 Lucy O. Keatts1*
Lucy O. Keatts1* Martin Robards2
Martin Robards2 Sarah H. Olson1
Sarah H. Olson1 Karsten Hueffer3
Karsten Hueffer3 Stephen J. Insley4,5
Stephen J. Insley4,5 Damien O. Joly6
Damien O. Joly6 Susan Kutz7
Susan Kutz7 David S. Lee8
David S. Lee8 Cheryl-Lesley B. Chetkiewicz4Stéphane Lair9Nicholas D. Preston10
Cheryl-Lesley B. Chetkiewicz4Stéphane Lair9Nicholas D. Preston10 Mathieu Pruvot1,7
Mathieu Pruvot1,7 Justina C. Ray4
Justina C. Ray4 Donald Reid4
Donald Reid4 Jonathan M. Sleeman11
Jonathan M. Sleeman11 Raphaela Stimmelmayr12,13
Raphaela Stimmelmayr12,13 Craig Stephen14,15Chris Walzer1,16
Craig Stephen14,15Chris Walzer1,16- 1Wildlife Conservation Society Health Program, Bronx, NY, United States
- 2Wildlife Conservation Society, Arctic Beringia Program, Fairbanks, AK, United States
- 3Department of Veterinary Medicine & Arctic and Northern Studies Program, University of Alaska Fairbanks, Fairbanks, AK, United States
- 4Wildlife Conservation Society Canada, Toronto, ON, Canada
- 5Department of Biology, University of Victoria, Victoria, BC, Canada
- 6Nyati Health Consulting, Nanaimo, BC, Canada
- 7Department of Ecosystem and Public Health, Faculty of Veterinary Medicine, University of Calgary, Calgary, AB, Canada
- 8Department of Wildlife and Environment, Nunavut Tunngavik Inc., Ottawa, ON, Canada
- 9Canadian Wildlife Health Cooperative, Université de Montréal, Montreal, QC, Canada
- 10Salmon Coast Field Station, Echo Bay, BC, Canada
- 11United States Geological Survey National Wildlife Health Center, Madison, WI, United States
- 12North Slope Department of Wildlife Management, Utqiagvik, AK, United States
- 13Institute of Arctic Biology, University of Alaska Fairbanks, Fairbanks, AK, United States
- 14University of British Columbia, Vancouver, BC, Canada
- 15Ross University School of Veterinary Medicine, Basseterre, Saint Kitts and Nevis
- 16Conservation Medicine Unit, Department of Interdisciplinary Life Sciences, Research Institute of Wildlife Ecology, University of Veterinary Medicine, Vienna, Austria
The COVID-19 pandemic has re-focused attention on mechanisms that lead to zoonotic disease spillover and spread. Commercial wildlife trade, and associated markets, are recognized mechanisms for zoonotic disease emergence, resulting in a growing global conversation around reducing human disease risks from spillover associated with hunting, trade, and consumption of wild animals. These discussions are especially relevant to people who rely on harvesting wildlife to meet nutritional, and cultural needs, including those in Arctic and boreal regions. Global policies around wildlife use and trade can impact food sovereignty and security, especially of Indigenous Peoples. We reviewed known zoonotic pathogens and current risks of transmission from wildlife (including fish) to humans in North American Arctic and boreal biomes, and evaluated the epidemic and pandemic potential of these zoonoses. We discuss future concerns, and consider monitoring and mitigation measures in these changing socio-ecological systems. While multiple zoonotic pathogens circulate in these systems, risks to humans are mostly limited to individual illness or local community outbreaks. These regions are relatively remote, subject to very cold temperatures, have relatively low wildlife, domestic animal, and pathogen diversity, and in many cases low density, including of humans. Hence, favorable conditions for emergence of novel diseases or major amplification of a spillover event are currently not present. The greatest risk to northern communities from pathogens of pandemic potential is via introduction with humans visiting from other areas. However, Arctic and boreal ecosystems are undergoing rapid changes through climate warming, habitat encroachment, and development; all of which can change host and pathogen relationships, thereby affecting the probability of the emergence of new (and re-emergence of old) zoonoses. Indigenous leadership and engagement in disease monitoring, prevention and response, is vital from the outset, and would increase the success of such efforts, as well as ensure the protection of Indigenous rights as outlined in the United Nations Declaration on the Rights of Indigenous Peoples. Partnering with northern communities and including Indigenous Knowledge Systems would improve the timeliness, and likelihood, of detecting emerging zoonotic risks, and contextualize risk assessments to the unique human-wildlife relationships present in northern biomes.
Introduction
Emerging infectious diseases (EIDs) are a significant burden on public health and economies, and are increasingly recognized as a global threat (1). EIDs are currently defined by the World Health Organization (WHO) as those that “have newly appeared in a population or have existed but are rapidly increasing in incidence or geographic range (2).” This definition does not clarify between different categories of emergence or re-emergence, nor does it clearly differentiate novel diseases with pandemic potential, such as COVID-19, from those that are variants of old pathogens, new detections of old pathogens with novel technologies, or re-emergence of old pathogens in new regions (2). Thus, the definition does not reflect the very different drivers and significance between diseases and pathogens in terms of global vs. local burden, threat and origin (2). The majority of EIDs (over 60%) are considered zoonotic, with zoonoses defined as “any infection that is naturally transmissible from vertebrate animals to humans” (3–7). There is, however, a need to better differentiate between diseases that originate in animals but are subsequently independently perpetuated in human populations, and those that require an animal host for pathogen persistence, to target research, control, policy and mitigation efforts (2). Infection of people with zoonotic pathogens occurs through contact with infected animals via a variety of mechanisms including: direct contact with bodily fluids (e.g., saliva, blood, urine, feces); indirect contact with surfaces contaminated with an animal's infectious secretions; vector-borne through biting arthropods; foodborne through consumption of contaminated raw or undercooked food; and waterborne, via contaminated drinking water (8). The definition of “risk” usually considers two dimensions: how likely the uncertainty is to occur (probability), and what the effect would be if it happened (impact) (9). Both components are important when establishing the local vs. global risks of zoonoses and emerging diseases of animal-origin, and in considering policy or other interventions.
Severe Acute Respiratory Syndrome (SARS)-CoV-2, considered a novel zoonotic coronavirus, and the causative agent for COVID-19, emerged in December of 2019 in Wuhan, China and rapidly spread globally, to devastating effect. Comparative genomic analysis indicates that SARS-CoV-2 evolved naturally, with bats the likely ancestral reservoir host (10). Given that the animal reservoir for SARS-CoV-2 is yet to be identified, others propose the virus be classified an “emerging infectious disease (EID) of probable animal origin” rather than a zoonosis (2). Many, but not all, early cases of COVID-19 were associated with a market in Wuhan that traded in wildlife (11), and plausible scenarios have been put forth to explain the origin of SARS-CoV-2: evolution in bats or an intermediate animal host before zoonotic spillover to humans in the market or market trade chain (12, 13); or natural evolution in humans following direct zoonotic transfer from bats (12), though purifying selection of SARS-CoV-2 in humans since the start of the pandemic has been weak compared with the significant positive diversifying selection that has occurred in bats since SARS-CoV-2 evolved from its closest known relative RmYN02 (13). SARS-CoV-2 shares some genetic similarity with SARS-CoV, and prevailing evidence suggests that SARS-CoV spilled over into humans via an intermediate host–likely masked palm civets (Paguma larvata)–at a wildlife market in southern China in 2002 (14–17). The most devastating pandemics in human history, the Black Death (in the 1300s), Spanish influenza (1918), and HIV/AIDS, all resulted from an initial zoonotic spillover from wildlife (18). Of currently defined emerging zoonoses, over 70% originate in wildlife (6, 19–21) however, very few diseases of wild animal origin persist with ongoing zoonotic transmission, with most human infections from such diseases being acquired through human-human transmission. The frequency of zoonotic disease spillover into humans is increasing (1, 19, 22, 23). This rise has been linked to changing human ecology, due to the growing global human population, and its demand for food, land, and natural resources (5, 19, 20, 24). Escalating anthropogenic activities are increasing contact rates between humans, domestic animals, and wildlife (19, 25–28).
Wild animal consumption-based food systems have been implicated in the emergence of diseases with zoonotic origins (including HIV, SARS, Ebola virus disease, Avian Influenza A), and mounting evidence indicates substantial human health risks from the trade in live wildlife (29–40). Members of the WHO team investigating the origins of the SARS-CoV-2 pandemic recently reported that wildlife farms were the most likely source (41). The trade of wild-harvested meat for food fills a continuum from subsistence-based rural consumption, to extensive commercial trade networks to meet growing urban and international demand for wild animal meat as a luxury product (42). There are calls from scientific, health, conservation organizations, and government officials to end commercial trade in wildlife for human consumption. Closing wild animal markets and the trade in wildlife, is viewed as an expedient measure to reduce the risk of future viral outbreaks, like the COVID-19 pandemic, that threaten human health, well-being, economies, and security at local, regional, and global scales (43, 44). Public health organizations, including the WHO, support the rigorous enforcement of bans on the sale and trade of wildlife for food (45). However, some populations are dependent on wild-harvested food to meet basic nutritional requirements. In the face of disruptions to food supply chains during the COVID-19 pandemic, harvesting and sharing of local foods has helped maintain food and nutrition security for isolated boreal and Arctic communities (46), and harvesting of wildlife, with secondary use of wildlife by-products (i.e., pelts, claws, skulls), forms an important part of traditional economies and socio-economic-cultural well-being of Indigenous Peoples. The World Organization for Animal Health (OIE) considers wildlife all of: (a) wild animals (b) free-ranging feral [domesticated] animals, and (c) non-domestic animals in captivity or farming, though these different categories and interfaces likely pose quite different risks for spillover to humans (2). Infection cycles in densely populated wildlife farms, along trade chains and live markets, are very different to those in natural, free-ranging populations. These crowded and stressful environments are much more likely to facilitate cross-species transmission of pathogens with pandemic potential (2, 32, 47). Based on past experiences, well-meaning initiatives aimed at halting the hunting, trapping, and use of wildlife risk violating the rights of Indigenous Peoples as outlined in the United Nations (UN) Declaration on the Rights of Indigenous Peoples (48), and also threaten the subsistence of non-Indigenous hunter communities.
Indigenous Peoples and local communities across the boreal and Arctic regions (or “northern communities”) have extensive and essential relationships with their environments, including through the harvest and sharing of nutritionally and spiritually important native plants, and wildlife, also known as traditional and country foods (49–53). Limited access to traditional and country foods is a strong predictor of health disparities in Indigenous Peoples across the North, and is correlated to diseases such as diabetes, cardiovascular disease and mental illness (54). Throughout boreal and Arctic ecosystems, subsistence economies support rich and diverse cultures that include the use, sharing, and consumption of wildlife. Subsistence is considered as: the personal consumption of wildlife for food, fuel, shelter, clothing, tools, or transportation; the barter, trade, or sharing of wildlife products in their harvested form with relatives, with others in the local community or with persons in locations other than the local community with whom local residents share familial, social, cultural, or economic ties; and the making and selling of handicrafts from wildlife products, when the wildlife are harvested for the purposes defined above (55). Subsistence, together with guided hunting and fishing, by Indigenous Peoples in the Arctic and boreal is best considered through the lens of food security and sovereignty, cultural security, and livelihoods (56, 57). Food security is considered to exist when “all people, at all times, have physical, social, and economic access to sufficient, safe, and nutritious food to meet their dietary needs, and food preferences, for a healthy and active life” (58). Food sovereignty refers to the “ability and right of people to define their own policies and strategies for sustainable production, distribution, and consumption of food that guarantees the right to food for the entire population” (59). The rights of Indigenous Peoples to determine and maintain these relationships with the environment are embedded in the UNDRIP (48), in the Canadian Constitution (60), and in the Truth and Reconciliation Commission of Canada's Calls to Action (61), but have received less formalized protections within the United States. The relationships between Indigenous Peoples in North America and the environment, particularly the harvest and use of wildlife, are the basis for historic numbered Treaties, and modern land claim agreements in Canada and the United States, Native to Native agreements, and Indigenous advisory and co-management institutions (see Table 1 for specific examples). Such agreements however, have not been as effective in addressing the needs of Indigenous Peoples and local communities as they are forced to adapt to the impacts of biodiversity loss and climate change within historically colonial approaches to environmental management (62). The use of wildlife by non-Indigenous local communities are largely managed through legislation and regulations developed, and enforced, by governments and regional authorities.

Table 1. Examples of formal agreements and co-management institutions protecting the rights of Indigenous Peoples in Arctic and boreal biomes across North America regarding land, hunting, and use of wildlife.
Indigenous Peoples in the Arctic and boreal are often at the forefront of protecting wild food systems, livelihoods, and cultural values (63, 64), and concerns have been raised about potential impacts to Indigenous rights and food sovereignty due to policy initiatives focused on hunting, consumption, sharing, and local trade of wildlife (55, 65–70). Negative impacts have occurred when socio-economic, cultural, and nutritional dimensions of Indigenous subsistence practices were not considered. For example, previous culturally insensitive and poorly developed communication outreach efforts regarding health and traditional and country foods (on contaminants) resulted in negative health consequences for affected populations from avoidance of traditional foods altogether, given limited healthy alternatives (71, 72). Zoonotic health risks and concerns are also relevant to local communities of hunters, trappers, and fishers across the region. This paper was written to respond to some of the issues related to northern food sovereignty, and to apprehension expressed by researchers and veterinarians working with these northern communities around potential routes for introduction of emerging zoonoses, such as COVID-19, into remote Indigenous and local communities where subsistence based on traditional and mixed economies remains a vital necessity. The manuscript aims to (i) review zoonotic pathogens of wildlife origin in Arctic and boreal systems, in the context of wildlife use by northern communities; (ii) provide a reference for northern communities, and wildlife disease and public health researchers, to consider and compare, the potential community and broader health implications of zoonoses transmissible via traditional use of wildlife; (iii) examine zoonotic pathogen spillover, amplification, epidemic or pandemic spread, and relevant dynamics in northern biomes; (iv) describe some future zoonotic concerns; and (v) discuss considerations for current and future monitoring, surveillance and risk reduction approaches. The results section, with Table 2, addresses aims 1 and 2, and the remaining aims are addressed in the discussion.
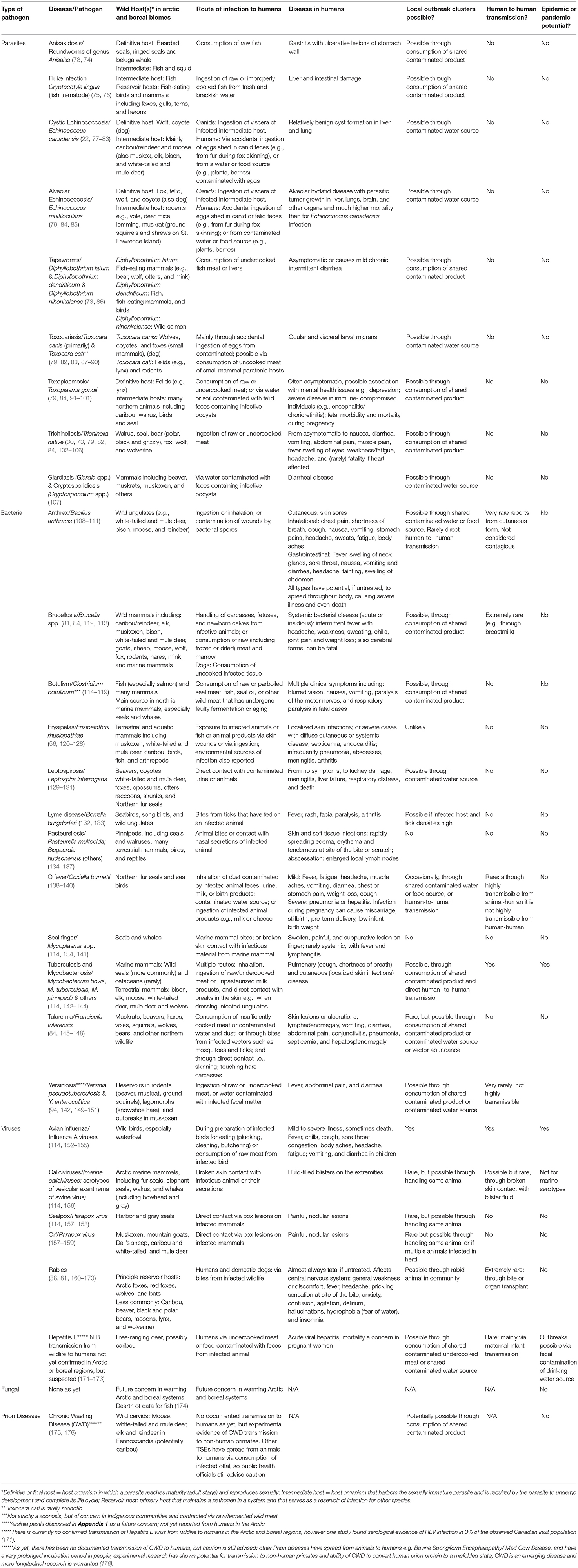
Table 2. Zoonotic pathogens of Arctic and boreal systems that can be transmitted to humans through hunting, consumption or other use of wildlife, noting pathogen potential for local outbreak clusters, human-to-human transmission, and epidemic/pandemic spread.
Materials and Methods
We carried out a qualitative literature review to identify publications focused on zoonotic diseases in Arctic and boreal biomes with a potential wildlife origin. The review focused on zoonoses associated with the hunting, trapping, butchering, sharing, use, and trade of wildlife (including fish), and only considered agricultural or domestic species if related to a sylvatic cycle. Given this focus, vector-borne zoonoses were not included in the review, but are considered in the discussion. In addition, we asked for the contributions of experts on the topics. Only published materials were included. Our search included articles, reviews, proceedings papers, reports and book chapters in the English language. The Web of Science database and Google Scholar search engine were utilized to conduct the search in late April, 2020. Publications from any year were considered.
To gain an overview of the literature pertaining to existing zoonotic disease reports for communities in Arctic and boreal regions associated with consumption and/or hunting of wildlife, search terms were conducted using the following keywords and phrases:
➢ “zoonoses” OR “zo>onotic”
➢ AND “wildlife” OR “fish” OR “hunter” OR “hunting” OR “hunted” OR “subsistence” OR “country food” OR “traditional food” OR “Indigenous”
➢ AND “Arctic” OR “boreal” OR “Alaska” OR “northern Canada”
Based on the titles and abstracts of the identified studies, we excluded those publications considered irrelevant to part one of our review, including those for which the full manuscript was not available, publications pertaining to vector-borne zoonoses in Northern regions; those describing zoonoses in other regions, or for which hunting, consumption, use or sharing of wildlife have not been reported to facilitate transmission; and those referring to experimental rather than natural infections. Relevance was attributed if the publications discussed case studies of zoonotic diseases transmitted to humans via the hunting, consumption, preparation or sharing of wildlife in northern regions of North America; or if they described the ecology and epidemiology of zoonoses with one or more wild hosts and potential transmission to humans through the hunting, consumption, preparation or sharing of wildlife in North American Arctic and boreal biomes; details on these relevant aspects were recorded in a table.
