 Jill N. Sutton1*
Jill N. Sutton1* Luc André2Damien Cardinal3
Luc André2Damien Cardinal3 Daniel J. Conley4,5
Daniel J. Conley4,5 Gregory F. de Souza6Jonathan Dean7,8
Gregory F. de Souza6Jonathan Dean7,8 Justin Dodd9
Justin Dodd9 Claudia Ehlert10
Claudia Ehlert10 Michael J. Ellwood11
Michael J. Ellwood11 Patrick J. Frings12,13
Patrick J. Frings12,13 Patricia Grasse14Katharine Hendry15Melanie J. Leng8,16Panagiotis Michalopoulos17
Patricia Grasse14Katharine Hendry15Melanie J. Leng8,16Panagiotis Michalopoulos17 Virginia N. Panizzo16,18
Virginia N. Panizzo16,18 George E. A. Swann16,18
George E. A. Swann16,18- 1Université de Brest - UMR 6539 CNRS/UBO/IRD/Ifremer, LEMAR - IUEM, Plouzané, France
- 2Earth Sciences Department, Royal Museum for Central Africa, Tervuren, Belgium
- 3LOCEAN Laboratory, Sorbonne Universités (UPMC, Univ Paris 06), Centre National de la Recherche Scientifique, IRD, MNHN, Paris, France
- 4Department of Geology, Lund University, Lund, Sweden
- 5Stellenbosch Institute for Advanced Study, Stellenbosch, South Africa
- 6Institute of Geochemistry and Petrology, ETH Zurich, Zurich, Switzerland
- 7School of Environmental Sciences, University of Hull, Hull, United Kingdom
- 8NERC Isotope Geosciences Facilities, British Geological Survey, Nottingham, United Kingdom
- 9Geology and Environmental Geosciences, Northern Illinois University, DeKalb, IL, United States
- 10Max Planck Research Group for Marine Isotope Geochemistry, Institute for Chemistry and Biology of the Marine Environment, University of Oldenburg, Oldenburg, Germany
- 11Research School of Earth and Ocean Sciences, Australian National University, Canberra, ACT, Australia
- 12Earth Surface Geochemistry, Helmholtz Centre Potsdam GFZ German Research Centre for Geosciences, Potsdam, Germany
- 13Department of Geoscience, Swedish Museum of Natural History, Stockholm, Sweden
- 14Ocean Circulation and Climate Dynamics, GEOMAR Helmholtz Centre for Ocean Research Kiel, Kiel, Germany
- 15School of Earth Sciences, University of Bristol, Bristol, United Kingdom
- 16Centre for Environmental Geochemistry, University of Nottingham, Nottingham, United Kingdom
- 17Hellenic Centre for Marine Research, Institute of Oceanography, Anavyssos, Greece
- 18School of Geography, University of Nottingham, Nottingham, United Kingdom
Silicon (Si) is the second most abundant element in the Earth's crust and is an important nutrient in the ocean. The global Si cycle plays a critical role in regulating primary productivity and carbon cycling on the continents and in the oceans. Development of the analytical tools used to study the sources, sinks, and fluxes of the global Si cycle (e.g., elemental and stable isotope ratio data for Ge, Si, Zn, etc.) have recently led to major advances in our understanding of the mechanisms and processes that constrain the cycling of Si in the modern environment and in the past. Here, we provide background on the geochemical tools that are available for studying the Si cycle and highlight our current understanding of the marine, freshwater and terrestrial systems. We place emphasis on the geochemistry (e.g., Al/Si, Ge/Si, Zn/Si, δ13C, δ15N, δ18O, δ30Si) of dissolved and biogenic Si, present case studies, such as the Silicic Acid Leakage Hypothesis, and discuss challenges associated with the development of these environmental proxies for the global Si cycle. We also discuss how each system within the global Si cycle might change over time (i.e., sources, sinks, and processes) and the potential technical and conceptual limitations that need to be considered for future studies.
Introduction
The global silicon (Si) cycle is of great interest due to the role that silicate weathering has played in maintaining climatic stability on geological time scales (Siever, 1991; Frings et al., 2016; Conley et al., 2017) and because Si is an important nutrient for many organisms in marine and freshwater ecosystems. It occurs as silicate minerals in association with all rock types (igneous, metamorphic, sedimentary). Weathering, biological and geochemical transformations, transport and interactions with other elements (in particular nutrients and carbon) form the basis of the global biogeochemical Si cycle (Frings et al., 2016). The global Si cycle has evolved through geologic time with overall declines in oceanic dissolved Si due to the uptake and deposition by organisms (Siever, 1991; Conley et al., 2017). While the modern Si biogeochemical cycle is influenced by anthropogenic forcing (Laruelle et al., 2009), the processes of weathering and burial as biogenic silica (bSiO2) are the dominant processes in the biogeochemical Si cycle.
The paper aims to review the use of Si stable isotopes and associated trace elements in order to address the following questions:
(1) What is the bio-geochemistry of biogenic silica?
(2) What is the current state of knowledge regarding the influence of biogenic silica on the global Si cycle?
(3) What is the influence of early to late sediment diagenesis on biogenic silica, and thus its utility as an environmental proxy for palaeoceanographic interpretation?
Although modeling is not being covered in the review, its importance cannot be underscored as it has been increasingly used to understand the mechanisms controlling Si isotope fractionation (e.g., Qin et al., 2016) and the development of the Si isotope composition as a tracer of biogeochemical silicon cycle in modern and past natural systems (e.g., Gao et al., 2016).
Silicon Reservoirs
The chemical weathering of silicate minerals and the eventual cycling of weathered products (clays, dissolved Si) provide the starting point of Si bio-geochemistry and its interaction with other elemental cycles such as carbon. Silicate weathering represents an important sink of atmospheric CO2 over geological time scales (Berner et al., 1983; Wollast and Mackenzie, 1989; Brady and Carroll, 1994) and depends on temperature and precipitation. Thus, the rate at which weathering occurs will be enhanced through changes in global temperature (White and Blum, 1995).
Silicon rarely occurs as the pure element in nature and appears most often in combination with oxygen to form solid silicate minerals or amorphous compounds (including biogenic Si), or in aqueous solutions it occurs as orthosilicic acid (Si(OH)4) (Iler, 1979). Silicon exists in major pools in dissolved and solid forms in all reservoirs: extra-terrestrial, continental (e.g., soil, vegetation, hydrothermal), freshwater (e.g., rivers, lakes, groundwater, organisms, sediment), atmospheric (e.g., aerosols), and oceanic (e.g., water column, organisms, sediment and pore-waters, oceanic crust, hydrothermal), (Figure 1). The dominant transformation processes are weathering of silicate rocks and the formation of secondary minerals and release of dissolved Si, uptake of dissolved Si by organisms for the biomineralization of biogenic Si, and the remineralization of Si. Coastal regions are of special importance here because they support a large fraction of the global primary production and control the transfer of dissolved and particulate nutrients from land to the open ocean (Conley et al., 1993; Rabouille et al., 2001).
