 Sheena Salvarrey1*†
Sheena Salvarrey1*† Loreley Castelli2,3
Loreley Castelli2,3 Ciro Invernizzi1Eugenia Suárez4Yulai Ramos1Joaquín Aldabe4Gabriela Bentancur5Juan P. Burla4Mónica Remedios5
Ciro Invernizzi1Eugenia Suárez4Yulai Ramos1Joaquín Aldabe4Gabriela Bentancur5Juan P. Burla4Mónica Remedios5 Ismael Díaz6Camila Fernández Nion6
Ismael Díaz6Camila Fernández Nion6 Estela Santos1Franco Teixeira de Mello7
Estela Santos1Franco Teixeira de Mello7 Natalia Arbulo4†
Natalia Arbulo4†- 1Sección Etología, Facultad de Ciencias, Universidad de la República, Montevideo, Uruguay
- 2Centro de Investigaciones en Ciencias Ambientales, Instituto de Investigaciones Biológicas Clemente Estable, Montevideo, Uruguay
- 3Laboratorio de Microbiología y Salud de las Abejas, Departamento de Microbiología, Instituto de Investigaciones Biológicas Clemente Estable, Montevideo, Uruguay
- 4Departamento de Sistemas Agrarios y Paisajes Culturales, Centro Universitario Regional Este, Universidad de la República, Rocha, Uruguay
- 5Sección Entomología, Facultad de Ciencias, Universidad de la República, Montevideo, Uruguay
- 6Laboratorio de Desarrollo Sustentable y Gestión Ambiental del Territorio, Instituto de Ecología y Ciencias Ambientales, Facultad de Ciencias, Universidad de la República, Montevideo, Uruguay
- 7Departamento de Ecología y Gestión Ambiental, Centro Universitario Regional Este, Universidad de la República, Maldonado, Uruguay
Introduction: The current pollination crisis underscores the urgent need for enhanced research and monitoring efforts on pollinator diversity, as well as the development of effective strategies to promote their conservation. In Uruguay, soybean cultivation has expanded by approximately 1,000% in the last 20 years, and monocultures and pesticide use threaten pollinator food and nesting resources. However, data on native pollinators remain limited.
Methods: In this study, the abundance and composition of pollinating insects were assessed in the San Salvador River basin, one of the country’s main agricultural areas, using color pan traps installed at six paired sites (cultivated vs. wild vegetation) over three seasons (summer 2022, spring 2022, and summer 2023). To assess the effects of land use and sampling season on insect diversity, two approaches were employed: insect abundance was analyzed using generalized linear mixed models (GLMMs), and community composition at the order and morphospecies levels was evaluated using the Bray–Curtis index.
Results: A total of 10,690 insects, belonging to the main orders of Hymenoptera, Diptera, Lepidoptera, and Coleoptera, which include pollinators, were identified. Total insect abundance was significantly higher in cultivated sites than in wild sites. This association was also observed in Coleoptera, although this group exhibited an interaction with the sampling season, with the most abundant numbers being observed in both summers. Beta diversity (Bray-Curtis) revealed significant differences by land use and season at the order and morphospecies levels.
Discussion: Land use appears to be the variable associated with differences in insect communities across the evaluated sites. Particularly noteworthy is the species A. atromaculatus (Coleoptera) which presented much higher abundances than all other taxa. These results provide new insights into the structure of pollinator communities in agroecosystems and highlight the need to incorporate them into sustainable production, monitoring, and conservation efforts in the region.
1 Introduction
Insects (Class: Hexapoda) are a key component of terrestrial biodiversity, contributing to essential ecological processes such as soil renewal, pest control, and pollination (Schowalter, 2006). Pollination is of great importance for plant reproduction, as 87.5% of angiosperm species depend on insects for pollination (Ollerton et al., 2011). Therefore, this insect’s activity is not only responsible for maintaining plant communities in natural ecosystems but also for producing 30% of the food that comprises our diet (Klein et al., 2007). Entomophilous pollination is attributed an economic value of $153 billion worldwide (Gallai et al., 2009), and in Latin America, this value is $23.7 billion (Basualdo et al., 2022).
The orders Coleoptera, Diptera, Lepidoptera, and Hymenoptera, also known as “The Big Four,” are the most prominent pollinator groups due to their feeding habits and behaviors, which enable them to interact with a wide variety of plant species (Wardhaugh, 2015; Ollerton et al., 2011). Coleoptera comprise the most diverse order within the animal kingdom, with over 350,000 species (Zhang, 2013). Some beetles are frequent floral visitors and show high specificity for certain plants; however, they are not always valued for their role as pollinators (Wardhaugh, 2015). Among the Diptera, several families are recognized as pollinators, highlighting the hoverflies (Syrphidae), with nearly 6,000 reported species (Rotheray and Gilbert, 2011; Stefanescu et al., 2018). Lepidoptera encompasses more than 157,000 species (van Nieukerken et al., 2011), with 90% of them feeding on the nectar of the flowers they visit (Wardhaugh, 2015). Hymenoptera, with approximately 125,000 described species, includes the bees of the superfamily Apoidea (20,000 species), which depend exclusively on pollen and nectar as food resources, visiting a wide diversity of flowers (Michener, 2007), and are considered the most important pollinators on Earth (Potts et al., 2010).
Despite this remarkable diversity, many pollinator species are in decline. We are experiencing what the FAO has termed a “pollinator crisis,” characterized by a decrease in abundance and diversity of various insect groups (FAO, 2019). This situation is the result of multiple synergistic human activities (Kearns et al., 1998). Currently, over 44% of the Earth’s land surface is used for agricultural production (IPBES, 2017; Millard et al., 2021), and agricultural land use has intensified in recent decades. Intensification is frequently associated to landscape simplification, increased use of agrochemicals, and mechanized tillage (Winfree et al., 2009; Beninde et al., 2015), all of which contribute to habitat loss and fragmentation, increased competition with exotic species, and increased vulnerability to parasites and pathogens (Goulson, 2010; Goulson et al., 2015; Cameron et al., 2011; Steffan-Dewenter and Westphal, 2008; IPBES, 2017; Soroye et al., 2020). Furthermore, the widespread application of pesticides, including insecticides, herbicides and fungicides, can have both direct and indirect effects on pollinators by compromising their health and disrupting ecological interactions (Goulson et al., 2015; Woodcock et al., 2017). Given their sensitivity to environmental pressures and close ecological links with flowering plants, pollinators are widely recognized as key bioindicators of agroecosystem health (Kevan, 1999).
Despite growing global evidence of pollinator decline, a notable knowledge gap persists, with most studies focusing on Europe and North America (Saunders et al., 2020). In South America, and particularly in Uruguay, few studies assess the diversity and composition of pollinating insect communities at the landscape scale, especially in agricultural areas.
In Uruguay, agricultural intensification in recent decades has focused on extensive crops, such as soybeans (Glycine max), corn (Zea mays), and sorghum (Sorghum spp.), which has reduced the proportion of natural habitats and increased the use of agrochemicals (MVOTMA-FAO, 2019). In 2024, more than 1.5 million hectares were allocated to summer crops, of which 83.2% correspond to soybeans (MGAP-DIEA, 2024). Pesticide imports reached 12 million kilograms in 2020, impacting not only terrestrial biodiversity but also water bodies (MVOTMA-FAO, 2019). The San Salvador River basin, located in Uruguay’s western littoral, is particularly one of the most intensively cultivated areas in the country. This region accounts for 10% of the country’s total agricultural area and has 70% of its land designated for agricultural use (MGAP-DIEA, 2024; Ministerio de Ambiente, 2023). Pesticide residues, including glyphosate, atrazine, and chlorpyrifos, have been detected in water since 2014, potentially impacting aquatic and terrestrial biodiversity (Fernández Nion and Díaz, 2024). This region constitutes a critical case of landscape transformation, where patches of wild vegetation coexist with intensive agricultural areas, with potential implications for the pollinator population. However, no published studies are addressing the situation regarding insect diversity and composition within this region. This gap hinders understanding of how current agricultural practices impact insect assemblages that support key ecosystem functions, such as pollination.
Studying the four insect groups together allows us to understand not only the taxonomic richness of pollinators but also their functional range, from specialist species to incidental pollinators (Wardhaugh, 2015).
