 Juan Luis Morales-Landa1
Juan Luis Morales-Landa1 Estefania Lazcano-Díaz2
Estefania Lazcano-Díaz2 Erandi Escamilla-García3
Erandi Escamilla-García3 Alan Gael Pérez-De la Rosa3
Alan Gael Pérez-De la Rosa3 Noé Luiz-Santos1*
Noé Luiz-Santos1*- 1Centro de Investigación y Asistencia en Tecnología y Diseño del Estado de Jalisco A.C. Subsede Noreste, Apodaca, Nuevo León, Mexico
- 2Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional Unidad Monterey, Apodaca, Nuevo León, Mexico
- 3Universidad Autónoma de Nuevo León, Facultad de Ciencias Biológicas, Instituto de Biotecnología, San Nicolás de los Garza, Nuevo León, Mexico
The growth behavior and lactic acid production capacity including both L-lactic and D-lactic acid isomers of five probiotic strains (Lactobacillus plantarum ATCC® 8014, L. rhamnosus ATCC® 53103, L. casei ATCC® 334, L. reuteri ATCC® 55730, and Bifidobacterium animalis ENCB-IPN) were systematically evaluated using defined carbon sources. Each strain was cultured for 24 h in modified MRS broth, where glucose was replaced with 1% (10 g/L) of individual substrates: monosaccharides (fructose, glycerol, rhamnose, arabinose, mannose, xylose, galactose), disaccharides (sucrose, cellobiose, lactose), a trisaccharide (raffinose), agave-derived fructooligosaccharides (FOS), and high-molecular-weight agave fructans (FAPM, DP > 10). Bacterial growth was monitored via optical density (OD595), and the specific growth rate (μ) and maximum OD were calculated. Fructose supported the highest total lactic acid production, particularly in L. plantarum and B. animalis, followed by sucrose, mannose, and cellobiose. Notably, L. rhamnosus achieved the highest L-lactic acid proportion (96%) with FAPM, while L. casei yielded 92.5% with cellobiose. In contrast, the highest D-lactic acid proportions were obtained with lactose (71.35% in L. plantarum, 55.88% in L. reuteri) and with glycerol in L. casei (62%). Principal component analysis (PCA) showed limited correlation between growth and acid production when monosaccharides were used, but strong positive correlations were found with disaccharides and FOS, particularly between μ and lactic acid yield. This study highlights the relevance of using defined carbon sources to elucidate the substrate-specific metabolic behavior of probiotic strains. The results provide a comparative framework for selecting or engineering strains for targeted lactic acid production (including optical purity) and lay the foundation for future bioprocess optimization using complex substrates such as agro-industrial residues or functional oligosaccharides.
1 Introduction
Lactic acid bacteria (LAB) are a group of microorganisms significant industrial relevance, particularly in the food and pharmaceutical sectors. They are widely used to produce fermented functional foods, enhancing the bioavailability of various bioactive compounds and contributing to the sensory and safety attributes of food products through their antimicrobial and biopreservative properties (Meruvu and Tellioglu Harsa, 2023; Evivie et al., 2017; Ayed et al., 2024). LAB naturally inhabits diverse ecological niches, including fermented foods, the oral cavity, and the gastrointestinal tract, where they promote host health by maintaining the balance of the intestinal microbiota. These attributes have led to their widespread use as probiotics (Abdel Tawab et al., 2023; Anjana and Tiwari, 2022; Bernal-Castro et al., 2024).
In recent years, the valorization of agro-industrial by-products for biotechnological applications has emerged as a sustainable alternative for waste management. These by-products, generated during food processing, are rich in essential nutrients and bioactive compounds, making them attractive substrates for the development of high-value-added products. Their use not only represents an economically viable alternative but also contributes to sustainability strategy for waste reduction and circular economy development (de la Rosa et al., 2019; Gonçalves et al., 2023). With global food waste estimated to exceed 1,052 million tons by 2022, contributing between 8 and 10% of global greenhouse gas emissions, innovative waste management strategies are urgently needed (Forbes et al., 2024).
LAB can produce lactic acid (LA) as the main product of carbohydrate fermentation. Agro-industrial residues containing lactose (from dairy) (Popova-Krumova et al., 2024; Turner et al., 2017), glycerol (from biodiesel) (Kano et al., 2022; Patel, 2012) fructans from agave (Martha-Lucero et al., 2025), and cellobiose (from cellulose hydrolysis) have shown promise as substrates for lactic acid production (Adsul et al., 2007), these options offer dual benefits: valorizing waste and supporting sustainable production processes.
Lactic acid is an organic acid recognized as GRAS (generally recognized as safe) by the FDA and has applications across multiple industries. In the food sector, it is used as a food preservative and sefety (Zapaśnik et al., 2022) and acidulant (Ameen and Caruso, 2017). In the chemical industry, it serves as a pH regulator (Zhou, 2023), solvent, cleaning agent (Kim et al., 2022), and precursor for biodegradable polymers like polylactic acid (PLA) (Banerjee et al., 2023; Swetha et al., 2023; de França et al., 2022). Additionally, it finds applications in cosmetics and pharmaceuticals, including skin treatments (Chuang et al., 2023), controlled drug delivery (Boisgard et al., 2017), and immunostimulants (Jiang et al., 2024).
Lactic acid exists as two optical isomers: L-lactic acid and D-lactic acid. In the context of biodegradable polymers, the mechanical properties and biodegradability of polylactic acid (PLA) are significantly influenced by the ratio of these enantiomers. Specifically, the D/L lactic acid ratio modulates the crystallinity, thermal stability, and degradation rate of the resulting polymer. Consequently, considerable research has been directed toward the selective production and purification of each isomer to meet the specific requirements of biopolymer applications (Augustiniene et al., 2022; Zhao et al., 2013). While chemical synthesis of lactic acid results in a racemic mixture, microbial fermentation offers a viable alternative for obtaining high-purity L-lactic acid. However, wild-type LAB strains, such as Lactobacillus plantarum, naturally produce a racemic mixture. Thus, it is critical to investigate the isomeric composition of lactic acid produced by various strains and substrates to optimize its industrial applicability.
Various species of Lactobacillus spp., including L. acidophilus, L. plantarum, L. casei, and L. rhamnosus, have been extensively applied in food biotechnology for the fermentation of diverse substrates, particularly in the development of symbiotic functional foods. Beyond their nutritional applications, these strains are gaining relevance in industrial and biomedical contexts due to their ability to synthesize bioactive metabolites, including lactic acid isomers, exopolysaccharides, and antioxidant compounds (Gao et al., 2025; Frumuzachi et al., 2024; Plessas et al., 2024; Palencia-Argel et al., 2024).
In addition to their recognized safety and probiotic status, certain LAB genera such as Lactiplantibacillus, Lacticaseibacillus, and Bifidobacterium exhibit notable antioxidant properties. These effects are mediated through the production of antioxidant enzymes, glutathione, and exopolysaccharides, which contribute to the mitigation of oxidative stress and the modulation of cellular signaling pathways such as Nrf2/NF-kB (Guerrero-Encinas et al., 2021; Ruiz-Gonzalez et al., 2024). These traits further support their suitability for health-oriented biotechnological applications.
To guide the rational selection of strains and conditions for targeted isomer production, the use of model substrates including monosaccharides, disaccharides, and oligosaccharides enables precise characterization of substrate preferences and fermentative capacities in LAB. This strategy is particularly relevant when designing processes that utilize complex carbon sources such as lignocellulosic hydrolysates or agro-industrial by-products rich in unconventional sugars.
Despite the extensive literature on glucose and lactose metabolism in LAB (Iskandar et al., 2019; Arefi et al., 2024; Wasewar et al., 2004), the fermentative behavior of probiotic strains in response to alternative carbon sources such as rhamnose, arabinose, agave-derived fructooligosaccharides (FOS), and high-degree polymerization fructans (FAMP) remains underexplored. These sugars are abundant in agave residues and other dietary fibers and offer great potential as functional substrates in sustainable fermentation processes.
Therefore, this study evaluates the growth performance and lactic acid isomer production (L- and D-lactic acid) of five probiotic strains cultured on selected model substrates derived from agro-industrial by-products. The results aim to support the development of efficient and sustainable bioprocesses aligned with circular economy principles.