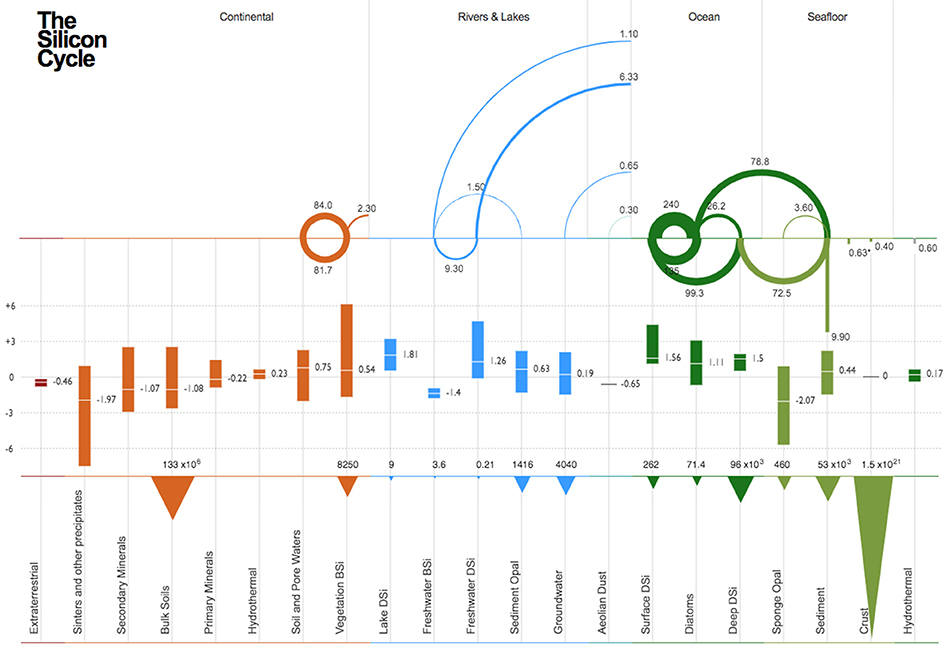
Figure 1. Schematic of the modern day global Si cycle grouped by reservoir and their associated δ30Si values (in ‰). The figure is divided into three parts. The upper part indicating the magnitude of the fluxes (in 1012mol yr−1; Tmol yr−1) between each reservoir, the middle part showing their associated δ30Si values, and the lower part showing reservoir size. The fluxes are numerically and figuratively represented, moving in a clockwise manner (e.g., In the Ocean compartment, there is a flux of 240 Tmol yr−1 from Surface DSi to Diatoms, after the Diatoms dissolve, there is 135 Tmol yr−1 flux back from the Diatoms to the Surface DSi in the upper water column). In the case where the flux points downwards (e.g., 9.90 Tmol yr−1 within the sediment reservoir), the flux is effectively removed from the system. * Value assumed to underestimate flux (see section Diagenetic Silica Precipitation and Reverse Weathering). Estimates for the Si fluxes were obtained from the following sources: Carey and Fulweiler (2012), Frings et al. (2016), Laruelle et al. (2009), Tréguer and De La Rocha (2013). Estimates for the range of δ30Si values were obtained from the following sources: Ehlert et al. (2016); Figure 2 from Frings et al. (2016) and references therein; Varela et al. (2016) and Panizzo et al. (2017). All data used to prepare this figure are available in Table S1.
Biomineralization, the Structure of Biogenic Silica, and Its Relevance to the Global Si Cycle
Many organisms use dissolved Si, including diatoms, silicoflagellates, radiolarians, sponges, and higher plants, to produce solid bSiO2 structures. Biogenic silica, also called biogenic opal, is the second most abundant mineral type formed by organisms after calcium carbonate (Brümmer, 2003). Biogenic silica is amorphous and its density, hardness, solubility, viscosity and composition may vary considerably (Perry et al., 2003). The silica structures are based upon a random network of SiO4 tetrahedral units connected through covalently linked Si-O-Si bonds of variable bond angle and bond lengths (Mann and Perry, 1986). All silicifying organisms utilize Si(OH)4; however, each silicifying organism has a unique biomineralizing pathway.
Details on the process of silicification can be found in the literature for diatoms (Hildebrand et al., 1997; Martin-Jezequel et al., 2000; Claquin et al., 2002; Hildebrand, 2008; Thamatrakoln and Hildebrand, 2008; Brunner et al., 2009; Thamatrakoln and Kustka, 2009), sponges (Wilkinson and Garrone, 1980; Reincke and Barthel, 1997; Cha et al., 1999; Maldonado et al., 1999, 2005; Brümmer, 2003; Uriz et al., 2003; Muller et al., 2007; Schröder et al., 2007), and higher plants (Takahashi et al., 1990; Ma et al., 2001, 2006).
Globally, diatoms (unicellular autotrophic algae) dominate the bSiO2 production with an estimate of 240 ± 40 Tmol yr−1 (Tréguer and De La Rocha, 2013) (Figure 1). The degree of bSiO2 production (i.e., silicification) of diatom frustules can vary significantly, even within the same species, and is primarily a function of ambient dissolved Si (DSi) concentration and cellular growth rate, whereby the bSiO2 content of a cell decreases during DSi limitation and/or accelerated growth (Martin-Jezequel et al., 2000; Claquin et al., 2002; Baines et al., 2010).
Until recently, the relevance of marine siliceous sponges on the Si cycle has been largely ignored and their role is currently being re-visited with a biogenic Si production estimated at 3.6 ± 3.7 Tmol yr−1 (Maldonado et al., 2005; Tréguer and De La Rocha, 2013) (Figure 1).
Higher plants produce roughly 84 (60–180) Tmol yr−1 of biogenic Si (phytoliths) (Figure 1), which is within a similar order of magnitude as diatom production (Conley, 2002). All plants that grow on soil contain Si in their tissues, whereby Si uptake and concentrations vary greatly among species depending on the different capacities for Si uptake by the roots (Takahashi et al., 1990; Ma et al., 2001). Active Si uptake has been suggested for Si-accumulating plants that are characterized by high Si concentrations (>1% dry weight in the leaves) and a molar Si:Ca ratio >1, whereas non-accumulating plants have a passive or discriminating Si uptake system (Takahashi et al., 1990; Ma et al., 2001). After uptake, DSi is rapidly transported through the plant to the different tissues within the transpiration stream. Due to transpirational water loss, DSi is gradually concentrated, and phytoliths are formed by precipitation in plant cells when the concentration increases above 2 mM. Thus, phytoliths vary in size and shape depending on the cell type, plant organ and plant species they are deposited in Madella et al. (2005).
Global Si Cycle Over Time
The global oceanic Si cycle has evolved through geologic time primarily due to the uptake of dissolved Si and subsequent biomineralization by organisms, especially sponges, radiolarians and diatoms (Siever, 1991). Substantial advances have occurred in our understanding of the evolution of the geological Si cycle through the use of the fossil record, silica isotope geochemistry and the phylogenomics of biosilicification (Conley et al., 2017; Hendry et al., 2018). The first biological impacts on the Si cycle are hypothesized to have occurred with the evolution of cyanobacteria in the Archean with further decreases in the mid-Proterozoic with evolution of eukaryotes capable of Si biomineralization. Decreases in oceanic DSi occurred prior to the start of the Phanerozoic with the evolution of widespread, large-scale skeletal bio-silicification. Due to the increased usage of dissolved Si by radiolarians in the lower to early Middle Ordovician, the loci of sponges shifted from shallow to deep-water basinal environments (Kidder and Tomescu, 2016). Finally, it is the appearance and subsequent proliferation of diatoms, with their superior ability to utilize low concentrations of DSi, that decreased DSi to the low levels observed in the global oceans today (Tréguer and De La Rocha, 2013). The impact of orogeny, e.g., the uplift of the Himalayas with periods of enhanced continental weathering fluxes in the Cenozoic (Misra and Froelich, 2012) could have increased DSi input to the oceans (Cermeño et al., 2015), although there is no evidence of large-scale changes in DSi concentrations in the geologic record (Fontorbe et al., 2016, 2017).
In the recent years, there have been a considerable number of studies on palaeo-environments of the Archean (4 – 2.5 Ga) and Proterozoic (2.5 – 0.5 Ga). Most use Si - O isotopes, elemental ratios (Ge/Si, REEs) or other isotopic systems (Fe, S…) associated to silica in Banded Iron Formations (e.g., André et al., 2006; Heck et al., 2011; Delvigne et al., 2012), cherts (e.g., Robert and Chaussidon, 2006; Chakrabarti et al., 2012) or paleosols (e.g., Delvigne et al., 2016). These studies all use the knowledge gained from modern, experimental and/or less ancient palaeo-records to interpret geochemical records, in order to reconstruct, for instance, the ocean temperature and oxygen levels, input of hydrothermal, and intensity of weathering. However, it is still challenging to provide a clear picture of these ancient environments because the studies have divergent results. A multi-proxy approach is therefore required. This is particularly needed to assess the level of preservation of the samples in order to take into account diagenesis while interpreting the geochemical signals into reconstruction of paleo-environments (Marin-Carbonne et al., 2014).
Modern day anthropogenic factors have a strong influence on the global Si cycle, for example through increases in CO2, temperature and changes in hydrological regimes and erosion, variations in agriculture and land use, eutrophication and changes in nutrient stoichiometry in coastal regions, and river damming (e.g., Rickert et al., 2002; Struyf et al., 2004, 2010; Laruelle et al., 2009; Clymans et al., 2011; Carey and Fulweiler, 2012). Anthropogenic perturbations of the global biogeochemical Si cycle are due to the gradual aggradation or depletion of the amorphous SiO2 pool held in continental soils (Barão et al., 2015; Vandevenne et al., 2015) and aquatic sediments (Frings et al., 2014b) in response to these changing environmental forcings (Struyf and Conley, 2012) and river damming (Conley et al., 1993). Our emerging understanding is that DSi inputs from the continents have potentially altered the magnitude and δ30Si composition of DSi supplied to the open ocean mostly because of changes occurring on the continent as well as changes of the silica sink on continental margins (Bernard et al., 2010; Frings et al., 2016). This includes short time scales such as anthropogenic impacts (not only on Si, but also on other nutrients e.g., N, P, Fe) and possibly to an extent large enough to impact whole-ocean isotopic signatures on the timescale of Quaternary glacial cycles. Thus, the sensitivity of biogeochemical Si cycling to anthropogenic pressure, especially in coastal regions, will likely be highlighted in the future (Laruelle et al., 2009; Bernard et al., 2010) and the continental Si cycle should not be neglected when interpreting pre-Quaternary long-term δ30SibSiO2 records from marine sediment records (Egan et al., 2013; Fontorbe et al., 2016).
