 Sebastian Soppelsa1*
Sebastian Soppelsa1* Michael Gasser1Alessia Castellan2Manuel Pramsohler2Massimo Zago1Aldo Matteazzi3
Michael Gasser1Alessia Castellan2Manuel Pramsohler2Massimo Zago1Aldo Matteazzi3 Markus Kelderer4
Markus Kelderer4- 1Research Group for Berries and Stonefruit, Laimburg Research Centre, Bolzano, Italy
- 2Research Group for Arable Crops and Aromatic Plants, Laimburg Research Centre, Bolzano, Italy
- 3Laboratory for Soil and Plant Analysis, Laimburg Research Centre, Bolzano, Italy
- 4Research Group for Organic Farming, Laimburg Research Centre, Bolzano, Italy
Intercropping is considered a biodiversity-promoting practice and consists of the simultaneous cultivation of multiple crops in the same field. Despite its ecological benefits, its adoption in specialized farming systems—such as strawberry monocultures—remains limited, as these systems typically focus on maximizing income from a single crop. The experiment investigated the effects of intercropping strawberry (Fragaria × ananassa) with herbs such as chives (Allium schoenoprasum L.), marigold (Calendula officinalis L.), peppermint (Mentha × piperita L.), strawberry mint (Mentha suaveolens Ehrh.) and common sage (Salvia officinalis L.), using a 2:1 strawberry-to-herb ratio. Conducted over two consecutive years (2021–2022) in the alpine environment of Martell Valley (South Tyrol, Italy), the study assessed plant growth, fruit production, biological and economic feasibility, and the impact of intercropping on beneficial species, including pollinators, pests, and predators. Our results showed significant year-to-year variation. Although strawberry plants intercropped with herbs in 2021 achieved a fruit production that was not significantly different from that of pure stand (on average 150 g plant−1), the intercropping systems outperformed monocropping as revealed by indices such as land equivalent ratio (LER) and income equivalent ratio (IER) both greater than one. However, in the second year of cultivation (2022), the main parameters showed a decline. For example, strawberry plant biomass was decreased (by an average of 33%) in intercropping combinations involving chives, calendula and peppermint, resulting in a reduction in strawberry production of over 50% compared to monoculture. Indeed, the interaction between different plant species analyzed with competitive indices showed that strawberry plant appeared as the dominated crop in those combinations (aggressivity for strawberry: Af < 0; competition ratio for strawberry: CRf < 1). In both years, leaf nutrient content in intercropped strawberry plants was statistically similar to that of monocropped plants. Conversely, significant differences were observed in intercropped herbs (e.g., K + 50% and S + 30% in marigold and chives leaves, respectively). Intercropping systems, especially with chives and marigold, increased the richness of arthropod species (+100% as compared to strawberry in pure crop). Our research work contributes to a better understanding of the biological and economic feasibility of intercropping as a diversified farming practice for a strawberry farm. Furthermore, our findings offer a foundation for further investigations into interspecies interactions, particularly in terms of enhancing pollinator attraction and pest repulsion.
Introduction
Agricultural expansion and biodiversity preservation have long been considered incompatible (Tscharntke et al., 2005). Indeed, certain forms of farming systems—such as conventional, intensive, unsustainable agriculture—have been well documented as contributing to the deterioration of biodiversity (Dudley and Alexander, 2017; Erisman et al., 2016). The term biodiversity refers to the multitude of plants and animals that positively interact with each other to maintain optimal balance within an ecosystem (Jarvis et al., 2007). Refuges for biodiversity conservation should be protected, preserved and expanded in future-oriented agriculture (Hole et al., 2005; Scherr and McNeely, 2007; Selwood and Zimmer, 2020; Tscharntke et al., 2021). The concept of biodiversity is undoubtedly one of the fundamental principles underlying organic farming (Rahmann, 2011).
Organic farming is generally characterized by lower crop yield levels than conventional farming, and the reasons are mainly due to the incidence of abiotic and biotic stresses that are not always easy to control (de Ponti et al., 2012; Röös et al., 2018; Rundlöf et al., 2016; Schrama et al., 2018). This negative issue can be overcome, or at least mitigated, by adopting sustainable and biodiversity-friendly agroecological practices (Bedoussac et al., 2015; Hole et al., 2005; Liu et al., 2016; Rundlöf et al., 2016).
The intercropping planting method best represents the concept of biodiversity in organic farming, attempting to bridge the production gap that still distinguishes a sustainable cultivation system from a traditional one. However, intercropping is not yet widespread among organic farmers (Fowler et al., 2004). The principle on which it is based is to cultivate plants of different species on the same plot of land at the same time (Bybee-Finley and Ryan, 2018). In other words, the main crop (called target crop) grows together with a secondary crop (called companion crop), so that the two crops gain a mutual benefit from increased biodiversity (Parker et al., 2013). Competitive relationships may arise in an intercropping system due to the limited availability of resources such as water, nutrients and light (Zhang and Li, 2003). This situation should be averted by the choice of optimal companion crops and thus by establishing a harmonious interaction between the two crops (Wang et al., 2021a).
The interaction of consociated crops with the rhizosphere creates mechanisms that modify the physico-chemical properties of the soil, as well as the microbiological communities are affected (Dai et al., 2019; Wang et al., 2021b). Studies have confirmed that the practice of intercropping promotes soil organic carbon content, improves soil structure, increases soil permeability and water holding capacity, reduces erosion phenomena, stimulates biological activity, regulates the emission of CO2 and N2O from the soil, and reduces the presence of phytophagous, pathogens and weeds (Cong et al., 2015; Mousavi and Eskandari, 2011; Romaneckas et al., 2020; Wang et al., 2014). As is well known, leguminous plants represent valuable companion crops to be included in an intercropping system, mainly due to their nitrogen-fixing activity (Maitra et al., 2021; Mazzafera et al., 2021). A clear example is the combination of peanut (Arachis hypogaea L.) or broad bean (Vicia faba L.) with maize (Zea mays L.), which has been shown to increase the presence of nitrogen-fixing microorganisms such as Rhizobium hainanense, Rhizobium leguminosarum, and Frankia, thus increasing the nitrogen use efficiency (NUE) parameter (Chen et al., 2018; Fan et al., 2006). Other mineral elements can also benefit from adopting the intercropping technique. In soils characterized by the low solubility and bioavailability of iron (Fe) or phosphorus (P), the exudates emitted by the roots of certain companion crops (e.g., Allium sativum L., Lupinus albus L.) can modify the soil microbial population and favor the mobilization of those nutrients, making them available to both crops (Dai et al., 2019; Dissanayaka et al., 2017; Gardner and Boundy, 1983; Wang et al., 2014; Xiao et al., 2013). In addition to providing an opportunity to reduce the anthropogenic input of mineral nutrients (Du et al., 2020), intercropping practice could find application in counteracting the effects of soil fatigue (LaMondia et al., 2002; McIntyre et al., 2001).
The technique of intercropping has proven to be efficient in controlling the development of weeds, harmful insects, pathogens, attracting pollinating insects, predators, parasitoids, and thus promoting harmonious development of cultivated crops (Bybee-Finley and Ryan, 2018). If a “trap effect” is attributed to the companion crop, it will attract the biotic threat to itself, resulting in the preservation of the main crop but inevitable loss of the companion crop. Conversely, a companion crop with a “repellent effect” will be able to ward off harmful organisms (Parker et al., 2013).
Strawberry is a relevant crop worldwide and particularly in the region, where there are few reports of intercropping. A bibliometric study of scientific literature was accomplished on 20 December 2024, applying the Web of Science database (Clarivate, 2024). The search keywords adopted in the query were: “strawberry” and “intercrop”. The search was not limited by the year of publication, but reviews and duplicate publications were excluded. Our research revealed that 23 research documents, including 10 conference proceedings, were found using the query strings: TITLE: [(strawberry *) AND (intercrop *)]. Our enquiry showed the presence of limited experiments in which strawberry crop was involved as the main crop in an intercropping system. Still, some experiences showed potential of implementing intercropping on strawberry. For example, (Hata et al. 2016) studied the effects of strawberry plants intercropped with some companion crops, such as chives (Allium schoenoprasum L.), coriander (Coriandrum sativum L.), fennel (Foeniculum vulgare Mill.), garlic (Allium sativum L.), oregano (Origanum vulgare L.), marjoram (Origanum majorana L.), containing the mite population (Tetranychus urticae Koch) below the damage threshold. Although most of the aromatic plants tested gave good results, combining strawberry with garlic plants resulted in a reduction of more than 50% in the number of Tetranychus urticae individuals, highlighting how the reduction is directly proportional to the increase in the density of the companion plants (Hata et al., 2016). Volatile sulfur compounds (VSCs) such as diallyl thiosulphinate (allicin), abundant components in garlic plants, have insect-repellent properties (Borlinghaus et al., 2021). In a follow-up study, Hata et al. (2019a) confirmed the efficacy of Allium sativum L. and Allium tuberosum Rottl. ex Sprengel used in intercropping (strawberry plants grown in soil) and undercropping (with strawberry plants above ground on support trellises), respectively, in effectively counteracting the presence of harmful insects, such as Neopamera bilobata Say.
We therefore hypothesized that strawberry intercropped with herbs would exhibit better performance than monoculture. The present study aimed to assess the implication of crop diversification in an alpine environment, for two consecutive years by exploring production parameters, fruit quality and nutrient allocation in plant tissues. Given the working environment located within the Stelvio National Park, one of the largest nature reserves in Europe, we paid attention to the conservation of biodiversity in a typical anthropic activity such as strawberry cultivation.
We selected the main intercropping indices, in terms of biological and economic efficiency, as well as competitiveness, and grouped them in the following work to publish an exhaustive paper. Our findings can thus contribute to the development of new planting models for crops such as strawberry, which is grown almost exclusively in monoculture.
Materials and methods
Site description
The trial was conducted over two growing seasons (2021 and 2022) in an experimental strawberry field managed by the Laimburg Research Centre and located in the municipality of Martell (46° 33′ 30.618′′ N; 10° 46′ 53.649′′ E; 1,361 m a.s.l.) in Alto Adige/South Tyrol, Italy. The Martell Valley, historically renowned for berry production, is a side valley of the Venosta Valley and is characterized by a typical alpine-mountain climate. The Martell Valley is part of the Stelvio National Park, which is listed in the Official List of Protected Natural Areas (EUAP), established under Italian Law 394/1991 (Framework Law on Italian Protected Areas).
The transplanting of cold-stored strawberry plants in spring (especially in May) in soil conditions is currently the most popular cultivation strategy adopted by farmers in the valley. Flowering begins in June (about a month after transplanting) and fruiting in July–August, depending on the cultivars and weather conditions. Some farmers opt for a second harvest the following year using the same plants. In this case, the harvest will occur earlier than in the first year, and precisely in late June-July.
Meteorological trends during the growing seasons (from May to August 2021 and 2022) were recorded by an iMETOS® weather station using the cloud platform “Field-Climate” (Pessl Instruments, Weiz, Austria) and the data are reported in Table 1.
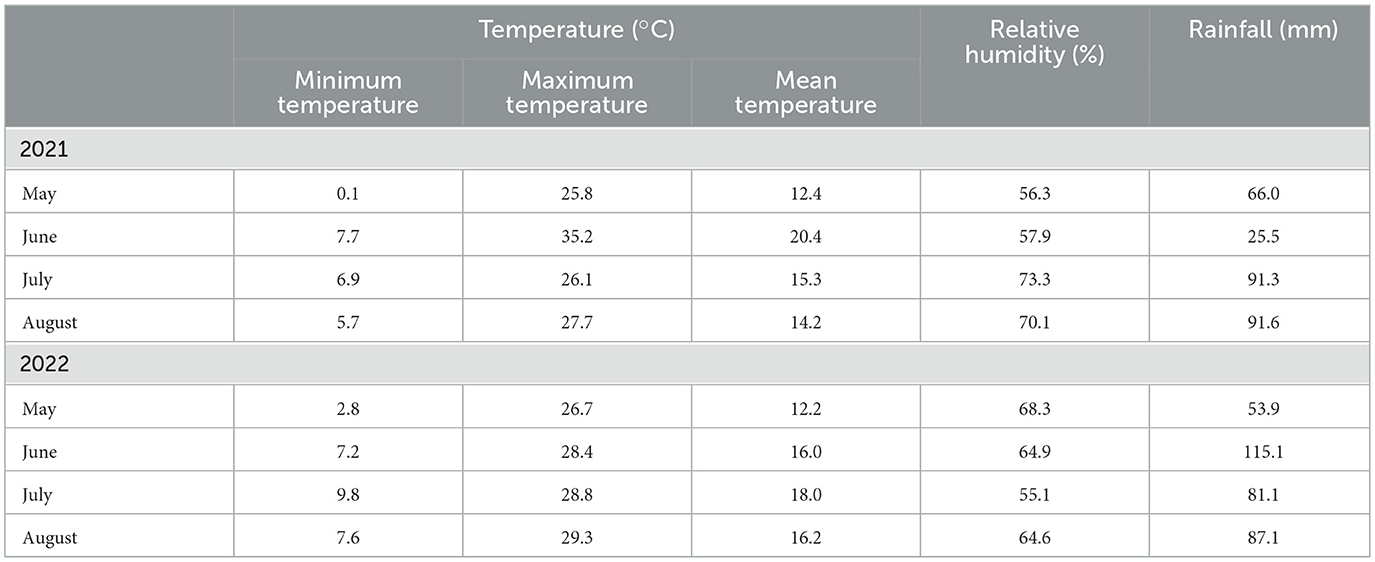
Table 1. Climatic conditions (monthly air temperatures, relative humidity, and rainfall) measured from May to August 2021 and 2022 during the first and second cropping year, respectively.