This study aims to assess the diversity and composition of pollinating insect communities, with a focus on the orders Coleoptera, Diptera, Lepidoptera, and Hymenoptera in both wild and cultivated environments within the San Salvador River basin. Wild environments are expected to have greater diversity of insect species, including those that are floral visitors and potentially effective pollinators.
2 Materials and methods
2.1 Study area
The study was conducted in the San Salvador River basin, located in the department of Soriano (−33.39811, −58.32177), on the western littoral of Uruguay. The area is primarily used for rainfed crops, predominantly soybean, wheat (Triticum aestivum), barley (Hordeum vulgare), corn and sorghum, and to a lesser extent, canola (Brassica napus), flax (Linum usitatissimum), and sunflower (Helianthus annuus) (Mondelli et al., 2015).
The study was carried out from the summer of 2022 to the summer of 2023, covering three sampling campaigns (summer 2022, spring 2022 and summer 2023). The region is climatically characterized by an average annual temperature of 17.8 °C, with an average annual precipitation of 1,214 mm (historical data: 1991–2020) (INUMET, 2023a). However, during the period in which this study was conducted (2022–2023), the region suffered a drought characterized by below-average rainfall, as reported by the National Meteorological Institute (INUMET, 2023b) in 2023. This drought significantly affected agricultural production, particularly during the 2022/2023 summer season.
2.2 Sampling design
To conduct insect sampling, six locations were selected across the upper, middle, and lower sections of the San Salvador River basin (Figure 1; Supplementary Table S1). The distance between consecutive locations ranged from 6.7 km to 29.7 km. Within each location, two nearby sites were chosen: one situated in a predominant crop area and the other in a non-crop area dominated by wild vegetation. This strategy resulted in six paired cultivated–wild sites, totaling 12 sampling sites (Supplementary Table S1). The distance between each paired site ranged from 1.7 to 5.9 km. To support the selection and categorization of crop and wild sites, the proportion of natural/semi-natural land surrounding each site was calculated. Using a database of land covers and uses for the San Salvador river basin generated for the study period (Fernández Nion and Díaz, 2024, 2025), the areas of different types of land covers and uses were determined for each site within radii of 500, 1,000, and 3,000 meters. Then, the proportion of natural/semi-natural land cover (natural forest, grassland and wetland) was calculated for each radius/area. To analyze differences in the proportion of natural/semi-natural land cover between sites, a generalized linear mixed model (GLMM) with beta distribution was used. Land use type (cultivated or wild) was used as an explanatory variable. Location identity was used as a random effect, to account for correlation within each locality. The analyses showed that sites categorized as wild had a significantly higher proportion of natural/semi-natural land cover than sites classified as cultivated at 500 meters (p < 0.001) and 1,000 meters (p < 0.001) (Supplementary Table S2) around them. This site selection suggests a clear distinction between wild and cultivated sites at these scales, confirming the classification used. However, there was no difference between wild and crop sites at 3000 meters (p > 0.1) (Supplementary Table S2), possibly due to the overlapping of the areas surrounding each paired site at this scale.
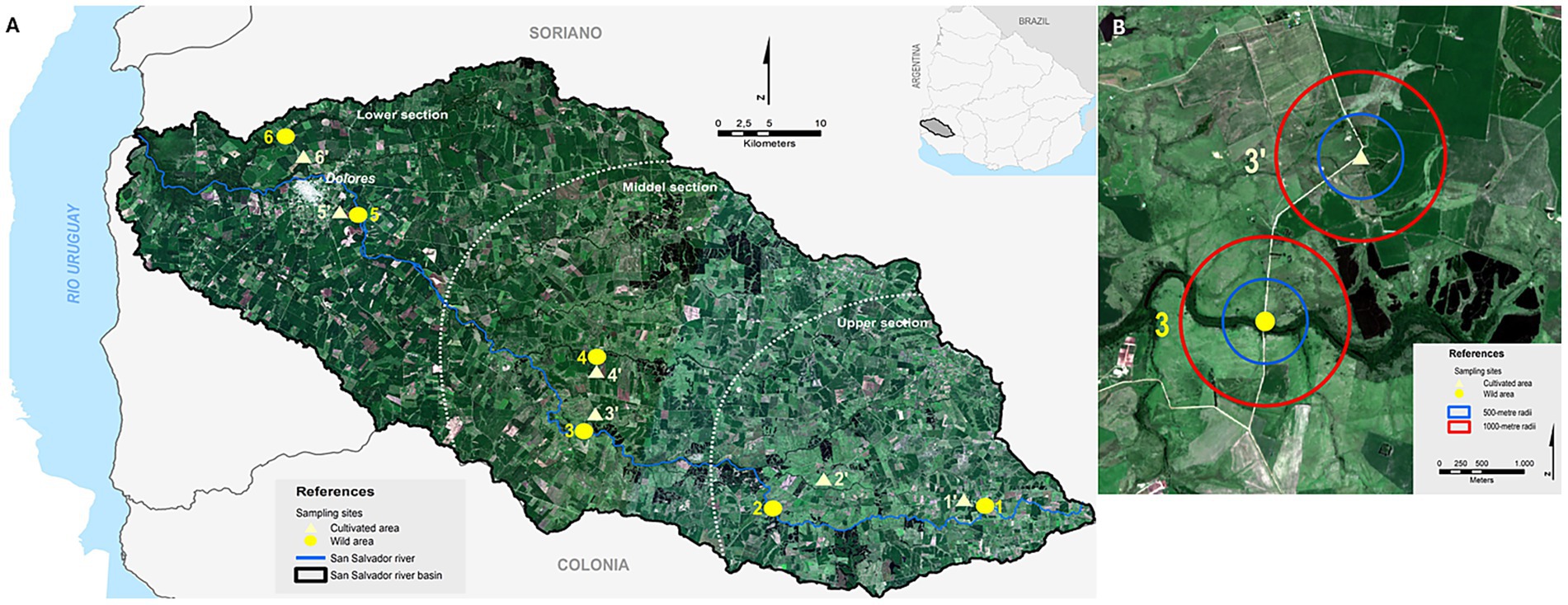
Figure 1. Study area in the San Salvador River basin, Uruguay. (A) Location of sampling sites classified as cultivated (triangles) and semi-natural (circles) environments. (B) Representative example of a sampling point showing the 500 m and 1,000 m buffers used for landscape characterization. Source: Sentinel-2 satellite image, March 2022, natural colour (B:4,3,2).
2.3 Insects sampling
At each site, a set of pan traps consisting of plates (16 cm in diameter, 5 cm high) in yellow, white, and blue was installed to attract different pollinators. Each set was secured by a metal frame 1.20 m high above ground level, thus remaining visible above the surrounding vegetation. In cultivated sites, the traps were placed at the edge of the crop to avoid potential inconveniences or losses due to tillage activities. In wild sites, areas far from human activity were chosen. Two-thirds of each container’s total volume was filled with a pre-made mixture of 600 mL of 10% ethylene glycol, 50 mL of detergent, and water, totaling 5 liters. To prevent overflows, small holes were made 1 cm below the rim of each container, allowing excess water from rainfall to drain without losing the contents (collected insects). The traps were active for 7–10 days. Sampling was conducted during summer 2022 (trap contents were removed on March 4th), spring 2022 (November 11th), and summer 2023 (March 3rd). During the sampling period, one set of traps was lost due to river overflow; this occurred in summer 2022 at wild site 1. Despite this, the analysis was based on 35 samples and a total of 12,216 individuals were captured.
The samples were collected from the field and stored in individual, properly labeled jars and 70% alcohol was added to preserve the contents. Once in the laboratory, the insects were identified to the order level. Following the criteria established in this study, only insects from the orders Coleoptera, Diptera, Hymenoptera, and Lepidoptera were considered. The remaining specimens were grouped under the “other” category and were excluded from the analysis. Similarly, microhymenoptera and microdipterans (< 5 mm) were not considered, assuming their potential contribution to pollination is not significant.
Species from the selected orders were classified into morphospecies. Photographic records were taken for each morphospecies, and between 1 and 3 specimens were stored in 95% alcohol in the Entomology lab (Centro Universitario Regional del Este) collection for subsequent identification. Using corresponding taxonomic keys, morphospecies were identified to the lowest possible taxonomic level (family, genus or species).
