Abstract
Introduction:
In order to effectively manage ecosystems, it is important to understand how the structure and function of the ecosystem are measured and interpreted, and ecosystem multifunctionality (EMF) is being used as an important indicator for providing sustainable ecosystem functions and managing qualitative stability. The objective of this study is to examine the biotic and abiotic mechanisms underlying the production and regulation of EMF within various forest stand types.
Methods:
To this end, we assessed the influence of biotic (tree species, functional, and stand structural diversity of trees), abiotic (elevation, aridity index), and stand age (mean age of the five dominant trees per plot) factors on EMF. A total of 2,859 natural forest plots—comprising coniferous, broadleaved, and mixed stands—were analyzed based on data from the 7th National Forest Inventory of South Korea. To determine the major factors influencing EMF, we applied a multi-model inference approach along with piecewise structural equation modeling.
Results:
Our results suggest that higher plant biodiversity was positively associated with EMF. In addition, older forests exhibit greater stand structural diversity, which in turn enhances the maintenance of EMF. The analysis of abiotic factors revealed that EMF increased with elevation. Furthermore, variables related to plant water stress consistently had negative direct and indirect effects across all forest stand types. Finally, the mechanisms controlling EMF differed among forest stand types.
Discussion:
These findings suggest that promoting forest succession, maintaining biodiversity, and enhancing stand structural diversity are essential forest management strategies for improving EMF. Furthermore, since the controlling factors of EMF varied among forest stand types, forest-type specific management strategies are required. Lastly, this study provides valuable insights for guiding sustainable forest management that enhances both EMF and specific ecosystem functions, while supporting human well-being.
1 Introduction
An ecosystem is a system in which biological components, such as plants, animals, and microorganisms, interact with abiotic components, such as climate, soil, and topography (Tansley, 1935). Organisms within an ecosystem closely interact through the use of solar energy and externally supplied nutrients, which is a key mechanism for maintaining both the structure and balance of the ecosystem (Tansley, 1935; DeAngelis, 1980). Through these mechanisms, humans have long benefited from various ecosystem services (Costanza et al., 1997). For example, forest ecosystems, which cover approximately 31% of the Earth’s land surface, account for about two-thirds of global photosynthesis. In doing so, they provide the ecosystem service of greenhouse gas mitigation by performing critical ecosystem functions, storing approximately 86% of terrestrial carbon and 73% of soil carbon (Sun and Liu, 2020). To effectively manage such ecosystems, it is essential to understand not only their structure but also how their functions are measured and interpreted (Garland et al., 2021). Accordingly, intensive research has been conducted on biodiversity and various ecosystem functions based on the Biodiversity–Ecosystem Functioning (BEF) framework (Grace et al., 2014; Mori et al., 2017; Gonzalez et al., 2020; Scherer-Lorenzen et al., 2022; Ali, 2023). Most studies on BEF have focused on carbon sinks for climate regulation, such as forest aboveground biomass (AGB), soil organic carbon (SOC), and soil microbial diversity, which are representative ecosystem functions (Chun et al., 2020; Lee et al., 2023; Lee et al., 2024b). Based on these studies, management strategies to enhance ecosystem functions have been proposed, while other studies have warned of functional declines resulting from forest area loss and ecosystem degradation caused by climate crises (Newbold et al., 2015; van der Plas et al., 2016; Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services [IPBES], 2019). However, recent research has reached a consensus that focusing on a single ecosystem function is insufficient to ensure the overall quality, stability, and sustainability of ecosystems, especially under intensified environmental change (Yuan et al., 2020; Garland et al., 2021; Dietrich et al., 2024). Consequently, enhancing multiple ecosystem functions in forests and developing comprehensive indicators to assess them has emerged as a key research priority (Byrnes et al., 2014; Manning et al., 2018). To achieve this goal and promote stable improvements in forest ecosystem functions, the concept of ecosystem multifunctionality (EMF), defined as the overall functioning of an ecosystem or its ability to provide multiple functions and services simultaneously, has been introduced (Hector and Bagchi, 2007; Gamfeldt et al., 2008; Manning et al., 2018). EMF, which considers the interactions and trade-offs among multiple ecosystem functions rather than a single one, is increasingly used as a crucial indicator for managing the sustainable delivery and qualitative stability of ecosystem functions (Manning et al., 2018).
As EMF gains attention as a key indicator for sustainable forest ecosystem management, understanding the major controlling factors that determine it is becoming increasingly important (Yuan et al., 2020; Wang et al., 2022). In particular, growing research efforts have aimed to uncover how various biotic and abiotic factors, such as biodiversity, forest structural attributes, and topographic characteristics, influence ecosystem functions and EMF. Previous studies have shown that EMF is regulated by multiple biotic factors across tropical, temperate, and various forest regions (Ouyang et al., 2023; Zhao et al., 2024). These findings suggest that aboveground biodiversity in forests plays a significant role in maintaining and enhancing both individual ecosystem functions and multifunctionality (Felipe-Lucia et al., 2018). In addition, the forest physical structure complexity (e.g., variation in breast height diameter and tree height) plays an important role in regulating ecosystem functions and is closely linked to biodiversity (Ehbrecht et al., 2021). Therefore, recent studies have focused on evaluating ecosystem multifunctionality and the relationship between biodiversity and stand structural diversity, as well as quantifying the ecosystem multifunctionality of forest structures (Pasari et al., 2013; Xie et al., 2018). Accordingly, a theoretical foundation for the main control factors affecting ecosystem multifunctionality has been established based on various research results (Yuan et al., 2019; Yuan et al., 2020).
Topographic factors, including elevation and slope, influence local microclimatic conditions, hydrological dynamics, and soil physical characteristics. In parallel, climatic factors such as precipitation and humidity are linked to water stress, which in turn plays a critical role in shaping plant species composition, growth patterns, and spatial distribution, ultimately regulating the physiological life cycles of plants (Becknell and Powers, 2014; Jucker et al., 2018). Moreover, biodiversity, one of the primary biotic factors, is widely acknowledged as a crucial driver that promotes resource-use efficiency through interspecific interactions, thereby supporting the maintenance and enhancement of EMF (Felipe-Lucia et al., 2018). In addition, research results have been reported that stand structural diversity, such as differences in canopy height and spatial arrangement of individual trees, is closely related to biodiversity and plays an important role in controlling ecosystem functions and maintaining stability (Ehbrecht et al., 2021). These findings collectively illustrate the diverse ways in which biotic factors shape EMF, providing the conceptual foundation for two main hypotheses proposed in previous studies: (1) niche complementarity and (2) the mass ratio hypothesis. Niche complementarity suggests that higher biodiversity leads to more efficient resource use among coexisting species, thereby enhancing the provision of ecosystem multifunctionality (Ali, 2019). Conversely, the mass ratio hypothesis suggests that EMF is primarily determined by the functional traits of dominant species, typically represented by the community-weighted mean (CWM) (Grime, 1998).
As previously explained, EMF is shaped by a range of interacting drivers, and the relative influence of these factors may differ across forest stand types, including coniferous, broadleaved, and mixed stands. This variation is due to differences in the dominance of tree functional traits across forest stand types (Lan et al., 2023). Therefore, it is important to elucidate the complex mechanisms by which these factors control EMF across different forest stand types; however, research in this area remains limited (Lan et al., 2023; Ding et al., 2024). Accordingly, this study aims to quantify ecosystem multifunctionality in three forest types (coniferous, broadleaved and mixed stands) and the entire forest using data from the 7th National Forest Inventory (NFI) of South Korea, and to identify the controlling mechanisms of biotic factors (tree biodiversity, stand structure diversity, and community-weighted mean of tree functional traits), abiotic factors (topography, climate), and factors related to forest succession stages (stand age) that control it.
To this end, this study established the following three hypotheses based on the conceptual model (Figure 1). (1) The biodiversity and stand structural diversity of trees are essential control factors for improving EMF. Increased tree diversity within forest stands is associated with improved ecosystem functions, such as more efficient resource utilization and greater carbon sequestration. Thus, structural heterogeneity at both the species and stand level is likely to promote higher levels of EMF (Ali, 2019; Yuan et al., 2020; Li et al., 2023). (2) Climatic factors represent key abiotic factors of EMF, with water stress in particular exerting direct influence on essential functions such as photosynthesis, plant growth, and hydrological processes. Accordingly, water stress is expected to be strongly associated with variations in EMF (Zhang et al., 2024). Finally, (3) Variations in species composition among forest stand types, along with associated differences in soil nutrient availability and water-use strategies, are likely to influence EMF and its underlying regulatory drivers (Tian et al., 2024).