The soil properties of the 0–20 cm layer were as follows: humic loamy sand, pH = 5.1, no free carbonate, organic carbon expressed as humus of 7.3%, phosphorus = 5.0 mg 100 g−1, potassium = 8.0 mg 100 g−1, and magnesium = 18.0 mg 100 g−1. For P and K, the plant-available fraction was determined with Method (ÖNORM L 1087:2019 A.5, 2019). Field preparation took place in April with the incorporation of soil amendments (e.g., manure from local livestock farms, the application dosage was 20.0 t ha−1). Tillage was performed at the beginning of May 2021, followed by the formation of raised beds and covering with white plastic mulch film. The beds were spaced 1.1 m apart (distance measured from the centerline of the beds) with a length of 22.0 m. Poly-covered high tunnels with a structure of stainless-steel rods were built (22.0 m length, 5.5 m width, and 3.0 m height). Given the width of 5.5 m, five beds per tunnel were covered. Each bed consisted of a staggered double row with strawberry plants 0.3 m apart in each row and 0.2 m between rows, in order to reach 60,000 plants ha−1 (an average plant density as indicated by Soppelsa et al., 2023).
Treatments and experimental design
The main crop represented by strawberry plants (Fragaria × ananassa Duch.) cv. Roxana (frigo plants A++ from the nursery: Geoplant Vivai s.r.l. Soc. Agr., Italy) were transplanted on the 1st of June 2021. Roxana is a rustic variety (e.g., low susceptibility to leaf pathogens) and performs particularly well in an organic farming system. Strawberry plants were subjected to different intercropping combinations with herbs (companion crops) such as Allium schoenoprasum L. (chives), Calendula officinalis L. var. Daisy Golden (marigold), Mentha x piperita L. var. Multimentha (peppermint), Mentha suaveolens Ehrh. (strawberry mint), Salvia officinalis L. var. Extrakta (common sage) which were planted within the double row (strawberry-herb combination ratio of 2:1) (Figure 1). A. schoenoprasum, C. officinalis, and S. officinalis were sown in a greenhouse at Laimburg Research Centre in March 2021. A standard substrate was used, and plants were transplanted about 4 weeks after sowing. Plantlets were transplanted again after 4 weeks in multipots with a diameter of 10 cm. Peppermint was propagated from root stolons and the plantlets were transplanted in multipots (10 cm diameter). Plantlets of M. suaveolens were purchased from a local nursery. The aromatic plants were planted in the field at the same time as the strawberry plants. Considering the plant life cycle, C. officinalis is an annual plant and was therefore replanted during the second cropping year (mid-May). Plants were managed in the same way in terms of fertilization and watering. No pesticides were used throughout the experiment. The experimental design was organized as a completely randomized block design. Each intercropping combination was replicated six times. Each replicate represented an experimental unit, consisting of 20 strawberry plants (main crop) and 10 plants (companion crop) (Supplementary Figure S1). The bed was subdivided into three experimental units (each 6.0 m long, separated by 1.0 m buffer zone). Monocropped crops were also included in the cropping system design (also 6 replicates). This means that strawberry plants were planted staggered in double rows, while complementary crops in single rows.

Figure 1. Strawberry plants planted in a double row on raised beds and subjected to different intercropping treatments. Treatments' legend: FRA, Sole Fragaria x ananassa; FRA-ALL, Fragaria-Allium schoenoprasum; FRA-CAL, Fragaria-Calendula officinalis; FRA-MENp, Fragaria-Mentha x piperita; FRA-MENs, Fragaria-Mentha suaveolens; FRA-SAL, Fragaria-Salvia officinalis.
Field data collection and measurement
Strawberry plant growth as affected by intercropping regime was determined at the end of the second cropping year by randomly selecting four plants per replicate. Plants in monoculture were also collected. Each selected plant was separated into roots and aerial part and weighted fresh (g fresh weight (FW) plant−1). Then, plants were put in an oven (ED 56, Binder GmbH, Tuttlingen, Germany) at 65 °C until they reached a stable weight, and the dry mass was recorded (g dry weight (DW) plant−1). This evaluation was not carried out in the first year as we wanted to preserve the plots for the following year.
Ripe strawberry fruits (uniformly red) were harvested every 3 days during the period from the end of July to mid-August 2021 (first harvest year) and throughout the month of July 2022 (second harvest year). Data were expressed as total production per plant (g fruit plant−1), calculated by dividing the harvested total fruit weight by the number of plants (considered 20 plants per experimental unit). The plant organs of companion crops with significant commercial interest are aerial parts including stems, leaves and flowers. Marigold flowers were taken starting from July to August at regular intervals (every 5 days) for a total of 12 times per year. The other companion crops were harvested at pre-flowering stage in late July and late August, precisely on 26th July, 30th August 2021, and 21st July, 30th August 2022. Only for chives a further harvest was carried out on 9th June 2022.
Strawberries were sampled at two intermediate picking times for each harvest year. Twenty healthy fruits per replicate were analyzed to determine parameters such as: -Flesh firmness, expressed with the Durofel index (ID) which represents the elasticity of the skin of the fruit (Agrosta® Winterwood instrument, Agrosta Sàrl, Serqueux, France); -Total soluble solids (°Brix), determined with a refractometer (RFM840, Bellingham-Stanley Ltd., Kent, UK); -Titratable acidity (g L−1 of citric acid), measured with a titrator (Flash Automatic Titrator, Steroglass, Perugia, Italy) by titrating strawberry pulp to pH 8.2 using 0.1 M NaOH; -External fruit color, assessed with a colorimeter (CR-400, Konica Minolta, Tokyo, Japan) by measuring the same fruits at three different positions around the equatorial side of each fruit. The colorimetric coordinates (L*, a*, b*) were used to calculate the color index (Equation 1; according to Tessmer et al., 2016).
The nutrient content analyses were carried out on samples of stems, leaves, flowers and fruits, randomly collected from the main and companion crop plants during the central harvest phase of each year. The dried samples were ground into fine powder and homogenized for elemental characterization. Nitrogen content was determined with an elementary analyzer method according Dumas DIN EN ISO_16634_1:2009 (2009) (LECO Mod. Truspec) and the other macro (P, K, Ca, Mg, S) and microelements (Fe, Cu, B, Zn, Mn) were analyzed with microwave-assisted acid digestion [EPA 3052 1996 (1996); Milestone Mod. UltraWave] using the inductively coupled plasma optical emission spectrometry [ICP-OES; EPA 6010D 2014 (2014); Agilent Model 720].
Biological efficiency of intercropping combinations
The Land Equivalent Ratio (LER) indicates the relative area of the monoculture crops required to produce an equivalent yield in an intercropping system. As proposed by Mead and Willey (1980), LER was assessed as indicated in Equation 2.
Where “Y” indicates the individual crop yield, “I” the crop production in intercropping, “M” the production in monocropping, “f ” the target crop (i.e., strawberry), “k” the companion crop (i.e., chives, marigold, mint, or sage). An LER of 1 indicates that the intercropped crops have similar production level whether they are intercropped or monocultured. If the LER is greater than 1, it means that the intercrop needs less land area than the monoculture to produce equal yields of the different products. On the contrary, if lower than 1, the intercropping system has a worsening effect on the crops and pure cultivation of the respective crops should be preferred (Bybee-Finley and Ryan, 2018).
As duration of the land occupation by a cropping system is not considered in the LER calculation, Hiebsch and McCollum (1987) proposed an Area Time Equivalency Ratio (ATER) in which the time factor of crops was included (Equation 3).
Where tf and tk refer to the period (days) between planting and harvesting for strawberries (90 days) and herbs (120 days), respectively. T indicates the duration of the whole intercropping system (120 days is the duration period of an intercropping combination per year).
According to Mason et al. (1986), the Land Utilization Efficiency (LUE) provides a more accurate assessment than LER and ATER taken individually, because they may lead to an over- or underestimation of the cropping system, respectively. LUE was computed as shown in Equation 4.
The System Productivity Index (SPI) allows the yield of complementary crop to be standardized in terms of the main crop, and to identify the crop combinations that use resources more efficiently (Odo, 1991). SPI was calculated by the following Equation 5 and expressed as megagrams per hectare.
Percentage Yield Difference (PYD) is related to the yield difference between a monocrop (100%) and an intercropping system (expressed in percentage) (Afe and Atanda, 2014). It presumes that yield reduction of one crop should be compensated by an increase in yield of the other crop. The lower the PYD value, the more efficient the intercropping system, and vice versa. Equation 6 was proposed by Afe and Atanda (2014).
Competition indices
Aggressivity (A) was proposed by McGilchrist (1965) as a measure of how much the relative yield of the main crop in the mixture was greater than that of a companion crop. This competitive index was computed as shown in Equations 7a and 7b (Willey, 1979).
Where Zf indicates the planting proportion (%) of strawberry crop (67%) in an intercropping system whereas Zk is the proportion of companion crop (33%). The value of aggressivity (A) zero means both crops are equally competitive. A positive value of A indicates the dominant crop, and a negative value the dominated one.
Competition Ratio (CR) is an indicator of competitive ability of intercropping components. It denotes the number of times by which one component crop is more competitive than the other, as expressed in Equations 8a and 8b (Willey and Rao, 1980).
If the value CRf is < 1, there is a positive benefit of combining intercrops, indicating that the main crop can be cultivated in combination with the companion crop. If CRf > 1, the competitive ability of the main crop is higher than that of the companion crop in the mixture.
The index named Actual Yield Loss (AYL) proposed by Banik (1996) represents the proportionate yield loss or gain of intercrops in comparison to the respective monoculture. Compared to the LER, the AYL considers the actual planting proportion of component crops. It was calculated according to Equation 9.
AYLf and AYLk indicate the actual yield loss of the main crop and companion crop, respectively. The value (positive or negative) of AYL denotes an advantage/disadvantage in adopting the consociation system (Banik et al., 2000).
Economic efficiency of intercropping combinations
The above indices do not take the economic aspect into account. Through the Monetary Advantage Index (MAI) proposed by Ghosh (2004) (see Equation 10), an economic approach was introduced to assess the intercropping combination.
Where Pf refers to the commercial value of strawberries (a plausible direct sales price on the local farmers' market is 5 EUR per kg) and Pk to the unit price of the companion crop (i.e., chives 10 EUR kg−1, marigold flowers 20 EUR kg−1, mint 5 EUR kg−1, sage 12 EUR kg−1 fresh weight). The higher the MAY value, the more profitable the mixture.
According to Edje (1982), the income equivalent ratio (IER) was the conversion of LER in monetary terms. In other words, it corresponds to the area under monocropping required to produce the equivalent gross income achieved in the intercropping system (Equation 11; Gitari et al., 2020). If the IER is greater than 1, it means that the intercropping system is advantageous.
Evaluation of arthropod community response to intercropping system
Pests and beneficial arthropods—including pollinators, predators, and parasitoids—were monitored on both the main crop and companion crops from May to August each year. Observations were conducted every 3 days between 09:00 and 12:00 using visual inspection of the aerial parts of the plants. Some arthropods are very sensitive to movement and the presence of an observer, so we avoided any contact with plants. Each experimental unit was inspected (approximately 2 min) considering all the plants (strawberries and/or complementary crops). Visual monitoring is a widely accepted method for assessing taxa that are identifiable in the field (Ambrosino et al., 2006; Cardona et al., 2021; Dalton et al., 2024; Montgomery et al., 2021; Thompson et al., 2021). This approach is particularly suitable within protected areas such as the Stelvio National Park, as it minimizes disturbance and avoids biodiversity impoverishment. The presence of arthropod specimens was documented through photography for subsequent taxonomic identification to the order, family, genus, and species levels.
Statistical analysis
Data normality and homoscedasticity were examined with the Shapiro-Wilk test and the Flinger–Killeen's test, respectively. A two-way ANOVA was performed on data collected from both years and mean separation of the dependent variables obtained with the post-hoc Tukey HSD test (p < 0.05). In case of significant interaction between “treatments” and “years”, results were presented separately for the 2 years in dedicated figures or tables. In the case of non-normal data, Kruskal-Wallis test was applied. A one-way ANOVA was performed on data (e.g., plant biomass) collected only in the last year of cultivation (2022). For comparisons between two groups, a two-tailed unpaired Student's t-test was performed. All analyses were carried out in R v. 4.4.2. (R foundation for statistical computing, Vienna, Austria). Values were expressed as mean ± standard deviation (SD).
Results
Effect of intercropping system on the main crop (Fragaria × ananassa)
Total strawberry production resulted significantly affected by the factor “year” (Figure 2). During the first year (Figure 2A), the amount of strawberries reached an average of 150 g per plant, with no significant differences among the cropping patterns (mono- and inter-cropping). In contrast, in 2022 (Figure 2B), strawberry plants cultivated in sole cropping were characterized by the highest fruit production (282 g plant−1). Overall, FRA-ALL, FRA-CAL, and FRA-MENp combinations significantly reduced strawberries by −56%, −34%, −27%, respectively, as compared to respective monoculture production. Conversely, FRA-MENs and FRA-SAL exhibited non-significant difference compared to strawberry in pure cropping (Figure 2B).
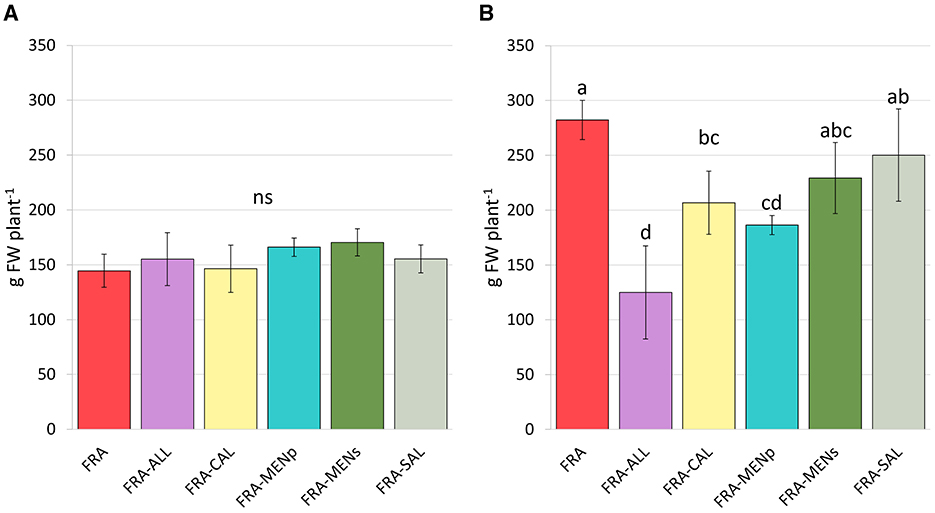
Figure 2. Strawberry total production, expressed as grams fresh fruit weight (FW) per plant, at the end of first (2021—A) and second (2022—B) cropping year, as affected by intercropping systems. Vertical bars indicate means ± SD (n = 6). The letters indicate significant differences according to Tukey's HSD test; p < 0.05 (ns, not significant).
