policy intervention. The study aims at floral diversity as the driver of social and economic sustainability. Healthy plant species influence agricultural productivity, food security, livelihood, and ecosystem stability.
The integration of local knowledge with sustainable agriculture and ecotourism activities is one method of reconciling conservation objectives with socioeconomic objectives. Ecotourism based on endemic flora can help to generate revenues among locals and foster a culture of conservation (Whitmee et al., 2015; Hautier et al., 2018). The variety of uses of floristic diversity makes it a fundamental driver to regional sustainability and also human health globally.
4.3 Hydrological interventions and their ecological impacts in the floristic diversity
Figure 7 presents the general ecological impacts of hydrological interventions according to existing bibliographic research. Hydrological alterations—like damming—have been found to have positive environmental effects in riparian zones.
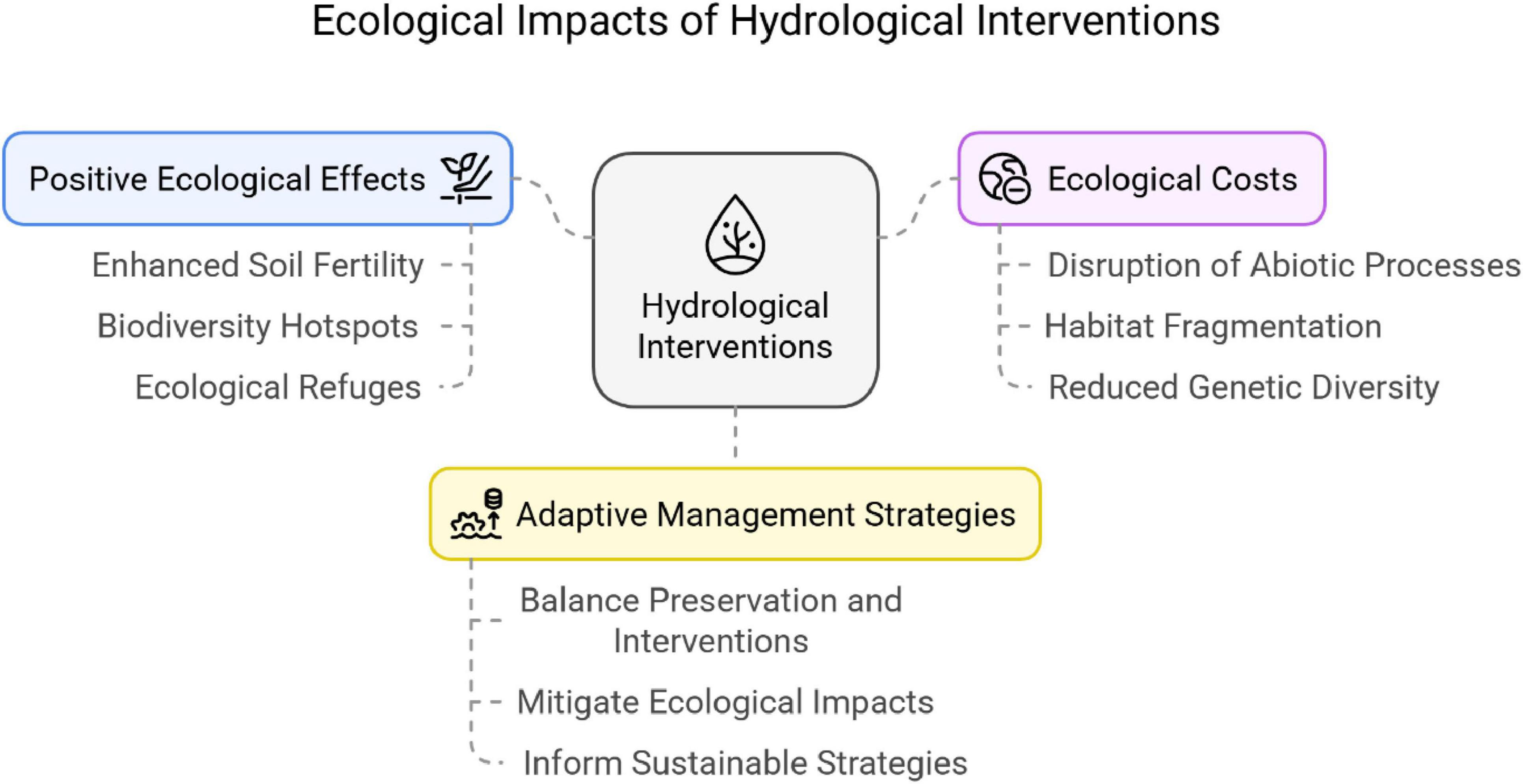
Figure 7. The overall ecological impacts of hydrological interventions based on present bibliographic analysis.
Research by Al-Munqedhi et al. (2022) demonstrates that repeated inundation and fertilizer input in such a setting enhance soil fertility and trigger higher species richness. These findings are consistent with the green cluster of the bibliometric analysis, which validates the importance of ecosystems and biodiversity conservation. Riparian zones in arid environments are hotspots of biodiversity due to the fact that they create essential habitats for water-dependent vegetation. The vegetation response within this zone, such as the formation of six new vegetation associations, illustrates the capacity of vegetation to acclimatize to altered hydrological regimens as shown in the red cluster on vegetation dynamics (Pütz et al., 2016; Renny et al., 2022).
These regions also provide environmental refuges that preserve essential ecosystem processes including nitrogen cycling and soil stabilization. Bibliometric analysis illustrates more clearly how they facilitate ecological connectivity, which is fundamental to species mobility and adaptation to changing climate conditions. Impounded water systems can potentially promote diversity in arid areas in line with international research that identifies habitat-based conservation initiatives.
But one must never overlook the environmental consequences of building dams. The blue cluster shows how dams disrupt abiotic processes naturally. Regulating flood regimes, for example, alters the propagule abundance of vegetation and hence reduces genetic heterogeneity and increases local extinctions. Al-Munqedhi et al. (2022) contend that altered patterns of water flow below degrade the biological integrity of affected ecosystems through soil erosion and nutrient removal.
Red and purple clusters indicate cascading impacts of habitat fragmentation and low genetic diversity that compromise ecosystem resilience at long-term large-scale levels. This is in line with worldwide patterns attained through the bibliometric analysis, where human action like damming enhances ecosystem vulnerability to climate change stress. Adaptive management strategies balancing hydrological intervention and ecological conservation are TOTALLY necessary if one is to mitigate such impacts (Braz Pires et al., 2024; Dullinger et al., 2020). The crossing point of habitat-specific resilience, hydrological intervention, and climatic variability points to the intricacy of biodiversity management in arid ecosystems. Case studies at the regional and bibliometric studies give a perfect understanding of these processes that inform sustainable ecosystem management strategies to respond to escalating environmental change (Blaus et al., 2021; Bordoloi et al., 2023).
4.4 Anthropogenic threats and ecological challenges in Saudi Arabia
4.4.1 Principal anthropogenic and climatic pressures
Figure 8 illustrates the principal threats to biodiversity and conservation in Saudi Arabia derived from the present bibliographic analysis. Overgrazing, agricultural expansion, urbanization, and alien species are the dominant human induced pressures. Urbanization has fragmented habitats and reduced local biodiversity, which is particularly evident in coastal counties along the Red Sea and in Jeddah. Industrial activities further degrade native vegetation by introducing pollutants that alter water and soil chemistry (Aljedaani and Fakhry, 2023; Al-Harbi, 2021, 2023). Habitat destruction is especially concerning in sensitive ecosystems such as the Gulf of Aqaba, where plants already persist under saline and hyper arid stressors (Basahi, 2018).
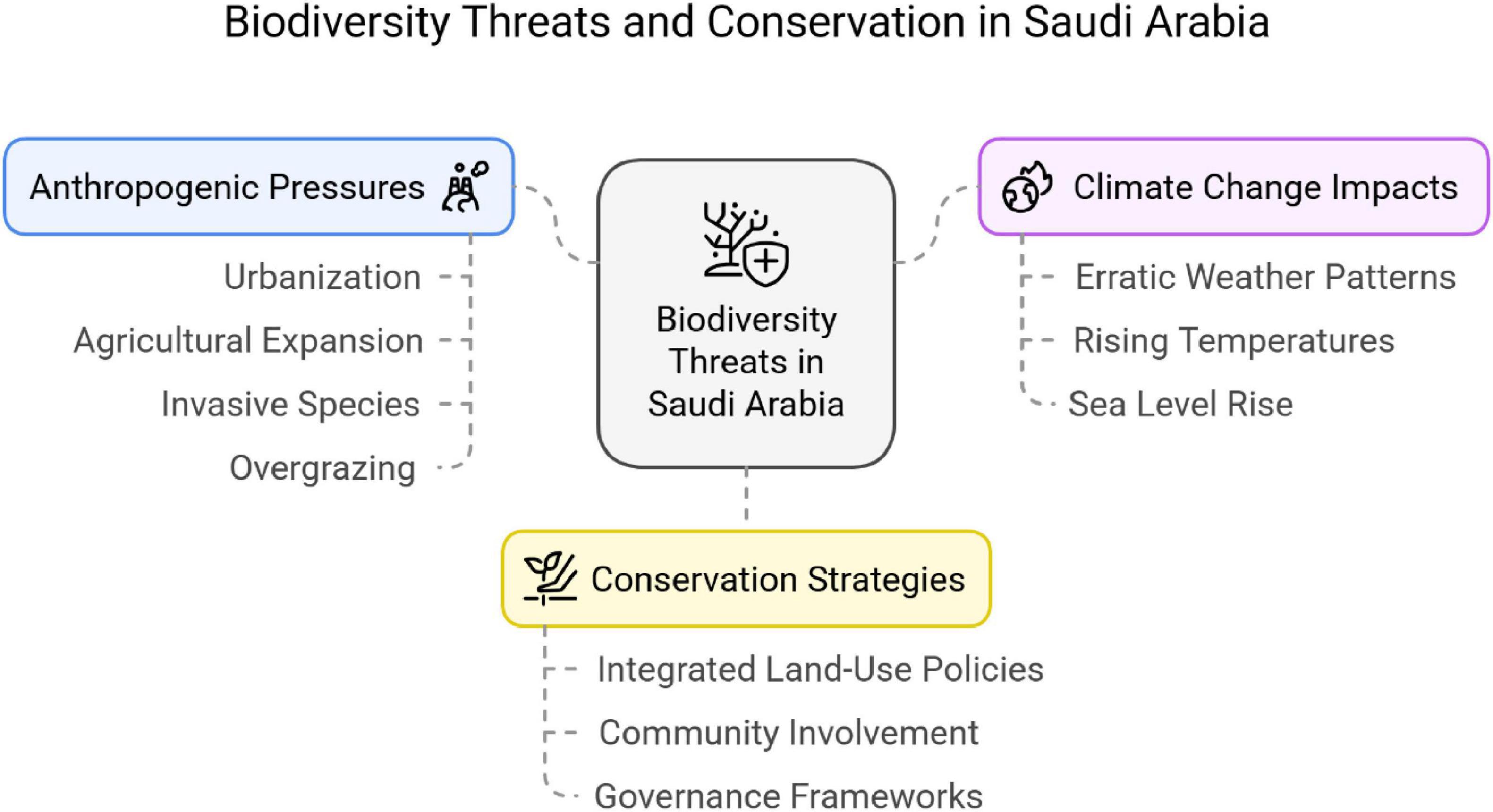
Figure 8. The overall biodiversity threats and conservation in Saudi Arabia based on present bibliographic analysis.