Stable Silicon Isotope Ratios to Study the Global Si Cycle
Silicon has three naturally occurring stable isotope with the following mean abundances: 28Si: 92.23%, 29Si: 4.67%, and 30Si: 3.10%. The Si isotope composition is expressed in delta notation (δnSi) and can be calculated according to the following formula:
where n can represent 29Si or 30Si, x refers to either bSiO2 or DSi and standard is the atomic ratio of the heavy and light Si isotopes of the quartz standard (NBS28).
Several processes have been shown to fractionate Si isotopes, which explain why this tool can be used to trace the Si biogeochemical cycle. The extent of isotopic fractionation is defined by the isotopic fractionation factor (α) that can be expressed by Hoefs (2009):
-for kinetic isotopic exchange (irreversible reaction)
Where k refers to the rate constants for the reaction of light and heavy isotopes.
-for equilibrium
Where A and B are two chemical substances, typically the substrate (A) and the product (B) of the reaction.
For practical and analytical reasons, Si isotope ratios and Si isotope fractionation are reported in delta notation with the permil (‰) scale. A useful way to think about Si isotope values is as fractionation of Si isotopes, or the difference between the δ30Si values of DSi and bSiO2 as described by ε in Equation (2) and calculated (Criss, 1999) as:
where αbSiO2−DSi is the isotopic fractionation factor between the product (bSiO2) and the substrate (DSi). Note the lack of redox speciation and organic complexation for silicon limits fractionation effects to predominately inorganic kinetic exchange factors, contrary to many other elements (Wiederhold, 2015).
The following is a description of the systems, processes and transformations that control the global silica cycle, and the known constraints on the δ30Si (see Figure 1).
Weathering
Weathering is a major process that fractionates Si isotopes in the critical zone. When primary minerals (−0.90 < δ30Si < +1.40‰, Figure 1) are weathered, light Si isotopes are preferentially incorporated into secondary minerals (−2.95 < δ30Si < +2.5‰) releasing a DSi pool that generally has an enriched isotope composition (−1 < δ30Si < +2‰) (cf. reviews of Opfergelt and Delmelle, 2012; Frings et al., 2016), Figure 1, Supplementary Table 1 and references therein). Adsorption of silicic acid onto Fe oxy-hydroxides (Delstanche et al., 2009) and Al hydroxides (Oelze et al., 2014) has also been demonstrated to preferentially immobilize the lighter Si isotopes. As discussed in section Rivers, all these processes are particularly complex on seasonal to geological timescales (Ziegler et al., 2005) and explain most of the higher δ30SiDSi of continental waters.
Vegetation
Significant fractionation has been observed during the uptake of Si by plant roots (Opfergelt et al., 2006a,b; Ding et al., 2008a,b; Sun et al., 2008) and exogenous material on the surface of vegetation (Engström et al., 2008), although the transporters behind both forms of uptake remain largely unknown. The δ30Si composition of these plants (Figure 1), and in particular of phytoliths is in the first instance regulated by the physical environment around the plant including the soluble Si concentration of the medium (Opfergelt et al., 2006b), the weathering of the soil substrate (Opfergelt et al., 2008) and soil organic matter (Ding et al., 2008a). Beyond this, there is clear evidence that significant isotopic fractionation can occur between plants with increased fractionation in heavy Si accumulators (Ding et al., 2005, 2008a,b; Opfergelt et al., 2006a,b). Rayleigh fractionation during the transportation of Si within plants also causes heavier isotopes to be concentrated within the xylem whilst lighter isotopes are preferentially deposited in phytoliths lower down the plant (Ding et al., 2005, 2008a,b; Opfergelt et al., 2006a,b; Hodson et al., 2008). Phytoliths have higher dissolution rates than other silicate materials (e.g., tephra, clay, feldspars, quartz), and provide a major source of DSi in some soil/terrestrial environments (Derry et al., 2005; Struyf et al., 2009; Cornelis et al., 2010; Opfergelt et al., 2010).
Freshwater
Rivers
Over the past two decades, several studies of δ30Si in DSi in freshwaters have been published, but we are still only beginning to understand the controls and variability of δ30Si in freshwaters. Since the first data of DSi in freshwaters by De La Rocha (De La Rocha et al., 2000), δ30Si has been analyzed from a number of rivers (and lakes) across the world including the Congo River and tributaries (Cardinal et al., 2010; Hughes et al., 2011), the Tana River (Hughes et al., 2012), the Yellow and Yangtze Rivers (Ding et al., 2004, 2011), the Kalix River in Sweden (Engström et al., 2010), Swiss alpine rivers (Georg et al., 2006), and most recently from the Nile River (Cockerton et al., 2013), the Amazon river and tributaries (Hughes et al., 2013), areas of central Siberia (Pokrovsky et al., 2013; Panizzo et al., 2017) and the Ganges (Fontorbe et al., 2013; Frings et al., 2015). The range in δ30Si from freshwaters, thus far analyzed, now stands at −0.17 to +4.66‰ (Figure 1).
In big-picture terms, (almost) every measurement ever made of river water DSi has been heavier than the parent material. When river water DSi concentrations are normalized to a conservative element (typically Na, which is common in silicate rocks), then it becomes clear that Si is removed from solution, either into secondary clay minerals or some form of bSiO2, and that this removal is associated with a discrimination against the heavier isotopes of Si. This is consistent with our understanding derived from the microscale (e.g., Steinhoefel et al., 2011; Schuessler and von Blanckenburg, 2014) to the soil-column scale (e.g., Opfergelt et al., 2012; Pogge von Strandmann et al., 2012). These all show that the new secondary mineral phases have less of the heavier isotopes relative to river waters. The same discrimination is also well established for plants (Ding et al., 2005, 2008a; Opfergelt et al., 2006a,b), freshwater diatoms (Alleman et al., 2005; Panizzo et al., 2016) and sponges, which preferentially utilize 28Si over 30Si (and 29Si), thus leading to an increase in δ30Si in the host water.
Broadly, δ30Si values in river waters reflect the weathering regime, which is ultimately a mass-balance constraint. We can conceptualize two end-member weathering regimes: “kinetically limited” and “supply limited” (almost equivalent to the geomorphological terms “transport limited” and “weathering limited”; see e.g., Stallard and Edmond, 1983). Kinetic limitation refers to a situation where the weathering flux (solute, DSi) is operating at maximum capacity for the conditions; increasing the factors that control the weathering rate (essentially temperature or water supply) will increase the rate of DSi export, because there is an excess of fresh material to be weathered. This could be e.g., the high Himalaya, the Andes, or (sub) glacial catchments (Georg et al., 2007; Fontorbe et al., 2013; Opfergelt et al., 2013) where physical erosion greatly outpaces chemical weathering. At very high erosion rates (low weathering intensity or congruency) river δ30Si is low: there is no time, and no thermodynamic driving force, for clays to form, soils to develop or biology to have a meaningful influence on the δ30Si measured in a stream.
The other end-member (supply limited) is where essentially all material leaves the catchment as a dissolved flux; the rate of solute generation is limited by how quickly material is supplied. Increasing the factors that control the weathering rate (temperature or water supply) will not increase Si export, because there is nothing left to be weathered. In this case the conversion of parent material (bedrock) to river solute is (near-) complete, then there can be no observable fractionation: the river takes on the composition of the bedrock, regardless of how much bSiO2 cycling is taking place in the catchment (and assuming steady-state, and no loss of bSiO2). This is the case in e.g., the lowland, blackwater tributaries of the Amazon (Hughes et al., 2013) and the Congo (Cardinal et al., 2010), where the interpretation is that the swampy, organic-matter-rich environments are conducive to the dissolution of previously formed clays.
