Abstract
Despite the vast area and large numbers of trees affected by drought- and bark beetle-induced tree mortality worldwide, relatively little is known about how post-mortality management practices affect forest recovery, particularly in forests historically adapted to frequent fire. Cutting and removing dead trees after a mass-mortality event provides an opportunity to salvage timber and lessen wildfire risk by reducing fuel loads, but the ecological impacts of this strategy extend beyond fuel reduction. A severe drought in California, USA (2012–2016) precipitated a mass die-off of conifers in the Sierra Nevada range. We examined how the management treatment of removing dead trees affected mixed-conifer forests in four key areas: fuels, tree regeneration, carbon stocks, and future fire behavior and severity. We collected data in 122 paired plots (22 m diameter) in treated and adjacent untreated areas spanning 300 km of the Sierra Nevada, one to 5 years after the removal of recently dead trees. We found that sound coarse woody debris mass was 51% higher with removal, but rotten coarse woody debris was not different. Litter depth and 1-h and 10-h fuel mass decreased with removal. Combined Pinus ponderosa and P. jeffreyi (shade-intolerant) seedling densities were 349% higher with removal, while P. lambertiana (moderately shade-tolerant) and Abies concolor (shade-tolerant) seedling densities were 64 and 55% lower with removal. Sapling density was 56% lower in treated plots but did not differ by species. Total forest carbon was 32% lower in treated sites, predominantly driven by dead-tree removal. Using the Forest Vegetation Simulator, we simulated future fire severity (% basal area mortality) and behavior (torch potential) over 100 years at 10-year intervals and predicted that after a 20-year lag, treatment persistently reduced simulated fire severity and behavior. Removal of dead trees initially enhanced regeneration of fire-adapted yellow pine species but negatively affected sugar pine regeneration, had varying effects on different fuel types, had predictable effects on on-site carbon stores, and lowered modeled future wildfire severity.
1 Introduction
Drought-induced mass tree mortality events are increasing due to a warming and drying climate (Bentz et al., 2010; Williams et al., 2013; Seidl et al., 2017) and forest densification (Young et al., 2019), with myriad consequences for ecosystems globally (Raffa et al., 2008; Allen et al., 2010; Williams et al., 2010; Maxwell et al., 2022). These altered disturbance regimes are reshaping forest ecosystems across diverse landscapes, disrupting ecosystem resilience. Impacts to biodiversity, forest structure, hydrological processes, ecosystem processes (e.g., fire), and ecosystem services are often poorly predicted or understood (Anderegg et al., 2013). Biotic forest disturbance agents, primarily insects, affect a larger land area than wildfire in the northern hemisphere (Kautz et al., 2017) and may be the primary source of forest mortality in temperate and boreal forests over the coming century (Safford and Vallejo, 2019; Maxwell et al., 2022). Yet, less attention in the literature is devoted to understanding the ecological effects of drought- and insect-induced tree mortality and how forest management may mediate or exacerbate those effects.
Increasing drought severity is exacerbating tree susceptibility to bark beetle outbreaks both through elevated moisture stress that reduces tree defenses to bark beetles, and to enhanced favorable conditions for bark beetle population growth (Robbins et al., 2022). Additionally, historical and current fire suppression practices in the western United States have led to increased forest densification, heightening competition for limited water resources and further stressing trees (Sohn et al., 2016; Bottero et al., 2017; Young et al., 2017; Vernon et al., 2018; Voelker et al., 2019; Keen et al., 2022). This densification may also amplify the susceptibility of forests to bark beetle infestations (Raffa et al., 2008; Hood et al., 2016; Jaime et al., 2022), as dense stands provide more hosts for beetles, potentially accelerating the spread and impact of these outbreaks. Such interactions between climate change, fire suppression, and drought underscore the urgent need for adaptive forest management strategies that can mitigate these intertwined threats (Bradford and Bell, 2017; Safford and Vallejo, 2019; Senf et al., 2020; Knapp et al., 2021; North et al., 2022).
The consequences of mass tree mortality extend beyond the immediate loss of tree life, posing significant challenges for fire risk management, forest health, and tree regeneration (Mueller et al., 2005; Wang et al., 2012; Wayman and Safford, 2021; Reed et al., 2023). The accumulation of dead trees increases the flammability of forest fuels and changes their spatial arrangements, likely elevating the risk of more severe and rapidly spreading wildfires at certain post-mortality time periods (Sieg et al., 2017; Wayman and Safford, 2021; Reed et al., 2023). Furthermore, the slow rate of decomposition of dry forest fuels (Harmon et al., 1987; Laiho and Prescott, 2004) coupled with the historically high tree density, is likely to result in fuel loads greater than the historical levels persisting until wildfire consumes them, potentially affecting fire behavior several decades into the future. Mass tree mortality changes forest understory conditions, with implications for tree regeneration and forest composition in future decades (Leverkus et al., 2021). Long-term carbon dynamics are affected by mass tree mortality as differing decomposition rates and changes to vegetation growth are induced (Kurz et al., 2008; Earles et al., 2014).
Dead-tree removal, often aimed at salvaging merchantable timber and reducing fuel loads, represents a potential management response to this complex suite of challenges posed by mass tree mortality. The ecological impacts of this strategy, however, extend beyond fuel reduction. The process involves disturbances to soil, creation of slash, incidental removal of live trees, and potential damage to surviving small trees (Leverkus et al., 2018), altering conditions influential to tree regeneration and future species composition. These changes can also alter immediate fire behavior and fire risk, while potentially affecting fire behavior over the course of a century through changes in fuel composition and structure (Fraver et al., 2011; Leverkus et al., 2020). Dead-tree removal likely reduces aboveground carbon stocks in the short term but may also reduce the long-term carbon efflux to the atmosphere by decreasing the size and severity of subsequent wildfire. This could change patterns of pyrogenic carbon production (Maestrini et al., 2017) while potentially increasing overall carbon stocks over time and across large areas of forest (Liang et al., 2017; Loudermilk et al., 2017). These combined changes can have profound effects on forest structure, regeneration, and future forest dynamics (Keyser et al., 2009; Seidl and Turner, 2022), necessitating a broad evaluation of dead-tree removal’s ecological outcomes.
A tree mortality event of unprecedented extent and severity that occurred in California, USA, during and after the exceptional 2012–2016 hot drought (National Drought Mitigation Center, 2016) provided an opportunity to evaluate the ecological effects of dead-tree removal. A historically high number of trees died due to drought-related causes statewide, with the largest concentration of mortality at middle elevations of the central and southern Sierra Nevada (Moore et al., 2018). We compared adjacent forest areas with and without dead-tree removal, with the aim of addressing critical questions regarding changes in forest regeneration, fuel dynamics, fire behavior, and carbon storage. Specifically, we ask how dead-tree removal affects: (1) the amount and composition of forest fuels; (2) tree regeneration in the form of seedling and sapling density and species composition; (3) carbon pools; and (4) fire behavior and severity and fuel loads over the following century. Answers to these questions will elucidate forest responses to the interacting disturbances of tree mortality and dead-tree removal, and will aid resource managers in making complex decisions affecting future forest composition, carbon stores, and wildfire.
2 Materials and methods
2.1 Study site
The study was conducted in the central and southern Sierra Nevada of California, USA. The forest type is mixed conifer (North et al., 2016; Safford and Stevens, 2017) and is dominated by white fir (Abies concolor), incense cedar (Calocedrus decurrens), ponderosa pine (Pinus ponderosa), Jeffrey pine (P. jeffreyi), sugar pine (P. lambertiana), and California black oak (Quercus kelloggii). Regional climate is Mediterranean-type, consisting of cold, wet winters and warm, dry summers. The mean annual precipitation of ~1,080 mm falls mostly between November and May, with approximately equal amounts of rain and snow (Minnich, 2007). Plot elevations ranged from 1,190 to 2,295 m. Soils are formed largely from granitic substrates except for approximately half the plots at Longbarn, where soils are derived from weathered glacial deposits. Soils are well-drained to somewhat excessively well-drained and deep to moderately deep (Giger and Schmitt, 1983; Hanes et al., 1996; UC Davis, 2022).
Where drought-related mortality occurred in the central and southern Sierra Nevada, an estimated 49% of trees died, with the highest concentrations of mortality in large-diameter ponderosa pine, mid- and large-diameter sugar pine, mid-diameter white fir, and small- and mid-diameter incense cedar (Fettig et al., 2019). The most common mortality agents were Western pine beetle (Dendroctonus brevicomis), Jeffrey pine beetle (D. jeffreyi), mountain pine beetle (D. ponderosae), and fir engraver beetle (Scolytus ventralis) (Moore et al., 2018; Fettig et al., 2019). We installed study plots at six sites where dead-tree removal was previously conducted to remove dead fuels, salvage merchantable timber, and/or remove hazard trees along roadsides. Sites spanned the Stanislaus, Sierra, and Sequoia National Forests, private lands owned and managed by Southern California Edison (SCE), and at Mountain Home State Demonstration Forest (Figure 1, Table 1). Study sites represent typical dry mixed-conifer forest conditions of the Sierra Nevada (Safford and Stevens, 2017).
Figure 1
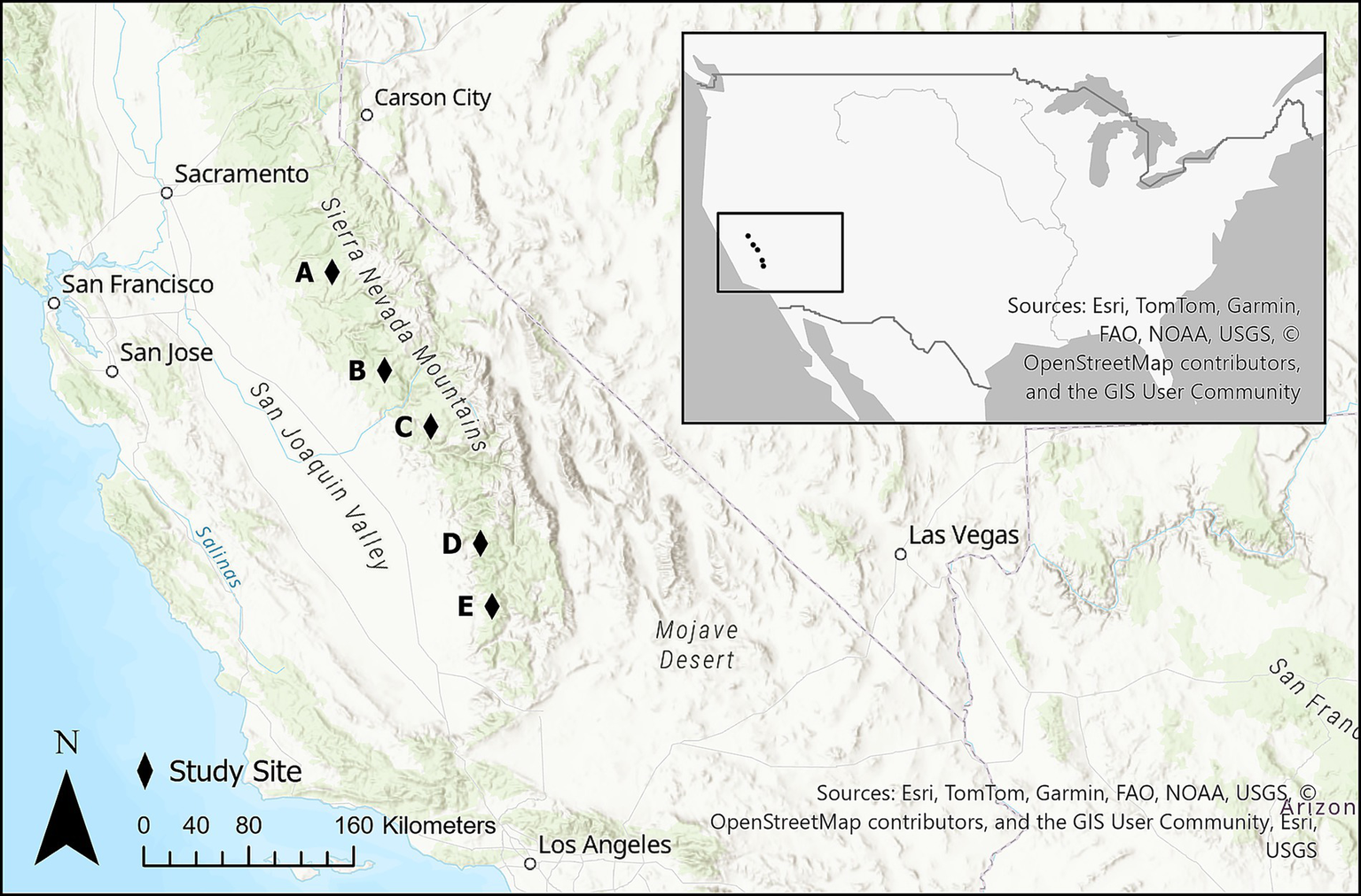
Distribution of field sites across the central and southern Sierra Nevada, California, USA: (A) Longbarn, (B) Bass Lake, (C) Dinkey Creek and Southern California Edison, (D) Mountain Home Demonstration State Forest, (E) Spear.