Invasive weeds such as P. juliflora reduce species diversity and compete strongly with native plants. Studies in agricultural fields at Tayma document how invasive weeds compete with crops, lower yields, and threaten food security (Al-Qahtani et al., 2023). Overgrazing by livestock further reduces plant cover, accelerates soil erosion, and weakens the capacity of desert ecosystems to maintain biodiversity.
Climate change compounds these pressures through altered rainfall regimes, rising temperatures, and more frequent extremes. These changes destabilize arid ecosystem processes and increase vulnerability to biological invasions and habitat loss. Coastal exploitation and sea level rise in Sharma threaten both terrestrial and aquatic biodiversity along the shoreline (Alharthi et al., 2020; Youssef et al., 2024).
Mitigating these impacts requires integrated land use planning that balances development with ecological safeguards and food security goals (Garcés-Pastor et al., 2022; Huang and Fu, 2023). Policy should prioritize sustainable land use, invasive species management, and climate smart agricultural practices, supported by community participation. Empowering local communities to co-manage landscapes can translate knowledge into sustainable action. Such whole of society programmes, grounded in public health and planetary health perspectives, can help secure Saudi Arabia’s unique biodiversity (Whitmee et al., 2015).
4.4.2 Floristic comparison of Tabuk and Khulais arid regions
Floristic comparison between regions of Tabuk and Khulais draws on species richness, numbers of genera and families, dominant families, life forms, and phytogeographical affinities. Reported species richness for Tabuk ranges from 96 to 198 species, reflecting differences in sampled area, methods, and ecological status across studies (Al-Mutairi et al., 2012, 2016; Al-Mutairi, 2022; Elkordy et al., 2022). Khulais region shows relatively higher and more balanced floristic diversity with 251 species recorded in a regional review (Alsherif and Almaghrabi, 2022). In Tabuk, the flora spans 75–124 genera and 38–47 families (Al-Mutairi et al., 2016; Al-Mutairi, 2022; Elkordy et al., 2022). Khulais comprises 160 genera in 50 families (Alsherif and Almaghrabi, 2022).
Dominant families overlap across both regions and include Asteraceae, Fabaceae, and Poaceae, which are consistently prominent in Tabuk (Al-Mutairi et al., 2012, 2016; Al-Mutairi, 2022; Elkordy et al., 2022). Khulais also shows strong representation of Poaceae, Papilionaceae, Euphorbiaceae, and Asteraceae, indicating a slightly more diversified dominance pattern (Alsherif and Almaghrabi, 2022). Life form spectra in both regions are dominated by therophytes. In Tabuk, therophytes constitute 37.0%–46.5% of species, followed by chamaephytes at 26.0%–32.0%, hemicryptophytes at 10.0%–13.0%, phanerophytes at 8.00%–10.0%, and geophytes at 6.90%–13.0% (Al-Mutairi et al., 2016; Elkordy et al., 2022). Khulais shows comparable proportions with therophytes at 41.2%, chamaephytes at 31.4%, hemicryptophytes at 13.7%, and phanerophytes at 10.0%, with geophytes least represented (Alsherif and Almaghrabi, 2022).
Phytogeographical composition also displays overlap with differences in proportions. Tabuk is dominated by Saharo Arabian elements at about 37.2%–38.5%, with Irano Turanian at 9.10%–11.6% and Sudanean at 10.5%–12.6% (Al-Mutairi et al., 2016; Elkordy et al., 2022). Khulais shows a combined Saharo Arabian and Sudanian representation of 43.6% (Alsherif and Almaghrabi, 2022). These patterns reflect the transitional biogeographic setting between African, Arabian, and Asian floras.
Overall, Khulais appears to hold greater species richness and taxonomic breadth than Tabuk while sharing arid adapted strategies. The diversity in Khulais likely reflects topographic variety, microhabitat heterogeneity, and possibly lower local disturbance in some sites. Both regions remain important reservoirs of desert adapted biodiversity and require region specific conservation plans. Indicator and endemic species are especially informative. Species such as A. collenettiae in Tabuk and R. palaestinum in Harrat ArRahah embody unique genetic resources and may be highly sensitive to habitat degradation and warming (Williams et al., 2021; Wróblewska and Mirski, 2018; Xia et al., 2022). Loss of such species not only reduces local diversity but also disrupts ecological processes including soil stabilization, nutrient cycling, and support for pollinators and other fauna (Yao et al., 2021; Zeng et al., 2016; Hautier et al., 2018). Conservation responses should include the protection of biodiversity hotspots such as Harrat ArRahah and Sharma, targeted habitat restoration, and research on physiological and genetic adaptation in rare and endemic taxa to guide management in comparable arid systems (Trisurat et al., 2023; Tyler et al., 2018; Schuldt et al., 2019).
Invasive and disturbance tolerant species that expand under aridification should be tracked explicitly. In Khulais there is evidence of increasing frequency of P. juliflora and T. portulacastrum in disturbed and agricultural contexts, which aligns with the broader pressures summarized above and reinforces the need for valley restoration, invasive species control, and careful water resource stewardship.
4.4.3 Global comparison and implications for Saudi arid floras
Placing Tabuk and Khulais in a wider arid land context shows both convergence and regional particularity. Tabuk reports 96 species from 75 genera and 38 families with strong representation of Asteraceae, Fabaceae, Zygophyllaceae, and Lamiaceae, and a life form spectrum dominated by therophytes and chamaephytes. The flora is largely Saharo Arabian with notable Irano Turanian and Sudanean elements and is increasingly affected by cutting of wood and urban expansion (Al-Mutairi et al., 2016; Van Eck and Waltman, 2017). Khulais has 251 species in 160 genera and 50 families with prominent Poaceae, Papilionaceae, Euphorbiaceae, and Asteraceae and similar life form dominance, confirming drought escape and drought resistance strategies in both regions (Alsherif and Almaghrabi, 2022).
Across other deserts, similar families dominate but richness and endemism vary widely. Middle Asia including Kazakhstan, Kyrgyzstan, and Uzbekistan supports 9,643 vascular taxa in 1,198 genera and 139 families, with endemism reaching 35.4% and strong dominance of Poaceae and Asteraceae, which mirrors Saudi patterns at family level while far exceeding them in absolute richness (Ma et al., 2024). The Irano Turanian province is one of the world’s largest arid floristic regions and a cradle of domestication with complex evolutionary history and high endemism, extending influence into northern Saudi Arabia (Manafzadeh et al., 2017; Lauterbach et al., 2019). In the Western Hemisphere, the coastal deserts of Peru hold around 215 plant species with richness increasing upslope and with notable endemism and threat levels, again demonstrating the ecological value and vulnerability of coastal and montane arid systems (Montenegro-Hoyos et al., 2022).
These comparisons underscore two messages for Saudi arid floras. First, functional convergence around annuals and dwarf shrubs reflects adaptation to water limitation and disturbance. Second, regional uniqueness in species pools and endemism requires place based conservation instruments. For Tabuk and Khulais this means climate aware hydrological stewardship in wadis and valleys, protection of rocky and coastal refugia, invasive species management with emphasis on P. juliflora and T. portulacastrum, and sustained biodiversity and genetic baselining that can support adaptive management under a warming and more variable climate.
4.5 Global comparison of floristic diversity between arid regions from Tabuk and Khulais, and tropical regions
To draw a comparison on the floristic diversity of tropical and desert habitats (Tabuk and Khulais), the composition of plant species, diversity indices, and environmental determinants under contrasting climatic conditions need to be considered. Desert ecosystems, such as in Saudi Arabia, have a different ecological setting compared to that of tropical rainforests and, therefore, have apparent differences in floral diversity and structure.
In the Tabuk desert area, 96 species of plants belonging to 75 genera and 38 families have been recorded. The most dominant families are Asteraceae, Fabaceae, Zygophyllaceae, and Lamiaceae (Al-Mutairi et al., 2016). The life form analysis shows the dominance of therophytes (37%) and chamaephytes (32%), an adaptation strategy to seasonal and xeric conditions. Its chorological composition is overwhelmingly Saharo-Arabian (37.21%), Irano-Turanian (11.63%), and Sudanean (10.47%) (Al-Mutairi et al., 2016). Moreover, β-diversity metrics exhibit high species turnover among habitats (Al-Mutairi et al., 2016; Van Eck and Waltman, 2017).
In contrast, the Khulais region has greater floristic richness, with 251 species in 160 genera and 50 families. The important contributing families include Poaceae, Papilionaceae, Euphorbiaceae, and Asteraceae (Alsherif and Almaghrabi, 2022). The classification of life-forms reveals that therophytes (41.2%) and chamaephytes (31.4%) dominate the flora, as in Tabuk. However, in Khulais, there is a wider representation of Saharo-Arabian and Sudanian chorological elements, which make up 43.6% of the flora (Alsherif and Almaghrabi, 2022).
As compared to these arid environments, tropical ecosystems such as Southeast Asia, the Neotropics, and tropical Africa are globally renowned for being exceedingly species-diverse. Latin America, for instance, boasts over 118,000 plant species (Raven et al., 2020). These ecosystems have an abundance of phanerophytes (trees and tall shrubs) due to the favorable climatic conditions of high rainfall and year-round warmth (Fine, 2015). The tropical floras generally contain a mixture of ancient and recently diversified lineages with strong Indo-Malaysian, Neotropical, and Afrotropical biogeographical elements (Fine, 2015; Hua, 2013).
When comparing the arid and tropical floristic profiles, a number of differences are evident. The arid areas possess limited species richness (96–251 species) and a life-form spectrum dominated by drought-tolerant chamaephytes and ephemeral therophytes. On the other hand, tropical areas have very high α and β diversity, partly caused by ecological heterogeneity, stability of the historical biome, and reduced extinction (Fine, 2015). Highly disparate geological and evolutionary backgrounds in relation to arid worlds are also the foundation for the diversity of tropical floras (Hua, 2013). Environmental forces are the predominant causes of such trends. Climatic stressors, soil, and anthropogenic disturbance such as woodcutting and urbanization govern the spatial pattern of vegetation in arid environments (Al-Mutairi et al., 2016; Van Eck and Waltman, 2017). Biodiversity is promoted in the tropical environments, however, due to long-term climatic consistency that allows speciation to surpass extinction and the establishment of complex ecological networks (Fine, 2015; Raven et al., 2020).
Generally, arid environments like Tabuk and Khulais share floristic rarity of species adapted to stressful environmental conditions. Tropical environments have unmatched species richness due to preferential environmental conditions and complex evolutionary history. Species composition differences, life forms, and chorological affinities emphasize the ecological uniqueness and conservation values of both arid and tropical ecosystems.
Because it functions as a critical bridge between terrestrial and marine ecosystems, the region around the Red Sea is critical to global health. The halophytes and mangroves in the region’s coastal flora provide an assemblage of ecosystem services upon which terrestrial and marine biodiversity are based. They provide shoreline stabilization, control the entry of nutrients into marine ecosystems, and carbon sequestration (Alharthi et al., 2020; Basahi, 2018). Degradation of these ecosystems by urbanization, sea level rise, or other human-induced factors can threaten fish stocks, coral reefs, and other Red Sea biodiversity hotspots (Jewitt et al., 2017; Kougioumoutzis et al., 2020). The interdependence of these systems underscores the necessity of joint conservation action. The health of marine ecosystems and terrestrial biodiversity depends on conservation of coastal ecosystems in Sharma and beyond. The ecosystems provide livelihoods and food security to numerous local communities, and therefore the conservation efforts need to include the socioeconomic consequences of biodiversity loss (Whitmee et al., 2015; Imanberdieva et al., 2018; Inague et al., 2021).