Along a gradient of weathering intensity (0 to 1, where 0 is all material exported by physical erosion, and 1 where all denudation occurs as dissolved fluxes), the offset between source material (bedrock) and river δ30Si should be zero at both ends and peak somewhere in the middle (Bouchez et al., 2013). The same is true for Li isotopes (Henchiri et al., 2016); in general, the isotopic separation along a weathering regime gradient is much clearer for Li (δ7Li) than δ30Si (see e.g., Dellinger et al., 2015; Frings et al., 2016). Superimposed on the above is the role of vegetation (see section Vegetation).
All the work published on the major catchment/riverine systems (e.g., Cockerton et al., 2013) highlight the complexity of lake/drainage basin systems, and clearly illustrate that both Si concentrations and δ30Si are closely controlled by a mixture of weathering/erosion processes [regulated by the wider climate system (De La Rocha et al., 2000; Opfergelt and Delmelle, 2012)], aquatic productivity and catchment soil, vegetation. Studies of the Congo Basin, for example, have shown that δ30SiDSi is mainly regulated by the intensity of silicate weathering/secondary mineral formation across the drainage basin with spatial variations further driven by the type of weathering regime and the extent to which clays are dissolved by organic matter (Cardinal et al., 2010). These abiotic processes are then superimposed by seasonal biological processes, in particular by diatom biomineralization in the dry season, which exports significant amounts of Si out of the water column (Hughes et al., 2011). Other examples also account for the downstream increase in Nile River δ30Si to the progressive uptake of DSi by diatoms and other Si-accumulating organisms (Cockerton et al., 2013). Indeed, Cockerton et al. (2013) showed significant seasonal variations in δ30Si from freshwaters, with higher δ30Si in the dry season (+1.54 to +4.66‰) than the wet season (+0.48 to +3.45‰) due to reduced mobilization of dissolved Si from the catchment relative to its aquatic demand. These examples demonstrate the superimposed (often seasonal) control that freshwater bSiO2 plays in regulating δ30Si signatures, where abiotic processes (chemical weathering and secondary mineral formation) otherwise dominate.
Additional examples also show the potential importance of vegetation in altering the Si concentration and/or δ30Si and regulating the flux of Si both into/out of river waters (Alleman et al., 2005; Ding et al., 2011; Hughes et al., 2011; Cockerton et al., 2013; Pokrovsky et al., 2013). For example, along the Yangtze River uptake of DSi by grasses in wetlands and rice in paddy fields drives a progressive increase in δ30SiDSi due to better phytolith preservation in areas of high phytolith production and/or low phytolith dissolution (Ding et al., 2004). In contrast the net impact of vegetation on δ30SiDSi seems to be minimal in the Okavango Delta, Congo and Amazon Basin due to the rapid dissolution/recycling of phytoliths (Cardinal et al., 2010; Hughes et al., 2013; Frings et al., 2014b).
Work on smaller aquatic systems has also shown significant temporal variations in δ30SiDSi. In a boreal river in North Sweden seasonal changes in δ30SiDSi of 0.8‰ were attributed to: (1) the release of plant derived bSiO2/phytoliths (low δ30Si) during snow melt; (2) changing inputs from headwaters; and (3) diatom biomineralization (Engström et al., 2010). Similarly, seasonal changes in δ30SiDSi of up to 0.6‰ were observed in a series of rivers in Switzerland (Georg et al., 2006), whilst variations in a series of Icelandic rivers have been linked to differential erosion rates and secondary minerals formation (Opfergelt et al., 2013).
Other studies have shown a significant anthropogenic impact on δ30SiDSi, often in response to land use changes and dam construction (Sun et al., 2011; Hughes et al., 2012; Delvaux et al., 2013). As a case study, along the River Nile, intensive water management through the irrigation/retention of waters behind dams exerts a significant control on δ30SiDSi (Cockerton et al., 2013). In the Tana River (Kenya), downstream increases (decreases) in δ30SiDSi (DSi) are attributed to both greater water retention time and the higher diatom DSi utilization in dams and reservoirs (Hughes et al., 2012), which leads to heavier (+0.54‰) downstream DSi compositions and a 41% decrease in concentrations. Similarly, outside of diatom growth season periods, the δ30SiDSi signature of Scheldt River (Belgium) tributaries (a human disturbed watershed) are negatively correlated to the percentage of forest cover (r2 = 0.95, p-value < 0.01) and positively correlated to the percentage of arable land cover (r2 = 0.70, p-value < 0.08) in the basin, suggesting that land use is a major control here on δ30SiDSi signatures (Delvaux et al., 2013). All of these examples document how temporal (namely seasonal) and spatial changes in riverine δ30SiDSi in both small and large drainage basins, can have great impact upon the delivery of DSi and δ30SiDSi to the oceans over time (Frings et al., 2016).
Challenges in Interpreting River Geochemistry
As outlined above, the cycling of DSi in river water is not a straightforward process. In general, we have a good conceptual understanding of the qualitative nature of the processes and the associated fractionations, but quantifying the fractionation at each step in the processes is often challenging. This is not a problem that is unique to Si isotope systematics (see Eiler et al., 2014).
The key steps in the generation of river water DSi include: (1) dissolution of the primary mineral, (2) incorporation of some fraction of this as a secondary phase, (3) bio-cycling and soil-column processes. Each of these steps is associated with multiple fractionations: We are beginning to understand that the (overall) Si isotope fractionation associated with any given process is extremely variable. Note that an analogy can be made with the ongoing work on biological Si fractionation (Sutton et al., 2013; Hendry et al., 2015). This range in observable isotope fractionation occurs because each phase transformation is typically several steps, each with their own fractionation. For example, the weathering of a primary silicate mineral to a secondary clay (e.g., plagioclase to kaolinite) will potentially involve breaking of bonds in mineral surface, diffusion across a leached layer and/or thin mineral-fluid boundary layer, de-solvation/coordination into and from solution, transfer to the precipitation site, and the forming of new bonds in the new mineral. Each step can involve isotope fractionation, plus various mass-balance constraints that regulate how much this fractionation can be expressed. They also involve fractionations of two fundamentally different manners: kinetic and equilibrium, which arise from Newtonian dynamics and bond-strength effects, respectively. Importantly, these fractionation events tend to cause opposing fractionation of the Si isotopes and complicate the interpretations of the resulting δ30Si values.
Laboratory experiments confirm that the combined fractionations associated with important processes (Si adsorption or precipitation) are both rate and temperature dependent (Geilert et al., 2014; Oelze et al., 2014, 2015; Roerdink et al., 2015), conclusions that are corroborated by well-designed field experiments (Geilert et al., 2015). In general, it is clear that there is no such thing as a single fractionation factor for a given process. This makes it hard to quantitatively interpret e.g., river DSi at a large scale, especially without a robust independent estimate of “fSi,” the fraction of Si remaining in solution. Catchment mass-balance models (e.g., Bouchez et al., 2013)) can help, but require that river sediment is collected and measured simultaneously and assume catchment steady-state. Predicting the magnitude, and even direction, of changes in the δ30Si values of river water during glacial-interglacial transitions is confounded by multiple parameters that are only qualitatively understood. It is not clear, for example, if these transitions would result in a congruent weathering regime.
Lakes
Lacustrine systems represent a key component of the continental Si cycle, although only a handful of results have been published to date (Figure 1). Whilst riverine inputs cause initial lake profiles to be primarily a function of upstream catchment processes (weathering of rock and productivity inputs), subsequent in-lake DSi biomineralization can significantly modify DSi/δ30SiDSi before waters continue through the outflow and eventually into the marine system (Alleman et al., 2005; Opfergelt et al., 2011). As with the oceans, siliceous productivity in lakes is primarily a function of light and nutrient availability. Accordingly, spatial/temporal patterns of DSi/δ30SiDSi can often be linked to factors including mixing/stratification regimes, which regulate the supply of nutrient rich deep waters to the photic zone and the retention of organisms in surface waters (Alleman et al., 2005).