Table 1
| Site name | Ownership | Number of plots | Sampling year |
|---|---|---|---|
| Longbarn | Stanislaus National Forest | 30 | 2021 |
| Bass Lake | Sierra National Forest | 28 | 2020, 2021 |
| Dinkey Creek | Sierra National Forest | 24 | 2021 |
| SCE | Southern California Edison | 8 | 2021 |
| Spear | Sequoia National Forest | 20 | 2020 |
| Mountain home demonstration state forest | State of California | 12 | 2020 |
Characteristics of sampling sites, all of which match those of Wayman and Sorenson (2025).
The number of plots in each site was limited by the sampling area. All sites are located within the Sierra Nevada range of California, USA. See Figure 1 for site locations.
We established plots at sites where targeted mechanical removal of dead trees was conducted between 2016 and 2020. Mountain Home and SCE lands have been actively managed for timber harvest, fuels reduction, and resistance to severe wildfire for many decades, including with prescribed fire. National Forest lands where our plots were located have a management history consisting of fire suppression and limited small-tree thinning and hazard-tree removal. We only sampled sites where contractors were required to remove felled trees as opposed to leaving them in place. Six plot pairs were within the 2016 Cedar Fire footprint in areas of low fire severity (as determined by field observations). Dead-tree removal at Mountain Home and SCE can be considered salvage logging, where trees were removed both for ecological and timber income purposes. Dead-tree removal on National Forest lands was limited to roadside hazard tree removal projects where removal bands were approved for ~185 m widths. Roadside dead-tree removal extended a minimum of 70 m from the road. Unlike many fuel reduction and timber harvest treatments, roadside hazard tree removal boundaries are not driven by ecologically relevant site conditions, which is an advantage for locating suitable paired control plots near the treatment. Because we sampled sites with a variety of salvage and hazard-tree prescriptions, we use the term “dead-tree removal” to describe both management actions.
2.2 Plot design and data collection
We established 122 paired plots with one plot per pair within an area of post-mortality dead-tree removal and one plot in an area with no dead-tree removal (“removal” or “no removal”; Figure 2). Plot pairs were established where removal treatment boundaries did not coincide with detectable physical or ecological differences that could influence our results. Plots within a pair were qualitatively similar in terms of rock cover and pre-drought overstory vegetation; they did not differ in slope gradient by more than 20% or aspect by more than 45°. Plot centers were a minimum of 35 m from roads and removal treatment boundaries for removal plots, and 60 m from removal treatment boundaries for no-removal plots. Within locations field-identified as meeting the above criteria, we used a random-number generator to establish plot pair coordinates, but restricted coordinates to at least 400 m from other pairs. Except that four plot pairs were > 330 m from other pairs due to the constrained sampling area at the site. Plots within a pair did not differ in pre-removal tree mortality intensity (Wayman and Sorenson, 2025). Plots were typically established 80–120 m and no more than 180 m from their pair. Some within-pair aspect differences exceeded 45° as measured by crews after plot establishment, but these were likely due to flat or undulating terrain.
Figure 2

Example photographs of sampling plots where no dead-tree removal occurred (a,c) and where dead-tree removal was performed (b,d) following severe drought- and bark beetle-induced tree mortality (Wayman and Sorenson, 2025). Photo Credits (R. B. Wayman, 2021).
In hazard tree removal areas, plots were typically located above roads to minimize downslope effects from the road. To examine possible effects of placing plots above roads, we surveyed five plot pairs below a road located 100–200 m from a matched pair above the road. For initial model runs, we included a fixed effect explaining position relative to the road (above, below, or away); it did not affect any response variable and was excluded for all analyses. Plots were sampled one to 5 years after the removal treatment was conducted. Time since removal was not considered because the precise year of removal treatment for each plot location was not available. The range of removal treatment ages included in our sample is suitable for our purposes because it captures the initial post-removal environment, which is the target timeframe of our study.
At each 11.35 m radius (~0.04 ha) circular plot, surveyors recorded a suite of topographic and ground cover metrics. Following methods in Brown (1974), coarse woody debris (1000-h fuels; sound = decay classes 1–3, rotten = decay classes 4–5; Thomas, 1979), fine fuels (1-, 10-, and 100-h fuels; diameter classes are: 1-h = 0cm to 0.6 cm; 10-h = 0.6cm to 2.5 cm; 100-h = 2.5cm to 7.6 cm), and litter depth were measured along four transects, starting at plot edge in each of the four cardinal directions and working inward. Fuel mass (Mg ha−1) was estimated using the equations in Brown (1974), using the mean tree species coefficients for ponderosa pine, white fir, and incense cedar from Van Wagtendonk et al. (1998). Five soil samples were collected at fixed locations within each plot using a 2.2 cm diameter soil core probe, with samples divided into 0cm–10 cm and 10cm–30 cm depths. Soils were combined into one sample per plot for each depth, then air-dried in paper bags. In the lab, roots were manually separated, oven-dried at 50 °C, and weighed. Soils were oven dried at 50 °C and weighed, then sieved through a 2-mm sieve. They were then analyzed for percent organic carbon and percent organic matter using the loss on ignition method (Miller et al., 2013). Litter and duff were collected within 10 cm× 10 cm quadrats by snipping the material along quadrat boundaries and depositing it into separate paper bags. Samples were air dried in the field and oven dried for 24 h at 50 °C in the lab, then weighed.
We recorded the species and diameter at breast height (diameter of tree bole 1.37 m from ground on uphill side; DBH) of each live and dead tree > 7.6 cm DBH. For each stump (defined as dead trees < 1.37 m tall), we recorded the species, diameter at 10 cm above the base of the litter layer, whether it was cut or broken, and decay class of one through five, from most sound to most rotten (Ganey and Vojta, 2010). The distance from the plot center to the nearest potential seed source for each tree species visible from the plot was recorded. Ponderosa and Jeffrey pines were combined into “yellow pines” for analysis due to low sample sizes, potential hybridization, and overlapping niches.
At the center of the ~0.04 ha main plot, a 60 m2 circular subplot was established for recording tree regeneration (which, within our study sites, was natural and not planted). Live seedlings (trees < 1.37 m or 4.5 ft. tall) and saplings (taller than seedlings but < 7.6 cm or 3 in DBH) were tallied by species and age (estimated by counting the bud scars and subtracting the current year; trees > 10 yr. recorded as > 10). Incense cedar age cannot be estimated by counting bud scars, so individuals were recorded as either age 0 (current-year seedlings), or > 0 with height recorded. Resprouts from hardwood stumps were tallied by species, and DBH was measured if > 1.37 m; resprouts > 1 m apart were considered separate individuals if not obviously belonging to the same stump.
Topographic Relative Moisture Index (an index ranging from 0 = xeric to 60 = mesic), which estimates relative soil moisture availability among sites in mountainous terrain, was calculated from four field metrics (Parker, 1982): transformed aspect, slope gradient, topographic position, and slope curvature.
2.3 Fire behavior and effects modeling and carbon estimation
To better understand the potential effects of dead-tree removal on future fuel loading, fire behavior and severity, and carbon stocks, we simulated snag and fuel dynamics using the Western Sierra variant of the Forest Vegetation Simulator (FVS) with Fire and Fuels Extension (FFE). FVS is a distance-independent, individual-tree growth and yield model used to simulate changes in forest structure, and FFE uses simulated forest structure to simulate fire behavior using fire-spread models (Rebain, 2015; Dixon, 2018; USDA Forest Service, 2023). For our paired plot design, simulations provide a valid comparison of the magnitude of difference between removal and no removal, even if exact values cannot be verified. Our simulations included plot coordinates and field-measured aspect and slope, all live and standing dead trees found in the study plots, harvested trees measured at cut height, and field-measured surface fuels. FVS-FFE models fuel effects using fire behavior predicted by a standardized set of fuel models, but the model – or combination of models—can be automatically determined based on quantitative data. We prepared our data by binning field-measured fuels into the FVS-ready categories of 3–6, 6–12, 12–20, 20–35, 35–50, and greater than 50- inch diameter fuels, with separate pools for “sound” versus “rotten” debris, and then converted these data into estimates of biomass (tons) per acre using the fuel mass methods described in Section 2.2.
Given that we were interested in the relative effects of dead-tree removal across a long time-horizon, we simulated growth with a common starting year 2020 and ending in 2110. Forest structure and fire effects are reported at 10-yr intervals, plus a 5-yr interval at year 2025. We focused our analysis on a “potential” wildfire burning at each of these intervals—a metric that captures the general flammability of the plot over time but does not have repercussions for subsequent time steps. The potential wildfire was simulated under “severe” conditions designed to mimic wildfire; we used default wind, temperature, and fuel moisture parameters to capture this condition. We chose percent basal area mortality as our metric of fire severity, and torch potential (p-torch) as our metric of fire behavior (Rebain, 2015).
We also used FVS-FFE to estimate total stand carbon, including aboveground and belowground sources that include live and dead trees as well as other vegetation and fuel sources (e.g., shrubs, herbs, surface fuels, litter, duff). FVS-FFE estimates above and belowground biomass from the stand structure in a given timestep using allometric equations that are specific to the regional variant and therefore the local species. It then uses a conversion factor to estimate the amount of carbon in this biomass (0.5 for most pools). The specific pools, equations, and multipliers used in the simulator are described in Rebain (2015).