Ninety five publications were identified for inclusion in the review. The lead author read each full article and extracted relevant information for inclusion in the manuscript. The other authors reviewed this selection and recommended additional papers, where available, that met inclusion criteria described above. All authors then considered together how the review findings combine with risk factors for zoonotic disease emergence, amplification and pandemic spread, and key cultural, socio-economic, political and ecological factors in Arctic and boreal regions, to inform on the risks of zoonotic disease emergence and amplification from Northern wildlife use and trade, and how such risks are being and could be better addressed.
Results
Our review identified 25 zoonotic bacterial (n = 12), parasitic (n = 9), and viral (n = 4) diseases described across Arctic and boreal regions of Canada and Alaska that can be transmitted to humans through the hunting, consumption, preparation, or other use of wildlife (22, 56, 73, 75–84, 86–95, 97, 99, 102–110, 112–117, 120–125, 129–134, 138–142, 145, 149–154, 157–170, 177–193). Table 2 summarizes the diseases, their causative agents, wild hosts, modes of transmission, and potential for local, epidemic or pandemic spread, with additional details given in the Appendix 1. Potential wild species hosts of these 25 zoonoses include many subsistence species important to northern Indigenous Peoples and local communities. These come from various taxa, including carnivores, ungulates, rodents, birds, marine mammals, and fish. In contrast to the global overrepresentation of viruses as emerging and pandemic human pathogens (21, 194–196), viruses comprise the lowest proportion of northern zoonoses. Of the 25 zoonotic diseases reported, 12 were identified as having potential to cause local outbreak clusters through the sharing of contaminated wild animal meat, or other products for consumption (anisakidosis, trematodosis, tapeworms, toxoplasmosis, trichinellosis, anthrax, botulism, brucellosis, mycobacteriosis including tuberculosis, Q fever, tularemia, yersiniosis); 8 have potential for human to human transmission, [anthrax, brucellosis (via breastmilk), influenzas, Q fever, tuberculosis/mycobacterial disease, yersiniosis, caliciviruses, and rabies]; and only 2 of these [influenzas and tuberculosis/mycobacterial disease] are considered as having true epidemic or pandemic potential. Many of these pathogens can threaten human health directly as zoonoses, but also indirectly via food and economic insecurity, by causing mortality events and declines in wildlife, or reduced quality of wildlife products on which northern Indigenous Peoples and local communities depend (120, 178, 179, 193). Thus, these zoonotic diseases are of local significance, even where EID risk of more global significance is low.
A study of 36 Inuit communities for 4 zoonotic parasites, found highest levels of exposure to Toxoplasma gondii (27.2%) and trichinella (18.6%), with overall seropositivity related to age, education, and consumption of marine mammals and seafood (180). Serological surveys of hunters and trappers in Cree communities in northern Québec (Eastmain, Wemindji, and Mistissini) found 44% to be positive for at least one zoonotic pathogen, with risk correlated to hunting, fishing, and trapping activities, as well as consumption of smoked game, and domestic dog ownership (91, 181). Free-ranging domestic dogs serve not only as sentinels for disease, but also as a conduit for diseases from wildlife to people e.g., for rabies and zoonotic parasites which dogs acquire from wild meat (78, 87, 182–184). The role of dogs as potential sources or amplifying hosts for zoonotic diseases highlights the importance of integrating improved domestic animal management and healthcare alongside that for humans in northern communities (197).
While wildlife use in Arctic and boreal biomes is not currently considered a major concern in terms of being the source of an epidemic or pandemic, zoonotic diseases of wildlife remain important for northern communities in the Arctic and boreal biomes: changes in quality and quantity of wildlife, access to traditional and country foods, and potential for disease risk, can create public health, food safety, and food security concerns. Indigenous and local communities in northern and remote areas of these biomes also tend to have limited access to health care services, along with suboptimal housing, infrastructure such as potable water, and sewage treatment facilities. These underlying factors contribute to the cumulative impact of additional stressors like zoonoses on the community (198).
Discussion
Zoonotic Pathogen Spillover, Amplification, Epidemic or Pandemic Spread, and Relevant Dynamics in Northern Biomes
Spillover and amplification of novel diseases is a rare, dynamic, and complex process with multiple factors at play (25, 47, 199–203). Zoonotic disease emergence events can result in dead-end “spillover” infections in which the pathogen is unable to establish stable onward transmission in the novel (human) host (27, 204). Despite this, spillover events are thought to be under-reported, as they can occur in remote regions where people have limited access to healthcare and reporting, with a lack of surveillance and diagnostic test services, and limited infrastructure, while also living in relative isolation from other human populations (47). Novel diseases are likely often mis-diagnosed, or not diagnosed at all in mild cases that resolve without treatment (47). The risk of pathogen spillover from reservoir hosts to humans, or other animals, depends upon the intensity of infection within reservoir host populations, and human contact with the reservoir population, and is affected by a cascade of events involving multiple factors associated with hosts (reservoir, intermediate/vector, recipient), the environment and the pathogen itself (205–208). This includes release of infectious material from reservoir hosts, pathogen survival in the environment, behaviors that drive exposure of a novel host, and biologically driven susceptibility of that host (199, 209). For a spillover event to result in epidemic or pandemic spread, it may need to overcome multiple barriers (210), and the pathogen must adapt to efficiently spread between individuals of the recipient population, with genetic, physiological and immunological attributes of the recipient host, together with dose and route of exposure, affecting the susceptibility to infection (206, 211). The pathogen can then be considered as transformed from a zoonotic EID to a pathogen specific for the recipient population (2). High densities, and gregariousness/sociability of the recipient host species can then facilitate pathogen exposure and transmission. Similarly, connectivity through travel, and trade networks facilitate regional, international, and global pandemic spread of pathogens in humans and domestic animals (212, 213).
Of human pathogens with potential for widespread transmission and global dissemination i.e., pandemic potential, a disproportionate number are viruses (20, 21, 26, 194–196, 214). Viruses transmitted to humans during practices that facilitate mixing of diverse animal species have significantly higher host plasticity, and higher pandemic potential (201). Zoonotic viruses capable of infecting a more diverse range of host species have higher pandemic potential in humans than those with a narrower host specificity: they are more likely to be amplified by human-to-human transmission, and spread on a global scale (201, 215). Host traits play a role in transmission, and the proportion of zoonotic viruses per mammal species has been linked with phylogenetic relatedness to humans, host taxonomy, host biomass and density, and opportunities for human contact (194, 211, 216–219). Certain mammalian groups (bats, rodents, primates) have been suggested as more likely to host zoonotic viruses (194, 215); though recent analysis suggest bats and rodents host high numbers of zoonoses simply due to the high degree of species richness within these orders, not as a result of intrinsic or ecological differences (220). Whilst wildlife are described as the source of the majority of emerging zoonotic diseases, the term “wildlife” is defined by OIE as “all free-roaming wild animals, feral animals and captive or farmed wildlife” (2). It is valuable to differentiate spillover risk posed by free-roaming wildlife, vs. captive-managed “farmed” or traded wildlife that may have wild origins, but have either been caught and maintained, or bred in captivity. Wildlife trade chains and markets that trade and process live animals or fresh meat from wildlife frequently represent high-risk interfaces due to high wildlife-human contact rates, and large numbers of admixed species (including mixing of wild and domestic animals), with their potential to shed and share viruses for extended periods prior to on-site slaughter or onward sale (221, 222). The proportion of wild rodents testing positive for coronaviruses has been shown to significantly increase along the live animal supply chain from local traders (21%), to large markets (32%), and to restaurants (56%) (32). Environmental and physiological stressors, such as the poor transportation and holding conditions along wildlife trade chains, impact animal immune function, and can lead to increased shedding and amplification of viruses (210, 223–225).
The risk of spillover of zoonotic diseases from wildlife increases when wildlife-human, and wildlife-domestic animal-human contact rates increase (19–21, 25, 42). Ecosystem degradation, land-use change for agricultural intensification or other industrial expansion, wildlife trade, and wildlife markets, all contribute to increasing contact rates, and therefore increase risk of disease emergence, particularly when they occur in areas of high biodiversity (19–21, 25, 47, 226–241). A higher diversity of zoonotic agents is found where the diversity of host animals is higher (239, 242). Although higher animal host and pathogen species diversity is associated with an increased risk of disease emergence (243), complex host, environmental and pathogen relationships ultimately determine spillover risk (206, 244). A high level of biodiversity can serve as a source of pathogens, but can also serve as a regulating factor (termed the “dilution” effect), where by loss of habitat specialists and predators due to habitat disturbance or hunting, can allow more generalist reservoir host species, such as some rodent species, to proliferate (240, 245). Zoonotic viral richness correlates strongly with mammalian species diversity and abundance (2), and domestic species harbor, on average, 19.3 zoonotic viruses compared to an average for free-ranging wildlife of 0.23 (2). Wildlife make up <1% of the world's non-human mammal biomass, thus, whilst wildlife is of concern as a source of uncommon but significant animal-origin EIDs, land-use change for agricultural intensification to supply human food systems, particularly for livestock, is of concern as a significant driver of EID risk (19, 24, 238). Higher biodiversity tends to be observed along ecosystem edges (e.g., along roads built into pristine ecosystems, or fragmented forest edges) where differing ecological systems meet, resulting in increased contact opportunities between a wider range of different host and pathogen species, and increased potential pathways for spillover (237, 246, 247). In biodiverse areas, livestock can become intermediate or amplifier hosts in which pathogens can evolve and spill over into humans (238) and over three-quarters of livestock pathogens are capable of infecting multiple host species (219).
Wildlife Use, Spillover and Emerging Zoonotic Diseases in North American Arctic and Boreal Regions: Social, Cultural, Political, and Ecological Considerations
Arctic and boreal socio-ecological systems are currently less predisposed for novel zoonotic disease emergence, due in part to a relatively low species diversity; low population density of human and livestock; limited and less intensive land uses, such as for commercial forestry and agriculture; and a lack of live wildlife markets, which are not a part of the cultural values of northern communities (19, 25, 206, 220, 239, 248–252). In addition, cold temperatures, photoperiod extremes, and geographic isolation restrict the diversity of pathogens, and often require specialized pathogen adaptations in order to persist in these environments (250, 253) (Figure 1). Recent mapping of global hotspots of relative risk for zoonotic EID events based on demographic, environmental, and biological correlates found Arctic and boreal biomes to be within the lowest risk index (25). Low human density, and reduced connectivity among human populations, also decrease the likelihood of onward pandemic spread of any newly emerged pathogen (28, 213).
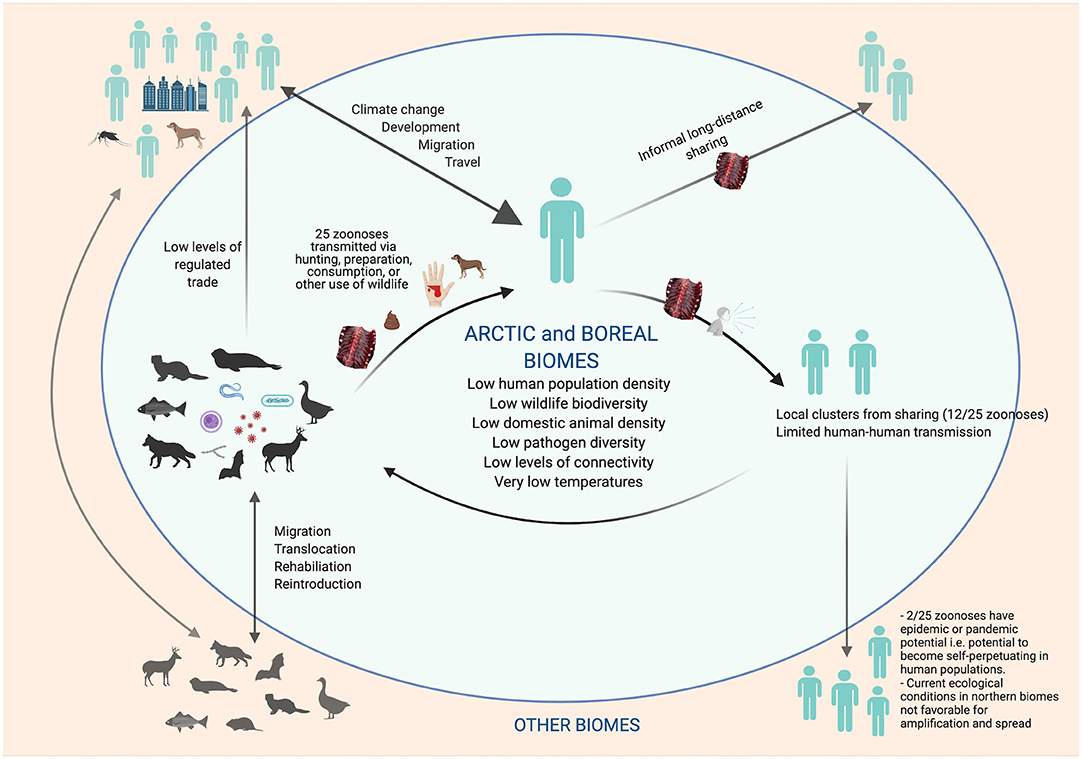
Figure 1. Transmission of Zoonoses of Wildlife Origin in Arctic and Boreal Regions: Characteristics That Lower Risk for Novel Pathogen Emergence and Pandemic Spread (Created with BioRender.com).
Commercialization of wildlife for food has been, and continues to be promoted in northern communities and regions. The type, and context, of sharing and trade of wildlife for food in northern communities is entirely different in character from the commercial trade in wildlife for human consumption, including wildlife markets, in parts of Asia and Africa. The trade is also not characterized by the same mixing of species carrying potential pandemic zoonotic pathogens as commonly occurs in wildlife trade chains throughout tropical regions. As an example, a Nunavut Wildlife Harvest Study found only 86 records (from over 145,000, and after removing the separately-monitored commercial harvest of muskoxen (Ovibos moschatus) and sales to fish or meat plants) indicating a harvest was sold commercially, vs. for personal use (254). Larger-scale commercial trade of wildlife from Arctic and boreal biomes is highly regulated, and falls under similar food safety inspection as meat from domestic animals, which not only lowers the risk of zoonotic disease transmission, but also protects the food and economic security of Indigenous and local communities who rely on wildlife (255–257). Whilst commercial trade still forms only a small component of wildlife use, the dual roles of wildlife harvesting in northern subsistence and mixed economies, including sport hunting, remain important (258–260). Maintaining and strengthening current surveillance systems for wildlife health and zoonoses, based on populations with the highest contact rates with wildlife, is an important component of an improved health infrastructure across the North. These systems can help detect, define, and control local human emergence of zoonoses while still geographically confined.
Risk for emergence of zoonotic-origin pandemic pathogens from wildlife use in Arctic and boreal biomes is currently minimal, indicating that new policies restricting these traditional and subsistence activities in the name of pandemic prevention would be unfounded at the current time.
Our review did find that endemic zoonotic concerns persist in northern communities (Table 2, Appendix 1), and increased risk is associated with consumption of raw meat, the practice of meat and fish fermentation (e.g., igunaq, muktuk), and exposure to the bodily fluids of animals through methods of harvesting and butchering (56, 118). For many Indigenous cultures around the Arctic, wildlife as food, and food sharing, are fundamental components of a cultural value system that emphasizes generosity, reciprocity, and cooperation, and usually operates within networks related to kinship and family social groups within the community (261–267). Alaska prohibits the sale of most wildlife hunted by Alaska Natives to non-Alaska Natives, but, across Canada, different jurisdictions recognize different Indigenous rights and responsibilities with respect to disposition of country food. For example, in Nunavut, an Inuk has the right to dispose freely to any person any wildlife lawfully harvested (268). Food exchanges are important because of the nutritional value of the items exchanged, and also because they carry cultural and economic values (53, 269, 270). Local sharing of foods can facilitate community zoonotic disease outbreaks (e.g., trichinella or botulism, as noted in Table 2); however, the limited potential for human-to-human transmission of current pathogens spread via sharing, and the low human population density, makes the likelihood of epidemics and pandemics originating from such activities very low. While this traditional way of sharing remains, there also exist other broader networks of sharing practices established to support urban hubs such as Anchorage, and others, across the United States (265, 266). Although low levels of consumption and sharing of traditional foods by Indigenous Peoples in urban centers in Canada have been previously reported (64, 271), relatively new sharing networks have emerged as a result of social media (272, 273) that facilitate the broader distribution of country food, often at the expense of local and traditional sharing practices and values (274), and potentially facilitating the spread of pathogens across larger geographic regions. Zoonotic disease risk is lowered through tight regulation of legal trade of wildlife and wildlife products from northern regions, however international spread of a zoonosis has been associated with sport hunting and the illegal export of wild meat (275).
Despite the perceived current low risk for zoonotic EID events, Arctic and boreal biomes are undergoing rapid ecological change. As the earth warms, and permafrost thaws, concerns have been raised for the potential release of dormant pathogens and contaminants that could affect wildlife and humans. Climate change, migration, introduction of industrial land-use, alongside increasing connectivity of people through development of tourism, travel and trade networks, will alter pathogen dynamics and could facilitate the future spread of emerging pathogens both to and from Arctic and boreal biomes (28, 213, 276, 277). These trends are already reflected in range expansion of existing pathogens (278, 279), and could promote reemergence of past diseases, novel host-pathogen relationships, and the emergence of new zoonotic diseases including vector-borne diseases (121, 185, 277, 280–282). Novel pathogens have recently emerged in northern regions: A second case of Alaskapox virus was reported in Fairbanks in October, 2020, with suspected, but as yet unknown, zoonotic origin (283), and Erysipelothrix has emerged as an apparently new disease causing agent in muskoxen in Canada, with a genotype distinct from strains found in other regions (121).
The greatest risk for pathogens of pandemic potential being introduced into northern communities is currently from people coming into the region from more densely populated southern areas (e.g., via cruise ship passengers). Travel and tourism were found to be the most significant and frequent drivers of epidemic events in Europe, and during the ongoing COVID-19 pandemic (284, 285). The current COVID-19 outbreak is an excellent example of travel-related introduction into northern regions, and subsequent community spread of a suspected wild-origin EID maintained by human-human transmission (286). As a result, many remote northern communities, who experienced impacts from previous epidemics such as influenzas, smallpox, and tuberculosis (287), have imposed strict travel restrictions and prohibited outsiders from entering their communities during the current pandemic (288). Northern residents traveling to and from regional hubs for medical procedures and other essential activities should be considered in future risk scenario planning, particularly during a pandemic. Severe Acute Respiratory Syndrome (SARS) like coronaviruses have not been identified in any wild animals in Arctic and boreal biomes. Natural and experimental infection indicates that SARS-CoV-2 appears to have the ability to infect a broad range of distantly related mammals (289). Analysis of Angiotensin-converting enzyme 2 (ACE2) receptors, suggests that some cetacean species are hypothetically susceptible, including beluga (Delphinapterus leucas) and narwhal (Monodon monoceros) (261) which are hunted by northern communities. However, entry of a virus into a cell is a complex biochemical process, with multiple other factors at play beyond the binding to a receptor, and actual infection of marine mammals is undocumented. Marine mammals are not deemed a risk based on current knowledge. Other coronaviruses have previously been identified in marine mammals, but there has been no evidence of zoonotic transmission from these animals to humans (290–293).
