Abstract
Recently, there is an urgent need for new drugs due to the emergence of drug resistant pathogenic microorganisms and new infectious diseases. Members of phylum Actinobacteria are promising source of bioactive compounds notably antibiotics. The search for such new compounds has shifted to extreme or underexplored environments to increase the possibility of discovery. Cave ecosystems have attracted interest of the research community because of their unique characteristics and the microbiome residing inside including actinobacteria. At the time of writing, 47 species in 30 genera of actinobacteria were reported from cave and cave related habitats. Novel and promising bioactive compounds have been isolated and characterized. This mini-review focuses on the diversity of cultivable actinobacteria in cave and cave-related environments, and their bioactive metabolites from 1999 to 2018.
Introduction
Caves are generally regarded as any natural underground chamber that is large enough for human entrance. They can be classified based on type of rock and formation method. The most common types of caves are limestone and other calcareous rocks (Northup and Lavoie, 2001). Though caves have been studied for hundreds of years, their microbiome are generally underexplored and overlooked. Caves are attracting the interests of microbiologists, in terms of microbial diversity, during the past decade (Laiz et al., 1999; Barton et al., 2004; Barton, 2006). It is believed that microbes collected from pristine sites that are unexplored or rarely visited by humans are likely to be novel taxa or strains which produce unique beneficial chemical compounds. Market demand for new drugs is on the rise due to the emergence of new diseases and drug resistant pathogens (Genilloud, 2017; Kemung et al., 2018; Takahashi and Nakashima, 2018). With a combination of unique conditions including high humidity, relatively low and stable temperature, and low nutrients, caves are expected to harbor novel microorganisms with biotechnological benefits. Members of actinobacteria are reported to be a dominant microbial population in several cave ecosystems (Groth and Saiz-Jimenez, 1999; Cheeptham et al., 2013; Tomczyk-Zak and Zielenkiewicz, 2016; Ghosh et al., 2017).
Actinobacteria are large group of high G+C Gram positive bacteria (Barka et al., 2016). They are regarded as the most prolific source of bioactive compounds in particular commercially available antibiotics. Actinobacteria produce approximately two-thirds of all know antibiotics in the market, most of these are from members of the genus Streptomyces (Barka et al., 2016). Several members of diverse actinobacterial taxa were also found to produce wide range of other biologically active compounds, for examples antibacterial, anticancer, or antifungal drugs (Barka et al., 2016; Genilloud, 2017; Castro et al., 2018; Takahashi and Nakashima, 2018). Isolation of actinobacteria from unique natural habitats is of interest to avoid re-isolation of strains that produce known bioactive metabolites and usually lead to highly diverse actinobacterial communities. The present mini-review provides evidence that actinobacteria from caves are expected to be a good source for drug discovery (Yücel and Yamac, 2010; Cheeptham et al., 2013; Kay et al., 2013; Ghosh et al., 2017; Riquelme et al., 2017).
Selective Isolation of Cave Actinobacteria
In the past decade, there are many reports on the discovery of novel actinobacteria in cave habitats. Successful isolation of actinobacteria from caves depend largely on factors of (1) media composition (Kim et al., 1998) (2) culture condition, and (3) pretreatment methods (Kim et al., 1998; Nakaew et al., 2009a,b; Duangmal et al., 2012; Niyomvong et al., 2012; Velikonja et al., 2014; Fang et al., 2017b; Adam et al., 2018). Media used for the isolation of cave actinobacteria range from routine cultivation media such as International Streptomyces Project medium 2 (yeast malt extract agar, ISP2) or tryptic soy agar (TSA) to selective media including humic acid vitamin agar (HV), starch casein agar (SC), starch casein nitrate agar (SCN), peptone-yeast extract/brain-heart infusion medium (PY-BHI), R2A medium, actinomycete isolation agar (AI), and Gauze's medium No.1. Moreover, isolation media that mimic the conditions of low concentration nutrients in caves such as tap water agar, 1/100 ISP2 and oligotrophic medium (M5) were also successfully used for the isolation of actinobacteria. (Lee et al., 2000b; Velikonja et al., 2014; Covington et al., 2018; Passari et al., 2018). High concentration of nutrients in standard cultivation media were reported to cause cell death in cave-associated bacteria due to osmotic stress (Barton, 2006; Ghosh et al., 2017).
Two important culture conditions for actinobacteria isolation are incubation temperature and incubation time. Four incubation temperatures (5°, 13°, 20°, and 28°C) were used for the isolation of soil bacteria including actinobacteria from three caves in Northern Spain (Laiz et al., 2003). The incubation temperature of 5°C was used to represent cave temperature and target psychrotrophs, 28°C as laboratory incubation temperature and 20°C as intermediate temperature between cave and laboratory conditions. The highest number of actinobacterial isolates (mostly sporoactinomycetes) was obtained at 28°C followed by 13°, 20° and 5°C, respectively. However, a higher diversity was observed from 13°C than 28°C. Therefore, these authors concluded that the isolation of actinobacteria is a temperature-dependent process. In addition, longer incubation time was successfully used to promote the recovery of slow-growing actinobacteria (Laiz et al., 2003).
Pretreatment, both chemical and physical methods are generally useful for isolation of various actinobacterial species. Physical pretreatments involve the use of air drying, moist heat, dry heat and electromagnetic wave. Moist heating (water bath at 50°C for 5–6 min) is useful for eliminating of fast growing bacteria (Niyomvong et al., 2012; Velikonja et al., 2014). Dry heating at 120°C for 1 h is effective in reducing number of unwanted bacteria and found to be an effective method for isolation of members of the genera Dactylosporangium, Streptosporangium and Microbispora, while growth of streptomycetes was limited (Jiang et al., 2016). In addition, dry heating with or without phenol treatment resulted in a reduction of bacteria and heat-labile Streptomyces, thus heat resistant rare actinobacteria were readily isolated (Kim et al., 1998; Nakaew et al., 2009a; Niyomvong et al., 2012). However, these treatments also affect the number of viable actinobacteria (Niyomvong et al., 2012). Pretreatment using microwave irradiation was effective for the isolation of rare actinobacteria (Niyomvong et al., 2012) and capable of inducing spore germination in some species of Streptomyces, Nocardia, Streptosporangium, Lentzea, Micromonospora, and Micropolyspora (currently transferred to Nocardia) (Bulina et al., 1997; Wang et al., 2013; Velikonja et al., 2014). For chemical pretreatment, the type and concentration of calcium salts are important for the isolation of actinobacteria (Fang et al., 2017b). Selective media supplemented with CaCO3 yield higher actinobacterial count than those supplemented with CaCl2 and (CH3COO)2Ca. The concentration of these three salts, at low concentration (0.1 and 0.01% (w/v) yield higher CFU of actinobacteria than in its absence or at high concentration. Calcium is important for environmental stress tolerance in actinobacteria because calcium forms a compound with dipicolinic acid as calcium dipicolinate and acts as secondary stabilizing agent for spore against environmental stress (Moir and Smith, 1990).
Novel Actinobacterial Taxa
Several novel actinobacterial taxa isolated from caves and cave related habitats during the period of 20 years from 1999 to 2018 were summarized in Table 1. In total, 47 species within 30 genera were described including 7 novel genera. The highest number of novel species was from genus Streptomyces (5) followed by Amycolatopsis (4) and Nocardia (4). The majority of these novel actinobacteria were isolated from cave soils including 6 novel genera, Antricoccus, Beutenbergia, Knoellia, Lysinibacter Spelaeicoccus and Sphaerimonospora. Only the genus Hoyosella was recovered from complex biofilm on the ceiling and wall of Altamira cave, Spain. The extreme conditions within the caves are expected to create stress for the inhabitant microorganisms at the genetic level, paving the way for the evolution of new species and their novel metabolites (Tawari and Gupta, 2013). Therefore, caves are considered as an attractive source for the isolation of novel actinobacterial taxa.