2.4 Statistical analyses
To assess the effects of land use and sampling season on insect diversity, two complementary analytical approaches were used.
First, generalized linear mixed models (GLMMs) with negative binomial distribution, suitable for modeling overdispersed count data, were used to analyse insect abundance. The response variables used were total insect abundance (Total_abundance) and total abundance for each insect order analyzed (Coleoptera_abundance, Hymenoptera_abundance, Diptera_abundance, Lepidoptera_abundance). Land use (cultivated, wild) and sampling season (summer 2022, spring 2022, summer 2023) were included as explanatory variables, while location identity was incorporated as a random effect to account for correlation within locations. For each response variable, different models were fitted, which included interactions between factors, additive effects and individual effects. In each case, the most appropriate model was selected using the Akaike information criteria (AIC) (Akaike, 1974).
Second, to assess diversity, we calculated richness and the Shannon index at the order and morphospecies levels and compare these parameters in relation to land use and sampling season, using non parametric Wilcoxon test. For community composition in relation to land use and sampling season, we analyzed beta diversity at the order and morphospecies level by using Bray–Curtis index (“vegdist” function) (Anderson, 2006; Anderson et al., 2006). Permutational multivariate analyses of variance (PERMANOVA) and tests for multivariate dispersion (betadisper) were used to evaluate variation in community composition (Anderson, 2006; Anderson et al., 2006). Only those comparisons that showed homogeneity of dispersion were considered (p > 0.05). The “metaMDS” function was used to plot ordinations. To identify taxa differentially associated with those variables, a differential abundance analysis was performed using DESeq2 (Love et al., 2014). Taxa with adjusted p < 0.05 (Benjamini–Hochberg correction) were considered significantly differentially abundant.
All analyses were performed using RStudio version 4.4.2 (Posit Team, 2024) using glmmTMB package (Brook et al., 2017) for mixed models and vegan package (Anderson, 2006; Anderson et al., 2006) for beta diversity analyses. Visual residual analysis was conducted to test model assumptions and model fit using “DHARMa” package (Hartig, 2024).
3 Results
A total of 10,690 individuals of 349 different morphospecies belonging to the orders Coleoptera (4,238), Diptera (5,065), Hymenoptera (1,286) and Lepidoptera (101) were analyzed. Within each group, 26 families were identified for the order Coleoptera, 21 for Diptera, 10 for Hymenoptera and 5 for Lepidoptera, leaving 17, 63, 58 and 15 morphospecies unclassified at the family level, respectively (Supplementary Table S3).
3.1 Insect abundance analyses
Total insect abundance was analyzed using negative binomial linear models, to assess the effects of land use, sampling season, and their interaction. This analysis was conducted for both total insect abundance and each insect order separately. Results showed that total insect abundance was significantly higher in cultivated sites than in wild sites (p = 0.038) (Figure 2A). The best-fitting model, based on the lowest AIC value (Table 1), includes land use as the sole predictor (Table 2).
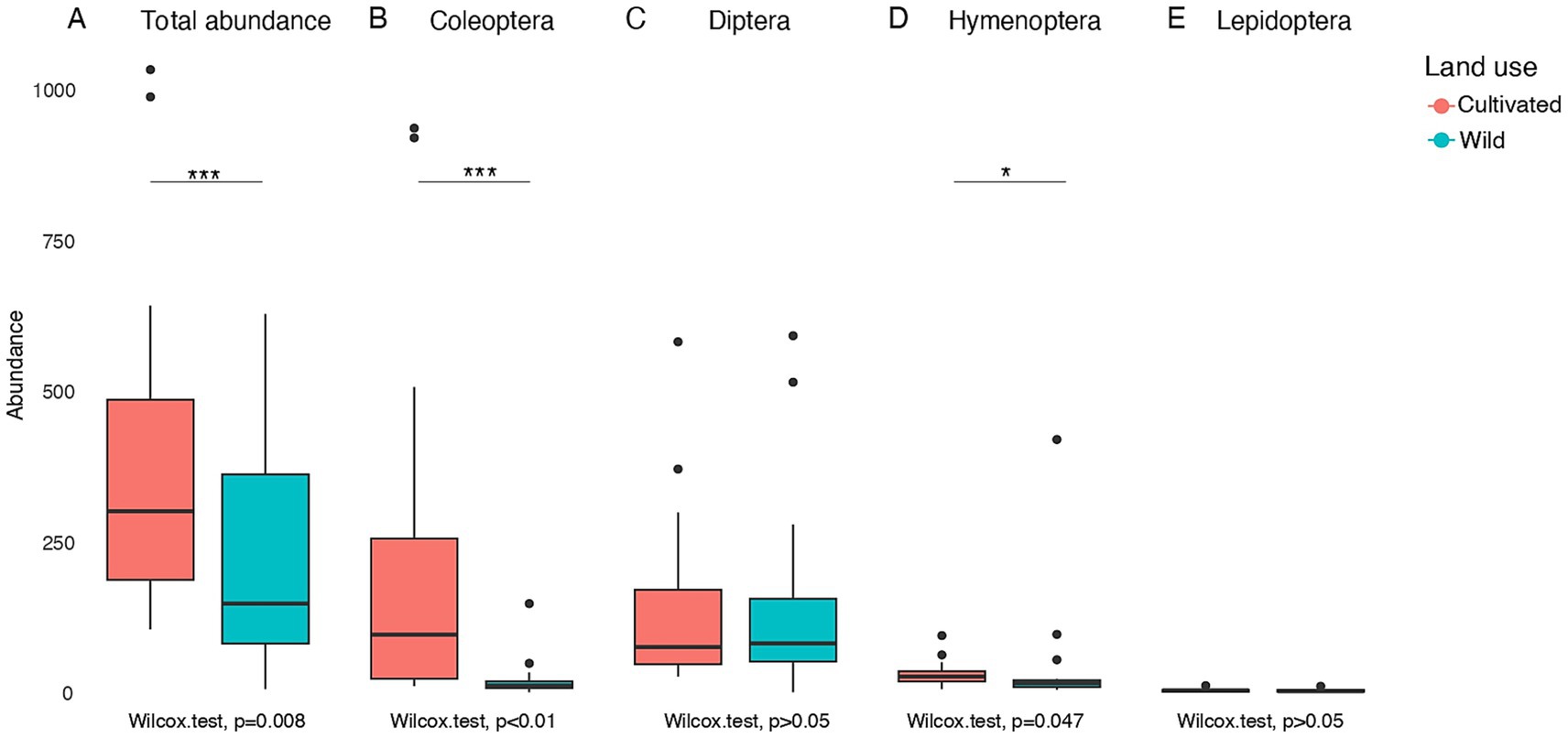
Figure 2. Total abundance of insect orders (Coleoptera, Diptera, Hymenoptera, Lepidoptera) across land use (A–E). Boxplots show the distribution of abundances in cultivated and wild areas for each insect order and total abundance. Results are shown as box plots, including median, 25 and 75% quartiles and outliers values.
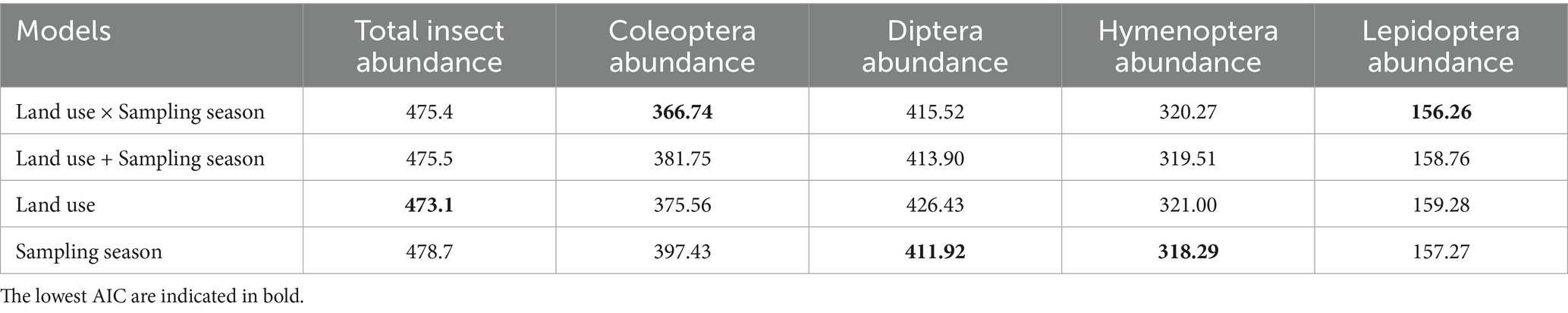
Table 1. Akaike information criteria (AIC) values were used to select the best fitted generalized linear mixed model for each response variable.