4.6 Knowledge gaps and future research directions
Figure 9 shows the overall concept of climate resilience of ecosystems based on the existing bibliographic study. In spite of the large body of knowledge gained, there are major information gaps in areas such as genetic studies, long-term biodiversity monitoring, and soil-plant interactions. Modern techniques such as remote sensing and DNA barcoding can potentially reveal dry plants’ ecological functions and evolutionary processes (El-Sheikh et al., 2019; Schuldt et al., 2019). Research in the future must also investigate the climate change impacts on endangered species and effective habitat restoration practices (Fakhry and Anazi, 2017; Tang et al., 2019; Trew et al., 2022). Elucidating the intricate interactions among these ecosystems requires synthesis between land and sea research. These holistic approaches can inform conservation interventions aligned with Saudi Arabia’s Vision 2030 goals as well as support sustainable ecosystem management and enhance global biodiversity and planetary health (Hautier et al., 2018; Ssali et al., 2023; Soliani et al., 2024). Some concepts to fill the knowledge gaps for future research are as follows:
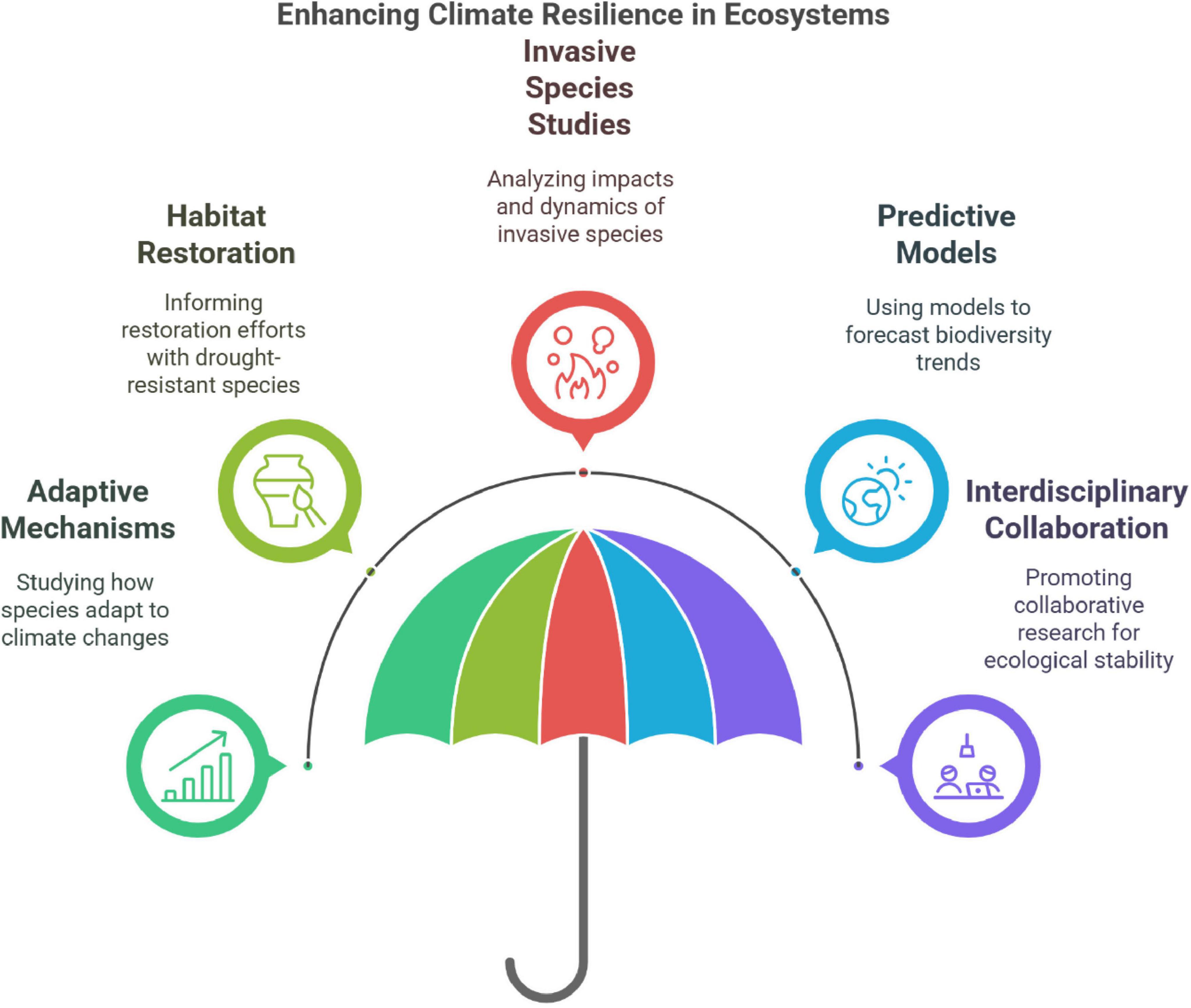
Figure 9. The overall concept of enhancing climate resilience in ecosystems based on present bibliographic analysis.
(a) Intensive monitoring systems
The bibliometric study suggests the need for intensive, long-term monitoring systems for biodiversity in drylands. The blue and green clusters reflect how new technologies such as satellite imaging, remote sensing, and Geographic Information Systems (GIS) might track the occurrence of invasive species, soil condition, and vegetation cover. Terms like “habitat dynamics” and “species richness” in the VOS study highlight the requirement of monitoring changes in ecosystems at different scales (Sun et al., 2021; Suggitt et al., 2019). Soil and water quality monitoring and ground-based biodiversity monitoring can also provide localized data apart from satellite-based monitoring. Long-term analysis of such records provide predictive insights on ecosystem responses to climate fluctuations and hydrological manipulations as well as biodiversity trends. Scaling up such monitoring and dealing with the complexities of ecosystems like valley, riparian, and rocky areas would need coordination among academia, governments, and local communities (Lu et al., 2021; Nabhan et al., 2022).
(b) Policy making
The red and purple clusters of the map indicate the important connection between policymaking and biodiversity preservation. Restoring valley ecosystems and preserving riparian zones are two habitat-related measures that should be the main priority of sound policy. Offering farmers and landowners financial and technical support encourages sustainable practices such as the restoration of indigenous vegetation and water-saving irrigation systems.
In order to manage invasive plants like P. juliflora, policy must commence concerted eradication, biosecurity, and early detection. The emphasis of the red cluster on worldwide ecosystems provides an example of how cross-border collaboration can enhance the sharing of resources and strategy development. Also, if conservation goals are integrated with broader development initiatives, biodiversity preservation harmonizes infrastructural development and economic growth (Ordonez and Svenning, 2017; Ortega et al., 2024).
(c) Community engagement
Investigation of the blue and green clusters identifies a crucial role for community engagement to realize sustainable conservation achievements. Local stakeholder monitoring programs, where local people document information on invasive species and species richness, can improve the coverage and efficacy of conservation action. Educational programs highlighting the need for native biodiversity and the dangers facing alien species can engender a sense of ownership among local people.
Locally driven restoration activities such as reforestation and soil stabilization yield cost-efficient advantages in the form of eco-tourism and job creation. Concerting conservation with actual community advantage brings sustainability to participation and long-term involvement toward biodiversity conservation.
(d) Innovative water management
The focus of the blue cluster on hydrodynamics of arid regions serves to underscore the compelling necessity for new water management technologies for dry regions. Regulated aquifer recharge, drip irrigation, and rainwater harvesting are sustainable practices that minimize the environmental impact and help alleviate the problem of water scarcity. The VOS network uses terms such as “riparian zones” and “water efficiency” to underscore the significance of water management in biodiversity and crop protection.
Through enhancing water efficiency in agriculture, drip irrigation systems guarantee riparian ecosystems are conserved. Similarly, managed aquifer recharge can store surplus water during wet seasons, thereby acting as a cushion against droughts. Integrating these technologies into conservation programs will guarantee long-term sustainability of arid ecosystems through an equilibrium of ecological preservation and human water demands (Peralta et al., 2024; Phelan et al., 2016). The blue cluster’s focus on hydrological dynamics underscores the need for new thinking to dryland water management. Regulated aquifer recharge, drip irrigation, and rainwater harvesting are sustainable practices that minimize environmental footprint and ease water scarcity. In the VOS network, words such as “riparian zones” and “water efficiency” underscore the significance of water management for biodiversity conservation and crop yields. By enhancing agricultural water use efficiency, drip irrigation systems avert riparian ecosystem degradation. Likewise, managed aquifer recharge can store surplus water during times of overabundant rainfall, hence acting as a drought buffer. Conservation programs involving such technologies will guarantee the long-term sustainability of arid ecosystems by ensuring ecological conservation balanced with human water needs (Peralta et al., 2024; Phelan et al., 2016).
(e) Research on climate resilience
The green and red clusters emphasize the importance of performing detailed studies on the way native and invasive species evolve to respond to rising temperatures. An understanding of the physiological and ecological traits enabling species to endure in desert environments can help habitat restoration processes like the selection of drought-resistant native flora for replanting (Ossa et al., 2017; Peng et al., 2023).
Extensive research on P. juliflora and other alien invasive species will be used to elucidate their ecological impacts, competitive interactions, and vulnerabilities. Conservation efforts may be informed through research on the interactions between alien and native biota to enable re-establishment of balance in the ecosystem. The use of predictive models that integrate climatic projections allows one to prioritize conservation efforts with utmost priority and forecast biodiversity patterns, as shown in the blue cluster. Climate resilience enhancement will be dependent on concerted, inter-disciplinary research.
4.7 SWOT analysis of floristic diversity of Saudi Arabia arid ecosystem
A SWOT analysis (Strengths, Weaknesses, Opportunities, Threats) provides a structured evaluation of the status of floristic diversity within arid ecosystems, considering their strengths and weaknesses and opportunities and threats facing them. Figure 10 displays the whole SWOT analysis of floristic diversity of Saudi Arabia ecosystem.

Figure 10. The overall SWOT (Strengths, Weaknesses, Opportunities, and Threats) analysis of floristic diversity in Saudi Arabia ecosystem.
(a) Strengths
Deserts in Saudi Arabia have resistant environments due to drought-resistant plant species and adaptive adaptations (Batanouny, 1987; Moawed and Ansari, 2015). The environments play crucial roles in genetic diversity and ecosystem resilience, with around 1,800 species, including endemic and endangered plants (Alsobeai, 2024). In African savannas that are semi-arid, gradients in terrain and soils regulate the distribution of woody plants, leading to species that are satisfied with survival under harsh conditions (Witkowski and O’Connor, 1996). Dry South American environments like Patagonia have floristic richness backed by adaptation processes that facilitate species to survive under grazing and aridity (Oliva et al., 2016). Red Sea and Gulf of Aqaba coastlines are carbon reservoirs where carbon is sequestered and integrity of the coast is ensured (Basahi, 2018). Desert rangelands of Tunisia sustain grazing- and drought-resistant plant communities that validate indigenous species’ resilience (Gamoun et al., 2012). Gulf of Gabès local species are tolerant to extreme environmental conditions due to disturbance gradients (Neji et al., 2018). These examples demonstrate that desert ecosystems can support vegetation communities under extreme environmental conditions. Brazilian semi-arid region studies exhibit the ecological significance of rocky outcrops as refugia for drought-tolerant vegetation (Silva Costa et al., 2015). The results confirm the importance of habitat heterogeneity in arid regions for guaranteeing ecological resilience.