Reverse zoonotic transmission of human pathogens to non-human animals (anthropozoonoses), including wildlife, occurs more frequently than previously thought (294), and this directional flow of infection for SARS-CoV-2 from humans to wildlife is currently more likely than the reverse in Arctic and boreal biomes. A recent assessment found a non-negligible risk of transmission of SARS-CoV-2 from humans to bats (295), and, whilst the COVID-19 pandemic is ongoing, suspension of field work that involves direct interaction with bats has been recommended (296). Preventing human-to-wildlife SARS-CoV-2 transmission is important for protecting animals from disease, but also to avoid establishment of reservoirs in wild animals (e.g., bats), complicating disease control efforts, with potential for future spillover to other wildlife (e.g., Mustelidae) and spillback to humans (297, 298). Both anthropozoonotic, and zoonotic transmission of SARS-CoV-2 has been reported on mink (Mustela vison) farms across Europe, and the United States (299, 300). Mustelids are highly susceptible to SARS-CoV-2, an important consideration given that trapping of marten (Martes americana) and wolverine (Gulo gulo) forms a critically important industry in many northern communities (301). Contaminated wastewater and refuse from human settlements, commercial vessels, and cruise ships could theoretically pose a risk of transmission of SARS-CoV-2 from humans to marine mammals in the Arctic (261, 302). However, it is unclear if the virus remains viable under these varying environmental conditions and risk likely remains low (303), and is further attenuated by the current ban on cruise ships in the Artic.
Future Zoonotic Disease Concerns
The circumpolar North is uniquely vulnerable to the health impacts of climate change, including, but not limited to, alterations in the distribution and ecology of infectious diseases, expansion of zoonotic disease vectors, changing migration patterns, impacts on food security, limited resources of northern communities to respond to medical emergencies, and changes in water availability and quality (65, 227).
Climate change can impact distribution, life cycle, and physiological status of hosts, pathogens, and vectors, and can drive novel cross-species viral transmission (276, 304, 305). For example, a marked increase in leptospirosis was observed in Ontario following the warmest and third wettest autumn in a decade (186). Leptospirosis has not yet been recorded in humans in the Arctic, but serosurveys of Alaskan wildlife found antibodies to Leptospira serovars in caribou (Rangifer tarandus), moose (Alces alces), and bears (Ursidae) (186). The cumulative impacts of environmental and climatic changes may be increasing the susceptibility to the bacteria Erysipelothrix rhusiopathiae in certain wild animal host populations, including multiple unusual mortality events of muskoxen in Nunavut, the Northwest Territories, and Alaska, and of moose and caribou in British Columbia (120–122). In addition to the health risk for people who interact with these animals or their environments, there is potential for reduced food security for northern communities, through the direct loss of animals from E. rhusiopathiae (56, 120, 123). Climate change is already and will lead to further changes in wildlife community dynamics, including range shifts and increasing overlap between marine and terrestrial ecosystems as a result of sea ice loss and other climate-related changes. Yersinia pestis, a bacterium that persists in rodent-flea communities and causes plague in humans, is not yet reported in the Arctic, though it has been identified twice from wildlife in Canada [in bushy-tailed woodrats (Neotoma cinerea) and a Prairie dog (genus Cynomys)] (149), and appears to occur enzootically in southern Alberta, Saskatchewan, and British Columbia (150). If warming leads to northern expansion of rodent reservoirs, the disease could affect Arctic and more boreal communities in the future. Northward expansion in mammal ranges are already noted, for example in beavers (Castor canadensis) increasingly colonizing the north. Such expansion impacts zoonotic disease ecology, with beavers, for e.g., capable of carrying tularemia, and amplifying and maintaining Giardia (249). Migratory terrestrial and marine intermediate hosts have been implicated in the introduction of other zoonoses to northern regions. For example, human seroprevalence for the protozoan parasite Toxoplasma gondii is high in some parts of the Canadian Arctic, with infection associated with consumption of under-cooked country foods (306). The parasite, thought originally to be of South American origin, but now ubiquitous around the globe, has felids as the only known definitive host. However, seropositive wildlife species have been detected in the Arctic, where wild felids do not occur, and hypotheses for arrival in the region include spread via migratory waterfowl or marine mammal intermediate hosts (306, 307). As environmental conditions change, bacterial infections routinely reported from direct contact with, or ingestion of, fish in more southern regions may also become a greater concern for northern communities who depend on fish for subsistence and as part of their food sovereignty (126). While there are currently no known zoonotic viruses of fish origin, there are many viruses among fish that share evolutionary history with modern human viral pathogens, and viruses have been shown to readily jump between species in the aquatic environment (308). The possibility of some diseases decreasing in relevance warrants consideration when assessing future potential risks from climate change (187, 253, 309).
Thawing of permafrost has already contributed to changing patterns of traditional food consumption, and forced some northern communities to abandon the use of traditional ice cellars and increasingly utilize preservation methods such as smoking, pickling, and salting, and other ways to store traditional and country foods (281).
If pathogens emerge from thawing permafrost, ice patches, glaciers, or graves, wildlife may be the first to be affected. Wildlife may be both important sentinels and amplifying hosts, and a food safety concern (281). In 2016 in the Russian Arctic, one fatality from anthrax occurred among native reindeer herders, and several were hospitalized. One proposed explanation for the outbreak was that warm temperatures melted permafrost, exposing the corpses of reindeer that had died of anthrax almost a 100 years earlier, and releasing infectious Bacillus anthracis spores into nearby waterways. However, another valid explanation is the large increase in reindeer herd sizes, discontinuation of routine anthrax vaccination of reindeer (110), loss of Indigenous Knowledge among herders, and lack of veterinary experience to recognize signs of the disease in reindeer (111). Certain zoonotic disease agents survive particularly well in cold northern climates, including spore-forming bacteria, Mycobacterium species, protozoan cysts/oocysts, some helminth eggs, prions, non-enveloped viruses, and pox viruses (281). Some fungi can survive in permafrost for extended periods, and nematodes can be viable after long-term cryobiosis in Arctic permafrost (310). Most viruses are rapidly inactivated outside host cells: while RNA from the 1918 influenza strain was detected from buried Arctic peoples almost 100 years after their death (18), the material was non-infectious. Giant viruses appear to be more resilient: a giant virus trapped for around 30,000 years was recently isolated from Siberian permafrost (311), and two plant viruses recovered from 700-yr-old caribou feces still demonstrated infectivity (312). In addition to revealing dormant pathogens, climate warming also leads to release of persistent environmental pollutants, heavy metals, carbon and other natural elements from soils and rocks (313). These have cumulative effects on the immune systems and health of wildlife and humans, increasing susceptibility to diseases, particularly when combined with additional climate-related stressors such as reduced or altered habitat and food availability for wildlife, and, in turn, for humans (186).
In terms of pathogens of epidemic or pandemic potential, climate-driven changes that influence wild Arctic water bird habitat use, distribution, and migration could be a factor in the global distribution of avian viral agents, and possibly the emergence of a new pandemic influenza strain (186). Several recent studies provide evidence that migratory birds serve as effective long-distance vectors of wildlife and zoonotic pathogens to the Arctic (282), with boreal regions providing stop-over sites for many migratory species (314–316).
Most vector-borne disease agents have limited pandemic potential, as their spread tends to be restrained by geographically and climatologically restricted vector habitats (317). However, with warming environmental temperatures, and changing precipitation levels, vector-borne pathogens will become of increasing concern for epidemics in Arctic and boreal biomes, as vector ranges expand. Such viruses include those spread through mosquito bites like West Nile virus and Sindbis virus, which are maintained in wild birds; California serogroup viruses, such as Jamestown Canyon virus (antibodies to which have been found in bison (Bison bison), Dall's sheep (Ovis dalli), snowshoe hare (Lepus americanus), and Arctic fox (Vulpes lagopus) in Alaska); and Snowshoe hare virus which is maintained in mammalian hosts including rodents, deer, and hares (186, 276). Tick borne-encephalitis virus can also infect multiple hosts including ungulates, birds, rodents, and carnivores, and is spread from animals to humans through tick bites. In the Russian Arctic, climate associated increases in mosquito and tick populations, and rodent-borne infections transmitted by arthropods are already having an impact on population health (281). Insect repellent and avoidance of tick bites will likely become increasingly important for people living in northern communities.
As existing agricultural regions are threatened by climate change, warming of high latitude regions, and increasing food demands may lead to northward expansion of global agriculture (318). Agriculture is associated with the emergence of more than 50% of zoonotic infectious diseases in humans (319), and future rates of zoonotic disease emergence and/or reemergence are predicted to be closely linked to the evolution of the agriculture–environment nexus (42, 202, 238, 320). As opportunities for agriculture, and other industries, increase with warming of Arctic and boreal biomes, there is potential for disruption and dramatic change of these ecosystems from an influx of people, domestic livestock, and pets, and the disease agents they harbor (226, 243, 321). Beyond prospective increases in infectious disease emergence, climate-driven agricultural expansion will have major impacts on biodiversity, on downstream water resources, and on carbon storage (322). Multiple links exist between human health and anthropogenic environmental degradation. Land-use modifications that lead to the loss of ecosystem integrity, and increased human-domestic animal-wildlife contact rates can increase susceptibility to emerging zoonoses directly, but also indirectly via impacts on the immune system, mental health issues, environmental contaminants, and endemic diseases, paired with reduced access to fundamental services such as timber, freshwater, wild foods, medicines, and decreased air quality (248). A systematic, interdisciplinary, and holistic approach to understand and address environmental health, zoonotic, and other disease concerns around agricultural and other developments are key to mitigating negative impacts (323–325).
Current and Future Considerations for Monitoring, Surveillance, and Risk Reduction Approaches
Given the changing socio-ecological, -economic, and -political systems of Arctic and boreal biomes, future research would be beneficial to understand how the increasing anthropogenic impacts across northern regions are altering ecological processes, and thereby potentially converting microbial hazards in naturally occurring pathogen diversity into risks to human health (277, 326, 327).
Human health monitoring for zoonotic diseases in Arctic and boreal biomes is typically coordinated through relevant local health authorities and health surveys (e.g., Inuit Health Survey). Outbreaks can lead to new sampling programs, such as the sampling of walrus (Odobenus rosmarus) meat before sharing and distribution in the community (189). In general, there has been minimal research on the risks associated with traditional and country food preparation practices, despite awareness of zoonotic pathogens found in harvested wildlife and a clear need for risk reduction (118). Good hygiene during butchering and skinning, and thorough cooking of food can prevent transmission of many of the endemic zoonotic pathogens reported (118). Most health status measures and outcomes are consistently poorer for Indigenous Peoples in comparison to the rest of the North American population (63, 328) and investing in health infrastructure across Arctic and boreal biomes is critical to meet the broader needs of these populations. These investments would enhance emerging infectious disease surveillance, contribute toward a better understanding of the patterns of exposure and immunity, and reduce risks from both endemic and emerging zoonoses (79, 248, 329). Ensuring ownership of health initiatives by local communities in general, and specifically in the necessary prioritization process, is essential, with ample evidence demonstrating that failure to engage and build trust with local political and thought leaders, Elders, traditional health workers, and community groups in disease detection and control, will delay both diagnosis and response for emerging diseases (330). Identifying priority pathogens is a valuable starting point given present-day perceived financial constraints (281), though broader monitoring systems capturing syndromic trends may be more sensitive to detect emerging pathogens. Furthermore, every community has location-specific, individual risk profiles that will help determine the discrete approaches needed. To provide timely, accurate, and pertinent information on zoonotic disease risks across northern regions, the current capacity for northern communities to establish or continue disease monitoring and diagnosis needs to be expanded (186, 193, 331).
National, regional, state and territorial level veterinarians and programs monitor the health of wildlife across North American Arctic and boreal biomes. These programs range from highly standardized to less formal efforts, and vary in their objectives, from basic data collection, to in-depth species specific research. Recommendations regarding the reduction of disease risk from wildlife are detailed in a variety of local guidance publications (see Table 3). Furthermore, many groups publish regular or as-needed health updates through websites, co-management meetings, and scientific gatherings. The Canadian Wildlife Health Cooperative (332) forms a cross-Canada network of collaborators dedicated to wildlife health and a national surveillance program for wildlife diseases, but the approach to date has been essentially reactive to emerging issues. A new “Pan-Canadian Approach to Wildlife Health” was initiated in 2018 (333), and represents a deliberate transition to proactive health promotion; however, increased funding commitments, and jurisdictional collaboration and coordination, are necessary to enable its execution. In Alaska, the U.S. Geological Survey's National Wildlife Health Center (NWHC) (334) supports wildlife disease detection, control, and prevention, and conducts wildlife disease outbreak investigations as part of a national, general surveillance program. The Alaska Department of Fish and Game has a longstanding wildlife disease monitoring program and wildlife health monitoring also occurs at local levels: the North Slope Borough Department of Wildlife Management has multiple harvest monitoring programs for subsistence species [e.g., bowhead whale (Balaena mysticetus), beluga, ice seals (Phocidae), walrus, polar bears (Ursus maritimus)], and has a well-developed wildlife health research program for these mammals. This program, and an increasing number of other programs in northern regions, are community-based, and work with hunters and communities to answer questions about health and diseases in wildlife in a changing Arctic environment. Some examples of current monitoring institutions and efforts regarding health of wildlife and traditional foods are shown in Table 4.
Table 3. Examples of Existing Guidance on Safety for Hunters.
Table 4. Examples of current health monitoring institutions and efforts for health of wildlife and traditional foods (not intended to be comprehensive).
Community-based monitoring, Indigenous Knowledge, participatory epidemiology, and citizen science have become increasingly relevant as ways of centering research on community needs and priorities, and obtaining invaluable information on health and the environment (190, 193, 197, 201, 331, 335–338). Rich knowledge, including on preventing certain food-borne illness, results from the long-standing relationships of Indigenous Peoples and local communities with their environment, including their harvesting of nutritionally and spiritually important native plants, fish, and wildlife (56, 57, 118, 339, 340). Indigenous knowledge; the prevention, monitoring, and surveillance of zoonotic agents; and education are considered the most important methods to reduce human health risks associated with the consumption of traditional and country foods (118). Many wildlife harvesters in Arctic and boreal biomes are keenly aware of concerns regarding wildlife health and food safety. In general, parasites, lesions (discoloration, tracts, fungal growths, etc.), or abnormal behavior of hunted animals are often observed by Indigenous and local communities. Indigenous Peoples were some of the first to draw attention to traditional and country food safety concerns related to environmental contaminants, because of changes they detected in quality of the animals and fish they hunted (118). Community-based monitoring programs contribute valuable information. For example, the Nunavik Trichinellosis Prevention Program provides rapid carcass-side testing for trichinella in country foods for Nunavik communities (341), giving community members control to obtain the information they need to make informed decisions about food preparation, consumption, and carcass disposal to prevent further transmission. Recent working groups assessing Unusual Mortality Events (UMEs) for marine mammals and other species around Alaska include Indigenous perspectives and observations (191, 342) and, across Canada, several Indigenous-led Guardians Programs monitor ecological health, maintain cultural sites, and protect sensitive areas and species, while also playing a vital role in creating conservation plans, and supporting Indigenous Protected and Conserved Areas (343). Table 3 lists some examples of the existing monitoring efforts for wildlife health, and country and traditional foods. Ongoing academic-government-community collaborative partnerships have resulted in several successful community-based wildlife health surveillance programs that aim to address community concerns about wildlife health and food safety while simultaneously addressing key research questions about wildlife ecology and dynamics (193, 197, 338).
Inuit, Métis, First Nations, and Alaskan Native societies across the Arctic and boreal biomes (and globally) maintain vast and holistic Indigenous Knowledge Systems, across generations, about the natural environment and how it is changing. Such awareness and presence of eyes-in-the-field is irreplaceable for the early detection of changes in wildlife populations, and the environment. Combining Indigenous Knowledge with scientific understanding improves wildlife surveillance, fosters reconciliation, and advances Indigenous Peoples' self-determination in research, while creating mutual health and conservation benefits (331, 344–348). However, a history of colonialism, relocations, residential schools, and loss of Indigenous languages has led to generational changes in diet, life-style and relationships with the environment for many Indigenous communities. Loss of transgenerational “hands-on” and oral transmission of knowledge, especially regarding harvesting, butchering, food safety and zoonoses, may increase zoonotic risk, increase meat wastage, and limit the ability to engage with “two-eyed-seeing” within participatory epidemiology (331). In addition to fostering reconciliation with Indigenous Peoples, programs that assist Elders to pass on Indigenous Knowledge to Indigenous youth, and brings communities together to prepare and share country and traditional foods, contributes to future monitoring of wildlife, human and environmental health, and supports a conservation economy focused on the land and wildlife. The participation and leadership of men and women involved in the harvest and preparation of wild foods, and youth in wildlife and community health monitoring and research, builds trust between researchers and local communities, particularly when focused on community-raised concerns and priorities (197, 331). Continued investment in existing community-based wildlife and human health monitoring efforts, paired with consistent and well-conveyed methodologies, would be a valuable approach to tracking wild harvest use and trade, zoonotic and other emerging pathogens, host species, and environmental impacts from the changing climate across northern communities.
Early recognition and intervention during an emerging infectious zoonotic disease event is essential to limit spread (349). Wild animals, and domestic dogs, can serve as sentinels for zoonotic diseases and other health concerns such as contaminants and it appears essential that existing veterinary and wildlife surveillance systems for zoonotic pathogens are closely integrated with public health surveillance, to better control such pathogens before they affect human health. Surveillance would benefit from the exploration and cooperative development with local and Indigenous leaders of practical ways to integrate and share surveillance-generated information for humans, domestic animals and wildlife. This would ensure that the surveillance systems are risk-based, spatially and temporally targeting species and geographic locations with the highest risk for spillover, and guaranteeing that the information gathered can be used by the local communities to deploy interventions in a timely manner. Expansion of regional, transdisciplinary One Health networks that involve community members and diverse stakeholders and collate real-time multifactorial data and explicit observations, (e.g., the Echo Network for health and conservation, the Circumpolar Climate Change and Infectious Diseases Workgroup, or the LEO network1) (350) and integration with public health networks such as the International Circumpolar Surveillance (ICS) project (192), can facilitate early-warning systems, rapid response, and mitigation for health threats to animals, humans, and the environment, while also advancing our understanding of endemic, and emerging zoonotic health risks for these communities.