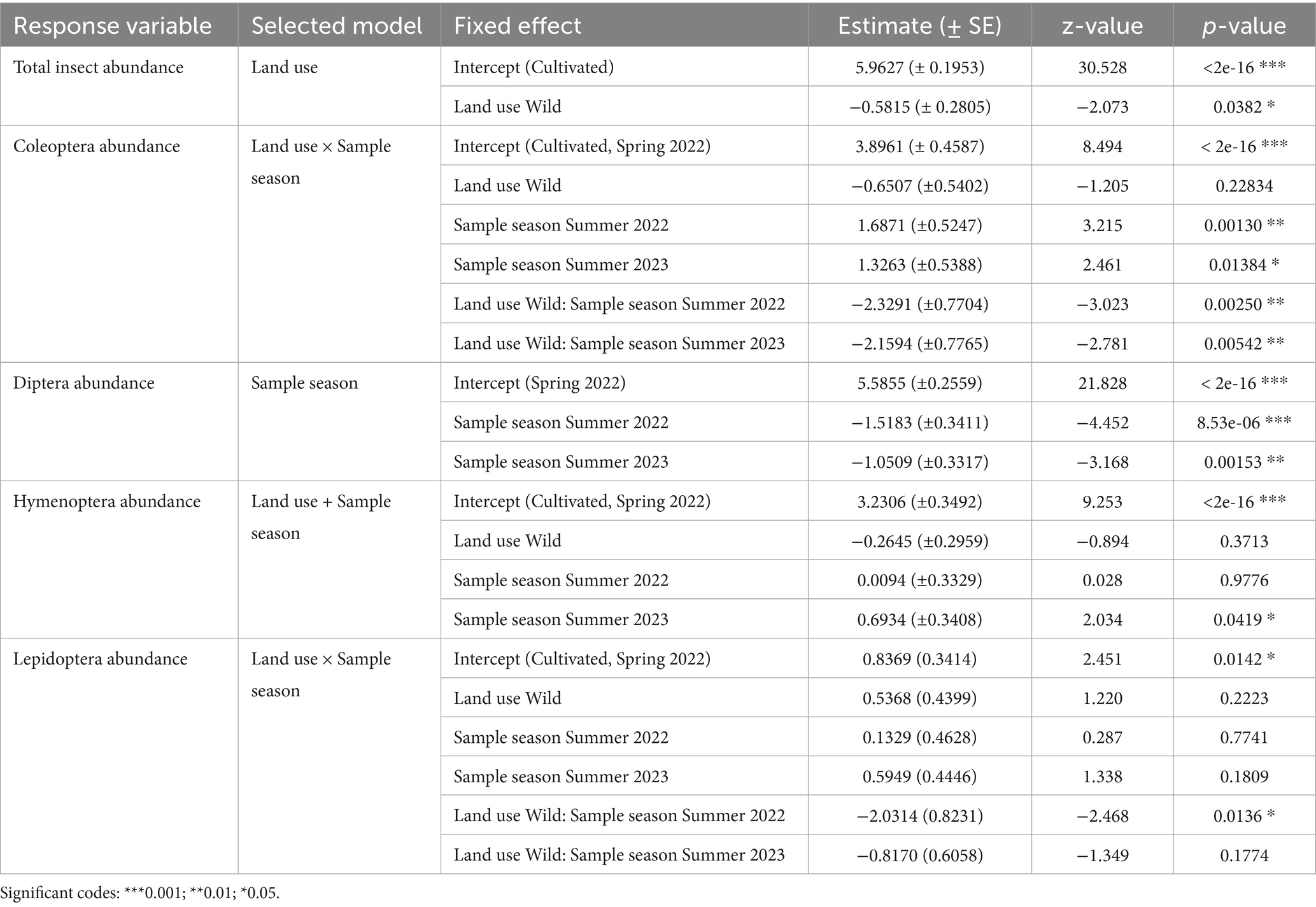
Table 2. Summary of the generalized linear mixed models (GLMMs) for each response variable.
3.2 Coleoptera
For Coleoptera abundance, the best-fitting model included the interaction between land use and sampling season (Land use × Sampling season), with the lowest AIC value (AIC = 366.7, Table 1). The interactions were significant in Summer 2022 (p = 0.0025) and Summer 2023 (p = 0.0054), indicating lower abundance in wild sites compared to cultivated sites during those seasons (Figure 2B; Table 2). These results suggest that the effect of land use on Coleoptera abundance varies seasonally, with cultivated sites being more favorable during summer.
3.3 Diptera
The model that included season as the sole predictor provided the best fit for the Diptera abundance (lowest AIC, Table 1). This analysis showed a significantly higher abundance of Diptera in Spring 2022 compared to Summer 2022 (p < 0.001) and Summer 2023 (p = 0.0015). Including land use as a predictor did not improve model performance in any of the evaluated cases. These results suggest that Diptera abundance varied primarily in response to seasonal factors, rather than land use (Figure 2C).
3.4 Hymenoptera
For Hymenoptera, when land use and sample season were included as additive effects, a significant difference was observed: in summer 2023, Hymenoptera abundance was significantly higher (p = 0.041) (Figure 2D; Table 2). Nevertheless, the model that provided the best fit based on AIC values was the one that considered season as the sole predictor (Table 1), although the higher abundance in Summer 2023 was only marginally significant (p = 0.051). In addition, considering the interaction model (Land use × Sampling season), a marginally higher abundance was observed in wild sites (p = 0.07). These results suggest that, during the season with the highest overall abundance, Hymenoptera were more closely associated with wild environments.
3.5 Lepidoptera
In Lepidoptera, the model that showed the best fit according to the AIC values (Figure 2E; Table 1) includes the interaction between land use and sampling season. This model revealed a higher abundance in cultivated sites during the summer of 2022 (Table 2).
3.6 Richness and Shannon index
At both the order and morphospecies levels, there were no significant differences in richness or the Shannon index between the different land uses (Supplementary Figure S1). The results were similar for each season, with no significant differences in these indices between cultivated and wild sites.
3.7 Community composition
To analyze community composition, a Bray–Curtis dissimilarity ordination (NMDS) was performed, showing community differentiation based on land use, sampling season, and their interaction (Figure 3).
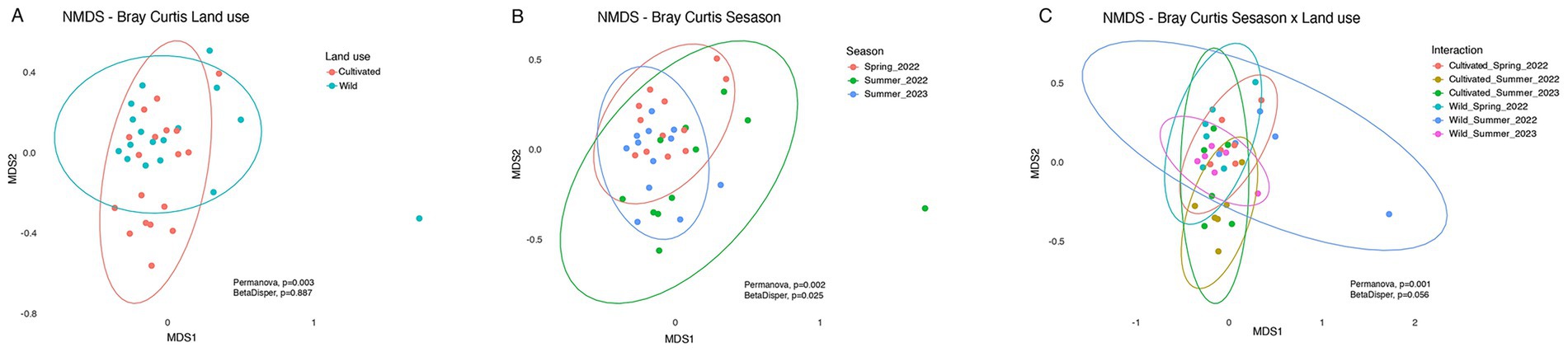
Figure 3. Ordination by non-metric multidimensional scaling (NMDS) based on Bray–Curtis dissimilarity of insect community composition (order level). NMDS plots illustrate differences in insect community structure according to land use (Cultivated vs. wild) (A), season (Spring_2022, Summer_2022 and Summer_2023) (B) and the interaction between land use and season (C).