(b) Weaknesses
Such environments are prone to soil loss and limited supply of water (Alsherif and Almaghrabi, 2022). Weather and human activities drain water resources in dry regions like Saudi Arabia. Overgrazing, especially in South America’s semi-arid region, interferes with species make-up, soils crumble, and kill natural cover (Oliva et al., 2016). Gamoun et al. (2012) have confirmed that grazing pressure and human intervention upsets plant community composition in Tunisia’s arid rangelands, reducing species richness and causing soil erosion. Exotic species like P. juliflora degrades biodiversity via competitive displacement of native vegetation (Yates et al., 2010). Invasive species are one of the primary drivers of biodiversity loss in arid ecosystems, such as Abu Dhabi’s desert offshore islands where invasive species encroach upon native plant populations (Sakkir et al., 2017). The Gulf of Gabès in Tunisia has been impacted by the ecological integrity of invasive species modifying plant dynamics (Neji et al., 2018). Urbanization, industrial pollution, and overgrazing exacerbate indigenous plant declines and interfere with ecological processes (Aljedaani and Fakhry, 2023). Urbanization in the semi-arid zones such as Bukhara, Uzbekistan, has fragmented habitats and generated non-indigenous species competing with native vegetation (Gafarova et al., 2024). South American drylands have been degraded by un-sustainable land use, causing loss of floristic richness and ecological functioning (López et al., 2006). Dams and water governance in Saudi Arabia have modified hydrological regimes, transforming riparian species richness and ecosystems (Al-Munqedhi et al., 2022).
(c) Opportunities
Tourists can witness the variety of drought-tolerant animals and distinctive scenery in deserts. Red Sea and Gulf of Aqaba ecotourism makes it possible for tourists to see desert vegetation, coral reefs, and mangroves (Basahi, 2018). Deserts offshore islands with limited vegetation attract nature lovers and raise Abu Dhabi conservation awareness (Sakkir et al., 2017). Conservation is possible through community participation and indigenous knowledge. Community can conserve land on a sustainable scale by implementing their indigenous knowledge of native plants to restore the ecosystem (El-Husseini et al., 2007). Bukhara urban conservation has proved that public participation plays an important role in indigenous plant conservation (Gafarova et al., 2024).
International cooperation can boost ecological restoration and adaptive strategy studies. Collaborative research can focus on indigenous species resistance in arid systems, for example, Acacia gerrardii adaptation in Saudi Arabian wadi ecosystems (Alatar et al., 2015). Collaborative studies across the Middle East, North Africa, and South America can support invasive species management, ecosystem rehabilitation, and sustainable farming (Schuchardt et al., 2023). Collaborations can ensure sound conservation plans and improve ecological insight. Collaboration might strengthen environmental research and ensure sound conservation.
(d) Threats
Desert plant communities face threats of rising temperature and declining rainfall. Desert plant communities are threatened by climate change (Alsherif and Almaghrabi, 2022). Long-term drought, low soil water, and decreased plant cover have modified species composition in semi-arid South America and Africa (Oliva et al., 2016; Witkowski and O’Connor, 1996). Rising temperatures in the Arabian Peninsula are exacerbating water scarcity, degrading native vegetation (Al-Mutairi et al., 2012; Basahi, 2018). Invasive species like P. juliflora push native species off native ranges and disrupt ecological processes within arid habitats (Yates et al., 2010). On Abu Dhabi’s offshore islands, invasive species disrupt native communities (Sakkir et al., 2017); in the Gulf of Gabès, they reduce species richness (Neji et al., 2018). Urban growth and industrial development lead to habitat fragmentation, which increases loss of diversity (Aljedaani and Fakhry, 2023). Development of arid ecosystems and hydrological alteration are a threat. These projects have disrupted water flows in Saudi Arabia, changing riparian ecosystems and diversity (Al-Munqedhi et al., 2022). Water management has altered plant cover in the Northern Cape of South Africa (Van Staden et al., 2020). Holistic policies and adaptive management are needed to prevent such threats.
4.8 TOWS analysis of floristic diversity of arid ecosystems of Saudi Arabia
Figure 11 shows the total TOWS strategies for Saudi Arabian arid ecosystem floristic diversity categorized from internal to external. A TOWS analysis refines the SWOT approach by highlighting strategic moves to leverage strengths and opportunities and mitigate weaknesses and threats.
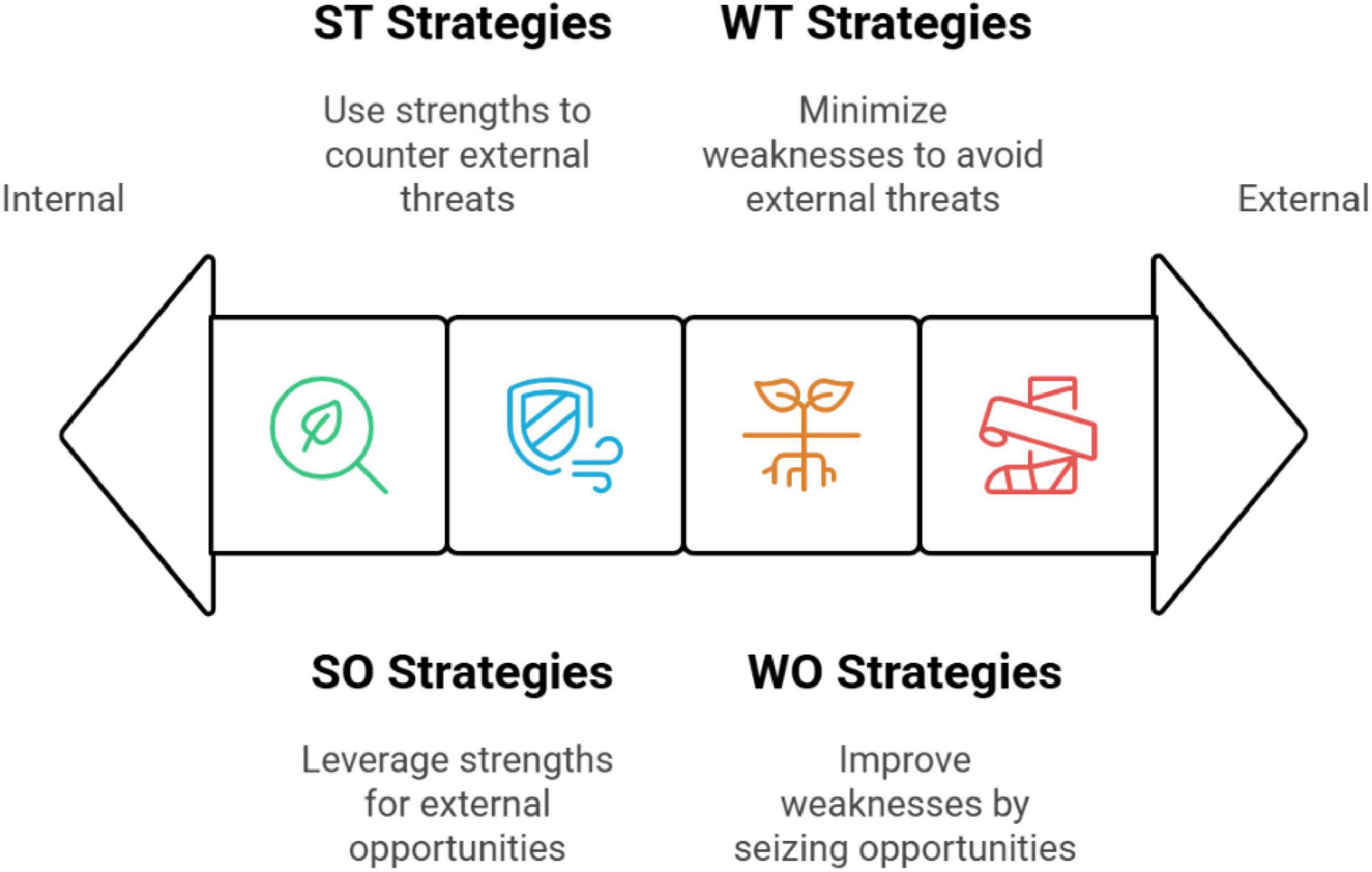
Figure 11. The overall TOWS (Threats, Opportunities, Weaknesses, and Strengths) strategies of floristic diversity in arid ecosystems of Saudi Arabia categorized from internal to external factors.
(a) Strength–opportunities (SO) strategies
The Saudi Arabian environments possess immense potential for the development of ecotourism through the utilization of the prevailing unique drought-resistant vegetation in areas such as Tabuk, Sharma, and other similar arid environments. The areas boast highly adapted vegetation to the harsh conditions, thereby presenting a natural attraction to nature enthusiasts, researchers, and nature preservers (Scaglia et al., 2024; Qureshi et al., 2011). Ecotourism initiatives can make tourists aware of the ecological importance of these animals while generating local economies through sustainable tourism.
The Red Sea coastal areas, with their extensive carbon-storing mangroves and halophytes, also have further learning opportunities for climate resilience. The coastal ecosystems, apart from their carbon-storing capabilities in preventing climate change, are also hotspots of biodiversity and uphold essential ecosystem services (Basahi, 2018). The regions can be utilized as learning sites, in which they raise awareness among visitors of the role played by coastal ecosystems in supporting efforts toward mitigating climate change.
Also, the native plant biodiversity of Saudi Arabia with environment-tolerant strains has potential genetic material for sustainable agriculture. Exploration of native drought-tolerant crops can consolidate agricultural sustainability and food security, particularly in dry regions (Imanbayeva et al., 2025; Hunter and Lechner, 2018). The use of this kind of genetic material for sustainable agriculture can build livelihoods at the local level and promote biodiversity conservation.
(b) Strengths–threats (ST) strategies
Conservation of endemic species in microhabitats is most important in alleviating the impacts of invasive species on Saudi desert ecosystems. There are certain habitats, such as mangroves on Red Sea shores and oases in deserts, which consist of endemic plant species having very high tolerance to stressful environments (Miara et al., 2025; El-Sheikh et al., 2021). Prioritized conservation of such microhabitats ensures native species continuity and does not permit the establishment of invasive species like P. juliflora, as it outcompetes native vegetation (Schuchardt et al., 2023).
Protection of the Red Sea coast needs to be enhanced to conserve carbon-sequestering ecosystems like mangroves and salt marshes. In addition to their role in climate resilience due to carbon sequestration, mangroves and salt marshes also serve as a natural barrier to coastal erosion (Basahi, 2018). Improved coastal protection will prevent the loss of these ecosystems from sea level rise and human pressures, sustaining their ecosystem services (Farid Ghazal, 2021; Li et al., 2018).