It is important that the desire to increase awareness of the potential risk of zoonotic infections be balanced with the unintended consequences on other determinants of health. Past experience with poorly designed communication strategies and materials with respect to country and traditional foods, such as contaminants in wild-caught foods, provides stark lessons on the need to avoid provoking unnecessary fear when addressing food safety risk in Indigenous and local communities (71, 72). For example, scientific communication to Nunavut Inuit communities in the late 1980s that women's breast milk had high levels of polychlorinated biphenyls (PCBs), failed to consider broader understanding of contamination issues in the region, and failed to include the Inuit directly on messaging (72). The resulting alarm and confusion across Inuit and other northern communities led many women to stop breastfeeding, and stopped consumption of country and traditional foods, leading to other more insidious health issues, poor nutrition, and food insecurity (72). Similar issues were associated with the research and communication of contaminants in wildlife in Alaska. Participation in subsistence activities, including the practice of hunting by Indigenous Peoples, is considered a protective mental health factor for Indigenous circumpolar youth (351), and there are different and pertinent health concerns associated with the consumption of store-bought, processed, imported market foods increasingly eaten by younger generations. Such foods are often low in nutritional value, high in sugar, and sometimes harbor contaminants including microplastics and pesticides (56, 352–354). In Alaska, the introduction of modern materials such as plastic bags, plastic pails and glass jars in the preparation of rendered and aged foods (muktuk and igunaq) actually increased outbreaks of botulism (118). Indigenous and non-Indigenous health professionals promoted the return to traditional methods thereby reducing fatality rates by leveraging knowledge from community members, Elders, and survivors of botulism. Communications and management approaches around zoonotic disease risk from traditional and country foods are better co-developed with Indigenous and other hunting/trapping communities, with mindfulness that hunters and their families are keenly aware of changes in the behavior of wildlife or condition of wildlife products; these foods are usually the most cost-effective and healthy sources of nutrition in northern and remote communities; and country and traditional foods have important cultural, spiritual, and social values to northern communities (355, 356).
Combining diverse approaches and ways of knowing, including Indigenous Knowledge, provides a more complete understanding of the socio-ecological system than applying Western science alone. Working together can help bridge gaps in scientific monitoring, and bring the best available knowledge to more effectively monitor, and respond to, the impacts of disease and climate change on the health of Arctic and boreal inhabitants (338).
Conclusion
An essential connection exists between subsistence activities, and well-being and resilience within Indigenous Peoples and local communities across the Arctic and boreal biomes in North America. The current threat of pandemic zoonotic disease emergence from hunting, consumption, and use of wildlife in the North American Arctic and boreal biomes is low, and policies restricting these traditional and subsistence activities in the name of pandemic prevention would be greatly misplaced. Health threats from endemic zoonotic diseases in northern biomes remain, with plausible hypotheses that environmental alterations from development, and climate change could alter epidemic and pandemic risks. Whilst commercialization of trade in wild food and products, and agricultural expansion have been identified as factors important in developing resilience in the North (351, 356, 357), they will bring a different set of challenges around zoonotic disease emergence, and disruption of ecosystems (238, 323). Co-development of any future policies and interventions with Indigenous Peoples and local communities may more effectively address zoonotic disease emergence in these rapidly changing northern regions. Combining Indigenous Knowledge Systems with science provides a more holistic understanding of zoonotic risks in Arctic and boreal ecosystems, informs opportunities for mitigation and monitoring, and improves disease risk communications, while avoiding potentially negative repercussions (331, 356, 358, 359). Monitoring, research, and response efforts would all benefit from employing more inclusive One Health approaches that draw on all knowledge systems and types of expertise, and proactively incorporate the complexity and interrelatedness of the environmental, biological, economic, political, cultural, and social dimensions of zoonotic disease emergence in northern biomes.
Author Contributions
LK, CW, MR and JR contributed to conception and design of the study. LK organized the database, performed the review and analysis and wrote the first draft of the manuscript. MR and C-LC also wrote sections of the manuscript. All authors contributed to manuscript revision and additions, further development of the research concept and read, and approved the submitted version.
Conflict of Interest
DJ was employed by the company Nyati Health Consulting, British Columbia, Canada. DL was employed by Nunavut Tunngavik Inc., Ottawa, Canada.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer AP declared a past co-authorship with one of the authors SO to the handling Editor.
Acknowledgments
The authors would like to acknowledge and thank Ms. Jean Allen, and Ms. Sharon Edmunds of Nunavut Tunngavik Inc., and Robert Suydam of North Slope Department of Wildlife Management, for their valuable input to the manuscript. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpubh.2021.627654/full#supplementary-material
Footnotes
1. ^The LEO Network collates and shares information on unusual events, climate and other drivers of environmental change. The online platform is multi-lingual, including Indigenous languages, and was designed to be accessible from remote locations, in order to gather input from local community members, using holistic observational data, and welcoming diverse knowledge systems. The network facilitates connectivity between communities, environmental and public health experts, and allows researchers and agencies to use data as a surveillance system for local updates on emerging issues and vulnerabilities, and to provide event specific consultations.
References
2. Haider N, Rothman-Ostrow P, Osman AY, Arruda LB, Macfarlane-Berry L, Elton L, et al. COVID-19—Zoonosis or emerging infectious disease? Front Public Health. (2020) 8:596944. doi: 10.3389/fpubh.2020.596944
3. Li HY, Zhu GJ, Zhang YZ, Zhang LB, Hagan EA, Martinez S, et al. A qualitative study of zoonotic risk factors among rural communities in southern China. Int Health. (2020) 12:77–85. doi: 10.1093/inthealth/ihaa001
4. López-Robles G, Montalvo-Corral M, Caire-Juvera G, Ayora-Talavera G, Hernández J. Seroprevalence and risk factors for swine influenza zoonotic transmission in swine workers from Northwestern Mexico. Transbound Emerg Dis. (2012) 59:183–8. doi: 10.1111/j.1865-1682.2011.01250.x
5. Morse SS. Factors in the emergence of infectious diseases. In: Price-Smith AT, editor. Plagues Politics. London: Palgrave Macmillan (2001). p. 8–26. doi: 10.1057/9780230524248_2
6. Taylor LH, Latham SM, Woolhouse MEJ. Risk factors for human disease emergence. Philos Trans R Soc London Ser B Biol Sci. (2001) 356:983–9. doi: 10.1098/rstb.2001.0888
7. Woolhouse MEJ, Haydon DT, Antia R. Emerging pathogens: the epidemiology and evolution of species jumps. Trends Ecol Evol. (2005) 20:238–44. doi: 10.1016/j.tree.2005.02.009
8. Loh EH, Zambrana-Torrelio C, Olival KJ, Bogich TL, Johnson CK, Mazet JAK, et al. Targeting transmission pathways for emerging zoonotic disease surveillance and control. Vector Borne Zoonotic Dis. (2015) 15:432–7. doi: 10.1089/vbz.2013.1563
9. Hillson D, Hulett D. Assessing Risk Probability: Impact Alternative Approaches. Prague: PMI® Global Congress 2004—EMEA (2004). Available online at: https://www.pmi.org/learning/library/assessing-risk-probability-impact-alternative-approaches-8444 (accessed March 10, 2021).
10. Zhang L, Shen F, Chen F, Lin Z. Origin and evolution of the 2019 novel coronavirus. Clin Infect Dis. (2020) 71:882–3. doi: 10.1093/cid/ciaa112
11. Zhang X, Chen X, Zhang Z, Roy A, Shen Y. Strategies to trace back the origin of COVID-19. J Infect. (2020) 80:e39–40. doi: 10.1016/j.jinf.2020.03.032
12. Andersen KG, Rambaut A, Lipkin WI, Holmes EC, Garry RF. The proximal origin of SARS-CoV-2. Nat Med. (2020) 26:450–2. doi: 10.1038/s41591-020-0820-9
13. MacLean OA, Lytras S, Weaver S, Singer JB, Boni MF, Lemey P, et al. Natural selection in the evolution of SARS-CoV-2 in bats created a generalist virus and highly capable human pathogen. PLOS Biol. (2021) 19:e3001115. doi: 10.1371/journal.pbio.3001115
14. Lau SKP, Woo PCY, Li KSM, Huang Y, Tsoi HW, Wong BHL, et al. Severe acute respiratory syndrome coronavirus-like virus in Chinese horseshoe bats. Proc Natl Acad Sci USA. (2005) 102:14040–5. doi: 10.1073/pnas.0506735102
15. Wang M, Yan M, Xu H, Liang W, Kan B, Zheng B, et al. SARS-CoV infection in a restaurant from palm civet. Emerg Infect Dis. (2005) 11:1860–5. doi: 10.3201/eid1112.041293
16. Wang LF, Eaton BT. Bats, civets and the emergence of SIRS. Curr Topics Microbiol Immunol. (2007) 315:325–344. doi: 10.1007/978-3-540-70962-6_13
17. Song H-D, Tu C-C, Zhang G-W, Wang S-Y, Zheng K, Lei L-C, et al. Cross-host evolution of severe acute respiratory syndrome coronavirus in palm civet and human. Proc Natl Acad Sic USA. (2005) 102:2430–5. doi: 10.1073/pnas.0409608102
18. Taubenberger JK, Hultin JV, Morens DM. Discovery and characterization of the 1918 pandemic influenza virus in historical context. Antivir Ther. (2007) 12:581–91.
19. Jones KE, Patel NG, Levy MA, Storeygard A, Balk D, Gittleman JL, et al. Global trends in emerging infectious diseases. Nature. (2008) 451:990–3. doi: 10.1038/nature06536
20. Karesh WB, Dobson A, Lloyd-Smith JO, Lubroth J, Dixon MA, Bennett M, et al. Ecology of zoonoses: natural and unnatural histories. Lancet. (2012) 380:1936–45. doi: 10.1016/S0140-6736(12)61678-X
21. Morse SS, Mazet JAK, Woolhouse M, Parrish CR, Carroll D, Karesh WB, et al. Prediction and prevention of the next pandemic zoonosis. Lancet. (2012) 380:1956–65. doi: 10.1016/S0140-6736(12)61684-5
22. Schurer JM, Bouchard E, Bryant A, Revell S, Chavis G, Lichtenwalner A, et al. Echinococcus in wild canids in Québec (Canada) and Maine (USA). PLoS Negl Trop Dis. (2018) 12:e0006712. doi: 10.1371/journal.pntd.0006712
23. Temmam S, Desnues C. Zoonotic viruses: how to monitor their emergence? Virologie. (2016) 20:231–44. doi: 10.1684/vir.2016.0660
24. McMichael AJ. Environmental and social influences on emerging infectious diseases: past, present and future. Philos Trans R Soc London Ser B Biol Sci. (2004) 359:1049–58. doi: 10.1098/rstb.2004.1480
25. Allen T, Murray KA, Zambrana-Torrelio C, Morse SS, Rondinini C, Di Marco M, et al. Global hotspots and correlates of emerging zoonotic diseases. Nat Commun. (2017) 8:1124. doi: 10.1038/s41467-017-00923-8
26. Carroll D, Daszak P, Wolfe ND, Gao GF, Morel CM, Morzaria S, et al. The global virome project. Science. (2018) 359:872–4. doi: 10.1126/science.aap7463
27. Geoghegan JL, Senior AM, Giallonardo F Di, Holmes EC. Virological factors that increase the transmissibility of emerging human viruses. Proc Natl Acad Sci USA. (2016) 113:4170–5. doi: 10.1073/pnas.1521582113
28. Pike J, Bogich T, Elwood S, Finnoff DC, Daszak P. Economic optimization of a global strategy to address the pandemic threat. Proc Natl Acad Sci USA. (2014) 111:18519–23. doi: 10.1073/pnas.1412661112
29. Cantlay JC, Ingram DJ, Meredith AL. A review of zoonotic infection risks associated with the wild meat trade in Malaysia. Ecohealth. (2017) 14:361–88. doi: 10.1007/s10393-017-1229-x
30. Chomel BB, Belotto A, Meslin FX. Wildlife, exotic pets, and emerging zoonoses. Emerg Infect Dis. (2007) 13:6–11. doi: 10.3201/eid1301.060480
31. Travis DA, Watson RP, Tauer A. The spread of pathogens through trade in wildlife. Rev Sci Tech OIE. (2011) 30:219–39. doi: 10.20506/rst.30.1.2035
32. Huong NQ, Nga NTT, Long N Van, Luu BD, Latinne A, Pruvot M, et al. Coronavirus testing indicates transmission risk increases along wildlife supply chains for human consumption in Viet Nam, 2013-2014. PLoS ONE. (2020) 15:e0237129. doi: 10.1371/journal.pone.0237129
33. Durand B, Lecollinet S, Beck C, Martínez-López B, Balenghien T, Chevalier V. Identification of hotspots in the european union for the introduction of four zoonotic arboviroses by live animal trade. PLoS ONE. (2013) 8:e70000. doi: 10.1371/journal.pone.0070000
34. Fèvre EM, Bronsvoort BMDC, Hamilton KA, Cleaveland S. Animal movements and the spread of infectious diseases. Trends Microbiol. (2006) 14:125–31. doi: 10.1016/j.tim.2006.01.004
35. Greatorex ZF, Olson SH, Singhalath S, Silithammavong S, Khammavong K, Fine AE, et al. Wildlife trade and human health in lao PDR: an assessment of the zoonotic disease risk in markets. PLoS ONE. (2016) 11:e0150666. doi: 10.1371/journal.pone.0150666
36. Hoffmann B, Tappe D, Höper D, Herden C, Boldt A, Mawrin C, et al. A variegated squirrel bornavirus associated with fatal human encephalitis. N Engl J Med. (2015) 373:154–62. doi: 10.1056/NEJMoa1415627
37. Karesh WB, Cook RA, Bennett EL, Newcomb J. Wildlife trade and global disease emergence. Emerg Infect Dis. (2005) 11:1000–2. doi: 10.3201/eid1107.050194
38. Kurpiers LA, Schulte-Herbrüggen B, Ejotre I, Reeder DM. Bushmeat and emerging infectious diseases: lessons from Africa. Probl Wildl. (2016) 507–51. doi: 10.1007/978-3-319-22246-2_24
39. Marano N, Arguin PM, Pappaioanou M. Impact of globalization and animal trade on infectious disease ecology. Emerg Infect Dis. (2007) 13:1807–9. doi: 10.3201/eid1312.071276
40. Pavlin BI, Schloegel LM, Daszak P. Risk of importing zoonotic diseases through wildlife trade, United States. Emerg Infect Dis. (2009) 15:1721–6. doi: 10.3201/eid1511.090467
41. ProMED. ProMED-Mail Post: Subject: PRO/AH/EDR> COVID-19 Update (102): China, Origin, Farmed Wild Animals susp. (2021). Available online at: https://promedmail.org/promed-post/?id=8251684 (accessed March 17, 2021).
42. Murray KA, Allen T, Loh E, Machalaba C, Daszak P. Emerging viral zoonoses from wildlife associated with animal-based food systems: risks and opportunities. In: Jay-Russell M, Doyle M, editors. Food Safety Risks from Wildlife. Cham: Springer International Publishing (2015). p. 31–57. doi: 10.1007/978-3-319-24442-6_2
43. Dobson AP, Pimm SL, Hannah L, Kaufman L, Ahumada JA, Ando AW, et al. Ecology and economics for pandemic prevention. Science. (2020) 369:379–81. doi: 10.1126/science.abc3189
44. Walzer C. COVID-19 and the curse of piecemeal perspectives. Front Vet Sci. (2020) 7:582983. doi: 10.3389/fvets.2020.582983
45. WHO. WHO Director-General's Opening Remarks at the Media Briefing on COVID-19–17 April 2020. (2020). Available online at: https://www.who.int/dg/speeches/detail/who-director-general-s-opening-remarks-at-the-media-briefing-on-covid-19-−17-april-2020 (accessed May 26, 2020).
46. Zavaleta-Cortijo C, Ford JD, Arotoma-Rojas I, Lwasa S, Lancha-Rucoba G, García PJ, et al. Climate change and COVID-19: reinforcing indigenous food systems. Lancet Planet Health. (2020) 4:e381–2. doi: 10.1016/S2542-5196(20)30173-X
47. Johnson CK, Hitchens PL, Pandit PS, Rushmore J, Evans TS, Young CCW, et al. Global shifts in mammalian population trends reveal key predictors of virus spillover risk. Proc R Soc B Biol Sci. (2020) 287:20192736. doi: 10.1098/rspb.2019.2736
48. United Nations. United Nations Declaration on the Rights of Indigenous Peoples. New York, NY: United Nations (2007).
49. Kuhnlein HV, Receveur O, Chan HM. Traditional food systems research with Canadian indigenous peoples. Int J Circumpolar Health. (2001) 60:112–22.
50. Walch A, Bersamin A, Loring P, Johnson R, Tholl M. A scoping review of traditional food security in Alaska. Int J Circumpolar Health. (2018) 77:1419678. doi: 10.1080/22423982.2017.1419678
51. Bjerregaard P, Kue Young T, Dewailly E, Ebbesson SOE. Review article: indigenous health in the Arctic: an overview of the circumpolar inuit population. Scand J Public Health. (2004) 32:390–5. doi: 10.1080/14034940410028398
52. Loring PA, Gerlach SC. Food, culture, and human health in Alaska: an integrative health approach to food security. Environ Sci Policy. (2009) 12:466–78. doi: 10.1016/j.envsci.2008.10.006
53. Collings P, Marten MG, Pearce T, Young AG. Country food sharing networks, household structure, and implications for understanding food insecurity in Arctic Canada. Ecol Food Nutr. (2016) 55:30–49. doi: 10.1080/03670244.2015.1072812
54. McGrath-Hanna NK, Greene DM, Tavernier RJ, Bult-Ito A. Diet and mental health in the Arctic: is diet an important risk factor for mental health in circumpolar peoples?–A review. Int J Circumpolar Health. (2003) 62:228–41. doi: 10.3402/ijch.v62i3.17560
55. Reeves RR. The origins and character of “aboriginal subsistence” whaling: a global review. Mamm Rev. (2002) 32:71–106. doi: 10.1046/j.1365-2907.2002.00100.x
56. CCA. Aboriginal Food Security in northern Canada: An Assessment of the State of Knowledge. Ottawa, ON: Council of Canadian Academies (2014).
57. ICC. Alaskan Inuit Food Security Conceptual Framework: How To Assess The Arctic From An Inuit Perspective. Anchorage, AK: Inuit Circumpolar Council-Alaska (2015).
58. Food and Agriculture Organisation. An Introduction to the Basic Concepts of Food Security. Rome: Food and Agriculture Organisation (2008).
59. World Forum on Food Sovereignty. Final Declaration of the World Forum on Food Sovereignty. Havana: World Forum on Food Sovereignty (2001).