2.4 Statistical analyses
2.4.1 Forest structure and forest floor
We evaluated differences between dead-tree removal and no removal by comparing: basal area and density of live and dead trees (> 7.6 cm DBH) and, more granularly, of live trees by size class: small (7.6–30 cm DBH), medium (30–60 cm DBH), and large (≥ 60 cm DBH), live tree cover, mean litter depth (i.e., O horizon and above), litter mass, soil organic carbon, coarse and fine fuel covers, and bare ground cover. We excluded one plot with a single live giant sequoia (Sequoia gigantea) with a DBH of 378 cm to reduce its statistical leverage and improve estimates. We analyzed tree densities using generalized linear mixed-effects models with Poisson distributions; all other variables were modeled using linear mixed-effects models. All models were structured with removal as a fixed effect and a site-level random effect with pair ID nested within site to account for the pair design and block structure of the data. When linear mixed-effects models produced non-normal, heteroscedastic residuals, the response variable was log-transformed after adding each variable’s minimum value to each measurement; model estimates and standard error were back-transformed when reported. This occurred for all response variables, except soil carbon. All models met assumptions of residual normalcy, homoscedasticity, and dispersion where appropriate. Analyses were conducted using R (R Core Team, 2023) with the package ‘lme4’ for mixed-effects models (Bates et al., 2023) using ‘lmerTest’ and ‘car’ to calculate p-values (Fox et al., 2019; Kuznetsova et al., 2020) and ‘performance’ to assess overdispersion (Lüdecke et al., 2021).
We also modeled the effect of dead-tree removal on live and dead tree density and basal area by each major species: white fir, incense cedar, ponderosa pine, Jeffrey pine, sugar pine, and California black oak. We modeled live and dead tree density using generalized linear mixed-effects models with Poisson distributions and live and dead tree total basal area using linear mixed-effects models. We structured each model to test for an interaction between removal and species identity. Again, to account for the paired design and spatial non-independence, we included a unique pair identifier nested within the site. Where density was a response variable, we included an observation-level random effect to account for overdispersion in the response variables. Tukey-adjusted post-hoc tests were performed for the effect of removal for each species to assess whether responses differed by species.
2.4.2 Tree regeneration
To examine the influence of dead-tree removal on seedling and sapling regeneration, we modeled seedling and sapling density in response to removal. To test for species-specific responses, we included an interaction between removal and species identity. For seedlings and saplings separately, we used generalized linear mixed-effects models with Poisson distributions where the response variable was the count per species per plot. To reduce unexplained variance in the model, we also included covariate environmental variables known to affect seedling and sapling density (Table 2). These included log-transformed distance to seed source (interacting with species to account for differing dispersal rates), elevation, topographic moisture index, percent bare ground, mean litter depth, mass of fine and coarse woody debris, percent live tree cover, and percent shrub cover. Random effects were included for each plot nested within a pair nested in a site to account for non-independence of each species within plot, the paired design, and site-level effects. To compensate for overdispersion, we also included an observation-level random effect. To improve model estimates, we removed all environmental variables that did not improve model fit by −2 AIC (Akaike Information Criterion). To determine differences in species responses, Tukey-adjusted post-hoc comparisons were conducted using estimated marginal means in the ‘emmeans’ package (Lenth et al., 2024) to test the effect of removal by species.
Table 2
| Variable | Source |
|---|---|
| Seed source | Distance (m) to nearest adult tree for each species |
| Elevation | Extracted from elevation raster in ArcGIS using bilinear interpolation (m) |
| Topographic relative moisture index | Index of relative soil moisture availability calculated from field measures of transformed aspect, slope, topographic position, and slope curvature according to Parker (1982). |
| Bare ground cover | Field estimate of exposed soil cover (%) |
| Litter depth | Mean litter depth of 8 measurements in plot (cm) |
| Fine woody debris mass | Estimated from 4 Brown’s fuel transects (Brown, 1974) per plot (Mg ha−1) |
| Coarse woody debris mass | Estimated from 4 Brown’s fuel transects (Brown, 1974) per plot (Mg ha−1) |
| Live tree cover | Field estimate of plot level cover of live trees >1.8 m tall (%) |
| Shrub cover | Field estimate of shrub cover (%) |
Environmental variables tested for associations with changes in conifer regeneration, followed by their definitions and units.
Most variables match those shown in Table 2 of Wayman and Sorenson (2025).
2.4.3 Carbon
To determine the effect of dead-tree removal on carbon stocks, we modeled carbon mass per ha as a response to removal. We used linear mixed-effects models to test for differences in total organic carbon across major pools (aboveground and belowground live and dead trees, downed dead wood, shrub and herb, and forest floor). Carbon masses were log-transformed to meet model assumptions of homoscedasticity and residual normality; model estimates and standard error were back-transformed when reported. To account for the paired-plot design and spatial non-independence of site, we included a random effect of pair nested in site.
2.4.4 Simulated future fire behavior, fire severity, and fuel loads
To evaluate the effect of dead-tree removal on potential future wildfire, we tested for within-year differences for five variables each year in the FVS-FFE simulation: percent basal area mortality, torch probability, surface fuel mass, and standing live and dead tree mass. For percent basal area mortality and torch probability, we employed a generalized linear mixed-effects model with a binomial distribution. Surface-fuel mass, standing live, and standing dead fuels were analyzed using a linear mixed-effects model with a log-transformed response variable to meet model assumptions of residual normalcy and homoscedasticity; model estimates and standard errors were back-transformed when reported. To test within-year differences, each model was structured with year as a categorical variable interacting with removal. Tukey-adjusted post-hoc tests were conducted for the effect of removal within the year to determine whether removal influenced fire behavior and fuel mass. Models also included a random effect for plot nested in pair nested in site.
Analyses were conducted in R version 4.3.1 (R Core Team, 2023) and RStudio version 2023.9.0.463 (RStudio Team, 2023).
3 Results
3.1 Forest structure and forest floor
Dead-tree removal, as expected, resulted in a substantial reduction in dead-tree density and dead-tree basal area (Table 3). For live trees, density was modestly lower with removal, while basal area was not affected. Of the live trees, only small trees (7.6–30.0 cm DBH) showed reduced density (Table 3).
Table 3
| Metric | Tree status | No removal | Dead-tree removal | p-value | ||
|---|---|---|---|---|---|---|
| Mean | SE | Mean | SE | |||
| Density (trees ha−1) | Live | 213 | +27/−24 | 164 | +21/−19 | 0.014* |
| Dead | 122 | +17/−15 | 45 | +7/−6 | 3.8E-09*** | |
| Small live | 111 | +15–13 | 75 | +11/−9 | 0.001** | |
| Medium live | 58 | +8/−7 | 52 | +8/−7 | 0.458 | |
| Large live | 43 | +7/−6 | 37 | +6/−5 | 0.308 | |
| Basal area (m2 ha−1) | Live | 37.4 | +3.5/−3.3 | 31.2 | +3.2/−3.0 | 0.081 |
| Dead | 24.5 | +2.4/−2.3 | 5.8 | +1.5/−1.4 | 2.2E-10*** | |
| Small live | 2.9 | +1.1/−1.1 | 2.0 | +1.1/−1.2 | 0.564 | |
| Medium live | 9.7 | +1.1/−1.0 | 8.5 | +1.3/−1.3 | 0.502 | |
| Large live | 23.7 | +2.0/−1.9 | 19.3 | +1.8/−1.7 | 0.072 | |
| Pretreatment dead | 25.3 | +3.0/−2.8 | 25.7 | +3.0/−2.8 | 0.912 | |
Model estimated density and basal area of live and dead trees subset by size: small [7.6–30 cm diameter at breast height (DBH; 1.37 m)], medium (30–60 cm DBH), and large (≥60 cm DBH), with an estimate for pre-removal dead tree basal area (Wayman and Sorenson, 2025).
Back-transformed means with upper and lower standard errors (SE) are shown. p values below 0.05 are shown in bold (p < 0.05*, < 0.01**, < 0.001***).
Within our study areas, white fir was dominant, followed by incense cedar, with sugar pine, yellow pines (ponderosa and Jeffrey pines), and black oak subdominant (Figure 3). For individual species, live tree density (trees > 7.6 cm DBH) did not differ with removal except for California black oak, which was 70% lower with removal (p = 0.036), while live tree basal area did not differ for any species (Figures 3a,c). Dead tree density was lower with removal for white fir (p = 0.007), sugar pine (p = 0.012), and yellow pines (p = 0.026), and dead basal area was lower with removal for white fir (p < 0.001), incense cedar (p = 0.055), and sugar pine (p = 0.011; Figures 3b,d).
Figure 3
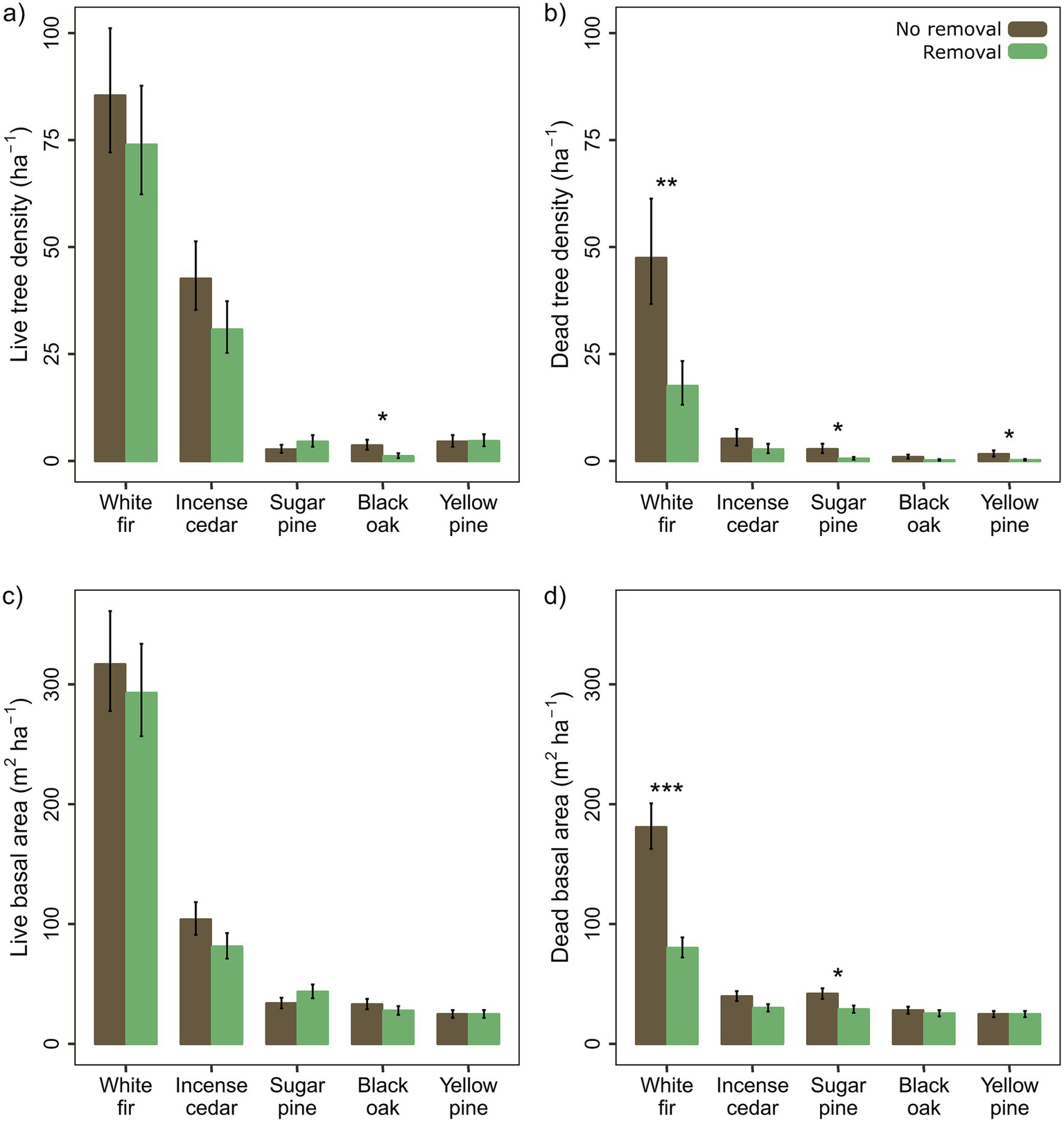
Treatment differences for overstory (>7.6 cm DBH) (a,b) tree density and (c,d) basal area for (a,c) live and (b,d) dead trees of the five most common tree species: white fir (Abies concolor), incense cedar (Calocedrus decurrens), sugar pine (Pinus lambertiana), California black oak (Quercus kelloggii), and yellow pines (P. jeffreyi and P. ponderosa). Means and standard errors are calculated from back-transformed model estimates and standard errors and were scaled to be per hectare based on the 0.0405 ha plot area (p < 0.05*, < 0.01**, < 0.001***).