Strawberry qualitative traits assessed as flesh firmness (FF), total soluble solid (TSS), titratable acidity (TA), and color index (CI) were not significantly affected by treatments (Table 2). The factor “year” was the only significantly relevant for the quality parameters. In general, FF, TSS and TA values were observed to be higher during the second cropping year, indicating more consistent, sweeter, and more acid-containing fruits. Fruit coloration appeared more intensely red in the first year (Table 2).
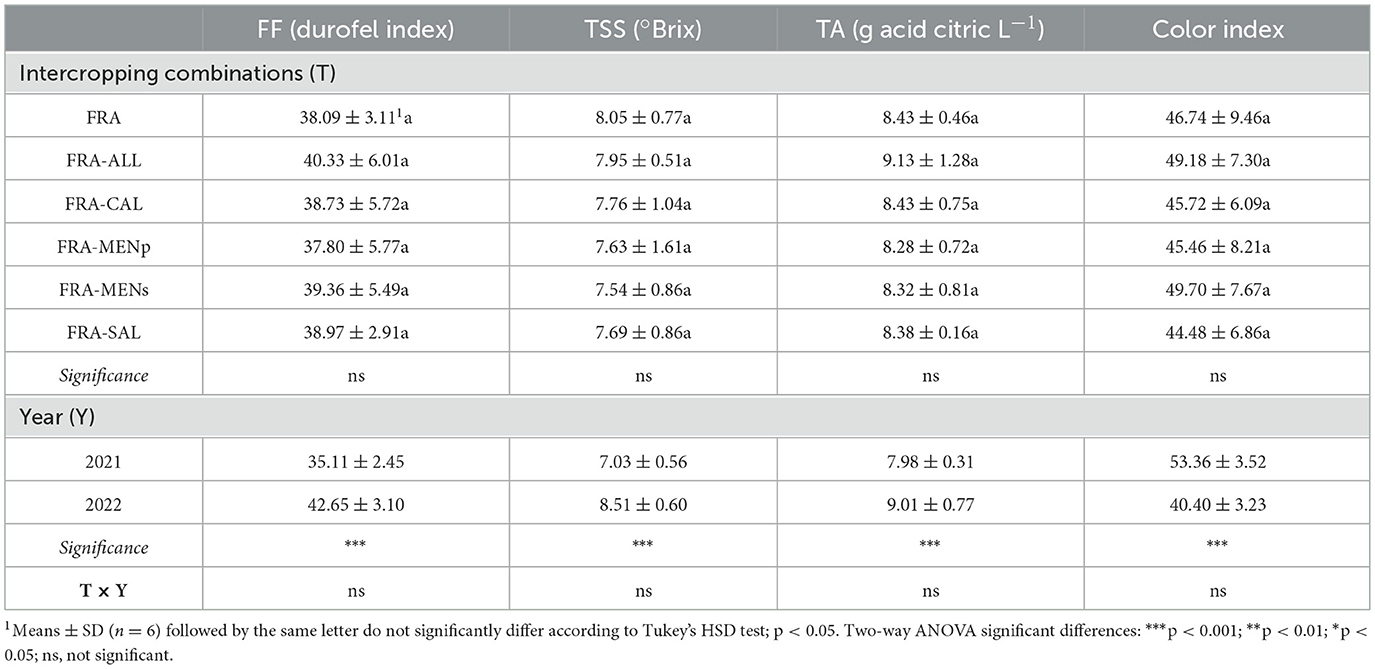
Table 2. Fruit quality traits (FF, firmness; TSS, total soluble solids; TA, titratable acidity, and CI, color index), as affected by intercropping systems.
Total biomass of strawberry plants under different cropping patterns at the end of the second year is presented in Figure 3. Intercropping with species from the genera Allium, Calendula, and Mentha (specifically M. piperita) resulted in a significant reduction in biomass, with an average decrease of 33% compared to monoculture (Figure 3). Consistent with the trends observed for fruit production data, the intercropping combinations FRA-MENs and FRA-SAL did not differ significantly from monoculture, indicating minimal impact on plant biomass.
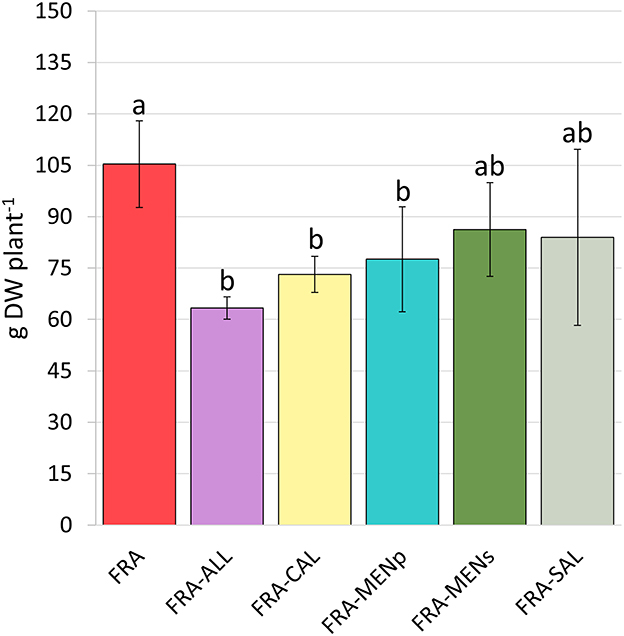
Figure 3. Total biomass (grams dry weight—DW per plant) of strawberry plants at the end of second cropping year (2022), as affected by intercropping systems. Vertical bars indicate means ± SD (n = 6). The letters on the top of the bar indicate significant differences according to Tukey's HSD test; p < 0.05.
Effect of intercropping system on the companion crops
Generally, intercropping combinations have almost always generated a significant reduction in production compared to the monoculture of those companion crops (xlimited in specialized farming systemsFigure 4). During the first year, the graph shows a deterioration in the performance of plants under a mixture system, with an average of −40% (Figure 4A). The gap between the two cultivation systems increased in the second year, except for peppermint (around 135 g in both mono- and inter-cropping systems) (Figure 4B).
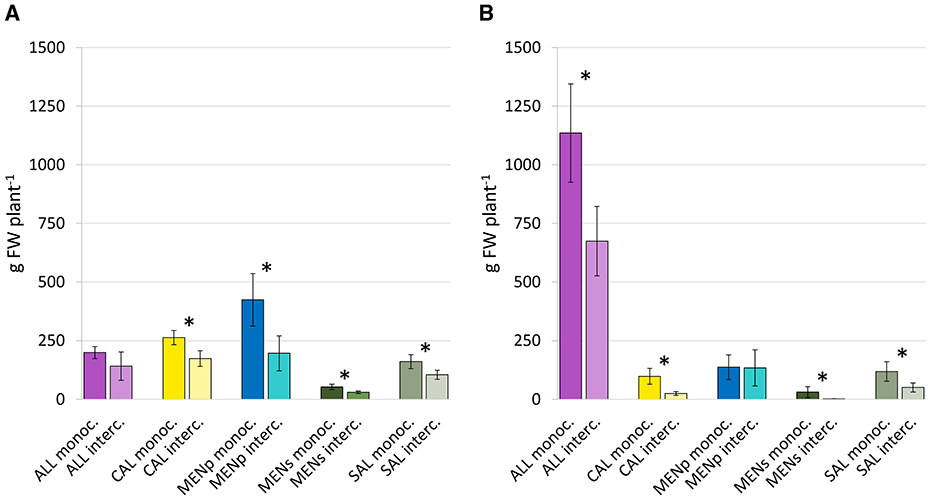
Figure 4. Herb total production, expressed as grams fresh weight (FW) per plant, at the end of first (2021—A) and second (2022—B) cropping year, as affected by intercropping systems. Vertical bars indicate means ± SD (n = 6). The abbreviation “monoc.” indicates a crop grown in monoculture, while “interc.” indicates the same crop in an intercropping system. Asterisk on the top between 2 bars indicates significant differences according to t-test (*p < 0.05).
Chives was the crop with the highest growth potential, quintupling production from the first to the second year. Marigold flower production, on the other hand, declined dramatically in 2022, as well as for peppermint in monoculture (−70%). The amount collected from strawberry mint and sage did not differ appreciably between cropping years.
Peppermint and chives were the companion crops with the highest production under pure stand in 2021 and 2022, respectively. Conversely, the companion crop with the lowest growth capacity in both harvest years was strawberry mint (less than 50 g per plant).
The concentrations of certain nutrients in leaves, stems and flowers of the companion crops were significantly (p < 0.05) influenced by the cropping year and the intercropping treatment (Figure 5). When compared with pure stands, intercropping treatments significantly increased the plant content of K (+50%) and S (+30%) in marigold and chives leaves, respectively, in both years (Figures 5A, B). In addition, the sulfur content also increased in other interplant treatments, e.g., in marigold (Figures 5B, C) and peppermint (Figure 5D), but this was significant only in the second year. Similarly, in 2022, the phosphorus content increased by +68, +28, and +39% in the leaves (Figure 5B) and flowers (Figure 5C) of marigold, and peppermint (Figure 5D), respectively, under a mixture regime. A higher magnesium content (+41%) was observed in the flowers of C. officinalis intercropped with strawberry in the second year (Figure 5C). The N and Ca contents of intercropped plants showed no significant differences compared to the respective monocultures (Figure 5). A high Ca content was evident in the marigold leaves, with values tending to exceed 5% DW (Figure 5B). For strawberry mint and sage treatments, macronutrient content in plant tissues was not significantly affected by cultivation regimes (Figures 5E, F).
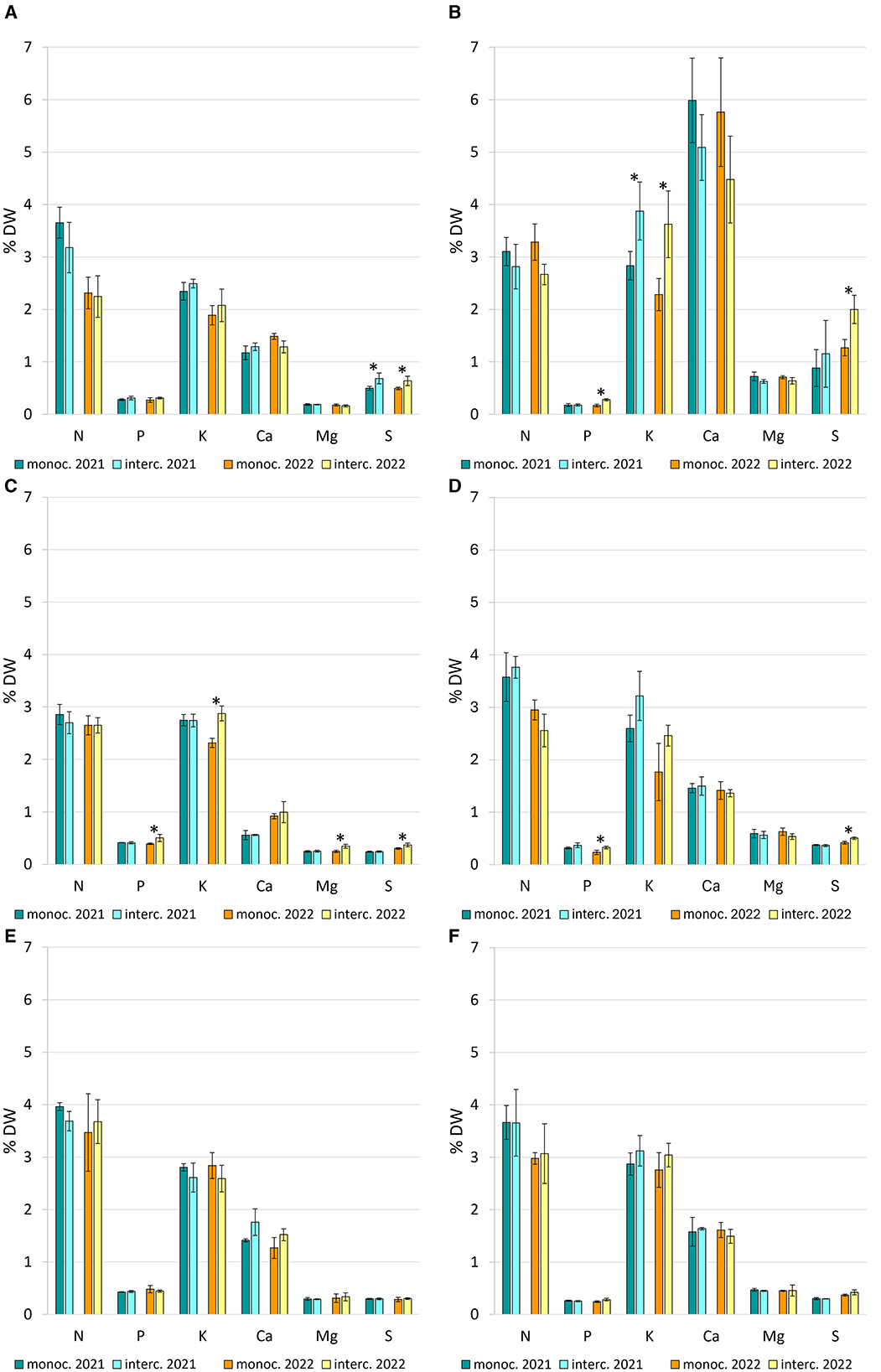
Figure 5. Macroelement content (N, P, K, Ca, Mg, and S), expressed as percentage dry weight—DW, in chives leaves (A), marigold leaves (B), marigold flowers (C), peppermint stems/leaves (D), strawberry mint stems/leaves (E), sage leaves (F), in plants grown in monoculture 2021 (turquoise bar) vs. intercropping 2021 (light blue bar) and monoculture 2022 (orange bar) vs. intercropping 2022 (yellow bar). Vertical bars indicate mean ± SD (n = 6). Asterisk on the top between 2 bars indicates significant differences according to t-test (*p < 0.05).