60. Government of Canada. Government of Canada Constitution Acts, 1867 to 1982: Rights of the Aboriginal Peoples of Canada. Ottawa, ON: Government of Canada (2016).
61. Truth and Reconciliation Commission. Truth and Reconciliation Commission of Canada: Calls to Action. Winnipeg, MB: Truth and Reconciliation Commission (2015).
62. Lougheed T. The changing landscape of arctic traditional food. Environ Health Perspect. (2010) 118:a386–93. doi: 10.1289/ehp.118-a386
63. Kuhnlein H, Erasmus B, Creed-Kanashiro H, Englberger L, Okeke C, Turner N, et al. Indigenous peoples' food systems for health: finding interventions that work. Public Health Nutr. (2006) 9:1013–19. doi: 10.1017/PHN2006987
64. Elliott B, Jayatilaka D, Brown C, Varley L, Corbett KK. “We are not being heard”: aboriginal perspectives on traditional foods access and food security. J Environ Public Health. (2012) 2012:1–9. doi: 10.1155/2012/130945
65. Farquhar SD. Inuit seal hunting in canada: emerging narratives in an old controversy. Arctic. (2020) 73:13–9. doi: 10.14430/arctic69833
66. Chan L, Receveur O, Batal M, Sadik T, Schwartz H, Ing A, et al. First Nations Food, Nutrition and Environment Study (FNFNES): Results from Saskatchewan (2015). Ottawa, ON: University of Ottawa (2018).
67. Goettke E, Reynolds J. “It's all interconnected… like a spider web”: a qualitative study of the meanings of food and healthy eating in an Indigenous community. Int J Circumpolar Health. (2019) 78:1648969. doi: 10.1080/22423982.2019.1648969
68. Hanemaayer R, Anderson K, Haines J, Lickers KRL, Lickers Xavier A, Gordon K, et al. Exploring the perceptions of and experiences with traditional foods among first nations female youth: a participatory photovoice study. Int J Environ Res Public Health. (2020) 17:2214. doi: 10.3390/ijerph17072214
69. Kuhnlein HV, Receveur O. Dietary change and traditional food systems of indigenous peoples. Annu Rev Nutr. (1996) 16:417–42. doi: 10.1146/annurev.nu.16.070196.002221
70. Moses SK, Whiting A V, Bratton GR, Taylor RJ, O'Hara TM. Inorganic nutrients and contaminants in subsistence species of Alaska: linking wildlife and human health. Int J Circumpolar Health. (2009) 68:53–74. doi: 10.3402/ijch.v68i1.18294
71. Furgal CM, Powell S, Myers H. Digesting the message about contaminants and country foods in the Canadian north: a review and recommendations for future research and action. Arctic. (2005) 58:103–14. doi: 10.14430/ARCTIC404
72. Krümmel EM, Gilman A, Krü EM. An update on risk communication in the Arctic. Int J Circumpolar Health. (2016) 75:33822. doi: 10.3402/ijch.v75.33822
73. Pufall EL, Jones-Bitton A, McEwen SA, Brown TM, Edge VL, Rokicki J, et al. Prevalence of zoonotic anisakid nematodes in inuit-harvested fish and mammals from the Eastern Canadian Arctic. Foodborne Pathog Dis. (2012) 9:1002–9. doi: 10.1089/fpd.2012.1186
74. Cawthorn M. Meat Consumption From Stranded Whales and Marine Mammals in New Zealand: Public Health and Other Issues (1997). Wellington: Department of Conservation.
75. Gordon JE, Frye WW, Babbott FL. Intestinal parasites of man in arctic greenland*. Am J Trop Med Hyg. (1961) 10:185–90. doi: 10.4269/ajtmh.1961.10.185
76. Chai JY, Jung BK. Fishborne zoonotic heterophyid infections: An update. Food Waterbor Parasitol. (2017) 8–9:33–63. doi: 10.1016/j.fawpar.2017.09.001
77. Schurer JM, Gesy KM, Elkin BT, Jenkins EJ. Echinococcus multilocularis and Echinococcus canadensis in wolves from western Canada. Parasitology. (2014) 141:159–63. doi: 10.1017/S0031182013001716
78. Rausch R. Cystic Echinococcosis in the Arctic and Sub-Arctic (2003). doi: 10.1017/s0031182003003664
79. Jenkins EJ, Castrodale LJ, de Rosemond SJC, Dixon BR, Elmore SA, Gesy KM, et al. Tradition and transition: parasitic zoonoses of people and animals in Alaska, northern Canada, and Greenland. Adv Parasitol. (2013) 82:33–204. doi: 10.1016/B978-0-12-407706-5.00002-2
80. Kutz SJ, Ducrocq J, Verocai GG, Hoar BM, Colwell DD, Beckmen KB, et al. Parasites in Ungulates of Arctic North America and Greenland. Adv Parasitol. (2012) 79:99–252. doi: 10.1016/B978-0-12-398457-9.00002-0
81. Lantis M. Zoonotic diseases in the Canadian and Alaskan North. Études Inuit Stud. (1981) 5:83–107.
82. Ross P, Olpinski S, Curtis M. Relationships between dietary practice and parasite zoonoses in Northern Québec Inuit communities. Études Inuit Stud. (1989) 13:33–47.
83. Schurer JM, Pawlik M, Huber A, Elkin B, Cluff HD, Pongracz JD, et al. Intestinal parasites of gray wolves (Canis lupus) in northern and western Canada. Can J Zool. (2016) 94:643–50. doi: 10.1139/cjz-2016-0017
84. Hueffer K, Parkinson AJ, Gerlach R, Berner J. Zoonotic infections in Alaska: disease prevalence, potential impact of climate change and recommended actions for earlier disease detection, research, prevention and control. Int J Circumpolar Health. (2013) 72. doi: 10.3402/ijch.v72i0.19562
85. Zhang W, Wen H, Li J, Lin R, McManus DP. Immunology and immunodiagnosis of cystic echinococcosis: an update. Clin Dev Immunol. (2012) 2012:1–10. doi: 10.1155/2012/101895
86. Kuchta R, Oros M, Ferguson J, Scholz T. Diphyllobothrium nihonkaiense tapeworm larvae in Salmon from North America. Emerg Infect Dis. (2017) 23:351–3. doi: 10.3201/eid2302.161026
87. Salb AL, Barkema HW, Elkin BT, Thompson RCA, Whiteside DP, Black SR, et al. Dogs as sources and sentinels of parasites in humans and wildlife, Northern Canada. Emerg Infect Dis. (2008) 14:60–3. doi: 10.3201/eid1401.071113
88. Bradbury RS, Panicker IS. Toxocara seroprevalence in Canada—climate, environment and culture. Adv Parasitol. (2020) 109:291–316. doi: 10.1016/bs.apar.2020.03.003
89. Wapenaar W, Barkema HW, O'Handley R. Fecal shedding of toxocara canis and other parasites in foxes and coyotes on prince Edward Island, Canada. J Wildl Dis. (2013) 49:394–7. doi: 10.7589/2012-04-113
90. Rausch RL, Fay FH. Toxascaris leonina in rodents, and relationship to eosinophilia in a human population. Comp Parasitol. (2011) 78:236–44. doi: 10.1654/4504.1
91. Lévesque B, Messier V, Bonnier-Viger Y, Couillard M, Côté S, Ward BJ, et al. Seroprevalence of zoonoses in a cree community (Canada). Diagn Microbiol Infect Dis. (2007) 59:283–6. doi: 10.1016/j.diagmicrobio.2007.06.002
92. Messier V, Lévesque B, Proulx J-F, Rochette L, Libman MD, Ward BJ, et al. Seroprevalence of toxoplasma gondii among nunavik inuit (Canada). Zoo Public Health. (2009) 56:188–97. doi: 10.1111/j.1863-2378.2008.01177.x
93. Dubey J, Lewis B, Beam K, Abbitt B. Transplacental toxoplasmosis in a reindeer (Rangifer tarandus) fetus. Vet Parasitol. (2002) 110:131–5. doi: 10.1016/S0304-4017(02)00320-5
94. Sørensen KK, Mørk T, Sigurð*ardóttir ÓG, Åsbakk K, Åkerstedt J, Bergsjø B, Fuglei E. Acute toxoplasmosis in three wild arctic foxes (Alopex lagopus) from Svalbard; one with co-infections of Salmonella enteritidis PT1 and Yersinia pseudotuberculosis serotype 2b. Res Vet Sci. (2005) 78:161–7. doi: 10.1016/j.rvsc.2004.07.010
95. Sandström CAM, Buma AGJ, Hoye BJ, Prop J, van der Jeugd H, Voslamber B, et al. Latitudinal variability in the seroprevalence of antibodies against Toxoplasma gondii in non-migrant and Arctic migratory geese. Vet Parasitol. (2013) 194:9–15. doi: 10.1016/j.vetpar.2012.12.027
96. Torrey EF, Yolken RH. Toxoplasma gondii and schizophrenia. Emerg Infect Dis. (2003) 9:1375–80. doi: 10.3201/eid0911.030143
97. Elmore SA, Huyvaert KP, Bailey LL, Milhous J, Alisauskas RT, Gajadhar AA, et al. Toxoplasma gondii exposure in arctic-nesting geese: a multi-state occupancy framework and comparison of serological assays. Int J Parasitol Parasites Wildl. (2014) 3:147–53. doi: 10.1016/j.ijppaw.2014.05.005
98. Gill CO. Microbiological conditions of meats from large game animals and birds. Meat Sci. (2007) 77:149–60. doi: 10.1016/j.meatsci.2007.03.007
99. Jenkins EJ, Simon A, Bachand N, Stephen C. Wildlife parasites in a one health world. Trends Parasitol. (2015) 31:174–80. doi: 10.1016/j.pt.2015.01.002
100. Massie GN, Ware MW, Villegas EN, Black MW. Uptake and transmission of Toxoplasma gondii oocysts by migratory, filter-feeding fish. Vet Parasitol. (2010) 169:296–303. doi: 10.1016/j.vetpar.2010.01.002
101. Chegeni TN, Sharif M, Sarvi S, Moosazadeh M, Montazeri M, Aghayan SA, et al. Is there any association between toxoplasma gondii infection and depression? A systematic review and meta-analysis. PLoS ONE. (2019) 14:e0218524. doi: 10.1371/journal.pone.0218524
102. McIntyre L, Pollock SL, Fyfe M, Gajadhar A, Isaac-Renton J, Fung J, et al. Trichinellosis from consumption of wild game meat. Can Med Assoc J. (2007) 176:449–51. doi: 10.1503/cmaj.061530
103. Seymour J, Horstmann-Dehn L, Rosa C, Lopez JA. Occurrence and genotypic analysis of Trichinella species in Alaska marine-associated mammals of the bering and chukchi seas. Vet Parasitol. (2014) 200:153–64. doi: 10.1016/j.vetpar.2013.11.015
104. Forbes LB. The occurrence and ecology of trichinella in marine mammals. Vet Parasitol. (2000) 93:321–34. doi: 10.1016/S0304-4017(00)00349-6
105. Sharma R, Thompson P, Elkin B, Mulders R, Branigan M, Pongracz J, et al. Trichinella pseudospiralis in a wolverine (Gulo gulo) from the Canadian North. Int J Parasitol Parasites Wildl. (2019) 9:274–80. doi: 10.1016/j.ijppaw.2019.06.005
106. Curtis MA, Rau ME, Tanner CE, Prichard RK, Faubert GM, Olpinski S, et al. Parasitic zoonoses in relation to fish and wildlife harvesting by inuit communities in Northern Quebec, Canada. Arctic Me Res. (1988) 47 (Suppl. 1):693–6.
107. Olson ME, Roach PD, Stabler M, Chan W. Giardiasis in ringed seals from the Western Arctic. J Wildl Dis. (1997) 33:646–8. doi: 10.7589/0090-3558-33.3.646
108. Leighton FA. Wildlife pathogens and diseases in Canada. In: Canadian Biodiversity: Ecosystem Status and Trends 2010, Technical Thematic Report No. 7. Ottawa, ON: Canadian Councils of Resource Ministers (2011). p. 53.
109. Shury TK, Frandsen D, O'Brodovich L. Anthrax in free-ranging bison in the prince albert national park area of Saskatchewan in 2008. Can Vet J. (2009) 50:152–4.
110. Hueffer K, Drown D, Romanovsky V, Hennessy T. Factors contributing to anthrax outbreaks in the circumpolar North. Ecohealth. (2020) 17:174–80. doi: 10.1007/s10393-020-01474-z
111. Timofeev V, Bahtejeva I, Mironova R, Titareva G, Lev I, Christiany D, et al. Insights from Bacillus anthracis strains isolated from permafrost in the tundra zone of Russia. PLoS ONE. (2019) 14:e0209140. doi: 10.1371/journal.pone.0209140
112. Tessaro SV. The existing and potential importance of brucellosis and tuberculosis in canadian wildlife: a review. Can Vet J. (1986) 27:119–24.
113. Shury TK, Nishi JS, Elkin BT, Wobeser GA. Tuberculosis and brucellosis in wood bison (bison bison athabascae) in northern Canada: a renewed need to develop options for future management. J Wildl Dis. (2015) 51:543–54. doi: 10.7589/2014-06-167
114. Tryland M. Zoonoses of arctic marine mammals. Infect Dis Rev. (2000) 2:55–64. Available online at: https://scholar.google.com/scholar_lookup?title=Zoonoses+of+arctic+marine+mammals&author=Tryland,+M.&publication_year=2000&journal=Infect.+Dis.+Rev.&volume=2&pages=55%E2%80%9364
115. Hauschild AHW, Gauvreau L. Food-borne botulism in Canada, 1971-84. Can Med Assoc J. (1985) 133:1141–6.
116. Leclair D, Fung J, Isaac-Renton JL, Proulx J-F, May-Hadford J, Ellis A, et al. Foodborne botulism in Canada, 1985–2005. Emerg Infect Dis. (2013) 19:961–8. doi: 10.3201/eid1906.120873
117. Shaffer N, Wainwright RB, Middaugh JP, Tauxe RV. Botulism among Alaska natives: the role of changing food preparation and consumption practices. West J Med. (1990) 153:390–3.
118. Food Safety Network. Safe Preparation and Storage of Aboriginal Traditional/Country Foods: A Review. Ottawa, ON: FSN (2008).
119. Franco DA. Botulism. In: Beran GW, Steele JH, Benenson AS, Torten M, Dreesen DW, Ristic M, Pier AC, editors. Handbook of Zoonoses. Boca Raton, FL: CRC Press (2009). p. 361–7. doi: 10.1201/9781003006107-27
120. Kutz S, Bollinger T, Branigan M, Checkley S, Davison T, Dumond M, et al. Erysipelothrix rhusiopathiae associated with recent widespread muskox mortalities in the Canadian Arctic. Can Vet J. (2015) 56:560–3.
121. Forde TL, Orsel K, Zadoks RN, Biek R, Adams LG, Checkley SL, et al. Bacterial genomics reveal the complex epidemiology of an emerging pathogen in arctic and boreal ungulates. Front Microbiol. (2016) 7:1759. doi: 10.3389/fmicb.2016.01759
122. Mavrot F, Orsel K, Hutchins W, Adams LG, Beckmen K, Blake JE, et al. Novel insights into serodiagnosis and epidemiology of Erysipelothrix rhusiopathiae, a newly recognized pathogen in muskoxen (Ovibos moschatus). PLoS ONE. (2020) 15:e0231724. doi: 10.1371/journal.pone.0231724
123. Chiu A, Goddard E, Parlee B. Caribou consumption in northern Canadian communities. J Toxicol Environ Heal Part A. (2016) 79:762–97. doi: 10.1080/15287394.2016.1174011
124. Groeschel M, Forde T, Turvey S, Joffe AM, Hui C, Naidu P, et al. An unusual case of Erysipelothrix rhusiopathiae prosthetic joint infection from the Canadian Arctic: whole genome sequencing unable to identify a zoonotic source. BMC Infect Dis. (2019) 19:282. doi: 10.1186/s12879-019-3913-7
125. Canadian Wildlife Health Cooperative. CWHC Manuals and Guides: Safety Manual for Harvesters of Fish and Wildlife in Nunavut. Saskatoon, SK: Canadian Cooperative Wildlife Health Centre (2012).
126. Washington State University IACUC. Zoonoses Associated with Fish | Institutional Animal Care and Use Committee. Washington State University. Available online at: https://iacuc.wsu.edu/zoonoses-associated-with-fish/ (accessed October 27, 2020).
127. Schwantje H, Macbeth JB, Kutz S, Elkin B. British Columbia Boreal Caribou Health Program Progress Report: Year 1 (November 1, 2013 – December 31, 2014), Victoria, BC: The British Columbia Oil and Gas Research and Innovation Society (OGRIS) (2014).
128. Brooke CJ, Riley TV. Erysipelothrix rhusiopathiae: bacteriology, epidemiology and clinical manifestations of an occupational pathogen. J Med Microbiol. (1999) 48:789–99. doi: 10.1099/00222615-48-9-789
129. Kassim SS, Dibernardo A, Lindsay LR, Wuerz TC. Locally acquired leptospirosis in expedition racer, manitoba, Canda. Emerg Infect Dis. (2018) 24:2386–8. doi: 10.3201/eid2412.181015
130. Smith AW, Brown RJ, Skilling DE, Bray HL, Keyes MC. Naturally-Occurring leptospirosis in Northern fur seals (Callorhinus ursinus). J Wildl Dis. (1977) 13:144–8. doi: 10.7589/0090-3558-13.2.144
131. Messier V, Lévesque B, Proulx JF, Rochette L, Serhir B, Couillard M, et al. Seroprevalence of seven zoonotic infections in Nunavik, Quebec (Canada). Zoo Public Health. (2012) 59:107–17. doi: 10.1111/j.1863-2378.2011.01424.x
132. Larsson C, Comstedt P, Olsen B, Bergström S. First record of lyme disease borrelia in the Arctic. Vector Borne Zoo Dis. (2007) 7:453–6. doi: 10.1089/vbz.2006.0644
133. Scott JD, Anderson JF, Durden LA. Widespread dispersal of borrelia burgdorferi–infected ticks collected from songbirds across Canada. J Parasitol. (2012) 98:49–59. doi: 10.1645/GE-2874.1
134. Sundeep S, Cleeve V. Isolation of Bisgaardia hudsonensis from a seal bite. Case report and review of the literature on seal finger. J Infect. (2011) 63:86–8. doi: 10.1016/j.jinf.2011.04.006
135. Hansen MJ, Bertelsen MF, Christensen H, Bisgaard M, Bojesen AM. occurrence of pasteurellaceae bacteria in the oral cavity of selected marine mammal species. J Zoo Wildl Med. (2012) 43:828–35. doi: 10.1638/2011-0264R1.1
136. Blanchong JA, Samuel MD, Goldberg DR, Shadduck DJ, Lehr MA. Persistence of pasteurella multocida in wetlands following avian cholera outbreaks. J Wildl Dis. (2006) 42:33–9. doi: 10.7589/0090-3558-42.1.33
137. Crawford RL, Blyde D, Blackall PJ, Forde BM, Beatson SA, Harris LM, et al. Novel insights into pasteurellosis in captive pinnipeds. Vet Microbiol. (2019) 231:232–7. doi: 10.1016/j.vetmic.2019.03.017
138. Miernyk KM, Bruden D, Parkinson AJ, Hurlburt D, Klejka J, Berner J, et al. Human Seroprevalence to 11 zoonotic pathogens in the U.S. Arctic, Alaska. Vector Borne Zoo Dis. (2019) 19:563–75. doi: 10.1089/vbz.2018.2390
139. Duncan C, Savage K, Williams M, Dickerson B, Kondas AV, Fitzpatrick KA, et al. Multiple strains of coxiella burnetii are present in the environment of St. Paul Island, Alaska. Transbound Emerg Dis. (2013) 60:345–50. doi: 10.1111/j.1865-1682.2012.01353.x
140. Duncan C, Dickerson B, Pabilonia K, Miller A, Gelatt T. Prevalence of Coxiella burnetii and Brucella spp. in tissues from subsistence harvested northern fur seals (Callorhinus ursinus) of St. Paul Island, Alaska. Acta Vet Scand. (2014) 56:67. doi: 10.1186/s13028-014-0067-x
141. Westley BP, Horazdovsky RD, Michaels DL, Brown DR. Identification of a novel mycoplasma species in a patient with septic arthritis of the hip and seal finger. Clin Infect Dis. (2015) 62:491–3. doi: 10.1093/cid/civ875
142. Wobeser G, Campbell GD, Dallaire A, McBurney S. Tularemia, plague, yersiniosis, and tyzzer's disease in wild rodents and lagomorphs in Canada: a review. Can Vet J. (2009) 50:1251–6.