Total mass of coarse woody debris (CWD) did not differ with removal, but sound CWD (decay classes 1–3) mass was 51% higher (p = 0.046) with removal (Figure 4). Total mass of fine fuels (1-, 10-, and 100-h combined) did not differ with removal, but 1-h and 10-h fuels were 21 and 20% lower, respectively, with removal (p = 0.006 and p = 0.004). Litter and duff depth decreased 38 and 51%, respectively, with removal (p > 0.001, p > 0.001), while cover of bare ground increased 15-fold (p > 0.001). Fuel depth also decreased 18% with removal (p = 0.035).
Figure 4
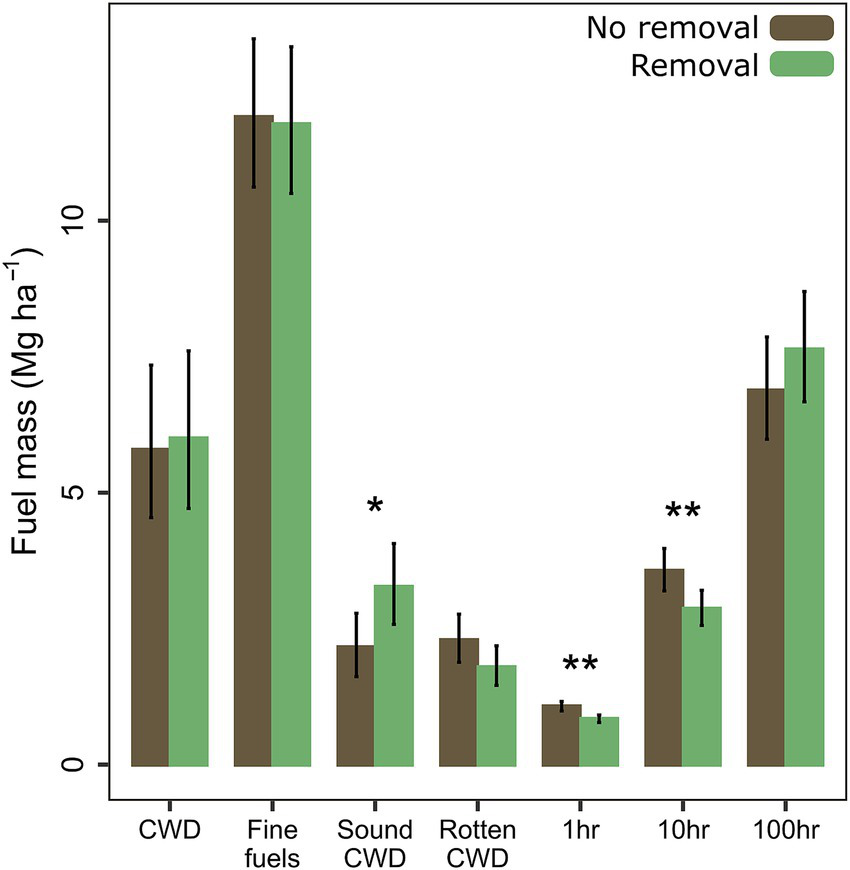
Fuel mass by fuel type. Total coarse woody debris (CWD) and fine fuels are compared first, followed by subgroups of each. Sound CWD = decay classes 1–3; rotten CWD = decay classes 4–5. Fine fuel diameter classes are: 1-h = 0 to 0.6 cm; 10-h = 0.6 to 2.5 cm; 100-h = 2.5 to 7.6 cm. Means and standard errors are calculated from back-transformed model estimates and standard errors (p < 0.05*, < 0.01**, < 0.001***).
Our estimate of pre-removal basal area mortality did not differ between plots with and without removal (p = 0.912; Table 3). Density of stumps in decay classes 1–2 was 516% higher with removal (p > 0.001), but density of stumps in decay classes 3–5 did not differ with removal (p = 0.61). Median tree height and height to live crown did not differ with removal (p = 0.43, p = 0.85).
3.2 Tree regeneration
The response of tree seedling density to removal differed by species: white fir and sugar pine seedling density were 55 and 64% lower, respectively, with removal (p = 0.013; p = 0.038), and yellow pine seedling density was 349% higher (p = 0.017) (Figure 5a). Incense cedar and black oak seedling density did not differ with removal. Total tree seedling density did not differ with removal. Sapling density was 56% lower with removal plots (p = 0.025), and the effect of removal on density did not differ by species (Figure 5b). Total seedling density was associated with higher elevations and higher topographic relative moisture index (p > 0.001), our proxy for soil moisture (p = 0.002), while sapling density was associated with higher elevations (p = 0.003).
Figure 5
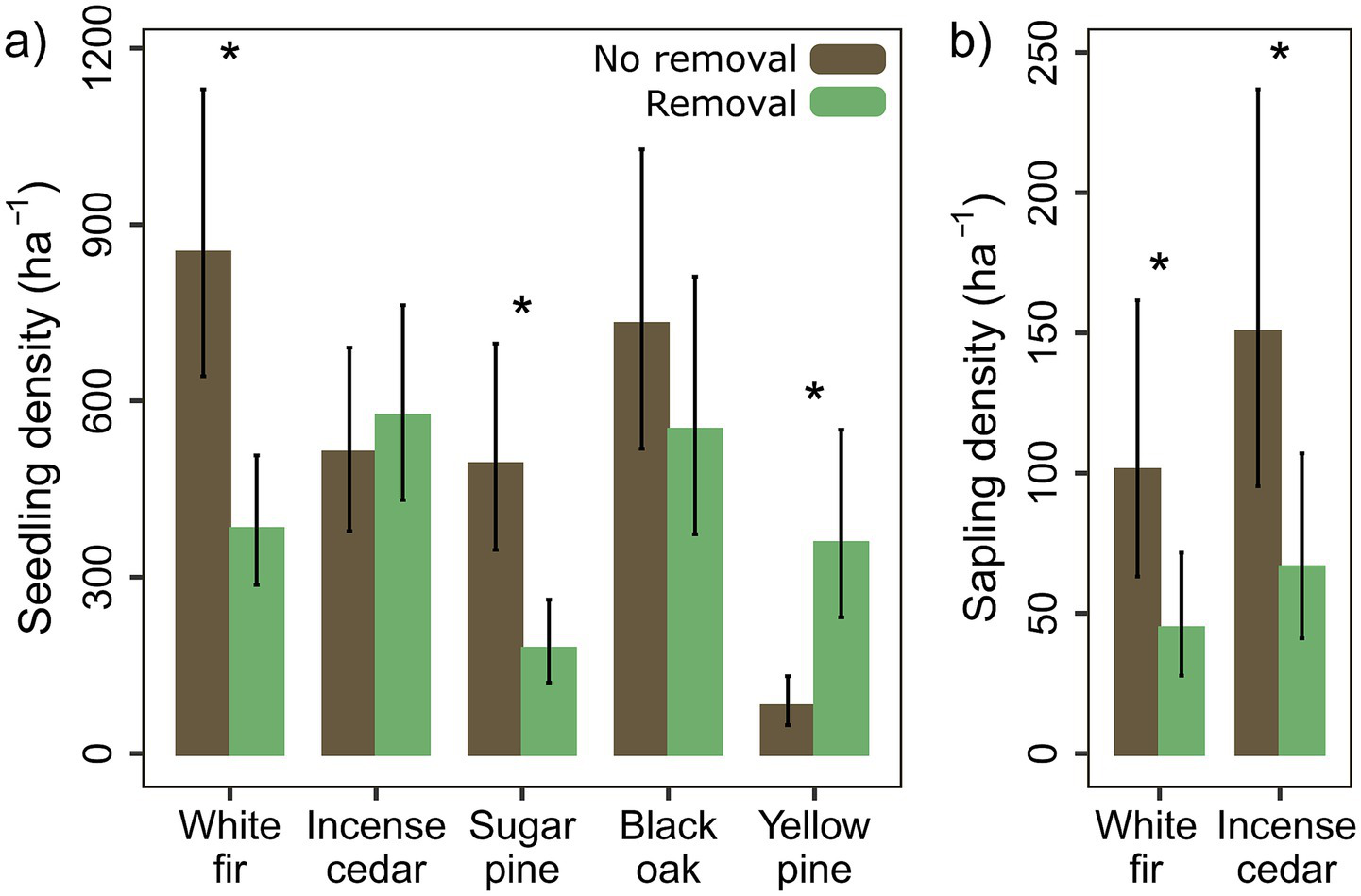
Mean (a) seedling (<1.34 m tall) and (b) sapling (>1.34 m tall, <7.6 cm DBH) density for the five most common tree species. Only white fir and incense cedar saplings were observed frequently enough to allow means testing. Means and standard errors are calculated from back-transformed model estimates and standard errors and were scaled to be per hectare from the 0.006 ha search area (p < 0.05*, < 0.01**, < 0.001***).
3.3 Carbon
Differences in carbon pools tracked with differences in surface fuels and standing live and dead trees (Figure 6). The total measured carbon mass was 32% lower with removal (p < 0.001). Aboveground standing live carbon was 25% (p = 0.028) lower with removal, while standing dead carbon was 83% (p < 0.001) lower. Belowground live carbon was 27% lower with removal (p = 0.042), but belowground dead carbon did not differ with removal (Figure 6). Carbon held in the surface layer of shrubs and herbs tended to be greater with removal, but the evidence was weak (p = 0.059). Carbon from downed wood and the forest floor did not differ between removal and no removal, and soil organic carbon was unaffected by removal at both depths.