Table 1
| Family | Genus | Species | Sources | Media | References |
|---|---|---|---|---|---|
| Brevibacteriaceae | Spelaeicoccus | Spelaeicoccus albus | Soil from natural cave in Jeju, Korea | Starch casein agar | Lee, 2013b |
| Conexibacteraceae | Conexibacter | Conexibacter stalactiti | Pieces of stalactites from Yongcheon cave in Jeju, Korea | Starch casein agar | Lee, 2017 |
| Glycomycetaceae | Stackebrandtia | Stackebrandtia cavernae | Rocks from karst cave, Guizhou, south-west China | R2A agar with 0.1% CaCO3 | Zhang et al., 2016 |
| Intrasporangiaceae | Fodinibacter | Fodinibacter luteus | Sample from wall of a salt mine in Yunnan, China | Marine agar 2216 | Wang et al., 2009 |
| Knoellia | Knoellia sinensis | Soils from the Reed Flute cave near Guilin, Guangxi, China | Casein mineral medium | Groth et al., 2002 | |
| Knoellia subterranea | Soils from the Reed Flute cave near Guilin, Guangxi, China | Peptone/yeast extract/brain- heart infusion medium (PY-BHI) | Groth et al., 2002 | ||
| Ornithinimicrobium | Ornithinimicrobium cavernae | Stalagmites from karst cave in Luoyang country, Henan, northern China | ISP 2 with nystatin and nalidixic acid | Zhang et al., 2018 | |
| Kineosporiaceae | Augustibacter | Augustibacter spluncae | Pieces of stalactites from Yongcheon cave in Jeju, Korea | Starch casein agar | Ko and Lee, 2017 |
| Microbacteriaceae | Agromyces | Agromyces subbeticus | Cyanobacterial biofilm from Cave of Bats, near Zuheros, Cordoba, southern Spain | Peptone/yeast extract/ brain- heart infusion medium (PY-BHI) | Jurado et al., 2005 |
| Humibacter | Humibacter antri | Clay soils from natural cave in Jeju, Korea | Starch casein agar | Lee, 2013a | |
| Lysinibacter | Lysinibacter cavernae | Soils from wild karst cave in the Wulong region, Chongqing, China | FA (fulvic acid) agar | Tuo et al., 2015 | |
| Micrococcaceae | Arthrobacter | Arthrobacter psychrophenolicus | Carbonate-rich deposit from Alpine ice cave, Salzburg, Austria | Soil-extract agar | Margesin et al., 2004 |
| Beutenbergia | Beutenbergia cavernae | Soils from the Reed Flute cave near Guilin, Guangxi, China | Casein mineral medium and peptone/yeast extract/brain-heart infusion medium | Groth et al., 1999 | |
| Micromonosporaceae | Catellatospora | Catellatospora koreensis | Soils from gold-mine cave in Kongju, Korea | Yeast extract; glucose; K2HPO4;Na2HPO4;KNO3; NaCl; MgSO4.7H2O; CaCl2.2H2O and trace mineral solution | Lee et al., 2000a |
| Micromonospora | Micromonospora kangleipakensisi | Sample from limestone quarry at Hundung, Manipur, India | Gauze's medium | Nimaichand et al., 2013c | |
| Mycobacteriaceae | Hoyosella | Hoyosella altamirensis | Complex bioflim on the cave ceiling and walls from Altamira cave, Cantabria, Spain | Starch casein agar | Jurado et al., 2009 |
| Nocardioidaceae | Jiangella | Jiangella alkaliphila | Soils from natural cave on Jeju island, Korea | Starch casein agar | Lee, 2008 |
| Nocardioides | Nocardioides cavernae | Soils from karst cave in Xingyi county, Guizhou, south-western China | R2A agar with cycloheximide and nalidixic acid | Han et al., 2017 | |
| Tenggerimyces | Tenggerimyces flavus | Soil from Shenxian cave, Henan, China | R2A agar with cycloheximide, nalidixic acid and potassium dichromate | Li et al., 2016 | |
| Norcardiaceae | Nocardia | Nocardia altamirensis | Complex microbial community forming a gray-colored colonization on the walls from Altamira cave, Cantabria, Spain | Tryptose soy agar | Jurado et al., 2008 |
| Nocardia cavernae | Soil from karst cave in Xingyi county, Guizhou, south-western China | Humic acid-vitamin agar with cycloheximide and nalidixic acid | Li et al., 2017 | ||
| Nocardia jejuensis | Soil from natural cave on Jeju island, Korea | Starch casein agar | Lee, 2006c | ||
| Nocardia speluncae | Soil from natural cave on Jeju island, Korea | Starch casein agar | Seo et al., 2007 | ||
| Rhodococcus | Rhodococcus antrifimi | Dried bat dung from natural cave in Jeju, Korea | Starch casein agar | Ko et al., 2015 | |
| Rhodococcus canchipurensis | Soil from limestone quarry at Hundung, Manipur, India | Starch casein nitrate agar | Nimaichand et al., 2013a | ||
| Propionibacteriaceae | Microlunatus | Microlunatus cavernae | Soil from Alu ancient cave, Yunnan, south-west China | R2A medium | Cheng et al., 2013a |
| Pseudonocardiaceae | Amycolatopsis | Amycolatopsis halotolerans | Soil from natural cave in Jeju island, Korea | Starch/casein agar | Lee, 2006b |
| Amycolatopsis jejuensis | Dried bat dung from natural cave in Jeju island, Korea | Starch casein agar | Lee, 2006b | ||
| Amycolatopsis jiguanensis | Soil from Ji Guan cave, Henan, middle-eastern China | ISP2 | Huang et al., 2016 | ||
| Amycolatopsis xuchangensis | Soil from Ji Guan cave, Henan, middle-eastern China | ISP2 | Huang et al., 2016 | ||
| Lentzea | Lentzea cavernae | Limestone from karst cave in Xingyi county, Guizhou, south-western China | Humic acid-vitamin agar with cycloheximide and nalidixic acid | Fang et al., 2017a | |
| Lentzea guizhouensis | Limestone from Puding karst ecosystem research station of the Chinese Academy of Sciences in Guizhou Province, south-west china | Modified ATCC-172 medium at 1/10 concentration | Cao et al., 2015 | ||
| Saccharothrix | Saccharothrix albidocapillata | Soil from gold mine cave in Kongju, Korea | Tap water agar and oligotrophic (M5) medium | Lee et al., 2000b | |
| Saccharothrix violacea | Soil from gold mine cave in Kongju, Korea | Tap water agar and oligotrophic (M5) medium | Lee et al., 2000b | ||
| Saccharopolyspora | Saccharopolyspora cavernae | Swallow cave, Yunnan, south-west China | Improved DSMZ medium 405 | Cheng et al., 2013b | |
| Streptomycataceae | Streptomyces | Streptomyces boninensis | Soil from limestone cave, Ogasawa islands, Tokyo, Japan | Humic acid-vitamin agar with benlate and nalidixic acid | Také et al., 2018 |
| Streptomyces canchipurensis | Soil from limestone quarry at Hundung, Manipur, India | Gauze's medium No. 1 | Li et al., 2014 | ||
| Streptomyces hundungensis | Soil from limestone quarry at Hundung, Manipur, India | Starch casein nitrate agar | Nimaichand et al., 2013b | ||
| Streptomyces lunaelactis | Moonmilk deposit from Grotte des Collemboles cave in Comblain-au-Pont, Belgium | ISP and starch nitrate (SN) medium with nalidixic acid and nystatin | Maciejewska et al., 2015 | ||
| Streptomyces manipurensis | Soil from limestone quarry at Hundung, Manipur, India | Starch casein Nitrate agar | Nimaichand et al., 2012 | ||
| Streptosporangiaceae | Sphaerimonospora | Sphaerimonospora thailandensis (formerly Microbispora thailandensis) | Soil from tropical limestone cave (Khao No-Khao Kaeo), Nakorn Sawan, Thailand | Humic acid-vitamin agar with nalidixic acid and ketoconazole | Duangmal et al., 2012 |
| Sphaerimonospora cavernae | Soil from tropical limestone cave (Khao No-Khao Kaeo), Nakorn Sawan, Thailand | - | Mingma et al., 2016 | ||
| Nonomuraea | Nonomuraea monospora | Soil from Pha Tup cave forest park, Nan, Thailand | Humic acid-vitamin agar with nystatin and cycloheximide | Nakaew et al., 2012 | |
| Nonomuraea indica | Soil from limestone open pit mine from Gulbarga region, Karnataka, India | Starch casein agar | Quadri et al., 2015 | ||
| Thermomonosporaceae | Actinocorallia | Actinocorallia cavernae | Natural cave in Jeju, Korea | Starch/casein agar | Lee, 2006a |
| Not assigned to family (Suborder Frankineae) | Antricoccus | Antricoccus suffuscus | Soil from natural cave in Jeju, Korea | Starch casein agar | Lee, 2015 |
Novel actinobacterial taxa isolated from cave and related habitats between 1999 and 2018.