FIGURE 1
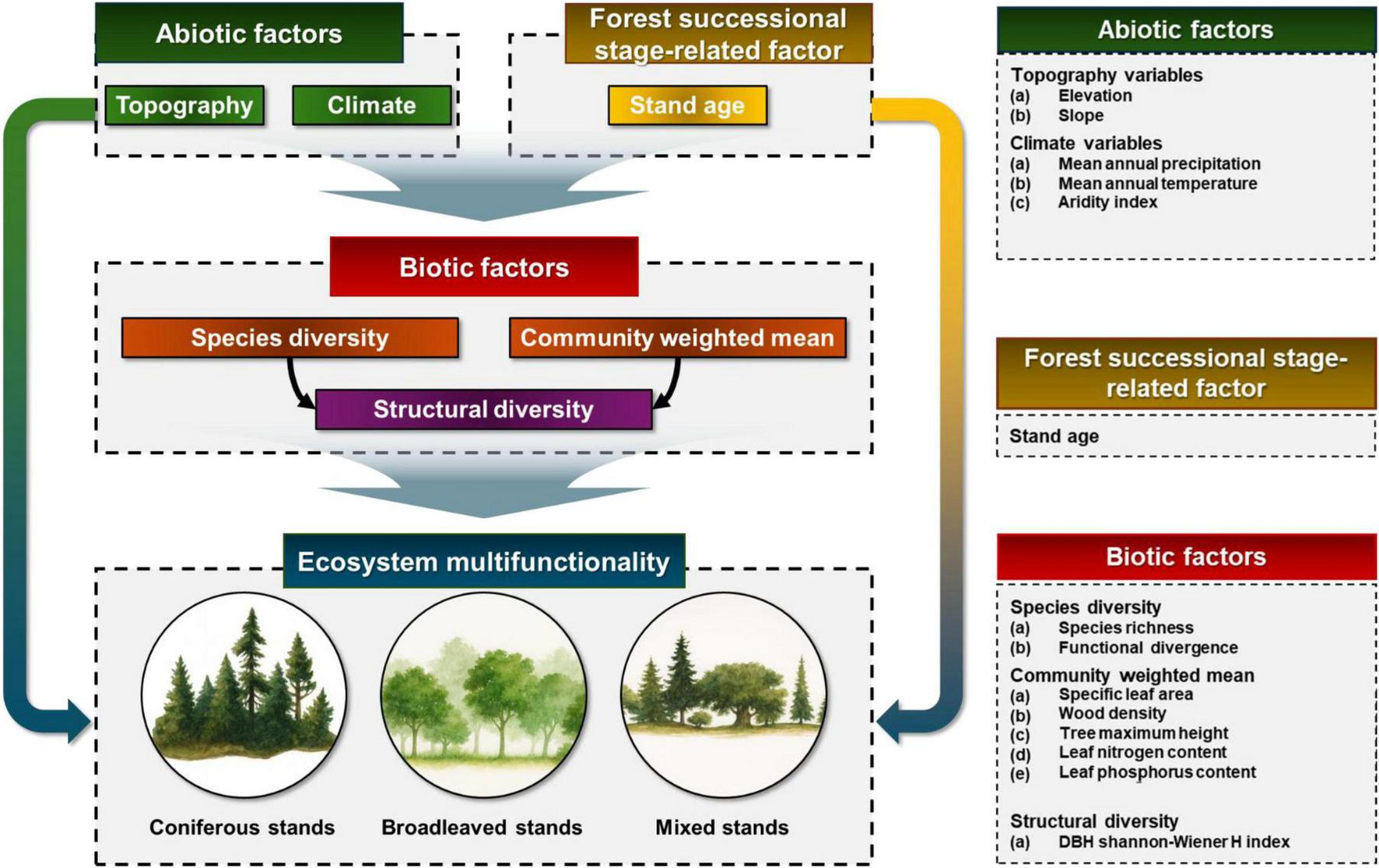
Conceptual model describing the biotic, abiotic, and forest successional stage-related factors that determine the ecosystem multifunctionality (EMF) index among forest stand types in temperate forests of South Korea. Based on this model, we test causal relationships among factors, and evaluate their direct and indirect effects on the EMF.
2 Materials and methods
2.1 Study sites and data acquisition
This study was conducted across the entire forested area of South Korea. As of 2020, forests covered approximately 62.6% of the national territory, making South Korea the country with the fourth-highest forest-to-land ratio among OECD member states (Korea Forest Service, 2022). The country extends longitudinally from north to south, encompassing a wide range of climatic zones that shape diverse vegetation distributions. Most regions fall within the temperate forest zone, with dominant species including oaks (Quercus spp.) and Korean red pine (Pinus densiflora Siebold & Zucc.) (Yun et al., 2011). Areas south of 35°N latitude lie within the warm-temperate to subtropical zones and are characterized by species such as Camellia japonica L. and Quercus acuta Thunb. (Yun et al., 2011). In addition, due to vertical vegetation zonation reaching elevations up to 1,305 meters, subalpine to alpine species become more prominent at higher elevation, including Abies nephrolepis (Trautv. ex Maxim.) Maxim. and Betula ermanii Cham. (Lee et al., 2024b). The mean annual temperature ranges from 5.88 °C to 15 °C (mean: 11.18 °C), and the annual precipitation ranges from 939 mm to 2,095 mm (mean: 1,329 mm) (Korean Meteorological Administration, 2021).
The National Forest Inventory (NFI) of South Korea has been conducted over seven cycles since its initiation in 1972. While the 1st to 4th inventories were carried out at irregular intervals of 2–7 years, the 5th inventory (2005–2010) marked the beginning of regular surveys conducted every 5 years (Korea Forest Service, 2017). In this study, we utilized data from the 7th NFI (2016–2020) to analyze the factors controlling EMF. The NFI employs a systematic sampling design, in which sample points are distributed across all forested areas nationwide at regular 4 km × 4 km intervals (2 km or 1 km for areas with small forest areas), using an arbitrary origin point (TM coordinates: X = 200,000, Y = 500,000) as a reference (Korea Forest Service, 2017). Among these grid points, those located within forested areas are designated as fixed sample plots and are subjected to field surveys for data collection. Each fixed sample plot is designed as a cluster plot composed of four circular subplots. One central plot is established at the origin point, and three additional plots are placed at 0° (true north direction), 120°, and 240° directions, each located 50 meters from the center. Nationwide, approximately 4,500 cluster plots have been installed, yielding a total of around 16,000 subplots (Supplementary Figure 1). Each subplot is a circular plot with a radius of 11.3 meters, equivalent to an area of approximately 0.04 ha. In this study, we selected 2,859 subplots from the central plot of each cluster in the 7th NFI, excluding those with missing biotic, abiotic, or stand age data or where ecosystem functions could not be assessed. In each subplot, all tree individuals with a diameter ≥6 cm were identified and measured for species, diameter, and height. Additionally, for understory vegetation, three 2 m × 2 m square quadrats (totaling 12 m2) were established within each circular subplot to count all herbaceous species and seedlings.
In addition, species composition, stand structural characteristics, and biodiversity vary across forest stand types, and such differences can have distinct effects on individual ecosystem functions and overall EMF (Matsuo et al., 2021; Lee et al., 2023). Therefore, in this study, all sample plots were categorized into three forest stand types to account for stand-specific differences: 660 coniferous plots (23.08%), 1,354 broadleaved plots (47.36%), and 845 mixed forest plots (29.56%) (Figure 2).