Intercropping patterns notably influenced manganese (Mn) accumulation in plant tissues (Figure 6). Overall, Mn content was reduced under intercropping for all companion crops, with the exception of S. officinalis. Particularly striking was the high Mn concentration in marigold (C. officinalis) leaves under sole cropping, where levels exceeded 1,300 mg kg−1 dry weight (DW). In contrast, intercropped marigold exhibited a significant reduction in leaf Mn content, with decreases exceeding 70% (Figure 6B). A similar trend was observed in marigold flowers, although absolute Mn values were substantially lower, ranging from 33 to 125 mg kg−1 DW (Figure 6C). In 2021, intercropping significantly increased boron (B) and iron (Fe) concentrations in strawberry mint (M. suaveolens), by 45% and 18%, respectively, compared to monoculture (Figure 6E). In 2022, B content in marigold leaves doubled under intercropping (Figure 6B). No significant differences in micronutrient content were detected between sole and intercropped systems for chives, peppermint, and sage across both years (Figures 6A, D, F).
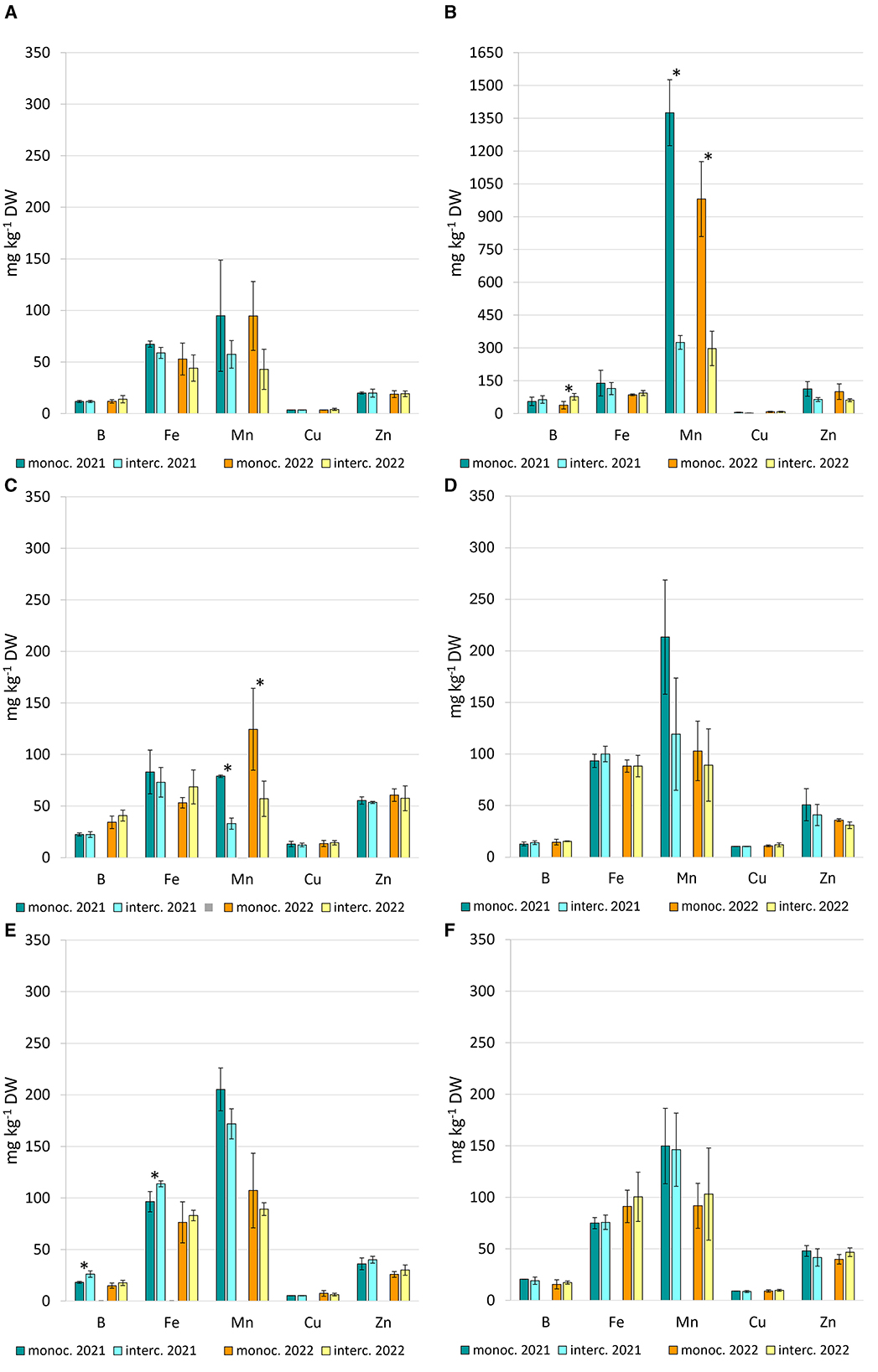
Figure 6. Microelement content (B, Fe, Mn, Cu, and Zn), expressed as mg per kg dry weight—DW, in chives leaves (A), marigold leaves (B), marigold flowers (C), peppermint stems/leaves (D), strawberry mint stems/leaves (E), sage leaves (F). Vertical bars indicate mean ± SD (n = 6). Asterisk on the top between 2 bars indicates significant differences according to t-test (*p < 0.05).
Biological efficiency of the Fragaria-herbs intercropping systems
The data reported in Figure 7 and Table 3 were presented separately for the 2 years, as the interaction between the factor “treatment” and the factor “year” was significant. LER, ATER, LUR, SPI, and PYD were significantly (p < 0.05) influenced by intercropping combinations with a substantial effect attributable to crop years. Generally, analyzing the indices separately for each year, we observed that during the first year the partial LER of target crop (i.e., strawberry) tended to be close to 1, corresponding to the theoretical value assumed for the pure stand (Figure 7A), while the partial LER values for companion crops ranged between 0.46 and 0.71. As consequence, the total LER 2021 values were higher than 1 in all cases (increased from +43 to +62% compared to sole cropping treatment), without significant differences among the intercropping treatments. On the contrary, intercropping combinations with total LER values around one occurred in 2022 (e.g., in FRA-ALL, FRA-CAL, and FRA-MENs) (Figure 7B). However, the highest LER value (above 1) in 2022 was recorded in FRA-MENp mix, with a value (1.64) significantly higher than the other intercropping combinations. A similar trend was shown for the ATER and LUE (Table 3). Regarding the SPI and PYD (Table 3), extreme values were recorded in the second cropping year, because the highest significant value was obtained in FRA-MENp combination, while the lowest in FRA-MENs.
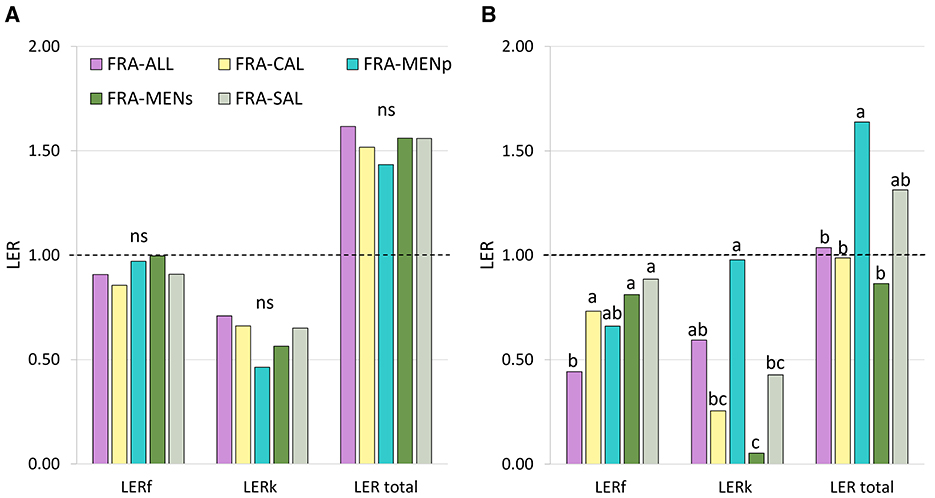
Figure 7. Land equivalent ratios (LERs) at the first (2021—A) and second (2022—B) cropping year, as affected by intercropping systems. Partial LERs are indicated with “f ” for strawberry crop and “k” for companion crops. (n = 6). The letters indicate significant differences according to Tukey's HSD test; p < 0.05 (ns, not significant). The dashed lines denote a LER equal to 1.
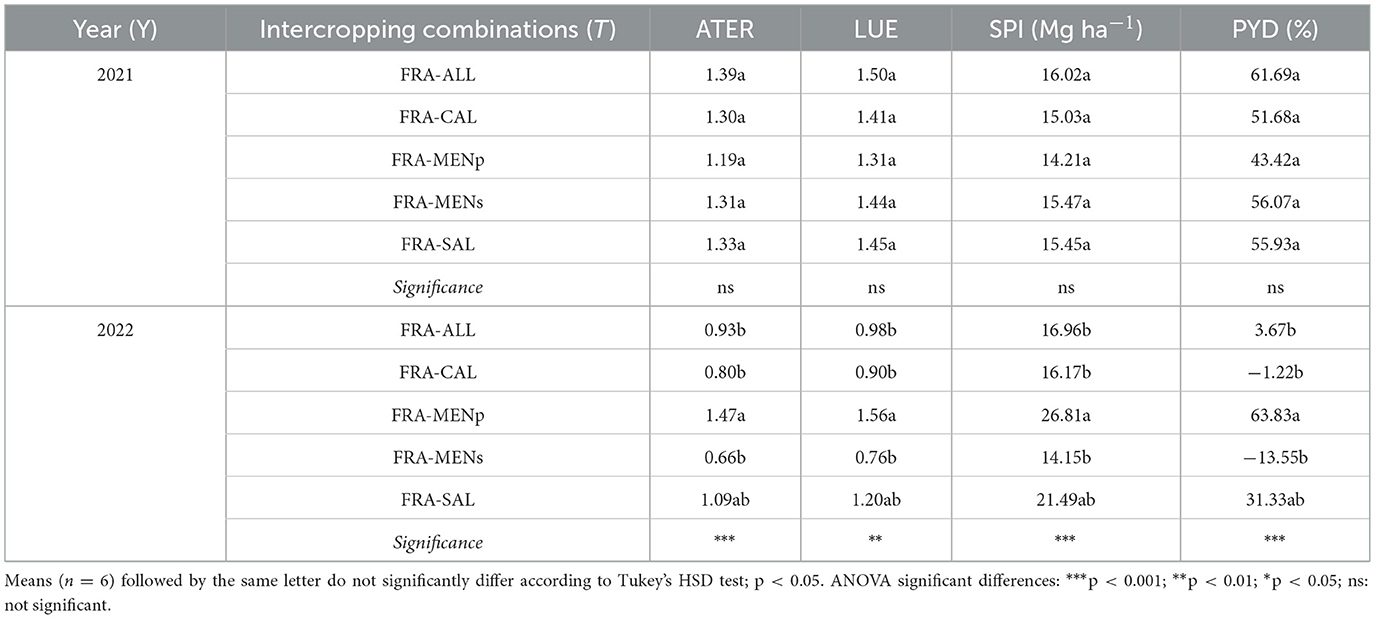
Table 3. Area time equivalent ratio (ATER), land utilization efficiency (LUE), system productivity index (SPI), percentage yield difference (PYD) as affected by strawberry-herb intercropping systems.
Competition indices under the Fragaria-herbs intercropping systems
As previously observed for the biological efficiency indices, significant differences emerged for competition indices depending on the factor “year” (Table 4). In our 2-year experiment, the aggressivity of companion crops exhibited consistently positive (up to +0.02) or equal to 0, and consequently Fragaria values were negative (up to −0.02) or zero, suggesting that strawberry was the dominated crop, or that both crops were equally competitive (A = 0), respectively. In only one combination was strawberry dominant (e.g., in FRA-MENs during the second year).