Most species were isolated from selective media that were designed for the isolation of actinobacteria such as humic acid vitamin agar, starch casein agar, starch casein nitrate agar. However, some novel species were isolated using general cultivation media such as ISP2 media (Amycolatopsis jiguanensis and A. xuchangensis) and TSA (Nocardia altamirensis). In addition, low nutrient media (tap water agar and oligotrophic M5 media) were preferable for the isolation of Saccharothrix violacea and S. albidocapillata. Most novel species were incubated at 28°–30°C for 1–6 weeks. However, Arthrobacter psychrophenolicus was isolated at 4°C, this may be because this species originated from Alpine ice cave in Salzburg, Austria (Margesin et al., 2004). Lysinibacter cavernae was isolated at 15°C from soil in a wild karst cave in the Wulong region, Chongqing, China (Tuo et al., 2015). Streptomyces lunaelactis was isolated at 17°C from a moonmilk deposit in the Grotte des Collemboles cave in Belgium (Maciejewska et al., 2015).
Pretreatment procedures were also useful for isolation of some novel species. For example, Microbispora thailandensis was isolated from soil pretreated with microwave radiation at a frequency of 2460 MHz and power setting of 100 W for 45 s (Duangmal et al., 2012). Nonomuraea monospora was isolated from soil treated with phenol (Nakaew et al., 2012). Streptomyces manipurensis was isolated from soil supplemented with 0.1 g of CaCO3 for 1 day to prevent the growth of fast growing bacteria (Nimaichand et al., 2012).
Bioactive Compounds From Cave Actinobacteria
Caves are extreme habitats with low nutrient, temperature and light intensity but have high humidity (Schabereiter-Gurtner et al., 2002). These unique characteristics may promote the production of bioactive substances in particular antibiotics by actinobacteria (Nakaew et al., 2009a). Bioactive metabolites from cave associated actinobacteria have been purified, their structure elucidated and reported in recent years (Table 2). These compounds mostly displayed anti-bacterial and/or anti-cancer activities. The most prolific producer is members of the genus Streptomyces.
Table 2
| Bioactivity | Compounds | Structure | Producing strain | Source of strain | References |
|---|---|---|---|---|---|
| Antibacterial | Cervimycins A, B, C, and D | 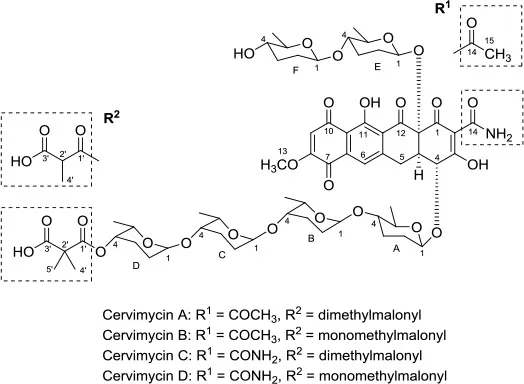 | Streptomyces tendae strain HKI 0179 | Rock wall from Ancient cave, The Grotta dei Cervi, Italy | Herold et al., 2005 |
| Undecylprodigiosin | 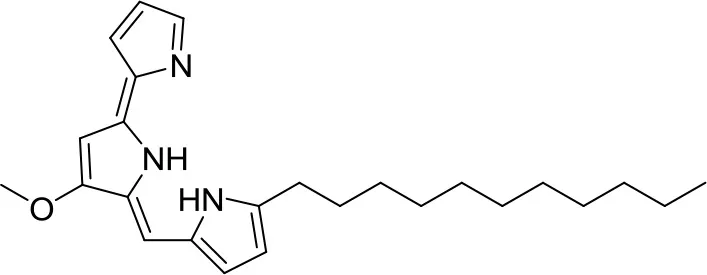 | Streptomyces sp. JS520 | Cave on mountain Miroc, Serbia | Stankovic et al., 2012 | |
| Xiakemycin A | 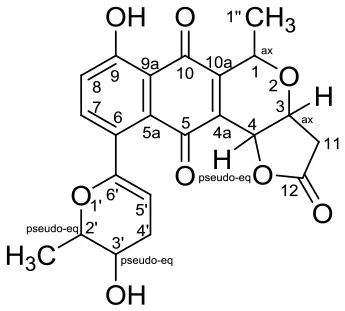 | Streptomyces sp. CC8-201 | Soil from karst cave,Chongqing city, China | Jiang et al., 2015 | |
| Chaxalactin B | 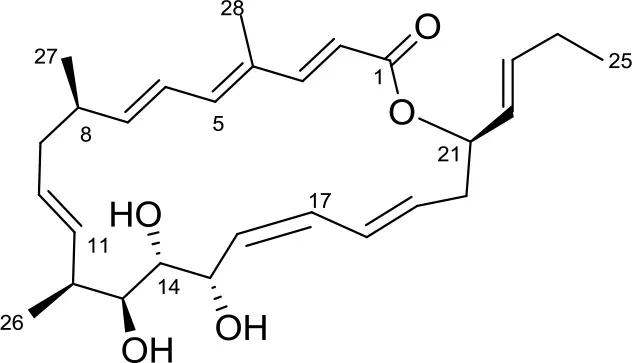 | Streptomyces sp. IB 2014/I/ 78-8 | Bolshaya Oreshnaya cave in the Mansk area of the Krasnoyaesk, Siberia, Russia | Axenov-Gibanov et al., 2016 | |
| Anticancer | Hypogeamicins A | 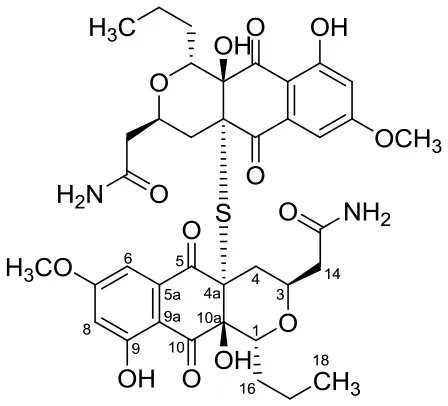 | Nonomuraea specus | Hardin's cave system located close to Ashland City, Tennessee | Derewacz et al., 2014 |
| Xiakemycin A | 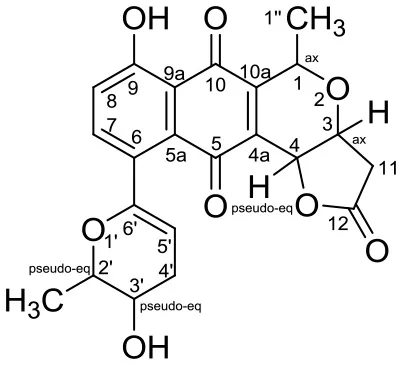 | Streptomyces sp. CC8-201 | Soil from karst cave,Chongqing city, China | Jiang et al., 2015 | |
| Huanglongmycin (HLM) A, | 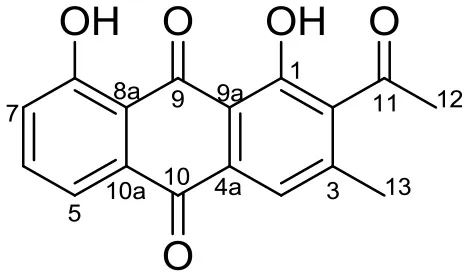 | Streptomyces sp. CB09001 | Soil from karstic cave in Xiangxi, China | Jiang et al., 2018 | |
| Antioxidative activity | Undecylprodigiosin | 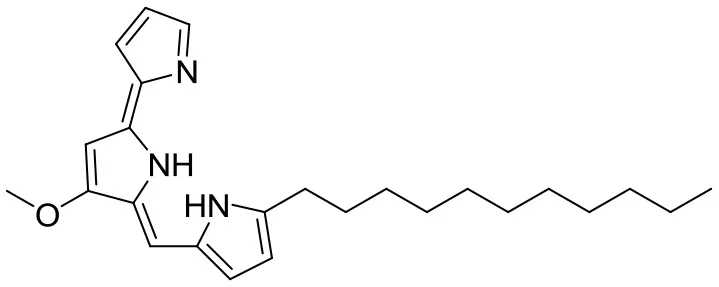 | Streptomyces sp. JS520 | Cave on mountain Miroc, Serbia | Stankovic et al., 2012 |
| Inhibitory activity against lipid metabolism | Gyrophoric acid | 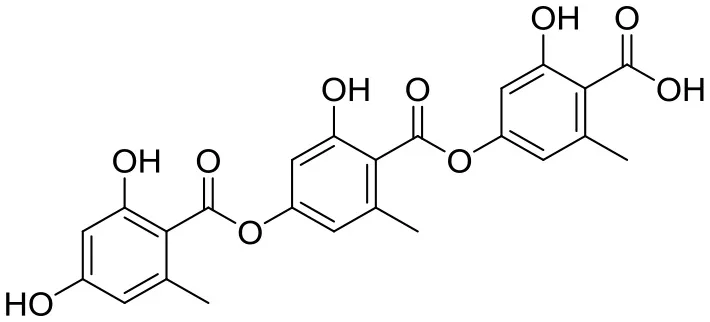 | Streptomyces sp. IB 2014/I/ 78-8 | Bolshaya Oreshnaya cave in the Mansk area of the Krasnoyaesk, Siberia, Russia | Axenov-Gibanov et al., 2016 Tomoda and Omura, 2001 |
Bioactive metabolites from cave actinobacteria.
Cervimycin A, B, C, and D were produced from Streptomyces tendae strain HKI 0179, isolated from a rock wall in an ancient cave, the Grotta dei Cervi in Italy. Cervimycins A and B are novel polyketide glycosides. However, cervimycin C and D have the same structure as known compounds A2121-3 and A2121-2. Cervimycins A-D are highly active against Gram positive bacteria (B. subtilis and S. aureus) and multi-drug-resistant S. aureus (MRSA), vancomycin-resistant Enterococcus faecalis (VRE) and efflux-resistant S. aureus EfS4 (Herold et al., 2005).
Xiakemycin A is a novel pyranonaphthoquinone (PNQ) antibiotics produced by Streptomyces sp. CC8-201 from remote karst soil in China. Xiakemycin A showed strong inhibitory activities against Gram positive bacteria (S. aureus, S. epidermidis, E. faecalis, and E. faecium) and cytotoxic against numerous cancer cell lines (human lung cancer A549 cells, breast cancer MCF-7 cells, hepatoma HepG-2 cell, cervical cancer HeLa cells, colon carcinoma HCT-116 cell p53 wt cells, neuroblastoma SH-SY5Y cells, and human prostate cancer PC-3) (Jiang et al., 2015).