FIGURE 2
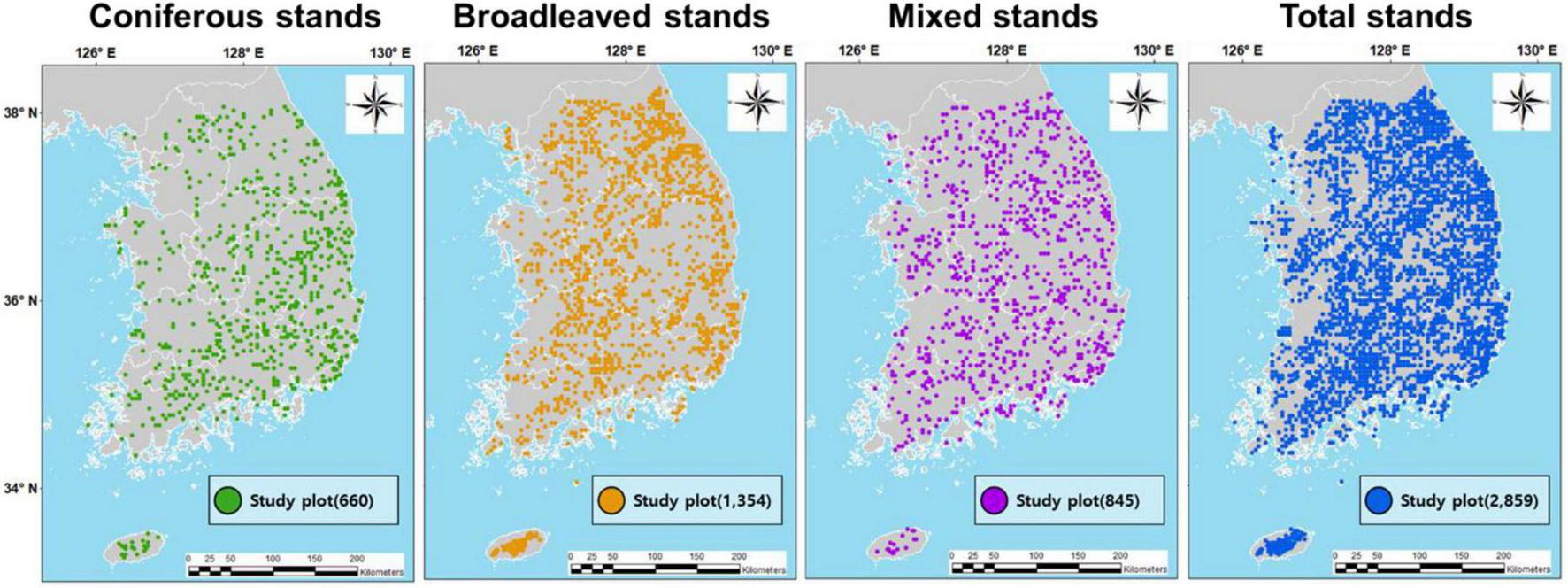
Location and distribution of 2,859 study plots consisting of 660 (23.08%), 1,354 (47.36%), and 845 (29.56%) plots of coniferous, broadleaved, and mixed stands, respectively, in temperate forests of South Korea.
The final dataset and summary statistics are provided in Supplementary Tables 1, 2.
2.2 Calculation of the ecosystem multifunctionality (EMF) index
Based on the classification criteria and case examples of ecosystem functions and services presented by the Millennium Ecosystem Assessment (Millenium Ecosystem Assessment [MEA], 2005) and Garland et al. (2021), we selected forest ecosystem functions for calculating the EMF index. A total of 11 ecosystem functions were selected, each representing a service from one of four ecosystem service categories (Table 1). In the provisioning service category, we quantified water and wood provision functions using water resources content and wood production indices. We also used the population of plants registered as edible and medicinal to represent the provisioning of food and medicinal production (Millenium Ecosystem Assessment [MEA], 2005). For the regulating service category, we selected variables representing atmospheric carbon capture functions (e.g., aboveground biomass, soil organic carbon), ultimately utilizing factors representing climate regulation functions (Lee et al., 2024a). Additionally, we utilized the forest disaster prevention index, which represents landslide and erosion prevention functions, and the soil bulk density, which represents soil structure (Temme, 2021). In the cultural services category, the forest recreation index, which represents potential forest recreation functions, and the population of ornamental plants were used (Garland et al., 2021; Roh et al., 2024). Finally, in the supporting services category, we utilized soil nitrogen content, a function related to plant growth and photosynthetic efficiency (Singh et al., 2022). Detailed descriptions of the data sources and quantification methods for each function are provided in Supplementary Material 2. To calculate the EMF index, we employed the averaging approach, which has been widely used in many previous studies (Byrnes et al., 2014). Although various methodologies, such as the threshold and multiple threshold approaches, have been introduced, the threshold approach has a drawback in that the choice of threshold is arbitrary (Byrnes et al., 2014). In addition, the multiple threshold approach also has limitations, as it produces a suite of metrics rather than a single simple value and provides only phenomenological patterns, making it necessary to compare with individual functions for mechanism interpretation (Byrnes et al., 2014). The averaging approach is considered the most intuitive method for integrating multiple ecosystem functions and has been widely applied in many studies (Wang et al., 2022; Wang et al., 2025) (Equation 1). It is calculated as follows (Maestre et al., 2012; Byrnes et al., 2014).
TABLE 1
| Service category | Ecosystem functions | Variables related to ecosystem functions | Unit | Data source |
| Provisioning | Food production | Number of edible plant population | – |
Korea Forest Service, 2017
Roh et al., 2024 |
| Medicinal production | Number of medicinal plant population | – | ||
| Water resources content | Water provision index | – | ||
| Wood production | Wood provision index | – | ||
| Regulating | Aboveground carbon storage | Above ground biomass | kg |
Korea Forest Service, 2017
Poggio et al., 2021 Roh et al., 2024 |
| Soil carbon storage | Soil organic carbon stock | t/ha | ||
| Soil structure stability | Soil bulk density | cg/cm3 | ||
| Landslide and erosion control | Forest disaster prevention index | – | ||
| Cultural Supporting |
Aesthetic value | Number of ornamental plant population | – |
Korea Forest Service, 2017
Roh et al., 2024 Poggio et al., 2021 |
| Providing forest recreation space | Forest recreation index | – | ||
| Plant growth and photosynthetic efficiency | Soil nitrogen content | cg/kg |
Ecosystem functions and data sources used to calculate ecosystem multifunctionality in this study.
where, F represents the number of measured ecosystem functions, and fi denotes the value of function i. The function g is a standardization formula used to rescale all function values to a standard scale between 0 and 1 (Byrnes et al., 2014). The EMF index was calculated using the multifunc package in R version 4.3.3 (R Development Core Team, 2023).
2.3 Quantification of biotic factors
We used key biotic factors—namely, taxonomic and functional diversity, stand structural diversity of trees, and community-weighted mean (CWM) values of functional traits—to assess their influence on EMF. Functional and structural diversity were derived from tree-level biological traits and stand structural attributes and thus were treated as biotic factors. Taxonomic diversity was measured using tree species richness (SR), defined as the number of species within each plot. Functional diversity was quantified using functional divergence (FDiv), an index that captures the degree of variation in tree functional traits within a community. To evaluate both FDiv we selected and measured five functional traits of trees. These traits included specific leaf area (SLA, mm2 g−1), leaf phosphorus content (P, mg g−1), leaf nitrogen content (N, mg g−1), maximum tree height (MH, m), and wood density (WD, g cmł). These traits are closely associated with plant growth, nutrient acquisition and use, and competitive ability (Wright et al., 2004; Anderegg et al., 2018). Trait data for the observed species were obtained through direct leaf and wood sampling and analysis conducted by Dr. Lee’s laboratory, following standardized procedures and publicly available trait databases (Perez-Harguindeguy et al., 2016). CWM values for five traits were calculated as the mean trait value of each plot, weighted by the relative basal area of each species (Equation 2). The calculation formula is presented as follows.
where, CWMx is the CWM for trait x, n is the total number of species in the study area, and pi and ti represent the relative breast height area and trait values of species i in the study area, respectively. FDiv and CWM of each study area were calculated using the FD package in R version 4.3.3 (R Development Core Team, 2023; Laliberté et al., 2014). To quantify the stand structural diversity and complexity of each plot, DBH diversity (DBH.Div) was calculated. This variable was derived using the Shannon-Wiener diversity index (Lee et al., 2024b). The DBH of all trees in the study area was divided into size classes with 5-cm intervals (e.g., 0–5 cm, 5–10 cm, etc., in DBH class). Then, the DBH diversity was derived using the number of individuals in each size class by applying the Shannon-Wiener index.
2.4 Quantification of abiotic factors and factors related to forest succession stages
To assess the influence and relative importance of abiotic factors and succession-related variables on EMF, we extracted and calculated topographic, climatic, and stand age factors. Topographic factors represent physical characteristics that directly affect both biotic and abiotic processes, such as vegetation distribution, soil moisture, and microclimate (Yuan et al., 2020). In this study, we derived elevation and slope from a digital elevation model (DEM) provided by the National Geographic Information Institute of South Korea (Kopecký et al., 2021).
Climatic factors were extracted using the Digital Climate Maps (DCM) developed by the Korea Meteorological Administration’s National Center for Agro-Meteorology, which provide spatial data at a 30 m resolution (Yun, 2010; Lee et al., 2021; Lee et al., 2023). From these maps, we obtained mean annual precipitation (MAP) and mean annual temperature (MAT). Furthermore, to assess vegetation water stress, we calculated the Aridity index, a humidity-related index based on evapotranspiration and MAP. These indices were derived from global climate datasets with a 1 km resolution provided by WorldClim.1 All topographic and climatic variables were processed using ArcGIS 10.5.
Stand age, a proxy for forest successional stage, was calculated as the average tree age of the five dominant individuals per plot, which were directly measured by increment coring at breast height as part of the NFI survey (Lee et al., 2024b).
