 Cathy Hawes*
Cathy Hawes* Andrew ChristieGillian BanksDavid BoldrinJacqueline Brandt
Andrew ChristieGillian BanksDavid BoldrinJacqueline Brandt Pietro IannettaIsabella SwystIzzy Turner
Pietro IannettaIsabella SwystIzzy Turner- The James Hutton Institute, Dundee, United Kingdom
Agricultural intensification has resulted in arable systems dominated by monocultures that are reliant on agrochemical inputs and frequent tillage to maintain high crop yields. This has caused significant decline in farmland biodiversity and soil health, further increasing the need for chemical inputs to regulate system processes. Regenerative practices aim to reverse this trend and capitalise on biodiversity-driven ecosystem functions that determine the long-term sustainability of agricultural production. However, despite a general acceptance of the potential benefits of this approach, there is currently very little supporting evidence from long-term, field scale experimental data. A whole-systems and nature-based approach for designing and implementing a regenerative cropping system at the Hutton’s Centre for Sustainable Cropping long-term platform has demonstrated the practical application of theoretical, outcomes- and biodiversity-based frameworks in a commercially realistic setting. Best practice management options were combined in a cropping system that, rather than conserving soil and biodiversity at the expense of crop production, aimed to maintain yields with less reliance on agrochemical inputs by simultaneously promoting soil health, crop fitness and biodiversity. Soil physical properties and biological processes were enhanced, plant diversity and the abundance of beneficial plant and invertebrate species were increased, and crop yield was maintained at levels comparable to the national average. A barrier to uptake of low input, regenerative practices is the perception of risk. Data-driven evidence for the positive and negative impacts of regenerative approaches on crop production and the environment is needed for farmers to make informed management decisions. Particularly important is an understanding of the balance between short-term costs and longer-term benefits as the system gradually stabilises and starts to deliver increased resilience to future environmental perturbation. This highlights the need for long-term, whole-system and field-scale studies to provide commercially realistic predictions of risks, costs and benefits for growers wishing to adopt regenerative cropping practices.
1 Introduction
Arable fields make up approximately 20% of the UK land surface area (Scottish Government, 2024). Diversification of this largely mono-cropped habitat represents a massive opportunity for biodiversity restoration and regeneration of agroecosystem functions and services (Hawes et al., 2021). Agricultural intensification has resulted in arable systems dominated by monocultures that are reliant on agrochemical inputs and frequent tillage for crop production, causing significant decline in farmland biodiversity and soil degradation, further increasing the need for inputs to regulate system processes (Tilman, 1999; Sanaullaha et al., 2020). Biodiversity maintains ecological functions that determine economic output and the sustainability of agricultural production. In annually disturbed arable systems, this biodiversity depends heavily on the strategic integration of non-crop plants and semi-natural vegetation. Their diversity supports a wide range of regulating ecosystem services. Flowering plants provide resources to support pollinator populations and therefore pollination of native plants and insect-pollinated crops (Nicholls and Altieri, 2013). These plants also provide nectar for adult forms of larval parasitoids, improving the control of crop pest populations (Crowder and Jabbour, 2014). Enhanced weed cover increases carbon inputs to soil, supporting earthworms and microbial communities (Gaba et al., 2020), and increasing nutrient availability and uptake (Aislabie and Deslippe, 2013). Finally, diverse weed communities can suppress the dominance of competitive weeds, creating a more even and stable community structure and a positive feedback loop to further reduce the need for blanket weed control (Storkey and Neve, 2018).
Regenerative cropping practices capitalise on these regulating ecosystem services that result from the complex network of ecological interactions within diversified agroecosystems. This supports crop production in the long-term whilst relying less on agrochemical inputs (Maeder et al., 2002; George et al., 2022). However, although the potential benefits of this approach are now widely recognised (Bless et al., 2023; British Ecological Society, 2025), scientific underpinning for these largely theoretical principles is scarce (Berthon et al., 2024). Recent developments in regenerative agriculture have tended to be farmer-led rather than driven by research, and based on the need for practical, on-farm solutions to restore natural processes, deliver the multiple benefits demanded by policy, and reduce costs. Very little quantitative data is available in support of these efforts and few studies take the whole-systems, integrative, field-scale and long-term approach necessary to understand the effects of management on environmental and economic outcomes.
This paper describes a long-term and ongoing agroecological experiment, the Centre for Sustainable Cropping (CSC) at Balruddery Farm, Scotland (Hawes et al., 2019). The CSC was established in 2009 based on a framework for regenerative crop system design, aiming to achieve multiple environmental and ecological benefits. The long-term, field-scale and whole-system impacts on diversity-driven ecosystem functions is assessed by monitoring systems indicators. Regenerative systems take a holistic, system-scale approach designed to capitalise on the complex and interconnected nature of the in-field ecological networks and the ecosystem services they provide (Hawes et al., 2021). At the CSC, the regenerative cropping system aims to achieve multiple benefits based on selection of management options that influence the in-field agroecological network (Figure 1): Organic matter amendments (1) and organic inputs from cover crops and weedy stubbles (2) are combined to increase soil carbon content (13). More soil organic matter, together with reduced tillage (3), improves soil structure (14), minimising erosion and run-off losses. Better soil structure and organic inputs provides resources and habitat for below-ground food webs (15, 16). These microbes, microarthropods and invertebrates enhance soil biological activity [particularly decomposition and nutrient cycling processes (17)]. This results in increases in nutrient availability (21) which, alongside nutrient inputs from legumes and sustainable nutrient management (20), and better soil physical conditions for root growth (18), improves resource uptake by crops (19). Above-ground biodiversity is harnessed to minimise reliance on crop protection inputs. IPM strategies (7, 8, 9), reduce non-target effects on beneficial insects (24) which are supported by diverse cover crop mixes and in-field weed communities (25). Natural enemies help regulate populations of crop pests (23), and pollinators (27) improve crop yield and quality of insect-pollinated crops. Taken together, these interactions minimise reliance on fertiliser and crop protection inputs to maintain crop yields (28) and create positive feedback loops for further enhancement of biodiversity, soil health and ecosystem function. This paper describes trends in indicators of these soil, biodiversity and production components of arable cropping systems, comparing the long-term impact of a regenerative approach relative to standard commercial practice over two 6-year crop rotations.
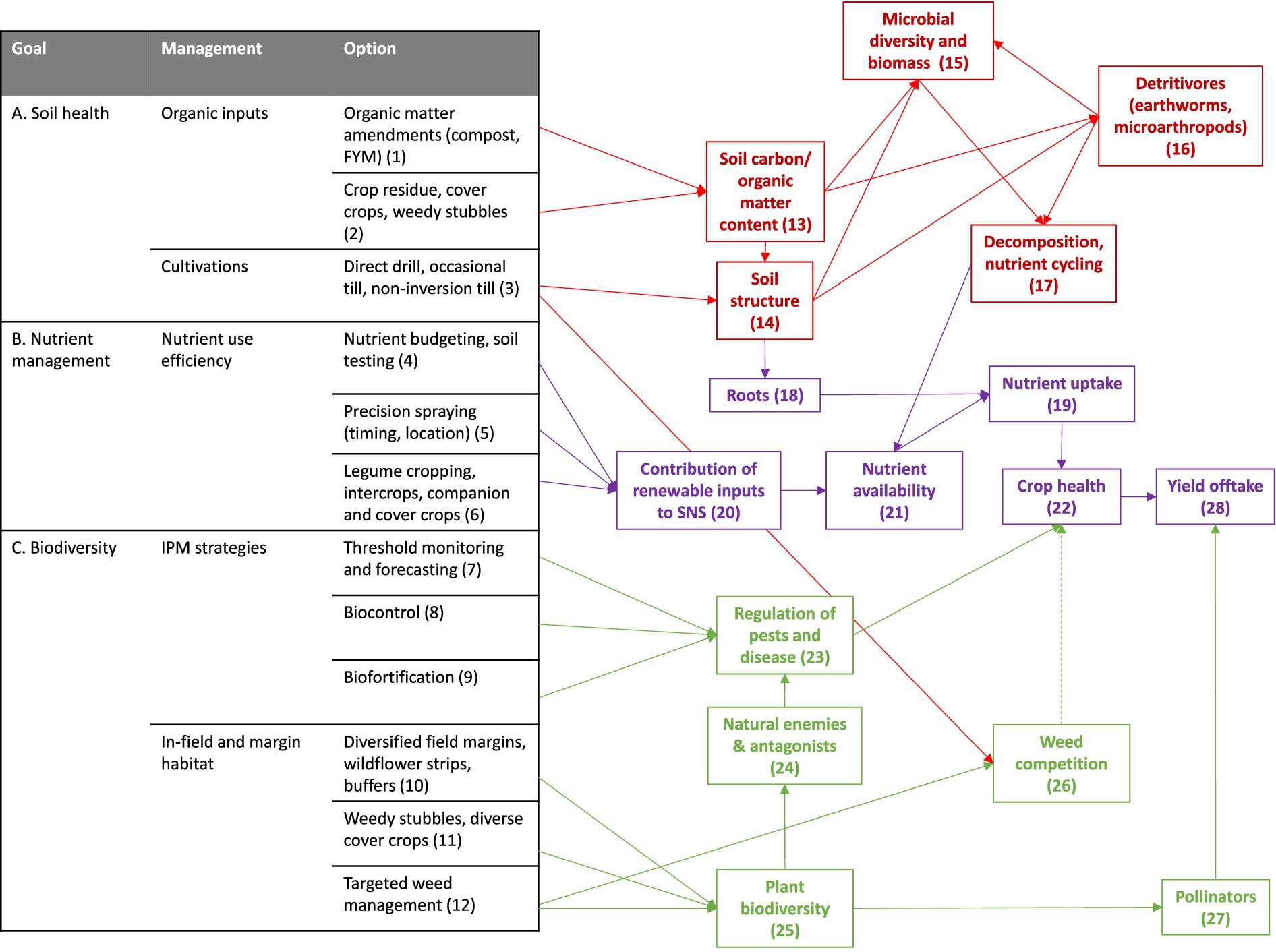
Figure 1. Framework for designing integrated cropping systems for multiple benefits (A–C) based on selection of management options (1–12) that influence the in-field agroecological network: soil biology and structure (red nodes, 13–17); crop nutrition and yield (blue nodes, 18–22, 28), and; biodiversity (green nodes, 23–27).
2 Methods
2.1 Site description and experimental design
The CSC site and experimental design have previously been described in Hawes et al. (2018). Briefly, the CSC long-term platform is a 42-hectare block of six arable fields, established in 2009 at Balruddery Farm near Dundee, Scotland (56.48 latitude, 3.13 longitude) (Figure 2). The farm is in a temperate maritime arable environment, with an average annual rainfall of 800 mm, an annual accumulated temperature of 1,100–1,375 day-degrees C (above 5.6 °C) and a mean annual potential water deficit of 50–75 mm. The area is moderately exposed (2.6–4.4 m s−1 wind speed) and has moderate winters of 50–110 day degrees C of accumulated frost. The soils are imperfectly draining Balrownie Series with an average pH of 5.7. Topsoil depths range from 25 cm to 40 cm, textures from sandy loam to sandy silt loam and stone contents of 10–20% volume.
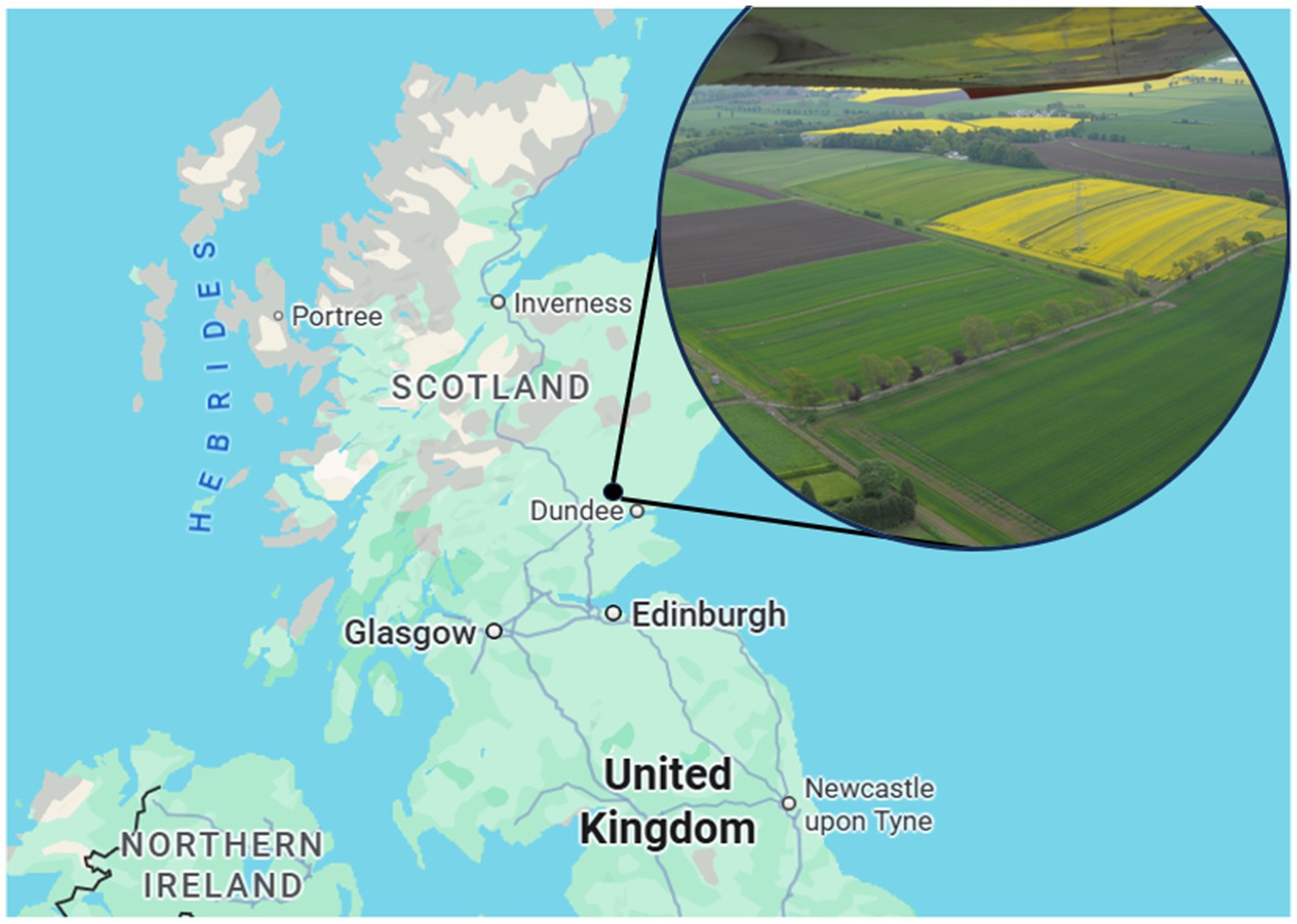
Figure 2. Location and aerial view of the centre for sustainable cropping long-term platform at Balruddery Farm, Dundee, Scotland.