Hypogeamicins A, B, C, and D were produced by Nonomuraea specus isolated from Hardin's cave system in Tennessee, USA. Hypogeamicin A showed cytotoxicity to colon cancer cell line TCT-1 while hypogeamicin B-D were active against B. subtilis with no cytotoxicity to TCT-1. However, hypogeamicin B-D are not as potent as erythromycin and gentamicin in terms of antimicrobial activity against B. subtilis (Derewacz et al., 2014).
Huanglongmycin A, B, and C are aromatic polyketides from Streptomyces sp. CB09001, isolated from karstic cave soil of Xiangxi, China. Huanglongmycin A showed a weak anti-Gram negative bacteria (Pseudomonas aeruginosa and Escherichia coli) and moderate cytotoxicity against A549 lung cancer cell line. Huanglongmycin B has weak antibacterial activity against S. aureus and multi-drug-resistant S. aureus (MRSA). Huanglongmycin C showed neither antibacterial nor anticancer activities (Jiang et al., 2018). Undecylprodigiosin was produced by Streptomyces sp. JS520 isolated from sediments in cave in the mountain Miroc, Serbia. Undecylprodigiosin is a deep red pigment with antibacterial activity against Micrococcus luteus, B. subtilis, and C. albicans. Moreover, undecylprodigiosin also showed antioxidative and UV-protective properties (Stankovic et al., 2012).
Four known compounds with bioactivity (cyclodysidin D, chaxalactin B, stylissazole B, and gyrophoric acid) were reported to produce by Streptomyces sp. IB 2014/I/ 78-8 from moonmilk speleothem of Bolshaya Oreshnaya cave in Siberia (Axenov-Gibanov et al., 2016). Cyclodysidin D is previously reported in marine sponge, Dysidea tupha associated Streptomyces sp. RV 15. This compound showed no activity against bacteria, fungi and parasites (Abdelmohsen et al., 2014). Chaxalactin B was produced from Streptomyces sp. C34 from a hyper-arid soil samples collected from the Atacama Desert, Chile. This compound has strong activity against Gram positive bacteria (Castro et al., 2018). Stylissazole B was isolated from the marine sponge Stylissa carteri collected in the Solomon islands but no report on bioactivity (Patel et al., 2010). Gyrophoric acid isolated from Humicola sp. FO-2942 is an inhibitor of diacylglycerol acyltransferase and a lipid-lowering agent (Inokoshi et al., 2010).
Bioactivity of Uncharacterized Compounds
Several cave actinobacteria have been screened for their biological activity such as antibacterial, anticancer and antifungal. However, no pure compound and their structure were reported in these studies. The screening of only bioactivity without the structure elucidation of bioactive metabolites may not useful for the discovery of new antibiotics (Hug et al., 2018). Nevertheless, these findings provide evidence which supports the potential of cave actinobacteria to be exploited for novel bioactive compounds.
Turkish karstic caves were reported to harbor actinobacteria, for which 62% of the isolates, were active against several microbial pathogens (Gram positive bacteria, Gram negative bacteria, yeast, and filamentous fungi). Streptomyces sp. 1492 had strong activity against clinical strains of MRSA, VRE, and Acinetobacter baumanii (Yücel and Yamac, 2010). Streptomyces E9 isolated from Helmcken Falls cave in British Columbia could inhibit the growth of Paenibacillus larvae, a causative agent of American foulbrood disease in honeybees (Kay et al., 2013). A moonmilk speleothems of limestone caves, Grotte des Collemboles in Belgium were investigated for antimicrobial producing cultivable actinobacteria. A collection of obtained Streptomyces displayed strong inhibitory activity against Gram positive and Gram negative bacteria (Maciejewska et al., 2016). In a study of cultivable actinobacteria from Azores volcanic caves in Portugal, 18.1% of 148 actinobacterial isolates have antibacterial activity against at least one of the following bacteria: Salmonella typhimurium, E. coli, P. aeruginosa, Proteus sp., Listeria monocytogenes, L. innocua, and S. aureus. Most of the active isolates belong to the genus Streptomyces (S. nojiriensis, S. spiroverticillatus, S. avidinii, and S. mauvecolor) followed by Arthrobacter (Riquelme et al., 2017). A total of 40 taxa belonging to the genera Agromyces, Amycolatopsis, Kocuria, Micrococcus, Micromonospora, Nocardia, Streptomyces, and Rhodococcus were recovered from moonmilk deposits inside the Grotte des Collemboles, Belgium. Antimicrobial activity was found in isolated strains against Gram positive bacteria (87%) and Gram negative bacteria (59%) (Adam et al., 2018). Sixteen isolates of Streptomyces spp. from Chaabe cave in Algeria were screened for their antimicrobial activity using agar cylinder method. All of them showed strong anti-Gram positive (S. aureus, M. luteus, L. monocytogenes, and B. subtilis) activity (Belyagoubi et al., 2018).