The net efficiency of the biological pump in transferring Si from the water column into the sediment record is often higher in lakes than marine systems, due to lower dissolution rates that are in turn influenced by faster rates of sinking and sediment accumulation. However, in instances where dissolution is significant, for example in response to elevated pH or salinity, the impacts are clearly observed with elevated DSi concentrations and lower δ30Si (Opfergelt et al., 2011). Importantly, the contemporary studies carried out to date all highlight the potential for δ30Si to be used to study temporal changes in Si cycling, for example through the use of sediment cores. Whilst several organisms (diatoms, radiolaria, sponges) have been explored for this purpose in marine environments, work on freshwater systems has so far focused on diatoms with few data for other organisms (e.g., sponges Hughes et al., 2011). Calibrations have replicated marine studies in demonstrating a fractionation factor of ca. −1.1‰ that is independent of temperature and species (Alleman et al., 2005). Furthermore, contemporary data of δ30Sidiatom from open sediment traps placed down the Lake Baikal water profile found a near constant δ30Sidiatom composition, suggesting the full preservation of the signal through the water column (Panizzo et al., 2016). More importantly, their comparison with δ30Sidiatom in surface sediments, where high diatom valve dissolution occurs, argued for the absence of a Si isotope fractionation associated with diatom dissolution (Panizzo et al., 2016).
Challenges in Interpreting Lake Geochemistry and Palaeolimnological Records
As outlined above, there have been a handful of studies to date that have examined the application of δ30SiDSi in limnological settings (Alleman et al., 2005; Opfergelt et al., 2011; Panizzo et al., 2016, 2017). Lakes, in addition to the soil-vegetation system, play an important role in buffering continental DSi export to the oceans. Previous assumptions were that this buffering capacity of the continental Si cycle was in steady-state; however, this has now been challenged (Frings et al., 2014a). Frings et al. (2014a) highlight the effectiveness of lacustrine environments in retaining DSi in sediments via the bSiO2 pump, which on a global scale can translate to a net export of between 21 and 27% of river DSi export. This equates to an estimation of 1.53 Tmol yr−1 (Frings et al., 2014a) (Figure 1). These data highlight the challenges in underpinning global estimates of lake biogeochemical cycling (particularly for large lakes and reservoirs) to better constrain the evolution of the global biogeochemical cycling of Si.
Marine
Silicon plays an important role in the marine realm as a vital macronutrient for silicifying organisms such as diatoms, radiolarians, siliceous sponges and silicoflagellates. Due to their short lifetimes and the dynamic, boom-bust nature of their populations, diatoms are dominantly responsible for the cycling of Si within the ocean, due to the large fluxes associated with their uptake and export of Si (Figure 1), although other siliceous organisms may contribute to sedimentary standing stocks of Si in some regions (Maldonado et al., 2011). Diatoms are thought to be responsible for up to ~40% of oceanic primary productivity (Nelson et al., 1995), and are especially efficient at exporting carbon from the surface ocean (Buesseler, 1998), giving them an important role in the oceanic biological pump of carbon. The marine cycles of Si and C are thus linked, albeit not in any simple manner, due to the differential cycling of Si; the release of Si from diatom frustules is slower relative to the remineralization of C from diatom cells (Bidle and Azam, 1999; Ragueneau et al., 2000; Bidle et al., 2002; Smetacek et al., 2012). The largest Si fluxes within the global ocean are the result of diatom uptake in and export from the surface ocean (Tréguer and De La Rocha, 2013) (Figure 1), most of which takes place in the upwelling regions of the high-latitude open ocean (Honjo et al., 2008) as well as in coastal upwelling regions. Diatoms preferentially take up the lighter isotopes of Si during silicification (De La Rocha et al., 1997; Milligan, 2004; Sutton et al., 2013), resulting in a higher δ30SiDSi signature in surface ocean DSi (Figure 1). Seawater δ30SiDSi variations are thus primarily created by the fractionation by diatom uptake in the euphotic zone of the ocean, where these photosynthesizers grow. This surface fractionation, combined with the oceanic circulation, is the major control on the global oceanic distribution of δ30SiDSi, which has highlighted some key features of the nature of marine Si cycling in recent years. However, other processes such as hydrothermal activity, and early sedimentary diagenesis are poorly quantified, and thus their impact on the interpretation of Si isotope and trace element signals in sedimentary bSiO2 remains unknown
Global Water Column Distribution
Since active fractionation of δ30Si by diatoms occurs within the euphotic zone of the ocean, the most extreme δ30SiDSi values are found within the uppermost 100 m of the water column, with values reaching above +3‰ both in highly nutrient-depleted oligotrophic gyres (with DSi concentrations <1 μM; Reynolds et al., 2006; Grasse et al., 2013) as well as in the slightly more Si-rich high-latitude upwelling regions (with DSi concentrations of 2–13 μM; Varela et al., 2004; Fripiat et al., 2011; de Souza et al., 2012a). These values are amongst the most highly fractionated δ30Si values reported in the literature (Reynolds, 2011), and at any given location within the ocean, the surface ocean represents the highest δ30SiDSi value found in the water column (e.g., Cardinal et al., 2005; Cavagna et al., 2011; Ehlert et al., 2012). Below the maximum in the surface ocean, depth profiles of δ30SiDSi typically show a monotonic decrease with depth, mirroring the increase in DSi concentration. The steepness of this isotopic gradient varies with oceanographic setting, with sharp δ30SiDSi decreases observed in upwelling regions where DSi concentrations increase quickly with depth (e.g., Cardinal et al., 2005; Ehlert et al., 2012). Elsewhere in the open ocean, where the silicicline is depressed and DSi concentrations increase only slowly through the uppermost 1,000 m of the water column, δ30SiDSi values remain elevated to intermediate depths, and can reach up to +1.8‰ at depths of ~800 m (de Souza et al., 2012a). These elevated values, which result from fractionation of δ30Si within the euphotic zone, provide robust evidence that the bSiO2-poor nature of the upper ocean is due to the spreading of DSi-depleted water masses that derive from the Southern Ocean, as initially suggested by Sarmiento et al. (2004). These water masses, Sub-Antarctic Mode Water (SAMW) and Antarctic Intermediate Water (AAIW), form from high-latitude surface water masses from which DSi has been stripped by the uptake of heavily silicified Southern Ocean diatoms. This DSi depletion is associated with an isotope fractionation, imparting SAMW and AAIW a high δ30SiDSi signature (de Souza et al., 2012a), a process that has been dubbed “Southern Ocean isotope distillation” by Brzezinski and Jones (Brzezinski and Jones, 2015). The high δ30SiDSi signature of SAMW and AAIW is exported to the upper ocean at a near-global scale (De La Rocha et al., 1998; de Souza et al., 2012a,b; Brzezinski and Jones, 2015; Singh et al., 2015). The upper-ocean δ30SiDSi distribution thus reflects the importance of Southern Ocean water-mass subduction in resupplying DSi, together with other major nutrients, to the nutrient-poor low-latitude upper ocean. The major exception to this strong Southern Ocean influence is the DSi-rich upper ocean of the North Pacific Ocean (Reynolds et al., 2006), which obtains its DSi inventory via upwelling in the subarctic Pacific and also influences the upper-ocean δ30SiDSi distribution of the equatorial Pacific (Beucher et al., 2008, 2011; de Souza et al., 2012a).
By far the most coherent large-scale δ30SiDSi variability in the global ocean is observed in the deep and abyssal ocean, below about 2,000 m. Here, DSi concentrations vary by a factor of ~20, from a little over 10 μM in the DSi-poor waters formed in the North Atlantic Ocean, to over 180 μM in the old, DSi-rich waters of the North Pacific Ocean (Garcia et al., 2014). The strongest DSi gradient is observed within the deep waters of the Atlantic Ocean, where DSi-poor North Atlantic Deep Water (NADW) overlies DSi-rich abyssal waters of Southern Ocean origin (Antarctic Bottom Water, AABW). This DSi gradient is associated with a coherent δ30SiDSi gradient between isotopically light AABW (+1.2‰) and isotopically heavy NADW with values up to +1.8‰ (de Souza et al., 2012b; Brzezinski and Jones, 2015). It has been argued that the elevated δ30SiDSi value of NADW is the result of the cross-equatorial transport of a heavy δ30SiDSi signature by SAMW and AAIW, which flow northward in the Atlantic Ocean to close the upper limb of the meridional overturning circulation (Ehlert et al., 2012; de Souza et al., 2015), illustrating the degree to which Southern Ocean Si utilization and the associated “distillation” of isotopes affects the δ30SiDSi distribution at the global scale even in the deep ocean. Indeed, this Southern Ocean influence would appear to extend as far north as the Arctic Ocean: recent work has shown that Arctic deep waters Ocean bear the most elevated δ30SiDSi values in the global ocean, with values averaging +1.9‰ (Varela et al., 2016). Since the Arctic Ocean receives most of its inflow from intermediate waters of the northern Atlantic Ocean, which are influenced by AAIW, the low-DSi, high-δ30SiDSi end-member characteristics of the deep Arctic Ocean may ultimately derive from Southern Ocean isotope “distillation,” although the role of more local processes such as Si cycling within the Arctic Ocean as well as Si input from riverine discharge need to be investigated in more detail.