2 Methodology
2.1 Probiotic bacterial strains, growth conditions and culture media
Five probiotic strains were evaluated, including four Lactobacillus species obtained from the American Type Culture Collection (ATCC®): Lacticaseibacillus casei ATCC® 334, Lactiplantibacillus plantarum ATCC® 8,014, Lactobacillus rhamnosus ATCC® 53,103, Lactobacillus reuteri ATCC® 55,730 (Limosilactobacillus reuteri), and Bifidobacterium animalis ENCB-IPN. The strains were propagated in 2% MRS-Glucose broth and agar (Man, Rogosa, Sharpe; BD Difco®, New Jersey, USA) at 37 ± 2 °C for 24 h in a HERATHERM bacteriological incubator (Thermo Scientific®) to confirm their viability and growth. The cultures were centrifuged at 6000 rpm for 5 min at 4 °C to obtain the bacterial pellet, which was washed twice with autoclave-sterilized 0.1% peptone water (pH 7.0 ± 0.1). The pellet was resuspended in 500 μL of sterile water and adjusted to an initial optical density of 0.125 at 595 nm (equivalent to 1 × 106 of CFU/mL, corresponding to tube 0.5 of the McFarland Nephelometer scale) using a Thermo Fisher Scientific™ Inc. Multiskan™ FC microplate photometer with incubator (Singapore).
The carbohydrate substrates included eight monosaccharides—glucose (GLU), fructose (FRU), maltose (MAL), glycerol (GLY), rhamnose (RHA), arabinose (ARA), mannose (MAN), xylose (XYL), and galactose (GAL); three disaccharides—cellobiose (CEL), lactose (LAC), and sucrose (SUC); and one trisaccharide—raffinose (RAF), all purchased from Merck (Darmstadt, Germany). Additionally, agave-derived fructooligosaccharides (FOS) and high-molecular-weight fructans (FAPM), obtained using membrane separation technologies (Luiz-Santos et al., 2020, 2022), were included in the analysis.
2.2 Growth behavior of the strains with different carbohydrates
Growth curves of L. paracasei, L. plantarum, L. rhamnosus, L. reuteri, and B. animalis were obtained in MRS broth modified to contain 1% (w/v) of the respective carbohydrate as the sole carbon source. Incubations were performed in 96-well plates at 37 ± 2 °C, with 200 μL fermentation volumes and internal shaking, using a Multiskan™ FC photometer (Thermo Fisher Scientific™, Singapore).
Substrate assimilation capacity was assessed by recording the optical density (OD) at 595 nm, a wavelength commonly used for turbidity-based growth monitoring in Lactobacillus and Bifidobacterium species period (Shiroda and Manning, 2020; Reale et al., 2015; Moya-Gonzálvez et al., 2021; Zhang et al., 2023). Measurements were taken every hour over a 24-h where time zero corresponded to cultures freshly inoculated with cells in exponential growth phase. Specific growth rates (μ) were estimated by plotting the natural logarithm of OD versus time. All experiments were conducted in triplicate.
2.3 Determination of production of lactic acid
Following incubation, cultures were centrifuged at 11,000 rpm for 10 min at 4 °C to remove cells. The supernatants were collected and stored in 1.5 mL Eppendorf tubes at −20 °C for subsequent analysis.
The concentrations of L-lactic acid and D-lactic acid were determined enzymatically using specific reagent kits from BioSystems (L-lactic acid, COD 12802; D-lactic acid, COD 12801). Both assays are based on the enzymatic oxidation of L- or D-lactate to pyruvate by L-lactate dehydrogenase (L-LDH) or D-lactate dehydrogenase (D-LDH), respectively, coupled to the reduction of NAD+ to NADH (Equation 1), which was quantified spectrophotometrically at 340 nm.
For each assay, 240 μL of Reagent A (enzyme solution in alkaline buffer, pH 9.5) and 3–15 μL of sample were incubated at 37 °C and subsequently mixed with 60–200 μL of Reagent B (NAD+). Reaction kinetics were monitored at 340 nm using a differential bi-reagent reading mode, which involves two absorbance measurements: an initial reading (A1) after the addition of Reagent A, and a second reading (A2) after the addition of Reagent B and incubation. This differential kinetic approach corrects for background absorbance and enables the accurate quantification of NADH formation, which is proportional to lactic acid concentration. Time points for absorbance readings were set at 72 s and 480 s for L-lactate, and 72 s and 408 s for D-lactate.
Calibration was performed using a multilevel aqueous standard solution. Results were expressed in g/L, and the system was configured to detect concentrations within the linear ranges of 0.02–3.00 g/L for L-lactate and 0.004–0.250 g/L for D-lactate. Coefficients of variation (CV) for repeatability and total imprecision were below 3.8% for L-lactate and below 1.5% for D-lactate, indicating good analytical performance.
2.4 Statistical analysis
The effects of carbohydrate type (monosaccharides, disaccharides, trisaccharides, and polysaccharides) and probiotic strain on lactic acid production were assessed using a multifactor categorical design.
L- and D-lactic acid production was compared using one-way ANOVA followed by Tukey’s post hoc test at a 95% confidence level (p < 0.05). All statistical analyses were performed using Statgraphics Centurion XVI. Principal component analysis (PCA) was conducted using XLSTAT to evaluate the correlation between kinetic growth parameters and lactic acid production.
3 Results
3.1 Growth behavior of probiotic strains with different carbohydrates
To illustrate the growth behavior of the tested probiotic strains, Figure 1 presents the growth curves of L. paracasei, L. plantarum, L. rhamnosus, L. reuteri, and B. animalis over a 24-h incubation period, measured as optical density at 595 nm, using various monosaccharides as sole carbon sources. Growth patterns varied notably among strains depending on the type of monosaccharide provided.
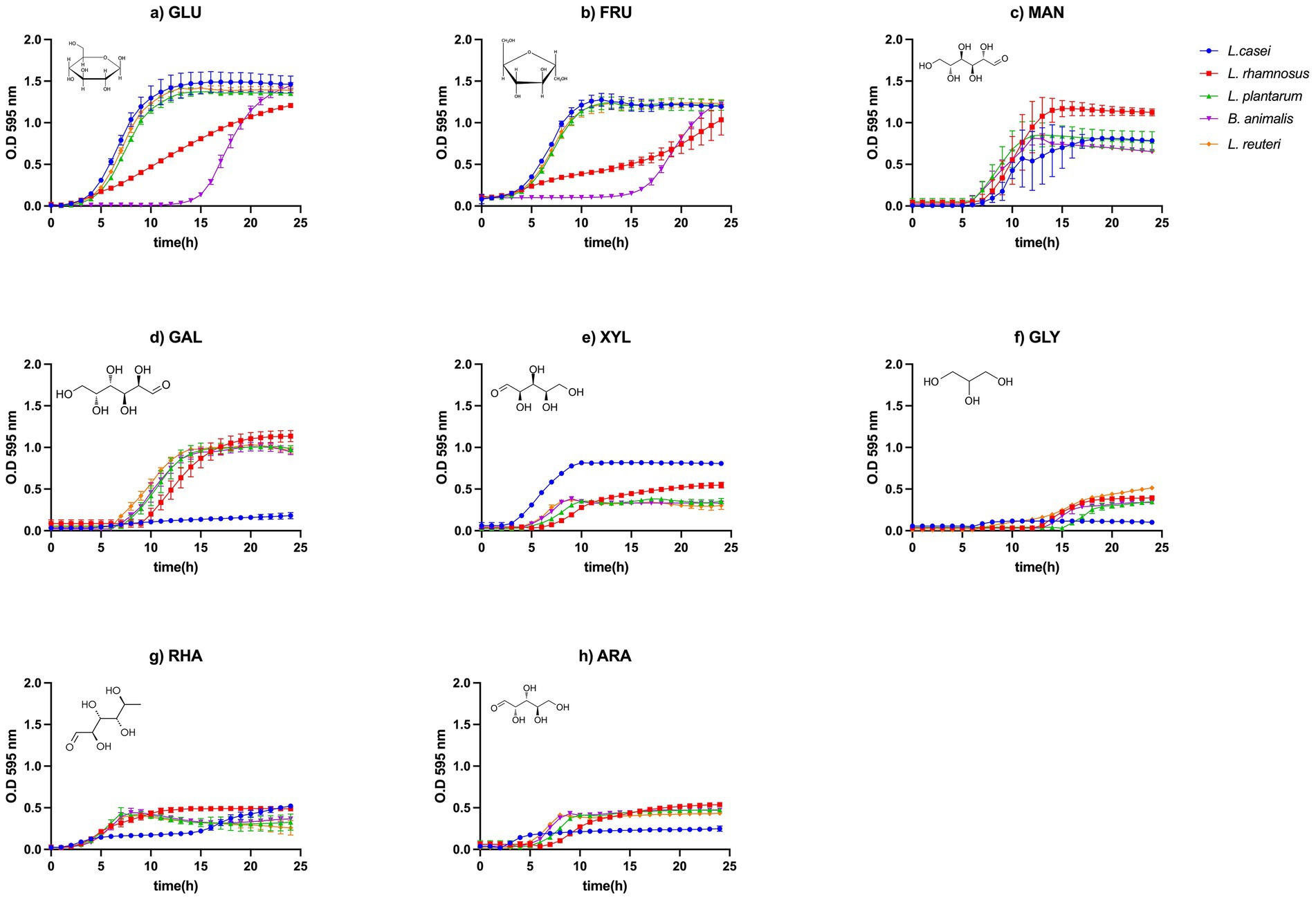
Figure 1. Growth curves of probiotic bacteria using monosaccharides as the sole carbon source: (a) Glucose (GLU), (b) Fructose (FRU), (c) Mannose (MAN), (d) Galactose (GAL), (e) Xylose (XYL), (f) Glycerol (GLY), (g) Rhamnose (RHA), and (h) Arabinose (ARA).