At the land use level, insect communities clustered differently according to land use, both at the order level (PERMANOVA, p = 0.006; BetaDisp p = 0.885) (Figure 3A) and at the morphospecies level (PERMANOVA, p = 0.003; BetaDisp p = 0.887). In both cases, Beta-disper values indicated no differences in within-group dispersion, supporting the observed differences in community composition. Seasonal analysis also revealed significant differences in composition at the order level (PERMANOVA, p = 0.002); however, these were accompanied by significant changes in dispersion (BetaDisp., p = 0.025), suggesting variability within seasons that may limit the interpretation of PERMANOVA results (Figure 3B). At the morphospecies level, seasonal differences in community were also observed (PERMANOVA p = 0.001; BetaDisp = 0.09). Pairwise PERMANOVA comparisons showed that Spring 2022 differed significantly from Summer 2022 (p = 0.039), while no significant differences were found between Spring 2022 and Summer 2023 (p = 0.114), or between Summer 2022 and Summer 2023 (p = 0.396).
Finally, when land use and season were considered simultaneously, the PERMANOVA analysis revealed a significant difference in community composition (p = 0.001). However, the BetaDisp value (p = 0.056) was marginally non-significant, suggesting some internal variability within each season depending on land use. The most clearly differentiated communities were observed in the summer of 2022, between cultivated and wild sites, showing the greatest dissimilarity in multivariate space (Figure 3C). This was supported by PERMANOVA results, which indicated a significant effect of land use during that season (F = 3.34, p = 0.014). In contrast, communities in the spring of 2022 showed the highest overlap and heterogeneity between land uses, with no significant differences detected (p = 0.613). Similarly, no significant differences were found in summer 2023 (p = 0.105), although a trend toward differentiation was observed.
Given the differences observed in abundance and species composition across seasons, a DEseq2 analysis was performed for each season (Figure 4). The results indicate that the predominant species in spring were the dipteran morphospecies Fannidae_sp. and the hymenopteran, Polybia scutellaris (Figure 4A), both associated with cultivated sites. In the summers of 2022 and 2023, the dominant species was the beetle, Astylus atromaculatus, which was also more abundant in cultivated sites (Figures 4A–C).
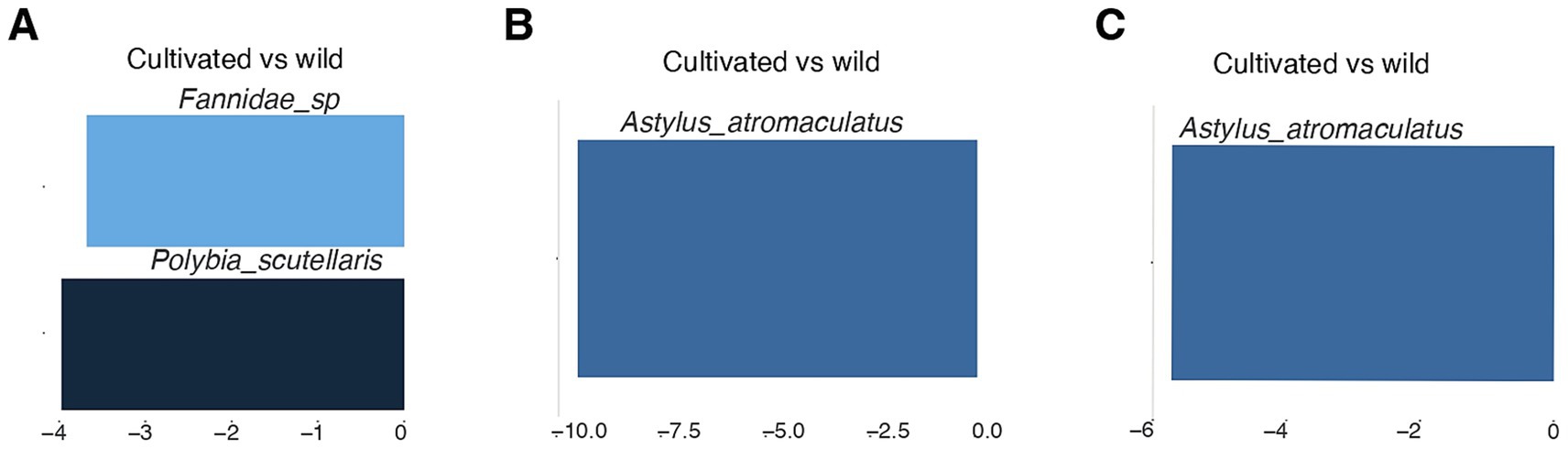
Figure 4. DESeq2 analysis of insect communities across different seasons (A-C) in sites with different land use (cultivated vs. wild). Fold change values < 0 indicate that the abundances were higher in cultivated areas.
As shown by the DESeq2 analysis, Astylus atromaculatus showed significantly higher abundance in cultivated sites during summer 2022, with approximately 1,024 times greater abundance compared to wild sites (log₂ fold change = 10) (Figure 4B). In summer 2023, its abundance remained higher in cultivated sites, with about 64 times greater abundance (log₂ fold change = 6), indicating a lower but still substantial seasonal difference in its response to land use (Figure 4C).
4 Discussion
Our results indicate that both land use and seasonality have a significant influence on the abundance and composition of pollinating insect communities in the San Salvador River basin. This result was not detected when comparing the richness and Shannon index. In general, the total abundance of insects was significantly higher in cultivated sites, especially during the summer. This suggests that agricultural environments may act as attraction zones for certain groups, possibly due to a greater availability of floral resources (Holzschuh et al., 2013) or they function as refuges for generalist species or those adapted to extensive systems (Westphal et al., 2003; da Silva et al., 2008; Aguilera et al., 2020). These patterns are not uniform across all pollinator taxa; instead, the response of insects to different land uses and climatic conditions appears to be largely taxon-dependent, as reported in previous studies (Cusser et al., 2018; Bottero et al., 2023). This may be the reason why we found no differences in richness and the Shannon index when we analysed all the orders together; an analysis of these indices for each order separately could reveal different responses. The composition analyses were consistent with these observations. The order Coleoptera showed a clear response to land use, with significantly higher abundances in cultivated sites during both summers. The presence of a dominant species, Astylus atromaculatus (Melyridae), significantly influenced this pattern, with its population reaching exceptionally high values in the summer of 2022, far exceeding the abundance recorded in 2023 (Figure 3B). This native species, whose adults feed on the pollen of various plant species, has been observed in association with several crops, exhibiting an opportunistic habit in agricultural landscapes (Cibils et al., 2023). It has also been highlighted as a pollinator in some crops (Pierre and Hofs, 2010). Among the crops it visits are sorghum, alfalfa, and soybean, the latter being especially abundant in this area (Cibils et al., 2023), reinforcing its role as a dominant species in cultivated environments (Figure 4).
DESeq2 analyses confirmed that a small set of dominant species determined the differences in abundance and composition. The population outbreak of Astylus atromaculatus during the summer of 2022 may be related to extreme climatic conditions, particularly the high temperatures and drought recorded that year, according to reports from official meteorological agencies (INUMET, 2022). Although this study did not include climatic variables in its analyses, it is important to acknowledge that such environmental conditions likely influenced the observed community dynamics, thereby shaping the responses of pollinating insects. Likewise, the massive presence of A. atromaculatus in the landscape during that period constitutes an example of ecological imbalance, with documented impacts at both productive and environmental levels. Its abundance caused public concern in the region (MGAP-DIEA, 2024), especially due to reported cases of livestock intoxication resulting from the accidental ingestion of these insects (Giannitti et al., 2024), reinforcing the importance of monitoring such events from an integrated ecological perspective.