The CSC experiment is a split-field comparison of regenerative cropping against standard agronomic practice over multiple 6-year rotations of potatoes, winter wheat, winter barley, winter oilseed rape, spring sown faba beans and spring barley. Each field was divided into two, with conventional and regenerative treatments randomly allocated to each half and then planted with one of the six crops of the rotation in 2010 (Figure 3).
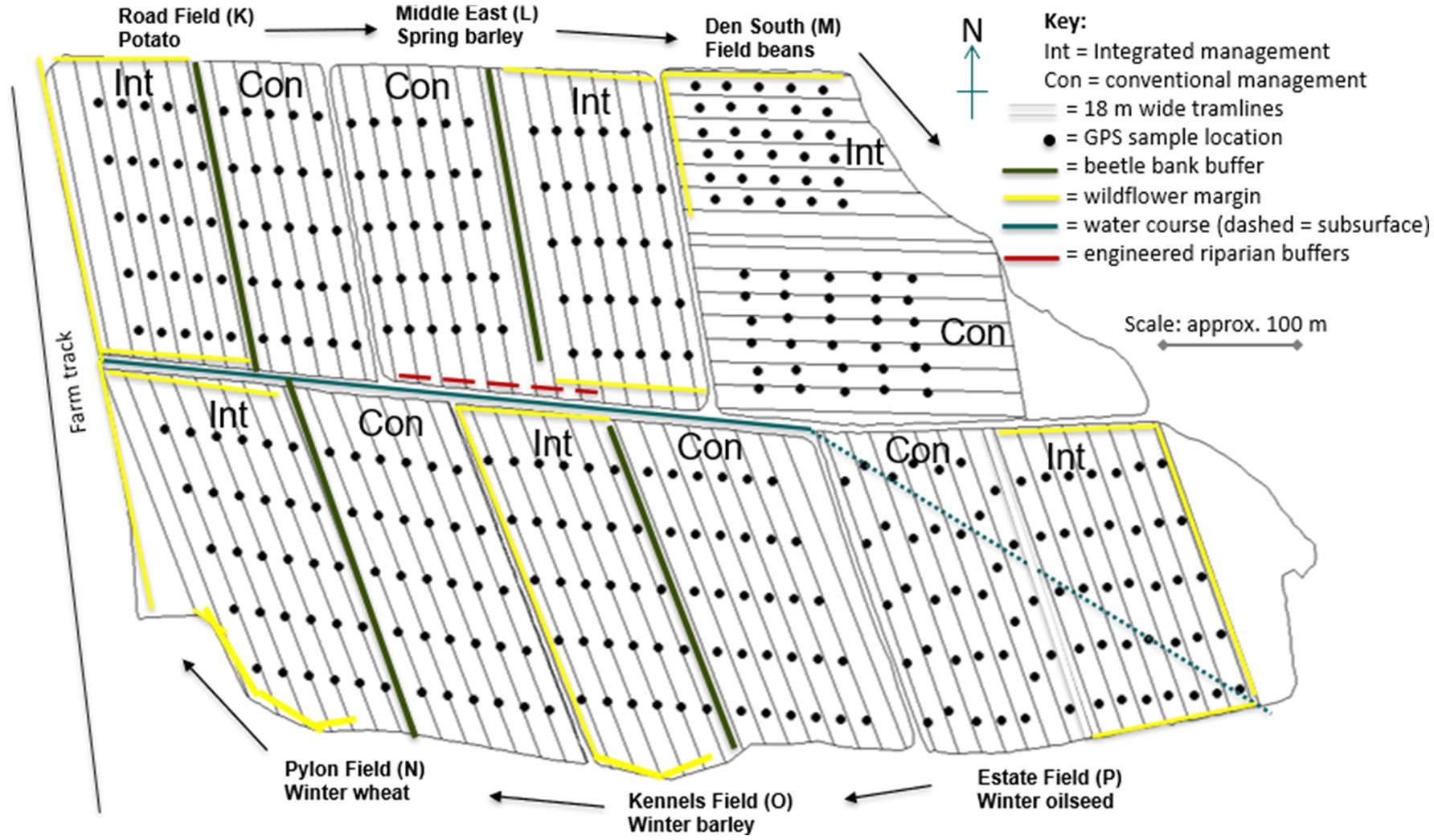
Figure 3. Field layout at the centre for sustainable cropping long-term experiment. Integrated (regenerative, Int) and conventional (Con) crop management systems are randomly allocated to each field half. Crops planted in each field are labelled for year 1 of each crop rotation and move in a clockwise direction (arrows) round the six fields each year. Black dots indicate position of 350 GPS sample locations (50 in Road field, 60 in each of the other five fields), spaced approximately 40 m apart along each tramline with 18 m between rows. Sown margins, buffers and water course are marked.
Replication of the 12 half-field plots (2 treatments × 6 crops) is year-on-year, providing 6 replicates of each crop/treatment combination per rotation. This necessitates more complex statistical analysis to handle potential confounding effect of year and field. However, it allows for large plot sizes (approximately 200 × 100 m blocks) which, unlike small, plot-scale experiments, enables the use of commercial farm machinery for sowing, management and harvest. Data generated on soil, biodiversity, crop and economics are therefore representative of field scale commercial situations, thus allowing realistic estimates of costs and benefits that are relevant to real-world commercial situations.
2.2 Treatments
The conventional cropping system is ploughed annually following standard cultivation practices for the region and each of the crops in the rotation. Cereal straw is baled and removed. Fertiliser inputs are based on Scottish Nitrate Vulnerable Zone recommendations (for N) and an estimate of offtake (for P and K). Crop protection treatments (fungicides, insecticides and herbicides) are prescriptive and prophylactic, following label recommendations and standard commercial agronomy (Hawes et al., 2018). Specific products, active ingredients and input rates vary year on year depending on the crop requirements, field conditions and weather, but agronomic decisions are made with the goal to optimise crop yield and profit margin as in standard commercial, intensive cropping practice.
The regenerative system aims to enhance soil health and biodiversity while maintaining commercially viable crop yields. The goal for soil management is to improve physical structure to minimise erosion and runoff losses, provide better conditions for root growth, and sustain diverse communities of soil organisms to support carbon turnover, nutrient cycling and more efficient resource uptake via fungal and microbial associations. To achieve these multiple targets, the regenerative system at the CSC combines six management strategies into a single cropping system:
• Reduced tillage: disturbance by ploughing occurs only 1 year in six for potato planting and harvest. Winter wheat is sown into the disturbed soil following potatoes, winter oilseed is minimum tilled to ensure good soil-seed contact, barley and bean are direct drilled. This minimises soil disturbance in 5 years of the six and provides the opportunity in the potato year to deal with any weed control or compaction issues that may have arisen in the no-till years.
• Organic matter amendments: green waste municipal compost from Dundee City Council applied at 35 t ha−1 yr.−1 for the first rotation to create a treatment difference between the regenerative and conventional systems, then reduced to a more commercially realistic 10 t ha−1 yr.−1 in the second rotation. Due to supply issues, applications are now at 5 t ha−1 yr.−1 and soil carbon content is being monitored to check that turnover can be maintained at this lower input rate. Additional organic matter inputs are derived internally - from crop residues where cereal straw is retained instead of baled and removed, winter cover crops, and weedy stubbles left over winter if harvest was too late for sowing a cover.
• Cover crops: to retain soil and nutrients over winter and provide additional organic matter inputs. Rye and mixed legume cover crops are sown after winter oilseed rape (before beans), oats are sown after beans (before spring barley), and rye is sown after spring barley (before potato). Field beans are generally harvested too late in this region for cover crop establishment. Instead, methods for undersowing the growing bean crop with clover to provide continuous post-harvest cover over winter are being investigated.
• Companion cropping: Berseem clover is undersown as a temporary companion crop for winter oilseed establishment. White clover is sown with spring barley to increase nitrogen supply from Biological Nitrogen Fixation. A diverse, legume-based species mix is sown after potato planting to reduce aphid colonisation and enhance natural enemy control in potato crops.
• Sustainable nutrient management: soil nitrogen supply (SNS) is measured as the concentration of soil mineral nitrogen in soil samples collected in March each year from each field. Mineral N concentration is converted to kg ha−1 to at 20 cm depth, accounting for bulk density, and used to calculate reduction in fertiliser rate. This optimises efficiency of fertiliser use, minimising the risk of surplus and therefore reducing losses through leaching and run-off.
• IPM strategies: Threshold crop protection inputs and disease forecasting (blight models using the Hutton Criteria) are used to reduce fungicide use. Mineral nutrition is applied to winter cereals to increase plant fitness and resilience to pests and disease, further reducing reliance on chemistry. Targeted herbicide applications aim to maintain weed densities at around 10% cover, below competition thresholds but sufficient to support beneficial insects.
As with the conventional treatment, this regenerative system is flexible rather than fixed, based on indicator monitoring and an iterative process of performance review against management targets. This enables improvements in sustainability outcomes over time through modification of existing interventions and incorporation of new best practice options within the overall regenerative strategy. Full details of all agronomic interventions can be made available on request.
2.3 Indicator monitoring
Long-term trends in indicators of soil, biodiversity and crop production are monitored to assess whole-systems impact of the regenerative system relative to conventional practice. Sampling was carried out annually between 2011 and 2024 following standardised assessment protocols at GPS-located sample points across the 12 treatment x crop combinations shown in Figure 3. Due to resource constraints, not all indicators were measured in all years at all sample locations. A list of the indicators reported here is given in Table 1, cross-referenced to the nodes in the agroecological network in Figure 1, along with their frequency and intensity of sampling.
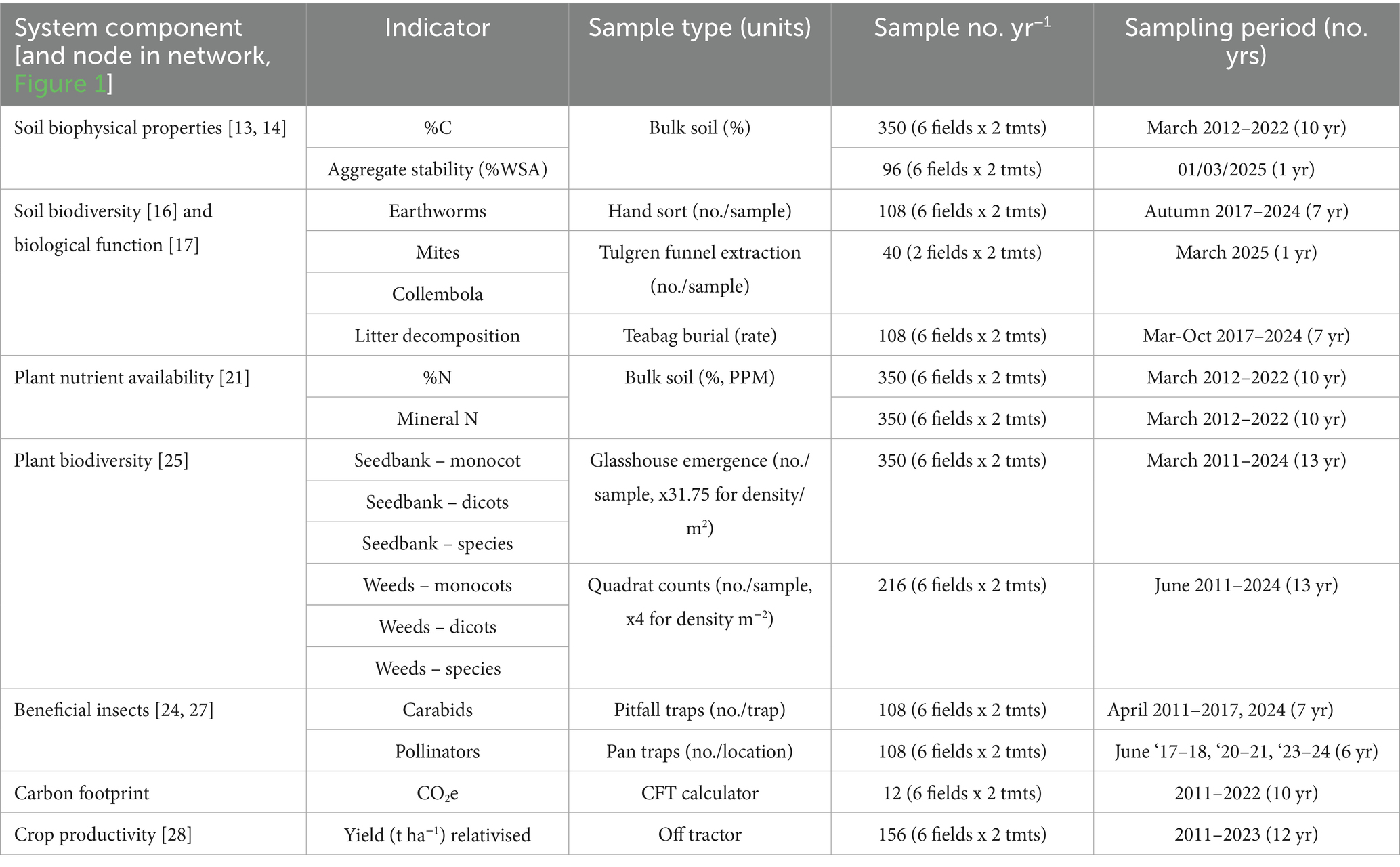
Table 1. Systems indicators monitored at the CSC long-term platform, grouped by system component (number refers to node in Figure 1) and showing sampling intensity and frequency.