For anticancer activity, a rare actinobacterium Spirillospora albida strain CMU-PNK470 was isolated from Phanangkhoi cave in northern Thailand (Nakaew et al., 2009a). This bacterium showed activity against human small lung cancer cell (NCI-H1870) with an IC50 value of 10.18 μg/ml. Similarly, Nonomurea roseola strain PT708 isolated from Phatup cave forest park in northern Thailand was tested positive for anticancer activity against human oral cavity cancer (KB) and human small lung cancer cells (NCI-H187) (Nakaew et al., 2009b). Moreover, these two strains are also active against some Gram positive pathogenic bacteria (B. cereus, MRSA, and Paenibacillus larvae).
Some examples of antifungal activity from cave actinobacteria have been reported. Antagonistic Streptomyces, Micromonospora, Streptosporangium, and Dactylosporangium were isolated from five caves (Cheondong, Kosoo, Nadong, Seonglyu, and Ssangyong) in Korea (Kim et al., 1998). They showed activity against at least one of plant pathogenic fungi (Alternaria solani, Colletotrichum gloeosporioides, Fusarium oxysporum f.sp. lycopersici, Magnaporthe grisea, Phytophthora capsici, and Rhizoctonia solani). Similarly, members of genera Streptomyces and Janibacter isolated from limestone deposit sites in Hundung, Manipur, India were reported to show anticandidal and biocontrol activities against rice fungal pathogens (Curvularia oryzae, F. oxysporum, Helminthosporum oryzae, Pyricularia oryzae, R. pryzae-sativae, and R. solani) as well as antibacterial activity (Nimaichand et al., 2015). However, Amycolatopsis, Rhodococcus, and Pseudonocardia isolates showed only biocontrol activity against rice fungal pathogen. Recently, five Streptomyces spp. from Chaabe cave in Algeria was reported to produce non-polyenic antifungal substances active against C. albicans (Belyagoubi et al., 2018).
Conclusion and Future Perspectives
Emerging and re-emerging infectious diseases are threatening human society at an alarming rate. It is a call of emergency to find an effective cure for these pathogens. Actinobacteria are proving again to be prolific producers of promising bioactive compounds with widely application. Cave and karst environments are underexplored microbiologically and should not be overlooked for the search and discovery of novel actinobacteria and their chemical diversity of useful compounds. It is evident from this mini-review that cave environments harbor novel and diverse actinobacteria (Table 1). These actinobacteria offer a rich source of bioactive compounds as exemplified in Table 2. We opine that in order to explore cave actinobacteria to their full potential, 2 major research area must be addressed. The first area of research should deal with the ability to isolate and cultivate actinobacteria of interest. It is well-accepted that most microorganisms could not be cultivated in laboratory. The isolation and cultivation of bioactive producing actinobacteria under laboratory conditions represent the first challenge. Currently, the isolation strategy specifically for cave actinobacteria is lacking. There is still an urgent need for an improved selective isolation to target specific actinobacterial taxa of interest and extended our ability to tap into the majority of these uncultivable bacteria. Modification of growth conditions and use of new culturing methods were proposed for cultivation of previously uncultivable microorganisms (Pham and Kim, 2012). A combination of enrichment techniques including heat-pretreatments of samples, adjusting media pH and calcium salts supplements were effectively applied to isolate rare actinobacteria from karstic caves (Fang et al., 2017b).
The advancement of next generation sequencing and accumulation of high quality whole genome data provide a powerful tool and useful information to support the search for novel bioactive metabolites for drug development. Currently, these genome data of actinobacteria reveal the presence of several biosynthetic gene clusters of secondary metabolites and reaffirm status of actinobacteria as prolific producers of bioactive compounds. However, these gene clusters are not normally expressed under laboratory conditions. Many secondary metabolites encoded by these gene clusters remain unidentified in fermentation broth (Scherlach and Hertweck, 2009; Ren et al., 2017). Therefore, the second challenge lies in our ability to activate these silent gene clusters. Recently, specific biological and chemical stimuli namely exposure to antibiotics, metals and mixed microbial culture, were successfully employed to activate secondary metabolites production in cave actinobacteria (Covington et al., 2018). Evidently, the study on cave actinobacteria and their bioactive compounds is still at an early stage. There still remains room for further study to guarantee cave actinobacteria as producers of new bioactive compounds for the benefit of human well-being.
Statements
Author contributions
PR contributed data for selective isolation, novel taxa, bioactive metabolites and Tables 1, 2. WP conceived the idea, wrote, and revised the whole manuscript.
Acknowledgments
This research work was supported by Center of Excellence on Biodiversity (BDC), Office of Higher Education Commission (Project BDC-PG3-161005) and partially supported by Chiang Mai University. PR is grateful to Graduate School, Chiang Mai University for TA/RA scholarship for the academic year 2018–2019.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1
AbdelmohsenU. R.BayarK.HentschelU. (2014). Diversity, abundance and natural products of marine sponge-associatd actinomycetes. Nat. Prod. Rep.31, 381–399. 10.1039/C3NP70111E
2
AdamD.MaciejewskaM.Naômé, A.MartinetL.CoppietersW.KarimL.et al. (2018). Isolation, characterization, and antibacterial activity of hard-to-culture actinobacteria from cave moonmilk deposits. Antibiotics7:28. 10.3390/antibiotics7020028
3
Axenov-GibanovD.VoytsekhovskayaI. V.TokovenkoB. T.ProtasovE. S.GamaiunovS. V.RabetsY. V.et al. (2016). Actinobacteria isolated from an underground lake moonmilk speleothem from the biggest conglomeratic karstic cave in Siberia as sources of novel biologically active compounds. PLoS ONE11:e0149216. 10.1371/journal.pone.0149216
4
BarkaE. A.VatsaP.SanchezL.Gaveau-VaillantN.JacquardC.KlenkH.-P.et al. (2016). Taxonomy, physiology, and natural products of actinobacteria. Microbiol. Mol. Biol. Rev.80, 1–43. 10.1128/MMBR.00019-15
5
BartonA. H.TaylorM. R.PaceN. R. (2004). Molecular phylogenetic analysis of a bacterial community in an oligotrophic cave environment. Geomicrobiol. J.21, 11–20. 10.1080/01490450490253428
6
BartonH. A. (2006). Introduction to cave microbiology: a review for the non-specialist. J. Cave Karst Stud.68, 43–54.
7
BelyagoubiL.Belyagoubi-BenhammouN.JuradaV.DupontJ.LacostS.DjebbahF.et al. (2018). Antimicrobial activities of culturable microorganisms (actinomycetes and fungi) isolated from Chaabe cave, Algeria. Int. J. Speleol.47, 189–199. 10.5038/1827-806X.47.2.2148
8
BulinaT. I.AlferovaI. V.TerkhovaL. P. (1997). A novel approach to isolation of actinomycetes involving irradiation of soil samples with microwaves. Microbiology66, 231–234.