Figure 6
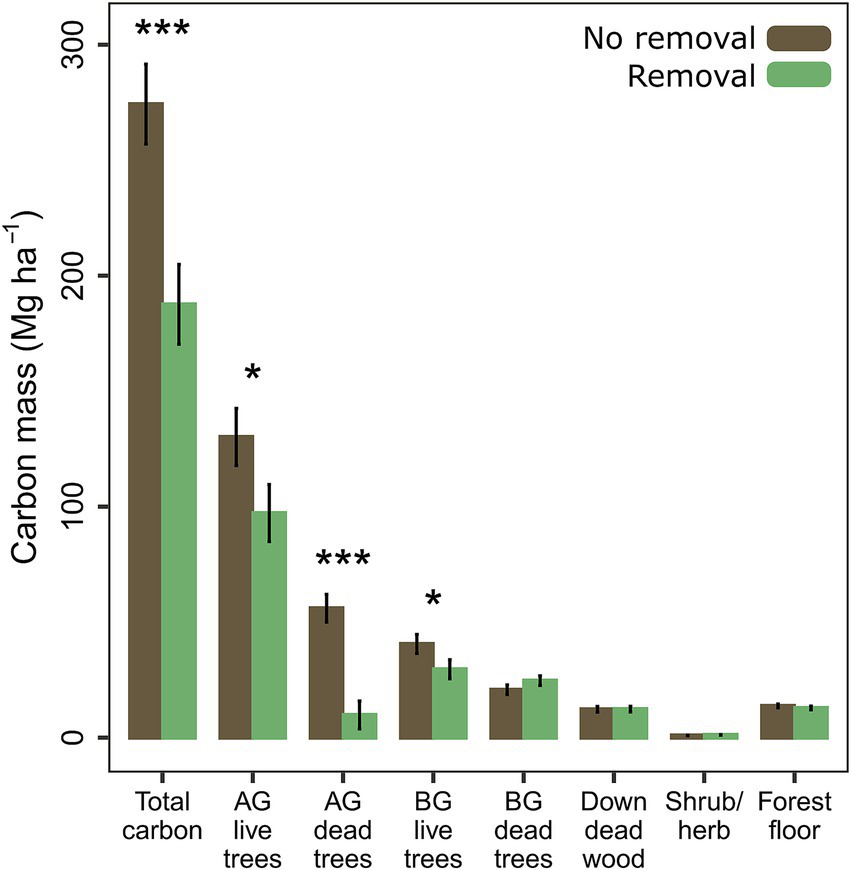
Mean mass of total carbon and carbon within major pools estimated by FVS-FFE. Means and standard errors are calculated from model estimates and standard errors (p < 0.05*, < 0.01**, < 0.001***). AG, aboveground; BG, belowground.
3.4 Simulated future fire behavior, fire severity, and fuel loads
Simulations of burn severity (% basal area mortality) and fire behavior (torch potential) over a 90-year period indicate a lag in removal effects: only after 20-year post-removal are burn severity and torching potential significantly reduced (p = 0.014 and p < 0.001, respectively). The 30 years following are marked by a widening gap in the effect of removal on burn severity and torch potential, ultimately requiring about 40 additional years for no removal to match the reductions in burn severity and torch potential seen after removal (Figures 7a,b). Modeled surface fuels were similar in 2020, but were projected to be lower with removal just 5 years after removal (p = 0.030). The simulation predicted the following 15 years to show a disproportionate increase in surface fuels in plots without removal, followed by an equilibrium state where surface fuels continue to increase overall but removal results in consistent ~35% lower surface fuel mass thereafter (Figure 7c). Simulated mass of standing dead fuels was lower with removal beginning at the time of sampling (p < 0.001) and continuing throughout the simulation period. Standing live fuel mass was marginally lower with removal (p = 0.091) at the time of sampling, but was not significantly different from plots with no removal for any future timestep (Figures 7d,e).
Figure 7
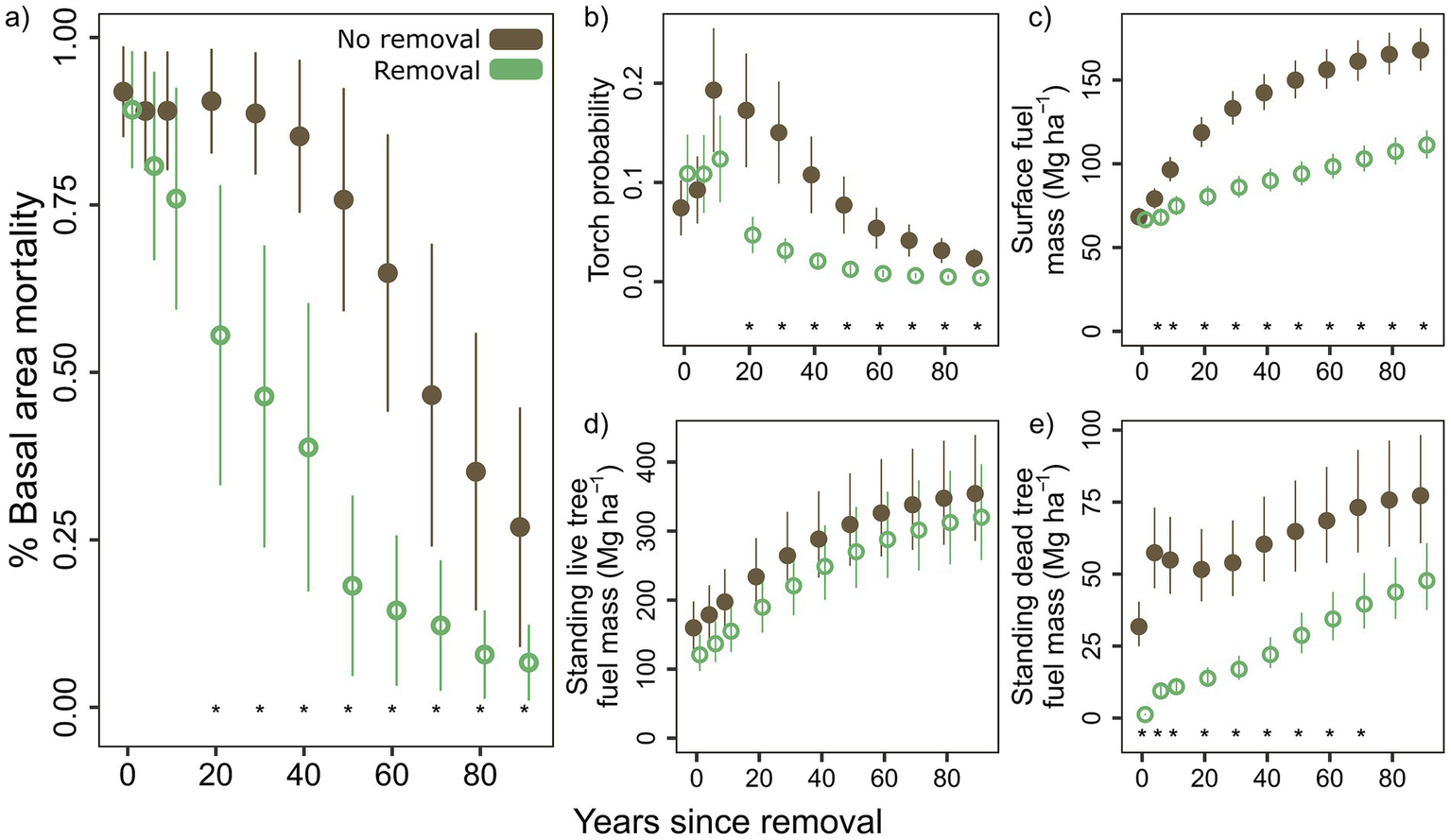
Changes in (a) post-fire percent basal area mortality, (b) torch probability, (c) surface fuel mass, (d) standing live tree fuel mass, and (e) standing dead tree fuel mass simulated by the Forest Vegetation Simulator with the Fire and Fuels Extension in decadal increments over 90 years. Percent basal area mortality (a) and torch probability (b) assume an average wildfire occurring in that year. Means and standard errors are calculated from back-transformed model estimates and standard errors (p < 0.05*).
4 Discussion
4.1 Summary of key findings
Dead-tree removal after severe drought- and bark beetle-induced mortality enhanced regeneration of the historically dominant shade-intolerant yellow pines (ponderosa and Jeffrey pine), while decreasing seedling regeneration of shade-tolerant white fir. However, removal also decreased regeneration of the moderately shade-tolerant sugar pine, and mature black oak density declined with removal. Simulations of future conditions after removal foretold meaningfully milder burn severity and fire behavior, likely as a result of massively reduced dead fuel loads persisting over a century and decreased live small tree density. Total on-site carbon at time of sampling was lower with removal, driven predominantly by the removal of aboveground dead-tree biomass, while live tree biomass was also reduced but to a smaller degree.
4.2 Regeneration
The effect of dead-tree removal on tree regeneration in the Rocky Mountains within lodgepole-pine forests aligns with our results: shade-intolerant pine regeneration was enhanced with removal, while fir regeneration was diminished (Collins et al., 2011; Rhoades et al., 2020). This outcome likely results from the effect of dead-tree removal on microsite conditions. The conditions created by over a century of fire exclusion have favored the more shade-tolerant white fir and incense cedar, while fire-adapted, shade-intolerant pine species declined (Stephens et al., 2015). The process of removing dead trees disturbs litter and duff, thereby increasing the prevalence of bare ground for seedling establishment. Furthermore, removal of snags and incidental live trees increases understory light availability, favoring establishment of fire-adapted, shade-intolerant species (e.g., ponderosa and Jeffrey pine; Agee, 1998; Filip et al., 2007; Safford and Stevens, 2017). Sugar pine seedlings are able to germinate in both bare mineral soil and litter but have higher soil moisture requirements than ponderosa and Jeffrey pines (Keeley and Zedler, 1998; Legras et al., 2010). It is possible that removal decreased soil moisture via increased sunlight and decreased protective duff and litter (Hood et al., 2017), thus negatively affecting sugar pine regeneration.
The stark shift in the relative proportion of white fir to yellow pine seedlings with removal underscores the strong impact of microsite conditions on tree regeneration. Mean seedling density of white fir was 10.7 times that of yellow pines without removal, but densities did not differ with removal (Figure 5), even though live overstory white fir density was nearly 20 times greater than yellow pine density. In this forest type, this suggests that in the absence of severe wildfire and given the availability of a seed source, favorable microsite conditions are a more significant factor for successful yellow pine recruitment than the abundance of overstory yellow pine individuals. Pine regeneration was not enhanced by post-drought mortality removal in montane hardwood conifer and ponderosa pine forest types at a southern Sierra Nevada site near the lower elevational range of our plots (Bisbing et al., 2025). In addition to dead-tree removal, treatments included mastication that heavily impacted live small trees and increased fine fuel loading, so microsite conditions may have been less favorable to pines than at our sites.