GLU, FRU, and MAN supported robust growth in all strains, with OD595 values approaching 1.5–2.0. XYL yielded moderate growth across most strains, while GLY, RHA, and ARA were the least effective substrates, resulting in significantly lower biomass accumulation.
When evaluating growth on disaccharides, SUC and CEL (Figures 2a,b) emerged as the most favourable substrates for the majority of strains. LAC also supported bacterial growth, although strain-dependent differences were observed. B. animalis and L. reuteri demonstrated the highest efficiency with LAC (Figure 2c), achieving OD595 values close to 1.5–2.0.
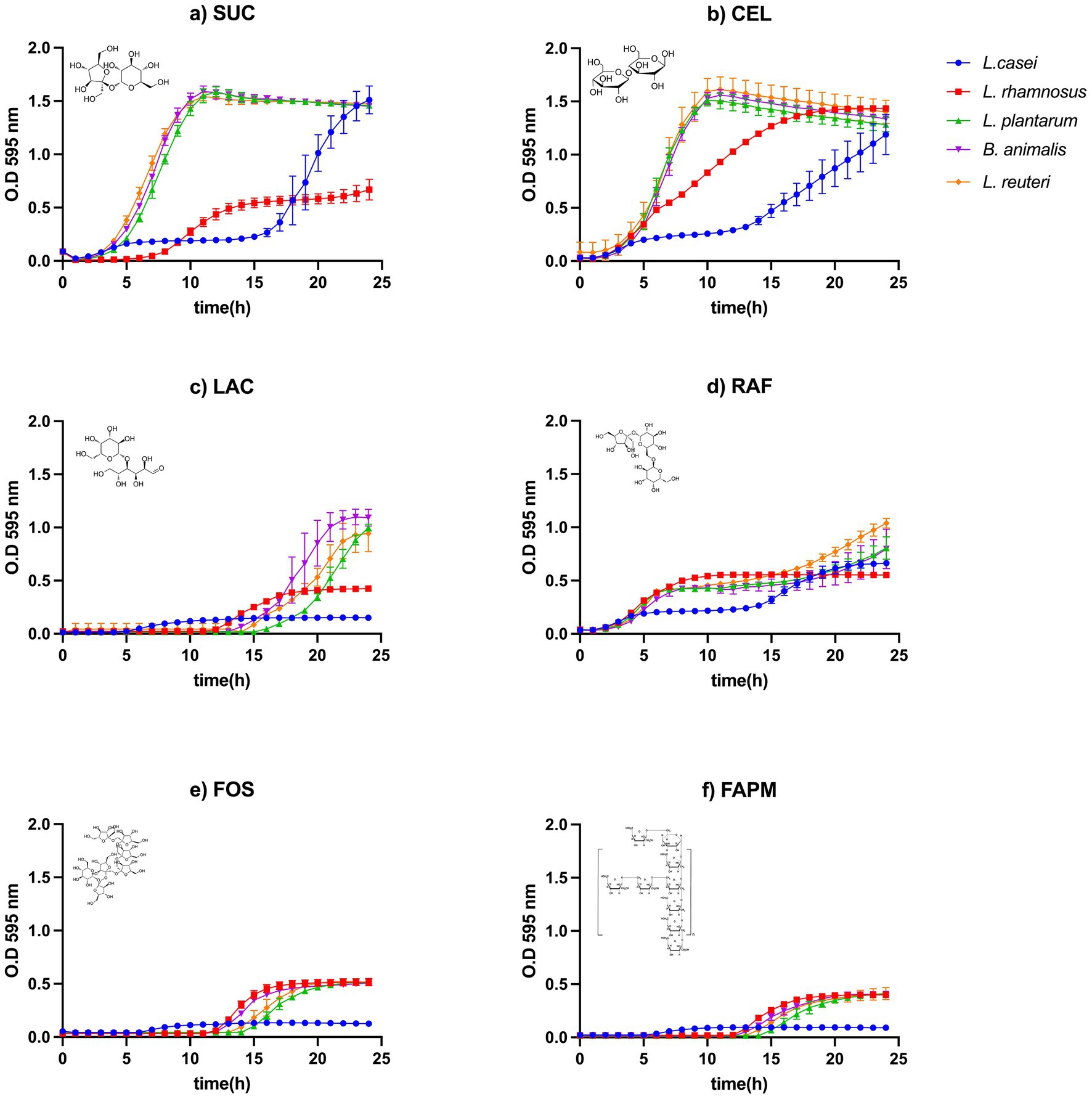
Figure 2. Growth curves of probiotic bacteria using disaccharides (a–c), trisaccharide (d), fructooligosaccharide (e), and polysaccharides (f) as carbon sources.
Fructooligosaccharides (FOS) supported limited growth compared to most monosaccharides and disaccharides, with the exception of rhamnose (RHA), glycerol (GLY), and arabinose (ARA), indicating that not all strains are capable of efficiently fermenting this carbohydrate. The maximum optical density observed was approximately 0.5–0.75 (Figure 2d), reflecting reduced substrate utilization. A similar growth pattern was observed with high-molecular-weight agave fructans (FAPM), which also showed minimal growth among the evaluated substrates (Figures 2e,f). This behavior was expected due to the structural complexity of these molecules. Such growth limitations for both FOS and FAPM have been previously reported (Morales-Landa et al., 2024; Martha-Lucero et al., 2025), showing lower productivity compared to glucose. However, a major advantage of fructans is that they do not require prior hydrolysis or pretreatment for fermentation.
To identify statistically significant factors influencing lactic acid production (Supplementary Table S1), a two-way analysis of variance (ANOVA) was conducted at a 95% confidence level (Figure 3). Both main effects (bacterial strain and carbohydrate source) and their interaction were found to be statistically significant (p < 0.05).