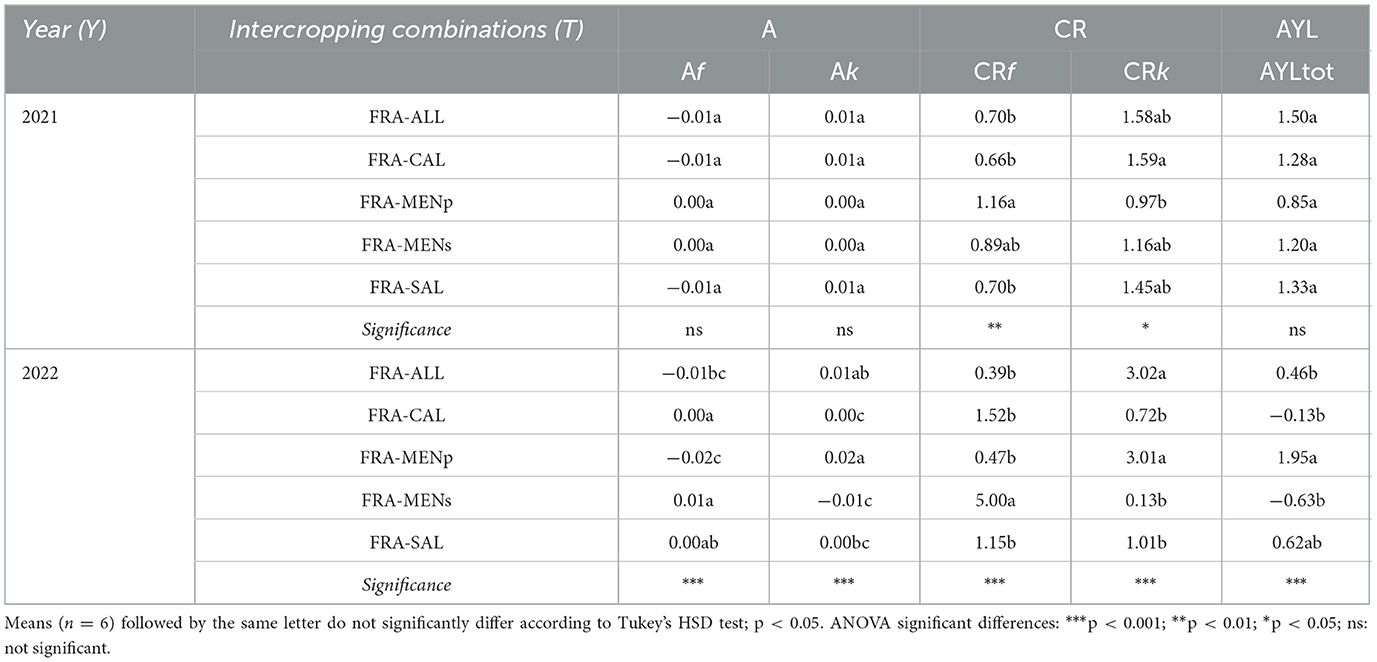
Table 4. Aggressivity (A), competition ratio (CR), actual yield loss (AYL) as affected by strawberry-herb intercropping systems.
In almost all cropping systems, the partial CRk was greater than one, meaning that companion crops were characterized by a higher ability of competition than Fragaria (Table 4). The highest and lowest CRf values were recorded in 2022 with the FRA-MENs and FRA-ALL combinations, respectively. In addition, the highest and the lowest CRk were obtained in the same intercropping combinations, but in the opposite way. That means that strawberry mint appeared as a poor competitor, while chives were a strong competitor.
In all treatments, an advantage in intercropping system was achieved as confirmed by the positive value of the AYL index, except for FRA-MENs (−0.63) and FRA-CAL (−0.13) in the second year (Table 4).
Economic efficiency of the intercropping systems
The calculation of the Income Equivalent Ratio (IER) and the Monetary Advantage Index (MAI) offers an effective means of assessing the economic efficiency of cropping systems tested (Table 5). The total IER, resulting from the sum of partial IERf and IERk, showed values above 1 in all intercropping combinations except in FRA-MENs.
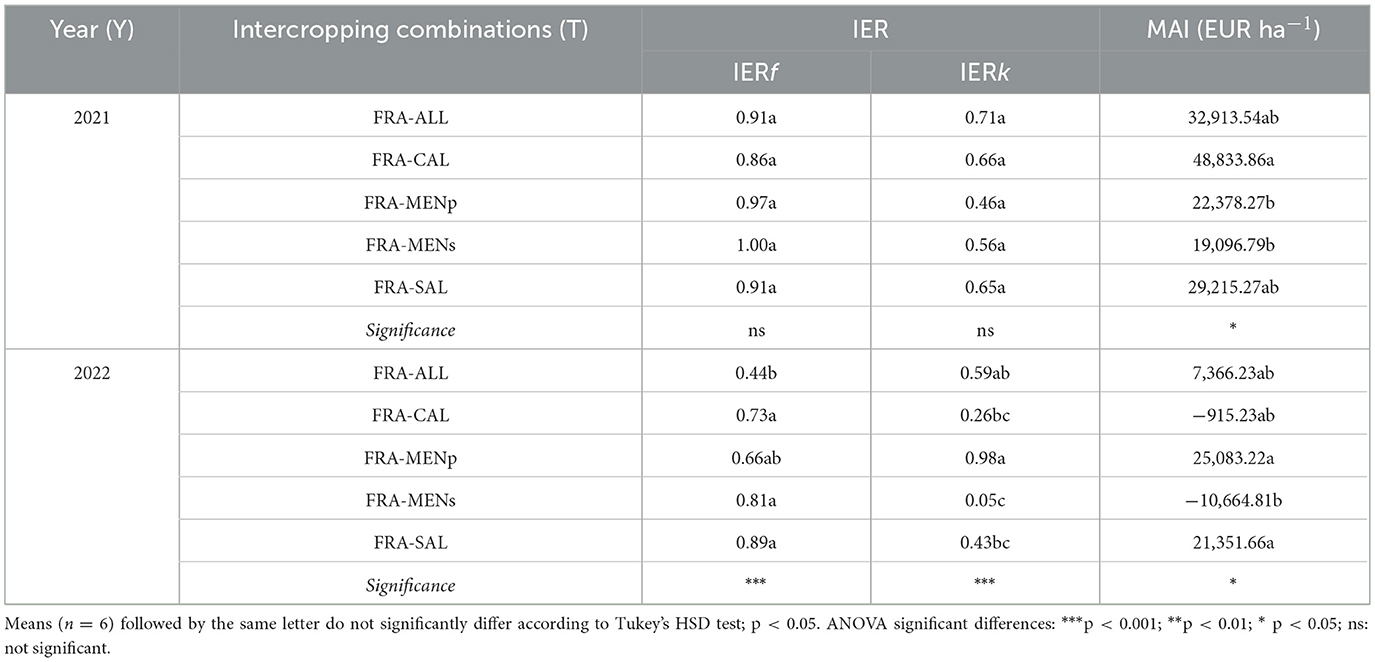
Table 5. Income equivalent ratio (IER), monetary advantage index (MAI), as affected by strawberry-herb intercropping systems.
In the second year, the MAI value decreased dramatically in 3 out of 5 combinations. For instance, FRA-CAL reached the highest MAI value in 2021 (EUR 48,833.86), while a negative value was observed in the following year. Similar differences were recorded for FRA-MENs (EUR−10,664.81, corresponding to the lowest MAI value), and for FRA-ALL (the MAI value in 2022 was lower than in the first year but with a positive sign). The MAI values remained stable (above EUR 20,000) for the combinations FRA-MENp and FRA-SAL (both years).
Evaluation of arthropod community response to intercropping system
The visual survey conducted during the experimental period recorded the highest species richness in the blocks subjected to intercropping treatments (+100% as compared to strawberry in pure crop; Figure 8). The number of individuals observed varied depending on the seasons. Arthropods belonging to six insect orders, including Coleoptera, Diptera, Hymenoptera, Lepidoptera, Thysanoptera, and Hemiptera, were identified and assigned to species level.

Figure 8. Arthropods categorized into four functional groups (pollinators, pests, predators, and neutral species). The crop on which they were observed is indicated in brackets. Arthropods identified in strawberry pure stand (left) and under intercropping systems (right).
Bees (especially Apis mellifera L.) were the most frequently observed insects throughout the blooming period, both on target and companion plants. In the group of pollinators, long hoverfly (Sphaerophoria scripta L.), thick-legged hoverfly (Syritta pipiens L.), common drone fly (Eristalis tenax L.) bumblebee (Bombus terrestris L.), small tortoiseshell (Aglais urticae L.) and peacock butterfly (Aglais io L.) were also detected but only on companion crops, particularly on chives and marigold.
Since no pest control treatments were applied, harmful insects such as strawberry-blossom weevil (Anthonomus rubi Herbst), garden foliage beetle (Phyllopertha horticola L.), western flower thrips (Frankliniella occidentalis Pergande), black vine weevil (Otiorhynchus sulcatus Fabricius), strawberry aphids (Chaetosiphon fragaefolii Cockerell) were observed on strawberry plants.
A well-known predator such as the common ladybug (Coccinella septempunctata L.) was found throughout the vegetative and productive phases of the plants.
Other arthropods identified were green tortoise beetle (Cassida viridis L. on peppermint), longhorn beetle (Leptura quadrifasciata L. on strawberry), bordered straw (Heliothis peltigera Denis and Schiffermüller on sage), click beetles (Athous haemorrhoidalis Fabricius on strawberry), and European garden spider (Araneus diadematus Clerck on peppermint).
Discussion
The strawberry is a high-value horticultural crop that requires intensive investment, and profitability is maximized through monoculture (Salaheen and Biswas, 2019; Simpson, 2018). The application of diversification practices such as intercropping still remains limited in specialized farming systems. According to the survey by Rodriguez et al. (2021), the barriers to crop diversification perceived by farmers can be listed as: (1) complex agricultural systems are difficult to manage due to lack of specific knowledge (e.g., best combinations of companion-target crop, suitable technology, etc.); (2) the lack of markets with dedicated infrastructures for alternative or minor crops; (3) high risk of failing to achieve the required quality (e.g., against unforeseen weather events, supporting policy and regulations are necessary). In this context, our study aimed to fill the first gap by conducting a comprehensive investigation into the implementation of the intercropping practice in strawberry farming.
The results from the two-year study indicate that the intercropping systems depend on the cropping year. The total LER values were greater than one during the first season (2021), suggesting the beneficial effect of all companion crops included in these intercropping patterns (Figure 7). Our results were consistent with previous research by Karlidag and Yildirim (2009) and Hata et al. (2019b), who reported that strawberry plants intercropped with vegetables (e.g., lettuce, radish, onion and garlic) significantly improved the land use efficiency (LER > 1) compared to strawberries grown in pure stand. On the other hand, in the second year (2022), reductions were recorded in FRA-ALL, FRA-CAL, and FRA-MENs, with values approaching 1 (Figure 7). Similar trends were shown for other biological efficiency indices (Table 3). The variation of these indices over the years was closely related to the production parameter.
Local strawberry production levels are around 250 g per plant during the first year, to double the following year (considering plants cultivated in soil conditions; Soppelsa et al., 2023). These values refer to crops managed according to Integrated Pest Management (IPM). In an organic or non-treatment regime, as in the current study, the production values are inevitably lower.
Analyzing the first year (2021), strawberry production data revealed a stable situation among the various combinations and no significant differences in comparison with pure stand (Figure 2A). Nurseries provide strawberry plants with appropriate floral induction conditions for optimal fruit production in future commercial fields (Bosc and Demené, 2009). This means strawberry plants maintain constant production levels during the first cropping year in the field, even though the plants are subjected to different agronomic growing conditions (Soppelsa et al., 2023). Nevertheless, plant-plant interactions in the first year can generate negative implications for the second harvest cycle (Soppelsa et al., 2023). The negative impact on strawberry plants in terms of production and total biomass was clearly visible in the second year for the intercropping combinations such as FRA-ALL, FRA-CAL, and FRA-MENp (Figures 2B, 3).
Strawberry crown and roots are considered a crucial source of carbohydrates and nutrients stored in reserve organs in late summer and then remobilized the following spring to support plant growth and reproductive organs (Albregts and Howard, 1980; Kirschbaum et al., 2012; Macías-Rodríguez et al., 2002; Tagliavini et al., 2005). Poor developed crowns directly affect plant vigor and production performance (Fagherazzi et al., 2021; Torres-Quezada et al., 2015). Intra- or inter-specific competition dependent on light and nutrient availability can lead plants to alter floral differentiation, the accumulation and distribution of storage compounds and nutrients in the reserve organs (Aerts, 1999; Craine and Dybzinski, 2013; Kobe, 1997; Soppelsa et al., 2023; Zhang et al., 2018).
Anyway, the lower biomass in strawberry plants observed in some of our cropping systems cannot be attributed to nutrient deficiencies because no changes in nutrient content were recorded in strawberry leaf tissue (Supplementary Table S1). The causes could be found in another factor such as light. Exposure to light is an essential environmental factor for plant development (Wu et al., 2025). The interaction between light and reproductive phenology in strawberry plants has been well studied as reported in some studies (Sidhu et al., 2022; Sønsteby and Hytonen, 2005; Wang et al., 2020). Planting plants close together inevitably leads to shading, resulting in poor photosynthetic capacity and reduced plant growth (Li et al., 2014; Pierson et al., 1990). In the first year, the size of the young companion plants was not sufficient to establish shading effects on the target plant, while the development of some herbs became excessive in the following year. The competition indices help us to explain the morphological changes in companion crops. The target crop appeared to be the dominated crop (negative A values), with a lower ability to compete than the companion crops (CRf < 1, especially in combination with chives and peppermint plants in 2022) (Table 4). Chives are perennial plants with thin, clustered bulbs, each with leaves in clumps of 2-5 (Chen, 2006). Leaves are cylindrical/filiform, and therefore a few bulbs, like our starting plants, do not have a great capacity for shade. Nevertheless, the bulbs multiply very quickly over time, creating a dense and large clump of leaves (Figure 4). Similarly, Mentha x piperita, a rhizomatous perennial plant with rapidly spreading above- and below-ground stolons, can quickly colonize new areas (Taneja and Chandra, 2012). The stems can reach a height of approximately 0.5 m (Oroian et al., 2017), decisively exceeding the height of a strawberry plant. In addition, the stems were not always well erect, thus shading any surrounding plants. Choosing complementary crops suitable for growing in the shade might be a valid solution (Prakhyath et al., 2024).