Additionally, invasive species early warning systems are necessary in such regions. In such systems, indigenous vegetation’s resilience must be tapped to identify changes in species composition and identify invasive threats at an early level (Pinheiro et al., 2010; Christie, 2014). Preventative surveillance with periodic monitoring through vegetation inventories and composition analysis can help detect modifications in plant communities, which is essential to prevent invasive species spread as well as guarantee ecological resilience (Hussein et al., 2021).
(c) Weaknesses–opportunities (WO) strategies
Strengthening community-conserved programs that involve indigenous knowledge is crucial to the achievement of sustainable land management in arid ecosystems. The programs can strengthen local communities through involvement in habitat conservation, native species cultivation, and restoration practices. For instance, integrating indigenous communities in Saudi Arabia would utilize their indigenous expertise on drought-resistant plant species, and this would enhance resilience in ecosystems (El-Husseini et al., 2007; Gomes and Alves, 2009).
International collaboration in research is also a major strategy for enhancing ecosystem restoration activities. Collaboration with other institutions from across the globe can aid in gaining knowledge on adaptive actions, sustainable agriculture, as well as biodiversity conservation. Collaborative research studies can target researching the ecological resilience of native plant species in various arid habitats, such as Tubiflorae in Egypt’s arid habitat (El-Husseini et al., 2007) and Acacia gerrardii in wadi ecosystems in Saudi Arabia (Alatar et al., 2015). Collaborative research studies can be used to innovate ecological restoration.
Public campaigns on sustainable agriculture are crucial to encouraging drought-resistant native species use. These programs have the ability to make local farmers aware of the benefits of cultivating species that are naturally suited for arid environments, reducing water consumption and enhancing crop resistance (Pashirzad et al., 2019). Promoting the use of such species can enhance food security in a manner that serves to conserve biodiversity. Urban biodiversity conservation has also been made possible by cities like Bukhara through campaigns for public awareness and education (Gafarova et al., 2024).
Strategic management policies for invasive species are critical in arid ecosystems. The policies should be directed toward early detection, rapid response, and invasive species eradication. For instance, the off-shore Abu Dhabi arid islands that are uniquely florally rich can be assisted by strict biosecurity to prevent introduction and invasive species spread (Sakkir et al., 2017). The policies should be dynamic in offering amendments based on monitoring data and emerging threats.
Sustainable land-use policy making is critical in the prevention of habitat degradation and fragmentation in ecologically sensitive ecosystems. South America’s arid regions have been shown through research to experience native vegetation loss and ecosystem function disruption due to unsustainable land use (López et al., 2006). In nations like Saudi Arabia, policy must chart boundaries for urban expansion, agricultural expansion, and industrialization to ensure the preservation of indigenous vegetation (Aljedaani and Fakhry, 2023).
Adaptive water management strategies can be used to mitigate negative impacts of dam construction on riparian systems. Adaptive water management has proved useful in maintaining biodiversity in nations like the Northern Cape of South Africa (Van Staden et al., 2020). For Saudi Arabia and other countries, managed water discharge, riparian area recovery, and buffer zone creation can be implemented to maintain vulnerable ecosystems (Al-Munqedhi et al., 2022; Akpalu et al., 2022). These measures can limit habitat disturbance, maintain species richness, and guarantee ecological connectivity within riparian corridors.
4.9 Linking bibliometric clusters with ecological mechanisms and conservation levers in arid floras
The bibliometric network revealed four coherent clusters that correspond to complementary domains of ecological understanding and management in arid floras: ecosystems and conservation (green), abiotic processes and grasslands (blue), plant dynamics and modeling (red), and phylogeny/genetics and conservation (purple). Taken together, these clusters outline a pathway from environmental drivers to population processes and, finally, to conservation action.
First, the blue cluster underscores how aridity, temperature rise, altered rainfall and soil constraints regulate productivity, recruitment, and survival in arid plant communities. Your Results already document regional warming and rainfall decline, and show how water limitation propagates through soil moisture to germination and seedling survival. This cluster provides the environmental baselines and thresholds that conservation planning must respect.
Second, the red cluster focuses on dynamics and modeling. Process-based and statistical models translate environmental signals into population- and community-level projections in space and time. In arid Saudi systems these tools help identify refugia, anticipate range shifts, and evaluate trade-offs in management, including hydrological interventions captured in Figure 7 and discussed in see section “4.3 Hydrological interventions and their ecological impacts in the floristic diversity.”
Third, the purple cluster integrates phylogeny and genetics with conservation. Genetic structure and evolutionary history inform priorities by identifying lineages, adaptive syndromes, and functional groups critical for persistence under climate stress. In arid floras with many short-lived therophytes but also lineages tied to specific substrates or microhabitats (as shown by the Tabuk–Khulais life-form and chorological comparisons), genetic and chorological signals help target conservation at both site and lineage levels.
Fourth, the green cluster aggregates ecosystem and conservation themes and is the natural “landing zone” where insights from the other clusters are operationalized. In this manuscript, that includes riparian management under dam operations (Figure 7), coastal–terrestrial coupling along the Red Sea (see section “4.8 TOWS analysis of floristic diversity of arid ecosystems of Saudi Arabia” text preceding Figure 9), and the SWOT profile of arid floristic diversity (Figure 10).
By reading the clusters together, the bibliometric map becomes an ecological blueprint: the blue cluster constrains the problem, the red cluster projects system responses, the purple cluster identifies evolutionary and functional priorities, and the green cluster organizes place-based actions. This linkage clarifies why effective arid conservation must combine climate-aware water management, genetic stewardship, and habitat connectivity.
4.10 Strategy implications for arid-zone floristic conservation in Saudi Arabia
Prioritizing these five gaps as strategic steps to convert bibliometric signals into deployable capacity.
(1) Climate-aware hydrological stewardship. The hydrological literature shows both benefits and risks of damming for arid riparian vegetation. Management that restores ecologically meaningful flow variability, creates buffer zones, and enforces soil-stabilizing practices can retain species richness and genetic heterogeneity while limiting abiotic degradation (Figure 7).
(2) Coastal–terrestrial coupling as a conservation axis. Along the Red Sea and Gulf of Aqaba, halophytes and mangroves support nutrient regulation, shoreline stability, and carbon sequestration. Safeguarding these coastal floras protects marine biodiversity and food security while sustaining terrestrial–marine linkages essential for regional resilience (see section “4.8 TOWS analysis of floristic diversity of arid ecosystems of Saudi Arabia”).
(3) Knowledge-gap triage to unlock conservation returns. The manuscript identifies needs for long-term biodiversity monitoring, soil–plant interaction studies, and genetic tools such as DNA barcoding, as well as remote sensing for scalable baselines (Figure 9).
(4) Floristic baselining by life-form and chorology. Comparative indices for Tabuk and Khulais show consistent therophyte dominance, differences in family-level representation, and distinct chorological compositions. These baselines guide site selection, restoration targets, and monitoring indicators that are tailored to arid-zone realities.
(5) From SWOT to policy. The SWOT analysis (Figure 10) situates arid floras within clear strengths and vulnerabilities, enabling policy that aligns Vision 2030 sustainability goals with ecological safeguards and local livelihoods (see sections “4.8 TOWS analysis of floristic diversity of arid ecosystems of Saudi Arabia”–“4.9 Linking bibliometric clusters with ecological mechanisms and conservation levers in arid floras”).
For the limitations of this review, the present synthesis is constrained by the temporal coverage and disciplinary balance of the underlying literature set and by uneven reporting across sites. This underlines the importance of the intensive monitoring and genetic baselining already highlighted in see section “4.8 TOWS analysis of floristic diversity of arid ecosystems of Saudi Arabia,” which are needed to refine model parameters and validate conservation outcomes in situ.
5 Conclusion
As salient outcomes, this review integrates bibliometric structure and ecological evidence into an operational pathway for arid floras in Saudi Arabia. The grouped patterns in the bibliometric co-occurrence network (Figure 3) and the thematic/bibliographic coupling structure (Figure 5) align with ecological drivers and forecasted responses (Figure 7), with phylogenetic and genetic considerations that identify irreplaceable lineages (Figure 9), and with the ecosystem-level conservation framework that organizes interventions (Figure 10). Applied to Saudi Arabia’s arid systems, this synthesis highlights three leverage points already developed in the manuscript: climate-aware riparian management, protection of coastal–terrestrial linkages along the Red Sea, and targeted closure of monitoring and genetic knowledge gaps. For clarity, we note that no separate “cluster analysis map” is presented; the term referred to the grouped patterns visualized in Figures 3, 5 and their cross-walk to Figures 7, 9, 10.
The strengths of this review lay in the combined PRISMA-based screening and VOSviewer mapping produced a transparent corpus and revealed stable thematic structure, enabling us to link clusters to actionable mechanisms and to triangulate across climate, hydrology, genetics, and conservation practice. However, the weaknesses of this review lay the limited coverage across subregions and habitats is uneven; genetic and long-term datasets are sparse; and reporting standards vary. Moreover, these features limit generalizability and complicate model validation, especially for rare lineages and microhabitats.
There are five recommendations for future strategies. Firstly, institutionalize climate-aware environmental flows in arid wadis to restore seasonal variability, protect riparian corridors, and maintain recruitment windows for arid plant guilds. Secondly, protect and restore coastal floras that anchor terrestrial–marine coupling and carbon storage along the Red Sea and Gulf of Aqaba, integrating livelihood co-benefits into conservation design. Thirdly, close the monitoring and genetic gaps by deploying remote sensing baselines, permanent plots, and DNA barcoding for key lineages to support model calibration and adaptive management. Fourthly, use life-form and chorological baselines to tailor site-specific restoration targets and indicators for Tabuk and comparable arid provinces (Tabuk–Khurais comparison). Fifthly, translate the SWOT profile to policy, aligning Vision 2030 biodiversity objectives with on-the-ground measures that reduce overgrazing pressure, control invasives, and prioritize connectivity.
Lastly, as a humble message, in arid Saudi landscapes, conserving floristic diversity under climate change is most effective when we tie environmental constraints to dynamic projections, anchor priorities in evolutionary and functional distinctiveness, and implement place-based actions in riparian corridors and coastal interfaces, all underpinned by sustained monitoring and genetic baselines. This bibliometric-to-ecology bridge provides a practical map for near-term decisions and a durable template for adaptive conservation in the decades ahead.