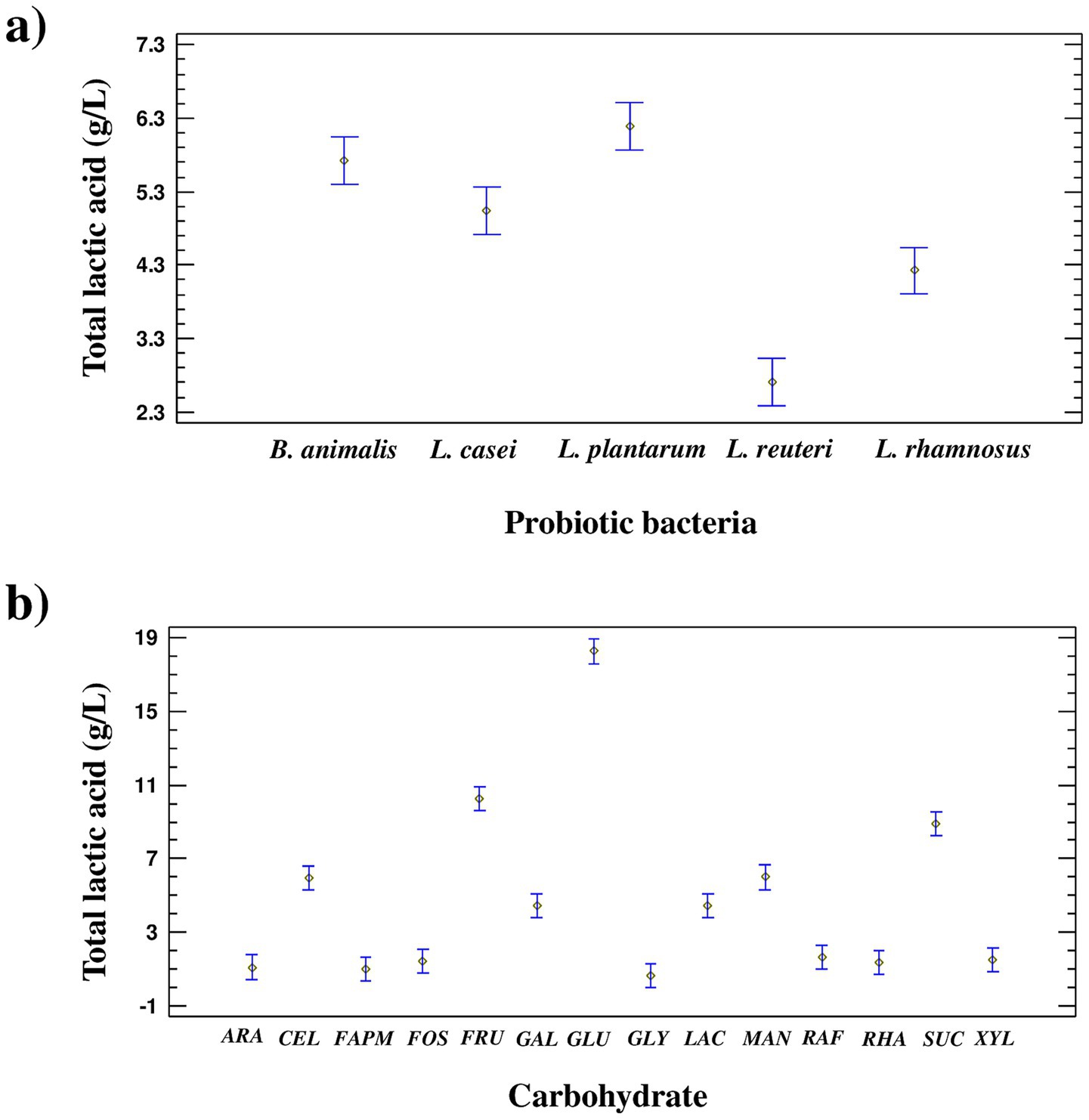
Figure 3. Analysis of variance (ANOVA) and Tukey’s test for the effects of (a) probiotic bacterial strain and (b) carbohydrate type on lactic acid production.
According to the mean comparison analysis, the highest lactic acid yields were obtained with L. plantarum and B. animalis. Among the substrates, the order of effectiveness was GLU > FRU > SUC > MAN ≈ CEL > LAC ≈ GAL. No significant differences were detected for RAF, XYL, FOS, RHA, ARA, FAPM, and GLY, which exhibited the lowest lactic acid production.
Multivariate relationships among growth performance, metabolite production, and substrate type were further explored through principal component analysis (PCA). Figure 4 presents three biplots derived from PCA, representing the relationships among maximum optical density (OD595), lactic acid production, and specific growth rate (μ) across different carbohydrate substrates. The primary axes, F1 and F2, represent the first and second principal components and indicate the percentage of variance explained. Blue dots correspond to different bacteria–substrate combinations, while red vectors represent the direction and magnitude of variables such as specific growth rate, doubling time, maximum OD, and total lactic acid. In Figure 4a, F1 and F2 account for 89.46% of the total variance (65.23 and 24.23%, respectively). F1 is primarily associated with specific growth rate and doubling time, whereas F2 correlates with lactic acid production and maximum OD. Specific growth rate and doubling time are positively correlated in strains such as L. plantarum with GAL, L. casei with MAN, and B. animalis with MAN. Conversely, maximum OD appears negatively correlated with these variables, particularly for L. casei grown on GAL, RHA, and GLY. A positive association between lactic acid production and growth was observed when GLU was used by L. casei, L. reuteri, and L. plantarum.
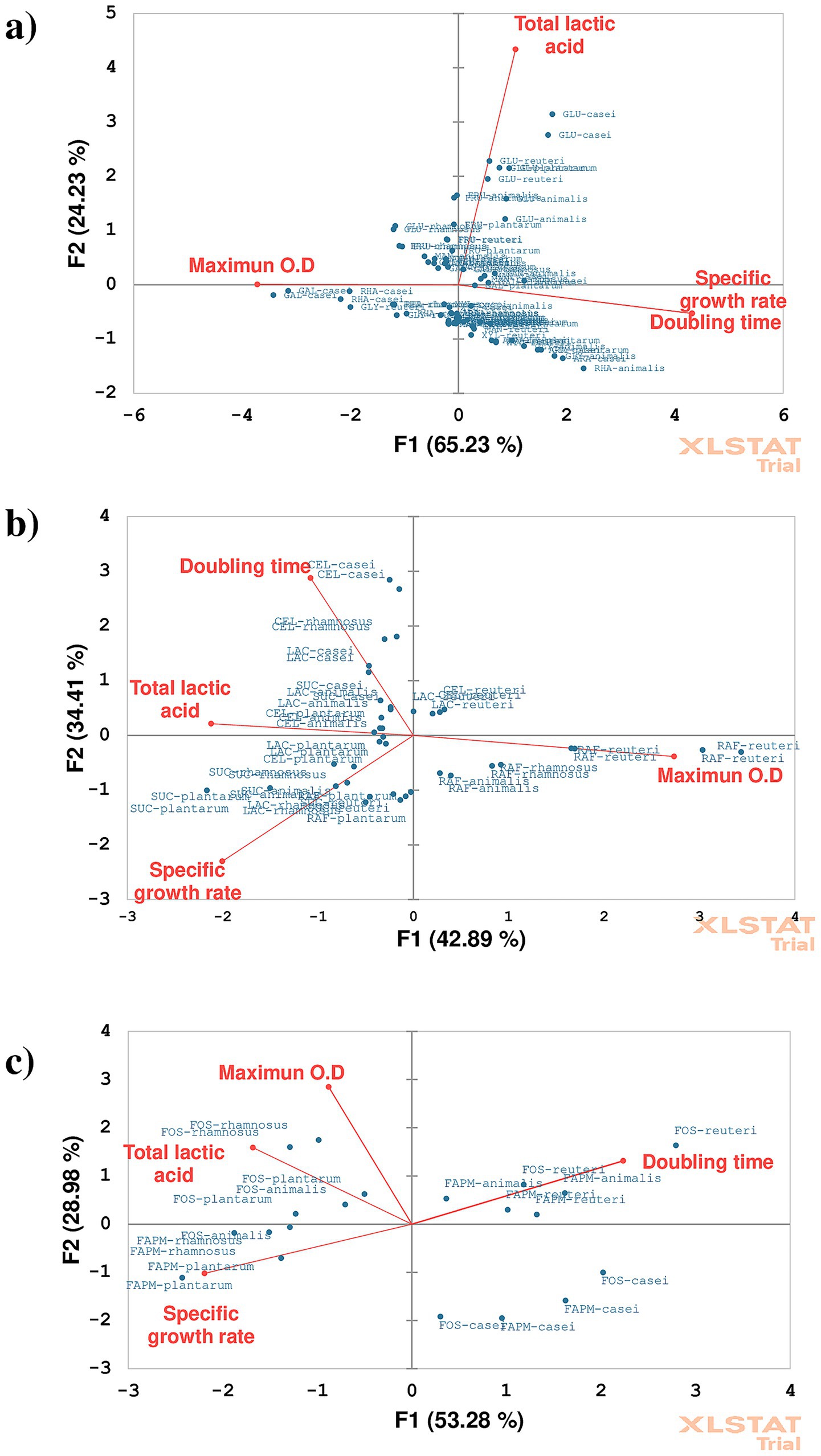
Figure 4. Biplots of principal components (PC1 and PC2) illustrate the correlations between growth and production parameters of probiotic bacteria cultivated on (a) monosaccharides, (b) disaccharides, and (c) polysaccharides as carbon sources.