The development of complementary plants was altered by intercropping treatments (Figure 4). These findings are in line with a study conducted by Ragab et al. (2014), who observed a more vigorous growth in sole crops than intercropping patterns. Several studies have determined that the intercropping practice modifies plant-soil interactions, affecting root architecture, metabolite and microbiome profiles of the rhizosphere compared to the profiles observed in monoculture (Jiang et al., 2024; Liu et al., 2023; Moţ et al., 2023; Shao et al., 2024). Increased microbial diversity and chemo-diversity of metabolites in intercropping soils can lead to an impact on nutrient uptake by plants (Chen et al., 2023; Jiang et al., 2024; Lu et al., 2025). It is interesting to note that the sulfur content increased significantly in the leaves of intercropped chives in both years, with a similar trend for the other intercropped herbs (Figure 5). Organic sulfur content in the soil is usually positively correlated with organic matter (Nguyen and Goh, 1992). Soil organic matter (SOM) mineralization gives rise to sulfate () which is the main source of available S for plants (Narayan et al., 2023). Sulfur-containing compounds such as cysteine (Cys) and methionine (Met) are involved in protein synthesis, while other metabolites (e.g., glutathione—GSH) contribute to oxidative stress protection and redox-control in plants (Godat et al., 2010; Sharma et al., 2024; Singh and Schwan, 2011). Sulfur interacts with other mineral elements, modifying the nutrient uptake and assimilation (Abdin et al., 2003), showing a synergistic relationship with K and P uptake (Mehmood et al., 2021; Singh et al., 2013) and an antagonistic effect for Mn assimilation (Kalbas et al., 1988; Klikocka, 2011). These findings were in agreement with the data shown in our study (Figures 5, 6). For instance, manganese content was drastically reduced in most intercropping systems (significantly in intercropped marigold plants; Figures 6B, C). Li et al. (2024) observed that the Mn uptake and extraction amount from soil were significantly decreased in intercropped maize plants compared to maize in pure stand, while increasing the Mn content in the associated plant (Sedum alfredii, Hance). In contrast, the results of our experiment did not reveal the strawberry to be a hyperaccumulator plant of that heavy metal (Supplementary Table S1).
According to several literature studies (Alcon et al., 2024; Rosa-Schleich et al., 2019; Sánchez et al., 2022; Stratton et al., 2021; Yogendra et al., 2024), a diversified farming system might lead to particularly favorable economic outcomes. Assessment of intercropping based on economic efficiency indicated that IER values were above 1, suggesting advantageous intercropping systems in both years with the only exception in FRA-MENs in 2022 (Table 5).
A higher MAI value means that the intercropping pattern is more profitable than another (Ghosh, 2004). The highest MAI value, close to EUR 50,000 per hectare, was observed in FRA-CAL cropping system in 2021 (Table 5). Calendula officinalis is a popular medicinal plant with edible flowers rich in nutraceutical compounds used in food and pharmaceutical industries (Kumari et al., 2021; Shahane et al., 2023). As consumer health consciousness has increased in recent years, the market for edible flowers has also grown (Fernandes et al., 2020; Jadhav et al., 2023). Pot marigold is considered a low-investment crop but high-profit (Popa, 2024). Nevertheless, the following year the MAI value collapsed for that combination because the flower yield of pot marigold intercrop was negligible (Figure 4B). This result can be explained due to planting time. While in the first year the young strawberry and pot marigold plants, planted at the same time, developed harmoniously together, in the second year the transplanting time of the marigold had a significant influence on its development. Please note that C. officinalis is an annual plant, which needs to be planted every year. Planting time is a crucial factor for intra-species competition, and if not optimized, has negative consequences on plant growth and yield (Ahmed et al., 2020). As reported by Santos et al. (2008), analyzing the different planting times for cucurbits included in some intercropping patterns with strawberry plants. In our experiment, the C. officinalis was probably planted too late in 2022 and consequently the already well-established strawberry plant, with its leaf canopy, prevented the young marigold plants from growing properly (Figure 4). In contrast to marigold, adopting perennial plants could be advantageous (e.g., no need to transplant new plants). Considering all planting patterns, great economic stability over the 2 years was achieved with strawberries intercropped with peppermint and sage (Table 5), both of which, however, developed less than in the first year (Figure 4). In the second year, we noted that parts of the sage plants were desiccated, confirming the limited tolerance of this Mediterranean species to alpine environments characterized by harsh winters. This observation is consistent with those of Carlen et al. (2009), which report that overwintering of Salvia officinalis L. can be problematic in mountain areas.
As shown in Figure 8, the intercropping patterns adopted in the trial were a promising approach to integrating islands of biodiversity into traditional farming. For instance, we found new and more beneficial arthropods attracted to the floral resources produced by the herbs in the intercropping systems (Figure 8). More specifically, marigold flowers proved particularly attractive to pollinators as confirmed by Kowalska et al. (2022). Although companion crops can provide supplemental food (e.g., nectar, pollen) for pollinators and natural enemies, the abundant and concomitant flowering of some species such as C. officinalis can act as a competitor in the pollination of the target crop (Azpiazu et al., 2020). In the field experiment conducted by Gowton et al. (2021), peppermint intercropped with berry crops increased the pollinator community and reduced the spotted-wing drosophila (Drosophila suzukii Matsumura) infestations. These results can be attributed to the volatile organic compounds (VOCs) released by mint, which have attraction or deterrence properties (Conboy et al., 2020; Gowton et al., 2021). Other studies have confirmed the ability of herbs to protect main crops against pests (Hata et al., 2016; Jankowska et al., 2009; Matsuura et al., 2006). Therefore, establishing attraction and/or repulsion zones in the field could represent a potential pest management practice.
The interference of intercropping treatments with arthropod pest pressure could not be established in our study. Although the strawberry plants were colonized by several individuals belonging to the pest category (e.g., blossom weevil, flower thrips), we only identified their presence, without counting the individuals. In future trials, the visitation rate of arthropods such as pollinators, predators and pests affected by the intercropping treatments will certainly be taken into account.
Conclusion
The present study is a broad investigation into the use of intercropping in strawberry cultivation in the alpine environment. Considering the parameters analyzed, the 2-year study revealed markedly different outcomes depending on the cropping year. All the different plant combinations in 2021 showed an advantage from the application of the intercropping practice, as illustrated by biological efficiency indices (e.g., LER above 1) and by positive values of the economic index (e.g., the highest MAI value was reached with C. officinalis). However, unlike the first year, companion crops such as chives, marigold and strawberry mint led to a deterioration of the cropping system in the following year. Greater economic stability over the 2 years was achieved with mint and sage.
Based on these findings, the following recommendations are proposed: evaluate whether to use annual or perennial companion crops, and whether the target crop will be maintained for one or more growing seasons; the selection of companion crops should prioritize species that are already present in the terroir and are tolerant or less susceptible to pathogens and pests.
This study also offers new insights into nutrient uptake dynamics, which warrant more in-depth analysis, in relation to the mechanisms of nutrient acquisition by plant roots in a mixed cropping system. Additionally, further research is needed to better understand how the arthropod community is influenced by intercropping, with the aim of leveraging this practice as a sustainable pest management strategy.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
SS: Conceptualization, Data curation, Investigation, Methodology, Supervision, Writing – original draft, Writing – review & editing. MG: Conceptualization, Investigation, Writing – review & editing. AC: Conceptualization, Investigation, Writing – review & editing. MP: Conceptualization, Investigation, Supervision, Writing – review & editing. MZ: Funding acquisition, Investigation, Supervision, Writing – review & editing. AM: Investigation, Writing – review & editing. MK: Methodology, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research activity was conducted in the framework of the “Action Plan for Mountain Agriculture and Food Sciences 2016–2022”, adopted by the Government of South Tyrol (Italy).
Acknowledgments
The authors thank the Department of Innovation, Research, University, and Museums of the Autonomous Province of Bozen/Bolzano for covering the Open Access publication costs.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsufs.2025.1645707/full#supplementary-material
References
Abdin, M. Z., Ahmad, A., Khan, N., Khan, I., Jamal, A., Iqbal, M., et al. (2003). “Sulphur interaction with other nutrients,” in Sulphur in Plants, eds. Y. P. Abrol and A. Ahmad (Berlin: Springer), 359–374.
Aerts, R. (1999). Interspecific competition in natural plant communities: mechanisms, trade-offs and plant-soil feedbacks. J. Exp. Bot. 50, 29–37. doi: 10.1093/jxb/50.330.29
Afe, A. I., and Atanda, S. (2014). Percentage yield difference, an Index for evaluating intercropping efficiency. J. Exp. Agricult. Int. 5, 459–465. doi: 10.9734/AJEA/2015/12405
Ahmed, S., Raza, M. A., Yuan, X., Du, Y., Iqbal, N., Chachar, Q., et al. (2020). Optimized planting time and co-growth duration reduce the yield difference between intercropped and sole soybean by enhancing soybean resilience toward size-asymmetric competition. Food Energy Secur. 9:e226. doi: 10.1002/fes3.226
Albregts, E. E., and Howard, C. M. (1980). Accumulation of nutrients by strawberry plants and fruit grown in annual hill culture1. J. Am. Soc. Horticult. Sci. 105, 386–388. doi: 10.21273/JASHS.105.3.386
Alcon, F., Albaladejo-García, J. A., Martínez-García, V., Rossi, E. S., Blasi, E., Lehtonen, H., et al. (2024). Cost benefit analysis of diversified farming systems across Europe: incorporating non-market benefits of ecosystem services. Sci. Total Env. 912:169272. doi: 10.1016/j.scitotenv.2023.169272
Ambrosino, M. D., Luna, J. M., Jepson, P. C., and Wratten, S. D. (2006). Relative frequencies of visits to selected insectary plants by predatory hoverflies (Diptera: Syrphidae), other beneficial insects, and herbivores. Environ. Entomol. 35, 394–400. doi: 10.1603/0046-225X-35.2.394
Azpiazu, C., Medina, P., Adán, Á., Sánchez-Ramos, I., del Estal, P., Fereres, A., et al. (2020). The role of annual flowering plant strips on a melon crop in central Spain: influence on pollinators and crop. Insects 11:66. doi: 10.3390/insects11010066
Banik, P. (1996). Evaluation of wheat (Triticum aestivum) and legume intercropping under 1:1 and 2:1 row-replacement series system. J. Agron. Crop Sci. 176, 289–294. doi: 10.1111/j.1439-037X.1996.tb00473.x
Banik, P., Sasmal, T., Ghosal, P. K., and Bagchi, D. K. (2000). Evaluation of Mustard (Brassica compestris Var. Toria) and legume intercropping under 1: 1 and 2: 1 row-replacement series systems. J. Agron. Crop Sci. 185, 9–14. doi: 10.1046/j.1439-037X.2000.00388.x
Bedoussac, L., Journet, E-. P., Hauggaard-Nielsen, H., Naudin, C., Corre-Hellou, G., Jensen, E. S., et al. (2015). Ecological principles underlying the increase of productivity achieved by cereal-grain legume intercrops in organic farming. A review. Agron. Sustain. Dev. 35, 911–935. doi: 10.1007/s13593-014-0277-7
Borlinghaus, J., Foerster, J., Kappler, U., Antelmann, H., Noll, U., Gruhlke, M. C. H., et al. (2021). Allicin, the odor of freshly crushed garlic: a review of recent progress in understanding Allicin's effects on cells. Molecules 26:1505. doi: 10.3390/molecules26061505
Bosc, J. P. H., and Demené, M. N. (2009). Floral induction duration, plant architecture and fruit production relations in strawberry cv. ‘Ciflorette'. Acta Hortic. 842, 667–670. doi: 10.17660/ActaHortic.2009.842.143
Bybee-Finley, K., and Ryan, M. (2018). Advancing intercropping research and practices in industrialized agricultural landscapes. Agriculture 8:80. doi: 10.3390/agriculture8060080
Cardona, A., Tchamitchian, M., Penvern, S., Dufils, A., Kramer Jacobsen, S., Korsgaard, M., et al. (2021). Monitoring methods adapted to different perceptions and uses of functional biodiversity: insights from a European qualitative study. Ecol. Indic. 129:107883. doi: 10.1016/j.ecolind.2021.107883
Carlen, C., Carron, C. A., Previdoli, S., and Baroffio, C. (2009). Salvia officinalis: influence of cutting frequency, cutting height and date of the last harvest before winter. Acta Hortic. 826, 25–30. doi: 10.17660/ActaHortic.2009.826.2
Chen, H. (2006). “19—Chives,” in Handbook of Herbs and Spices, ed. K. V. Peter (Cambridge: Woodhead Publishing), 337–346.
Chen, J., Arafat, Y., Wu, L., Xiao, Z., Li, Q., Khan, M. A., et al. (2018). Shifts in soil microbial community, soil enzymes and crop yield under peanut/maize intercropping with reduced nitrogen levels. Appl. Soil Ecol. 124, 327–334. doi: 10.1016/j.apsoil.2017.11.010
Chen, X., Chen, J., and Cao, J. (2023). Intercropping increases soil N-targeting enzyme activities: a meta-analysis. Rhizosphere 26:100686. doi: 10.1016/j.rhisph.2023.100686
Clarivate (2024). Web of Science [Database]. Available online at: https://clarivate.com/academia-government/scientific-and-academic-research/research-discovery-and-referencing/web-of-science/ (Retrieved December 20, 2024).
Conboy, N. J. A., McDaniel, T., George, D., Ormerod, A., Edwards, M., Donohoe, P., et al. (2020). Volatile organic compounds as insect repellents and plant elicitors: an integrated pest management (IPM) strategy for glasshouse whitefly (Trialeurodes vaporariorum). J. Chem. Ecol. 46, 1090–1104. doi: 10.1007/s10886-020-01229-8
Cong, W-. F., Hoffland, E., Li, L., Six, J., Sun, J-. H., Bao, X-. G., et al. (2015). Intercropping enhances soil carbon and nitrogen. Glob. Chang. Biol. 21, 1715–1726. doi: 10.1111/gcb.12738
Craine, J. M., and Dybzinski, R. (2013). Mechanisms of plant competition for nutrients, water and light. Funct. Ecol. 27, 833–840. doi: 10.1111/1365-2435.12081
Dai, J., Qiu, W., Wang, N., Wang, T., Nakanishi, H., Zuo, Y., et al. (2019). From leguminosae/gramineae intercropping systems to see benefits of intercropping on iron nutrition. Front. Plant Sci. 10:605. doi: 10.3389/fpls.2019.00605
Dalton, D., Berger, V., Kirchmeir, H., Adams, V., Botha, J., Halloy, S., et al. (2024). A Framework for Monitoring Biodiversity in Protected Areas and Other Effective Area-Based Conservation Measures: Concepts, Methods and Technologies (1st Edn.). Gland: IUCN, International Union for Conservation of Nature.
de Ponti, T., Rijk, B., and van Ittersum, M. K. (2012). The crop yield gap between organic and conventional agriculture. Agric. Syst. 108, 1–9. doi: 10.1016/j.agsy.2011.12.004
DIN EN ISO_16634_1:2009 (2009). Food products—Determination of the total nitrogen content by combustion according to the Dumas principle and calculation of the crude protein content.Berlin: Beuth Verlag.