Data availability statement
The original contributions presented in this study are included in this article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
KA-M: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that Generative AI was used in the creation of this manuscript. I used Grammarly to check for English language and ChatGPT to check the References format in APA format.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2025.1669742/full#supplementary-material
References
Akpalu, S. E., Anglaaere, L., Damnyag, L., Dawoe, E. K., Abunyewa, A. A., and Akpalu, M. M. (2022). Floristic composition of agroforestry parklands in the semi-arid zone of Ghana: A special focus on Faidherbia albida (Delile) A. Chev. Trees For. People 9:100310. doi: 10.1016/j.tfp.2022.100310
Al-Anazi, F. S., and Al-Qahtani, S. M. (2018). Floristic composition and plant diversity of Rawdhat Al Masodi, Saudi Arabia. JKAU Sci. 30, 27–33. doi: 10.4197/Sci.30-2.3
Al-Anazi, F. S., and Al-Qahtani, S. M. (2019). Influence of selected soil physical and chemical variables on distribution of perennial plant species in different plant communities of Rawdhat Al Masodi, Saudi Arabia. Pak. J. Bot. 51, 1375–1380. doi: 10.30848/PJB2019-4(27)
Alatar, A. A., El-Sheikh, M. A. R., Thomas, J., Hegazy, A. K., and El Adawy, H. A. (2015). Vegetation, floristic diversity, and size-classes of Acacia gerrardii in an arid wadi ecosystem. Arid Land Res. Manage. 29, 335–359. doi: 10.1080/15324982.2014.968692
Al-Harbi, N. A. (2021). A comparative study on the kinds of weeds of palm plantations in Tabuk and Al-Qassim regions in Saudi Arabia. Iraqi J. Agric. Sci. 52, 763–773. doi: 10.36103/ijas.v52i3.1463
Al-Harbi, N. A. (2022). Ecology of indigenous plants in abandoned agricultural areas in Tabuk Region, Saudi Arabia. Open J. Ecol. 12, 162–174. doi: 10.4236/oje.2022.122010
Al-Harbi, N. A. (2023). Ecological and floristic study on the medicinal plants in Al-Qassim region, Saudi Arabia. Pak. J. Bot. 55, 185–197. doi: 10.30848/PJB2023-1(2)
Alharthi, A., El-Sheikh, M. A., Elhag, M., and Alatar, A. (2020). Remote sensing of 10 years changes in the vegetation cover of the northwestern coastal land of Red Sea, Saudi Arabia. Saudi J. Biol. Sci. 27, 3169–3179. doi: 10.1016/j.sjbs.2020.04.021
Alharthi, S. T., El-Shiekh, M. A., and Alfarhan, A. A. (2023). Alien plant invasions of the natural habitat in the western region of Saudi Arabia: Floristic diversity and vegetation structure. Diversity 15:309. doi: 10.3390/d15030309
Ali, A., Molau, U., Bai, Y., Jägerbrand, A. K., and Alatalo, J. M. (2016). Diversity-productivity dependent resistance of an alpine plant community to different climate change scenarios. Ecol. Res. 31, 935–945. doi: 10.1007/s11284-016-1403-6
Aljedaani, G. S., and Fakhry, A. M. (2023). Floristic diversity of Jeddah: An arid desert, western region of Saudi Arabia. Egypt. J. Bot. 63, 57–68. doi: 10.21608/ejbo.2022.126880.1933
Aljeddani, G. S., Al-Harbi, N. A., Al-Qahtani, S. M., El-Absy, K. M., Abdullatif, B. M., and Dahan, T. E. (2021). Inventory of some introduced and invasive plant species in some governorates of the Kingdom of Saudi Arabia. Appl. Ecol. Environ. Res. 19, 4373–4388. doi: 10.15666/aeer/1906_43734388
Al-Masri, M. A., and Alsharif, A. H. (2021). Climate change impacts on plant biodiversity in arid regions: A case study from Saudi Arabia. Environ. Sci. Policy 119, 123–130. doi: 10.1016/j.sjbs.2022.01.043
Al-Munqedhi, B. M., El-Sheikh, M. A., Alfarhan, A. H., Alkahtani, A. M., Arif, I. A., Rajagopal, R., et al. (2022). Climate change and hydrological regime in arid lands: Impacts of dams on the plant diversity, vegetation structure and soil in Saudi Arabia. Saudi J. Biol. Sci. 29, 3194–3206. doi: 10.1016/j.sjbs.2022.01.043
Al-Mutairi, K. A. (2017). Influence of soil physical and chemical variables on the species composition and richness of plants in the arid region of Tabuk, Saudi Arabia. Ekol 36, 112–120. doi: 10.1515/eko-2017-0010
Al-Mutairi, K. A. (2022). Do spatially structured soil variables influence the plant diversity in Tabuk arid region, Saudi Arabia? Sustainability 14:2611. doi: 10.3390/su14052611
Al-Mutairi, K. A., El-Bana, M., Mansor, M., Al-Rowaily, S., and Mansor, A. (2012). Floristic diversity, composition, and environmental correlates on the arid, coralline islands of the Farasan archipelago, Red Sea, Saudi Arabia. Arid Land Res. Manage. 26, 137–150. doi: 10.1080/15324982.2012.656179
Al-Mutairi, K., Al-Shami, S. A., Khorshid, Z., and Moawed, M. M. (2016). Floristic diversity of Tabuk Province, North Saudi Arabia. J. Anim. Plant Sci. 26, 1019–1025.
Al-Qahtani, S. M., El-Hashash, E., El-Absy, E., and Al-Taymanl, A. (2023). Diversity of weed species in grape farms of Tayma (Tabuk, Saudi Arabia): Implication for invasive species ecology. Biosci. J. 39:e39088. doi: 10.14393/BJ-v39n0a2023-67975
Alsherif, E. A., and Almaghrabi, O. A. (2022). Consequences of climate change on the floristic composition of Khulais region, Saudi Arabia. Appl. Ecol. Environ. Res. 20, 2683–2695. doi: 10.15666/aeer/2003_26832695
Alsobeai, S. M. (2024). Floristic diversity, life form, and plant life chorology at Al-Fawzan Reserve, Kingdom of Saudi Arabia. Appl. Ecol. Environ. Res. 22, 3709–3726. doi: 10.15666/aeer/2204_37093726
Anju, T., and Kumar, A. (2024). Traditional ecological knowledge and medicinal plant diversity usage among the Mullu Kuruman tribes of Wayanad district of Kerala, India, and its implications for biodiversity conservation in the face of climate change. Trees For. People 16:100595. doi: 10.1016/j.tfp.2024.100595
Ansari, A. A. (2016). Seasonal dynamics in the relative density of aquatic flora along some coastal areas of the Red Sea, Tabuk, Saudi Arabia. Egypt. J. Aquat. Res. 42, 273–280. doi: 10.1016/j.ejar.2016.06.001
Ansari, A. A., Siddiqui, Z. H., Alatawi, F. A., Alharbi, B. M., and Alotaibi, A. S. (2022). An assessment of biodiversity in Tabuk region of Saudi Arabia: A comprehensive review. Sustainability 14:10564. doi: 10.3390/su141710564
Azeem, A., Haq, M. Z. U., Rasheed, H. U., Uddin, S., Dullu, M. U. D., Azeem, S., et al. (2023). “Bridging the gap between climate change and plant biology: Enhancing resilience through genetic diversity and agroecological practices,” in Water-Soil-Plant-Animal Nexus in the Era of Climate Change, 173–193, ed. K. Ahmed (Pennsylvania: IGI Global), doi: 10.4018/978-1-6684-9838-5.ch008
Bai, L., Wang, Z., Lu, Y., Tian, T., and Peng, Y. (2022). Monthly rather than annual climate variation determines plant diversity change in four temperate grassland nature reserves. Environ. Sci. Pollut. Res. 29, 10357–10365. doi: 10.1007/s11356-021-16473-y
Bakker, W., Morel, T., Ozinga, W., Scheper, J., and Vergeer, P. (2024). The relative importance of nitrogen deposition and climate change in driving plant diversity decline in roadside grasslands. Sci. Total Environ. 955:176962. doi: 10.1016/j.scitotenv.2024.176962
Basahi, R. A. (2018). Plant diversity of the coastal regions of Gulf of Aqaba, Saudi Arabia. Ann. Res. Rev. Biol. 26, 1–11. doi: 10.9734/ARRB/2018/41212
Batanouny, K. H. (1987). Current knowledge of plant ecology in the Arab Gulf countries. Catena 14, 291–316. doi: 10.1016/0341-8162(87)90022-1
Belyazid, S., Phelan, J., Nihlgård, B., Sverdrup, H., Driscoll, C., Fernandez, I., et al. (2019). Assessing the effects of climate change and air pollution on soil properties and plant diversity in northeastern U.S. hardwood forests: Model setup and evaluation. Water Air Soil Pollut. 230:106. doi: 10.1007/s11270-019-4145-6
Blaus, A., Reitalu, T., Poska, A., Vassiljev, J., and Veski, S. (2021). Mire plant diversity change over the last 10,000 years: Importance of isostatic land uplift, climate and local conditions. J. Ecol. 109, 3634–3651. doi: 10.1111/1365-2745.13742
Bordoloi, R., Sharma, A. R., Das, B., Teron, G., Thungon, L. T., Paul, A., et al. (2023). Plant diversity and variation in soil properties of selected land use types of Arunachal Pradesh: A local climate change perspective. Vegetos 36, 950–960. doi: 10.1007/s42535-022-00484-2
Braz Pires, M., Kougioumoutzis, K., Norder, S., Dimopoulos, P., Strid, A., and Panitsa, M. (2024). The future of plant diversity within a Mediterranean endemism centre: Modelling the synergistic effects of climate and land-use change in Peloponnese, Greece. Sci. Total Environ. 947:174622. doi: 10.1016/j.scitotenv.2024.174622
Cardinale, B. J., Duffy, J. E., Gonzalez, A., Hooper, D. U., Perrings, C., Venail, P., et al. (2012). Biodiversity loss and its impact on humanity. Nature 486, 59–67. doi: 10.1038/nature11148
Christie, K. (2014). Floristic dynamics across a semi-arid volcanic chronosequence in northern Arizona. Southwestern Naturalist 59, 364–371. doi: 10.1894/FMO-11.1
Dullinger, I., Gattringer, A., Wessely, J., Moser, D., Plutzar, C., Willner, W., et al. (2020). A socio-ecological model for predicting impacts of land-use and climate change on regional plant diversity in the Austrian Alps. Glob. Change Biol. 26, 2336–2352. doi: 10.1111/gcb.14977
El-Husseini, N., Abd El-Ghani, M. M., and El-Naggar, S. I. (2007). Floristic analysis and biogeography of Tubiflorae in some arid regions. Int. J. Ecol. Environ. Sci. 33, 1–21.
Elkordy, A., Nour, I. H., Ellmouni, F. Y., Al Shaye, N. A., Al-Bakre, D. A., and El-Banhawy, A. (2022). Floristic diversity of Jabal Al-Ward, southwest Tabuk region, Kingdom of Saudi Arabia. Agronomy 12:2626. doi: 10.3390/agronomy12112626
El-Sheikh, M. A., Al-Shehri, M. A., Alfarhan, A. H., Alatar, A. A., Rajakrishnan, R., and Al-Rowaily, S. L. (2019). Threatened Prunus arabica in an ancient volcanic protected area of Saudi Arabia: Floristic diversity and plant associations. Saudi J. Biol. Sci. 26, 325–333. doi: 10.1016/j.sjbs.2018.08.004
El-Sheikh, M. A., Thomas, J., Arif, I. A., and El-Sheikh, H. M. (2021). Ecology of inland sand dunes “nafuds” as a hyper-arid habitat, Saudi Arabia: Floristic and plant associations diversity. Saudi J. Biol. Sci. 28, 1503–1513. doi: 10.1016/j.sjbs.2020.12.002
Fakhry, A. M., and Anazi, H. K. (2017). Floristic diversity and vegetation structure of Al-Dissah area, northwestern Saudi Arabia. Egypt. J. Exp. Biol. 13, 447–457. doi: 10.5455/egyjebb.20171009105256
Fakhry, A. M., and El-Kenany, E. T. (2019). Floristic diversity and endangered species in Harrat ArRahah solidified lava area, southern Tabuk, Saudi Arabia. Egypt. J. Exp. Biol. 15, 217–226. doi: 10.5455/egyjebb.20190619084913
Farid Ghazal, A. M. (2021). Assessment of potential functional use of floristic compositions in landscape restoration of habitats in arid regions. Pak. J. Bot. 53, 2143–2155. doi: 10.30848/PJB2021-6(38)
Fine, P. V. A. (2015). Ecological and evolutionary drivers of geographic variation in species diversity. Annu. Rev. Ecol. Evol. Syst. 46, 369–392. doi: 10.1146/annurev-ecolsys-112414-054102
Gafarova, S. M., Gulamov, M. I., Esanov, H. K., and Umedov, A. M. (2024). Urban floristic diversity in the arid zone: A case study of Bukhara city. Acta Biol. Sibirica 10, 197–213. doi: 10.5281/zenodo.10934573
Gamoun, M., Ouled Belgacem, A., Hanchi, B., Neffati, M., and Gillet, F. (2012). Impact of grazing on the floristic diversity of arid rangelands in south Tunisia. Revue d’Ecologie 67, 271–282.