Soil carbon (%C) and aggregate stability (%WSA) were measured as indicators of soil structure, both predicted to increase in the regenerative cropping systems relative to standard practice. Soil carbon was measured across the six fields in March before spring crops were sown from 2012 to 2022 (10 years data). Approximately 1.5 kg of bulk soil was sampled to a depth of 15 cm at each of 350 permanent GPS sample locations every year. Samples were sieved to 10 mm and stones exceeding 10 mm in diameter were removed. A sub-sample of sieved soil was then put through a 2 mm sieve, dried at 70 °C for 24 h, milled using a Retsch mill and 5 mg weighed for analysis using an Exeter Analytical CE440 Elemental Analyser (EAI, Coventry, UK) which determines the carbon, hydrogen, and nitrogen content in organic and inorganic compounds by combustion.
Aggregate stability was measured as the proportion of 2-8 mm diameter soil aggregates that remain intact when immersed in water (Le Bissonais, 2016). Eight bulk soil samples were collected from each treatment across the six CSC fields in March 2025. Soil from each of the 96 sample locations was air dried and a 5 g sub-sample was passed through an 8 mm sieve. The 8 mm sieved soil subsequently retained on a 2 mm sieve was used to test for water stable aggregates (%WSA). In a wet sieving apparatus (Eijelkamp Soil and Water, Netherlands), 2 mm-mesh sieves filled with the air-died soil are mechanically moved up and downward (up-down strokes = 1.3 cm) in a can filled with distilled water for 3 min ± 5 s (34 times/min). Unstable aggregates disintegrate and pass through the 2 mm sieve into the water-filled can under the sieve. The cans with soil from unstable aggregates were then removed and replaced by new water-filled cans. All stable aggregates retained on the sieve mesh were then fully disintegrated and soil from the stable aggregates was collected into the second set of water cans. Note that gravel, stones, plant residuals and roots remain on the sieve and only the soil from the aggregates passed into the water cans. After drying the two sets of cans with the soil from unstable and stable aggregates, %WSA is calculated as the proportion of stable aggregates in the sample (weight of the stable aggregates divided by the sum of both the stable and unstable aggregate fractions).
Earthworms were sampled from 9 locations in each treatment across all 6 fields in spring (April, after spring crops were sown) and autumn (October/November, after autumn crops were sown) each year from 2017 to 2024 (108 samples each year for 7 years). Sampling was carried out when soil was damp but not waterlogged or frozen. A 20 × 20 × 20 cm (8,000 cm3) soil pit was dug out at each location, placed on a white tray and hand sorted for earthworms. Juveniles were counted and returned to the pit. Adult earthworms were separated into three ecological functional groups following Stroud (2019): epigeic (surface active red coloured, small <8 mm long, found in leaf litter), anecic (topsoil dwellers, small-medium sized, pink, grey, green or mottled yellow), or endogeic deep vertical burrows, large >8 mm with red or black head. Numbers in each group were recorded, specimens photographed for future reference and then returned to the pit.
Soil microarthropods were collected from two pooled soil cores, using cylinders of 10 cm depth by 5 cm diameter (sample volume 392 cm3), taken at 10 GPS sample locations in each half of fields L (winter wheat) and K (winter barley), one- and two-years following potato cultivation, respectively. All 50 soil samples were collected within a week of each other at the end of March 2025. Soil was stored in a cool box after sampling and placed in Tullgren funnels (Capinera, 2008) within 4 h of collection. Soil microarthropod extraction followed similar procedures to Crossley and Blair (1991) but used 25 Watt bulb Tullgren funnel banks. Extraction into collecting jars of 70% ethanol occurred over 6 days and counts of Acari and Collembola were conducted under a stereo microscope.
Decomposition rates were estimated using the teabag method (Keuskamp et al., 2013) where 3 replicate green tea (Lipton EAN no.: 8722700 055525) and Rooibos tea (Lipton EAN no.: 8722700 188,438) teabags were dried at 70 °C for 48 h, weighed, labelled and buried 10 cm deep in 2 rows of 3 tea bags at 9 GPS sample locations in each of the 2 treatments across the 6 crops in the rotation. Sampling was carried out during the cropping seasons 2017, 2018, 2019, 2021 and 2024, 6 to 13 years from conversion to regenerative cropping. Tea bags were installed in April after spring crops were sown and retrieved approximately 90 days later after crop harvest. Loose soil and roots were gently removed, and the tea bags were then dried at 70 °C for 48 h before re-weighing. Initial weight, final weight and exact number of days of burial were used to calculate rate of litter decomposition for each tea bag following the formula described in Keuskamp et al. (2013). Higher values represent faster decomposition rates by microbes. This procedure measures decomposition by microbial activity since the tea is contained within a mesh too small to allow access by microarthropods or earthworms.
Taxonomic and functional diversity of species in the seedbank are an indicator of the long-term filtering effects of management on biodiversity. At the CSC, the arable weed seedbank was monitored by seedling emergence from soil samples taken across all 350 GPS sample locations in March over a 13-year period (2011–2024). Soil samples were sieved to 10 mm and approximately 1.2 L of the sieved sample placed in a 15 × 21 cm seed tray to a depth of 4 cm. Seed trays were arranged randomly in an unheated glasshouse on benches fitted with capillary matting, kept moist via an overhead automatic misting system. The volume and depth of soil and the general conditions for emergence, were similar to those in historical studies (Roberts, 1958) and more recent assessments carried out at the Hutton (Heard et al., 2003; Hawes et al., 2010). As seedlings emerged, they were identified to species, recorded and removed. The first flush of seedlings was considered complete after about 4 weeks, when there was no further emergence for a period of 14 days. The soil was then re-sieved, placed back in the seed trays and the procedure repeated for a second flush of emergence to ensure that the majority of viable seeds in each sample were recorded. The total number of weed seedlings per species per sample was converted to number per m2 field surface to a depth of 0.15 m using the conversion factor 31.75 based on tray surface area.
The arable weed flora, germinating from annually disturbed soil seedbanks, provide resources for above- and below-ground foodwebs, supporting arable biodiversity within cropped fields. Weed densities were assessed in June each year from 2011 to 2024. The number of individual plants per species were recorded in 18 × 0.5m2 quadrats per half field (providing 216 data points per year for 13 years).
Pollinating insects were sampled using nine sets of three 20 cm diameter coloured pan traps (blue, white and yellow) within the cropped area of each field half (integrated and conventional management) across 6 crops in June 2017, 2018, 2020, 2021, 2023, and 2024. Pan traps were positioned at the height of the crop canopy and filled with 100 mL per trap of dilute Teepol® detergent. Traps were set at 10 a.m. on a sunny day, not less than15 °C with a wind speed under 5 on the Beaufort scale. Traps were collected at 4 p.m. on the day of sampling and the insects sampled from the three traps at each location were combined into a single sample and stored in 70% ethanol until processing. Numbers of specimens in all pollinator groups (bumblebees, solitary bees, honey bees and hoverflies) were counted and recorded.
Carabids were sampled by pitfall trapping in April every year from 2011 to 2017 and again in 2024. In each field half, nine pitfall traps were installed 10 m apart in a line perpendicular to the field margin into the cropped area, starting 20 m from the field edge. This provides a sample number of 108 for the cropped area each year across all 6 fields. Pitfall traps comprised white plastic cups 8 cm deep and 7 cm diameter containing 80 mL antifreeze and inserted into the ground such that the rim of the cup was level with the ground surface. A rain cover was positioned over the top of each trap raised 5 cm above the ground surface on wire legs to reduce the risk of overflowing in heavy rain. Traps were left to run for 14 days before collection. All arthropods sampled were then transferred to tubes of 70% ethanol for storage, carabids were identified to species and counted.
Finally, agronomic indicators included crop yield and carbon footprint. Crop yields were recorded as weight of grain from the combine and potato offtake by tonne box for each 18 m wide tramline (five in each half of Road field, and six in each half of the remaining five fields). Data were converted to tonnes ha−1 yr.−1 based on sown area. Agronomic impacts on carbon footprint were estimated using the Cool Farm Tool (coolfarm.org version 2.29.0). This online calculator estimates CO2 equivalents (CO2e in kg ha−1 yr.−1) for all inputs, fuel use and agronomic operations to assess relative differences between crops and treatments in on-farm (direct) greenhouse gas emissions and soil carbon sequestration. Input data comprised basic field information (cultivated area, soil type), soil organic matter, fertiliser type (NPK, compost etc), fertiliser and compost application rates, crop protection inputs (fungicide, insecticide and herbicide type, application rates and % active ingredient), and field operations energy use (machine type (drill, plough, sprayer, combine etc) and number of operations). Based on these data, the carbon footprint was calculated for every crop, year and treatment providing 144 comparisons over two crop rotations.
2.4 Data handling and statistical analysis
All datasets were double-punched, checked, validated and entered into an SQL relational database for archiving. Data were analysed using Genstat 23rd edition and data used in this analysis will be made available via Zenodo.
To account for year-on-year replication in trial design, a Generalised Linear Mixed Model (GLMM) was used to test for treatment and crop or field effects under the composite null hypotheses:
1. Total crop yield, soil % carbon, weed seedbank monocot and dicot seed densities and seedbank species richness within a field are not significantly affected by the cumulative effects of regenerative cropping relative to standard commercial practices over two crop rotations.
2. Total soil % nitrogen and plant available nitrogen, earthworms, litter decomposition rates, emerged weeds, carabids, pollinator abundance and carbon footprint are not significantly affected by regenerative management practices relative to standard commercial cropping in any of the six crops in the rotation.
For (1), the effect of management treatment on each indicator can vary with field and the cumulative effects of previous cropping and management history are more important than current crop type in determining the treatment response. For these indicator variables, Treatment and Field were therefore declared as fixed effects, and Year and Crop included as random effects to account for any confounding effect of year and crop on the interaction. The application of this model is indicated by (F) in Table 2. For (2), the current crop type and its management is more important than the field itself in determining immediate responses of each indicator to treatment. In these cases, Treatment and Crop were declared as fixed effects, and Year and Field as random effects. This model is applied to the indicators marked (C) in Table 2. Aggregate stability and the abundance of mites and collembola were only measured in a single year. In these cases, the confounding effect of crop and field cannot be distinguished (indicated with * in Table 2). This remains a limitation for interpretation of results for these three indicators until further years’ data can be included in the analysis.