2.5 Statistical analysis
Before conducting statistical analyses, all dependent and independent variables used in this study were log- or square-root-transformed and standardized to improve linearity and normality. To minimize multicollinearity among variables, those with an absolute correlation coefficient greater than 0.65 were excluded from the analysis. |r| ≥ 0.7 is typically used to eliminate multicollinearity among variables (Lee et al., 2024a). However, because our model represents an overall ecosystem that includes various variables, including abiotic and biotic factors, we adopted a more conservative criterion (|r| ≥ 0.65) to minimize potential collinearity among predictor variables. As a result, MAT was excluded due to high correlations with elevation. Similarly, MAP and CWM.N for all forest stands except broadleaf stands were excluded due to strong correlations with Aridity and CWM.P, respectively. Furthermore, FDiv in coniferous stands was strong correlated with SR and was therefore excluded from further analysis (Supplementary Figure 2).
To assess the presence of spatial autocorrelation, we employed generalized least squares (GLS) modeling (Legendre and Legendre, 2012). Specifically, we compared the model fit between a spatial GLS model that included the geographic coordinates of each plot and a non-spatial GLS model that did not incorporate spatial information. Model fit was evaluated using the Akaike Information Criterion (AIC), and the results indicated that spatial autocorrelation had a significant effect in both coniferous and mixed stands (Supplementary Table 3). Therefore, in subsequent analyses, we used a spatial GLS model that included geographic coordinates in both coniferous and mixed stands. The multimodel inference (MuMIn) approach was applied to identify topographic and CWM factors that significantly influence EMF (Lee et al., 2023; Ouyang et al., 2023) (Supplementary Figure 3). This method determines the most influential predictors of EMF based on standardized regression coefficients (β). Before model evaluation, we calculated variance inflation factors (VIFs) to assess multicollinearity in the multiple regression models. In general, VIF values greater than 10 indicate that multicollinearity may reduce the model’s explanatory power (Graham, 2003). However, in this study, factors with VIF values exceeding 3 were excluded to eliminate the potential influence of multicollinearity. As a result, the VIF for total stands CWM.SLA exceeded 3 and were therefore excluded from subsequent analyses. Subsequently, the VIF values for all models remained below 3, indicating that multicollinearity among independent variables did not affect the model results.
To structurally represent the causal relationships among various factors based on the conceptual model, we employed piecewise structural equation modeling (pSEM) (Lefcheck, 2016). Using pSEM, we analyzed both the direct and indirect effects, as well as the causal pathways, between EMF and biotic, abiotic factors, and stand age. The initial model was constructed as a comprehensive structure including all possible pathways. It was then optimized by removing statistically non-significant paths identified through d-separation tests (Lee et al., 2024b). Model fit was evaluated using Fisher’s C statistic, p-values, and Akaike Information Criterion (AIC).
Multimodel inference and pSEM analyses were conducted using the MuMIn and piecewiseSEM packages in R version 4.5.1 (R Development Core Team, 2023; Burnham and Anderson, 2004; Lefcheck, 2016).
3 Results
The results of the piecewise structural equation modeling (pSEM) revealed that, in coniferous stands, maximum tree height (CWM.MH), and stand structural diversity (DBH_Div) had direct positive effects on ecosystem multifunctionality (EMF) (Figure 3a). In addition, elevation affected EMF indirectly through CWM.MH, while stand age, species richness, and CWM.MH further exerted an indirect effect on EMF via stand structural diversity. The model evaluation statistics for coniferous stands were AIC = 5099.65, Fisher’s C = 8.14, df = 10, and P = 0.616, indicating a reasonably good balance between parsimony and model fit based on AIC, and a statistically acceptable fit to the data based on Fisher’s C and p-values.
FIGURE 3
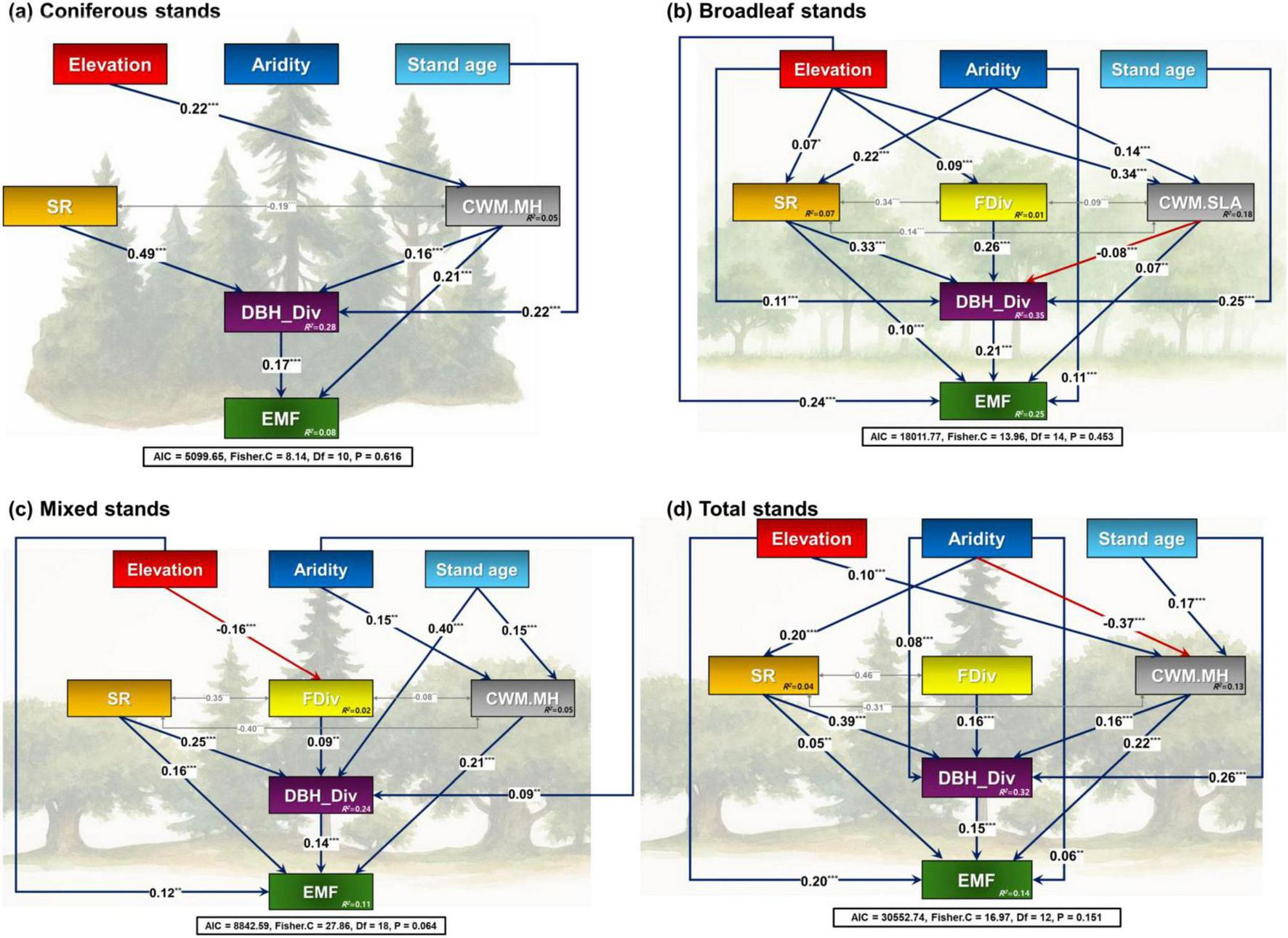
Structural equation models accounting for the effects of biotic, abiotic, and forest successional stage-related factors for ecosystem multifunctionality in panel (a) coniferous, (b) broadleaved, (c) mixed, and (d) total stands of temperate forests in South Korea. Solid blue and red lines represent positive and negative effects, respectively. Dotted two-way arrows indicate the covariance between two variables. Standardized coefficients are shown for each path. Statistics to evaluate the goodness of fit for the structural equation models are provided. Aridity, aridity index; SR, species richness; FDiv, functional divergence; DBH_Div, diameter at breast height diversity index; CWM, community weighted mean; SLA, specific leaf area; MH, mature tree height; EMF, ecosystem multifunctionality; AIC, Akaike Information Criterion; Fisher’s C, Fisher chi-square; Df, degrees of freedom.
In the broadleaved stand, pSEM results showed that elevation, aridity, species richness, specific leaf area (CWM.SLA), and stand structural diversity had direct positive effects on EMF (Figure 3b). Also, elevation, stand age, species richness, FDiv, and CWM.SLA indirectly controlled EMF through stand structural diversity. Furthermore, elevation and aridity appeared to control EMF through species richness and CWM.SLA indirectly. The model evaluation statistics for broadleaved stands were AIC = 18011.77, Fisher’s C = 13.96, df = 14, and P = 0.453, indicating a good model fit.