Dissanayaka, D. M. S. B., Wickramasinghe, W. M. K. R., Marambe, B., and Wasaki, J. (2017). Phosphorus-mobilization strategy based on carboxylate exudation in lupins (Lupinus, Fabaceae): a mechanism facilitating the growth and phosphorus acquisition of neighbouring plants under phosphorus-limited conditions. Exp. Agricult. 53, 308–319. doi: 10.1017/S0014479716000351
Du, Q., Zhou, L., Chen, P., Liu, X., Song, C., Yang, F., et al. (2020). Relay-intercropping soybean with maize maintains soil fertility and increases nitrogen recovery efficiency by reducing nitrogen input. Crop J. 8, 140–152. doi: 10.1016/j.cj.2019.06.010
Dudley, N., and Alexander, S. (2017). Agriculture and biodiversity: a review. Biodiversity 18, 45–49. doi: 10.1080/14888386.2017.1351892
Edje, C. L. (1982). “Comparative development and yield and other agronomic characteristics of maize and groundnut in monoculture and in association,” in Intercropping: Proceedings of the second Symposium on Intercropping in Semi-Arid Areas, held at Morogoro, Tanzania, 4-7 August 1980. Symposium on Intercropping in Semi-Arid Areas, Ottawa. Intern, ed. C. L. Keswani and B. J. Ndunguru. Ottawa: Development Research Centre.
EPA 3052 1996 (1996). Microwave Assisted Acid Digestion of Siliceous and Organically Based Matrics. Washington, D.C.: U.S. Environmental Protection Agency.
EPA 6010D 2014 (2014). Inductively Coupled Plasma—Optical Emission Spectrometry. Washington, D.C.: U.S. Environmental Protection Agency.
Erisman, J. W., van Eekeren, N., de Wit, J., Koopmans, C., Cuijpers, W., Oerlemans, W., et al. (2016). Agriculture and biodiversity: a better balance benefits both. AIMS Agricult. Food 1, 157–174. doi: 10.3934/agrfood.2016.2.157
Fagherazzi, A. F., Suek Zanin, D., Soares dos Santos, M. F., Martins de Lima, J., Welter, P. D., Francis Richter, A., et al. (2021). Initial crown diameter influences on the fruit yield and quality of strawberry Pircinque. Agronomy 11:184. doi: 10.3390/agronomy11010184
Fan, F., Zhang, F., Song, Y., Sun, J., Bao, X., Guo, T., et al. (2006). Nitrogen fixation of faba bean (Vicia faba L.) interacting with a non-legume in two contrasting intercropping systems. Plant Soil 283, 275–286. doi: 10.1007/s11104-006-0019-y
Fernandes, L., Casal, S., Pereira, J. A., Saraiva, J. A., and Ramalhosa, E. (2020). An overview on the market of edible flowers. Food Rev. Int. 36, 258–275. doi: 10.1080/87559129.2019.1639727
Fowler, C., Condron, L., and McLenaghen, R. (2004). Effect of green manures on nitrogen loss and availability in organic cropping system. N. Z. J. Agricult. Res. 47, 95–100. doi: 10.1080/00288233.2004.9513575
Gardner, W. K., and Boundy, K. A. (1983). The acquisition of phosphorus by Lupinus albus L. IV. The effect of interplanting wheat and white lupin on the growth and mineral composition of the two species. Plant Soil 70, 391–402. JSTOR. doi: 10.1007/BF02374894
Ghosh, P. K. (2004). Growth, yield, competition and economics of groundnut/cereal fodder intercropping systems in the semi-arid tropics of India. Field Crops Res. 88, 227–237. doi: 10.1016/j.fcr.2004.01.015
Gitari, H. I., Nyawade, S. O., Kamau, S., Karanja, N. N., Gachene, C. K. K., Raza, M. A., et al. (2020). Revisiting intercropping indices with respect to potato-legume intercropping systems. Field Crops Res. 258:107957. doi: 10.1016/j.fcr.2020.107957
Godat, E., Madalinski, G., Muller, L., Heilier, J-. F., Labarre, J., Junot, C., et al. (2010). “Chapter 2—Mass spectrometry-based methods for the determination of sulfur and related metabolite concentrations in cell extracts,” in Methods in Enzymology, Vol. 473, eds. E. Cadenas and L. Packer (Cambridge, MA: Academic Press), 41–76.
Gowton, C. M., Cabra-Arias, C., and Carrillo, J. (2021). Intercropping with peppermint increases ground dwelling insect and pollinator abundance and decreases Drosophila suzukii in fruit. Front. Sustain. Food Syst. 5:700842. doi: 10.3389/fsufs.2021.700842
Hata, F. T., Ventura, M. U., Béga, V. L., Camacho, I. M., and de Paula, M. T. (2019a). Chinese chives and garlic in intercropping in strawberry high tunnels for Neopamera bilobata Say (Hemiptera: Rhyparochromidae) control. Bull. Entomol. Res. 109, 419–425. doi: 10.1017/S0007485318000299
Hata, F. T., Ventura, M. U., Carvalho, M. G., Miguel, A. L. A., Souza, M. S. J., Paula, M. T., et al. (2016). Intercropping garlic plants reduces Tetranychus urticae in strawberry crop. Exp. Appl. Acarol. 69, 311–321. doi: 10.1007/s10493-016-0044-3
Hata, F. T., Ventura, M. U., de Paula, M. T., Shimizu, G. D., de Paula, J. C. B., Kussaba, D. A. O., et al. (2019b). Intercropping garlic in strawberry fields improves land equivalent ratio and gross income. Cienc. Rural 49:e20190338. doi: 10.1590/0103-8478cr20190338
Hiebsch, C. K., and McCollum, R. E. (1987). Area- × -Time equivalency ratio: a method for evaluating the productivity of intercrops1. Agron. J. 79, 15–22. doi: 10.2134/agronj1987.00021962007900010004x
Hole, D. G., Perkins, A. J., Wilson, J. D., Alexander, I. H., Grice, P. V., Evans, A. D., et al. (2005). Does organic farming benefit biodiversity? Biol. Conserv. 122, 113–130. doi: 10.1016/j.biocon.2004.07.018
Jadhav, H. B., Badwaik, L. S., Annapure, U., Casanova, F., and Alaskar, K. (2023). A Review on the Journey of edible flowers from farm to consumer's plate. Appl. Food Res. 3:100312. doi: 10.1016/j.afres.2023.100312
Jankowska, B., Poniedziałek, M., and Jedrszczyk, E. (2009). Effect of intercropping white cabbage with French Marigold (Tagetes patula nana L.) and Pot Marigold (Calendula officinalis L.) on the colonization of plants by pest insects. Folia Horticulturae 21, 95–103. doi: 10.2478/fhort-2013-0129
Jarvis, D. I., Padoch, C., and Cooper, H. D. (2007). “Biodiversity, agriculture, and ecosystem services,” in Managing Biodiversity in Agricultural Ecosystems, eds. D. I. Jarvis, C. Padoch, and H. D. Cooper (Columbia: Columbia University Press), 1–12.
Jiang, P., Wang, Y., Zhang, Y., Fei, J., Rong, X., Peng, J., et al. (2024). Intercropping enhances maize growth and nutrient uptake by driving the link between rhizosphere metabolites and microbiomes. New Phytol. 243, 1506–1521. doi: 10.1111/nph.19906
Kalbas, M., Filsoof, F., and Rezai-Nejad, Y. (1988). Effect of sulfur treatments on yield and uptake of Fe, Zn, and Mn by corn, sorghum, and soybeans. J. Plant Nutr. 11, 1353–1360. doi: 10.1080/01904168809363892
Karlidag, H., and Yildirim, E. (2009). Strawberry intercropping with vegetables for proper utilization of space and resources. J. Sustain. Agric. 33, 107–116. doi: 10.1080/10440040802587462
Kirschbaum, D., Larson, K., Weinbaum, S., and Dejong, T. (2012). Accumulation pattern of total nonstructural carbohydrate in strawberry runner plants and its influence on plant growth and fruit production. Afr. J. Biotechnol. 11, 16253–16262. doi: 10.5897/AJB12.2567
Klikocka, H. (2011). The effect of sulphur kind and dose on content and uptake of micro-nutrients by potato tubers (Solanum tubersosum L.). Acta Scientiarum Polonorum, Hortorum Cultus, 10, 65–75.
Kobe, R. K. (1997). Carbohydrate allocation to storage as a basis of interspecific variation in sapling survivorship and growth. Oikos 80, 226–233. doi: 10.2307/3546590
Kowalska, J., Antkowiak, M., and Sienkiewicz, P. (2022). Flower strips and their ecological multifunctionality in agricultural fields. Agriculture 12:1470 . doi: 10.3390/agriculture12091470
Kumari, P., and Ujala, Bhargava, B. (2021). Phytochemicals from edible flowers: opening a new arena for healthy lifestyle. J. Funct. Foods 78:104375. doi: 10.1016/j.jff.2021.104375
LaMondia, J. A., Elmer, W. H., Mervosh, T. L., and Cowles, R. S. (2002). Integrated management of strawberry pests by rotation and intercropping. Crop Prot. 21, 837–846. doi: 10.1016/S0261-2194(02)00050-9
Li, T., Liu, L-. N., Jiang, C-. D., Liu, Y-. J., and Shi, L. (2014). Effects of mutual shading on the regulation of photosynthesis in field-grown sorghum. J. Photochem. Photobiol. B Biol. 137, 31–38. doi: 10.1016/j.jphotobiol.2014.04.022
Li, Z., Shang, Q., Zou, L., Xing, Z., Chen, G., Chen, Z., et al. (2024). The dynamic response mechanism of crops to manganese uptake and transfer mediated by different intercropping crop attributes. J. Sci. Food Agric. 104, 9706–9718. doi: 10.1002/jsfa.13795
Liu, H., Meng, J., Bo, W., Cheng, D., Li, Y., Guo, L., et al. (2016). Biodiversity management of organic farming enhances agricultural sustainability. Sci. Rep. 6:23816. doi: 10.1038/srep23816
Liu, X., Jiao, Y., Zhao, X., Yu, X., Zhang, Q., Li, S., et al. (2023). Root architecture of forage species varies with intercropping combinations. Agronomy 13:2223 . doi: 10.3390/agronomy13092223
Lu, M., Zhao, J., Lu, Z., Li, M., Yang, J., Fullen, M., et al. (2025). Maize–soybean intercropping increases soil nutrient availability and aggregate stability. Plant Soil 506, 441–456. doi: 10.1007/s11104-023-06282-2
Macías-Rodríguez, L., Quero, E., and López, M. G. (2002). Carbohydrate differences in strawberry crowns and fruit (Fragaria × ananassa) during plant development. J. Agric. Food Chem. 50, 3317–3321. doi: 10.1021/jf011491p
Maitra, S., Hossain, A., Brestic, M., Skalicky, M., Ondrisik, P., Gitari, H., et al. (2021). Intercropping—A low input agricultural strategy for food and environmental security. Agronomy 11:343. doi: 10.3390/agronomy11020343
Mason, S. C., Leihner, D. E., and Vorst, J. J. (1986). Cassava-cowpea and cassava-peanut intercropping. I. yield and land use efficiency. Agron. J. 78, 43–46. doi: 10.2134/agronj1986.00021962007800010010x
Matsuura, S., Hoshino, S., and Koga, H. (2006). Verbena as a trap crop to suppress thrips-transmitted Tomato spotted wilt virus in chrysanthemums. J. Gener. Plant Pathol. 72, 180–185. doi: 10.1007/s10327-005-0264-6
Mazzafera, P., Favarin, J. L., and de Andrade, S. A. L. (2021). Editorial: intercropping systems in sustainable agriculture. Front. Sustain. Food Syst. 5:634361. doi: 10.3389/fsufs.2021.634361
McGilchrist, C. A. (1965). Analysis of competition experiments. Biometrics 21, 975–985. doi: 10.2307/2528258
McIntyre, B., Gold, C., Kashaija, I., Ssali, H., Night, G., Bwamiki, D., et al. (2001). Effects of legume intercrops on soil-borne pests, biomass, nutrients and soil water in banana. Biol. Fertil. Soils 34, 342–348. doi: 10.1007/s003740100417
Mead, R., and Willey, R. W. (1980). The concept of a ‘Land Equivalent Ratio' and advantages in yields from intercropping. Exp. Agricult. 16, 217–228. doi: 10.1017/S0014479700010978
Mehmood, M. Z., Afzal, O., Ahmed, M., Qadir, G., Kheir, A. M. S., Aslam, M. A., et al. (2021). Can sulphur improve the nutrient uptake, partitioning, and seed yield of sesame? Arab. J. Geosci. 14:865. doi: 10.1007/s12517-021-07229-6
Montgomery, G. A., Belitz, M. W., Guralnick, R. P., and Tingley, M. W. (2021). Standards and best practices for monitoring and benchmarking insects. Front. Ecol. Evol. 8. doi: 10.3389/fevo.2020.579193
Moţ, A., Frîncu, M., Vlad, I., Ivan, E. Ş., Bujor, O.-C., Mihalcea, B., et al. (2023). Influence of intercropping on physico-chemical and biological soil properties in organic strawberry crop. Sci. Pap. Ser. B Horticult. 67, 132–137.
Mousavi, S. R., and Eskandari, H. (2011). A general overview on intercropping and its advantages in sustainable agriculture. J. Appl. Environ. Biol. Sci. 1, 482–486.