9
CaoC. L.ZhouX. Q.QinS.TaoF. X.JiangJ. H.LianB. (2015). Lentzea guizhouensis sp. nov., a novel lithophilous actinobacterium isolated from limestone from the karst area, Guizhou, China. Antonie van Leeuwenhoek108, 1365–1372. 10.1007/s10482-015-0589-x
10
CastroJ. F.RazmilicV.Gomez-EscribanoJ. P.AndrewsB.AsenjoJ.BibbM. (2018). The ‘gifted’ actinomycetes Streptomyces leeuwenhoekii. Antonie van Leeuwenhoek111, 1433–1488. 10.1007/s10482-018-1034-8
11
CheepthamN.SadowayT.RuleD.WatsonK.MooteP.SolimanL. C.et al. (2013). Cure from the cave: volcanic cave actinomycetes and their potential in drug discovery. Int. J. Speleol.42, 35–47. 10.5038/1827-806X.42.1.5
12
ChengJ.ChenW.ZhangB. H.NimaichandS.ZhouE. M.LuX. H.et al. (2013a). Microlunatus cavernae sp. nov., a novel actinobacterium isolated from Alu ancient cave, Yunnan, south-west China. Antonie van Leeuwenhoek104, 95–101. 10.1007/s10482-013-9929-x
13
ChengJ.ZhangY. G.ChenW.LiL.ZhangD. F.WangH. F.et al. (2013b). Saccharopolyspora cavernae sp. nov., a novel actinomycete isolated from the Swallow cave in Yunnan, south-west China. Antonie van Leeuwenhoek104, 837–843. 10.1007/s10482-013-9996-z
14
CovingtonB. C.SpragginsJ. M.Ynigez-GutierrezA. E.HyltonZ. B.BachmannB. O. (2018). Response of secondary metabolism of hypogean actinobacterial genera to chemical and biological stimuli. Appl. Environ. Microbiol.84, e01125–e01118. 10.1128/AEM.01125-18
15
DerewaczD. K.McNeesC. R.ScalmaniG.CovingtonC. L.ShanmugamG.MarnettL. J.et al. (2014). Structure and stereochemical determination of hypogeamicins from a cave-derived actinomycete. J. Nat. Prod.77, 1759–1763. 10.1021/np400742p
16
DuangmalK.MingmaR.Pathom-areeW.NiyomvongN.InahashiY.MatsumotoA.et al. (2012). Microbispora thailandensis sp. nov., an actinomycete isolate from cave soil. J. Antibiot.65, 491–494. 10.1038/ja.2012.57
17
FangB. Z.HanM. X.LiuL.ZhangZ. T.LiuW. L.ShenJ. T.et al. (2017a). Lentzea covernae sp. nov., an actinobacterium isolated from a karst cave sample, and emended description of the genus Lentzea. Int. J. Syst. Evol. Microbiol.67, 2357–2362. 10.1099/ijsem.0.002364
18
FangB. Z.NimaichandS.HanM. X.JiaoJ. Y.ChengJ.WeiD. Q.et al. (2017b). Insights on the effects of heat pretreatment, pH, and calcium salts on isolation of rare Actinobacteria from karstic caves. Front. Microbiol.8:1535. 10.3389/fmicb.2017.01535
19
GenilloudO. (2017). Actinomycetes: still a source of novel antibiotics. Nat. Prod. Rep.34, 1203–1232. 10.1039/C7NP00026J
20
GhoshS.KuisieneN.CheepthamN. (2017). The cave microbiome as a source for drug discovery: reality or pipe dream?Biochem. Pharmacol.134, 18–34. 10.1016/j.bcp.2016.11.018
21
GrothI.Saiz-JimenezC. (1999). Actinomycetes in hypogean environments. Geomicrobiol. J.16, 1–8. 10.1080/014904599270703
22
GrothI.SchumannP.SchuetzeB.AugstenK.KramerI.StackebrandtE. (1999). Beutenbergia cavernae gen. nov., sp. nov., an L-lysine-containing actinomycete isolated from a cave. Int. J. Syst. Bacteriol.49, 1733–1740. 10.1099/00207713-49-4-1733
23
GrothI.SchumannP.SchutzeB.AugstenK.StackebrandtE. (2002). Knoellia sinensis gen. nov., sp. nov. and Knoellia subterranea sp. nov., two novel actinocacteria isolated from a cave. Int. J. Syst. Evol. Microbiol.52, 77–84. 10.1099/00207713-52-1-77
24
HanM. X.FangB. Z.TianY.ZhangW. Q.JiaoJ. Y.LiuL.et al. (2017). Nocardioides cavernae sp. nov., an actinobacterium isolated from a karst cave. Int. J. Syst. Evol. Microbiol.67, 633–639. 10.1099/ijsem.0.001676
25
HeroldK.GollmickF. A.GrothI.RothM.MenzelK. D.MollmannU.et al. (2005). Cervimycin A-D: a polyketide glycoside complex from a cave bacterium can defeat vancomycin resistance. Chem. Eur. J.11, 5523–5530. 10.1002/chem.200500320
26
HuangJ. R.MingH.LiS.ZhaoZ. L.MengX. L.ZhangJ. X.et al. (2016). Amycolatopsis xuchangensis sp. nov. and Amycolatopsis jiguanensis sp. nov., isolated from soil. Antonie van Leeuwenhoek109, 1423–1431. 10.1007/s10482-016-0742-1
27
HugJ. J.BaderC. D.RemskarM.CirnskiK.MullerR. (2018). Concepts and methods to access novel antibiotics from actinomycetes. Antibiotics7:44. 10.3390/antibiotics7020044
28
InokoshiJ.TakagiY.UchidaR.MasumaR.OmureS.TomodaH. (2010). Production of a new type of amidepsine with a sugar moiety by static fermentation of Humicola sp. FO-2942. J. Antibiot.63, 9–16. 10.1038/ja.2009.110
29
JiangL.PuH.XiangJ.SuM.YanX.YangD.et al. (2018). Huanglongmycin A-C, cytotoxic polyketides biosynthesized by a putative type II polyketide synthase from Streptomyces sp. CB09001. Front. Chem.6:254. 10.3389/fchem.2018.00254
30
JiangY.LiQChenX.JiangC. (2016). Isolation and cultivation methods of Actinobacteria in Actinobacteria- Basics and Biotechnological Applications, ed. DhanasekaranD. (Rijeka: InTech), 39–57.
31
JiangZ. K.GuoL.ChenC.LiuS. W.ZhangL.DaiS. J.et al. (2015). Xiakemycin A, a novel pyranonaphthoquinone antibiotic, produced by the Streptomyces sp. CC8-201 from the soil of a karst cave. J. Antibiot.68, 771–774. 10.1038/ja.2015.70
32
JuradoV.KroppenstedtR. M.JimenezC. S.KlenkH.-P.MounieéD.LaizL.et al. (2009). Hoyosella altamirensis gen. nov., sp. nov., a new member of the order Actinomycetales isolated from a cave bioflim. Int. J. Syst. Evol. Microbiol.59, 3105–3110. 10.1099/ijs.0.008664-0
33
JuradoV.BoironP.KroppenstedtR. M.LaurentF.CoubleA.LaizL.et al. (2008). Nocardia altamirensis sp. nov., isolated from Altamira cave, Cantabria, Spain. Int. J. Syst. Evol. Microbiol.58, 2210–2214. 10.1099/ijs.0.65482-0
34
JuradoV.GrothI.GonzalezJ. M.LaizL.JimenezC. S. (2005). Agromyces subbeticus sp. nov., isolated from a cave in southern Spain. Int. J. Syst. Evol. Microbiol.55, 1897–1901. 10.1099/ijs.0.63637-0
35
KayS.Pathom-areeW.CheepthamN. (2013). Screening of volcanic cave actinomycetes for antimicrobial activity against Paenibacillus larvae, a honey bee pathogen. Chiang Mai J. Sci.40. 26–33.
36
KemungH. M.TanL. T. H.KhanT. M.ChanK. G.PusparajahP.GohB. H.et al. (2018). Streptomyces as a prominent resource of future anti-MRSA drugs. Front. Microbiol.9:2221. 10.3389/fmicb.2018.02221
37
KimB. S.LeeJ. Y.HwangB. K. (1998). Diversity of actinomycetes antagonistic to plant pathogenic fungi in cave and sea-mud soils of Korea. J. Microbiol.36, 86–92.