How the removal’s effect on recruitment translates to mature overstory species composition will depend on future demographics. Young conifer seedlings commonly suffer high levels of mortality (Fowells and Stark, 1965), and seed production is highly variable from year to year (McDonald, 1992). Successful pine seedling establishment depends on the coincidence of infrequent masting events with favorable climatic conditions for germination and survival (Keyes and Manso González, 2015), thus adding further uncertainty to future species composition. Additionally, pine sapling abundance was very low across all surveyed plots (three Jeffrey pines and five sugar pines); therefore, any possible release of young, suppressed pines as a result of removal would be severely limited. In their re-sampling of Sierra Nevada mixed-conifer plots initially sampled before and after logging in 1928–9, Brodie et al. (2023) found that zero current overstory pines established after logging, even though skid trails, and bare ground were positively associated with initial post-logging regeneration of pines (and other species). They speculate the cause to be the absence of fire as a disturbance process. Future sampling at our sites will be necessary to evaluate whether the initial increase in pine seedling abundance translates into meaningful shifts in mature tree species composition.
Black oak seedling density did not change significantly with removal, but as with post-removal yellow pines, overall black oak seedling density was high relative to mature black oak density, which did decrease with removal (Figures 3, 5). Black oaks were resistant to mortality from this extreme drought and may increase in dominance in lower and middle elevation Sierra Nevada forests with a changing climate (Young et al., 2019; Bisbing et al., 2025). Again, future sampling will be needed to track any such shifts.
4.3 Removal impacts on forest carbon
Our analysis only looks at a current snapshot of forest carbon and does not attempt to quantify the off-site carbon storage or effects on future carbon storage within the forest. Removal impacts on on-site carbon pools were largely as expected, with the largest differences occurring in the standing dead carbon pool, which removal was designed to remove. The material was removed from the site as merchantable timber; therefore, a large portion of it remains stored as wood product and was not transformed to atmospheric carbon. Importantly, if reductions in current forest carbon on site translate to reduced fire severity in the future, total net stored carbon would actually be increased over the longer term with dead-tree removal (Liang et al., 2017; Loudermilk et al., 2017).
Recovery to pre-drought carbon storage levels may be inhibited by future bark beetle outbreaks (Robbins et al., 2023), but with removal, the decrease in both sapling and small tree density may reduce susceptibility to both wildfire and bark beetle outbreaks in the future (Vernon et al., 2018; Restaino et al., 2019; Knapp et al., 2021). Simulation modeling of density-reduction treatment effects on subsequent insect outbreaks under future climate scenarios has indicated that pre-outbreak treatments (as modeled) may not substantially reduce tree mortality in the Sierra Nevada as the climate warms (Maxwell et al., 2022). It is possible, however, that the combination of severe tree mortality plus post-mortality dead-tree removal that we observed would lower tree densities substantially enough to positively impact water availability, and thus forest resistance to insect-related mortality, under future drought.
4.4 Fuels and implications for future wildfire
The different responses by fuel type in forest floor fuel mass demonstrate that not all downed fuels respond similarly to removal. The process of tree felling left behind large pieces of sound material, thereby increasing sound coarse woody debris, but rotten coarse woody debris was unchanged. The 100-h surface fuels were unchanged with removal, but it is possible that both crushing by heavy machinery and the addition of new material took place. The two smallest fuel size classes were lower with removal, possibly due to being crushed or pulverized by heavy machinery. This is in contrast to results from Hood et al. (2017), who found that in beetle-affected lodgepole pine forests, salvage treatments substantially increased 1-h and 10-h fuels.
Sound coarse surface fuels increased with removal in the near-term, but in no-removal plots, those equivalent fuels remained tied up in standing dead trees that will eventually fall to the forest floor, if not consumed by fire first. Coarse fuel mass will substantially increase without removal as snags fall (Reed et al., 2023; Northrop et al., 2024). Large loads of coarse woody debris across extensive areas can foster mass fire behavior, driving faster fire spread and more intense burning (Stephens et al., 2018, 2022). Dead-tree removal may increase surface fuel ignition probabilities and fire spread rates due to decreased fuel moisture from increased solar radiation, surface temperatures, and herbaceous fuels (Hood et al., 2017). In contrast, the decrease in litter cover and depth and reduced mass of 1-h and 10-h fuels would likely reduce the speed of surface fire spread in areas with removal.
Updated fire behavior modeling, fully incorporating these complex fuel dynamics, is needed to appropriately assess net benefits and costs of dead-tree removal on future wildfire severity and behavior. Current operational models of fire behavior do not appropriately account for the substantial energy release and long burning times of large diameter standing dead and downed fuels found in areas of severe drought- and bark beetle-induced tree mortality (Stephens et al., 2022); only fine fuel classes influence fire behavior in the model (Rebain, 2015). Even so, our models predict higher fire severity and more extreme fire behavior without removal within 20 years and continuing into the future, possibly due to the gradual contribution of standing dead trees to coarse woody debris and eventually to smaller fuel classes that, in turn, contribute to fire behavior and spread patterns in the FVS-FFE models. The differences in fire behavior with removal would likely be even larger if the direct combustion of larger-diameter fuels (snags, coarse woody debris) could be incorporated in the fire model (see Stephens et al., 2022). Notwithstanding the limitations of the simulation models, it is reasonable to accept the general prediction of future decreased fire severity and behavior after the removal of large amounts of fuel.
The actual timeframe of removal impacts on fire behavior and severity could differ from FVS-FFE predictions depending on real-world snag-fall rates, but FVS-FFE estimates for our most abundant species do align well with snag-fall rates observed in our study system (Reed et al., 2023). FVS-FFE estimates that <10% of 50.8 cm DBH snags will have fallen within five years of mortality, and about half within 20 years of mortality, with only small divergences from this estimate based on species (Rebain, 2015). After the same mortality event that we studied, Reed et al. (2023) observed that 4% of white fir snags had fallen by five years since death, and 10% of incense cedar snags had fallen by four years since death (they did not report snag-fall rates beyond five years). White fir and incense cedar make up the vast majority of live and dead trees at our sites (Figure 3). Still, there are large discrepancies in the wider literature regarding snag-fall rates of these two species (Reed et al., 2023), and the fast rates they observed for ponderosa pine (40% of ponderosa pine snags fell by five years since death) and the fast snag-fall rates reported in our study system by Northrop et al. (2024) do not align with FVS-FFE assumptions. Monitoring for additional years post-mortality would be needed to evaluate alignment with modeled snag-fall rates.
4.5 Management implications
Even with the uncertainties surrounding future drought-related tree mortality in a changing climate, proactive forest restoration treatments are likely resource managers’ best hope for facilitating forest resistance to stressors such as severe wildfire and insect outbreaks (Bottero et al., 2017; Restaino et al., 2019). Where severe insect outbreaks and drought mortality do occur, our research demonstrates that frequent-fire forests long departed from historic disturbance conditions likely will receive benefits from dead-tree removal, although the decreases in mature black oak density and sugar pine seedling density with removal may be counter to management goals. Removal had positive initial effects on the species composition of regenerating trees by favoring some fire-resistant conifers, but if restoration goals include maintaining or increasing the abundance of sugar pine, replanting or maintaining microsites with higher soil moisture via more heterogeneous treatments may be necessary. Even the initial increase in yellow pine regeneration we observed may not translate into future overstory composition without low-severity fire (Brodie et al., 2023). To maintain or enhance the persistence of black oak, a species with high Indigenous cultural value and wildlife habitat value, restoration of frequent fire and targeted pruning of post-disturbance resprouts may be needed (Stephens et al., 2023; Ritchie et al., 2024), along with fuel reduction treatment prescriptions that specify maintenance of live mature individuals.
Dead-tree removal reduced modeled wildfire severity and behavior beginning two decades after disturbance, and this is likely a conservative estimate, since the models do not fully incorporate the large future increase in downed woody fuels in sites without removal. In terms of carbon storage, forest managers will need to weigh the short-term loss of standing dead carbon and small live trees with removal against possible protections from future carbon loss via severe wildfire and insect outbreaks.
The choice to remove dead trees should consider the ecological benefits of standing snags and downed coarse wood. Snags are used by wildlife for nesting, denning, and roosting, and provide important food sources for many species, while downed wood provides habitat and food and contributes to forest nutrient cycling and microbial activity (Knapp, 2015). The challenge is to find the appropriate balance between reducing the risk of catastrophic wildfire and drought mortality, promoting forest health and vigor, maintaining long-term carbon stocks, and enhancing biodiversity.
5 Conclusion
In Sierra Nevada mixed-conifer forests, removal of dead trees in areas of severe drought- and bark beetle-induced tree mortality initially enhanced regeneration of fire-adapted yellow pine species, but future sampling will be needed to confirm whether this cohort of young trees survives beyond the very vulnerable seedling stage. Future modeled wildfire severity was unaffected by removal initially but was reduced with removal beginning in the year 2040 (20 years post-removal), likely due to eventual inputs of fine fuels from initially standing dead trees in sites without removal. However, this modeling does not fully incorporate the massive amounts of standing and downed coarse fuels remaining on site in different spatial arrangements and quantities as a result of removal, and this question should be re-examined if more appropriate simulation models become available. Forest carbon pools followed predictable patterns, with removal having a large reduction in the standing dead carbon pool. Dead-tree removal’s effect on future forest carbon sequestration and storage will depend on the extent to which they facilitate more fire-adapted tree species composition and whether they increase resistance to future severe wildfire and insect outbreaks.
Statements
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://doi.org/10.5061/dryad.q573n5ttg Dryad accession number: dryad.q573n5ttg.
Author contributions
QS: Writing – original draft, Visualization, Writing – review & editing, Investigation, Methodology, Supervision, Formal analysis, Data curation. RW: Methodology, Writing – review & editing, Investigation, Data curation, Funding acquisition, Formal analysis, Conceptualization, Writing – original draft, Supervision, Project administration, Visualization. TU: Writing – review & editing, Writing – original draft, Formal analysis. HS: Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This project was funded by the California Department of Forestry and Fire Protection’s Forest Health Research Program as part of the California Climate Investments Program, grant # 8GG18801. Funding for open access publication fees was provided by University of California.
Acknowledgments
We are grateful to the outstanding field crew members who collected data for this project: Marta DeGuili, Greg Pelaez, Molly Radosevich, Cindy Rodriguez-Batres, Brittany Rossetti, Dylan Stephens, and George Tomal. Thanks to Amarina Wuenschel (USFS Region 5 Ecology Program), Ryan Stewart (Southern California Edison), Jim Kral (Mountain Home Demonstration State Forest), and staff of the Stanislaus, Sierra, and Sequoia National Forests and Sequoia National Monument.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1
Agee J. K. (1998). “Fire and pine ecosystems” in Ecology and biogeography of Pinus. ed. RichardsonD. M. (Cambridge, UK: The Press Syndicate of the University of Cambridge), 193–218.
2
Allen C. D. Macalady A. K. Chenchouni H. Bachelet D. McDowell N. Vennetier M. et al . (2010). A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag.259, 660–684. doi: 10.1016/j.foreco.2009.09.001
3
Anderegg W. R. L. Kane J. M. Anderegg L. D. L. (2013). Consequences of widespread tree mortality triggered by drought and temperature stress. Nat. Clim. Chang.3, 30–36. doi: 10.1038/nclimate1635
4
Bates D. Maechler M. Bolker B. Walker S. (2023). lme4: linear mixed-effects models using “eigen” and S4. Available online at: https://CRAN.R-project.org/package=lme4 (Accessed March 22, 2023).