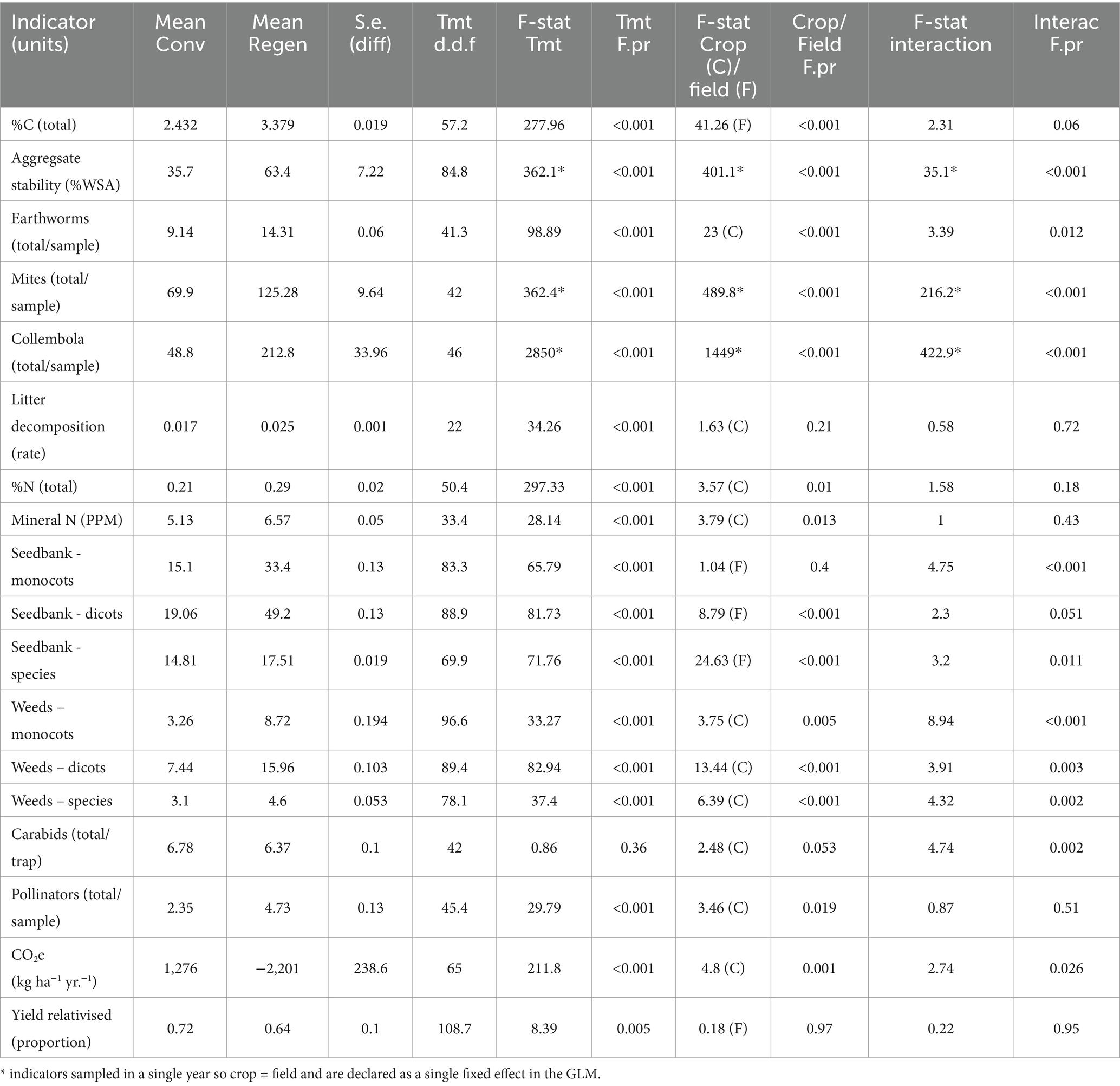
Table 2. Back-transformed means, standard errors of treatment effects, treatment degrees of freedom and F-statistics for fixed effects from Generalised Linear Mixed Models of each indicator variable from Table 1.
One-sample Kolmogorov–Smirnov (K-S) tests were applied to each indicator dataset to determine degree of conformity to a normal distribution. For all indicators K-S p-values were <0.05 suggesting significant difference from a normal distribution. Apart from yield, no data transformations were performed. Instead, a Poisson distribution with log-link function was applied in each GLMM analysis to avoid assumptions of normality. To account for inherent differences in tonnes ha−1 offtake of tubers, grain and oilseed, yield comparisons were made by converting t ha−1 for each harvested strip in each year to the proportion of the maximum yield recorded for that crop across all years and both treatments. This relativised data was used to compare the effect of cropping system on overall crop productivity. Actual yields for each crop over time are shown separately.
Back-transformed mean values for each indicator by treatment, along with standard errors of the treatment difference, degrees of freedom, the F-statistics and probabilities for fixed effects and their interactions are given in Table 2.
To test for associations between weed density and pollinator activity, and between weed density and crop yield, Generalised Linear Regression models were applied with a Poisson distribution and log-link function. The fitted models were Dicots + Treatment + dicots.treatment with crop and year as random effects and pollinators or relativised yield as the response variables.
Finally, the whole-systems response to cropping system, incorporating all indicators together, was assessed using Canonical Variance Analysis (Genstat 23rd edition) with Treatment combined with crop rotational stage as the grouping factor. Rotational stage was defined on the basis of results from the GLMM where significant interaction between treatment and crop type indicated different responses with time since potato cultivation for many of the indicators measured. Rotation stage 1 was therefore classified as the potato crop and following winter wheat crop (sown into ploughed field soil). Rotation stage 2 were the remaining cereal, oilseed and bean crops (direct drilled 3 years or more after potato cultivation). This gave 4 groups (conventional vs. regenerative each divided into two rotational stages, early and late).
3 Results and discussion
The effects of 15 years of regenerative cropping on indicators of within-field biodiversity, soil health, carbon footprint and crop production are presented in Table 2 and discussed below in relation to the agroecological functions represented in Figure 1.
3.1 Soil structure
Soil structure is determined largely by soil type, level of disturbance and organic matter content. Reduced tillage and organic matter inputs in the regenerative treatment resulted in soils with higher carbon content, averaging 3.4% in the regenerative treatments compared to 2.4% under standard practice across all six fields. Significant differences were also detected between fields with K, L and M fields having less %C than N, O and P fields (Figure 3), possibly due to topography resulting in more erosion risk in the former compared to the latter. No significant interaction between treatment and field was detected over the ten-year period.
Increased soil carbon and reduced disturbance by tillage resulting in significantly greater aggregate stability (%WSA) in the regenerative soils (63.4%) compared to conventional (35.7%). Although this indicator was only sampled in 1 year, %WSA was significantly lower in the potato field and following winter wheat crop than in crops with a longer period since the last potato cultivation (Figure 4A). Intensive soil disturbance for potato planting and harvest had a negative impact on soil structural quality, bringing the %WSA down to just 6–7% in the first 2 years following conventional potato cultivation. This effect was less in the regenerative soils (16 %WSA in the first year recovering to 42% in the second year and >90% thereafter), indicating the potential for regenerative practices to mitigate some of the negative effects of potato planting and harvest on soil structure. Further monitoring is required to separate field from crop effects, but this interpretation is supported by Bartlová et al. (2015) who demonstrated positive associations between reduced tillage and %WSA, humic content and crop yields. They proposed a 5-point classification scale of structural quality from very low to very high based on %WSA. Averaged across all fields, CSC conventional soils fall into the “medium” category (34.1–50 %WSA) and the regenerative soils had a %WSA falling within the “very high” category (>66 %WSA) after 3 years of no-till. This suggests that regenerative practices incorporating more organic matter and minimising soil disturbance result in better structural quality.
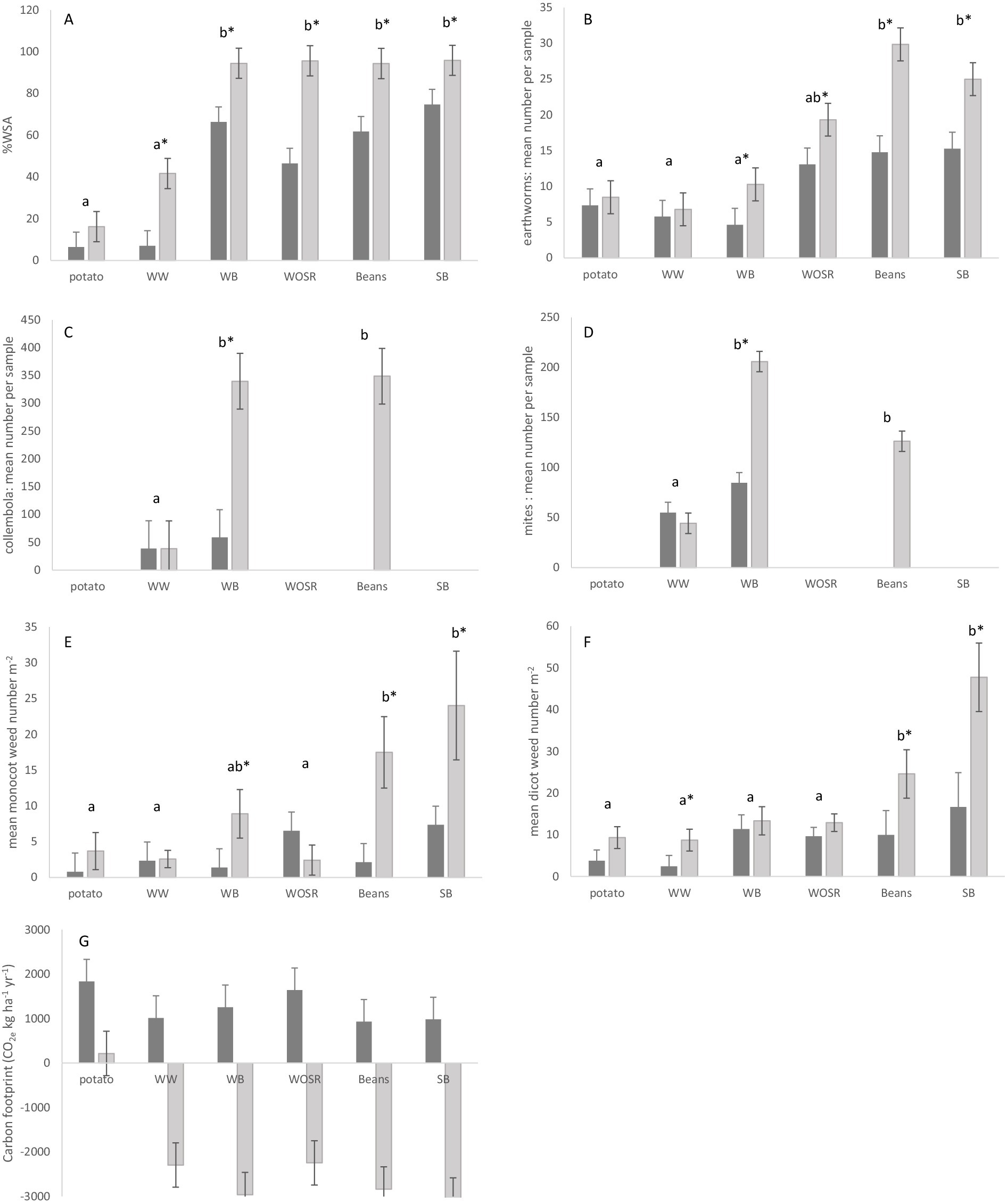
Figure 4. Differences in indicators of soil structure and biodiversity between treatments over the crop rotation: aggregate stability (A), abundance of earthworms (B), Collembola (C) and mites (D), and densities of monocot (E), dicot (F) weeds and (G) carbon footprint. Conventional treatment (dark shaded bars); Integrated treatment (light shaded bars). Crop rotation: yr. 1 potato, yr. 2 winter wheat (WW), yr. 3 winter barley (WB), yr. 4 winter oilseed (WOSR), yr. 5 faba beans and yr. 6 spring barley (SB). Data are back transformed mean values per sample (with standard errors). * = significant treatment differences. Letters denote significant differences between crops (p < 0.005).
Structural quality is important for water infiltration and water holding capacity, lessening risk of both waterlogging and drought, minimising erosion losses and improving the soil conditions for root growth (temperature, moisture and strength) (Franzluebbers, 2002). Informal observations at the CSC indicate that water logging over winter in the regenerative treatments may be less than in conventionally managed soil. However, direct measurement of root growth parameters in relation to these variables are needed to test this hypothesis and determine whether improvements in the regenerative system are agronomically significant in terms of improving crop resilience to extreme weather events.
3.2 Soil biodiversity and biological processes
Soil structural quality not only improves conditions for plant growth but also provides a better environment for soil microbes and arthropods: organic matter fuels below-ground foodwebs and increased soil pore size diversity creates environmental conditions for a diversity of soil organisms responsible for a range of soil functions (Hartmann and Six, 2023). Beneficial soil organisms include arthropod detritivores which break down dead plant and animal matter, microbial decomposers which are important in nutrient cycling processes, and symbiotic fungi and bacteria which improve plant nutrient availability and uptake. Microbial foodweb diversity also helps regulate pathogen populations by competition with neutral antagonists, potentially enhancing the pest suppressive properties of the soil (Bonanomi et al., 2018). In cultivated soils, these functions are reduced compared to natural, undisturbed ecosystems (Creamer et al., 2015). However, the impact of agricultural intensification on soil biota can be minimised in less intensively managed systems, depending on specific management options in relation to species’ functional traits [life history, body size, burrowing depth, habitat and food preferences (Postma-Blaauw et al., 2010)].
Earthworms contribute to soil functions such as structural maintenance, nutrient cycling, organic matter formation, decomposition and water infiltration (Frazão et al., 2017) and can increase crop yields up to 25% (van Groenigen et al., 2014). Non-inversion tillage can increase earthworm abundance to levels comparable to those found in undisturbed field margins (van Groenigen et al., 2014). Less intensive management options particularly reduced tillage and retention of crop residue are therefore needed to enhance earthworm populations and earthworm-related functions within arable fields (Stroud et al., 2017).
At the CSC, total number of earthworms averaged 9.1 per sample (225 m−2) in conventional and 14.3 (358 m−2) in regenerative soils across crops, years and seasons, but with differences between treatment response and overall abundance according to crop sequence. The most abundant category of earthworms were juveniles sampled in autumn which made up 60% of all earthworms sampled. This group showed the largest difference between treatments and crops through the rotation (Figure 4B). Fields cultivated for potato and those in the following winter wheat crops had the lowest number of earthworms and no significant difference between cropping systems. Two years after potato, in the winter barley crops, earthworm numbers were still significantly lower than in beans and spring barley at the end of the crop rotation but showing recovery in the regenerative system and a significant difference between treatments. By the fourth year after potato cultivation, the number of earthworms appeared to have recovered from disturbance during potato planting and harvest, and numbers sampled were higher in the regenerative system and remained higher than the conventional crops for the rest of the crop rotation.