Dipterans showed a marked seasonal response, with higher abundance during spring 2022, regardless of land use type. The absence of a significant land use effect suggests that this group responds primarily to life cycle requirements, which are closely linked to climatic conditions and the temporal availability of resources (Davis et al., 2023). Within this group, a morphospecies of the Fanniidae family stood out, associated with cultivated environments in spring, possibly using the vegetation as a refuge and/or nectar source. This family, common in Neotropical regions, includes members with diverse feeding habits, although many species in their adult stage feed on nectar and visit various flowers (Grisales and de Carvalho, 2019).
Hymenopterans generally showed an intermediate response, with a slight preference for wild environments during the summer of 2023. This could be due to the high functional diversity of the group, which includes bees and wasps with different degrees of sociability, specialization, and feeding strategies, allowing them to adapt to both wild and cultivated environments. Nevertheless, seasonal DESeq2 analyses revealed a high abundance of the native social wasp Polybia scutellaris in cultivated environments during spring. This species, as other wasps, exhibit both phytophagous and predatory habits, highlighting their dual role as pollinators and biological control agents. Their low dispersal capacity may explain their association with crops, where they find accessible food sources (Fernández Corujo et al., 2010).
On the other hand, national-level studies have shown a relationship between bee diversity and landscape characteristics, finding a positive association with environments of lower agricultural intensity (Santos et al., 2020a). However, bees such as Apis mellifera have been found in extensive crops, including soybeans, an important source of nectar and pollen, which can even have a positive impact on honey production (Santos et al., 2021). Similarly, native bees of the Megachilidae family use these crops as a resource for nesting (Santos et al., 2020b), thus demonstrating the adaptability of the group. The diversity of life histories and ecological strategies within this order may explain the differentiated responses observed to land use and seasons.
Lepidopterans were more abundant in summer and showed a slight preference for cultivated environments, possibly linked to the presence of soybean, the dominant crop in that season. This group was the least abundant of the four orders analyzed, with only 101 individuals recorded. This low detection may be due to limitations in the sampling methodology, as color pan traps may not be efficient for capturing this group, especially for larger butterfly species, as reported in other studies (Campbell and Hanula, 2007; Vrdoljak and Samways, 2012). Despite this limitation, which may occur for several other taxa, the same bias is expected across all comparative samples. Taking this into account, it is worth noting that color pan traps are a relatively low-cost, simple, and effective method for sampling relative abundances and species richness of several flower-visiting insect groups, especially suitable for comparative and monitoring purposes (Campbell and Hanula, 2007; Vrdoljak and Samways, 2012).
The higher abundance observed in cultivated sites does not necessarily imply greater habitat quality, functional diversity, or improved ecosystem service provision (Isbell et al., 2011; Tscharntke et al., 2012). The composition analyses revealed that agricultural environments were dominated by a few species (e.g., Astylus atromaculatus), in contrast to wild sites, which, although showing lower insect abundance, tended to harbor more even, and potentially more diverse communities. These findings highlight that a higher number of individuals does not equate to ecological balance or resilience. Pollinator diversity plays a crucial role in maintaining stable and resilient pollination services (Garibaldi et al., 2011; 2013, Cariveau et al., 2013), as diverse assemblage can pollinate a broader range of plants, buffer against species losses, and enhance the ecosystem’s ability to support disturbances (Kennedy et al., 2013; Maggi et al., 2023). Thus, although specific agricultural environments may attract large numbers of pollinators, as illustrated by the dominance of A. atromaculatus discussed above, such patterns may reflect reduced ecological integrity and reduced functional redundancy rather than enhanced pollination potential. In this context, although wild sites did not show the highest abundances, they may harbor key species with ecologically relevant functions that are not detected by simple abundance metrics. The specific composition and evenness observed in these environments reinforce the need for more complex studies that integrate functional aspects and interaction networks, along with landscape composition and configuration (Fijen et al., 2025).
It is worth noting that the study region is fully embedded within an intensively farmed matrix, so the contrast between “cultivated” and “wild” sites manifests at relatively small spatial scales (Kennedy et al., 2013; Landis, 2017). This is evidenced by the significant differences in wild cover observed between paired points at distances of 500 and 1,000 meters, but not at 3000 meters, suggesting that at larger scales, wild environments may function as isolated patches lacking functional connectivity. In this context, our study area has undergone substantial landscape changes over the past few decades, which may have shaped the local pollinator communities, reducing differences between cultivated and wild zones or diminishing the influence of the latter. To assess the effects of biodiversity, long-term monitoring efforts in these regions must be strengthened, as the impacts of land use are more significant when sustained over time (Cusser et al., 2018, 2021).
In this study, we assessed landscape composition based on the relative proportion of wild cover in the surroundings; however, we did not address structural heterogeneity or the spatial configuration of patches, key aspects for understanding ecological connectivity. This methodological limitation should be addressed in future work to account for the sensitivity of pollinator communities to subtle changes in landscape structure and organization. Likewise, it is necessary to acknowledge the limitations inherent to passive sampling, which does not discriminate between floral visitors and effective pollinators. The incorporation of complementary methods, such as direct observations on flowers and the analysis of plant–pollinator interaction networks, would enable a more comprehensive and accurate characterization of the ecological roles of these communities.
Finally, this study differs from other approaches focused exclusively on bees (Potts et al., 2010; Winfree et al., 2009; Garibaldi et al., 2011; Kennedy et al., 2013; Cariveau et al., 2013) by incorporating a community-level perspective on insects. The inclusion of orders relevant to pollination (Coleoptera, Diptera, Hymenoptera, and Lepidoptera) enabled the detection of broader responses to environmental and landscape factors, thus providing valuable evidence for understanding ecological functioning in agricultural landscapes of southern South America. In a context of high intensification, where the expansion of soybean monoculture and intensive grazing have significantly reduced ecosystem services in our landscapes (Modernell et al., 2016), understanding pollinator dynamics can contribute to the design of conservation and biodiversity-use strategies, fostering more resilient environments.
Future research should aim to increase the resolution of land use categories, incorporating detailed information on pesticide applications, crop types, and management practices, which can significantly influence pollinator communities. In addition, the integration of tools such as remote sensing and geographic information systems (GIS) could improve our ability to detect landscape features that act as barriers or corridors for pesticide dispersal, and that may limit or facilitate pollinator movement and survival. These advances would enhance the development of more effective and targeted conservation strategies in agroecosystems.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
SS: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Resources, Supervision, Visualization, Writing – original draft, Writing – review & editing. LC: Data curation, Formal analysis, Methodology, Software, Visualization, Writing – review & editing. CI: Conceptualization, Investigation, Project administration, Writing – review & editing. EuS: Data curation, Writing – review & editing. YR: Data curation, Writing – review & editing. JA: Formal analysis, Writing – review & editing. GB: Data curation, Writing – review & editing. JB: Data curation, Writing – review & editing. MR: Data curation, Writing – review & editing. ID: Methodology, Visualization, Writing – review & editing. CFN: Methodology, Writing – review & editing. EsS: Data curation, Writing – review & editing. FTM: Conceptualization, Funding acquisition, Project administration, Writing – review & editing. NA: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by Food and Agriculture Organization of the United Nations (FAO) through project “Fortalecimiento de las capacidades para la gestión ambientalmente adecuada de plaguicidas incluyendo COPs” UTF/URU/035/URU. The publication was supported by Programa de Desarrollo de la Ciencias Básicas (PEDECIBA) de la Universidad de la República.