5
Bentz B. J. Regniere J. Fettig C. J. Hansen E. M. Hayes J. L. Hicke J. A. et al . (2010). Climate change and bark beetles of the Western United States and Canada: direct and indirect effects. Bioscience60, 602–613. doi: 10.1525/bio.2010.60.8.6
6
Bisbing S. M. Nagelson P. B. Cox L. E. Phelan C. N. (2025). Post-mortem silviculture meets fuel reduction objective in drought-impacted southern Sierra Nevada forests. For. Ecol. Manag.595:123035. doi: 10.1016/j.foreco.2025.123035
7
Bottero A. D’Amato A. W. Palik B. J. Bradford J. B. Fraver S. Battaglia M. A. et al . (2017). Density-dependent vulnerability of forest ecosystems to drought. J. Appl. Ecol.54, 1605–1614. doi: 10.1111/1365-2664.12847
8
Bradford J. B. Bell D. M. (2017). A window of opportunity for climate-change adaptation: easing tree mortality by reducing forest basal area. Front. Ecol. Environ.15, 11–17. doi: 10.1002/fee.1445
9
Brodie E. G. Knapp E. E. Latimer A. M. Safford H. D. Vossmer M. Bisbing S. M. (2023). The century-long shadow of fire exclusion: historical data reveal early and lasting effects of fire regime change on contemporary forest composition. For. Ecol. Manag.539:121011. doi: 10.1016/j.foreco.2023.121011
10
Brown J. K. (1974) Handbook for inventorying downed woody material. USDA Forest Service GTR INT‐16. Ogden, UT. U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station.
11
Collins B. J. Rhoades C. C. Hubbard R. M. Battaglia M. A. (2011). Tree regeneration and future stand development after bark beetle infestation and harvesting in Colorado lodgepole pine stands. For. Ecol. Manag.261, 2168–2175. doi: 10.1016/j.foreco.2011.03.016
12
Dixon G. (2018). Essential FVS: A user’s guide to the Forest vegetation simulator. Ft. Collins, CO: USDA Forest Service.
13
Earles J. M. North M. P. Hurteau M. D. (2014). Wildfire and drought dynamics destabilize carbon stores of fire-suppressed forests. Ecol. Appl.24, 732–740. doi: 10.1890/13-1860.1
14
Fettig C. J. Mortenson L. A. Bulaon B. M. Foulk P. B. (2019). Tree mortality following drought in the central and southern Sierra Nevada, California, U.S.For. Ecol. Manag.432, 164–178. doi: 10.1016/j.foreco.2018.09.006
15
Filip G. M. Maffei H. Chadwick K. (2007). Forest health decline in a Central Oregon mixed-conifer forest revisited after wildfire: a 25-year case study. West. J. Appl. For.22, 278–284. doi: 10.1093/wjaf/22.4.278
16
Fowells H. A. Stark N. B. (1965). Natural regeneration in relation to environment in the mixed conifer forest type of California. Berkeley: USDA Forest Service Research.
17
Fox J. Weisberg S. Price B. Adler D. Bates D. Baud-Bovy G. et al . (2019). CAR: Companion to Applied Regression. Available online at: https://CRAN.R-project.org/package=car (Accessed March 22, 2023).
18
Fraver S. Jain T. Bradford J. B. D’Amato A. W. Kastendick D. Palik B. et al . (2011). The efficacy of salvage logging in reducing subsequent fire severity in conifer-dominated forests of Minnesota, USA. Ecol. Appl.21, 1895–1901. doi: 10.1890/11-0380.1
19
Ganey J. L. Vojta S. C. (2010). Coarse woody debris assay in northern Arizona mixed-conifer and ponderosa pine forests. Ft. Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station.
20
Giger D. R. Schmitt G. J. (1983). Soil survey of the Sierra National Forest Area. California: USDA Forest Service.
21
Hanes R. O. Plocher S. E. Martynn D. Z. (1996). Soil survey of Sequoia National Forest. California: USDA Forest Service Pacific Southwest Region.
22
Harmon M. E. Cromack K. Smith B. G. (1987). Coarse woody debris in mixed-conifer forests, Sequoia National Park, California. Can. J. For. Res.17, 1265–1272. doi: 10.1139/x87-196
23
Hood S. M. Baker S. Sala A. (2016). Fortifying the forest: thinning and burning increase resistance to a bark beetle outbreak and promote forest resilience. Ecol. Appl.26, 1984–2000. doi: 10.1002/eap.1363
24
Hood P. R. Nelson K. N. Rhoades C. C. Tinker D. B. (2017). The effect of salvage logging on surface fuel loads and fuel moisture in beetle-infested lodgepole pine forests. For. Ecol. Manag.390, 80–88. doi: 10.1016/j.foreco.2017.01.003
25
Jaime L. Batllori E. Ferretti M. Lloret F. (2022). Climatic and stand drivers of forest resistance to recent bark beetle disturbance in European coniferous forests. Glob. Chang. Biol.28, 2830–2841. doi: 10.1111/gcb.16106
26
Kautz M. Meddens A. J. H. Hall R. J. Arneth A. (2017). Biotic disturbances in northern hemisphere forests – a synthesis of recent data, uncertainties and implications for forest monitoring and modelling. Glob. Ecol. Biogeogr.26, 533–552. doi: 10.1111/geb.12558
27
Keeley J. E. Zedler P. H. (1998). “Evolution of life histories in Pinus” in Ecology and biogeography of Pinus. ed. RichardsonD. M. (Cambridge: Cambridge University Press), 219–250.
28
Keen R. M. Voelker S. L. Wang S. Y. S. Bentz B. J. Goulden M. L. Dangerfield C. R. et al . (2022). Changes in tree drought sensitivity provided early warning signals to the California drought and forest mortality event. Glob. Change Biol.28, 1119–1132. doi: 10.1111/gcb.15973
29
Keyes C. R. Manso González R. (2015). Climate-influenced ponderosa pine (Pinus ponderosa) seed masting trends in western Montana, USA. For. Syst.24:021. doi: 10.5424/fs/2015241-05606
30
Keyser T. L. Smith F. W. Shepperd W. D. (2009). Short-term impact of post-fire salvage logging on regeneration, hazardous fuel accumulation, and understorey development in ponderosa pine forests of the Black Hills, SD, USA. Int. J. Wildland Fire18, 451–458. doi: 10.1071/wf08004
31
Knapp E. E. (2015). Long-term dead wood changes in a Sierra Nevada mixed conifer forest: habitat and fire hazard implications. For. Ecol. Manag.339, 87–95. doi: 10.1016/j.foreco.2014.12.008
32
Knapp E. E. Bernal A. A. Kane J. M. Fettig C. J. North M. P. (2021). Variable thinning and prescribed fire influence tree mortality and growth during and after a severe drought. For. Ecol. Manag.479:118595. doi: 10.1016/j.foreco.2020.118595
33
Kurz W. A. Dymond C. C. Stinson G. Rampley G. J. Neilson E. T. Carroll A. L. et al . (2008). Mountain pine beetle and forest carbon feedback to climate change. Nature452, 987–990. doi: 10.1038/nature06777
34
Kuznetsova A. Brockhoff P. B. Christensen R. H. B. Jensen S. P. (2020). LmerTest: tests in linear mixed effects models. Available online at: https://CRAN.R-project.org/package=lmerTest (Accessed March 22, 2023).
35
Laiho R. Prescott C. E. (2004). Decay and nutrient dynamics of coarse woody debris in northern coniferous forests: a synthesis. Can. J. For. Res.34, 763–777. doi: 10.1139/x03-241
36
Legras E. C. Vander Wall S. B. Board D. I. (2010). The role of germination microsite in the establishment of sugar pine and Jeffrey pine seedlings. For. Ecol. Manag.260, 806–813. doi: 10.1016/j.foreco.2010.05.039
37
Lenth R. V. Bolker B. Buerkner P. Giné-Vázquez I. Herve M. Jung M. et al (2024). Emmeans: Estimated Marginal Means, aka Least-Squares Means. Available online at: https://cran.r-project.org/web/packages/emmeans/index.html (Accessed March 21, 2024).
38
Leverkus A. B. Gustafsson L. Lindenmayer D. B. Castro J. Rey Benayas J. M. Ranius T. et al . (2020). Salvage logging effects on regulating ecosystem services and fuel loads. Front. Ecol. Environ.18, 391–400. doi: 10.1002/fee.2219
39
Leverkus A. B. Polo I. Baudoux C. Thorn S. Gustafsson L. Rubio de Casas R. (2021). Resilience impacts of a secondary disturbance: Meta-analysis of salvage logging effects on tree regeneration. J. Ecol.109, 3224–3232. doi: 10.1111/1365-2745.13581
40
Leverkus A. B. Rey Benayas J. M. Castro J. Boucher D. Brewer S. Collins B. M. et al . (2018). Salvage logging effects on regulating and supporting ecosystem services — a systematic map. Can. J. For. Res.48, 983–1000. doi: 10.1139/cjfr-2018-0114
41
Liang S. Hurteau M. D. Westerling A. L. (2017). Response of Sierra Nevada forests to projected climate–wildfire interactions. Glob. Chang. Biol.23, 2016–2030. doi: 10.1111/gcb.13544
42
Loudermilk E. L. Scheller R. M. Weisberg P. J. Kretchun A. (2017). Bending the carbon curve: fire management for carbon resilience under climate change. Landsc. Ecol.32, 1461–1472. doi: 10.1007/s10980-016-0447-x
43
Lüdecke D. Ben-Shachar M. S. Patil I. Waggoner P. Makowski D. (2021). Performance: an {R} package for assessment, comparison and testing of statistical models. Available online at: https://cran.r-project.org/package=performance (Accessed March 22, 2023).
44
Maestrini B. Alvey E. C. Hurteau M. D. Safford H. Miesel J. R. (2017). Fire severity alters the distribution of pyrogenic carbon stocks across ecosystem pools in a Californian mixed-conifer forest. J. Geophys. Res. Biogeosciences122, 2338–2355. doi: 10.1002/2017JG003832
45
Maxwell C. J. Scheller R. M. Wilson K. N. Manley P. N. (2022). Assessing the effectiveness of landscape-scale forest adaptation actions to improve resilience under projected climate change. Front. For. Glob. Change5:740869. doi: 10.3389/ffgc.2022.740869
46
McDonald P. M. (1992). Estimating seed crops of conifer and hardwood species. Can. J. For. Res.22, 832–838. doi: 10.1139/x92-112
47
Miller R. O. Gavlak R. Horneck D. (2013). “Western region method S - 9.20” in Soil, plant, and water reference methods for the Western region - 4th edition. pp. 77–78.
48
Minnich R. A. (2007). “Climate, paleoclimate, and paleovegetation” in Terrestrial vegetation of California. eds. BarbourM. G.Keeler-WolfT.SchoenherrA. A. (Berkeley, CA: University of California Press), 43–70.
49
Moore J. Pope J. Woods M. Ellis A. (2018). 2017 aerial survey results: California. Davis, CA: USDA Forest Service Forest Health Monitoring Program.