In Figure 4b, PCA of disaccharides (SUC, CEL, LAC) and RAF reveals that F1 and F2 explain 42.89 and 34.41% of the variance, respectively. The highest OD was observed for L. reuteri and L. rhamnosus cultured on RAF. However, increased OD did not always correlate with higher lactic acid production. Similarly, a shorter doubling time did not consistently correspond with a higher specific growth rate. Nevertheless, a positive correlation between lactic acid production and specific growth rate was evident, suggesting that these parameters increase in tandem.
The PCA for polysaccharides shows that F1 and F2 explain 53.28 and 28.98% of the total variance, respectively (Figure 4c). A clear distinction was observed between bacterial growth on FOS and FAPM, indicating metabolic differences in substrate utilization. Higher lactic acid production was more closely associated with specific growth rate and maximum OD when FOS was used as the carbon source.
The distribution of L- and D-lactic acid production according to the influence of the type of carbon substrate and the probiotic bacterial strain is shown in Figure 5. Overall, no statistically significant differences were observed in the L-/D-lactic acid ratios across all substrates for each bacterium. However, certain combinations exhibited a strong preference for L-lactic acid production. Notably, L. rhamnosus grown on FAPM achieved the highest L-lactic acid proportion at 96%, followed by L. casei with cellobiose at 92.5%. Additionally, B. animalis grown on glucose produced 90.38% L-lactic acid, and lactose fermentation by L. casei resulted in 79.78% L-lactic acid. In contrast, the highest D-lactic acid proportions were observed with lactose as the substrate, reaching 71.35% for L. plantarum and 55.88% for L. reuteri. Furthermore, L. casei produced 62% D-lactic acid when utilizing GLY as the carbon source (Supplementary Table S2).
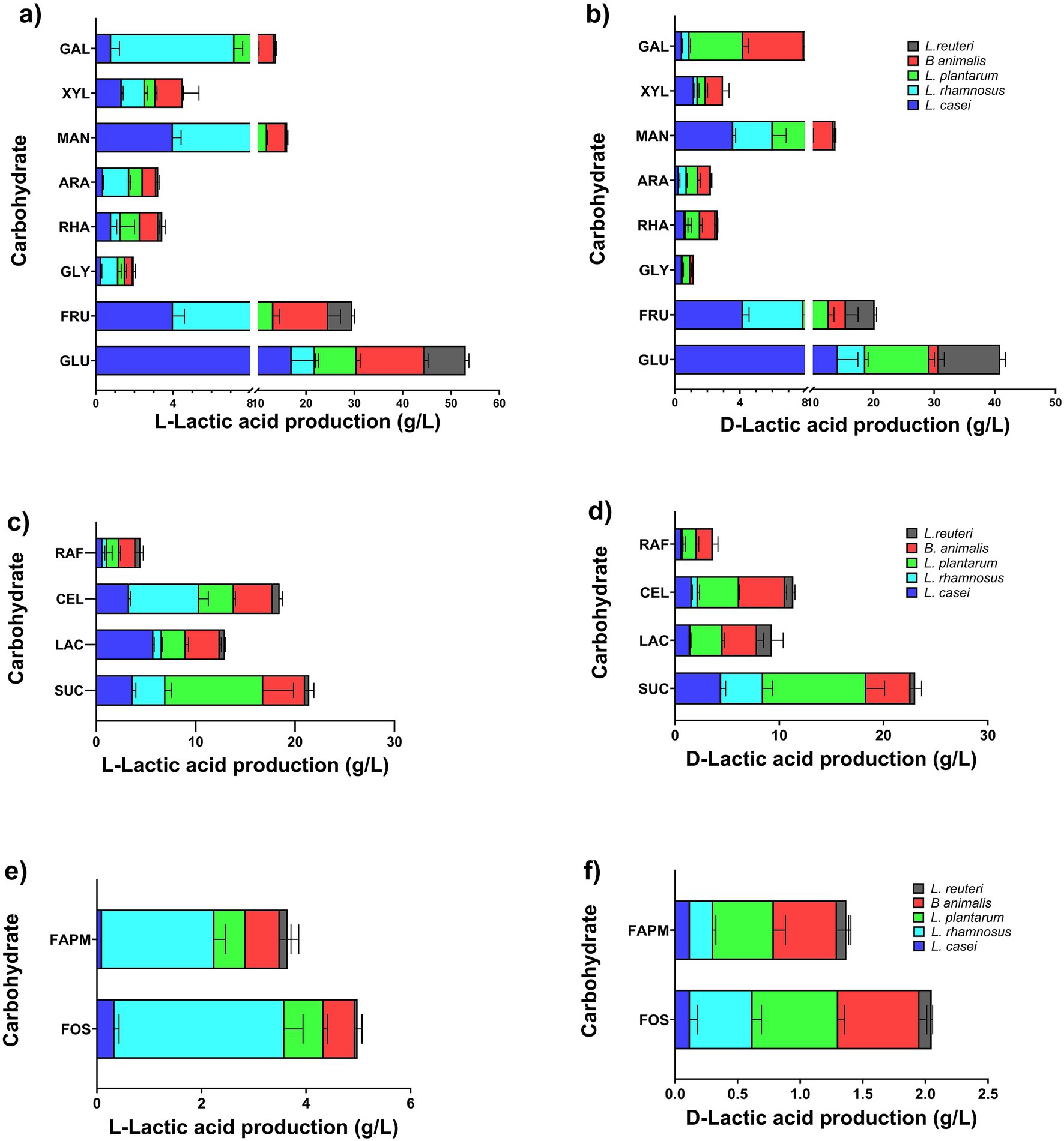
Figure 5. Production of L- and D-lactic acid using monosaccharides (a,b), disaccharides and trisaccharides (c,d), fructooligosaccharides (FOS), high molecular weight agave fructans (FAPM) (e,f).
4 Discussion
Lactic acid bacteria (LAB) can produce lactic acid as a primary metabolite through the fermentation of hexoses. Homofermentative LAB uses glycolysis (Embden-Meyerhof-Parnas pathway) to convert a single hexose molecule into two molecules of lactic acid, generating two ATP molecules while maintaining a balanced NAD+/NADH redox state. This could explain the higher lactic acid production observed in L. casei (Cui and Qu, 2021), L. plantarum (Gänzle, 2015), and B. animalis (Cui et al., 2019).
In contrast, heterofermentative LAB such as Lactobacillus reuteri (Årsköld et al., 2008) and L. rhamnosus (Morita et al., 2009) utilise the pentose phosphate pathway due to the absence of fructose-1,6-bisphosphate aldolase. As a result, these strains produce not only lactic acid but also CO₂, ATP, and either ethanol or acetic acid per glucose molecule. In this context, the conversion of acetyl phosphate to ethanol requires the oxidation of two NADH molecules, maintaining redox balance. Conversely, the formation of acetic acid allows for the regeneration of NAD+ and the production of an additional ATP molecule.
Our findings suggest that lactic acid isomer production (L- or D-form) is influenced by the bacterial strain and the type of substrate fermented. Certain strain-substrate combinations favour specific metabolic pathways that result in high-purity L-lactic acid, while others promote D-lactic acid production. On the other hand, the low performance in the proliferation or growth of some heterofermentative LAB, such as L. reuteri, may be due not only to the metabolic implications of assimilating a complex carbohydrate source, but also to the acidic characteristics of the lactate produced by LAB that may act as growth inhibitor, like the case studied in the strain L. reuteri JCM1112 (Ichinose et al., 2020).