Narayan, O. P., Kumar, P., Yadav, B., Dua, M., and Johri, A. K. (2023). Sulfur nutrition and its role in plant growth and development. Plant Signal. Behav. 18:2030082. doi: 10.1080/15592324.2022.2030082
Nguyen, M. L., and Goh, K. M. (1992). Sulphur mineralization and release of soluble organic sulphur from camp and non-camp soils of grazed pastures receiving long-term superphosphate applications. Biol. Fertil. Soils 14, 272–279. doi: 10.1007/BF00395463
Odo, P. E. (1991). Evaluation of short and tall Sorghum varieties in mixtures with cowpea in the Sudan savanna of Nigeria: land equivalent ratio, grain yield and system productivity index. Exp. Agricult. 27, 435–441. doi: 10.1017/S0014479700019426
ÖNORM L 1087:2019 A.5. (2019). Chemical Analysis of Soils - Determination of “plant-available” Phosphorus and Potassium by the Calcium-Acetate-Lactate (CAL)-Method. Austrian Standards International.
Oroian, C., Covrig, I., Malinas, C., Moldovan, C., and Fleseriu, A. (2017). Effects of cultivation systems and environmental conditions on peppermint (Mentha × piperita L.) biomass yield and oil content. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 45:576. doi: 10.15835/nbha45210857
Parker, J. E., Snyder, W. E., Hamilton, G. C., and Rodriguez-Saona, C. (2013). “Companion planting and insect pest control,” in Weed and Pest Control Conventional and New Challenges, eds. S. Soloneski and M. Larramendy (Rijeka: InTech), 1–30.
Pierson, E. A., Mack, R. N., and Black, R. A. (1990). The effect of shading on photosynthesis, growth, and regrowth following defoliation for Bromus tectorum. Oecologia 84, 534–543. doi: 10.1007/BF00328171
Popa, R-. G. (2024). Calendula officinalis culture—Business with low investment, efficient production and high profit. Ann. “Constantin Brancusi” Univ. Targu Jiu, Eng. Ser. 2, 45–53.
Prakhyath, K. M., Yogendra, N. D., Arul Prakash, T., Kumar, D., and Anandakumar, T. M. (2024). Resource use efficiency and system productivity of patchouli (Pogostemon cablin (Blanco) Benth) intercropping with food crops. Ind. Crops Prod. 222:119868. doi: 10.1016/j.indcrop.2024.119868
Ragab, M. E., Youssef, S. M., Nasr, S. M., and Al-arabi, H. A. (2014). Strawberry intercropping with some vegetables for economic productivity. Acta Hortic. 1049, 503–508. doi: 10.17660/ActaHortic.2014.1049.74
Rahmann, G. (2011). Biodiversity and organic farming: what do we know? Agricult. Forst. Res. 3, 189–208.
Rodriguez, C., Dimitrova Mårtensson, L-. M., Zachrison, M., and Carlsson, G. (2021). Sustainability of diversified organic cropping systems—challenges identified by farmer interviews and multi-criteria assessments. Front. Agron. 3:698968. doi: 10.3389/fagro.2021.698968
Romaneckas, K., Adamavičiene, A., Šarauskis, E., and Balandaite, J. (2020). The impact of intercropping on soil fertility and sugar beet productivity. Agronomy 10:1406. doi: 10.3390/agronomy10091406
Röös, E., Mie, A., Wivstad, M., Salomon, E., Johansson, B., Gunnarsson, S., et al. (2018). Risks and opportunities of increasing yields in organic farming. A review. Agron. Sustain. Dev. 38:14. doi: 10.1007/s13593-018-0489-3
Rosa-Schleich, J., Loos, J., Mußhoff, O., and Tscharntke, T. (2019). Ecological-economic trade-offs of diversified farming systems—a review. Ecol. Econ. 160, 251–263. doi: 10.1016/j.ecolecon.2019.03.002
Rundlöf, M., Smith, H. G., and Birkhofer, K. (2016). Effects of Organic Farming on Biodiversity. John Wiley and Sons Ltd (Ed.), eLS (pp. 1–7). John Wiley and Sons, Ltd. doi: 10.1002/9780470015902.a0026342
Salaheen, S., and Biswas, D. (2019). “Chapter 2—Organic farming practices: integrated culture vs. monoculture,” in Safety and Practice for Organic Food, eds. D. Biswas and S. A. Micallef (Cambridge, MA: Academic Press), 23–32.
Sánchez, A. C., Kamau, H. N., Grazioli, F., and Jones, S. K. (2022). Financial profitability of diversified farming systems: a global meta-analysis. Ecol. Econ. 201:107595. doi: 10.1016/j.ecolecon.2022.107595
Santos, B. M., Esmel, C. E., Slamova, S., and Golden, E. A. (2008). Optimum planting dates for intercropping cucumber, squash, and muskmelon with strawberry. HortTechnology Hortte 18, 656–659. doi: 10.21273/HORTTECH.18.4.656
Scherr, S. J., and McNeely, J. A. (2007). Biodiversity conservation and agricultural sustainability: towards a new paradigm of ‘ecoagriculture' landscapes. Philos. Transac. Royal Soc. B Biol. Sci. 363, 477–494. doi: 10.1098/rstb.2007.2165
Schrama, M., de Haan, J. J., Kroonen, M., Verstegen, H., and Van der Putten, W. H. (2018). Crop yield gap and stability in organic and conventional farming systems. Agricult. Ecosyst. Environ. 256, 123–130. doi: 10.1016/j.agee.2017.12.023
Selwood, K. E., and Zimmer, H. C. (2020). Refuges for biodiversity conservation: a review of the evidence. Biol. Conserv. 245:108502. doi: 10.1016/j.biocon.2020.108502
Shahane, K., Kshirsagar, M., Tambe, S., Jain, D., Rout, S., Ferreira, M. K., et al. (2023). An updated review on the multifaceted therapeutic potential of Calendula officinalis L. Pharmaceuticals 16:611. doi: 10.3390/ph16040611
Shao, Z., Zheng, C., Postma, J. A., Gao, Q., and Zhang, J. (2024). More N fertilizer, more maize, and less alfalfa: Maize benefits from its higher N uptake per unit root length. Front. Plant Sci. 15:1338521. doi: 10.3389/fpls.2024.1338521
Sharma, R. K., Cox, M. S., Oglesby, C., and Dhillon, J. S. (2024). Revisiting the role of sulfur in crop production: a narrative review. J. Agricult. Food Res. 15:101013. doi: 10.1016/j.jafr.2024.101013
Sidhu, V., Bernier-English, V., Lamontagne-Drolet, M., and Gravel, V. (2022). Effect of light quality and extended photoperiod on flower bud induction during transplant production of day-neutral strawberry cultivars. Can. J. Plant Sci. 102, 356–367. doi: 10.1139/cjps-2021-0081
Simpson, D. (2018). “The economic importance of strawberry crops,” in The Genomes of Rosaceous Berries and Their Wild Relatives, eds. T. Hytönen, J. Graham, and R. Harrison (Berlin: Springer International Publishing), 1–7.
Singh, P., Panotra, N., Singh, K. N., and Kanht, R. (2013). Interaction effect of sulphur and potassium on nutritional quality and profitability of brown sarson (Brassica rapa var brown sarson) in Temperate Kashmir. Indian J. Agron. 58, 91–95. doi: 10.59797/ija.v58i1.4151
Singh, S. P., and Schwan, A. L. (2011). “4.20—Sulfur metabolism in plants and related biotechnologies,” in Comprehensive Biotechnology (Second Edition), ed. M. Moo-Young (Cambridge, MA: Academic Press), 257–271.
Sønsteby, A., and Hytonen, T. (2005). Manipulating flower induction through temperature and photoperiod fluctuations. Int. J. Fruit Sci. 5, 17–27. doi: 10.1300/J492v05n01_03
Soppelsa, S., Gasser, M., and Zago, M. (2023). Optimizing planting density in alpine mountain strawberry cultivation in Martell Valley, Italy. Agronomy 13:1422. doi: 10.3390/agronomy13051422
Stratton, A. E., Wittman, H., and Blesh, J. (2021). Diversification supports farm income and improved working conditions during agroecological transitions in southern Brazil. Agron. Sustain. Dev. 41:35. doi: 10.1007/s13593-021-00688-x
Tagliavini, M., Baldi, E., Lucchi, P., Antonelli, M., Sorrenti, G., Baruzzi, G., et al. (2005). Dynamics of nutrients uptake by strawberry plants (Fragaria × Ananassa Dutch.) grown in soil and soilless culture. Eur. J. Agron. 23, 15–25. doi: 10.1016/j.eja.2004.09.002
Taneja, S. C., and Chandra, S. (2012). “20—Mint,” in Handbook of Herbs and Spices (Second Edition), ed. K. V. Peter (Cambridge: Woodhead Publishing), 366–387.
Tessmer, M. A., Appezzato-da-Glória, B., and Antoniolli, L. R. (2016). Influence of growing sites and physicochemical features on the incidence of lenticel breakdown in ‘Gala' and ‘Galaxy' apples. Sci. Hortic. 205, 119–126. doi: 10.1016/j.scienta.2016.04.027
Thompson, A., Frenzel, M., Schweiger, O., Musche, M., Groth, T., Roberts, S. P. M., et al. (2021). Pollinator sampling methods influence community patterns assessments by capturing species with different traits and at different abundances. Ecol. Indic. 132:108284. doi: 10.1016/j.ecolind.2021.108284
Torres-Quezada, E. A., Zotarelli, L., Whitaker, V. M., Santos, B. M., and Hernandez-Ochoa, I. (2015). Initial crown diameter of strawberry bare-root transplants affects early and total fruit yield. HortTechnology Hortte 25, 203–208. doi: 10.21273/HORTTECH.25.2.203
Tscharntke, T., Grass, I., Wanger, T. C., Westphal, C., and Batáry, P. (2021). Beyond organic farming—harnessing biodiversity-friendly landscapes. Trends Ecol. Evol. 36, 919–930. doi: 10.1016/j.tree.2021.06.010
Tscharntke, T., Klein, A. M., Kruess, A., Steffan-Dewenter, I., and Thies, C. (2005). Landscape perspectives on agricultural intensification and biodiversity—ecosystem service management. Ecol. Lett. 8, 857–874. doi: 10.1111/j.1461-0248.2005.00782.x
Wang, C., Zhou, L., Zhang, G., Gao, J., Peng, F., Zhang, C., et al. (2021a). Responses of photosynthetic characteristics and dry matter formation in waxy sorghum to row ratio configurations in waxy sorghum-soybean intercropping systems. Field Crops Res. 263:108077. doi: 10.1016/j.fcr.2021.108077
Wang, G., Bei, S., Li, J., Bao, X., Zhang, J., Schultz, P. A., et al. (2021b). Soil microbial legacy drives crop diversity advantage: linking ecological plant–soil feedback with agricultural intercropping. J. Appl. Ecol. 58, 496–506. doi: 10.1111/1365-2664.13802
Wang, R., Eguchi, M., Gui, Y., and Iwasaki, Y. (2020). Evaluating the effect of light intensity on flower development uniformity in strawberry (Fragaria × ananassa) under early induction conditions in forcing culture. HortScience Horts 55, 670–675. doi: 10.21273/HORTSCI14917-20
Wang, Z-. G., Jin, X., Bao, X-. G., Li, X-. F., Zhao, J-. H., Sun, J-. H., et al. (2014). Intercropping enhances productivity and maintains the most soil fertility properties relative to sole cropping. PLoS ONE 9:e113984. doi: 10.1371/journal.pone.0113984
Willey, R. W. (1979). Intercropping—Its importance and research needs: part I. Competition and yield advantages. Field Crop Abstracts 32, 1–10.
Willey, R. W., and Rao, M. R. (1980). A competitive ratio for quantifying competition between intercrops. Exp. Agricult. 16, 117–125. doi: 10.1017/S0014479700010802
Wu, W., Chen, L., Liang, R., Huang, S., Li, X., Huang, B., et al. (2025). The role of light in regulating plant growth, development and sugar metabolism: a review. Front. Plant Sci. 15:1507628. doi: 10.3389/fpls.2024.1507628
Xiao, X., Cheng, Z., Meng, H., Liu, L., Li, H., Dong, Y., et al. (2013). Intercropping of green garlic (Allium sativum L.) induces nutrient concentration changes in the soil and plants in continuously cropped cucumber (Cucumis sativus L.) in a plastic tunnel. PLoS ONE 8:e62173. doi: 10.1371/journal.pone.0062173
Yogendra, N. D., Keerthi, P. E., Nazeer, M., Jnanesha, A. C., Verma, R. S., Sundaresan, V., et al. (2024). Livelihood enhancement and resource use efficiency under lemongrass intercropping with food crops. Ecol. Front. 44, 266–274. doi: 10.1016/j.chnaes.2023.06.001
Zhang, F., and Li, L. (2003). Using competitive and facilitative interactions in intercropping systems enhances crop productivity and nutrient-use efficiency. Plant Soil 248, 305–312. doi: 10.1023/A:1022352229863
Keywords: Fragaria x ananassa, land equivalent ratio, competition indices, companion plants, polyculture, alpine climate, biodiversity
Citation: Soppelsa S, Gasser M, Castellan A, Pramsohler M, Zago M, Matteazzi A and Kelderer M (2025) Strawberry-herb intercropping: a 2-year study toward sustainable intensification and diversification. Front. Sustain. Food Syst. 9:1645707. doi: 10.3389/fsufs.2025.1645707
Received: 12 June 2025; Accepted: 15 September 2025;
Published: 17 October 2025.
Edited by:
Santiago Dogliotti, Universidad de la República, UruguayReviewed by:
Mariana Scarlato, Universidad de la República, UruguayN. D. Yogendra, Central Institute of Medicinal and Aromatic Plants, Council of Scientific and Industrial Research (CSIR), India
Copyright © 2025 Soppelsa, Gasser, Castellan, Pramsohler, Zago, Matteazzi and Kelderer. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sebastian Soppelsa, U2ViYXN0aWFuLlNvcHBlbHNhQGxhaW1idXJnLml0