38
KoD. H.LeeS. D. (2017). Augustibacter spluncae sp. nov., isolated from a lava cave stalactite. Int. J. Syst. Evol. Microbiol.67, 3283–3288. 10.1099/ijsem.0.002108
39
KoK. S.KimY.SeongC. N.LeeS. D. (2015). Rhodococcus antrifimi sp. nov., isolated from dried bat dung of a cave. Int. J. Syst. Evol. Microbiol.65, 4043–4048. 10.1099/ijsem.0.000534
40
LaizL.Gonzalez-DelvalleM.HermosinB.Ortiz-MartinezA.Saiz-JimenezC. (2003). Isolation of cave bacteria and substrate utilization at different temperature. Geomicrobiol. J.20, 479–489. 10.1080/713851125
41
LaizL.GrothI.GonzalezI.Saiz-JimenezC. (1999). Microbiological study of the dripping waters in Altamira cave (Santillana del mar, Spain). J. Microbiol. Methods36, 129–138. 10.1016/S0167-7012(99)00018-4
42
LeeS. D. (2006a). Actinocorallia cavernae sp. nov., isolated from a natural cave in Jeju, Korea. Int. J. Syst. Evol. Microbiol.56, 1085–1088. 10.1099/ijs.0.63895-0
43
LeeS. D. (2006b). Amycolatopsis jejuensis sp. nov. and Amycolatopsis halotolerans sp. nov., novel actinomycetes isolated from a natural cave. Int. J. Syst. Evol. Microbiol.56, 549–553. 10.1099/ijs.0.63881-0
44
LeeS. D. (2006c). Nocardia jejuensis sp. nov., novel actinomycetes isolated from a natural cave on Jeju island, Republic of Korea. Int. J. Syst. Evol. Microbiol.56, 559–562. 10.1099/ijs.0.63866-0
45
LeeS. D. (2008). Jiangella alkaliphila sp. nov., an actinobacterium isolated from a cave. Int. J. Syst. Evol. Microbiol.58, 1176–1179. 10.1099/ijs.0.65479-0
46
LeeS. D. (2013a). Humibacter antri sp. nov., an actinobactrium isolated from a natural cave, and emended description of the genus Humibacter. Int. J. Syst. Evol. Microbiol.63, 4315–4319. 10.1099/ijs.0.050708-0
47
LeeS. D. (2013b). Spelaeicoccus albus gen. nov., sp. nov., an actinobacterium isolated from a natural cave. Int. J. Syst. Evol. Microbiol.63, 3958–3963. 10.1099/ijs.0.050732-0
48
LeeS. D. (2015). Antricoccus suffuscus gen. nov., sp. nov., isolated from a natural cave. Int. J. Syst. Evol. Microbiol.65, 4410–4416. 10.1099/ijsem.0.000588
49
LeeS. D. (2017). Conexibacter stalactiti sp. nov., isolated from stalactites in a lava cave and emended description of the genus Conexibacter. Int. J. Syst. Evol. Microbiol.67, 3214–3218. 10.1099/ijsem.0.002083
50
LeeS. D.KangS. O.HahY. C. (2000a). Catellatospora koreensis sp. nov., a novel actinomycete isolated from a gold-mine cave. Int. J. Syst. Evol. Microbiol.50, 1103–1111. 10.1099/00207713-50-3-1103
51
LeeS. D.KimE. S.RoeJ. H.KimJ. H.KangS. O.HahY. C. (2000b). Saccharothrix violacea sp. nov., isolated from a gold mine cave, and Saccharothrix albidocapillata comb. nov. Int. J. Syst. Evol. Microbiol.50, 1315–1323. 10.1099/00207713-50-3-1315
52
LiQ. Q.HanM. X.FangB. Z.JiaoJ. Y.LiuL.YangZ. W.et al. (2017). Nocardia cavernae sp. nov., a novel actinobacterium isolated from a karst cave sample. Int. J. Syst. Evol. Microbiol.67, 2998–3003. 10.1099/ijsem.0.002072
53
LiW. J.NimaichandS.JiangZ.LiuM. J.KhieuT. N.KimC. J.et al. (2014). Streptomyces canchipurensis sp. nov., isolated from a limestone habitat. Antonie van Leeuwenhoek106, 1119–1126. 10.1007/s10482-014-0281-6
54
LiX. J.DaiS. J.LiuS. W.LiuJ. M.ChenL.HuL.et al. (2016). Tenggerimyces flavus sp. nov., isolated from soil in a karst cave, and emended description of the genus Tenggerimyces. Int. J. Syst. Evol. Microbiol.66, 1499–1505. 10.1099/ijsem.0.000908
55
MaciejewskaM.AdamD.MartinetL.NaomeA.CalusiriskaM.DelfosseP.et al. (2016). A phenotypic and genotypic analysis of the antimicrobial potential of cultivable Streptomyces isolated from cave moonmilk deposits. Front. Microbiol.7:1455. 10.3389/fmicb.2016.01455
56
MaciejewskaM.PessiI. S.AriasA. A.NoirfaliseP.LuisG.OngenaM.et al. (2015). Streptomyces lunaelactis sp. nov., a novel ferroverdin A-producing Streptomyces species isolated from a moonmilk speleothem. Antonie van Leeuwenhoek107, 519–531. 10.1007/s10482-014-0348-4
57
MargesinR.SchumannP.SpröerC.GounotA. M. (2004). Arthrobacter psychrophenolicus sp. nov., isolated from an alpine ice cave. Int. J. Syst. Evol. Microbiol.54, 2067–2072. 10.1099/ijs.0.63124-0
58
MingmaR.DuangmalK.TakéInahashiY.ŌmuraS.TakahashiY.MatsumotoA. (2016). Proposal of Sphaerimonospora cavernae gen. nov., sp. nov. and transfer of Microbispora mesophila (Zhang et al., 1984). Sphaerimonospora mesophila comb. nov. and Microbispora thailandensis (Duangmal et al., 2012) to Sphaerimonospora thailandensis comb. nov. Int. J. Syst. Evol. Microbiol.66, 1735–1744. 10.1099/ijsem.0.000935
59
MoirA.SmithD. A. (1990). The genetics of bacterial spore germination. Annu. Rev. Microbiol.44, 531–553. 10.1146/annurev.mi.44.100190.002531
60
NakaewN.Pathom-areeW.LumyongS. (2009a). First record of the isolation, identification and biological activity of a new strain of Spirillospora albida from Thai cave soils. Actinomycetologica23, 1–7. 10.3209/saj.SAJ230102
61
NakaewN.Pathom-areeW.LumyongS. (2009b). Generic diversity of rare actinomycetes from Thai cave soils and their possible use as new bioactive compounds. Actinomycetologica23, 21–26. 10.3209/saj.SAJ230201
62
NakaewN.SungthongR.YokotaA.LumyongS. (2012). Nonomuraea monospora sp. nov., an actinomycete isolated from cave soil in Thailand, and emended description of the genus Nonomuraea. Int. J. Syst. Evol. Microbiol.62, 3007–3012. 10.1099/ijs.0.035220-0
63
NimaichandS.DeviA. M.TamreihaoK.NingthoujamD. S.LiW.-J. (2015). Actinobacterial diversity in limestone deposit sites in Hundung, Manipur (India) and their antimicrobial activities. Front. Microbiol6:413. 10.3389/fmicb.2015.00413
64
NimaichandS.SanasamS.ZhengL.-Q.ZhuW.-Y.YangL.-L.TangS.-K.et al. (2013a). Rhodococcus canchipurensis sp. nov., an actinomycete isolated from a limestone deposit site. Int. J. Syst. Evol. Microbiol.63, 114–118. 10.1099/ijs.0.036087-0
65
NimaichandS.TamrihaoK.YangL. L.ZhuW. Y.ZhangY. G.LiL.et al. (2013b). Streptomyces hundungensis sp. nov., a novel actinomycete with antifungal activity and plant growth promoting traits. J. Antibiot.66, 205–209. 10.1038/ja.2012.119
66
NimaichandS.ZhangY.-G.ChengJ.LiL.ZhangD.-F.ZhouE.-M.et al. (2013c). Micromonospora kangleipakensis sp. nov., isolated from a sample of limestone quarry. Int. J. Syst. Evol. Microbiol.63, 4546–4551. 10.1099/ijs.0.052746-0
67
NimaichandS.ZhuW. Y.YangL. L.MingH.NieG. X.TangS. K.et al. (2012). Streptomyces manipurensis sp. nov., a novel actinomycete isolated from a limestone deposit site in Manipur, India. Antonie van Leeuwenhoek102, 133–139. 10.1007/s10482-012-9720-4
68
NiyomvongN.Pathom-areeW.ThamchaipenetA.DuangmalK. (2012). Actinomycetes from tropical limestone caves. Chiang Mai J. Sci.39, 373–388.