In mixed stands, elevation, species richness, maximum tree height (CWM.MH), and stand structural diversity directly enhanced EMF (Figure 3c). Aridity, stand age, species richness, and FDiv indirectly controlled EMF through stand structural diversity. Aridity and stand age further exerted indirect effects on EMF through CWM.MH. The model evaluation statistics for mixed stands were AIC = 8842.59, Fisher’s C = 27.86, df = 18, and P = 0.064, suggesting acceptable parsimony and fit.
The total forest stand types, pSEM results showed that elevation, aridity, species richness, maximum tree height (CWM.MH), and stand structural diversity directly increased EMF (Figure 3d). Aridity, stand age, species richness, FDiv, and CWM.MH indirectly controlled EMF through stand structural diversity. Elevation and stand age were found to influence EMF through CWM.MH, and aridity indirectly influenced EMF through species richness and CWM.MH. The total forest stand type evaluation statistics were AIC = 30552.74, Fisher’s C = 16.97, df = 12, and P = 0.151, indicating a well-fitting and statistically appropriate model.
In addition, bivariate relationships between EMF and individual controlling factors by forest type, as well as the results of multimodel inference and relative importance analysis, supported the pSEM results (Figure 4 and Supplementary Figures 4–7).
FIGURE 4
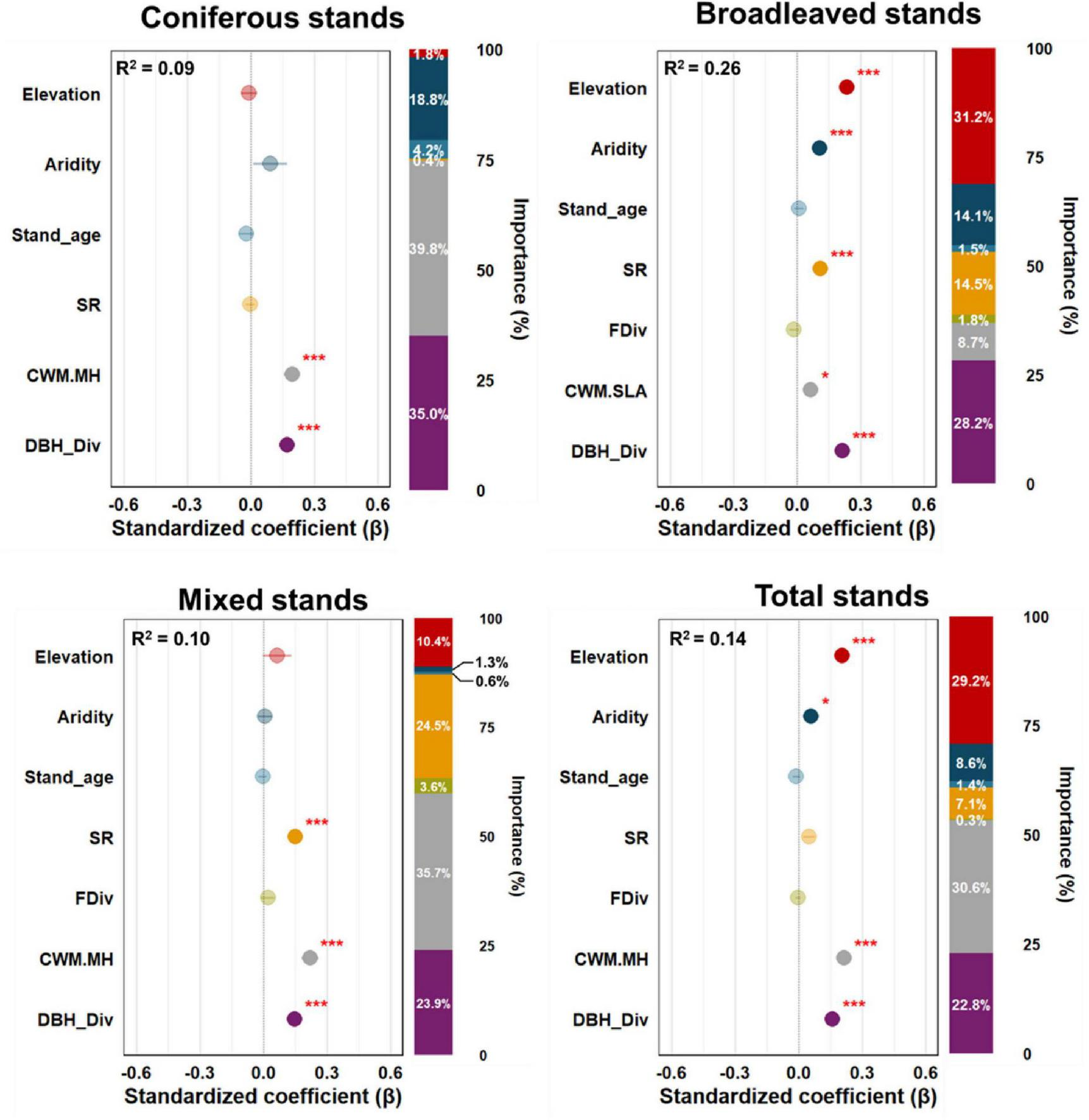
Standardized parameter estimates of the effects size (circle) and standard error (bar) of biotic, abiotic, and forest successional stage-related factors on ecosystem multifunctionality in panel coniferous, broadleaved, mixed, and total stands of temperate forests in South Korea. The closed and open circles indicate significant (P < 0.05) and nonsignificant relationships, respectively. The relative importance of each factor was calculated as the ratio between the parameter estimate of the factor and the sum of all parameters estimates and then presented as a percentage. Abbreviations for the variables are shown in Figure 3.
4 Discussion
In this study, based on the analysis of the relationships between EMF in each forest stand type and the biotic, abiotic factors, and stand ages that control it, we will discuss three major perspectives to understand the mechanisms that maintain and regulate EMF. (1) Higher plant species richness and older forests have higher stand structural diversity of the stand, which can better maintain EMF. (2) The results of abiotic factor analysis showed that factors related to water stress of plants had direct and indirect negative effects across total forest stand types. (3) The mechanisms that control EMF in each forest stand type were different. In the following, we will discuss the research results based on ecological theory.
4.1 Biotic factors controlling forest ecosystem multifunctionality
In this study, we confirmed that EMF across the entire forest is primarily controlled by species richness and stand structural diversity, elevation, and climatic conditions associated with water stress. In particular, stand age and species richness were found to indirectly enhance EMF by increasing stand structural diversity. Generally, species richness and structural diversity within a stand—such as variation in diameter at breast height and tree height—is strongly associated with key ecosystem functions, including carbon storage, nutrient cycling, and productivity (Zeller and Pretzsch, 2019). It also contributes to regulating or mediating the interactions between biotic and abiotic factors (Ali, 2019). Forest tree species differ in their specific requirements for water and light, which vary depending on their physical size and the characteristics of their growth environment (Ali, 2019). Therefore, high species richness and stand structural diversity can enhance the efficiency of light capture and resource utilization, thereby enhancing the benefits of ecological niche complementarity (Ali et al., 2016). It has been reported that increasing stand structural diversity within forest stands improves ecosystem functions like aboveground biomass production, and concurrently leads to greater diversity of coarse woody debris and leaf litter, thereby fostering microbial activity and enhancing nutrient supply (Lange et al., 2015; Sanaei et al., 2021). Similarly, a study conducted in the Changbai Mountain region of northeastern China also found that tree biodiversity and structural diversity increased EMF (Sanaei et al., 2021). Furthermore, increased biodiversity and structural complexity within forest stands have been shown to strengthen human–nature relationships by contributing to greater aesthetic appeal, recreational opportunities, and microhabitat heterogeneity (Gao et al., 2014; Pretzsch et al., 2024). Thus, increasing tree species richness—thereby enhancing stand structural diversity—appears to play a significant role in promoting EMF (Yuan et al., 2020). In addition, stand structural diversity gradually increases with stand age, which in turn enhances EMF. In forest ecosystems, stands progress through several successional stages over time, with early stages dominated by fast-growing, small-sized tree species, while later stages are characterized by slow-growing, large-sized tree species (Lienard et al., 2015). During this succession, the range of tree sizes changes, which ultimately has a positive impact on ecosystem function and EMF (Matsuo et al., 2021).