Lactic acid is an important bio-based platform compound, which can be divided into D-lactic acid and L-lactic acid according to its optical rotation (Wang et al., 2021). The high proportion of L-lactic acid obtained from L. rhamnosus fermenting agave fructans, L, casei with cellobiose and lactose, and B. animalis with glucose, may be attributed to two key factors: (1) enzyme specificity and (2) metabolic regulation, indicating dominant L-lactate dehydrogenase (L-LDH) activity under favourable substrate conditions. Under these circumstances, the activity or expression of D-lactate dehydrogenase (D-LDH) is reduced, leading to the near-exclusive production of L-lactic acid.
Metabolic regulation may also dictate the dominance of homolactic versus heterolactic fermentation depending on the substrate. In L. rhamnosus, L. casei, and B. animalis, homolactic fermentation is prevalent (Cui et al., 2019; Gänzle, 2015; Cui and Qu, 2021), meaning most pyruvate is channelled toward L-lactic acid formation rather than alternative products such as ethanol or acetic acid.
Agave fructans are fructose-based polymers that must first be hydrolyzed into fructooligosaccharides (FOS), free fructose, and glucose for microbial utilization (Morales-Landa et al., 2024; Cui et al., 2021; Biedrzycka and Bielecka, 2004). L. rhamnosus and L. casei possess fructofuranosidase or β-fructosidase enzymes capable of breaking down these polymers. The resulting monosaccharides enter glycolysis via the fructose-6-phosphate (F6P) pathway, bypassing the metabolic branching of the pentose phosphate pathway and favouring L-lactic acid production. Similarly, cellobiose, a disaccharide composed of two glucose units linked by a β-1,4 bond, is hydrolyzed by β-glucosidase in L. casei, enabling glucose entry into glycolysis via glucose-6-phosphate (G6P) and promoting rapid L-lactate formation.
In B. animalis, glucose metabolism primarily follows the bifid shunt, also known as the fructose-6-phosphate phosphoketolase pathway, which is unique to the genus Bifidobacterium. In this route, fructose-6-phosphate is cleaved by F6PPK into acetyl phosphate and erythrose-4-phosphate. The conversion of acetyl phosphate to acetate or lactate depends on NADH availability and L-LDH activity. Under conditions of high NADH regeneration such as during glucose fermentation, B. animalis favors lactate over acetate production, improving conversion efficiency (González-Rodríguez et al., 2013; Pokusaeva et al., 2011).
In contrast, L. reuteri and L. plantarum appear to exhibit greater D-LDH activity when fermenting lactose, favoring D-lactic acid production (Morita et al., 2008; Goffin et al., 2004). This behavior may result from differences in LDH gene regulation or intracellular conditions such as NAD+/NADH ratios and carbon flux. These factors can shift the metabolic equilibrium toward D-lactic acid synthesis.
The L and D forms of lactic acid produced by LAB fermentation play a significant role in the synthesis of biodegradable polylactic acid. There is a great interest in the biomedical area in the fields of tissue regeneration and repair, in the application of biomaterial technologies (Capuana et al., 2022; Alavi et al., 2023), in the cosmetic area for skin care and treatment of melanogenesis (Huang et al., 2020), to mention a few examples.
Overall, these findings highlight the metabolic complexity of lactic acid biosynthesis in probiotic strains and underscore the importance of strain-substrate selection in directing fermentation toward high-purity L-lactic acid.
5 Conclusion
The growth behavior and lactic acid production of probiotic bacteria are strongly influenced by the type of substrate used. Glucose, fructose, and disaccharides such as cellobiose and sucrose are the most efficient in promoting both bacterial growth and lactic acid synthesis. Furthermore, enzymatic specificity and metabolic regulation play critical roles in determining the L-/D-lactic acid ratio. Notably, L. rhamnosus, L. casei, and B. animalis show a clear preference for L-lactic acid production. These results provide valuable insight into fermentation processes through strategic selection of both LAB strains and carbon sources, with implications for food biotechnology but for the medical field. Our findings ensure a follow-up in the study with substrates originating from by-products of the food industry, aimed at promising uses and biomedical applications, thus promoting the utilisation of these sources, which are important actions in the circular economy.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
JM-L: Conceptualization, Formal analysis, Funding acquisition, Methodology, Supervision, Writing – original draft, Writing – review & editing. EL-D: Formal analysis, Funding acquisition, Investigation, Visualization, Writing – original draft, Writing – review & editing. EE-G: Conceptualization, Funding acquisition, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing. AP-D: Data curation, Investigation, Methodology, Writing – original draft, Writing – review & editing. NL-S: Conceptualization, Data curation, Formal analysis, Funding acquisition, Project administration, Supervision, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was supported by I2T2-RETOS-2024-010: Proceso de pasteurización en frío y reducción de carbohidratos con tecnología de membranas en alimentos: caso leche bronca deslactosada, RETOS de Innovación PIIT 2024, Instituto de Innovación y Transferencia de Tecnología (I2T2) - Gobierno del Estado de Nuevo León.
Acknowledgments
The authors acknowledge L.Q. Carlos Eduardo Muñiz Cervantes, Business Development Manager at BioSystems México (https://biosystems.global/es), for his technical assistance with the use of the Y15 Automatic Analyzer (Y211bml6QGJpb3N5c3RlbXMuY29tLm14). They also thank Dr. Oscar Rodolfo Rodas Suárez, research professor at the Escuela Nacional de Ciencias Biológicas (ENCB) – Instituto Politécnico Nacional (IPN) (https://www.encb.ipn.mx/), for providing the Bifidobacterium animalis ENCB-IPN strain for research purposes (b3JvZGFzc0BpcG4ubXg=).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that Gen AI was used in the creation of this manuscript. The authors used an AI language model (ChatGPT, OpenAI) to assist with English language editing, including improvements in grammar, structure, and clarity. All content was critically reviewed, verified, and approved by the authors.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsufs.2025.1613809/full#supplementary-material
References
Abdel Tawab, F. I., Abd Elkadr, M. H., Sultan, A. M., Hamed, E. O., El-Zayat, A. S., and Ahmed, M. N. (2023). Probiotic potentials of lactic acid Bacteria isolated from Egyptian fermented food. Sci. Rep. 13:16601. doi: 10.1038/s41598-023-43752-0
Adsul, M., Khire, J., Bastawde, K., and Gokhale, D. (2007). Production of lactic acid from Cellobiose and Cellotriose by Lactobacillus Delbrueckii mutant Uc-3. Appl. Environ. Microbiol. 73, 5055–5057. doi: 10.1128/AEM.00774-07
Alavi, M. S., Memarpour, S., Pazhohan-Nezhad, H., Salimi Asl, A., Moghbeli, M., Shadmanfar, S., et al. (2023). Applications of poly(lactic acid) in bone tissue engineering: a review article. Artif. Organs 47, 1423–1430. doi: 10.1111/aor.14612
Ameen, S. M., and Caruso, G.. (2017). Lactic acid in the food industry. Cambridge, MA: Academic Press.