3.2.1 Microarthropods
Two main groups of microarthropods, Acari and Collembola are highly abundant in soils (Petersen and Luxton, 1982) and perform a wide range of functions within the soil foodweb; as predators, microbivores, herbivores and detritivores (Crossley et al., 1992), contributing to ecosystem services such as nutrient cycling and pest control (Neher and Barbercheck, 2019; Bonfanti et al., 2025). They occupy preexisting pore spaces in the soil and surface litter (Rusek, 1998) and so are positively affected by improved soil structure where pore size diversity is greater and ground cover present. Ploughing, fertiliser and insecticide use negatively impact microarthropod abundance (Cortet et al., 2002; Cao et al., 2011) which are therefore predicted to be lower in the conventional treatments.
The mite community was dominated by Prostigmata which made up 76% of the total number sampled, followed by Mesostigmata at 11% with the remaining groups occurring at frequencies less than 5% of the total. Collembola comprised primarily Entomobryidae (80% of the total number of Collembola) and Poduromorpha (17% of the total). Low numbers of Acari and Collembola sampled in the winter wheat following potato cultivation suggests that both groups are negatively affected by soil disturbance in the previous year and the absence of a treatment difference indicates that long-term regenerative practices do not mitigate this negative impact (Figures 4C,D). However, in the regenerative treatments, collembola and mite populations were significantly greater (8x and 4x respectively) 2 years after potatoes than the field 1 year after potato harvest, and densities were comparable to reference samples taken in the regenerative treatment 4 years following potato. This contrasts with a difference of just 1.5x in the conventional field halves between years one and two after potato cultivation. Although treatment effects are statistically significant, more years’ data are required to confirm temporal trends through the rotation and control for any confounding effect of field. However, these microarthropods appeared to show a positive response to reduced tillage and more rapid population recovery than earthworms which took 4 years to return to pre-cultivation numbers.
3.2.2 Litter decomposition
A key agroecosystem function needed to maintain yields in low input cropping systems is the decomposition and mineralisation of nutrients from dead organic matter (Beare et al., 1992). Earthworms and microarthropod detritivores break down dead organic matter, providing material for microbial decomposers. The increased numbers of earthworms and microarthropods in regenerative soils at the CSC is therefore likely to be associated with faster rates of litter decomposition driven by higher levels of soil microbial activity. Assessment of the CSC soil microbiome and quantification of specific microbial responses to field management requires dedicated resources and is being addressed separately. Here, estimates of microbial-driven litter decomposition rates are used as a simple indicator of soil biological function.
Decomposition rates were significantly faster in the regenerative system compared to conventional soils (Table 2). However, unlike earthworm and microarthropod numbers, there was no significant difference between crop types, nor any interaction between crop and treatment. Greater concentrations of soil organic matter and increased detritivore populations in the regenerative treatments therefore appears to be associated with increased rates of decomposition by microbes. However, unlike the microarthropods and earthworms, soil disturbance during potato planting and harvest had no negative effect. Microbial-driven rates of litter decomposition was therefore a good indicator of soil organic matter content but was not impacted by intensity of disturbance.
Greater microbial activity in the regenerative treatments across all crops is expected to increase the rate of nitrification and conversion of ammonia to plant available nitrate, thus contributing to maintaining crop productivity with less reliance on mineral fertiliser. Mineral N (NO3 mg N kg−1 + NH4 mg N kg−1) from soil samples taken across all fields in March each year from 2012 to 2022 and analysed by Yara Analytical commercial laboratory services, averaged 5.13 mg N kg−1 in conventional soils and 6.57 mg N kg−1 in regenerative systems. Total soil %N was measured from the Elemental Analyser alongside %C (section 2.3). Both mineral N and total %N were significantly greater in the regenerative soils compared to conventional, following the same trend across treatments through the rotation as soil carbon and maintaining a constant C: N ratio. Soil Nitrogen Supply (SNS) was therefore improved in regenerative treatment and these values were used to calculate field and crop specific mineral fertiliser requirements at the start of each growing season. This enabled a reduction in mineral nitrogen inputs to the crops in the regenerative treatments of 40–60% of the standard fertiliser rates applied to the conventional crops. Similar results were found in a meta-analysis of long-term studies where crop diversification, cover crops and organic matter inputs were able to substitute nitrogen supply and increase yields in systems with low fertiliser inputs (Maclaren et al., 2022).
3.3 Above-ground biodiversity
3.3.1 Arable weeds
Within cultivated fields, weeds are the primary source of plant diversity and crucial to the functioning of arable systems. They provide floral, seed and vegetative resources for above and below ground foodwebs and associated ecosystem services (Marshall et al., 2003; Smith et al., 2020). Weed seeds can persist in the soil for decades, providing a degree of resilience to disturbance and a reservoir for regenerating biodiversity following agricultural intensification (Brenchley and Warington, 1933; Rotchés-Ribalta et al., 2020). Although weeds are an important component of biodiversity, delivering a range of ecosystem services, they also represent a significant burden to crop yield. Traditional management aims at eradication, but this tends to select for highly competitive and resistant species which are difficult to control, resulting in a disproportionate impact on beneficial species, declining biodiversity and loss of ecosystem function (von Redwitz et al., 2025).
Arable cropping makes up approximately 20% of the total UK land area and, being largely mono-cropped with little or no understorey flora, represents a massive and currently underutilised opportunity for reversing biodiversity loss and enhancing system functions at a national scale. Tolerance of some weed cover within fields is therefore essential for biodiversity conservation and the maintenance of within-field processes (Franke et al., 2009). The challenge is to define the optimal density and composition of the weed flora that supports a healthy agroecosystem but without detrimental impact on crop yield or product quality.
In annually disturbed cropped systems, the in-field flora is dependent on annual regeneration from the soil seedbank. It is therefore important to apply management filters to the above-ground vegetation that result in seedbank communities comprising primarily of beneficial species with low numbers of pernicious weeds (Ryan et al., 2010). In theory, this can be achieved by varying the timing of disturbance (cultivation and herbicide) in relation to periods of flowering and seed set by the beneficial and competitive weed species present. In practice, this is difficult to achieve without better knowledge of the impact of community-level functional trait composition on weed-crop competition thresholds. At the CSC, the integrated crop system currently uses a semi-targeted approach, omitting pre-emergence herbicides to allow a low understorey of weeds, and using post-emergence sprays only where the emerged weed density exceeds around 10% ground cover mid-season. This generally occurs in fields where the seedbank populations of competitive weeds are particularly high, or after several years of direct drilling where weed densities exceed threshold levels at the soil surface before cultivation for potato provides an opportunity for burial.
Seedbank species richness and the densities of monocot and dicot weeds were greater in regenerative crops compared to conventional practice across all six fields of the CSC (Table 2). Field effects were also observed with fewer broadleaved weed seeds in K field and fewer grass weeds in both K and L fields. These fields may be subject to greater risk of erosion and losses of soil and associated seedbank over time (Lewis et al., 2013). Some significant interactions between crop and treatment were observed with smaller treatment differences in fields with low seedbank densities, even though the direction of effect was consistent across fields.
Seedbank species appeared to form distinct community compositions, with different sets of species dominating in each field: Brassica napus volunteers and Senecio vulgaris (field K), Veronica arvensis and Myosotis arvensis (L field), Matricaria spp. and Viola arvensis (M field), Gnaphalium uliginosum and Veronica persica (N field), Capsella bursa-pastoris and Veronica arvensis (O field), Matricaria species (P field). Differences in the functional response and effect traits of these species (time to flowering, shade tolerance, germination requirements, resource provision for pollinators etc.) in relation to timing and intensity of disturbance will form the focus of further analysis of these datasets on the relation between management filters, species composition and functional diversity.
Overall, mean weed densities across all fields and crops were greater in the regenerative cropping systems compared to conventional practice (Table 2). Weed densities in the conventional treatments remained fairly constant through the crop rotation (Figures 4E,F). In the regenerative treatments, a gradual build-up of weeds was observed from low numbers in the potato crop through to highest densities in beans and spring barley after 4 years of no-till cropping.
These observations suggest that occasional tillage (ploughing 1 year in six) is an effective rotational strategy for maintaining a balance between managing weeds below competition threshold (see section 3.4 for yield effects) while tolerating some weed understorey for biodiversity gain. Further research is needed to identify reliable and cost-effective cultural methods for targeted management of specific beneficial weed functional groups. For example, varying the type, intensity and timing of disturbance events could selectively target competitive species while supporting functionally diverse weed assemblages. Communities made up of a diversity of beneficial species may also have an antagonistic effect on dominant, competitive weeds (Adeux et al., 2019) and, with application of appropriate weed management filters, can further reduce the impact of difficult to control species to achieve a long-term balance between biodiversity and crop productivity.
3.3.2 Pollinators and natural enemies
The abundance and diversity of pollinators and nectar feeding adults of parasitoid and diptera larvae natural enemies are dependent on the provision of floral resources in and around arable fields. Intensive cropping practices have negative consequences for natural enemy population dynamics and plant-pollinator networks (Powney et al., 2019). This results in poorer natural control of crop pests and lower pollination rates (Biesmeijer et al., 2006). Pollinator diversity is also crucial for the survival of native plants, particularly rare and declining species. Rare plants share pollinators with more common plant species, the latter providing insects with continuity and extended seasonal resource abundance of nectar and pollen through the year (Gibson et al., 2006), emphasising the need for a diverse and balanced arable plant community (Simba et al., 2018).
Low-intensity management systems that increase within-field and field margin resource for pollinators are therefore likely to promote pollinator diversity (Nicholls and Altieri, 2013). Provision of floral resources as part of Agri-Environment Schemes have been largely successful in terms of promoting pollinator abundance within semi-natural habitats (Carvell et al., 2007), depending on landscape context (Krimmer et al., 2019). However, dispersal of beneficial insects from field margins into the cropped area depends on provision of in-field plant resources, without which pollinator and natural enemy activity decline rapidly with distance into the crop (Woodcock et al., 2016). At the CSC, diversified field margins together with increased in-field weed diversity are predicted to enhance the activity of beneficial insects in the regenerative cropping systems relative to conventional practice where plant diversity in the cropped area and surrounding field margin is low.
Most of the pollinators sampled were honeybees (58%), followed by bumblebees (38%) with solitary bees occurring less frequently (6% of the total). Overall, pollinator activity was higher in the regenerative cropping systems than in conventionally managed crops (Table 2). A significant crop effect was also detected due to high numbers of pollinators visiting the flowering oilseed rape crops, but no interaction between treatment and crop was detected. Apart from in flowering crops of oilseed, positive associations were observed between dicot weed densities and pollinator number across treatments, years and crop/rotation stage (p < 0.001, deviance ratio = 55.6, d.f. = 78, Figure 5). This suggests that the greater abundance of flowering weeds in the regenerative treatments is associated with an increase in the activity of pollinators within the cropped fields.
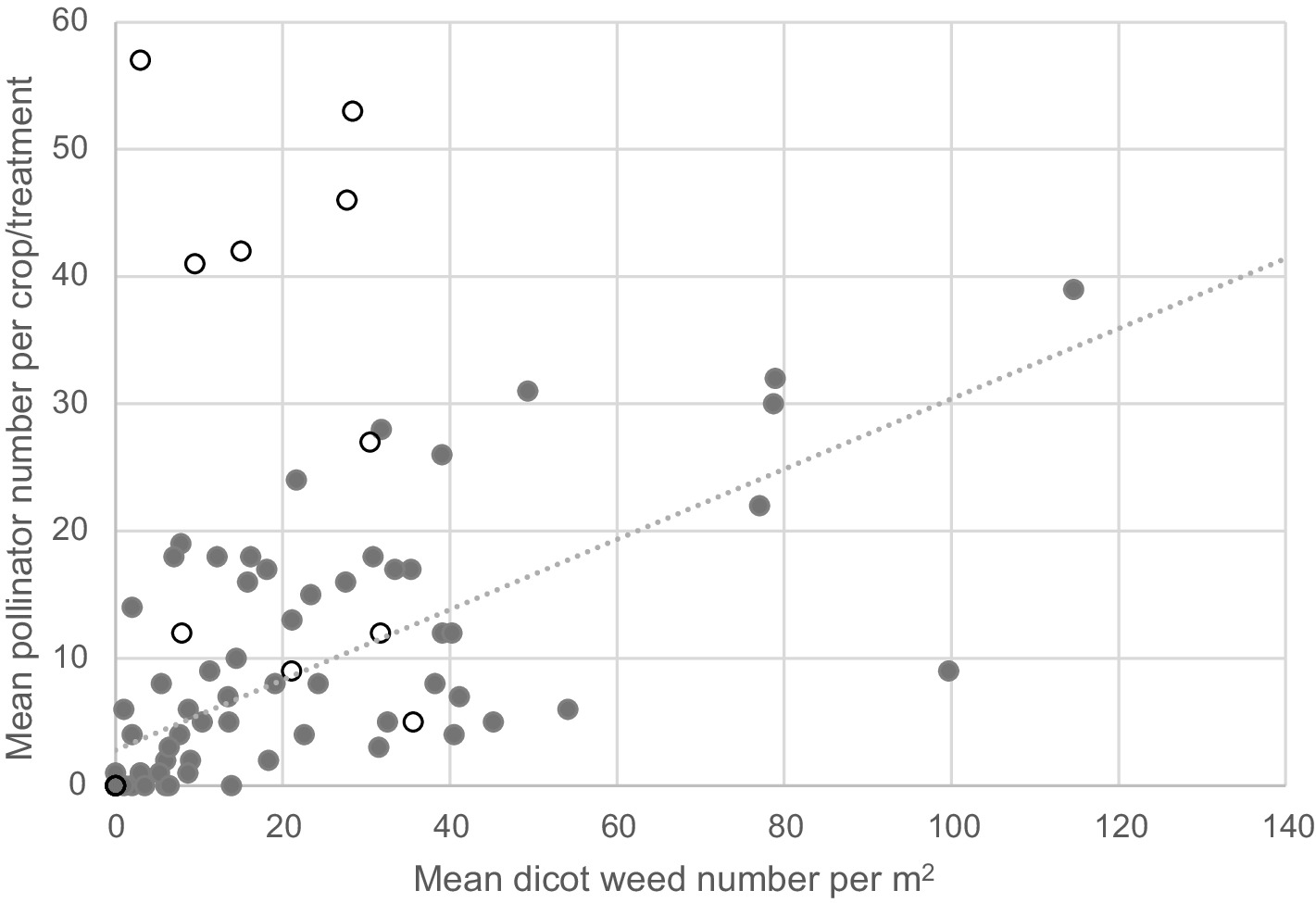
Figure 5. Relation between mean number of dicot weeds m−2 and mean pollinator number across all crops and treatments. Open symbols are winter oilseed crops and the 5 points with pollinator numbers over 40 per sample were crops in flower at the time of sampling, showing the over-riding effect of a flowering crop on the weed-pollinator relation.