143. Barnett JEF, Woodley AJ, Hill TJ, Turner L. Conditions in grey seal pups (Halichoerus grypus) presented for rehabilitation. Vet Rec. (2000) 147:98–104. doi: 10.1136/vr.147.4.98
144. Sunstrum J, Shoyinka A, Power LE, Maxwell D, Stobierski MG, Signs K, et al. Notes from the field: zoonotic mycobacterium bovis disease in deer hunters — michigan, 2002–2017. MMWR Morb Mortal Wkly Rep. (2019) 68:807–8. doi: 10.15585/mmwr.mm6837a3
145. Hansen CM, Vogler AJ, Keim P, Wagner DM, Hueffer K. Tularemia in Alaska, 1938 - 2010. Acta Vet Scand. (2011) 53:61. doi: 10.1186/1751-0147-53-61
146. Desvars A, Furberg M, Hjertqvist M, Vidman L, Sjöstedt A, Rydén P, et al. Epidemiology and ecology of tularemia in Sweden, 1984–2012. Emerg Infect Dis. (2015) 21:32–9. doi: 10.3201/eid2101.140916
147. Zellner B, Huntley JF. Ticks and tularemia: do we know what we don't know? Front Cell Infect Microbiol. (2019) 9:146. doi: 10.3389/fcimb.2019.00146
148. Posautz A, Gyuranecz M, Dénes B, Knauer F, Dier H, Walzer C. Tularemia - possible increase and new risk factors. Int J Infect Dis. (2019) 79:60. doi: 10.1016/j.ijid.2018.11.155
149. Antonation KS, Shury TK, Bollinger TK, Olson A, Mabon P, Van Domselaar G, et al. Sylvatic plague in a canadian black-tailed prairie dog (Cynomys ludovicianus). J Wildl Dis. (2014) 50:699–702. doi: 10.7589/2013-08-215
150. Liccioli S, Stephens T, Wilson SC, McPherson JM, Keating LM, Antonation KS, et al. Enzootic maintenance of sylvatic plague in Canada's threatened black-tailed prairie dog ecosystem. Ecosphere. (2020) 11:e03138. doi: 10.1002/ecs2.3138
151. Blake JE, McLean BD, Gunn A. Yersiniosis in free-ranging muskoxen on banks island, northwest territories, Canada. J Wildl Dis. (1991) 27:527–33. doi: 10.7589/0090-3558-27.4.527
152. Reed C, Bruden D, Byrd KK, Veguilla V, Bruce M, Hurlburt D, et al. Characterizing wild bird contact and seropositivity to highly pathogenic avian influenza A (H5N1) virus in Alaskan residents. Influ Other Respi Viruses. (2014) 8:516–23. doi: 10.1111/irv.12253
153. USDA. USDA APHIS | USDA Confirms Highly Pathogenic H5N2 Avian Influenza in a Wild Mallard Duck in Alaska. (2016). Available online at: https://www.aphis.usda.gov/aphis/newsroom/stakeholder-info/sa_by_date/stakeholder-announcements-2016/sa-08/hpai-ak (accessed June 27, 2020).
154. Nielsen O, Clavijo A, Boughen JA. Serologic evidence of influenza a infection in marine mammals of Arctic Canada. J Wildl Dis. (2001) 37:820–5. doi: 10.7589/0090-3558-37.4.820
155. Webster R, Geraci J, Petursson G, Skirnisson K. Conjunctivitis in human beings caused by influenza a virus of seals. N Engl J Med. (1981) 304:911. doi: 10.1056/NEJM198104093041515
156. Smith AW, Berry ES, Skilling DE, Barlough JE, Poet SE, Berke T, et al. In vitro isolation and characterization of a calicivirus causing a vesicular disease of the hands and feet. Clin Infect Dis. (1998) 26:434–9. doi: 10.1086/516311
157. Tomaselli M, Dalton C, Duignan PJ, Kutz S, van der Meer F, Kafle P, et al. Contagious ecthyma, rangiferine brucellosis, and lungworm infection in a muskox (Ovibos moschatus) from the Canadian Arctic, 2014. J Wildl Dis. (2016) 52:719–24. doi: 10.7589/2015-12-327
158. Vikøren T, Lillehaug A, Åkerstedt J, Bretten T, Haugum M, Tryland M. A severe outbreak of contagious ecthyma (orf) in a free-ranging musk ox (Ovibos moschatus) population in Norway. Vet Microbiol. (2008) 127:10–20. doi: 10.1016/j.vetmic.2007.07.029
159. Tryland M, Beckmen KB, Burek-Huntington KA, Breines EM, Klein J. Orf virus infection in Alaskan mountain goats, dall's sheep, muskoxen, caribou and sitka black-tailed deer. Acta Vet Scand. (2018) 60:12. doi: 10.1186/s13028-018-0366-8
161. Hueffer K, Murphy M. Rabies in Alaska, from the past to an uncertain future. Int J Circumpolar Health. (2018) 77:1475185. doi: 10.1080/22423982.2018.1475185
162. Rausch RL. Some observations on rabies in alaska, with special reference to wild canidae. J Wildl Manage. (1958) 22:246–280.
163. Ducrocq J, Proulx J, Lévesque B, De Serres G, Wood H, Lemire M. Assessment of naturally acquired neutralizing antibodies against rabies lyssavirus in a subset of nunavik's inuit population considered most at risk of being exposed to rabid animals. Zoo Public Health. (2019) 66:533–9. doi: 10.1111/zph.12561
164. Mørk T, Prestrud P. Arctic rabies - a review. Acta Vet Scand. (2004) 45:1–9. doi: 10.1186/1751-0147-45-1
165. Aenishaenslin C, Simon A, Forde T, Ravel A, Proulx J-F, Fehlner-Gardiner C, et al. Characterizing rabies epidemiology in remote inuit communities in Québec, Canada: a “one health” approach. Ecohealth. (2014) 11:343–55. doi: 10.1007/s10393-014-0923-1
167. Alaska Department of Fish and Game. Rabies in Alaska: Rabid Wolverine Found on North Slope. (2012). Available online at: http://www.adfg.alaska.gov/index.cfm?adfg=wildlifenews.view_article&articles_id=582 (accessed October 27, 2020).
168. Odegaard O, Krogsrud J. Rabies in Svalbard: infection diagnosed in arctic fox, reindeer and seal. Vet Rec. (1981) 109:141–2. doi: 10.1136/vr.109.7.141
169. Rausch R. Tropical problems in the Arctic: infectious and parasitic diseases, a common denominator. Ind Trop Health. (1974) 8:63–70.
170. Tabel H, Corner AH, Webster WA, Casey CA. History and epizootiology of rabies in Canada. Can Vet J. (1974) 15:271–81.
171. Minuk G, Sun A, Sun D, Uhanova J, Nicolle L, Larke B, et al. Serological evidence of hepatitis e virus infection in an indigenous north american population. Can J Gastroenterol. (2007) 21:439–42. doi: 10.1155/2007/289059
172. Weger S, Elkin B, Lindsay R, Bollinger T, Crichton V, Andonov A. Hepatitis E virus seroprevalence in free-ranging deer in Canada. Transbound Emerg Dis. (2017) 64:1008–11. doi: 10.1111/tbed.12462
173. Teshale EH, Hu DJ. Hepatitis E: epidemiology and prevention. World J Hepatol. (2011) 3:285–91. doi: 10.4254/wjh.v3.i12.285
174. Gozlan RE, Marshall WL, Lilje O, Jessop CN, Gleason FH, Andreou D. Current ecological understanding of fungal-like pathogens of fish: what lies beneath? Front Microbiol. (2014) 5:62. doi: 10.3389/fmicb.2014.00062
175. Güere ME, Våge J, Tharaldsen H, Benestad SL, Vikøren T, Madslien K, et al. Chronic wasting disease associated with prion protein gene (PRNP) variation in Norwegian wild reindeer (Rangifer tarandus). Prion. (2020) 14:1–10. doi: 10.1080/19336896.2019.1702446
176. Waddell L, Greig J, Mascarenhas M, Otten A, Corrin T, Hierlihy K. Current evidence on the transmissibility of chronic wasting disease prions to humans-a systematic review. Transbound Emerg Dis. (2018) 65:37–49. doi: 10.1111/tbed.12612
177. Revich B, Tokarevich N, Parkinson AJ. Climate change and zoonotic infections in the Russian Arctic. Int J Circumpolar Health. (2012) 71:18792. doi: 10.3402/ijch.v71i0.18792
178. Davidson R, Simard M, Kutz SJ, Kapel CMO, Hamnes IS, Robertson LJ. Arctic parasitology: why should we care? Trends Parasitol. (2011) 27:239–45. doi: 10.1016/j.pt.2011.02.001
179. Ferguson MAD. rangiferine brucellosis on Baffin Island. J Wildl Dis. (1997) 33:536–43. doi: 10.7589/0090-3558-33.3.536
180. Goyette S, Cao Z, Libman M, Ndao M, Ward BJ. Seroprevalence of parasitic zoonoses and their relationship with social factors among the Canadian inuit in Arctic regions. Diagn Microbiol Infect Dis. (2014) 78:404–10. doi: 10.1016/j.diagmicrobio.2013.08.026
181. Campagna S, Lévesque B, Anassour-Laouan-Sidi E, Côté S, Serhir B, Ward BJ, et al. Seroprevalence of 10 zoonotic infections in 2 Canadian cree communities. Diagn Microbiol Infect Dis. (2011) 70:191–9. doi: 10.1016/j.diagmicrobio.2011.01.009
182. Schurer JM, Quewezance H, Ndao M, Elmore SA, Jenkins EJ. People, pets, and parasites: one health surveillance in Southeastern Saskatchewan. Am J Trop Med Hyg. (2014) 90:1184–90. doi: 10.4269/ajtmh.13-0749
183. Schurer JM, Ndao M, Skinner S, Irvine J, Elmore SA, Epp T, et al. Parasitic zoonoses: one health surveillance in Northern Saskatchewan. PLoS Negl Trop Dis. (2013) 7:e2141. doi: 10.1371/journal.pntd.0002141
184. Jenkins EJ, Schurer JM, Gesy KM. Old problems on a new playing field: Helminth zoonoses transmitted among dogs, wildlife, and people in a changing northern climate. Vet Parasitol. (2011) 182:54–69. doi: 10.1016/j.vetpar.2011.07.015
185. Parkinson AJ, Berner J. Climate change and impacts on human health in the Arctic: an international workshop on emerging threats and the response of Arctic communities to climate change. Int J Circumpolar Health. (2009) 68:84–91. doi: 10.3402/ijch.v68i1.18295
186. Bradley MJ, Kutz SJ, Jenkins E, O'Hara TM. The potential impact of climate change on infectious diseases of Arctic fauna. Int J Circumpolar Health. (2005) 64:468–77. doi: 10.3402/ijch.v64i5.18028
187. Gallana M, Ryser-Degiorgis MP, Wahli T, Segner H. Climate change and infectious diseases of wildlife: altered interactions between pathogens, vectors and hosts. Curr Zool. (2013) 59:427–37. doi: 10.1093/czoolo/59.3.427
188. Elmore SA, Jenkins EJ, Huyvaert KP, Polley L, Root JJ, Moore CG. Toxoplasma gondii in circumpolar people and wildlife. Vector Borne Zoo Dis. (2012) 12:1–9. doi: 10.1089/vbz.2011.0705
189. Proulx J, MacLean JD, Gyorkos TW, Leclair D, Richter A, Serhir B, et al. Novel prevention program for trichinellosis in inuit communities. Clin Infect Dis. (2002) 34:1508–14. doi: 10.1086/340342
190. Tomaselli M, Elkin B, Kutz S, Harms NJ, Nymo HI, Davison T, et al. A transdisciplinary approach to brucella in muskoxen of the western Canadian Arctic 1989–2016. Ecohealth. (2019) 16:488–501. doi: 10.1007/s10393-019-01433-3
191. Bodenstein B, Beckmen K, Sheffield G, Kuletz K, Van Hemert C, Berlowski B, et al. Avian cholera causes marine bird mortality in the bering sea of Alaska. J Wildl Dis. (2015) 51:934–7. doi: 10.7589/2014-12-273
192. Bruce M, Zulz T, Koch A. Surveillance of infectious diseases in the Arctic. Public Health. (2016) 137:5–12. doi: 10.1016/j.puhe.2016.06.014
193. Tomaselli M, Kutz S, Gerlach C, Checkley S. Local knowledge to enhance wildlife population health surveillance: conserving muskoxen and caribou in the Canadian Arctic. Biol Conserv. (2018) 217:337–48. doi: 10.1016/j.biocon.2017.11.010
194. Olival KJ, Hosseini PR, Zambrana-Torrelio C, Ross N, Bogich TL, Daszak P. Host and viral traits predict zoonotic spillover from mammals. Nature. (2017) 546:646–50. doi: 10.1038/nature22975
195. Woolhouse M, Gaunt E. Ecological origins of novel human pathogens. Crit Rev Microbiol. (2007) 33:231–42. doi: 10.1080/10408410701647560
196. Woolhouse M, Scott F, Hudson Z, Howey R, Chase-Topping M. Human viruses: discovery and emergence. Philos Trans R Soc B Biol Sci. (2012) 367:2864–71. doi: 10.1098/rstb.2011.0354
197. Brook RK, Kutz SJ, Millins C, Veitch AM, Elkin BT, Leighton T. Evaluation and delivery of domestic animal health services in remote communities in the Northwest territories: a case study of status and needs. Can Vet J. (2010) 51:1115–22.