69
NorthupD. E.LavoieK. H. (2001). Geomicrobiology of caves: a review. Geomicrobiol. J.18, 199–222. 10.1080/01490450152467750
70
PassariA. K.MishaV. K.GuptaV. K.SinghB. P. (2018). Chapter 1: methods used for the recovery of culturable endophytic actinobacteria: an overview, in Actinobacteria: Diversity and Biotechnological Applications: New and Future Developments in Microbial Biotechnology and Bioengineering, eds SinghA. K.PassariB. P.GuptaV. K. (Amsterdam: Elsevier), 1–11.
71
PatelK.LailleR.MartinM. T.TilviS.MoriouC.GallardJ. F.et al. (2010). Unprecedented stylissazoles A-C from Stylissa carteri: another dimension for marine pyrrole-2-aminoimadazole metabolite diversity. Angew. Chem. Int. Ed.49, 4775–4779. 10.1002/anie.201000444
72
PhamV. H.KimJ. (2012). Cultivation of unculturable soil bacteria. Trends Biotechnol.30, 475–484. 10.1016/j.tibtech.2012.05.007
73
QuadriS. R.TianX. P.ZhangJ.LiJ.NieG. X.TangS. K.et al. (2015). Nonomuraea indica sp. nov., novel actinomycetes isolated from lime-stone open pit mine, India. J. Antibiot.68, 491–495. 10.1038/ja.2015.24
74
RenH.WangB.ZhaoH. (2017). Breaking the silence: New strategies for discovering novel natural products. Curr. Opin. Biotechnol.48, 21–27. 10.1016/j.copbio.2017.02.008
75
RiquelmeC.DapkeviciusM. L. E.MillerA. Z.Charlop-PowersZ.BradyS.MasonC.et al. (2017). Biotechnological potential of actinobacteria from Canadian and Azorean volcanic caves. Appl. Microbiol. Biotechnol.101, 843–857. 10.1007/s00253-016-7932-7
76
Schabereiter-GurtnerC.Saiz-Jimenez„CPinarG.LubitzW.RöllkeS. (2002). Altamira cave paleolithic paintings harbor partly unknown bacterial communities. FEMS Microbiol. Lett.211, 7–11. 10.1111/j.1574-6968.2002.tb11195.x
77
ScherlachK.HertweckC. (2009). Triggering cryptic natural product biosynthesis in microorganisms. Org. Biomol. Chem.7, 1753–1760. 10.1039/b821578b
78
SeoJ. P.YunY. W.LeeS. D. (2007). Nocardia speluncae sp. nov., isolated from a cave. Int. J. Syst. Evol. Microbiol.57, 2932–2935. 10.1099/ijs.0.65085-0
79
StankovicN.RadulovicV.PetkovicM.VuckovicI.JadraninM.VasiljevicB.et al. (2012). Streptomyces sp. JS520 produces exceptionally high quantities of undecylprodigiosin with antibacterial, antioxidative, and UV-protective properties. Appl. Microbiol. Biotechnol.96, 1217–1231. 10.1007/s00253-012-4237-3
80
TakahashiY.NakashimaT. (2018). Actinomycetes, an inexhaustible source of naturally occurring antibiotics. Antibiotics7:45. 10.3390/antibiotics7020045
81
TakéA.InahashiY.ŌmuraS.TakahashiY.MatsumotoA. (2018). Streptomyces boninensis sp. nov., isolated from soil from a limestone cave in the Ogasawara islands. Int. J. Syst. Evol. Microbiol.68, 1795–1799. 10.1099/ijsem.0.002753
82
TawariK.GuptaR. K. (2013). Diversity and isolation of rare actinomyctes: an overview. Crit. Rev. Microbiol.39, 256–294. 10.3109/1040841X.2012.709819
83
Tomczyk-ZakK.ZielenkiewiczU. (2016). Microbial diversity in caves. Geomicrobiol. J.33, 20–38. 10.1080/01490451.2014.1003341
84
TomodaH.OmuraS. (2001). Screening for inhibitors of lipid metabolism, in Enzyme Technologies for Pharmaceutical and Biotechnological Applications, eds. KirstH.A.YehW.K.ZmijewskiM.J.Jr. (New York, NY: Marcel Dekker Inc), 343–378.
85
TuoL.LiuS. W.LiuJ. M.ZhangY. Q.JiangZ. K.LiuX. F.et al. (2015). Lysinibacter cavernae gen. nov., sp. nov., a new member of the family Microbacteriaceae isolated from a karst cave. Int. J. Syst. Evol. Microbiol.65, 3305–3312. 10.1099/ijsem.0.000415
86
VelikonjaB. H.TkaveR.PasicL. (2014). Diversity of cultivable bacteria involved in the formation of macroscopic microscopic microbial colonies (cave silver) on the walls of a cave in Slovenia. Int. J. Speleol.43, 45–46. 10.5038/1827-806X.43.1.5
87
WangD. S.XueQ. H.ZhuW. J.ZhaoJ.DuanJ. L.ShenG. H. (2013). Microwave irradiation is a useful tool for improving isolation of actinomycetes from soil. Microbiology82, 102–110. 10.1134/S0026261712060161
88
WangZ. G.WangY. X.LiuJ. H.ChenY. G.ZhangX. X.WenM. L.et al. (2009). Fodinibacter luteus gen. nov., sp. nov., an actinobactrium isolated from a salt mine. Int. J. Syst. Evol. Microbiol.59, 2185–2190. 10.1099/ijs.0.006882-0
89
YücelS.YamacM. (2010). Selection of Streptomyces isolates from Turkish karstic caves against antibiotic resistant microorganisms. Pak. J. Pharm. Sci.23, 1–6.
90
ZhangL. Y.MingH.MengX. L.FangB. Z.JiaoJ. Y.NimaichandS.et al. (2018). Ornithinimicrobium cavernae sp. nov., an actinobactrium isolated from a karst cave. Antonie van Leeuwenhoek.112, 179–186. 10.1007/s10482-018-1141-6
91
ZhangW. Q.LiY. Q.LiuL.NimaichandS.FangB. Z.WeiD. Q.et al. (2016). Stackebrandtia cavernae sp. nov., a novel actinobacterium isolated from a karst cave sample. Int. J. Syst. Evol. Microbiol.66, 1206–1211. 10.1099/ijsem.0.000859
Summary
Keywords
actinobacteria, cave, karst, bioactive compounds, diversity, antimicrobial, anticancer, natural products
Citation
Rangseekaew P and Pathom-aree W (2019) Cave Actinobacteria as Producers of Bioactive Metabolites. Front. Microbiol. 10:387. doi: 10.3389/fmicb.2019.00387
Received
15 November 2018
Accepted
13 February 2019
Published
22 March 2019
Volume
10 - 2019
Edited by
Learn-Han Lee, Monash University Malaysia, Malaysia
Reviewed by
Sheng Qin, Jiangsu Normal University, China; Joachim Wink, Helmholtz Center for Infection Research, Helmholtz Association of German Research Centers (HZ), Germany
Updates
Copyright
© 2019 Rangseekaew and Pathom-aree.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Wasu Pathom-aree wasu.p@cmu.ac.th
This article was submitted to Antimicrobials, Resistance and Chemotherapy, a section of the journal Frontiers in Microbiology
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.