3.3.3 Ground beetles
Arable fields and their immediate surroundings can support a diversity of invertebrates (particularly predatory ground beetles, rove beetles and spiders) active in leaf litter and on the ground surface. Many of these species play important ecological roles as food for mammals and birds, generalist predators of pests (slugs and aphids) and consumers of arable weed seeds (Holland, 2002). These invertebrate groups are sensitive to local habitat conditions, availability of their preferred food and the intensity of management, especially crop protection inputs. Much is known about their ecology, and they are easy to sample across many different habitats. The abundance of these invertebrate groups therefore has potential as an indicator of farmland habitat quality (Makwela et al., 2023).
Unlike other indicators, carabid abundance and species richness showed no significant effect of treatment. Numbers were lowest in the potato and following winter wheat but highly variable through the rest of the rotation, so the crop effect was not statistically significant. However, there was a significant interaction between crop and treatment due to lower numbers trapped in regenerative spring barley and higher numbers in regenerative winter barley relative to the conventional crop comparison. Pitfall trapping is a measure of activity as well as population density and therefore the numbers trapped are influenced by a range of factors not controlled as part of the CSC experimental platform (Lang, 2000). Where ground cover vegetation is dense (e.g., in weedy fields such as the spring barley), carabid movement may be obstructed and lower ground temperatures result in lower activity levels. Both factors reduce the likelihood of being trapped even in areas where population densities may have been positively impacted by low input management. In areas with little ground cover (e.g., winter barley), likelihood of trapping may be higher and a positive effect of management therefore more detectable. Further work is required to separate activity from density and generate more reliable estimates of the effect of cropping system on carabid communities.
Overall, results from 14 years of monitoring at the CSC suggest that the regenerative cropping system has a positive effect on most soil health and biodiversity indicators. Whether these benefits are sufficient to provide the regulating ecosystem services required to maintain crop production with less fertiliser and crop protection inputs was assessed by comparing the impact on yields (Figure 6).
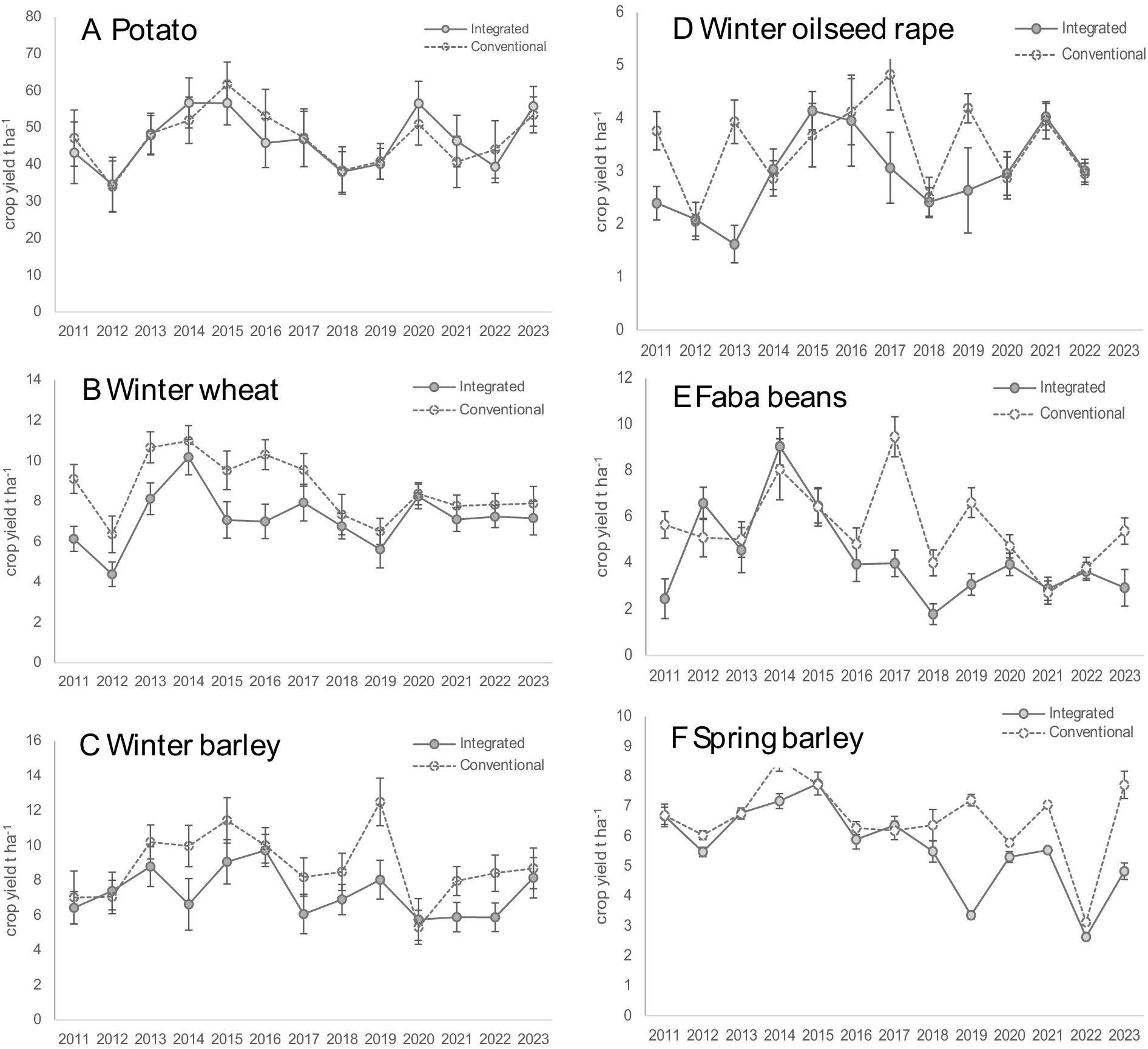
Figure 6. Mean crop yields over 13 years at the CSC long-term platform following two rotations of A potato, B winter wheat, C winter barley, D winter oilseed rape, E faba beans and F spring barley. Solid line = integrated crops, dashed line = conventional crops, error bars are standard errors of the mean.
3.4 Crop production
Annual average yield for potato crops from 2011 to 2023 was 43.9 tonnes ha−1 in the regenerative cropping system and 44.3 tonnes ha−1 under conventional management. This difference was not statistically significant when tested using a linear mixed model on data from the first six-year rotation (Hawes et al., 2018). Potato yields were comparable to the national average for Scotland (40–55 t ha−1) indicating good overall performance in both management systems and no apparent yield penalty in the regenerative treatment.
Winter wheat yields were significantly lower in the regenerative cropping system in the first rotation, showing a consistent 1 t ha−1 yr.−1 yield penalty with an average of 7.1 t ha−1 compared to 8.3 t ha−1 under standard commercial practice (Hawes et al., 2018). In the second rotation, mineral fertiliser requirement was calculated from soil nitrogen supply rather than applied at a standard reduced rate. This helped reduce the yield gap between treatments from 2017 onwards to an average of 7.4 t ha−1 (regenerative) and 7.7 t ha−1 (conventional). The ten-year winter wheat national average is 8.6 t ha−1, comparable to the conventional cropping system over all years.
Winter barley yields averaged 8.3 t ha−1 in conventional treatments compared to 7.2 t ha−1 in regenerative systems, but despite the difference in the first rotation being non-significant (Hawes et al., 2018), variability was high and conventional crops appeared to perform better in 2014, 2019, 2021 and 2022 compared to those in regenerative systems. In the second rotation, from 2017, this could be due to a build-up of winter wheat volunteers with direct drilling, resulting in competitive effects in some years. Further investigation is required to determine possible causes. Both systems produced yields comparable to the national average of 7.5 t ha−1.
Yields of oilseed were not statistically different between treatments in the first rotation (Hawes et al., 2018) and, over the whole period, averaged 3.7 and 3.2 t ha−1 in the conventional and regenerative treatments, respectively. However, lower yields were recorded in some years for regenerative crops due to poorer establishment when direct drilled through the crop residue from the previous cereal crop and, in some fields, disproportionate pest damage where pigeons favoured the no-till side of the field. Issues with establishment were resolved in later years using non-inversion till rather than direct drilling into stubble, providing better seed-soil contact.
Regenerative management had no significant impact on bean yields in the first crop rotation, averaging 5.5 t ha−1 compared to 5.8 t ha−1 for conventional crops (Hawes et al., 2018). However, the move from non-inversion tillage to direct drilling in the second rotation may have contributed to a decline in the bean yield in three of the 6 years to just 3.3 t ha−1 relative to standard crop management. The combination of direct drilling, dry soil conditions during establishment and weedy conditions in some fields can detrimentally affect final yield.
Spring barley yields were 6.1 t ha−1 in the regenerative system compared to 6.5 t ha−1 in the conventional and a national average of 6.7 t ha−1. This difference was not statistically significant for the first rotation (Hawes et al., 2018). Lower yields in regenerative cropping in some years were most likely caused by competition with the clover companion crop (in 2014), and with weeds (in 2019) where under-sown clover limits the weed control options available. Soil compaction could also be an issue in the second rotation where direct drilling replaced non-inversion tillage. Spring barley is the last crop in the rotation before potato and therefore the longest in no-till. Some varietal effects were detected: cv. Sassy has a better rooting structure than other varieties tested and performed better in the regenerative system than the conventional, particularly in dry seasons.
To enable direct comparison across all indicators, mean yields per crop, treatment and year were converted to a relative value across all crops (Table 2). On average, yields in regenerative systems were 64% of the maximum compared to 72% in conventional systems (p < 0.01). No differences in relative yields were detected between fields and there was no treatment.field interaction. Weed competition can have a significant negative effect on crop yield, and reduced tillage systems are at risk of generating a weed burden over time. The CSC regenerative treatments had significantly greater weed densities than the conventional crops, but a Generalised Linear Regression model showed no negative correlation between weed density and relativised crop yield (p = 0.94, deviance ratio 0.14, d.f.126). This, together with the positive association between weed densities and pollinator numbers, suggests that occasional tillage and the targeted weed management approach used in the regenerative system at the CSC may be sufficient to minimise the trade-off between biodiversity and crop yield.
The small yield penalties observed for regenerative crops were likely to be due to a range of extraneous factors including, in some years, extreme weather (drought conditions in spring, water logging in winter), pest damage (pigeon grazing) and establishment issues (direct drilled OSR and beans). These uncontrolled variables represent risk factors faced by commercial growers in the real-world and highlight the importance of experimental testing at commercially realistic spatial and temporal scales to provide quantitative data on the actual risks, costs and benefits of alternative cropping strategies for growers. Based on these results, improvements to system design should focus on: crop residue management (to improve OSR seedling establishment), ameliorating compaction issues after 4 or 5 years with no tillage (cereals), further reducing reliance on mineral fertilisers (e.g., by increased use of legume companion crops), better integration of IPM strategies, and screening for crop varieties that perform better in no-till and reduced input systems.
3.5 Carbon footprint and whole-systems impact
The regenerative system had a lower carbon footprint than the conventional system (Table 2) due to increased soil carbon storage, reduced mineral fertiliser and crop protection inputs and, apart from potato crops, less tillage disturbance. With the exception of potato, all the regenerative crops produced a negative carbon footprint through carbon sequestration. This carbon footprint improved through the rotation with lowest values in the spring sown faba beans and spring barley crops after 4 years no-till (Figure 4G). Conventionally cropped beans had a lower carbon footprint than other conventional crops. The highest CO2e across all crops was recorded for conventionally managed potato. The regenerative system therefore had less environmental impact than the conventional and an overall positive effect on greenhouse gas emission reductions.