Acknowledgments
We thank the Ministerio de Medio Ambiente, Fundación para el Desarrollo de las Ciencias Básicas (FUNDACIBA); the institutions and collaborators who facilitated access to field sites and provided logistical support throughout the study. SS and FTM thank Sistema Nacional de Investigadores (SNI) and Programa de Desarrollo de las Ciencias Básicas (PEDECIBA). Additionally, part of the R code used in the statistical analyses was developed with assistance from the AI tool ChatGPT (OpenAI, 2025), which was used to refine scripts and resolve coding issues. All analyses and interpretations were conducted and validated by the authors.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that Gen AI was used in the creation of this manuscript. Additionally, part of the R code used in the statistical analyses was developed with assistance from the AI tool ChatGPT (OpenAI, 2025), which was used to refine scripts and resolve coding issues. All analyses and interpretations were conducted and validated by the authors.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsufs.2025.1672127/full#supplementary-material
References
Aguilera, G., Roslin, T., Miller, K., Tamburini, G., Birkhofer, K., Caballero-Lopez, B., et al. (2020). Crop diversity benefits carabid and pollinator communities in landscapes with semi-natural habitats. J. Appl. Ecol. 57, 2170–2179. doi: 10.1111/1365-2664.13712
Akaike, H. (1974). A new look at the statistical model identification. IEEE Trans. Autom. Control 19, 716–723. doi: 10.1109/TAC.1974.1100705
Anderson, M. J. (2006). Distance-based tests for homogeneity of multivariate dispersions. Biometrics 62, 245–253. doi: 10.1111/j.1541-0420.2005.00440.x
Anderson, M. J., Ellingsen, K. E., and McArdle, B. H. (2006). Multivariate dispersion as a measure of beta diversity. Ecol. Lett. 9, 683–693. doi: 10.1111/j.1461-0248.2006.00926.x
Basualdo, M., Cavigliasso, P., de, R. Jr., Aldea-Sánchez, P., Correa-Benítez, A., Harms, J., et al. (2022). Current status and economic value of insect-pollinated dependent crops in Latin America. Ecol. Econ. 196:107395. doi: 10.1016/j.ecolecon.2022.107395
Beninde, J., Veith, M., and Hochkirch, A. (2015). Biodiversity in cities needs space: a meta-analysis of factors determining intra-urban biodiversity variation. Ecol. Lett. 18, 581–592. doi: 10.1111/ele.12427
Bottero, I., Dominik, C., Schweiger, O., Albrecht, M., Attridge, E., Brown, M. J. F., et al. (2023). Impact of landscape configuration and composition on pollinator communities across different European biogeographic regions. Front. Ecol. Evol. 11:1128228. doi: 10.3389/fevo.2023.1128228
Brook, M., Bolker, B., Kristensen, K., Mächler, M., Magnusson, A., Skaug, H., et al. (2017). glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. The R J. 9, 378–400. doi: 10.32614/RJ-2017-066
Cameron, S. A., Lozier, J. D., Strange, J. P., Koch, J. B., Cordes, N., Solter, L. F., et al. (2011). Patterns of widespread decline in north American bumble bees. Proc. Natl. Acad. Sci. USA 108, 662–667. doi: 10.1073/pnas.1014743108
Campbell, J. W., and Hanula, J. L. (2007). Efficiency of malaise traps and colored pan traps for collecting flower visiting insects from three forested ecosystems. J. Insect Conserv. 11, 399–408. doi: 10.1007/s10841-006-9055-4
Cariveau, P. D., Williams, N. M., Benjamin, F. E., and Winfree, R. (2013). Response diversity to land use occurs but does not consistently stabilize ecosystem services provided by native pollinators. Ecol. Lett. 16, 903–911. doi: 10.1111/ele.12126
Cibils, X., Calistro, P., Baráibar, N., Giambiasi, M., Giannitti, F., Machado, M., et al. (2023). ESCARABAJO SIETE DE ORO (Astylus atromaculatus): actualización y recomendaciones en vísperas del verano 2024. Revista INIA 75, 35–39.
Cusser, S., Helms, J. IV, Bahlai, C. A., and Haddad, N. M. (2021). How long do population level field experiments need to be? Utilising data from the 40-year-old LTER network. Ecol. Lett. 24, 1103–1111. doi: 10.1111/ele.13710
Cusser, S., Neff, J. L., and Jha, S. (2018). Land-use history drives contemporary pollinator community similarity. Landsc. Ecol. 33, 1335–1351. doi: 10.1007/s10980-018-0668-2
da Silva, P. M., Aguiar, C. A. S., Niemelä, J., Sousa, J. P., and Serrano, A. R. M. (2008). Diversity patterns of ground-beetles (Coleoptera: Carabidae) along a gradient of land-use disturbance. Agric. Ecosyst. Environ. 124, 270–274. doi: 10.1016/j.agee.2007.10.007
Davis, A. E., Bickel, D. J., Saunders, M. E., and Rader, R. (2023). Crop-pollinating diptera have diverse diets and habitat needs in both larval and adult stages. Ecol. Appl. 33:e2859. doi: 10.1002/eap.2859
Fernández Corujo, V. L., Basilio, M. A., and Galati, B. G. (2010). Pollen content accumulated in nests of Polybia scutellaris (hymenoptera, vespidae). Grana 49, 308–313. doi: 10.1080/00173134.2010.525663
Fernández Nion, C., and Díaz, I. (2024). Spatial distribution of pesticide use based on crop rotation data in La Plata River basin: a case study from an agricultural region of Uruguay. Environ. Monit. Assess. 196:633. doi: 10.1007/s10661-024-12759-z
Fernández Nion, C., and Díaz, I. (2025). Wetland pesticide exposure in agricultural regions: a case study from Uruguay. Integr. Environ. Assess. Manag. 21, 858–869. doi: 10.1093/inteam/vjaf028
Fijen, T. P. M., Eeraerts, M., Osterman, J., Beyer, N., Hass, A., Lundin, O., et al. (2025). Crop diversification for pollinator conservation. Landsc. Ecol. 40:19. doi: 10.1007/s10980-024-02027-3
Gallai, N., Salles, J.-M., Settele, J., and Vaissière, B. E. (2009). Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 68, 810–821. doi: 10.1016/j.ecolecon.2008.06.014
Garibaldi, L. A., Steffan-Dewenter, I., Kremen, C., Morales, J. M., Bommarco, R., Cunningham, S. A., et al. (2011). Stability of pollination services decreases with isolation from natural areas despite honey bee visits. Ecol. Lett. 14, 1062–1072. doi: 10.1111/j.1461-0248.2011.01669.x
Giannitti, F., Machado, M., Silva Silveira, C. D., Cibils-Stewart, X., Baráibar, N., Queiroz-Machado, C., et al. (2024). Experimental oral administration of pollen beetle (Astylus atromaculatus) to cattle results in an acute lethal gastrointestinal disease. Vet. Pathol. 61, 590–603. doi: 10.1177/03009858241231557
Goulson, D. (2010). Impacts of non-native bumblebees in Western Europe and North America. Appl. Entomol. Zool. 45, 7–12. doi: 10.1303/aez.2010.7
Goulson, D., Nicholls, E., Botías, C., and Rotheray, E. L. (2015). Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 347:1255957. doi: 10.1126/science.1255957
Grisales, D., and de Carvalho, C. J. B. (2019). Highland biodiversity of Fanniidae (Insecta, Diptera): fourteen new species from the Andes and Central America. Zootaxa 4551, 330–360. doi: 10.11646/zootaxa.4551.3.4
Hartig, F. (2024). DHARMa: residual diagnostics for hierarchical (multi-level / mixed) regression models. R package version 0.4.7. Available online at: https://CRAN.R-project.org/package=DHARMa (Accessed July 23, 2025)
Holzschuh, A., Dormann, C. F., Tscharntke, T., and Steffan-Dewenter, I. (2013). Mass-flowering crops enhance wild bee abundance. Oecologia 172, 477–484. doi: 10.1007/s00442-012-2515-5
INUMET. (2022). Boletín anual de precipitaciones 2022. Available online at: https://www.inumet.gub.uy/ (accessed July 13, 2025).
INUMET. (2023a). Boletín anual de precipitaciones 2023. Available online at: https://www.inumet.gub.uy/ (accessed July 13, 2025).
INUMET. (2023b). Sequía meteorológica en Uruguay 2020–2023. Available online at: https://www.inumet.gub.uy/ (accessed July 13, 2025).
IPBES. (2017). The assessment report on pollinators, pollination and food production. Available online at: https://www.ipbes.net (accessed July 13, 2025).
Isbell, F., Calcagno, V., Héctor, A., Connolly, J., Harpole, W. S., Reich, P. B., et al. (2011). High plant diversity is needed to maintain ecosystem services. Nature 477, 199–202. doi: 10.1038/nature10282
Kearns, C. A., Inouye, D. W., and Waser, N. M. (1998). Endangered mutualisms: the conservation of plant–pollinator interactions. Annu. Rev. Ecol. Syst. 29, 83–112. doi: 10.1146/annurev.ecolsys.29.1.83
Kennedy, C. M., Lonsdorf, E., Neel, M. C., Williams, N. M., Ricketts, T. H., Winfree, R., et al. (2013). A global quantitative synthesis of local and landscape effects on wild bee pollinators in agroecosystems. Ecol. Lett. 16, 584–599. doi: 10.1111/ele.12082
Kevan, P. G. (1999). Pollinators as bioindicators of environmental status: species, activity, and diversity. Agri. Ecosys. Environ. 74, 373–393.