50
Mueller R. C. Scudder C. M. Porter M. E. Talbot Trotter R. Iii Gehring C. A. Whitham T. G. (2005). Differential tree mortality in response to severe drought: evidence for long-term vegetation shifts. J. Ecol.93, 1085–1093. doi: 10.1111/j.1365-2745.2005.01042.x
51
National Drought Mitigation Center (2016) U.S. drought monitor. Available online at: https://droughtmonitor.unl.edu/DmData/GISData.aspx (Accessed August 15, 2022).
52
North M. P. Collins B. M. Safford H. D. Stephenson N. L. (2016). “Montane forests” in Ecosystems of California. eds. MooneyH. A.ZavaletaE. (Berkeley, CA: University of California Press), 553–577.
53
North M. P. Tompkins R. E. Bernal A. A. Collins B. M. Stephens S. L. York R. A. (2022). Operational resilience in western US frequent-fire forests. For. Ecol. Manag.507:120004. doi: 10.1016/j.foreco.2021.120004
54
Northrop H. Axelson J. N. Das A. J. Stephenson N. L. Vilanova E. Stephens S. L. et al . (2024). Snag dynamics and surface fuel loads in the Sierra Nevada: predicting the impact of the 2012–2016 drought. For. Ecol. Manag.551:121521. doi: 10.1016/j.foreco.2023.121521
55
Parker A. J. (1982). The topographic relative moisture index: an approach to soil-moisture assessment in mountain terrain. Phys. Geogr.3, 160–168. doi: 10.1080/02723646.1982.10642224
56
R Core Team (2023). R: a language and environment for statistical computing. Vienna: R Foundation for Statistical Computing.
57
Raffa K. F. Aukema B. H. Bentz B. J. Carroll A. L. Hicke J. A. Turner M. G. et al . (2008). Cross-scale drivers of natural disturbances prone to anthropogenic amplification: the dynamics of bark beetle eruptions. Bioscience58, 501–517. doi: 10.1641/B580607
58
Rebain S. (2015). The fire and fuels extension to the Forest Vegetation Simulator: updated model documentation. Ft. Collins, CO: USDA Forest Service, Forest Management Service Center.
59
Reed C. C. Hood S. M. Cluck D. R. Smith S. L. (2023). Fuels change quickly after California drought and bark beetle outbreaks with implications for potential fire behavior and emissions. Fire Ecol.19:16. doi: 10.1186/s42408-023-00175-6
60
Restaino C. Young D. J. N. Estes B. Gross S. Wuenschel A. Meyer M. et al . (2019). Forest structure and climate mediate drought-induced tree mortality in forests of the Sierra Nevada, USA. Ecol. Appl.29:e01902. doi: 10.1002/eap.1902
61
Rhoades C. C. Hubbard R. M. Elder K. Fornwalt P. J. Schnackenberg E. Hood P. R. et al . (2020). Tree regeneration and soil responses to management alternatives in beetle-infested lodgepole pine forests. For. Ecol. Manag.468:118182. doi: 10.1016/j.foreco.2020.118182
62
Ritchie M. Berrill J.-P. Hammett E. Long J. W. (2024). Early responses to crown modification of California black oak sprouts initiated by high-severity wildfire. J. For.122, 70–78. doi: 10.1093/jofore/fvad038
63
Robbins Z. J. Xu C. Aukema B. H. Buotte P. C. Chitra-Tarak R. Fettig C. J. et al . (2022). Warming increased bark beetle-induced tree mortality by 30% during an extreme drought in California. Glob. Chang. Biol.28, 509–523. doi: 10.1111/gcb.15927
64
Robbins Z. J. Xu C. Jonko A. Chitra-Tarak R. Fettig C. J. Costanza J. et al . (2023). Carbon stored in live ponderosa pines in the Sierra Nevada will not return to pre-drought (2012) levels during the 21st century due to bark beetle outbreaks. Front. Environ. Sci.11:1112756. doi: 10.3389/fenvs.2023.1112756
65
RStudio Team (2023) RStudio: integrated development for R. Available online at: http://www.rstudio.com/
66
Safford H. D. Stevens J. T. (2017). Natural range of variation for yellow pine and mixed-conifer forests in the Sierra Nevada, southern cascades, and Modoc and Inyo National Forests, California, USA. Gen. Tech. Rep. PSW-GTR-256. Albany, CA: U.S. Department of Agriculture.
67
Safford H. D. Vallejo V. R. (2019). “Ecosystem management and ecological restoration in the Anthropocene: integrating global change, soils, and disturbance in boreal and Mediterranean forests” in Developments in soil science. Eds. M. Busse, C. P. Giardina, D. M. Morris, and D. S. Page-Dumroese (Cambridge, MA, USA: Elsevier), 259–308.
68
Seidl R. Thom D. Kautz M. Martin-Benito D. Peltoniemi M. Vacchiano G. et al . (2017). Forest disturbances under climate change. Nat. Clim. Chang.7, 395–402. doi: 10.1038/nclimate3303
69
Seidl R. Turner M. G. (2022). Post-disturbance reorganization of forest ecosystems in a changing world. Proc. Natl. Acad. Sci.119:e2202190119. doi: 10.1073/pnas.2202190119
70
Senf C. Buras A. Zang C. S. Rammig A. Seidl R. (2020). Excess forest mortality is consistently linked to drought across Europe. Nat. Commun.11:6200. doi: 10.1038/s41467-020-19924-1
71
Sieg C. H. Linn R. R. Pimont F. Hoffman C. M. McMillin J. D. Winterkamp J. et al . (2017). Fires following bark beetles: factors controlling severity and disturbance interactions in ponderosa pine. Fire Ecol.13, 1–23. doi: 10.4996/fireecology.130300123
72
Sohn J. A. Saha S. Bauhus J. (2016). Potential of forest thinning to mitigate drought stress: a meta-analysis. For. Ecol. Manag.380, 261–273. doi: 10.1016/j.foreco.2016.07.046
73
Stephens S. L. Bernal A. A. Collins B. M. Finney M. A. Lautenberger C. Saah D. (2022). Mass fire behavior created by extensive tree mortality and high tree density not predicted by operational fire behavior models in the southern Sierra Nevada. For. Ecol. Manag.518:120258. doi: 10.1016/j.foreco.2022.120258
74
Stephens S. L. Collins B. M. Fettig C. J. Finney M. A. Hoffman C. M. Knapp E. E. et al . (2018). Drought, tree mortality, and wildfire in forests adapted to frequent fire. Bioscience68, 77–88. doi: 10.1093/biosci/bix146
75
Stephens S. L. Hall L. Stephens C. W. Bernal A. A. Collins B. M. (2023). Degradation and restoration of indigenous California black oak (Quercus kelloggii) stands in the northern Sierra Nevada. Fire Ecol.19:12. doi: 10.1186/s42408-023-00172-9
76
Stephens S. L. Lydersen J. M. Collins B. M. Fry D. L. Meyer M. D. (2015). Historical and current landscape-scale ponderosa pine and mixed conifer forest structure in the southern Sierra Nevada. Ecosphere6, 1–63. doi: 10.1890/es14-00379.1
77
Thomas J. W. (1979). Wildlife habitats in managed forests the Blue Mountains of Oregon and Washington. Washington, DC: US Department of Agriculture, Forest Service.
78
UC Davis (2022). Soil Data Explorer. Available online at: https://casoilresource.lawr.ucdavis.edu/sde/ (Accessed April 18, 2022).
79
USDA Forest Service (2023). Forest vegetation simulator (FVS-online). Available online at: https://charcoal2.cnre.vt.edu/FVSOnline/ (Accessed September 20, 2023).
80
Van Wagtendonk J. W. Benedict J. M. Sydoriak W. M. (1998). Fuel bed characteristics of Sierra Nevada conifers. West. J. Appl. For.13, 73–84. doi: 10.1093/wjaf/13.3.73
81
Vernon M. J. Sherriff R. L. van Mantgem P. Kane J. M. (2018). Thinning, tree-growth, and resistance to multi-year drought in a mixed-conifer forest of northern California. For. Ecol. Manag.422, 190–198. doi: 10.1016/j.foreco.2018.03.043
82
Voelker S. L. Merschel A. G. Meinzer F. C. Ulrich D. E. M. Spies T. A. Still C. J. (2019). Fire deficits have increased drought sensitivity in dry conifer forests: fire frequency and tree-ring carbon isotope evidence from Central Oregon. Glob. Chang. Biol.25, 1247–1262. doi: 10.1111/gcb.14543
83
Wang W. Peng C. Kneeshaw D. D. Larocque G. R. Luo Z. (2012). Drought-induced tree mortality: ecological consequences, causes, and modeling. Environ. Rev.20, 109–121. doi: 10.1139/a2012-004
84
Wayman R. B. Safford H. D. (2021). Recent bark beetle outbreaks influence wildfire severity in mixed-conifer forests of the Sierra Nevada, California, USA. Ecol. Appl.31:e02287. doi: 10.1002/eap.2287
85
Wayman R. B. Sorenson Q. M. (2025). Dead tree removal after drought mortality increases understory plant diversity in a mixed conifer forest. Ecosphere16:e70215. doi: 10.1002/ecs2.70215
86
Williams A. P. Allen C. D. Macalady A. K. Griffin D. Woodhouse C. A. Meko D. M. et al . (2013). Temperature as a potent driver of regional forest drought stress and tree mortality. Nat. Clim. Chang.3, 292–297. doi: 10.1038/nclimate1693
87
Williams A. P. Allen C. D. Millar C. I. Swetnam T. W. Michaelsen J. Still C. J. et al . (2010). Forest responses to increasing aridity and warmth in the southwestern United States. Proc. Natl. Acad. Sci.107, 21289–21294. doi: 10.1073/pnas.0914211107
88
Young D. J. N. Meyer M. Estes B. Gross S. Wuenschel A. Restaino C. et al . (2019). Forest recovery following extreme drought in California, USA: natural patterns and effects of pre-drought management. Ecol. Appl.30:e02002. doi: 10.1002/eap.2002
89
Young D. J. Stevens J. T. Earles J. M. Moore J. Ellis A. Jirka A. L. et al . (2017). Long-term climate and competition explain forest mortality patterns under extreme drought. Ecol. Lett.20, 78–86. doi: 10.1111/ele.12711
Summary
Keywords
bark beetle, tree regeneration, salvage logging, mixed-conifer forest, fuels, drought mortality, fire severity
Citation
Sorenson QM, Wayman RB, Ursell TL and Safford HD (2025) Removing dead trees after mass drought mortality enhances fire-adapted tree recruitment, reduces future fire severity, and has mixed effects on carbon stocks. Front. For. Glob. Change 8:1691015. doi: 10.3389/ffgc.2025.1691015
Received
22 August 2025
Accepted
20 October 2025
Published
10 November 2025
Volume
8 - 2025
Edited by
José Manuel Fernández-Guisuraga, University of León, Spain
Reviewed by
John Bailey, Oregon State University, United States; Luiza Gonçalves Lazzaro, The Pennsylvania State University, United States
Updates
Copyright
© 2025 Sorenson, Wayman, Ursell and Safford.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rebecca Bewley Wayman, rbwayman@ucdavis.edu
‡These authors have contributed equally to this work and share first authorship
† Present addresses: Rebecca Bewley Wayman, California Department of Forestry and Fire Protection, Fire and Resource Assessment Program, Sacramento, CA, United States Tara L. Ursell, California Air Resources Board, Sacramento, CA, United States
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.