Anjana,, and Tiwari, S. K. (2022). Bacteriocin-producing probiotic lactic acid Bacteria in controlling Dysbiosis of the gut microbiota. Front. Cell. Infect. Microbiol. 12:851140. doi: 10.3389/fcimb.2022.851140
Arefi, A., Sturm, B., Babor, M., Horf, M., Hoffmann, T., Höhne, M., et al. (2024). Digital model of biochemical reactions in lactic acid bacterial fermentation of simple glucose and biowaste substrates. Heliyon 10:e38791. doi: 10.1016/j.heliyon.2024.e38791
Årsköld, E., Lohmeier-Vogel, E., Cao, R., Roos, S., Rådström, P., and Van Niel, E. W. J. (2008). Phosphoketolase pathway dominates in Lactobacillus Reuteri ATCC 55730 containing dual pathways for glycolysis. J. Bacteriol. 190, 206–212. doi: 10.1128/JB.01227-07
Augustiniene, E., Valanciene, E., Matulis, P., Syrpas, M., Jonuskiene, I., and Malys, N. (2022). Bioproduction of l- and d-lactic acids: advances and trends in microbial strain application and engineering. Crit Rev Biotechnol. 42, 342–360. doi: 10.1080/07388551.2021.1940088
Ayed, L., M’hir, S., Nuzzolese, D., Di Cagno, R., and Filannino, P. (2024). Harnessing the health and techno-functional potential of lactic acid bacteria: a comprehensive review. Foods 13:1538. doi: 10.3390/foods13101538
Banerjee, A., Jha, K., Petru, M., Kumar, R., Sharma, S., Saini, M. S., et al. (2023). Fabrication and characterization of weld attributes in hot gas welding of alkali treated hybrid flax Fiber and pine cone fibers reinforced poly-lactic acid (PLA) based biodegradable polymer composites: studies on mechanical and morphological properties. J. Mater. Res. Technol. 27, 272–297. doi: 10.1016/j.jmrt.2023.09.252
Bernal-Castro, C., Espinosa-Poveda, E., Gutiérrez-Cortés, C., and Díaz-Moreno, C. (2024). Vegetable substrates as an alternative for the inclusion of lactic acid Bacteria with probiotic potential in food matrices. J. Food Sci. Technol. 61, 833–846. doi: 10.1007/s13197-023-05779-z
Biedrzycka, E., and Bielecka, M. (2004). Prebiotic effectiveness of Fructans of different degrees of polymerization. Trends Food Sci. Technol. 15, 170–175. doi: 10.1016/j.tifs.2003.09.014
Boisgard, A. S., Lamrayah, M., Dzikowski, M., Salmon, D., Kirilov, P., Primard, C., et al. (2017). Innovative drug vehicle for local treatment of inflammatory skin diseases: ex vivo and in vivo screening of five topical formulations containing poly(lactic acid) (PLA) nanoparticles. Eur. J. Pharm. Biopharm. 116, 51–60. doi: 10.1016/j.ejpb.2016.09.021
Capuana, E., Lopresti, F., Ceraulo, M., and La Carrubba, V. (2022). Poly-L-lactic acid (PLLA)-based biomaterials for regenerative medicine: a review on processing and applications. Polymers 14:1153. doi: 10.3390/polym14061153
Chuang, F. J., Wang, Y. W., Chang, L. R., Chang, C. Y., Cheng, H. Y., and Kuo, S. M. (2023). Enhanced skin Neocollagenesis through the transdermal delivery of poly-L-lactic acid microparticles by using a needle-free supersonic atomizer. Biomater. Adv. 154:213619. doi: 10.1016/j.bioadv.2023.213619
Cui, S., Sadiq, F. A., Mao, B., Xu, Z., Liu, Z., Zhao, J., et al. (2019). High-density cultivation of Lactobacillus and Bifidobacterium using an automatic feedback feeding method. LWT 112:108232. doi: 10.1016/j.lwt.2019.05.130
Cui, Y., Wang, M., Zheng, Y., Miao, K., and Qu, X. (2021). The carbohydrate metabolism of Lactiplantibacillus plantarum. Int. J. Mol. Sci. 22:13452. doi: 10.3390/ijms222413452
Cui, Y., and Qu, X. (2021). Genetic mechanisms of prebiotic carbohydrate metabolism in lactic acid bacteria: emphasis on Lacticaseibacillus casei and Lacticaseibacillus paracasei as flexible, diverse and outstanding prebiotic carbohydrate starters. Trends Food Sci. Technol. 115, 486–499. doi: 10.1016/j.tifs.2021.06.058
França, J. O. C.de, Valadares, D. da S., Paiva, M. F., Dias, S. C. L., and Dias, J. A. (2022). “Polymers based on PLA from synthesis using D,L-lactic acid (or racemic Lactide) and some biomedical applications: a short review” Polymers 14:2317. doi: 10.3390/polym14122317.
de la Rosa, O., Flores-Gallegos, A. C., Muñíz-Marquez, D., Nobre, C., Contreras-Esquivel, J. C., and Aguilar, C. N. (2019). Fructooligosaccharides production from agro-wastes as alternative low-cost source. Trends Food Sci. Technol. 91, 139–146. doi: 10.1016/j.tifs.2019.06.013
Evivie, S. E., Huo, G. C., Igene, J. O., and Bian, X. (2017). Some current applications, limitations and future perspectives of lactic acid Bacteria as probiotics. Food Nutr. Res. 61:1318034. doi: 10.1080/16546628.2017.1318034
Forbes, H., Peacock, E, Abbot, N, Jones, M, and O’Connor, C. (2024). “Think eat save tracking progress to halve global food waste, waste food waste index report 2024.” Available online at: https://www.unep.org/resources/publication/food-waste-index-report-2024
Frumuzachi, O., Rohn, S., and Mocan, A. (2024). Fermented black chokeberry (Aronia Melanocarpa (Michx.) Elliott) products—a systematic review on the composition and current scientific evidence of possible health benefits. Food Res. Int. 196:115094. doi: 10.1016/j.foodres.2024.115094
Gänzle, M. G. (2015). Lactic metabolism revisited: metabolism of lactic acid Bacteria in food fermentations and food spoilage. Curr. Opin. Food Sci. 2, 106–117. doi: 10.1016/j.cofs.2015.03.001
Gao, Y., Liang, Q., Sun, J., Xiaomeng, W., Song, Y., Yunpeng, X., et al. (2025). Probiotic potential of lactic acid Bacteria with antioxidant properties in modulating health: mechanisms, applications, and future directions. Food Biosci. 64:106181. doi: 10.1016/j.fbio.2025.106181
Goffin, P., Lorquet, F., Kleerebezem, M., and Hols, P. (2004). Major role of NAD-dependent lactate dehydrogenases in aerobic lactate utilization in Lactobacillus Plantarum during early stationary phase. J. Bacteriol. 186, 6661–6666. doi: 10.1128/JB.186.19.6661-6666.2004
Gonçalves, D. A., González, A., Roupar, D., Teixeira, J. A., and Nobre, C. (2023). How prebiotics have been produced from agro-industrial waste: an overview of the enzymatic technologies applied and the models used to validate their health claims. Trends Food Sci. Technol. 135, 74–92. doi: 10.1016/j.tifs.2023.03.016
González-Rodríguez, I., Gaspar, P., Sánchez, B., Gueimonde, M., Margolles, A., and Neves, A. R. (2013). Catabolism of glucose and lactose in Bifidobacterium animalis subsp. lactis, studied by 13C nuclear magnetic resonance. Appl. Environ. Microbiol. 79, 7628–7638. doi: 10.1128/AEM.02529-13
Guerrero-Encinas, I., González-González, J. N., Santiago-López, L., Muhlia-Almazán, A., Garcia, H. S., Mazorra-Manzano, M. A., et al. (2021). Protective effect of Lacticaseibacillus Casei CRL 431 Postbiotics on mitochondrial function and oxidative status in rats with aflatoxin B1–induced oxidative stress. Probiotics Antimicrob. Proteins 13, 1033–1043. doi: 10.1007/s12602-021-09747-x
Huang, H.-C., Lee, I. J., Huang, C., and Chang, T.-M. (2020). Lactic acid bacteria and lactic acid for skin health and melanogenesis inhibition. Curr. Pharm. Biotechnol. 21, 566–577. doi: 10.2174/1389201021666200109104701
Ichinose, R., Fukuda, Y., Yamasaki-Yashiki, S., and Katakura, Y. (2020). Suppression of lactate production of Lactobacillus reuteri JCM1112 by co-feeding glycerol with glucose. J. Biosci. Bioeng. 129, 110–115. doi: 10.1016/j.jbiosc.2019.07.004
Iskandar, C. F., Cailliez-Grimal, C., Borges, F., and Revol-Junelles, A.-M. (2019). Review of lactose and galactose metabolism in lactic acid bacteria dedicated to expert genomic annotation. Trends Food Sci. Technol. 88, 121–132. doi: 10.1016/j.tifs.2019.03.020