198. Palmater P. Priority Pandemic Response Needed for First Nations. Toronto, ON: Policy Options (2020).
199. Alexander KA, Carlson CJ, Lewis BL, Getz WM, Marathe MV, Eubank SG, et al. The ecology of pathogen spillover and disease emergence at the human-wildlife-environment interface. Adv Environ Microbiol. 5:267–98. doi: 10.1007/978-3-319-92373-4_8
200. Brierley L, Vonhof MJ, Olival KJ, Daszak P, Jones KE. Quantifying global drivers of zoonotic bat viruses: a process-based perspective. Am Nat. (2016) 187:E53–64. doi: 10.1086/684391
201. Kreuder Johnson C, Hitchens PL, Smiley Evans T, Goldstein T, Thomas K, Clements A, et al. Spillover and pandemic properties of zoonotic viruses with high host plasticity. Sci Rep. (2015) 5:14830. doi: 10.1038/srep14830
202. White RJ, Razgour O. Emerging zoonotic diseases originating in mammals: a systematic review of effects of anthropogenic land-use change. Mamm Rev. (2020) 50:336–52. doi: 10.1111/mam.12201
203. Borland S, Gracieux P, Jones M, Mallet F, Yugueros-Marcos J. Influenza A virus infection in cats and dogs: a literature review in the light of the “one health” concept. Front Public Health. (2020) 8:83. doi: 10.3389/fpubh.2020.00083
204. Heeney JL. Zoonotic viral diseases and the frontier of early diagnosis, control and prevention. J Intern Med. (2006) 260:399–408. doi: 10.1111/j.1365-2796.2006.01711.x
205. Wardeh M, Sharkey KJ, Baylis M. Integration of shared-pathogen networks and machine learning reveals the key aspects of zoonoses and predicts mammalian reservoirs. Proc R Soc B Biol Sci. (2020) 287:20192882. doi: 10.1098/rspb.2019.2882
206. Plowright RK, Parrish CR, McCallum H, Hudson PJ, Ko AI, Graham AL, et al. Pathways to zoonotic spillover. Nat Rev Microbiol. (2017) 15:502–10. doi: 10.1038/nrmicro.2017.45
207. Carver S, Mills JN, Parmenter CA, Parmenter RR, Richardson KS, Harris RL, et al. Toward a mechanistic understanding of environmentally forced zoonotic disease emergence: sin nombre hantavirus. Bioscience. (2015) 65:651–66. doi: 10.1093/biosci/biv047
208. Fedeli C, Moreno H, Kunz S. Novel insights into cell entry of emerging human pathogenic arenaviruses. J Mol Biol. (2018) 430:1839–52. doi: 10.1016/j.jmb.2018.04.026
209. Sokolow SH, Nova N, Pepin KM, Peel AJ, Pulliam JRC, Manlove K, et al. Ecological interventions to prevent and manage zoonotic pathogen spillover. Philos Trans R Soc B Biol Sci. (2019) 374:20180342. doi: 10.1098/rstb.2018.0342
210. Plowright RK, Peel AJ, Streicker DG, Gilbert AT, McCallum H, Wood J, et al. Transmission or within-host dynamics driving pulses of zoonotic viruses in reservoir–host populations. PLoS Negl Trop Dis. (2016) 10:e0004796. doi: 10.1371/journal.pntd.0004796
211. Guth S, Visher E, Boots M, Brook CE. Host phylogenetic distance drives trends in virus virulence and transmissibility across the animal–human interface. Philos Trans R Soc B Biol Sci. (2019) 374:20190296. doi: 10.1098/rstb.2019.0296
212. Tatem AJ, Hay SI, Rogers DJ. Global traffic and disease vector dispersal. Proc Natl Acad Sic USA. (2006) 103:6242–7. doi: 10.1073/pnas.0508391103
213. Bajardi P, Poletto C, Ramasco JJ, Tizzoni M, Colizza V, Vespignani A. Human mobility networks, travel restrictions, and the global spread of 2009 H1N1 pandemic. PLoS ONE. (2011) 6:e16591. doi: 10.1371/journal.pone.0016591
214. Lycett SJ, Duchatel F, Digard P. A brief history of bird flu. Philos Trans R Soc B Biol Sci. (2019) 374:20180257. doi: 10.1098/rstb.2018.0257
215. Cleaveland S, Haydon DT, Taylor L. Overviews of pathogen emergence: which pathogens emerge, when and why? Curr Top Microbiol Immunol. (2007) 315:85–111. doi: 10.1007/978-3-540-70962-6_5
216. Wang B, Harms D, Yang XL, Bock CT. Orthohepevirus C: an expanding species of emerging hepatitis e virus variants. Pathogens. (2020) 9:154. doi: 10.3390/pathogens9030154
217. Washburne AD, Crowley DE, Becker DJ, Olival KJ, Taylor M, Munster VJ, et al. Taxonomic patterns in the zoonotic potential of mammalian viruses. PeerJ. (2018) 6:e5979. doi: 10.7717/peerj.5979
218. Luis AD, Hayman DTS, O'Shea TJ, Cryan PM, Gilbert AT, Pulliam JRC, et al. A comparison of bats and rodents as reservoirs of zoonotic viruses: are bats special? Proc R Soc B Biol Sci. (2013) 280:20122753. doi: 10.1098/rspb.2012.2753
219. Cleaveland S, Laurenson MK, Taylor LH. Diseases of humans and their domestic mammals: pathogen characteristics, host range and the risk of emergence. Philos Trans R Soc London Ser B Biol Sci. (2001) 356:991–9. doi: 10.1098/rstb.2001.0889
220. Mollentze N, Streicker DG. Viral zoonotic risk is homogenous among taxonomic orders of mammalian and avian reservoir hosts. Proc Natl Acad Sic USA. (2020) 117:9423–30. doi: 10.1073/pnas.1919176117
221. Achenbach JE, Bowen RA. Transmission of avian influenza A viruses among species in an artificial barnyard. PLoS ONE. (2011) 6:e17643. doi: 10.1371/journal.pone.0017643
222. Bosco-Lauth AM, Bowen RA, Root JJ. Limited transmission of emergent H7N9 influenza A virus in a simulated live animal market: do chickens pose the principal transmission threat? Virology. (2016) 495:161–66. doi: 10.1016/j.virol.2016.04.032
223. Gervasi SS, Burgan SC, Hofmeister E, Unnasch TR, Martin LB. Stress hormones predict a host superspreader phenotype in the West Nile virus system. Proc R Soc B Biol Sci. (2017) 284:20171090. doi: 10.1098/rspb.2017.1090
224. Hing S, Narayan EJ, Thompson RCA, Godfrey SS. The relationship between physiological stress and wildlife disease: consequences for health and conservation. Wildl Res. (2016) 43:51. doi: 10.1071/WR15183
225. Seeber PA, Quintard B, Sicks F, Dehnhard M, Greenwood AD, Franz M. Environmental stressors may cause equine herpesvirus reactivation in captive grévy's zebras (Equus grevyi). PeerJ. (2018) 6:e5422. doi: 10.7717/peerj.5422
226. Patz JA, Daszak P, Tabor GM, Aguirre AA, Pearl M, Epstein J, et al. Unhealthy landscapes: policy recommendations on land use change and infectious disease emergence. Environ Health Perspect. (2004) 112:1092–8. doi: 10.1289/ehp.6877
227. Pike BL, Saylors KE, Fair JN, LeBreton M, Tamoufe U, Djoko CF, et al. The origin and prevention of pandemics. Clin Infect Dis. (2010) 50:1636–40. doi: 10.1086/652860
228. Wolfe ND, Daszak P, Kilpatrick AM, Burke DS. Bushmeat hunting, deforestation, and prediction of zoonotic disease. Emerg Infect Dis. (2005) 11:1822–7. doi: 10.3201/eid1112.040789
229. Valitutto MT, Aung O, Tun KYN, Vodzak ME, Zimmerman D, Yu JH, et al. Detection of novel coronaviruses in bats in Myanmar. PLoS ONE. (2020) 15:e0230802. doi: 10.1371/journal.pone.0230802
230. Purse BV, Darshan N, Kasabi GS, Gerard F, Samrat A, George C, et al. Predicting disease risk areas through co-production of spatial models: the example of kyasanur forest disease in india's forest landscapes. PLoS Negl Trop Dis. (2020) 14:e0008179. doi: 10.1371/journal.pntd.0008179
231. Tijjani M, Majid RA, Abdullahi SA, Unyah NZ. Detection of rodent-borne parasitic pathogens of wild rats in Serdang, Selangor, Malaysia: a potential threat to human health. Int J Parasitol Parasites Wildl. (2020) 11:174–82. doi: 10.1016/j.ijppaw.2020.01.008
232. Grimm K, Rivera NA, Fredebaugh-Siller S, Weng H-Y, Warner RE, Maddox CW, et al. Evidence Of leptospira serovars in wildlife and leptospiral DNA in water sources in a natural area in east-central Illinois, USA. J Wildl Dis. (2020) 56:316. doi: 10.7589/2019-03-069
233. Adesiyun AA, Knobel DL, Thompson PN, Wentzel J, Kolo FB, Kolo AO, et al. Sero-Epidemiological study of selected zoonotic and abortifacient pathogens in cattle at a wildlife-livestock interface in South Africa. Vector-Borne Zoonotic Dis. (2020) 20:258–67. doi: 10.1089/vbz.2019.2519
234. Lönker NS, Fechner K, Abd El Wahed A. Horses as a crucial part of one health. Vet Sci. (2020) 7:28. doi: 10.3390/vetsci7010028
235. Narat V, Alcayna-Stevens L, Rupp S, Giles-Vernick T. Rethinking human–nonhuman primate contact and pathogenic disease spillover. Ecohealth. (2017) 14:840–50. doi: 10.1007/s10393-017-1283-4
236. Reaser JK, Clark EE, Meyers NM. All creatures great and minute: a public policy primer for companion animal zoonoses. Zoo Public Health. (2008) 55:385–401. doi: 10.1111/j.1863-2378.2008.01123.x
237. Gibb R, Redding DW, Chin KQ, Donnelly CA, Blackburn TM, Newbold T, et al. Zoonotic host diversity increases in human-dominated ecosystems. Nature. (2020) 584:398–402. doi: 10.1038/s41586-020-2562-8
238. Jones BA, Grace D, Kock R, Alonso S, Rushton J, Said MY, et al. Zoonosis emergence linked to agricultural intensification and environmental change. Proc Natl Acad Sic USA. (2013) 110:8399–404. doi: 10.1073/pnas.1208059110
239. Keesing F, Belden LK, Daszak P, Dobson A, Harvell CD, Holt RD, et al. Impacts of biodiversity on the emergence and transmission of infectious diseases. Nature. (2010) 468:647–52. doi: 10.1038/nature09575
240. Morand S, Owers K, Bordes F. Biodiversity and emerging zoonoses. In: Yamada A, Kahn LH, Kaplan B, Monath TP, Woodall J, Conti L, editors. Confronting Emerging Zoonoses. Tokyo: Springer (2014). p. 27–41. doi: 10.1007/978-4-431-55120-1_3
241. Murray KA, Daszak P. Human ecology in pathogenic landscapes: two hypotheses on how land use change drives viral emergence. Curr Opin Virol. (2013) 3:79–83. doi: 10.1016/j.coviro.2013.01.006
242. Anthony SJ, Johnson CK, Greig DJ, Kramer S, Che X, Wells H, et al. Global patterns in coronavirus diversity. Virus Evol. (2017) 3:vex012. doi: 10.1093/ve/vex012
243. Horby PW, Hoa NT, Pfeiffer DU, Wertheim HFL. Drivers of emerging zoonotic infectious diseases. Confr Emerg Zoonoses. (2014) 13–26. doi: 10.1007/978-4-431-55120-1_2
244. Roberts MG, Heesterbeek JAP. Quantifying the dilution effect for models in ecological epidemiology. J R Soc Interface. (2018) 15:20170791. doi: 10.1098/rsif.2017.0791
245. Khalil H, Ecke F, Evander M, Magnusson M, Hörnfeldt B. Declining ecosystem health and the dilution effect. Sci Rep. (2016) 6:31314. doi: 10.1038/srep31314
246. Borremans B, Faust C, Manlove KR, Sokolow SH, Lloyd-Smith JO. Cross-species pathogen spillover across ecosystem boundaries: mechanisms and theory. Philos Trans R Soc B Biol Sci. (2019) 374:20180344. doi: 10.1098/rstb.2018.0344
247. Wilkinson DA, Marshall JC, French NP, Hayman DTS. Habitat fragmentation, biodiversity loss and the risk of novel infectious disease emergence. J R Soc Interface. (2018) 15:20180403. doi: 10.1098/rsif.2018.0403
248. Evans T, Olson S, Watson J, Gruetzmacher K, Pruvot M, Jupiter S, et al. Links between ecological integrity, emerging infectious diseases originating from wildlife, and other aspects of human health -an overview of the literature. Wildl Conservat Soc. (2020) 4:e303. doi: 10.13140/RG.2.2.34736.51205
249. Kutz SJ, Jenkins EJ, Veitch AM, Ducrocq J, Polley L, Elkin B, et al. The arctic as a model for anticipating, preventing, and mitigating climate change impacts on host–parasite interactions. Vet Parasitol. (2009) 163:217–28. doi: 10.1016/j.vetpar.2009.06.008
250. Kutz SJ, Hoberg EP, Molnár PK, Dobson A, Verocai GG. A walk on the tundra: host-parasite interactions in an extreme environment. Int J Parasitol Parasites Wildl. (2014) 3:198–208. doi: 10.1016/j.ijppaw.2014.01.002
251. Meltofte H, Barry T, Berteaux D, Bültmann H, Christiansen JS, Cook JA, et al. Arctic Biodiversity Assessment: Status and Trends in Arctic Biodiversity; Synthesis. Akureyri: Conservation of Arctic Flora and Fauna (2013).
252. Ruviniyia K, Abdullah DA, Sumita S, Lim YAL, Ooi PT, Sharma RSK. Molecular detection of porcine Enterocytozoon bieneusi infection in Peninsular Malaysia and epidemiological risk factors associated with potentially zoonotic genotypes. Parasitol Res. (2020) 119:1663–74. doi: 10.1007/s00436-020-06648-w
253. Hueffer K, O'Hara TM, Follmann EH. Adaptation of mammalian host-pathogen interactions in a changing arctic environment. Acta Vet Scand. (2011) 53:17. doi: 10.1186/1751-0147-53-17
254. Nunavut Wildlife Management Board. The Nunavut Wildlife Harvest Study Feb 2004. Iqaluit, NU: Nunavut Wildlife Management Board (2004).
255. Aarluk Consulting. Review of Intersettlement Trade Opportunities for Arctic Food Products in Nunavut Final Report. (2005).
256. Gombay N. The commoditization of country foods in nunavik: a comparative assessment of its development, applications, and significance. Arctic. (2005) 58:115–28. Available online at: https://www.jstor.org/stable/40512686
257. Turner NJ, Plotkin M, Kuhnlein H. Indigenous Peoples' food systems and well-being: interventions and policies for healthy communities. In: Kuhnlein HV, Erasmus B, Spigelski D, Burlingame B, editors. Indigenous Peoples' Food Systems and Well-Being. Rome: Food and Agriculture Organization (2013).
258. Dowsley M. The value of a polar bear: evaluating the role of a multiple-use resource in the nunavut mixed economy. Arctic Anthropol. (2010) 47:39–56. doi: 10.1353/arc.0.0035
259. Natcher D. Subsistence and the social economy of canada's aboriginal north | northern review. North Rev. (2009) 30:83–98. Available online at: https://thenorthernreview.ca/index.php/nr/article/view/6 (accessed April 12, 2021).
260. Reeves RR. The commerce in maktaq at Arctic Bay, northern Baffin Island, NWT. Arctic Anthropol. (1993) 30:79–93. doi: 10.2307/40316330
261. Damas J, Hughes GM, Keough KC, Painter CA, Persky NS, Corbo M, et al. Broad host range of SARS-CoV-2 predicted by comparative and structural analysis of ACE2 in vertebrates. bioRxiv [Preprint]. (2020). doi: 10.1101/2020.04.16.045302
262. Damas D. Central eskimo systems of food sharing. Ethnology. (1972) 11:220. doi: 10.2307/3773217
263. Wenzel GW. Ningiqtuq: resource sharing and generalized reciprocity in Clyde River, Nunavut. Arctic Anthropol. (1995) 32:43–60. doi: 10.2307/40316386
264. Collings P, Wenzel G, Condon RG. Modern food sharing networks and community integration in the central Canadian Arctic. Arctic. (1998) 51:301–14.
265. Kofinas G, BurnSilver SB, Magdanz J, Stotts R, Okada M. Subsistence Sharing Networks and Cooperation: Kaktovik, Wainwright, and Venetie, Alaska. BOEM Report 2015-023; AFES Report MP 2015-02. School of Natural Resources and Extension, University of Alaska Fairbanks.
266. Braund SR, Associates Anchorage Alaska. Description of Alaskan Eskimo Bowhead Whale Subsistence Sharing Practices. Final report submitted to the Alaska Eskimo Whaling Commission (2018).
267. Dombrowski K, Khan B, Channell E, Moses J, McLean K, Misshula E. Kinship, family, and exchange in a labrador inuit community. Arctic Anthropol. (2013) 50:89–104. doi: 10.3368/aa.50.1.89
268. Justice Law. Nunavut Land Claims Agreement Act: Article 5. Iqaluit, NU: Government of Canada (1993).
269. Bodenhorn B. It's good to know who your relatives are but we were taught to share with everybody: shares and sharing among inupiaq households. Senri Ethnol Stud. (2000) 53:27–60. Available online at: https://www.semanticscholar.org/paper/Meating-the-social%3A-sharing-atiku-euiash-in-Castro/3e1a8517b622aa4b1f0ebb332f707424cc06791d
270. Berkes F, Jolly D. Adapting to climate change: social-ecological resilience in a Canadian western arctic community. Ecol Soc. (2002) 5:18. doi: 10.5751/es-00342-050218
271. Bhawra J, Cooke MJ, Hanning R, Wilk P, Gonneville SLH. Community perspectives on food insecurity and obesity: focus groups with caregivers of métis and off-reserve first nations children. Int J Equity Health. (2015) 14:96. doi: 10.1186/s12939-015-0232-5
272. Dunn L, Gross P. Food-Sharing practices online in the facebook group cambridge bay news. Études Inuit Stud. (2019) 40:225–43. doi: 10.7202/1055440ar
273. BurnSilver S, Magdanz J, Stotts R, Berman M, Kofinas G. Are mixed economies persistent or transitional? Evidence using social networks from Arctic Alaska. Am Anthropol. (2016) 118:121–9. doi: 10.1111/aman.12447
275. Ancelle T, De Bruyne A, Poisson DM, Dupouy-Camet J. Outbreak of trichinellosis due to consumption of bear meat from Canada, France, September 2005. Wkly Releas. (2005) 10:pii2809. doi: 10.2807/esw.10.41.02809-en
276. Parkinson AJ, Evengard B, Semenza JC, Ogden N, Børresen ML, Berner J, et al. Climate change and infectious diseases in the Arctic: establishment of a circumpolar working group. Int J Circumpolar Health. (2014) 73:25163. doi: 10.3402/ijch.v73.25163
277. Dudley JP, Hoberg EP, Jenkins EJ, Parkinson AJ. Climate change in the north american arctic: a one health perspective. Ecohealth. (2015) 12:713–25. doi: 10.1007/s10393-015-1036-1
278. Kafle P, Peller P, Massolo A, Hoberg E, Leclerc L-M, Tomaselli M, et al. Range expansion of muskox lungworms track rapid arctic warming: implications for geographic colonization under climate forcing. Sci Rep. (2020) 10:17323. doi: 10.1038/s41598-020-74358-5
279. Wallenhammar A, Lindqvist R, Asghar N, Gunaltay S, Fredlund H, Davidsson Å, et al. Revealing new tick-borne encephalitis virus foci by screening antibodies in sheep milk. Parasit Vectors. (2020) 13:185. doi: 10.1186/s13071-020-04030-4
280. Brandt JP, Flannigan MD, Maynard DG, Thompson ID, Volney WJA. An introduction to Canada's boreal zone: ecosystem processes, health, sustainability, and environmental issues. Environ Rev. (2013) 21:207–226. doi: 10.1139/er-2013-0040
281. National Academies of Science Engineering Medicine. Understanding Responding to Global Health Security Risks from Microbial Threats in the Arctic. Everett L, editor. Washington, DC.: National Academies Press (2020). doi: 10.17226/25887
282. Van Hemert C, Pearce JM, Handel CM. Wildlife health in a rapidly changing North: focus on avian disease. Front Ecol Environ. (2014) 12:548–56. doi: 10.1890/130291
283. Gigante CM, Gao J, Tang S, McCollum AM, Wilkins K, Reynolds MG, et al. Genome of Alaskapox virus, a novel orthopoxvirus isolated from Alaska. Viruses. (2019) 11:708. doi: 10.3390/v11080708
284. Lee EC, Wada NI, Grabowski MK, Gurley ES, Lessler J. The engines of SARS-CoV-2 spread. Science. (2020) 370:406–7. doi: 10.1126/science.abd8755
285. Semenza JC, Lindgren E, Balkanyi L, Espinosa L, Almqvist MS, Penttinen P, et al. Determinants and drivers of infectious disease threat events in Europe. Emerg Infect Dis. (2016) 22:581–9. doi: 10.3201/eid2204.151073
286. Alaska State Government. First case of COVID-19 confirmed by Alaska State Public Health Laboratory is an international resident – Mike Dunleavy. (2020). Available online at: https://gov.alaska.gov/newsroom/2020/03/12/first-case-of-covid-19-confirmed-by-alaska-state-public-health-laboratory-is-an-international-resident/ (accessed October 1, 2020).
287. Doxey M, Chrzaszcz L, Dominguez A, James RD. A forgotten danger. J Public Heal Manag Pract. (2019) 25:S7–10. doi: 10.1097/PHH.0000000000001062
288. ProPublica. Remote Alaska Villages Isolate Themselves Further in Effort to Shield Against Coronavirus. Anchorage, AK: ProPublica (2020)
289. Gryseels S, De Bruyn L, Gyselings R, Calvignac-Spencer S, Leendertz FH, Leirs H. Risk of human-to-wildlife transmission of SARS-CoV-2. Mam Rev. (2021) 51:272–92. doi: 10.1111/mam.12225
290. Bossart GD, Schwartz JC. Acute necrotizing enteritis associated with suspected coronavirus infection in three harbor seals (Phoca vitulina). J Zoo Wildl Med. (1990) 21:84–7.