Canonical Variates Analysis of all indicators together show a separation between conventional and regenerative systems and between crops through the rotation (Figure 7). The first axis (explaining 65% of the variation in the indicator data) separates the late-stage regenerative crops (min-till oilseed and direct drilled beans and spring barley) from all conventional ploughed crops and from early-stage regenerative crops (fields ploughed for potato and the following winter cereals planted into disturbed soil). The second axis (explaining a further 31% variation) separates the early-stage regenerative crops from all annually ploughed conventional crops. A clear distinction was therefore demonstrated between conventionally managed potato and winter cereal crops with low values for most indicators, and the regenerative spring sown beans and barley that had the longest time since potato cultivation and highest values for most indicators. Intermediate between these two extremes were the conventional spring crops (at the upper end of the conventionally managed range), and the regenerative potato and winter crops (at the lower end of the regenerative range). This suggests that, over multiple rotations, the combination of practices that comprise the CSC regenerative system can have a positive impact on soil health and biodiversity relative to standard commercial practice despite periodic disturbance for potato cropping.
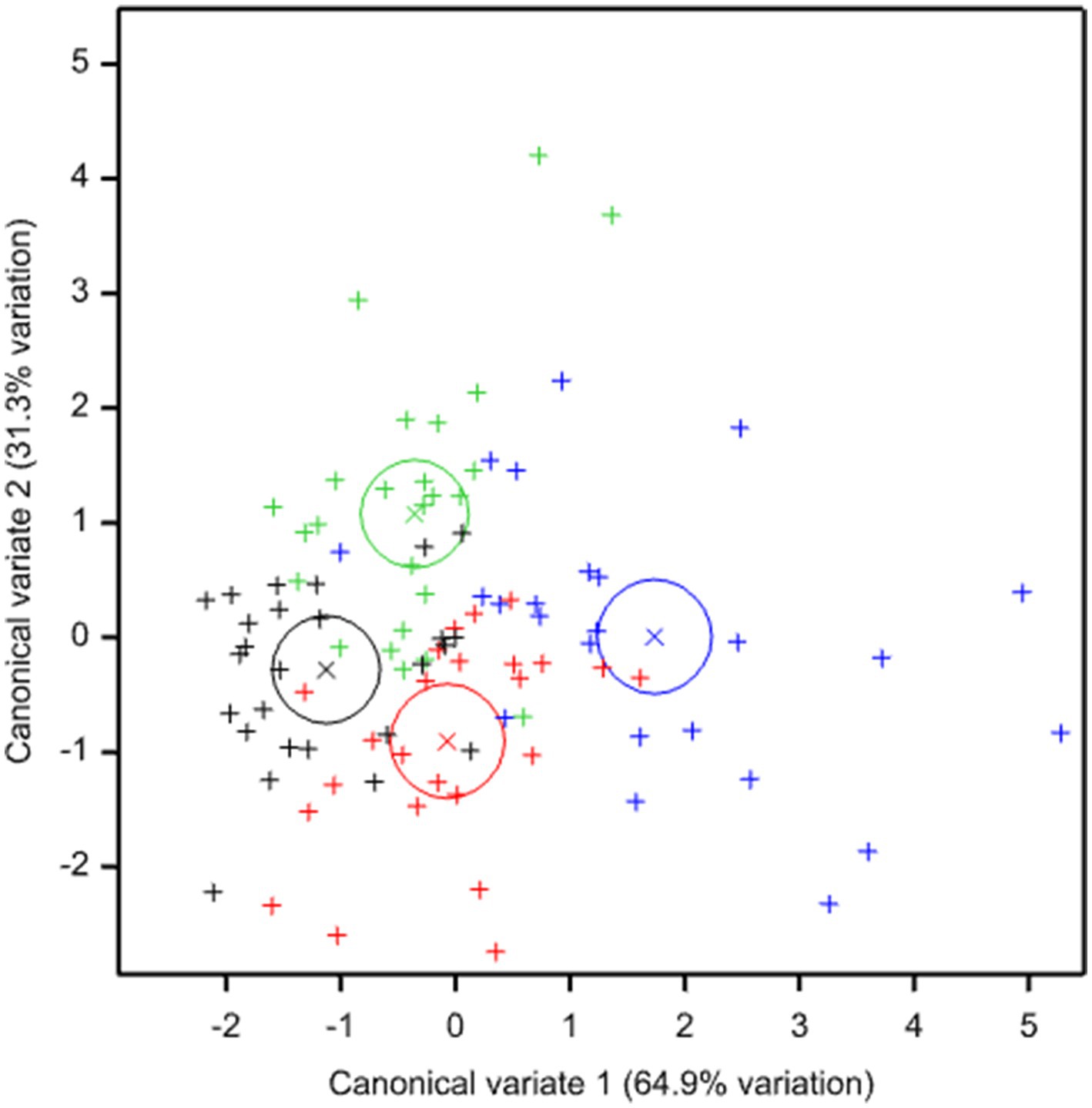
Figure 7. Canonical Variates scores showing 95% confidence limits for all indicator variables, grouped by treatment and time from potato cultivation: conventional, years 1–3 (black); conventional, years 4–6 (red); regenerative, years 1–3 years (green); regenerative, years 4–6 years (blue).
4 Conclusion
The primary goal of arable cropping is to grow high quality produce for food, feed, fuel or fibre. The challenge for regenerative agriculture systems is to do this without damaging the environment in which the crop is grown and therefore the prospects of continuing to crop the same land in perpetuity. Trends in indicators from the CSC long-term experiment suggest that, over 15 years of regenerative cropping, the original goals have largely been met. Organic matter inputs, cover cropping and occasional tillage over two crop rotations were associated with an increase soil carbon content and improved structural quality. Better soil structure and greater organic matter content promotes populations of earthworms and microarthropods, stimulating soil microbial activity and decomposition of organic matter to give greater concentrations of plant available nutrients for crop growth. Combined with these soil properties, targeted weed, pest and disease management resulted in higher scores for in-field weed biodiversity and greater activity levels of beneficial pollinating insects. In general, crop yields in the lower input, regenerative systems were largely maintained, demonstrating that management for environmental benefit is not necessarily in conflict with crop production and strategic combinations of best practice options can improve crop production efficiency, maintaining yields with less impact (Maeder et al., 2002).
Management to minimise or prevent environmental damage can incur a yield penalty [e.g., in organic systems where yields are generally around 80% that of conventional (de Ponti et al., 2012; Maeder et al., 2002)]. Organic systems are often designed around replacing chemical control with biological alternatives. The result is often a less polluting but less efficient version of conventional practice (Leifeld, 2012; Connor, 2008). Unlike these organic systems, regenerative and integrated approaches combine technology and agroecological practices to enhance soil health and biodiversity to support in-field processes. This has the potential to increase resilience and efficiency, thereby overcoming the apparent trade-off between environment and yield in low input systems (Röös et al., 2018; British Ecological Society, 2025). By adopting a long-term, whole-systems approach which encompasses key ecological and agronomic elements of the arable ecosystem (Figure 1), the CSC has developed, applied and demonstrated a framework for optimising these trade-offs to achieve multiple benefits for arable cropping.
Long-term studies are necessary to provide data on risks, costs and benefits at a temporal scale that is relevant to farmers and provide insight into opportunities to promote positive feedback loops and internal system regulation for increased production efficiency (George et al., 2022; Maclaren et al., 2022). For example, soil organic matter inputs improve soil structure which reduces loss by erosion and therefore maintains soil carbon content. Diverse weed understorey, maintained through targeted rather than blanket/eradication weed control strategies, can suppress dominance by competitive types. This further reduces the need for weed control and extends the life of existing crop protection chemistry. Minimising crop protection inputs reduces non-target effects, increasing non-pest herbivore densities which support natural enemy populations to regulate crop pest populations and mitigate over-reliance on crop protection chemicals.
However, these feedback loops take time to develop. In the initial stages of transition, the direct impact of management intervention on the system component may occur reasonably quickly (within a season or two, depending on the indicator in question). The second stage of the transition relies on a recovery from the initial perturbation caused by the introduction of a new management strategy to eventually reach a new stable state. This new stable state then generates the positive feedback loops to maintain and regulate the system process with less need for external intervention. Understanding these processes in the transition towards more sustainable production is essential for long-term success (Lavandero et al., 2025). Temporal trends in systems indicators from the CSC will be used to develop tools for farmers based on the agroecological network illustrated in Figure 1. This will enable farmers and land managers to explore impacts on system resilience and stability under future climate and cropping scenarios, facilitating informed judgements on how different management options may affect in-field ecology, how ecological processes impact sustainability outcomes, how to minimise trade-offs and monitor impact.
Data availability statement
Indicator datasets are available here: 10.5281/zenodo.17177640.
Author contributions
CH: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Supervision, Visualization, Writing – original draft, Writing – review & editing. AC: Data curation, Investigation, Methodology, Writing – review & editing. GB: Investigation, Methodology, Writing – review & editing. DB: Investigation, Methodology, Supervision, Writing – review & editing. JB: Investigation, Methodology, Writing – review & editing. PI: Funding acquisition, Supervision, Writing – review & editing. IS: Investigation, Methodology, Writing – review & editing. IT: Investigation, Methodology, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was funded by the Scottish Government Rural and Environment Science and Analytical Services Divisions, Strategic Research Programs: 2011–2016, 2016–2021 and 2021–2026 and supported by Projects: ‘N efficient plants for Climate Smart arable cropping systems’ (NCS) Project (Grant Agreement number 10043778), funded by DEFRA [Department for Environment, Food, and Rural Affairs (DEFRA; UK Government)], and supported by Innovate UK; the European Commission funded ‘Research and Innovation Action’ projects: www.RADIANT-project.eu, www.econutri-project.eu, and, www.legumESproject.eu (Grant Agreement numbers 101000622, 101081858, and 101135512, respectively).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adeux, G., Vieren, E., Carlesi, S., Barberi, P., Munier-Jolain, N., and Cordeau, S. (2019). Mitigating crop yield losses through weed diversity. Nat. Sustain. 2, 1018–1026. doi: 10.1038/s41893-019-0415-y
Aislabie, J., and Deslippe, J. R. (2013). “Soil microbes and their contribution to soil services” in Ecosystems Services in new Zealand Conditions and Trends. ed. J. R. Dymond (Lincoln: Manaaki Press), 143–161.
Bartlová, J., Badalíková, B., Pospíšilová, L., Pokorný, E., and Šarapatka, B. (2015). Water stability of soil aggregates in different systems of tillage. Soil Water Res. 10, 147–154. doi: 10.17221/132/2014-SWR
Beare, M. H., Parmelee, P. W., Hendrix, P. F., Cheng, W., Coleman, D. C., and Crossley, D. A. (1992). Microbial and faunal interactions and effects on litter nitrogen and decomposition in agroecosystems. Ecol. Monogr. 62, 569–591. doi: 10.2307/2937317
Berthon, K., Jaworski, C. C., Beacham, J. D., Jackson, P., Leake, J., McHugh, N. M., et al. (2024). Measuring the transition to regenerative agriculture in the UK with a co-designed experiment: design, methods and expected outcomes (2024). Environ. Res. Food Syst. 1:025007.
Biesmeijer, J. C., Roberts, S. P. M., Reemer, M., Ohlemüller, R., Edwards, M., Peeters, T., et al. (2006). Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 313, 351–354. doi: 10.1126/science.1127863
Bless, A., Davila, F., and Plant, R. (2023). A genealogy of sustainable agriculture narratives: implications for the transformative potential of regenerative agriculture. Agric. Hum. Values 40, 1379–1397.
Bonanomi, G., Cesarano, G., Antignani, V., Maio, C. D., Filippis, F. D., and Scala, F. (2018). Conventional farming impairs Rhizoctonia solani disease suppression by disrupting soil food web. J. Phytopathol. 166, 663–673. doi: 10.1111/jph.12729
Bonfanti, J., Potapov, A. M., Angst, G., Ganault, P., Briones, M. J. I., Calderón-Sanou, I., et al. (2025). Linking effect traits of soil fauna to processes of organic matter transformation. Funct. Ecol. 39, 446–461. doi: 10.1111/1365-2435.14720
Brenchley, W. A., and Warington, K. (1933). The weed seed population of arable soil. II. Influence of crop, soil and methods of cultivation upon the relative abundance of viable seeds. J. Ecol. 21, 103–127.
British Ecological Society (2025). Regenerative agriculture in the UK: an ecological perspective. London, UK:British Ecological Society.