4.2 Abiotic factors controlling forest ecosystem multifunctionality
Our findings indicate that abiotic control factors play a critical role in determining EMF. Specifically, aridity—a key climatic variable—was found to influence EMF both directly and indirectly across all forest stand types, except in coniferous stands. Aridity used in this study refers to a dryness index, where a higher value indicates a more humid climate (Zomer et al., 2022). In this study, EMF increased as Aridity increased, suggesting that EMF decreased as water stress increased. These findings align with those of previous studies and can be attributed to two underlying mechanisms: enhanced resource availability and the mitigation of water stress (Guo et al., 2023; Zhang et al., 2024). As precipitation and moisture levels increase, the resources available to plant species increase, which strengthens the complementarity of different ecological niches of species, improves productivity, and ultimately enhances ecosystem function (Guo et al., 2023). In addition, increased soil moisture and the alleviation of water stress promote plant growth and stimulate soil microbial activity, thereby enhancing ecosystem functions such as soil carbon and nitrogen storage, as well as habitat provision (Zhang et al., 2024). Accordingly, abiotic factors associated with forest moisture availability can directly contribute to the improvement of EMF. Conversely, under intensified water stress, plant survival and reproduction may be compromised, and species interactions can be disrupted, ultimately diminishing ecosystem stability (Hu et al., 2021). Such conditions may inhibit fundamental ecosystem functions—including carbon uptake, soil nutrient cycling, moisture retention, and wood production—thereby leading to a decline in EMF (Garland et al., 2021).
The results of this study further demonstrated that EMF increases with elevation across all forest types. This pattern can largely be explained by the elevation–climate relationship, in which temperature and evapotranspiration decrease with elevation, thereby alleviating water stress (Zhang et al., 2024). Consistent with this, our findings showed that MAT declined with elevation, whereas aridity and MAP increased (Supplementary Figure 2). These results suggest that EMF enhancement along elevational gradients is mediated by mechanisms similar to those through which aridity regulates EMF. In addition, plant species adapted to high-elevation environments are typically specialized for extreme conditions, and their functionally complementary roles contribute to maintaining ecosystem stability (Lefcheck et al., 2015). This indicates that interspecific complementarity becomes a critical factor for sustaining EMF at high elevation. Overall, the positive relationship between elevation and EMF revealed in this study can be interpreted as the combined outcome of climatic stress alleviation and the functional complementarity of high-elevation vegetation assemblages.
4.3 Factors controlling ecosystem multifunctionality by forest stand types
In this study, EMF was found to increase with the community weighted mean of maximum tree height (CWM.MH) in coniferous and mixed stands. In coniferous stands, taller trees generally indicate greater stand maturity and biomass accumulation, which directly enhance ecosystem functions related to long-term carbon storage (Stephenson et al., 2014). Furthermore, in mixed stands, increases in CWM.MH not only reflects increased biomass but also contributes to enhanced vertical stratification, which is expected to promote resource allocation and functional complementarity among species. In particular, mixed stands are forests where conifers and broad-leaved trees coexist, so the more species with high maximum height dominate, the more space can be divided through crown plasticity to optimize stratification (Jucker et al., 2015). In other words, CWM.MH increases, mixed stands develop multi-layered canopies that create diverse microhabitats and light environments, supporting a greater diversity of understory species and enhancing functional complementarity.
On the other hand, in broadleaf stands, EMF was found to increase as the community weighted mean of specific leaf area (CWM.SLA) increased. Broadleaf trees generally have thin and broad leaves, and many species exhibit greater variation and higher SLA than conifers. Therefore, in broadleaf forests, SLA appears to be a key indicator explaining functional differences between species, and thus, a clear relationship with EMF appears. Species with high SLA generally exhibit an acquisitive resource utilization strategy characterized by rapid growth, high photosynthetic rates, and efficient light capture (Reich, 2014). Therefore, it appears that higher SLA enhances primary productivity and nutrient cycling, directly contributing to increased EMF.
This study observed that, unlike other stands, aridity did not significantly affect EMF, either directly or indirectly, in coniferous stands. Compared to broadleaf trees, conifers have relatively low transpiration rates and thick leaf structures, leading to high water use efficiency (Choat et al., 2012). Therefore, even in arid environments, they are less sensitive to water stress than broadleaf trees, suggesting that aridity has a relatively limited impact on functional characteristics and EMF. Notably, coniferous stands in Korea are predominantly comprised of pine (Pinus densiflora) (25% of all Korean forests), and pine is a representative tree species with high drought tolerance, enabling it to maintain growth even in harsh environments (Park et al., 2016). These characteristics likely contributed to the limited impact of aridity in Korean coniferous forests. This suggests that the relative importance of aridity varies across stand types, and that, particularly in coniferous forests, ecological constraints other than water stress drive EMF fluctuations. This suggests the need to consider forest stand types of adaptation strategies in climate change scenarios.
5 Conclusion
This study revealed that biotic and abiotic factors, together with stand age, influence EMF through distinct pathways across coniferous, broadleaved, and mixed forest stand types. These findings underscore the importance of forest management strategies that facilitate smooth ecological succession, conserve biodiversity, and enhance stand structural diversity to promote EMF, ecosystem quality, and long-term stability. In particular, coniferous stands benefit from preserving large trees to secure carbon storage, broadleaved stands from maintaining species with high specific leaf area to enhance productivity, and mixed forests from fostering vertical stratification and functional complementarity. Therefore, (1) in coniferous stands, large trees with high maximum height should be preserved to enhance long-term carbon storage and structural stability. Management should place greater emphasis on addressing ecological constraints such as disturbance and pests rather than water stress. (2) In broadleaved stands, maintaining and expanding the proportion of species with high specific leaf area can enhance productivity and nutrient cycling, while long-term EMF can be supported through the accumulation of soil organic matter and the promotion of microbial community activity. (3) In mixed forests, vertical stratification and functional complementarity among species should be strengthened by incorporating tall tree species that promote canopy plasticity and a multi-layered structure. Beyond these management implications, EMF contributes directly to human well-being through climate regulation and community resilience. Future research should be conducted to maintain and preserve EMF’s spatiotemporal stability under extreme external disturbances such as climate change and land use change. Also, this study did not account for other biotic groups (e.g., insects and microorganisms), anthropogenic and climatic drivers (e.g., forest fires, logging, rainfall, soil type), or historical disturbances (e.g., past management activities). Future research need to incorporate these factors to provide a more comprehensive understanding of EMF in forest ecosystems. Such efforts will help refine sustainable forest management strategies that simultaneously enhance EMF and human welfare.