Jiang, M., Wang, Y., Zhao, X., and Yu, J. (2024). From metabolic byproduct to immune modulator: the role of lactate in tumor immune escape. Front. Immunol. 15:1492050. doi: 10.3389/fimmu.2024.1492050
Kano, E., Aihara, T., Ghampson, I. T., Nogami, T., Miura, H., and Shishido, T. (2022). Continuous production of lactic acid from glycerol over bifunctional catalysts under base-free conditions using a liquid-phase flow reactor. ACS Sustain. Chem. Eng. 10, 12072–12081. doi: 10.1021/acssuschemeng.2c00129
Kim, J., Kim, Y. M., Lebaka, V. R., and Wee, Y. J. (2022). Lactic acid for green chemical industry: Recent advances in and future prospects for production technology, recovery, and applications. Fermentation 8:609. doi: 10.3390/fermentation8110609
Luiz-Santos, N., Prado-Ramírez, R., Arriola-Guevara, E., Camacho-Ruiz, R. M., and Moreno-Vilet, L. (2020). Performance evaluation of tight ultrafiltration membrane Systems at Pilot Scale for Agave Fructans fractionation and purification. Membranes 10, 1–19. doi: 10.3390/membranes10100261
Luiz-Santos, N., Prado-Ramírez, R., Camacho-Ruíz, R. M., Guatemala-Morales, G. M., Arriola-Guevara, E., and Moreno-Vilet, L. (2022). Effect of operating conditions and Fructans size distribution on tight ultrafiltration process for Agave Fructans fractionation: optimization and modeling. Membranes 12:575. doi: 10.3390/membranes12060575
Martha-Lucero, N., Cruz-Guerrero, A., Favela-Torres, E., Viniegra-González, G., Cira-Chávez, L. A., and Estrada-Alvarado, M. I. (2025). Lactic acid production by direct fermentation of Agave fructans. J. Chem. Technol. Biotechnol. 100, 935–941. doi: 10.1002/jctb.7828
Meruvu, H., and Tellioglu Harsa, S. (2023). Lactic acid Bacteria: isolation–characterization approaches and industrial applications. Crit. Rev. Food Sci. Nutr. 63, 8337–8356. doi: 10.1080/10408398.2022.2054936
Morales-Landa, J. L., Moreno-Vilet, L., Cordero-Quiñones, L. M., González-Montemayor, J. A., García-Fajardo, J. A., and Luiz-Santos, N. (2024). Effect of the molecular structure and degree of polymerization of fructans on the growth behavior of four lactic acid bacteria. ACS Food Sci. Technol. 4, 1929–1936. doi: 10.1021/acsfoodscitech.4c00251
Morita, H., Hidehiro, T. O. H., Fukuda, S., Horikawa, H., Oshima, K., Suzuki, T., et al. (2008). Comparative genome analysis of Lactobacillus renteri and Lactobacillus fermentum reveal a Genomic Island for reuterin and cobalamin production. DNA Res. 15, 151–161. doi: 10.1093/dnares/dsn009
Morita, H., Toh, H., Oshima, K., Murakami, M., Taylor, T. D., Igimi, S., et al. (2009). Complete genome sequence of the probiotic Lactobacillus rhamnosus ATCC 53103. J. Bacteriol. 191, 7630–7631. doi: 10.1128/JB.01287-09
Moya-Gonzálvez, E. M., Rubio-del-Campo, A., Rodríguez-Díaz, J., and Yebra, M. J. (2021). Infant-gut associated Bifidobacterium dentium strains utilize the galactose moiety and release lacto-N-triose from the human Milk oligosaccharides lacto-N-Tetraose and lacto-N-Neotetraose. Sci. Rep. 11:23328. doi: 10.1038/s41598-021-02741-x
Palencia-Argel, M., Rodríguez-Villamil, H., Bernal-Castro, C., Díaz-Moreno, C., and Fuenmayor, C. A. (2024). Probiotics in anthocyanin-rich fruit beverages: research and development for novel synbiotic products. Crit. Rev. Food Sci. Nutr. 64, 110–126. doi: 10.1080/10408398.2022.2104806
Patel, M. R. (2012). “Microbial production of lactic acid by using crude glycerol from biodiesel.” J. Microbiol. Biotechnol. Res. 2:90–93. Available online at: https://www.researchgate.net/publication/278683927
Plessas, S., Mantzourani, I., Terpou, A., and Bekatorou, A. (2024). Assessment of the physicochemical, antioxidant, microbial, and sensory attributes of yogurt-style products enriched with probiotic-fermented Aronia melanocarpa berry juice. Foods 13. doi: 10.3390/foods13010111
Pokusaeva, K., Fitzgerald, G. F., and Van Sinderen, D. (2011). Carbohydrate metabolism in Bifidobacteria. Genes Nutr. 6, 285–306. doi: 10.1007/s12263-010-0206-6
Popova-Krumova, P., Danova, S., Atanasova, N., and Yankov, D. (2024). Lactic acid production by Lactiplantibacillus plantarum AC 11S—kinetics and modeling. Microorganisms 12:739. doi: 10.3390/microorganisms12040739
Reale, A., Di Renzo, T., Rossi, F., Zotta, T., Iacumin, L., Preziuso, M., et al. (2015). Tolerance of Lactobacillus casei, Lactobacillus paracasei and Lactobacillus rhamnosus strains to stress factors encountered in food processing and in the gastro-intestinal tract. LWT 60, 721–728. doi: 10.1016/j.lwt.2014.10.022
Ruiz-Gonzalez, C., Cardona, D., Rueda-Ruzafa, L., Rodriguez-Arrastia, M., Ropero-Padilla, C., and Roman, P. (2024). Cognitive and emotional effect of a multi-species probiotic containing Lactobacillus rhamnosus and Bifidobacterium lactis in healthy older adults: a double-blind randomized placebo-controlled crossover trial. Probiotics Antimicrob. Proteins. doi: 10.1007/s12602-024-10315-2
Shiroda, M., and Manning, S. D. (2020). Lactobacillus strains vary in their ability to interact with human endometrial stromal cells. PLoS One 15:e0238993. doi: 10.1371/journal.pone.0238993
Swetha, T., Angelin, V., Ananthi,, Bora, A., Sengottuvelan, N., Ponnuchamy, K., et al. (2023). A review on biodegradable Polylactic acid (PLA) production from fermentative food waste - its applications and degradation. Int. J. Biol. Macromol. 234:123703. doi: 10.1016/j.ijbiomac.2023.123703
Turner, T. L., Kim, E., Hwang, C. H., Zhang, G. C., Liu, J. J., and Jin, Y. S. (2017). Short communication: conversion of lactose and whey into lactic acid by engineered yeast. J. Dairy Sci. 100, 124–128. doi: 10.3168/jds.2016-11784
Wang, Y., Wu, J., Lv, M., Shao, Z., Hungwe, M., Wang, J., et al. (2021). Metabolism characteristics of lactic acid Bacteria and the expanding applications in food industry. Front. Bioeng. Biotechnol. 9:612285. doi: 10.3389/fbioe.2021.612285
Wasewar, K. L., Yawalkar, A. A., Moulijn, J. A., and Pangarkar, V. G. (2004). Fermentation of glucose to lactic acid coupled with reactive extraction: a review. Ind. Eng. Chem. Res. 43, 5969–5982. doi: 10.1021/ie049963n
Zapaśnik, A., Sokołowska, B., and Bryła, M. (2022). Role of lactic acid bacteria in food preservation and safety. Foods 11:1283. doi: 10.3390/foods11091283
Zhang, G., Sun, H., Xu, Z., Tan, Z., Xiao, L., He, M., et al. (2023). Screening of Bifidobacteria with probiotic potential from healthy infant feces by using 2′-Fucosyllactose. Foods 12:858. doi: 10.3390/foods12040858
Zhao, J., Liyuan, Xu, Wang, Y., Zhao, X., Wang, J., Garza, E., et al. (2013). “Homofermentative production of optically pure L-lactic acid from xylose by genetically engineered Escherichia Coli B.” Available online at: http://www.microbialcellfactories.com/content/12/1/57
Keywords: biotechnology, probiotic bacterial, sugar sources, functional foods, biomass, lactic acid production
Citation: Morales-Landa JL, Lazcano-Díaz E, Escamilla-García E, Pérez-De la Rosa AG and Luiz-Santos N (2025) Comparative study of L/D lactic acid production in cultures of probiotic strains supplemented with carbohydrates from agave and other agroindustrial resources. Front. Sustain. Food Syst. 9:1613809. doi: 10.3389/fsufs.2025.1613809
Edited by:
José Antonio Teixeira, University of Minho, PortugalReviewed by:
Jagrani Minj, Washington State University Spokane, United StatesSujitta Raungrusmee, Kasetsart University, Thailand
Copyright © 2025 Morales-Landa, Lazcano-Díaz, Escamilla-García, Pérez-De la Rosa and Luiz-Santos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Noé Luiz-Santos, bnNhbnRvc0BjaWF0ZWoubXg=