Garcés-Pastor, S., Coissac, E., Lavergne, S., and Schwörer, C. (2022). High resolution ancient sedimentary DNA shows that alpine plant diversity is associated with human land use and climate change. Nat. Commun. 13:6559. doi: 10.1038/s41467-022-34010-4
Ghesquière, M., Bourgoin, T., Durand, J.-L., Volaire, F., and Sampoux, J.-P. (2016). Confronting climate change using plant genetic diversity: Available margin of improvement in forage grasses. Fourrages 2016, 29–38.
Gomes, P., and Alves, M. (2009). Floristic and vegetational aspects of an inselberg in the semi-arid region of northeast Brazil. Edinburgh J. Bot. 66, 329–346. doi: 10.1017/S0960428609005241
Hautier, Y., Isbell, F., Borer, E. T., Seabloom, E. W., Harpole, W. S., Lind, E. M., et al. (2018). Local loss and spatial homogenization of plant diversity reduce ecosystem multifunctionality. Nat. Ecol. Evol. 2, 50–56. doi: 10.1038/s41559-017-0395-0
Hua, Z. (2013). The floras of southern and tropical southeastern Yunnan have been shaped by divergent geological histories. PLoS One 8:e64213. doi: 10.1371/journal.pone.0064213
Huang, S., and Fu, U. (2023). Impacts of climate change and human activities on plant species α-diversity across the Tibetan grasslands. Remote Sens. 15:2947. doi: 10.3390/rs15112947
Hunter, J. T., and Lechner, A. M. (2018). A multiscale, hierarchical, ecoregional, and floristic classification of arid and semi-arid ephemeral wetlands in New South Wales, Australia. Mar. Freshwat. Res. 69, 418–431. doi: 10.1071/MF17006
Hussein, E. A., El-Ghani, M. M. A., Hamdy, R. S., and Shalabi, L. F. (2021). Do anthropogenic activities affect floristic diversity and vegetation structure more than natural soil properties in hyper-arid desert environments? Diversity 13:157. doi: 10.3390/d13040157
Imanbayeva, A., Ishmuratova, M., Duisenova, N., Sagyndykova, M., Orazov, A., and Tuyakova, A. (2025). Floristic and anatomical diversity of Crataegus ambigua C.A.Mey. ex A.K.Becker populations in different areas of the arid Mangystau region (Kazakhstan). Forests 16:585. doi: 10.3390/f16040585
Imanberdieva, N., Imankul, B., Severoglu, Z., Altay, V., and Öztürk, V. (2018). “Potential impacts of climate change on plant diversity of Sary-chelek biosphere reserve in Kyrgyzstan,” in Vegetation of Central Asia and Environs: 349–364, eds D. Egamberdieva and M. Öztürk (Berlin: Springer), doi: 10.1007/978-3-319-99728-5_15
Inague, G. M., Zwiener, V. P., and Marques, M. C. M. (2021). Climate change threatens the woody plant taxonomic and functional diversities of the Restinga vegetation in Brazil. Perspect. Ecol. Conserv. 19, 53–60. doi: 10.1016/j.pecon.2020.12.006
Jewitt, D., Goodman, P. S., Erasmus, B. F. N., O’Connor, T. G., and Witkowski, E. T. F. (2017). Planning for the maintenance of floristic diversity in the face of land cover and climate change. Environ. Manag. 59, 792–806. doi: 10.1007/s00267-017-0829-0
Kougioumoutzis, K., Kokkoris, I. P., Panitsa, M., Trigas, P., Strid, V., and Dimopoulos, P. (2020). Plant diversity patterns and conservation implications under climate-change scenarios in the Mediterranean: The case of Crete (Aegean, Greece). Diversity 12:270. doi: 10.3390/d12070270
Kürschner, H., and Neef, R. (2011). A first synthesis of the flora and vegetation of the Tayma oasis and surroundings (Saudi Arabia). Plant Divers. Evol. 129, 27–58. doi: 10.1127/1869-6155/2011/0129-0033
Kürschner, H., and Neef, R. (2024). “Taymā’ Oasis (Saudi Arabia) and its surroundings – A first synthesis of the flora, vegetation, natural resources, and floral history,” in Taymā’: Archaeological exploration, palaeoenvironment, cultural contacts, eds A. Hausleiter, R. Eichmann, and M. Al-Najem (Bicester: Archaeopress), 87–102.
Lauterbach, M., Veranso-Libalah, M. C., Sukhorukov, A. P., and Kadereit, G. (2019). Biogeography of the xerophytic genus Anabasis L. (Chenopodiaceae). Ecol. Evol. 9, 3539–3552. doi: 10.1002/ece3.4987
Lelieveld, J., Proestos, Y., Hadjinicolaou, P., Tanarhte, M., Tyrlis, E., and Zittis, G. (2016). Strongly increasing heat extremes in the Middle East and North Africa (MENA) in the 21st century. Climatic Change 137, 245–260. doi: 10.1007/s10584-016-1665-6
Li, E., Huang, Y., Chen, H., and Zhang, J. (2018). Floristic diversity analysis of the Ordos Plateau, a biodiversity hotspot in arid and semi-arid areas of China. Folia Geobot. 53, 405–416. doi: 10.1007/s12224-018-9331-6
Liu, Y., Su, X., Shrestha, N., Xu, X., Wang, S., Li, Y., et al. (2019). Effects of contemporary environment and Quaternary climate change on drylands plant diversity differ between growth forms. Ecography 42, 334–345. doi: 10.1111/ecog.03698
López, R. P., Alcázar, D. L., and Macía, M. J. (2006). The arid and dry plant formations of South America and their floristic connections: New data, new interpretation? Darwiniana 44, 18–31.
Lu, J., Lu, H., Brusseau, M. L., He, L., Gorlier, A., Yao, T., et al. (2021). Interaction of climate change, potentially toxic elements (PTEs), and topography on plant diversity and ecosystem functions in a high-altitude mountainous region of the Tibetan Plateau. Chemosphere 275:130099. doi: 10.1016/j.chemosphere.2021.130099
Ma, S., Li, W., Tojibaev, K. S., Turginov, O., Yang, W., and Ma, K. (2024). Regionwide and nationwide floristic richness reveal vascular plant diversity in central Asia. Plants 13:2275. doi: 10.3390/plants13162275
Maestre, F. T., Quero, J. L., Gotelli, N. J., Escudero, A., Ochoa, V., Delgado-Baquerizo, M., et al. (2012). Plant species richness and ecosystem multifunctionality in global drylands. Science 335, 214–218. doi: 10.1126/science.1215442
Manafzadeh, S., Staedler, Y. M., and Conti, E. (2017). Visions of the past and dreams of the future in the Orient: The Irano-Turanian region from classical botany to evolutionary studies. Biol. Rev. 92, 1365–1388. doi: 10.1111/brv.12287
Miara, M. D., Youssef, S., Fendane, Y., Marris, L., Sallio, R., Boukcim, H., et al. (2025). Phytogeographic transition areas hide floristic diversity in hyper-arid environments (Sharaan Natural Reserve, NW-Saudi Arabia). Diversity 17:30. doi: 10.3390/d17010030
Moawed, M. M. (2016). Plant flora of Alaqan region, Tabuk province, Saudi Arabia, Egypt. J. Exp. Biol. 12, 107–113. doi: 10.12345/ejeb.2016.107113
Moawed, M. M., and Ansari, A. A. (2015). Wild plants diversity of Red Sea coastal region, Tabuk, Saudi Arabia. J. Chem. Pharm. Res. 7, 220–227.
Moher, D., Liberati, A., Tetzlaff, J., and Altman, D. G.PRISMA Group. (2009). Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 6:e1000097. doi: 10.1371/journal.pmed.1000097
Montenegro-Hoyos, A., Vega, N., and Linares-Palomino, R. (2022). Plant diversity and structure in desert communities of the Andean piedmont in Ica, Peru. Vegetation Classification Survey 3, 53–66. doi: 10.3897/VCS.68006
Nabhan, G. P., Colunga-García, M. P., and Zizumbo-Villarreal, D. (2022). Comparing wild and cultivated food plant richness between the arid American and the Mesoamerican centers of diversity, as means to advance indigenous food sovereignty in the face of climate change. Front. Sustain. Food Syst. 6:840619. doi: 10.3389/fsufs.2022.840619
Neji, M., Serbaji, M. M., Hardy, O., and Chaieb, M. (2018). Floristic diversity and vegetation patterns along disturbance gradient in arid coasts in southern Mediterranean: Case of the Gulf of Gabès, southern Tunisia. Arid Land Res. Manage. 32, 291–315. doi: 10.1080/15324982.2018.1431332
Oliva, G., Ferrante, D., Paredes, P., Humano, G., and Cesa, V. (2016). A conceptual model for changes in floristic diversity under grazing in semi-arid Patagonia using the State and Transition framework. J. Arid Environ. 127, 120–127. doi: 10.1016/j.jaridenv.2015.10.018
Ordonez, A., and Svenning, J. C. (2017). Consistent role of Quaternary climate change in shaping current plant functional diversity patterns across European plant orders. Sci. Rep. 7:42988. doi: 10.1038/srep42988
Ortega, M. A., Cayuela, L., Griffith, D. M., and Camacho, A. (2024). Climate change increases threat to plant diversity in tropical forests of Central America and southern Mexico. PLoS One 19:e0297840. doi: 10.1371/journal.pone.0297840
Ossa, P. G., Armesto, J. J., and Pérez, F. (2017). Assessing the influence of life form and life cycle on the response of desert plants to past climate change: Genetic diversity patterns of an herbaceous lineage of Nolana along western South America. Am. J. Bot. 104, 1533–1545. doi: 10.3732/ajb.1700101
Pashirzad, M., Ejtehadi, H., Vaezi, J., and Shefferson, R. P. (2019). Multiple processes at different spatial scales determine beta diversity patterns in a mountainous semi-arid rangeland of Khorassan-Kopet Dagh floristic province, NE Iran. Plant Ecol. 220, 829–844. doi: 10.1007/s11258-019-00957-6
Peng, Y., Xin, J., and Peng, N. (2023). Climate change alters the spatial pattern of plant spectral diversity across forest types. Front. Ecol. Evol. 11:1137111. doi: 10.3389/fevo.2023.1137111
Peralta, P. A., Nores, M. J., Bach, H. G., and Robbiati, F. O. (2024). Facing climate change: Range dynamics and chromosome diversity in Hedeoma multiflora Benth., a South American aromatic-medicinal plant at risk. Flora 315:152519. doi: 10.1016/j.flora.2024.152519
Phelan, J., Belyazid, S., Jones, P., Cajka, J., Buckley, J., and Clark, C. (2016). Assessing the effects of climate change and air pollution on soil properties and plant diversity in sugar maple-beech-yellow birch hardwood forests in the Northeastern United States: Model simulations from 1900 to 2100. Water Air Soil Pollut. 227:84. doi: 10.1007/s11270-016-2762-x
Pinheiro, K., Rodal, M. J. N., and Alves, M. (2010). Floristic composition of different soil types in a semi-arid region of Brazil. Rev. Caatinga 23, 68–77.