Apart from the Atlantic and Arctic Oceans, the deep water δ30SiDSi distribution shows markedly little variability, with Indian and Pacific Ocean deep waters exhibiting δ30SiDSi values of around +1.2‰ to +1.3‰ (Beucher et al., 2008, 2011; de Souza et al., 2012a; Grasse et al., 2013; Singh et al., 2015), i.e., essentially invariant considering the degree of analytical consistency between laboratories (Reynolds et al., 2007; Grasse et al., 2017). Although there are some hints of heterogeneity in the North Pacific, e.g., in the Cascadia Basin proximal to the North American continent (Beucher et al., 2008), when compared to the ~0.5‰ meridional gradient observed in the Atlantic Ocean, δ30SiDSi variability in the Indian and Pacific Oceans is neither particularly significant nor systematic. A possible explanation for this homogeneity comes from a modeling study of the controls on the deep ocean δ30SiDSi distribution (de Souza et al., 2014), which found that deep water δ30SiDSi values outside the Atlantic and Arctic Oceans are very strongly governed by the export of isotopically light DSi-rich abyssal water from the Southern Ocean.
Hydrothermal Vents
There are two types of hydrothermal vents that influence the marine Si cycle: (A) on-ridge hot (near critical point) black smoker fluids and, (B) warm (>45°C) and cool (<20°C) Ridge Flank Hydrothermal Fluids (RFHF). They are inherently similar in terms of the Si transfer to the ocean because (a) reactions at high and low temperature leach Si from the oceanic crust, resulting in high (17 ± 3 mmol/kg for hot) and moderate (~0.5 ± 0.2 mmol/kg for warm and cool) DSi hydrothermal fluids and (b) cooling of the hot fluids or warming of the cool and warm fluids removes DSi through precipitation of smectite-like clays before their venting from the seabed (Wheat and McManus, 2005). At a global scale, high temperature hydrothermal DSi fluxes can be constrained at a maximal of 0.14 ± 0.02 Tmol yr−1 using a Si input of 17 ± 3 mmol/kg (the average concentration in hydrothermal vents at an exit temperature of 300°C) with a water flux at 8 × 1012 kg yr−1 (Coogan and Dosso, 2012). However, the influence of hydrothermal inputs on the global marine Si cycle remain uncertain due to the paucity of δ30Si data available. To date, the only published data are two data points (−0.4‰ and −0.2‰) for hydrothermal vents on the East Pacific Rise (De La Rocha et al., 2000).
Sedimentary Processes
Upon deposition on the seabed, recycling of SiO2 during early diagenesis in marine sediments involves: (a) initial dissolution of bSiO2 and/or detrital siliceous phases and consequent DSi build-up in marine pore waters (b) flux of dissolved Si out of the sediments and (c) bSiO2 reconstitution and/or precipitation of SiO2-rich diagenetic solid phases. The three recycling processes take place simultaneously and ultimately control DSi fluxes and diagenetic SiO2 deposition and storage in marine sediments (Dixit et al., 2001; Aller, 2014) (Figure 1). When early diagenetic siliceous precipitates are rich in seawater-derived cations (e.g., K, Mg) the precipitation process is called “reverse weathering” and the cation-rich precipitates “reverse weathering products” (Mackenzie and Garrels, 1966; Michalopoulos and Aller, 1995).
Pore water and biogenic silica preservation
The δ30Si of pore waters from marine sediments is an indicator of diagenetic turnover of Si and its impact on the preservation of environmental signals in bSiO2 (Ehlert et al., 2016). In fine-grained terrigenous sediments of the Amazon delta these authigenic Al-Si phases form at a rate as high as 280 μmol Si cm−2 yr−1 (Michalopoulos and Aller, 2004). In shelf sediments the rate is thought to be substantially lower at around 56 μmol Si cm−2 yr−1, which, however, means that approximately 24% of the total dissolving bSiO2 re-precipitates in the upper few centimeters of the sediment (Ehlert et al., 2016). Several experimental studies have shown that such Si precipitation is associated with strong enrichment of light Si isotopes up to −4.5‰ in the product although they were not focused on sediment—pore water diagenesis (e.g., Geilert et al., 2014; Oelze et al., 2014, 2015; Roerdink et al., 2015).
To date, there have been only a few studies that investigated the effect of diagenesis on preservation of the primary environmental signal in bSiO2 and they found potentially differing results. Demarest et al. (2009) reported a preferential release of light Si isotopes during partial dissolution of bSiO2 in batch reactors, whereas leaching experiments with sediments showed no fractionation (Wetzel et al., 2014; Tatzel et al., 2015). In young hemi-pelagic sediments, the early diagenetic formation of new Al-Si phases is focused in the topmost layer of the sediment column (ca. 0–10 cm sediment depth). Ehlert et al. (2016) found pore waters from the Peruvian shelf with high δ30Si (average 1.46 ± 0.22‰) with the highest values occurring close to the sediment–water interface. These values were enriched compared to bSiO2 in the cores (average 0.85 ± 0.28‰) and overlying bottom water (<1.5‰), which is consistent with the formation of authigenic Al-Si phases from the dissolving bSiO2. The fractionation factor between the precipitates and the pore waters was estimated at −2.0‰. However, the isotope composition of the bSiO2 seemed to remain constant within the reactive surface layer where most of the dissolution and precipitation occurred, which makes isotopic fractionation during dissolution unlikely. From leaching experiments on older Pleistocene and Pliocene sediments, Tatzel et al. (2015) report that the formation of authigenic Al-Si phases should be 2‰ lighter than the pore waters they formed from, in agreement with previous experimental results (Delstanche et al., 2009; Geilert et al., 2014; Oelze et al., 2014, 2015). However, the authors observed an increase of the Si isotope composition of the preserved SiO2 during phase transformation from opal-A to opal-CT. The reason for this was an isotope exchange between SiO2 and pore water, and the Si isotope increase was higher with an increase in the amount of detrital material in the sediment.
Diagenetic silica precipitation and reverse weathering
It has been proposed that reverse weathering reactions in marine sediments are significant oceanic sinks for K, Mg, and alkalinity (Mackenzie and Garrels, 1966; Rude and Aller, 1994; Michalopoulos and Aller, 1995, 2004; Sun et al., 2016). The precipitation process can also involve the uptake of minor elements (Li, Ge, F) into new authigenic siliceous phases, which can act as primary sinks and thus balance the oceanic mass and isotopic budget. The incorporation of redox-sensitive elements (e.g., Fe) depends on the availability and early diagenetic cycling of the relevant element in dissolved form.
Currently, the flux of reverse weathering on the global Si cycle is estimated at −0.63 ± 0.6 Tmol yr−1 (Frings et al., 2016) or 1.5 ± 0.5 (Tréguer and De La Rocha, 2013) (Figure 1, Table S1), however, it is assumed that this flux is under-estimated. It has been proposed that reactive bSiO2 that has undergone reconstitution/dissolution/re-precipitation transformations remains partially unaccounted for (Michalopoulos and Aller, 2004; Presti and Michalopoulos, 2008) using the traditional operational leach method of DeMaster (1981). However, measurements of cosmogenic 32Si incorporated in reactive SiO2 products indicate that even the modified alkaline leach methods used to date in high sediment accumulation rates underestimate the amount of reconstituted bSiO2 and authigenic silicates stored in deltaic sediments (Rahman et al., 2016). Thus the conservative estimates for the net storage of altered bSiO2 and authigenic silicates in deltaic sediments range from 2–3x in the Mississippi (Presti and Michalopoulos, 2008) to 5–10x in the Amazon (Michalopoulos and Aller, 2004). Based on 32Si, actual storage in the Amazon delta may be 2–3x greater (i.e., 3–4.5 Tmol yr−1) than the best recent conservative estimates (Rahman et al., 2016).
In general, the impact of dissolution, diagenesis and reverse weathering on the stable isotope analysis of bSiO2 are poorly understood. Whilst it is possible to assess the degree of recrystallization of bSiO2 through geological time (De La Rocha, 2003), there are some early stage dissolution and diagenetic processes that can occur in the water column and shallow sediments that may be more challenging to detect. However, studies seem to indicate that with high bSiO2 concentrations in the sediments, the original δ30Si of the bSiO2 is more likely to be preserved because exchange with pore water and the formation of authigenic Al-Si phases is limited.