Statements
Data availability statement
The original contributions presented in this study are included in this article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
M-KL: Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft. J-HC: Funding acquisition, Investigation, Methodology, Writing – original draft. C-BL: Conceptualization, Funding acquisition, Methodology, Project administration, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was conducted with support from the R&D Program for Forest Science Technology provided by Korea Forest Service (Korea Forestry Promotion Institute) (Project No. RS-2024-00404816), National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (Project No. RS-2024-00358413), and the National Institute of Forest Science, Republic of Korea (Project No. FE0100-2021-01).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The authors declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2025.1675167/full#supplementary-material
Footnotes
References
1
Ali A. (2019). Forest stand structure and functioning: Current knowledge and future challenges.Ecol. Indic.98665–677. 10.1016/j.ecolind.2018.11.017
2
Ali A. (2023). Biodiversity–ecosystem functioning research: Brief history, major trends and perspectives.Biol. Conserv.285:110210. 10.1016/j.biocon.2023.110210
3
Ali A. Yan E. R. Chen H. Y. Chang S. X. Zhao Y. T. Yang X. D. et al (2016). Stand structural diversity rather than species diversity enhances aboveground carbon storage in secondary subtropical forests in Eastern China.Biogeosciences134627–4635. 10.5194/bg-13-4627-2016
4
Anderegg W. R. L. Konings A. G. Trugman A. T. Yu K. Bowling D. R. Gabbitas R. et al (2018). Hydraulic diversity of forests regulates ecosystem resilience during drought.Nature561538–541. 10.1038/s41586-018-0539-7
5
Becknell J. M. Powers J. S. (2014). Stand age and soils as drivers of plant functional traits and aboveground biomass in secondary tropical dry forest.Can. J. For. Res.44604–613. 10.1139/cjfr-2013-0331
6
Burnham K. P. Anderson D. R. (2004). Multimodel inference: Understanding AIC and BIC in model selection.Sociol. Methods Res.33261–304. 10.1177/0049124104268644
7
Byrnes J. E. Gamfeldt L. Isbell F. Lefcheck J. S. Griffin J. N. Hector A. et al (2014). Investigating the relationship between biodiversity and ecosystem multifunctionality: Challenges and solutions.Methods Ecol. Evol.5111–124. 10.1111/2041-210X.12143
8
Choat B. Jansen S. Brodribb T. J. Cochard H. Delzon S. Bhaskar R. et al (2012). Global convergence in the vulnerability of forests to drought.Nature491752–755. 10.1038/nature11688
9
Chun J. H. Ali A. Lee C. B. (2020). Topography and forest diversity facets regulate overstory and understory aboveground biomass in a temperate forest of South Korea.Sci. Total Environ.744:140783. 10.1016/j.scitotenv.2020.140783
10
Costanza R. d’Arge R. De Groot R. Farber S. Grasso M. Hannon B. et al (1997). The value of the world’s ecosystem services and natural capital.Nature387253–260. 10.1038/387253a0
11
DeAngelis D. L. (1980). Energy flow, nutrient cycling, and ecosystem resilience.Ecology61764–771. 10.2307/1936746
12
Dietrich P. Ebeling A. Meyer S. T. Asato A. E. B. Bröcher M. Gleixner G. et al (2024). Plant diversity and community age stabilize ecosystem multifunctionality.Glob. Chang Biol.30:e17225. 10.1111/gcb.17225
13
Ding L. J. Ren X. Y. Zhou Z. Z. Zhu D. Zhu Y. G. (2024). Forest-to-Cropland conversion reshapes microbial hierarchical interactions and degrades ecosystem multifunctionality at a national scale.Environ. Sci. Technol.5811027–11040. 10.1021/acs.est.4c01203
14
Ehbrecht M. Seidel D. Annighöfer P. Kreft H. Köhler M. Zemp D. C. et al (2021). Global patterns and climatic controls of forest structural complexity.Nat. Commun.12:519. 10.1038/s41467-020-20767-z
15
Felipe-Lucia M. R. Soliveres S. Penone C. Manning P. van der Plas F. Boch S. et al (2018). Multiple forest attributes underpin the supply of multiple ecosystem services.Nat. Commun.9:4839. 10.1038/s41467-018-07082-4
16
Gamfeldt L. Hillebrand H. Jonsson P. R. (2008). Multiple functions increase the importance of biodiversity for overall ecosystem functioning.Ecology891223–1231. 10.1890/06-2091.1
17
Gao T. Hedblom M. Emilsson T. Nielsen A. B. (2014). The role of forest stand structure as biodiversity indicator.For. Ecol. Manag.33082–93. 10.1016/j.foreco.2014.07.007
18
Garland G. Banerjee S. Edlinger A. Miranda Oliveira E. Herzog C. Wittwer R. et al (2021). A closer look at the functions behind ecosystem multifunctionality: A review.J. Ecol.109600–613. 10.1111/1365-2745.13511
19
Gonzalez A. Germain R. M. Srivastava D. S. Filotas E. Dee L. E. Gravel D. et al (2020). Scaling-up biodiversity-ecosystem functioning research.Ecol. Lett.23757–776. 10.1111/ele.13456
20
Grace J. B. Adler P. B. Stanley Harpole W. Borer E. T. Seabloom E. W. (2014). Causal networks clarify productivity–richness interrelations, bivariate plots do not.Funct. Ecol.28787–798. 10.1111/1365-2435.12269
21
Graham M. H. (2003). Confronting multicollinearity in ecological multiple regression.Ecology842809–2815. 10.1890/02-3114
22
Grime J. P. (1998). Benefits of plant diversity to ecosystems: Immediate, filter and founder effects.J. Ecol.86902–910. 10.1046/j.1365-2745.1998.00306.x
23
Guo H. Zhou X. Tao Y. Yin J. Zhang L. Guo X. et al (2023). Precipitation preferences alter the relative importance of herbaceous plant diversity for multifunctionality in the drylands of China. Front. Ecol. Evol.11:1084949.
24
Hector A. Bagchi R. (2007). Biodiversity and ecosystem multifunctionality.Nature448188–190. 10.1038/nature05947
25
Hu W. Ran J. Dong L. Du Q. Ji M. Yao S. et al (2021). Aridity-driven shift in biodiversity–soil multifunctionality relationships.Nat. Commun.12:5350. 10.1038/s41467-021-25641-0
26
Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services [IPBES] (2019). Global assessment report on biodiversity and ecosystem services of the intergovernmental science-policy platform on biodiversity and ecosystem services.Bonn: IPBES.
27
Jucker T. Bongalov B. Burslem D. F. R. P. Nilus R. Dalponte M. Lewis S. L. et al (2018). Topography shapes the structure, composition and function of tropical forest landscapes.Ecol. Lett.21989–1000. 10.1111/ele.12964
28
Jucker T. Bouriaud O. Coomes D. A. (2015). Crown plasticity enables trees to optimize canopy packing in mixed-species forests.Funct. Ecol.291078–1086. 10.1111/1365-2435.12428
29
Kopecký M. Macek M. Wild J. (2021). Topographic Wetness Index calculation guidelines based on measured soil moisture and plant species composition.Sci. Total Environ.757:143785. 10.1016/j.scitotenv.2020.143785
30
Korea Forest Service (2017). Guide to the seventh national forest resources Inventory and field survey of forest ecosystem health and vitality.Incheon: Korea Forest Service.
31
Korea Forest Service (2022). Korea forest service: Statistical yearbook of forest.Incheon: Korea Forest Service.
32
Korean Meteorological Administration (2021). Weather information. Available online at: https://data.kma.go.kr(accessed November 1, 2022).
33
Laliberté E. Legendre P. Shipley B. (2014). FD: Measuring functional diversity from multiple traits, and other tools for functional ecology. R Package Version 1.0-12.
34
Lan J. Lei X. He X. Gao W. Guo H. (2023). Multiple mechanisms drive biodiversity-ecosystem service multifunctionality but the dominant one depends on the level of multifunctionality for natural forests in northeast China.For. Ecol. Manag.542:121101. 10.1016/j.foreco.2023.121101
35
Lange M. Eisenhauer N. Sierra C. A. Bessler H. Engels C. Griffiths R. I. et al (2015). Plant diversity increases soil microbial activity and soil carbon storage.Nat. Commun.6:6707. 10.1038/ncomms7707
36
Lee M. K. Chun J. H. Lee C. B. (2021). Prediction of distribution changes of Carpinus laxiflora and C. tschonoskii based on climate change scenarios using MaxEnt model.Kor. J. Agricultural For. Meteorol.2355–67. 10.5532/KJAFM.2021.23.1.55
37
Lee Y. J. Lee C. B. Lee M. K. (2023). Tree size variation induced by stand age mainly regulates aboveground biomass across three major stands of temperate forests in South Korea.Front. For. Global Change6:1229661. 10.3389/ffgc.2023.1229661
38
Lee Y. J. Lee H. I. Lee C. B. Lee K. H. Kim R. H. Ali A. (2024a). Abiotic and stand age-induced changes in tree diversity and size inequality regulate aboveground biomass and soil organic carbon stock in temperate forests of South Korea.Catena237:107827. 10.1016/j.catena.2024.107827
39
Lee Y. J. Park G. E. Lee H. I. Lee C. B. (2024b). Stand age-driven tree size variation and stand type regulate aboveground biomass in alpine-subalpine forests, South Korea.Sci. Total Environ.915:170063. 10.1016/j.scitotenv.2024.170063
40
Lefcheck J. S. (2016). piecewiseSEM: Piecewise structural equation modelling in r for ecology, evolution, and systematics.Methods Ecol. Evol.7573–579. 10.1111/2041-210X.12512
41
Lefcheck J. S. Byrnes J. E. K. Isbell F. Gamfeldt L. Griffin J. N. Eisenhauer N. et al (2015). Biodiversity enhances ecosystem multifunctionality across trophic levels and habitats.Nat. Commun.6:6936. 10.1038/ncomms7936
42
Legendre P. Legendre L. (2012). Numerical ecology. Amsterdam: Elsevier.
43
Li F. Shi Z. Liu S. Xu G. Zhang M. Cao X. et al (2023). Soil properties and plant diversity co-regulate ecosystem multifunctionality of subalpine primary dark coniferous forest on the eastern Qinghai-Tibetan Plateau.Plant Soil493207–219. 10.1007/s11104-023-06222-0
44
Lienard J. Florescu I. Strigul N. (2015). An appraisal of the classic forest succession paradigm with the shade tolerance index.PLoS One10:e0117138. 10.1371/journal.pone.0117138
45
Maestre F. T. Castillo-Monroy A. P. Bowker M. A. Ochoa-Hueso R. (2012). Species richness effects on ecosystem multifunctionality depend on evenness, composition and spatial pattern.J. Ecol.100317–330. 10.1111/j.1365-2745.2011.01918.x
46
Manning P. Van Der Plas F. Soliveres S. Allan E. Maestre F. T. Mace G. et al (2018). Redefining ecosystem multifunctionality.Nat. Ecol. Evol.2427–436. 10.1038/s41559-017-0461-7
47
Matsuo T. Martínez-Ramos M. Bongers F. van der Sande M. T. Poorter L. (2021). Forest structure drives changes in light heterogeneity during tropical secondary forest succession.J. Ecol.1092871–2884. 10.1111/1365-2745.13680
48
Millenium Ecosystem Assessment [MEA] (2005). Ecosystems and human well-being:multiscale assessment, millennium ecosystemassessment series, 4.Washington, DC: Island Press.