Pütz, T., Kiese, R., Wollschläger, U., Groh, J., Rupp, H., Zacharias, S., et al. (2016). TERENO-SOILCan: A lysimeter-network in Germany observing soil processes and plant diversity influenced by climate change. Environ. Earth Sci. 75:1242. doi: 10.1007/s12665-016-6031-5
Qureshi, R., Bhatti, G. R., and Shabbir, G. (2011). Floristic inventory of Pir Mehr Ali Shah Arid Agriculture University Research farm at Koont and its surrounding areas. Pak. J. Bot. 43, 1679–1684.
Raven, P. H., Gereau, R. E., Phillipson, P. B., Chatelain, C., Jenkins, C. N., and Ulloa Ulloa, C. (2020). The distribution of biodiversity richness in the tropics. Sci. Adv. 6:eabc6228. doi: 10.1126/sciadv.abc6228
Renny, M., Acosta, M. C., and Sérsic, A. N. (2022). Ancient climate changes and Andes uplift, rather than Last Glacial Maximum, affected distribution and genetic diversity patterns of the southernmost mycoheterotrophic plant Arachnitis uniflora Phil. (Corsiaceae). Glob. Planet. Change 208:103701. doi: 10.1016/j.gloplacha.2021.103701
Sakkir, S., Shah, J. N., Soorae, P., Al Mehairbi, M., and Al Dhaheri, S. (2017). Floristic composition of arid off-shore islands in Abu Dhabi, United Arab Emirates. J. Coast. Conserv. 21, 189–195. doi: 10.1007/s11852-016-0490-4
Scaglia, J. A., Flores, D. G., Tapia, R., and Martinelli, M. (2024). Effects of geomorphology and distribution of water sources for livestock on the floristic composition and livestock receptivity of the Arid Chaco. Rev. Fac. Cienc. Agrar. 56, 12–25. doi: 10.48162/rev.39.119
Schuchardt, M. A., Berauer, B. J., Duc, A. L., Ingrisch, J., Niu, Y., Bahn, M., et al. (2023). Increases in functional diversity of mountain plant communities is mainly driven by species turnover under climate change. Oikos 2023:e09922. doi: 10.1111/oik.09922
Schuldt, A., Ebeling, A., Kunz, M., Staab, M., Guimarães-Steinicke, C., Bachmann, D., et al. (2019). Multiple plant diversity components drive consumer communities across ecosystems. Nat. Commun. 10:1460. doi: 10.1038/s41467-019-09335-3
Sekar, K. C., Thapliyal, N., Bhojak, P., Bisht, K., Pandey, A., Mehta, P., et al. (2024). Early signals of climate change impacts on alpine plant diversity in Indian Himalaya. Biodivers. Conserv. 34, 207–233. doi: 10.1007/s10531-024-02966-y
Sillmann, J., Kharin, V. V., Zwiers, F. W., Zhang, X., and Bronaugh, D. (2013). Climate extremes indices in the CMIP5 multimodel ensemble: Part 2. Future climate projections. J. Geophys. Res Atmospheres 118, 2473–2493. doi: 10.1002/jgrd.50188
Silva Costa, E. C., Lopes, S. F., and De Melo, J. I. M. (2015). Floristic similarity and dispersal syndromes in a rocky outcrop in semi-arid northeastern Brazil. Rev. Biol. Trop. 63, 827–843. doi: 10.15517/rbt.v63i3.15929
Soliani, C., Ceccarelli, V., Lantschner, M. V., Thomas, E., and Marchelli, P. (2024). Correction to: Predicting the distribution of plant species from southern South America: Are the hotspots of genetic diversity threatened by climate change? Biodivers. Conserv. 33, 1869–1870. doi: 10.1007/s10531-024-02836-7
Ssali, F., Mugerwa, B., van Heist, M., Sheil, D., Kirunda, B., Musicante, M., et al. (2023). Plant diversity and composition vary with elevation on two equatorial high mountains in Uganda: Baselines for assessing the influence of climate change. Alp. Bot. 133, 149–161. doi: 10.1007/s00035-023-00301-9
Stoica, I.-A., Hodor, N., Tudose, T., and Coldea, G. (2017). Expected floristic changes in hygro-cryophilic and snowbed plant communities caused by climate change and human impact in the Romanian Carpathians. Contrib. Bot. 52, 163–181. doi: 10.24193/Contrib.Bot.52.12
Suggitt, A. J., Lister, D. G., and Thomas, C. D. (2019). Widespread effects of climate change on local plant diversity. Curr. Biol. 29, 2905–2911.e2. doi: 10.1016/j.cub.2019.06.079
Sun, Y., Yao, S., Hu, W., and Wang, T. (2021). Predicting future changes in plant species richness under climate change scenarios: A random forest approach applied to drylands in China and implications for Saudi Arabia. Ecol. Indic. 132, 1470–1160. doi: 10.1016/j.ecolind.2021.108288
Tang, L., Wang, R., He, K. S., Shi, C., Yang, T., Huang, Y., et al. (2019). Throwing light on dark diversity of vascular plants in China: Predicting the distribution of dark and threatened species under global climate change. PeerJ 2019:e6731. doi: 10.7717/peerj.6731
Trew, B. T., Early, R., Duffy, J. P., Chown, S. L., and Maclean, I. (2022). Using near-ground leaf temperatures alters the projected climate change impacts on the historical range of a floristic biodiversity hotspot. Divers. Distrib. 28, 1282–1297. doi: 10.1111/ddi.13540
Trisurat, Y., Sutummawong, N., Roehrdanz, P. R., and Chitechote, A. (2023). Climate change impacts on species composition and floristic regions in Thailand. Diversity 15:1087. doi: 10.3390/d15101087
Tyler, T., Herbertsson, L., Olsson, P. A., Fröberg, L., Olsson, K.-A., Svensson, Å, et al. (2018). Climate warming and land-use changes drive broad-scale floristic changes in Southern Sweden. Glob. Change Biol. 24, 2607–2621. doi: 10.1111/gcb.14031
Van Eck, N. J., and Waltman, L. (2010). Software survey: VOSviewer, a computer program for bibliometric mapping. Scientometrics 84, 523–538. doi: 10.1007/s11192-009-0146-3
Van Eck, N. J., and Waltman, L. (2014). “Visualizing bibliometric networks,” in Measuring scholarly impact: Methods and practice, eds Y. Ding, R. Rousseau, and D. Wolfram (Berlin: Springer), 285–320. doi: 10.1007/978-3-319-10377-8_13
Van Eck, N. J., and Waltman, L. (2017). Citation-based clustering of publications using CitNetExplorer and VOSviewer. Scientometrics 111, 1053–1070. doi: 10.1007/s11192-017-2300-7
Van Staden, N., Siebert, S. J., Cilliers, D. P., Wilsenach, D., and Frisby, A. W. (2020). Floristic analysis of semi-arid mountain ecosystems of the Griqualand West Centre of Plant Endemism, Northern Cape, South Africa. Biodiversitas 21, 1989–2002. doi: 10.13057/biodiv/d210526
Whitmee, S., Haines, A., Beyrer, C., Boltz, F., and Capon, A. G. (2015). Safeguarding human health in the Anthropocene epoch: Report of the Rockefeller Foundation-Lancet Commission on planetary health. Lancet 386, 1973–2028. doi: 10.1016/S0140-6736(15)60901-1
Williams, T. M., Schlichting, C. D., and Holsinger, K. E. (2021). Herbarium records demonstrate changes in flowering phenology associated with climate change over the past century within the Cape Floristic Region, South Africa. Clim. Change Ecol. 1:100006. doi: 10.1016/j.ecochg.2021.100006
Witkowski, E. T. F., and O’Connor, T. G. (1996). Topo-edaphic, floristic and physiognomic gradients of woody plants in a semi-arid African savanna woodland. Vegetatio 124, 9–23. doi: 10.1007/BF00045140
Wróblewska, A., and Mirski, P. (2018). From past to future: Impact of climate change on range shifts and genetic diversity patterns of circumboreal plants. Reg. Environ. Change 18, 409–424. doi: 10.1007/s10113-017-1208-3
Xia, C., Huang, Y., Qi, Y., Yang, X., Xue, T., Hu, R., et al. (2022). Developing long-term conservation priority planning for medicinal plants in China by combining conservation status with diversity hotspot analyses and climate change prediction. BMC Biol. 20:89. doi: 10.1186/s12915-022-01285-4
Yang, X., Liu, B., Bussmann, R. W., Guan, X., Xu, W., Xue, T., et al. (2021). Integrated plant diversity hotspots and long-term stable conservation strategies in the unique karst area of southern China under global climate change. For. Ecol. Manag. 498:119540. doi: 10.1016/j.foreco.2021.119540
Yao, S., Sun, Y., and Hu, W. (2021). Future trends of plant species richness in arid regions based on predictive ecological models. Ecol. Indic. 132:108288.
Yates, C. J., McNeill, A., Elith, J., and Midgley, G. F. (2010). Assessing the impacts of climate change and land transformation on Banksia in the South West Australian Floristic Region. Divers. Distrib. 16, 187–201. doi: 10.1111/j.1472-4642.2009.00623.x
Youssef, S., Miara, M., Boivin, S., Sallio, R., Nespoulous, J., Boukcim, H., et al. (2024). Recovery of perennial plant communities in disturbed hyper-arid environments (Sharaan Nature Reserve, Saudi Arabia). Land 13:2033. doi: 10.3390/land13122033
Zeng, Y., Liu, T., Zhou, X.-B., Sun, Q.-M., Han, Z.-Q., and Liu, K. (2016). Effects of climate change on plant composition and diversity in the Gurbantünggüt Desert of northwestern China. Ecol. Res. 31, 427–439. doi: 10.1007/s11284-016-1352-0
Keywords: floristic diversity, climate change, conservation strategies, hydrological interventions, Saudi Arabia
Citation: Al-Mutairi KA (2025) From desert margins to global insights: floristic diversity and conservation strategies in the arid regions of Tabuk and Khulais, Saudi Arabia – a bibliometric and ecological synthesis. Front. For. Glob. Change 8:1669742. doi: 10.3389/ffgc.2025.1669742
Received: 20 July 2025; Accepted: 07 October 2025;
Published: 20 October 2025.
Edited by:
Mohammad Imam Hasan Reza, Centre for Environment and Sustainability, Presidency Education, BangladeshReviewed by:
Sumit Chakravarty, Uttar Banga Krishi Viswavidyalaya, IndiaAmal M. Fakhry, Alexandria University, Egypt
Copyright © 2025 Al-Mutairi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Khalid Awadh Al-Mutairi, a211dGFpcmlAdXQuZWR1LnNh
†ORCID: Khalid Awadh Al-Mutairi, orcid.org/0000-0003-2356-0724