291. Mihindukulasuriya KA, Wu G St. Leger J, Nordhausen RW, Wang D. Identification of a novel coronavirus from a beluga whale by using a panviral microarray. J Virol. (2008) 82:5084–8. doi: 10.1128/JVI.02722-07
292. Nollens HH, Wellehan JFX, Archer L, Lowenstine LJ, Gulland FMD. Detection of a respiratory coronavirus from tissues archived during a pneumonia epizootic in free-ranging pacific harbor seals Phoca vitulina richardsii. Dis Aquat Organ. (2010) 90:113–20. doi: 10.3354/dao02190
293. Woo PCY, Lau SKP, Lam CSF, Tsang AKL, Hui S-W, Fan RYY, et al. Discovery of a novel bottlenose dolphin coronavirus reveals a distinct species of marine mammal coronavirus in gammacoronavirus. J Virol. (2014) 88:1318–31. doi: 10.1128/JVI.02351-13
294. Messenger AM, Barnes AN, Gray GC. Reverse zoonotic disease transmission (zooanthroponosis): A systematic review of seldom-documented human biological threats to animals. PLoS ONE. (2014) 9:e89055. doi: 10.1371/journal.pone.0089055
295. Runge MC, Campbell Grant EH, Coleman JTH, Reichard JD, Gibbs SEJ, Cryan PM, et al. Assessing the Risks Posed by SARS-CoV-2 in and Via North American Bats—Decision Framing and Rapid Risk Assessment. Reston, VA: U.S. Geological Survey (2020). doi: 10.3133/ofr20201060
296. Canadian Wildlife Health Cooperative. Wildlife Health and COVID-19 in Canada: Bats. (2020). Available online At: http://cwhc-rcsf.ca/docs/miscellaneous/CWHC Bat health and Covid-19 (Version 1.0 April 23 2020).pdf
297. Olival KJ, Cryan PM, Amman BR, Baric RS, Blehert DS, Brook CE, et al. Possibility for reverse zoonotic transmission of SARS-CoV-2 to free-ranging wildlife: a case study of bats. PLOS Pathog. (2020) 16:e1008758. doi: 10.1371/journal.ppat.1008758
298. CDC. Guidance to Reduce the Risk of SARS-CoV-2 Spreading between People and Wildlife. (2021). Available online at: https://www.cdc.gov/coronavirus/2019-ncov/your-health/wildlife.html (accessed March 17, 2021).
299. Oude Munnink BB, Sikkema RS, Nieuwenhuijse DF, Molenaar RJ, Munger E, Molenkamp R, et al. Jumping back and forth: anthropozoonotic and zoonotic transmission of SARS-CoV-2 on mink farms. bioRxiv [Preprint]. (2020). doi: 10.1101/2020.09.01.277152
300. USDA. USDA Confirms SARS-CoV-2 in Mink in Utah. (2020) Available online at: https://www.aphis.usda.gov/aphis/newsroom/stakeholder-info/sa_by_date/sa-2020/sa-08/sare-cov-2-mink
301. Lanen JVM, Stevens C, Brown CL, Maracle B, Koster DS. Subsistence Land Mammal Harvests and Uses, Yukon Flats, Alaska: 2008–2010 Harvest Report and Ethnographic Update. Alaska Department of Fish and Game, Division of Subsistence Technical Paper No. 377. Anchorage (2012).
302. Wurtzer S, Marechal V, Mouchel J-M, Maday Y, Teyssou R, Richard E, et al. Evaluation of lockdown impact on SARS-CoV-2 dynamics through viral genome quantification in Paris wastewaters. medRxiv [Preprint]. (2020). doi: 10.1101/2020.04.12.20062679
303. Lodder W, de Roda Husman AM. SARS-CoV-2 in wastewater: potential health risk, but also data source. Lancet Gastroenterol Hepatol. (2020) 5:533–4. doi: 10.1016/S2468-1253(20)30087-X
304. Carlson CJ, Albery GF, Merow C, Trisos CH, Zipfel CM, Eskew EA, et al. Climate change will drive novel cross-species viral transmission. bioRxiv [Preprint]. (2020). doi: 10.1101/2020.01.24.918755
305. Polley L, Thompson RCA. Parasite zoonoses and climate change: molecular tools for tracking shifting boundaries. Trends Parasitol. (2009) 25:285–91. doi: 10.1016/j.pt.2009.03.007
306. Reiling SJ, Dixon BR. Toxoplasma gondii: how an amazonian parasite became an inuit health issue. Can Commun Dis Rep. (2019) 45:183–90. doi: 10.14745/ccdr.v45i78a03
307. Prestrud KW, Åsbakk K, Fuglei E, Mørk T, Stien A, Ropstad E, Tryland M, et al. Serosurvey for Toxoplasma gondii in arctic foxes and possible sources of infection in the high Arctic of Svalbard. Vet Parasitol. (2007) 150:6–12. doi: 10.1016/j.vetpar.2007.09.006
308. Zhang Y-Z, Wu W-C, Shi M, Holmes EC. The diversity, evolution and origins of vertebrate RNA viruses. Curr Opin Virol. (2018) 31:9–16. doi: 10.1016/j.coviro.2018.07.017
309. Estrada-Peña A, Ostfeld RS, Peterson AT, Poulin R, de la Fuente J. Effects of environmental change on zoonotic disease risk: an ecological primer. Trends Parasitol. (2014) 30:205–14. doi: 10.1016/j.pt.2014.02.003
310. Shatilovich AV, Tchesunov AV, Neretina TV, Grabarnik IP, Gubin SV, Vishnivetskaya TA, et al. Viable nematodes from late pleistocene permafrost of the kolyma river lowland. Dokl Biol Sci. (2018) 480:100–2. doi: 10.1134/S0012496618030079
311. Legendre M, Bartoli J, Shmakova L, Jeudy S, Labadie K, Adrait A, et al. Thirty-thousand-year-old distant relative of giant icosahedral DNA viruses with a pandoravirus morphology. Proc Natl Acad Sci USA. (2014) 111:4274–9. doi: 10.1073/pnas.1320670111
312. Ng TFF, Chen L-F, Zhou Y, Shapiro B, Stiller M, Heintzman PD, et al. Preservation of viral genomes in 700-y-old caribou feces from a subarctic ice patch. Proc Natl Acad Sci USA. (2014) 111:16842–7. doi: 10.1073/pnas.1410429111
313. Rydberg J, Klaminder J, Rosén P, Bindler R. Climate driven release of carbon and mercury from permafrost mires increases mercury loading to sub-arctic lakes. Sci Total Environ. (2010) 408:4778–83. doi: 10.1016/j.scitotenv.2010.06.056
314. Erskine AJ. Birds In Boreal Canada: Communities, Densities And Adaptations. Canadian Wildlife Services Report. Ottawa, ON: Ministry of Supply and Services (1977).
315. Virkkala R, Rajasärkkä A. Climate change affects populations of northern birds in boreal protected areas. Biol Lett. (2011) 7:395–8. doi: 10.1098/rsbl.2010.1052
316. Post D, Taylor J, Kitchell J, Olson M, Schindler D, Herwig B. The role of migratory waterfowl as nutrient vectors in a managed wetland. Conserv Biol. (1998) 12:910–20.
317. Adalja AA, Watson M, Toner ES, Cicero A, Inglesby T V. The Characteristics of Pandemic Pathogens. Baltimore, MD: Johns Hopkins Bloomberg School of Public Health, Center for Health Security (2018).
318. King M, Altdorff D, Li P, Galagedara L, Holden J, Unc A. Northward shift of the agricultural climate zone under 21st-century global climate change. Sci Rep. (2018) 8:7904. doi: 10.1038/s41598-018-26321-8
319. Rohr JR, Barrett CB, Civitello DJ, Craft ME, Delius B, DeLeo GA, et al. Emerging human infectious diseases and the links to global food production. Nat Sustain. (2019) 2:445–56. doi: 10.1038/s41893-019-0293-3
320. Morand S. Emerging diseases, livestock expansion and biodiversity loss are positively related at global scale. Biol Conserv. (2020) 248:108707. doi: 10.1016/j.biocon.2020.108707
321. Charron DF. Potential impacts of global warming and climate change on the epidemiology of zoonotic diseases in Canada. Can J Public Health. (2002) 93:334–35. doi: 10.1007/BF03404563
322. Hannah L, Roehrdanz PR, Fraser EDG, Donatti CI, Saenz L, Wright TM, et al. The environmental consequences of climate-driven agricultural frontiers. PLoS ONE. (2020) 15:e0228305. doi: 10.1371/journal.pone.0228305
323. Coker R, Rushton J, Mounier-Jack S, Karimuribo E, Lutumba P, Kambarage D, et al. Towards a conceptual framework to support one-health research for policy on emerging zoonoses. Lancet Infect Dis. (2011) 11:326–31. doi: 10.1016/S1473-3099(10)70312-1
324. Liverani M, Waage J, Barnett T, Pfeiffer DU, Rushton J, Rudge JW, et al. Understanding and managing zoonotic risk in the new livestock industries. Environ Health Perspect. (2013) 121:873–7. doi: 10.1289/ehp.1206001
325. Sonne C, Letcher RJ, Jenssen BM, Desforges J-P, Eulaers I, Andersen-Ranberg E, et al. A veterinary perspective on one health in the arctic. Acta Vet Scand. (2017) 59:84. doi: 10.1186/s13028-017-0353-5
326. Hosseini PR, Mills JN, Prieur-Richard A-H, Ezenwa VO, Bailly X, Rizzoli A, et al. Does the impact of biodiversity differ between emerging and endemic pathogens? The need to separate the concepts of hazard and risk. Philos Trans R Soc B Biol Sci. (2017) 372:20160129. doi: 10.1098/rstb.2016.0129
327. Jones BA, Betson M, Pfeiffer DU. Eco-social processes influencing infectious disease emergence and spread. Parasitology. (2017) 144:26–36. doi: 10.1017/S0031182016001414
328. Canadian Population Health Initiative. Improving the Health of Canadians. Ottawa, ON: Canadian Population Health Initiative (2004).
329. Munster VJ, Bausch DG, de Wit E, Fischer R, Kobinger G, Muñoz-Fontela C, et al. Outbreaks in a rapidly changing central Africa — lessons from ebola. N Engl J Med. (2018) 379:1198–201. doi: 10.1056/NEJMp1807691
330. Bird BH, Mazet JAK. Detection of emerging zoonotic pathogens: an integrated one health approach. Annu Rev Anim Biosci. (2018) 6:121–39. doi: 10.1146/annurev-animal-030117-014628
331. Kutz S, Tomaselli M. “Two-eyed seeing” supports wildlife health. Science. (2019) 364:1135–7. doi: 10.1126/science.aau6170
332. Canadian Wildlife Health Cooperative. CWHC-RCSF - Canadian Wildlife Health Cooperative/Réseau Canadien Pour la Santé de la Faune. Available online at: http://www.cwhc-rcsf.ca/ (accessed October 26, 2020).
334. National Wildlife Health Center. USGS National Wildlife Health Center. Available online at: https://www.usgs.gov/centers/nwhc (accessed May 21, 2020).
335. Carlsson A, Veitch A, Popko R, Behrens S, Studies SKO. Monitoring wildlife health for conservation food security in the Canadian Arctic-a case study from the Sahtu settlment area in the Northwest Territories. In: Cork S, Hall D, Liljebjelke K, editors. One Health Case Studies: Addressing Complex Problems in a Changing World. Sheffield: 5m Publishing (2016).
336. McKay AJ, Johnson CJ. identifying effective and sustainable measures for community-based environmental monitoring. Environ Manage. (2017) 60:484–95. doi: 10.1007/s00267-017-0887-3
337. Mosites E, Lujan E, Brook M, Brubaker M, Roehl D, Tcheripanoff M, et al. Environmental observation, social media, and one health action: a description of the local environmental observer (LEO) network. One Health. (2018) 6:29–33. doi: 10.1016/j.onehlt.2018.10.002
338. Peacock SJ, Mavrot F, Tomaselli M, Hanke A, Fenton H, Nathoo R, et al. Linking co-monitoring to co-management: bringing together local, traditional, and scientific knowledge in a wildlife status assessment framework. Arct Sci. (2020) 6:247–66. doi: 10.1139/as-2019-0019
339. Huntington HP. Using traditional ecological knowledge in science: methods and applications. Ecol Appl. (2000) 10:1270–4. doi: 10.2307/2641282
340. Tagalik S. Inuit Qaujimajatuqangit: The Role of Indigenous Knowledge Supporting Wellness in Inuit Communities in Nunavut. British Columbia, CA: National Collaborating Center for Aboriginal Health, Prince George (2010).
341. Larrat S, Simard M, Lair S, Bélanger D, Proulx JF. From science to action and from action to science: the nunavik trichinellosis prevention program. Int J Circumpolar Health. (2012) 71:18595. doi: 10.3402/ijch.v71i0.18595
342. Stimmelmayr R, Sheffield G, Garlich-Miller J, Metcalf V, Goodwin J, Raverty S. The alaska Northern Pinniped unusual mortality event: 2011–2012. in Proceedings of the 44th Conference of the International Association for Aquatic Animal Medicine. Sausalito, CA (2012).
343. Indigenous Leadership Initiative. Indigenous Guardians Program – Indigenous Leadership Initiative. Available online at: https://www.ilinationhood.ca/our-work/guardians/ (accessed October 26, 2020).
344. Parlee BL, Goddard E, First Nation ŁKD, Smith M. Tracking change: traditional knowledge and monitoring of wildlife health in Northern Canada. Hum Dimens Wildl. (2014) 19:47–61. doi: 10.1080/10871209.2013.825823
345. Robards MD, Huntington HP, Druckenmiller M, Lefevre J, Moses SK, Stevenson Z, et al. Understanding and adapting to observed changes in the Alaskan Arctic: actionable knowledge co-production with alaska native communities. Deep Sea Res Part II Top Stud Oceanogr. (2018) 152:203–13. doi: 10.1016/j.dsr2.2018.02.008
346. Ostertag SK, Loseto LL, Snow K, Lam J, Hynes K, Gillman DV. “That's how we know they're healthy”: the inclusion of traditional ecological knowledge in beluga health monitoring in the Inuvialuit Settlement Region. Arct Sci. (2018) 4:292–320. doi: 10.1139/as-2017-0050
347. Moore SE, Hauser DDW. Marine mammal ecology and health: finding common ground between conventional science and indigenous knowledge to track arctic ecosystem variability. Environ Res Lett. (2019) 14:075001. doi: 10.1088/1748-9326/ab20d8
348. Thompson S, Thapa K, Whiteway N. Sacred harvest, sacred place: mapping harvesting sites in wasagamack first nation. J Agric Food Syst Community Dev. (2019) 9:1–29. doi: 10.5304/jafscd.2019.09B.017
349. National Research Council (US) Committee on Achieving Sustainable Global Capacity for Surveillance Response to Emerging Zoonotic Diseases. 3. Drivers of zoonotic diseases. In: Keusch GT, Pappaioanou M, Gonzalez MC, Scott KA, editors. Sustaining Global Surveillance Response to Emerging Zoonotic Diseases. Washing, DC: National Academies Press (2009).
350. LEO Network. About LEO Network - LEO Network. Available online at: https://www.leonetwork.org/en/docs/about/about (accessed October 26, 2020).
351. MacDonald JP, Ford JD, Willox AC, Ross NA. A review of protective factors and causal mechanisms that enhance the mental health of indigenous circumpolar youth. Int J Circumpolar Health. (2013) 72:21775. doi: 10.3402/ijch.v72i0.21775
352. O'Hara TM, Hoekstra PF, Hanns C, Backus SM, Muir DCG. Concentrations of selected persistent organochlorine contaminants in store-bought foods from Northern Alaska. Int J Circumpolar Health. (2005) 64:303–13. doi: 10.3402/ijch.v64i4.18008
353. DiNicolantonio JJ, O'Keefe JH. The introduction of refined carbohydrates in the Alaskan Inland inuit diet may have led to an increase in dental caries, hypertension and atherosclerosis. Open Hear. (2018) 5:e000776. doi: 10.1136/openhrt-2018-000776
354. Willows N, Johnson-Down L, Kenny T-A, Chan HM, Batal M. Modelling optimal diets for quality and cost: examples from inuit and first nations communities in Canada. Appl Physiol Nutr Metab. (2019) 44:696–703. doi: 10.1139/apnm-2018-0624
355. Kuhnlein H V, Chan HM. Environment and contaminants in traditional food systems of Northern indigenous peoples. Annu Rev Nutr. (2000) 20:595–626. doi: 10.1146/annurev.nutr.20.1.595
356. Chatwood S, Francois P, Baker GR, Eriksen AMA, Hansen KL, Eriksen H, et al. E. Indigenous values and health systems stewardship in circumpolar countries. Int J Env Res Public Heal. (2017) 14:1462. doi: 10.3390/ijerph14121462
357. Duhaime G, Bernard N. (eds). Arctic Food Security. Edmonton, AB: Canadian Circumpolar Institute Press (2008). p. 344. Available online at: https://www.researchgate.net/publication/276232632_DUHAIME_Gerard_and_Nick_BERNARD_eds_2008_Arctic_Food_Security_Edmonton_Canadian_Circumpolar_Institute_Press_344_pages (accessed April 13, 2021).
358. Ban NC, Frid A, Reid M, Edgar B, Shaw D, Siwallace P. Incorporate Indigenous perspectives for impactful research and effective management. Nat Ecol Evol. (2018) 2:1680–3. doi: 10.1038/s41559-018-0706-0
Keywords: wildlife, hunting, zoonotic, pandemic, Arctic, boreal, Indigenous, One Health
Citation: Keatts LO, Robards M, Olson SH, Hueffer K, Insley SJ, Joly DO, Kutz S, Lee DS, Chetkiewicz C-L, Lair S, Preston ND, Pruvot M, Ray JC, Reid D, Sleeman JM, Stimmelmayr R, Stephen C and Walzer C (2021) Implications of Zoonoses From Hunting and Use of Wildlife in North American Arctic and Boreal Biomes: Pandemic Potential, Monitoring, and Mitigation. Front. Public Health 9:627654. doi: 10.3389/fpubh.2021.627654
Received: 09 November 2020; Accepted: 30 March 2021;
Published: 05 May 2021.
Edited by:
Pierre Echaubard, SOAS University of London, United KingdomReviewed by:
Simon Rodrigo Rüegg, University of Zurich, SwitzerlandAlison J. Peel, Griffith University, Australia
Copyright © 2021 Keatts, Robards, Olson, Hueffer, Insley, Joly, Kutz, Lee, Chetkiewicz, Lair, Preston, Pruvot, Ray, Reid, Sleeman, Stimmelmayr, Stephen and Walzer. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lucy O. Keatts, bGtlYXR0c0B3Y3Mub3Jn