Cao, Z. (2011). Changes in the abundance and structure of a soil mite (Acari) community under long-term organic and chemical fertilizer treatments’, Applied Soil Ecology, 49, 131–138. doi: 10.1016/j.apsoil.2011.06.0031
Carvell, C., Meek, W. R., Pywell, R. F., Goulson, D., and Nowakowski, M. (2007). Comparing the efficacy of Agri-environment schemes to enhance bumble bee abundance and diversity on arable field margins. J. Appl. Ecol. 44, 29–40. doi: 10.1111/j.1365-2664.2006.01249.x
Connor, D. (2008). Organic agriculture cannot feed the world. Field Crop Res. 106, 187–190. doi: 10.1016/j.fcr.2007.11.010
Cortet, J., Ronce, D., Poinsot-Balaguer, N., Beaufreton, C., Chabert, A., Viaux, P., et al. (2002). Impacts of different agricultural practices on the biodiversity of microarthropod communities in arable crop systems. Eur. J. Soil Biol. 38, 239–244. doi: 10.1016/S1164-5563(02)01152-4
Creamer, R. E., Hannula, S. E., Van Leeuwen, J. P., Stone, D., Rutgers, M., Schmelz, R. M., et al. (2015). Ecological network analysis reveals the inter-connection between soil biodiversity and ecosystem function as affected by land use across Europe. Appl. Soil Ecol. 97, 112–124. doi: 10.1016/j.apsoil.2015.08.006
Crossley, D. A., and Blair, J. M. (1991). A high-efficiency, “low-technology” Tullgren-type extractor for soil microarthropods. Agric. Ecosyst. Environ. 34, 187–192. doi: 10.1016/0167-8809(91)90104-6
Crossley, D. A., Mueller, B. R., and Perdue, J. C. (1992). Biodiversity of microarthropods in agricultural soils: relations to processes. Agric. Ecosyst. Environ. 40, 37–46. doi: 10.1016/0167-8809(92)90082-M
Crowder, D. W., and Jabbour, R. (2014). Relationships between biodiversity and biological control in agroecosystems: current status and future challenges. Biol. Control 75, 8–17. doi: 10.1016/j.biocontrol.2013.10.010
de Ponti, T., Rijk, B., and van Ittersum, M. K. (2012). The crop yield gap between organic and conventional agriculture. Agric. Syst. 108, 1–9. doi: 10.1016/j.agsy.2011.12.004
Franke, A. C., Lotz, L. A. P., vander Burg, W. J., and van Overbeek, L. (2009). The role of arable weed seeds for agroecosystem functioning. Weed Res. 49, 131–141.
Franzluebbers, A. J. (2002). Water infiltration and soil structure related to organic matter and its stratification with depth. Soil Tillage Res. 66, 197–205. doi: 10.1016/S0167-1987(02)00027-2
Frazão, J., de Goede, R. G. M., Brussaard, L., Faber, J. H., Groot, J. C. J., and Pulleman, M. M. (2017). Earthworm communities in arable fields and restored field margins as related to management practices and surrounding landscape diversity. Agric. Ecosyst. Environ. 248, 1–8. doi: 10.1016/j.agee.2017.07.014
Gaba, S., Cheviron, N., Perrot, T., Piutti, S., Gautier, J.-L., and Bretagnolle, V. (2020). Weeds enhance multifunctionality in arable lands in south-west of France. Front. Sustain. Food Syst. 4:71. doi: 10.3389/fsufs.2020.00071
George, T. S., Hawes, C., Valentine, T. A., Karley, A. J., Iannetta, P. P. M., and Brooker, R. W. (2022). Harnessing ecological principles and physiological mechanisms in diversifying agricultural systems for sustainability: experience from studies deploying nature-based solutions in Scotland. Front. Agric. Sci. Eng. 9, 214–237.
Gibson, R. H., Nelson, I. L., Hopkins, G. W., Hamlett, B. J., and Memmott, J. (2006). Pollinator webs, plant communities and the conservation of rare plants: arable weeds as a case study. J. Appl. Ecol. 43, 246–257. doi: 10.1111/j.1365-2664.2006.01130.x
Hartmann, M., and Six, J. (2023). Soil structure and microbiome functions in agroecosystems. Nat. Rev. Earth Environ. 4, 4–18. doi: 10.1038/s43017-022-00366-w
Hawes, C., Alexander, C. J., Begg, G. S., Iannetta, P. P. M., Karley, A. J., Squire, G. R., et al. (2018). Plant responses to an integrated cropping system designed to maintain yield whilst enhancing soil properties and biodiversity. Agronomy 8:229. doi: 10.3390/agronomy8100229
Hawes, C., Iannetta, P. P. M., and Squire, G. R. (2021). Agroecological practices for whole system sustainability. CABI Rev. 5:16. doi: 10.1079/PAVSNNR202116005
Hawes, C., Squire, G. R., Hallett, P. D., Watson, C. A., and Young, M. (2010). Arable plant communities as indicators of farming practice. Agric. Ecosyst. Environ. 138, 17–26. doi: 10.1016/j.agee.2010.03.010
Hawes, C., Young, M. W., Banks, G., Begg, G. S., Christie, A., Iannetta, P. P. M., et al. (2019). Whole-systems analysis of environmental and economic sustainability in arable cropping systems: a case study. Agronomy 9:438. doi: 10.3390/agronomy9080438
Heard, M. S., Hawes, C., Champion, G. T., Clark, S. J., Firbank, L. G., Haughton, A. J., et al. (2003). Non-crop plants in fields with contrasting conventional and genetically modified herbicide-tolerant crops. 1. Main effects of treatments. Philos. Trans. R. Soc. Lond. B 358, 1819–1832.
Holland, J. M. (2002). “Carabid beetles: their ecology, survival and use in agroecosystems” in The Agroecology of Carabid Beetles. ed. J. M. Holland (Andover, UK: Intercept Ltd.), 1–40.
Keuskamp, J. A., Dingemans, B. J. J., Lehtinen, T., Sarneel, J. M., and Hefting, M. M. (2013). Tea bag index: a novel approach to collect uniform decomposition data across ecosystems. Methods Ecol. Evol. 4, 1070–1075. doi: 10.1111/2041-210X.12097
Krimmer, E., Martin, E. A., Krauss, J., Holzschuh, A., Steffan,, and Derwenter, I. (2019). Size age and surrounding semi-natural habitats modulate the effectiveness of flower-rich agrienvironment schemes to promote pollinator visitation in crop fields. Agric. Ecosyst. Environ. 284:106590.
Lang, A. (2000). The pitfalls of pitfalls: a comparison of pitfall trap catches and absolute density estimates of epigeal invertebrate predators in arable land. J. Pest. Sci. 73, 99–106. doi: 10.1007/BF02956438
Lavandero, B., González-Chang, M., Jara-Rojas, R., Gallardo, I., and Wyckhuys, K. (2025). Enhancing agroecological transitions: from locally adapted protocols to a global transdisciplinary applied approach. Farming Syst. 3:100154. doi: 10.1016/j.farsys.2025.100154
Le Bissonais, Y. (2016). Aggregate stability and assessment of soil crustability and erodibility: I. Theory and methodology. Eur. J. Soil Sci. 67, 1–21.
Leifeld, J. (2012). How sustainable is organic farming? Agric. Ecosyst. Environ. 150, 121–122. doi: 10.1016/j.agee.2012.01.020
Lewis, T. D., Rowan, J. S., Hawes, C., and McKenzie, B. M. (2013). Assessing the significance of soil erosion for arable weed seedbank diversity in agro-ecosystems. Prog. Phys. Geogr. 37, 622–641. doi: 10.1177/0309133313491131
MacLaren, C., Mead, A., van Balen, D., Claessens, L., Etana, A., de Haan, J., et al. (2022). Long-term evidence for ecological intensification as a pathway to sustainable agriculture. Nat. Sustain. 5, 770–779. doi: 10.1038/s41893-022-00911-x
Maeder, P., Fliessbach, A., Dubois, D., Gunst, L., Fried, P., and Niggli, U. (2002). Soil fertility and biodiversity in organic farming. Science 296, 1694–1697. doi: 10.1126/science.1071148
Makwela, M. M., Slotow, R., and Munyai, T. C. (2023). Carabid beetles (Coleoptera) as indicators of sustainability in agroecosystems: a systematic review. Sustainability 15:3936. doi: 10.3390/su15053936
Marshall, E. J. P., Brown, V. K., Boatman, N. D., Lutman, P. J. W., Squire, G. R., and Ward, L. K. (2003). The role of weeds in supporting biological diversity within crop fields. Weed Res. 43, 77–89. doi: 10.1046/j.1365-3180.2003.00326.x
Neher, D. A., and Barbercheck, M. E. (2019). Soil microarthropods and soil health: intersection of decomposition and Pest suppression in agroecosystems. Insects 10:414. doi: 10.3390/insects10120414
Nicholls, C. I., and Altieri, M. A. (2013). Plant biodiversity enhances bees and other insect pollinators in agroecosystems. A review. Agron. Sustain. Dev. 33, 257–274. doi: 10.1007/s13593-012-0092-y
Petersen, H., and Luxton, M. (1982). A comparative analysis of soil Fauna populations and their role in decomposition processes. Oikos 39, 288–388. doi: 10.2307/3544689
Postma-Blaauw, M. B., de Goede, R. G. M., Bloem, J., Faber, J. H., and Brussaard, L. (2010). Soil biota community structure and abundance under agricultural intensification and extensification. Ecology 91, 460–473. doi: 10.1890/09-0666.1
Powney, G. D., Carvell, C., Edwards, M., Morris, R. K. A., Roy, H. E., Woodcock, B. A., et al. (2019). Widespread losses of pollinating insects in Britain. Nat. Commun. 10:1018. doi: 10.1038/s41467-019-08974-9
Roberts, H. A. (1958). Studies on the weeds of vegetable crops. I. Initial effects of cropping on the weed seeds in the soil. J. Ecol. 46, 759–768. doi: 10.2307/2257549
Röös, E., Mie, A., Wivstad, M., Salomon, E., Johansson, B., Gunnarsson, S., et al. (2018). Risks and opportunities of increasing yields in organic farming. A review. Agron. Sustain. Dev. 38:14. doi: 10.1007/s13593-018-0489-3
Rotchés-Ribalta, R., Sans, F. X., Mayer, J., and Mäder, P. (2020). Long-term farming systems and last crop sown shape the species and functional composition of the arable weed seed bank. Appl. Veg. Sci. 23, 428–440. doi: 10.1111/avsc.12496
Rusek, J. (1998). Biodiversity of Collembola and their functional role in the ecosystem’, Biodiversity & Conservation. 7, 1207–1219. doi: 10.1023/A:1008887817883
Ryan, M. R., Smith, R. G., Mirsky, S. B., Mortensen, D. A., and Seidel, R. (2010). Management filters and species traits: weed community assembly in long-term organic and conventional systems. Weed Sci. 58, 265–277. doi: 10.1614/WS-D-09-00054.1
Sanaullaha, M., Usman, M., Wakeela, A., Cheema, S. A., Ashraf, I., and Farooq, M. (2020). Terrestrial ecosystem functioning affected by agricultural management systems: a review. Soil Tillage Res. 196, 1–11.
Scottish Government (2024). Available online at: https://www.gov.scot/publications/results-from-the-scottish-agricultural-census-june-2024/ (Accessed May 9, 2025).
Simba, L. D., Foord, S. H., Thebault, E., and van Veen, F. J. F. (2018). Indirect interactions between crops and natural vegetation through flower visitors: the importance of temporal as well as spatial spill over. Agric. Ecosyst. Environ. 253, 148–156. doi: 10.1016/j.agee.2017.11.002
Smith, B. M., Aebischer, N. J., Ewald, J., Moreby, S., Potter, C., and Holland, J. M. (2020). The potential of arable weeds to reverse invertebrate declines and associated ecosystem services in cereal crops. Front. Sustain. Food Syst. 3:118. doi: 10.3389/fsufs.2019.00118
Storkey, J., and Neve, P. (2018). What good is weed diversity? Weed Res. 58, 239–243. doi: 10.1111/wre.12310
Stroud, J. L. (2019). Soil health pilot study in England: outcomes from an on-farm earthworm survey. PLoS One 14:e0203909. doi: 10.1371/journal.pone.0203909
Stroud, J., Irons, D. E., Watts, C. W., Storkey, J., Morris, N. I., Stobart, R. M., et al. (2017). Cover cropping with oilseed radish (Raphanus sativus) alone does not enhance deep burrowing earthworm (Lumbricus terrestris) midden counts. Soil Tillage Res. 165, 11–15.
Tilman, D. (1999). Global environmental impacts of agricultural expansion: the need for sustainable and efficient practices. Proc. Natl. Acad. Sci. USA 96, 5995–6000. doi: 10.1073/pnas.96.11.5995
van Groenigen, J. W., Lubbers, I. M., Vos, H. M. J., Brown, G. G., de Deyn, G. B., and van Groenigen, K. J. (2014). Earthworms increase plant production: a meta-analysis. Nat. Sci. Rep. 4:6365. doi: 10.1038/srep06365
von Redwitz, C., Andert, S., Bensch, J., Forster, R., Schatke, M., Strehlow, B., et al. (2025). Enhancing arable weed diversity by reduced herbicide use? J. Crop Health. 77:65. doi: 10.1007/s10343-025-01127-7
Keywords: functional diversity, regenerative agriculture, agroecological networks, biodiversity, integrated cropping, whole system design
Citation: Hawes C, Christie A, Banks G, Boldrin D, Brandt J, Iannetta P, Swyst I and Turner I (2025) Long-term regenerative practices enhance in-field biodiversity and soil health for sustainable crop yields. Front. Sustain. Food Syst. 9:1651686. doi: 10.3389/fsufs.2025.1651686
Edited by:
Liming Ye, Ghent University, BelgiumReviewed by:
Mauricio Gonzalez-Chang, Austral University of Chile, ChileMashford Zenda, University of the Free State, South Africa
Copyright © 2025 Hawes, Christie, Banks, Boldrin, Brandt, Iannetta, Swyst and Turner. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cathy Hawes, Q2F0aHkuSGF3ZXNAaHV0dG9uLmFjLnVr