Additional Challenges in Interpreting Marine Data
In addition to environmental alteration (see sections Pore Water and Biogenic Silica Preservation and Diagenetic Silica Precipitation and Reverse Weathering) and any analytical challenges associated with the separation and cleaning (see section Multi-proxy Geochemical Approaches in Palaeoceanography) of bSiO2 from sediment cores, ensuring the effective removal of any contaminant phases (Morley et al., 2004; Egan et al., 2012), there are a number of different biological and ecological assumptions that are required for the interpretation of both Si and oxygen isotopes (δ18O) in diatom bSiO2.
Firstly, many downcore studies assume a constant δ30Si fractionation factor of −1.1 ± 0.4‰, which was based on the initial culture studies carried out by De La Rocha et al. (1997). However, more recent studies show that this fractionation factor may be more variable, and potentially species-specific, based on both field studies (e.g., Varela et al., 2004; Beucher et al., 2007; Ehlert et al., 2012; Fripiat et al., 2012) and culture experiments (Sutton et al., 2013).
Secondly, the mathematical models of DSi utilization also assume a starting value of δ30SiDSi for the upwelling water, which supplies the essential nutrients to diatoms in the surface layer. To a certain extent, this can be estimated using simple box models (e.g., Beucher et al., 2007), and/or deep-water archives [see section Palaeoceanographic (and Palaeolimnological) Applications below Horn et al., 2011; Egan et al., 2013]. There is a potential that changes in water mass circulation, or end-member composition, could impact the distribution of silicon isotope ratios in the oceans (Hendry and Robinson, 2012) (see section Global Water Column Distribution). Furthermore, changes in inputs and—potentially—biological productivity and diatom bSiO2 burial could change the whole ocean Si isotope budget over periods of time longer than the residence time of Si in the ocean, 10–15 thousand years (Frings et al., 2016; Hawkings et al., 2017).
Palaeoceanographic (and Palaeolimnological) Applications
The stable isotopic composition of biogenic bSiO2, together with the concentration and stable isotope composition of trace elements within the bSiO2, provide a useful source of palaeoceanographic data. The remains of silicifiers, isolated from marine sediments, are effective archives of marine Si cycling at different depths within the water column, yielding key information about DSi supply and utilization by marine algae. Si cycling by algae is important to quantify if we are to understand the processes that are critical to the cycling of carbon in the Earth's climate system, such as changes in organic carbon production and export, oceanic circulation and chemical weathering (Frings et al., 2016). Biogenic SiO2 has the potential to provide records of marine Si cycling throughout the Cenozoic in the case of diatoms, and further back into the Meso- and Palaeozoic in the case of radiolarians and sponges, as it is well-preserved and widespread in sediments, and can be readily assessed for diagenesis through XRD analysis (e.g., De La Rocha, 2003). In addition, bSiO2 archives are useful in climatically important locations where the more traditional, carbonate-based palaeoceanographic archives (principally foraminifera) are poorly preserved, such as the Quaternary Southern Ocean.
In the following sections, we will provide a summary of the bSiO2-based palaeoceanographic tools available, including trace element geochemistry (section Trace Element Geochemistry of Diatoms and Sponges) and major element stable isotope methods (section Major Element Stable Isotope Ratios in Diatoms, Sponges and Radiolarians). These sections emphasize the value of multi-archive and –proxy approaches to better constrain changes in silicon (bio)geochemistry and examples of such applications are provided within the context of palaeoceanography in section Multi-proxy Geochemical Approaches in Palaeoceanography (e.g., Silicic Acid Leakage Hypothesis). Although we will focus on the more widely studied silicifying organisms (diatoms, sponges and radiolarians), there is a wealth of other silicifying organisms in the oceans that are yet to be explored for their potential as oceanographic archives.
Trace Element Geochemistry of Diatoms and Sponges
Biogenic silica incorporates low quantities of trace elements into the structure. Although often in parts per million—or less—the concentrations of these elements (often normalized to Si) can reveal information about the environmental conditions during growth. Many of these elements show strong correlations with DSi in seawater and exhibit “refractory nutrient-like” profiles.
Aluminum
Aluminum is one of the most abundant, but highly variable, trace elements in bSiO2 ranging from 0.0001 to 0.1 g/g (e.g., Hendry et al., 2011). Although Al uptake occurs by adsorption and is present in associated clay phases (van Bennekom and van der Gaast, 1976; Moran and Moore, 1988), Al is also incorporated into living diatom bSiO2 that is still protected by an organic matrix (Gehlen et al., 2002). X-ray absorption spectra at the Al-K edge show that Al occurs in 4-fold co-ordination, with tetrahedra inserted inside the bSiO2 framework, although samples of diatom bSiO2 taken from natural marine waters show both 4- and 6-fold co-ordination, possibly as a result of clay contamination (Gehlen et al., 2002). Whilst there appear to be some climatic signals in Al/Si in diatom bSiO2 (e.g., Hendry et al., 2011), Al/Si appears to be highly susceptible to rapid alteration during early sedimentation processes, most likely sourced from clay material (Ren et al., 2013) increasing an order of magnitude or more between water column or sediment trap samples and sediment core tops in both laboratory and field studies (Koning et al., 2002, 2007; Loucaides et al., 2010; Hendry et al., 2011). In addition to Al concentration analysis, diatom bSiO2 26Al and 10Be have also been explored as archives of marine cosmogenic nuclide concentrations (Lal et al., 2006). Less is known about the Al content of other silicifiers, although limited data indicate that the Al/Si of fresh sponge bSiO2 is lower than that of diatoms (<0.0002 g/g; Hendry and Andersen, 2013).
Germanium
In the ocean, dissolved inorganic germanium (Ge) cycles in a manner similar to that of DSi, with Ge uptake and regeneration from diatom frustules being the main control on its oceanic distribution (Froelich and Andreae, 1981; Froelich et al., 1989; Mortlock and Froelich, 1996; Sutton et al., 2010). Although Ge mimics Si, differences in their geochemical behavior can occur reflecting differences in the oceanic inputs and losses and subtle differences in the biogeochemical cycling of these two elements within the ocean. Because of differences in molecular weight and subtle differences in chemistries, the uptake, incorporation and regeneration of Ge from diatom bSiO2 is also non-ideal. These differences are especially apparent at low DSi and Ge concentrations where there is apparent discrimination of Ge during Si uptake via bSiO2 formation (Sutton et al., 2010), which lead to significant non-zero Ge intercept and slightly non-linear behavior of the global Ge-Si relationship ([Ge] = 0.76±0.00[Si] + 0.08±0.17, r2 = 1.00, n = 248, Ge in pmol L−1 and Si in μmol L−1). The fractionation of Ge during uptake with Si is readily apparent in deep sea sponges which have a lower affinity for Si uptake (Froelich and Barthel, 1997; Reincke and Barthel, 1997; Maher et al., 2006).
The two main sources of Ge and Si to the ocean are mineral weathering and hydrothermal fluids (Mortlock and Froelich, 1987; Mortlock et al., 1993; Kurtz et al., 2002). Weathering produces low Ge:Si ratios (Ge/Si) input ratios as reflected in the Ge/Si for rivers (0.3–1.2 μmol/mol), while hydrothermal fluids have higher Ge/Si input ratios (8–14 μmol/mol) (Mortlock and Froelich, 1987; Mortlock et al., 1993; Kurtz et al., 2002). The two main sinks for Ge removal from the ocean are through incorporation into bSiO2 and Ge lost via in non-opal phases (Murnane et al., 1989; Hammond et al., 2000, 2004; King et al., 2000; McManus et al., 2003). Currently, there is debate about the magnitude and loss via the ‘missing Ge sink’ (Hammond et al., 2000, 2004; King et al., 2000; McManus et al., 2003; Baronas et al., 2016; Rouxel and Luais, 2017). Recently Baronas et al. (2016) have suggested that Ge loss from the ocean appears to be dependent on pore-water oxidation–reduction reactions and the formation authigenic aluminosilicate (see section Sedimentary Processes) minerals within marine sediments. Note, a recent study looking at the links between Ge isotopes and silicon in Hawaiian hydrothermal vents has shown the potential of this new isotopic system to trace the fate of hydrothermal elements released into the ocean (Escoube et al., 2015).