49
Mori A. S. Lertzman K. P. Gustafsson L. (2017). Biodiversity and ecosystem services in forest ecosystems: A research agenda for applied forest ecology.J. Appl. Ecol.5412–27. 10.1111/1365-2664.12669
50
Newbold T. Hudson L. N. Hill S. L. Contu S. Lysenko I. Senior R. A. et al (2015). Global effects of land use on local terrestrial biodiversity.Nature52045–50. 10.1038/nature14324
51
Ouyang S. Gou M. Lei P. Liu Y. Chen L. Deng X. et al (2023). Plant functional trait diversity and structural diversity co-underpin ecosystem multifunctionality in subtropical forests.For. Ecosyst.10:100093. 10.1016/j.fecs.2023.100093
52
Park M. J. Yoon S. J. Yoon H. M. Jang H. N. Han S. H. Ahn J. A. et al (2016). Effects of artificial warming and precipitation manipulation on the physiological characteristics and growth of Pinus densiflora seedlings.J. Climate Change Res.79–17. 10.15531/ksccr.2016.7.1.9
53
Pasari J. R. Levi T. Zavaleta E. S. Tilman D. (2013). Several scales of biodiversity affect ecosystem multifunctionality.Proc. Natl. Acad. Sci. U. S. A.11010219–10222. 10.1073/pnas.1220333110
54
Perez-Harguindeguy N. Diaz S. Garnier E. Lavorel S. Poorter H. Jaureguiberry P. et al (2016). Corrigendum to: New handbook for standardised measurement of plant functional traits worldwide.Aus. J. Botany64715–716. 10.1071/BT12225_CO
55
Poggio L. De Sousa L. M. Batjes N. H. Heuvelink G. B. Kempen B. Ribeiro E. et al (2021). SoilGrids 2.0: Producing soil information for the globe with quantified spatial uncertainty. Soil7, 217–240.
56
Pretzsch H. Hilmers T. del Río M. (2024). The effect of structural diversity on the self-thinning line, yield level, and density-growth relationship in even-aged stands of Norway spruce.For. Ecol. Manag.556:121736. 10.1016/j.foreco.2024.121736
57
R Development Core Team (2023). R Version 4.2.3.Vienna: R Foundation for Statistical Computing.
58
Reich P. B. (2014). The world-wide ‘fast–slow’plant economics spectrum: A traits manifesto.J. Ecol.102275–301. 10.1111/1365-2745.12211
59
Roh Y. H. Song J. E. Park Y. S. Son E. H. Seong H. J. Kim H. H. (2024). Establishment of a new forest function assessment system and development of forest function classification maps.Kor. J. For. Sci.113482–492. 10.14578/jkfs.2024.113.4.482
60
Sanaei A. Ali A. Yuan Z. Liu S. Lin F. Fang S. et al (2021). Context-dependency of tree species diversity, trait composition and stand structural attributes regulate temperate forest multifunctionality.Sci. Total Environ.757:143724. 10.1016/j.scitotenv.2020.143724
61
Scherer-Lorenzen M. Gessner M. O. Beisner B. E. Messier C. Paquette A. Petermann J. S. et al (2022). Pathways for cross-boundary effects of biodiversity on ecosystem functioning.Trends Ecol. Evol.37454–467. 10.1016/j.tree.2021.12.009
62
Singh A. Hidangmayum A. Tiwari P. Singh B. N. Dwivedi P. (2022). “How the soil nitrogen nutrient promotes plant growth—a critical assessment,” in New and Future developments in microbial biotechnology and bioengineering, 99–118.
63
Stephenson N. L. Das A. J. Condit R. Russo S. E. Baker P. J. Beckman N. G. et al (2014). Rate of tree carbon accumulation increases continuously with tree size.Nature50790–93. 10.1038/nature12914
64
Sun W. Liu X. (2020). Review on carbon storage estimation of forest ecosystem and applications in China.For. Ecosyst.7:4. 10.1186/s40663-019-0210-2
65
Tansley A. G. (1935). The use and abuse of vegetational concepts and terms.Ecology16284–307. 10.2307/1930070
66
Temme A. J. (2021). Relations between soil development and landslides.Hydrogeol. Chem. Weather. Soil Form.177–185. 10.1002/9781119563952.ch9
67
Tian J. Ni T. Miao Z. Hao Y. Ma A. Dong L. et al (2024). Effects of biotic and abiotic factors on ecosystem multifunctionality of plantations.Ecol. Proc.13:44. 10.1186/s13717-024-00525-5
68
van der Plas F. Manning P. Soliveres S. Allan E. Scherer-Lorenzen M. Verheyen K. et al (2016). Biotic homogenization can decrease landscape-scale forest multifunctionality.Proc. Natl. Acad. Sci. U. S. A.1133557–3562. 10.1073/pnas.1517903113
69
Wang W. Bi S. Li F. Degen A. A. Li S. Huang M. et al (2025). Soil organic matter composition affects ecosystem multifunctionality by mediating the composition of microbial communities in long-term restored meadows.Environ. Microb.20:22. 10.1186/s40793-025-00678-6
70
Wang Y. Liu B. Zhao J. Ye C. Wei L. Sun J. et al (2022). Global patterns and abiotic drivers of ecosystem multifunctionality in dominant natural ecosystems.Environ. Int.168:107480. 10.1016/j.envint.2022.107480
71
Wright I. J. Reich P. B. Westoby M. Ackerly D. D. Baruch Z. Bongers F. et al (2004). The worldwide leaf economics spectrum.Nature428821–827. 10.1038/nature02403
72
Xie H. Wang G. G. Yu M. (2018). Ecosystem multifunctionality is highly related to the shelterbelt structure and plant species diversity in mixed shelterbelts of eastern China.Glob. Ecol. Conserv.16:e00470. 10.1016/j.gecco.2018.e00470
73
Yuan Z. Ali A. Jucker T. Ruiz-Benito P. Wang S. Jiang L. et al (2019). Multiple abiotic and biotic pathways shape biomass demographic processes in temperate forests.Ecology100:e02650. 10.1002/ecy.2650
74
Yuan Z. Ali A. Ruiz-Benito P. Jucker T. Mori A. S. Wang S. et al (2020). Above-and below-ground biodiversity jointly regulate temperate forest multifunctionality along a local-scale environmental gradient.J. Ecol.1082012–2024. 10.1111/1365-2745.13378
75
Yun J. I. (2010). Agroclimatic maps augmented by a GIS technology. Korean J. Agricult. For. Meteorol.12:63.
76
Yun C. W. Kim H. J. Lee B. C. Shin J. H. Yang H. M. Lim J. H. (2011). Characteristic community type classification of forest vegetation in South Korea.J. Kor. Soc. For. Sci.100504–521.
77
Zeller L. Pretzsch H. (2019). Effect of forest structure on stand productivity in Central European forests depends on developmental stage and tree species diversity. For. Ecol. Manage.434, 193–204.
78
Zhang S. Chen Y. Zhou X. Zhang Y. (2024). Climate and human impact together drive changes in ecosystem multifunctionality in the drylands of China.Appl. Soil Ecol.193:105163. 10.1016/j.apsoil.2023.105163
79
Zhao X. Xu Y. Pu J. Tao J. Chen Y. Huang P. et al (2024). Achieving the supply-demand balance of ecosystem services through zoning regulation based on land use thresholds.Land Use Policy139:107056. 10.1016/j.landusepol.2024.107056
80
Zomer R. J. Xu J. Trabucco A. (2022). Version 3 of the global aridity index and potential evapotranspiration database.Sci. Data9:409. 10.1038/s41597-022-01493-1
Summary
Keywords
biotic and abiotic factors, ecosystem multifunctionality, forest stand type, stand age, structural diversity
Citation
Lee M-K, Chun J-H and Lee C-B (2025) Relative importance of biotic, abiotic and stand age factors in influencing ecosystem multifunctionality across forest stand types in South Korea. Front. For. Glob. Change 8:1675167. doi: 10.3389/ffgc.2025.1675167
Received
29 July 2025
Accepted
24 September 2025
Published
13 October 2025
Volume
8 - 2025
Edited by
Manoj Kumar Jhariya, Sant Gahira Guru Vishwavidyalaya, India
Reviewed by
Donald Mlambo, National University of Science and Technology, Zimbabwe
Bhimappa Kittur, Indian Institute of Forest Management, India
Updates
Copyright
© 2025 Lee, Chun and Lee.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chang-Bae Lee, kecolee@kookmin.ac.kr
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.