Klein, A. M., Vaissière, B. E., Cane, J. H., Steffan-Dewenter, I., Cunningham, S. A., Kremen, C., et al. (2007). Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B 274, 303–313. doi: 10.1098/rspb.2006.3721
Landis, D. A. (2017). Designing agricultural landscapes for biodiversity-based ecosystem services. Basic Appl. Ecol. 18, 1–12. doi: 10.1016/j.baae.2016.07.005
Love, M. I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15:550. doi: 10.1186/s13059-014-0550-8
Maggi, T., Pardo, L., and Chreil, R. (2023). Pollinator diversity: a key to ecosystem resilience and food security. Available online at: https://www.researchgate.net/publication/371864583_Pollinator_Diversity_A_Key_to_Ecosystem_Resilience_and_Food_Security
MGAP-DIEA. (2024). Encuesta agrícola: Invierno 2024. Ministerio de Ganadería, Agricultura y Pesca. Available online at: https://www.gub.uy/ministerio-ganaderia-agricultura-pesca/comunicacion/noticias/diea-presenta-resultados-encuesta-agricola-invierno-2024 (accessed July 13, 2025).
Millard, J., Outhwaite, C. L., Kinnersley, R., Freeman, R., Gregory, R. D., Adedoja,, et al. (2021). Global effects of land-use intensity on local pollinator biodiversity. Nat. Commun. 12:2902. doi: 10.1038/s41467-021-23228-3
Ministerio de Ambiente. (2023). Plan de Monitoreo de. Río San Salvador, Informe de Actividades 2021. División Calidad Ambiental. Available online at: www.ambiente.gub.uy (accessed July 13, 2025).
Modernell, P., Rossing, W. A. H., Corbeels, M., Dogliotti, S., Picasso, V., and Tittonell, P. (2016). Land use change and ecosystem service provision in pampas and Campos grasslands of southern South America. Environ. Res. Lett. 11:3002. doi: 10.1088/1748-9326/11/11/113002
Mondelli, M., Simeto, S., and Rivas, M. (2015). Inventario de cultivos en la cuenca del río San Salvador. Uruguay: Instituto Nacional de Investigación Agropecuaria (INIA).
Ollerton, J., Winfree, R., and Tarrant, S. (2011). How many flowering plants are pollinated by animals? Oikos 120, 321–326. doi: 10.1111/j.1600-0706.2010.18644.x
Pierre, J., and Hofs, J. L. (2010). Astylus atromaculatus (Coleoptera: Melyridae): Abundance and role in pollen dispersal in Bt and non-Bt cotton in South Africa. Environ. Entomol. 39, 1523–1531. doi: 10.1603/EN09142
Posit Team (2024). RStudio: Integrated development environment for R. Posit software, PBC. Boston, MA: Posit Team.
Potts, S. G., Biesmeijer, J. C., Kremen, C., Neumann, P., Schweiger, O., and Kunin, W. E. (2010). Global pollinator declines: trends, impacts and drivers. Trends Ecol. Evol. 25, 345–353. doi: 10.1016/j.tree.2010.01.007
Rotheray, G. E., and Gilbert, F. S. (2011). The natural history of hoverflies. Reino Unido: Forrest Text, Cardigan, 333.
Santos, E., Daners, G., Morelli, E., and Galván, G. A. (2020a). Diversity of bee assemblage (family Apidae) in natural and agriculturally intensified ecosystems in Uruguay. Environ. Entomol. 49, 1232–1241. doi: 10.1093/ee/nvaa078
Santos, E., Morelli, E., and Galván, G. A. (2020b). Recortes de hoja de soja (Glycine max Merril) son utilizados para construcción de nidos de abejas nativas (Hymenoptera: Apidae), Megachilinae de Uruguay. Bol. Soc. Zool. Urug. 29, 183–186. doi: 10.26462/29.2.17
Santos, E. I., Morelli, E., and Galván, G. A. (2021). Dependence of soybeans (Glycine max Merril) to entomophile pollination and study of food resources of honey bees (Apis mellifera L.) in contact with the crop. Agrocienc. Urug. 25:e1523. doi: 10.31285/AGRO.25.1523
Saunders, M. E., Janes, J. K., and O'Hanlon, J. C. (2020). Semantics of the insect decline narrative: recommendations for communicating insect conservation to peer and public audiences. Insect Conserv. Diversity 13, 211–213. doi: 10.1111/icad.12406
Soroye, P., Newbold, T., and Kerr, J. T. (2020). Climate change contributes to widespread declines among bumble bees across continents. Science 367, 685–688. doi: 10.1126/science.aax8591
Stefanescu, C., Asís, J. D., Baños-Picón, L., Cerdà, X., García, M. A. M., Micó,, et al. (2018). Diversidad de insectos polinizadores en la península ibérica. Ecosistemas 27, 9–22. doi: 10.7818/ECOS.1391
Steffan-Dewenter, I., and Westphal, C. (2008). The interplay of pollinator diversity, pollination services and landscape change. J. Appl. Ecol. 45, 737–741. doi: 10.1111/j.1365-2664.2008.01483.x
Tscharntke, T., Tylianakis, J. M., Rand, T. A., Didham, R. K., Fahrig, L., Batáry, P., et al. (2012). Landscape moderation of biodiversity patterns and processes - eight hypotheses. Biol. Rev. 87, 661–685. doi: 10.1111/j.1469-185X.2011.00216.x
van Nieukerken, J., Kaila, L., Kitching, I. J., Kristensen, N. P., Lees, D. C., Minet, J., et al. (2011). Order Lepidoptera Linnaeus, 1758. Zootaxa 3148, 212–221. doi: 10.11646/zootaxa.3148.1.41
Vrdoljak, S. M., and Samways, M. J. (2012). Optimising coloured pan traps to survey flower visiting insects. J. Insect Conserv. 16, 345–354. doi: 10.1007/s10841-011-9420-9
Wardhaugh, C. W. (2015). How many species of arthropods visit flowers? Arthropod Plant Interact. 9, 547–565. doi: 10.1007/s11829-015-9398-4
Westphal, C., Steffan-Dewenter, I., and Tscharntke, T. (2003). Mass flowering crops enhance pollinator densities at a landscape scale. Ecol. Lett. 6, 961–965. doi: 10.1046/j.1461-0248.2003.00523.x
Winfree, R., Aguilar, R., Vázquez, D. P., LeBuhn, G., and Aizen, M. A. (2009). A meta-analysis of bees' responses to anthropogenic disturbance. Ecology 90, 2068–2076. doi: 10.1890/08-1245.1
Woodcock, B. A., Isaac, N. J. B., Bullock, J. M., Roy, D. B., Garthwaite, D. G., Crowe, A., et al. (2017). Impacts of neonicotinoid use on long-term population changes in wild bees in England. Nat. Commun. 8:1093. doi: 10.1038/s41467-017-02269-3
Keywords: insect pollinators, diversity, agricultural landscape, Coleoptera, Diptera, Hymenoptera, Lepidoptera
Citation: Salvarrey S, Castelli L, Invernizzi C, Suárez E, Ramos Y, Aldabe J, Bentancur G, Burla JP, Remedios M, Díaz I, Fernández Nion C, Santos E, Teixeira de Mello F and Arbulo N (2025) Agricultural intensification favors dominant species while changing pollinator community compositions in a subtropical watershed of Uruguay. Front. Sustain. Food Syst. 9:1672127. doi: 10.3389/fsufs.2025.1672127
Edited by:
Liming Ye, Ghent University, BelgiumReviewed by:
Justin Bastow, Eastern Washington University, United StatesSarah Nanyiti, National Crops Resources Research Institute (NaCRRI), Uganda
Copyright © 2025 Salvarrey, Castelli, Invernizzi, Suárez, Ramos, Aldabe, Bentancur, Burla, Remedios, Díaz, Fernández Nion, Santos, Teixeira de Mello and Arbulo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sheena Salvarrey, c3NhbHZhcnJleUBmY2llbi5lZHUudXk=
†These authors have contributed equally to this work