While inorganic Ge can be lost from solution through incorporation into biogenic silica and association with authigenic mineral formation, there is a large unreactive organic Ge pool within the dissolved phase; monomethyl- and dimethyl-germanium. The formation of these two species is relatively unknown, but it is speculated to be continentally derived and lost under anoxic conditions (Lewis et al., 1985, 1988, 1989). The residence time for these methylated forms of Ge are on the order of 100,000+ years, which is considerably longer than that of inorganic Ge.
In addition, inorganic Ge can form stable complexes with organic molecules (e.g., humic acids) containing di- and polyfunctional carboxylic acids, polyalcohols and ortho-diphenols functional groups. Complexation of Ge by humic acids in freshwater regimes needs to be considered with interpreting variations in the Ge/Si ratio for the global ocean through time with respect to terrestrial inputs (Pokrovski and Schott, 1998). The organic Ge complexation has also been suggested to explain the low Ge/Si ratio in phytoliths due to different pathways of Ge relative to Si in the plant: Ge is not discriminated against at the root–soil solution interface but it is organically trapped in roots, in contrast to Si (Delvigne et al., 2009).
The Ge/Si imprinted into diatom frustules has proven useful for palaeo-reconstructions but it gives no information regarding the sizes of the Si and Ge oceanic pools. The overall Ge/Si for the diatom bSiO2 from the Southern Ocean is relatively constant and appears to reflect seawater Ge/Si (Shemesh et al., 1988; Froelich et al., 1989); however, the late Pleistocene diatom Ge/Si record shows clear, systematic variations between interglacial (Ge/Si = 0.70–0.78 μmol/mol) and glacial periods (Ge/Si = 0.45–0.60 μmol/mol) (Froelich et al., 1989; Mortlock et al., 1991; Bareille et al., 1998) suggesting that size of either the Si or the Ge pool has varied (Murnane et al., 1989; Froelich et al., 1992; Hammond et al., 2000, 2004; King et al., 2000; McManus et al., 2003). To date, little progress has been made to follow up on this earlier work and our understanding on what drives the large variations has little advanced since the work of King et al. (2000), Hammond et al. (2004), and McManus et al. (2003) (see Rouxel and Luais, 2017 for more detail).
Zinc
Zinc is incorporated into diatom bSiO2 in approximately 1–20 ppm levels. Culture studies show that the Zn/Si content of diatom bSiO2 relate to the concentration of free Zn2+ ions in the ambient seawater (Ellwood and Hunter, 1999). However, more recent observations of Zn speciation in the Southern Ocean question this link, because the variability in free Zn2+ ion concentrations are not captured by diatom bSiO2 Zn/Si (Baars and Croot, 2011). Field studies support a link between diatom bSiO2 Zn/Si and productivity and diatom bSiO2 burial rates, although the mechanism behind this relationship is less clear and could relate to nutrient uptake vs. supply, growth rate or salinity (Hendry and Rickaby, 2008; Andersen et al., 2011). The Zn/Si ratio of diatom bSiO2 from sediment cores, which appears to be less susceptible to initial alteration of the bSiO2 as compared to Al/Si (Hendry and Rickaby, 2008), has been used in a small number of studies as a proxy for changes in Zn supply to surface waters in the open ocean and coastal Southern Ocean regions (Ellwood and Hunter, 2000; Hendry and Rickaby, 2008). Recent studies indicate that a large proportion of Zn uptake by diatoms is into the organic material in the cell rather than the bSiO2, challenging its use as a palaeoceanographic proxy (Twining et al., 2003). Thorough cleaning protocols are required to ensure only the Zn incorporated into the silica structure is analyzed (Hendry and Rickaby, 2008; Andersen et al., 2011).
Zinc isotopes (denoted by δ66Zn) in diatom bSiO2 ranges from 0.7 to 1.5‰ and appears to relate to changes in seawater δ66Zn composition because of biological drawdown of isotopically light Zn by phytoplankton. If this is the case, then δ66Zn in diatom bSiO2 from sediment cores has the potential to be used as an archive of Zn biogeochemical cycling in the past (Andersen et al., 2011). However, Zn isotopic fractionation during incorporation into diatoms appears to be determined by whether the diatom is utilizing a high or low-affinity Zn transport mechanism, in addition to ambient free Zn2+ concentrations, and is also heavily influenced by surface adsorption and organic matter incorporation (John et al., 2007; Zhao et al., 2014).
One study has shown that Zn/Si in sponges relates to the flux of particulate organic carbon (POC) to sediments (Ellwood et al., 2004), leading to its potential use as an export production proxy (Ellwood et al., 2005). Sponge δ66Zn systematics appear to differ between major groups of silicifying sponges: δ66Zn reflects seawater zinc isotopic values in Hexactinellids, whereas Demosponge δ66Zn, most likely reflects a combination of internal isotopic fractionation and fractionation of isotopes in dietary organic matter (Hendry and Andersen, 2013).
Major Element Stable Isotope Ratios in Diatoms, Sponges and Radiolarians
Given that bSiO2 is high-purity SiO2, the most straightforward targets for geochemical analysis are stable Si and oxygen isotope ratios (denoted by δ30Si and δ18O, respectively). However, other major elements, including C and N that are encased within the bSiO2, have also been analyzed. Here, we review recent developments in the use of major isotope systems in bSiO2 produced by diatoms, sponges and radiolarians as palaeoceanographic proxies.
Carbon
Increasingly, studies of carbon isotopes are turning to the analysis of specific organisms rather than bulk organic matter so that changes in the carbon cycle can be better investigated without the complication of a mixed source. For example, δ13C analysis can be undertaken on the cellular (occluded) organic matter in diatoms, which is well protected from degradation by the frustule. During photosynthesis, organic carbon matter is formed from both and CO2(aq) and incorporated into diatoms (Tortell and Morel, 2002). The main control on δ13Cdiatom is the balance between supply and demand for Dissolved Inorganic Carbon (DIC), driven by variations in biological productivity or carbon cellular concentrations. Organisms, including diatoms, preferentially use 12C over 13C, so as photosynthetic demand for DIC rises, increasing amounts of 12C are removed from the DIC pool, leading to progressively higher δ13Cdiatom (Laws et al., 1995). Other factors that could influence δ13Cdiatom include changes in the δ13CDIC of inputs to the water column, particularly in the lacustrine environment where the sources of the carbon supplied will be more variable (Barker et al., 2013), dissolution of diatoms, which releases carbon into the ambient water, and changes in CO2(aq) concentration (Laws et al., 1995; Rau et al., 1996, 1997). Therefore, if factors such as inter-species vital effects can be controlled for (Des Combes et al., 2008), or if samples of a single species are analyzed (Swann and Snelling, 2015), then changes in δ13Cdiatom can be used to reconstruct environmental changes. In marine sediments, this is usually primary productivity (e.g., Shemesh et al., 1995; Panizzo et al., 2014; Swann and Snelling, 2015) and/or changes in CO2 (Heureux and Rickaby, 2015; Stoll et al., 2017). In the case of lake sediments, δ13Cdiatom is used to reconstruct changes in the balance between the source and amount of carbon supply and the productivity within the lake ecosystem (e.g., Barker et al., 2013).
Nitrogen
The measurement of variations in diatom-bound stable isotopes of nitrogen (expressed as δ15N) is a powerful tool to reconstruct past nitrogen (N) utilization from palaeo-archives (e.g., Sigman et al., 1999; Robinson et al., 2004). To date, ~10 studies have used diatom-bound δ15N obtained from sediment cores to reconstruct palaeo N cycling, with a strong focus on the Southern Ocean (e.g., Crosta and Shemesh, 2002; Robinson et al., 2004, 2005; Robinson and Sigman, 2008; Horn et al., 2011). Shemesh et al. (1993) was the first to develop a method that used the organic matter contained within the hydrated SiO2 matrix of the diatom, making it possible to measure diatom-bound N stable isotopes (15N and 14N). However, during the last decade the chemical preparation has systematically improved (e.g., Sigman et al., 1999; Robinson et al., 2004). During nitrate uptake, diatoms preferentially incorporate the lighter isotope (14N) into the organic matter (e.g., Altabet et al., 1991; Montoya and McCarthy, 1995), which affects the isotope composition of both the diatom and the residual nitrate. An enzymatic pathway catalyzes this process, where nitrate is reduced to nitrite by nitrate reductase (e.g., Needoba et al., 2003), which can lead to high fractionation factors of up to 14.0‰ in cultured polar diatoms (Horn et al., 2011). Generally, the fractionation of diatom-bound δ15N shows a broad range (1–14‰), with high δ15N variations among diatom species (1.9–11.2‰) with weak relationship between δ15N and cellular size and/or surface area (Horn et al., 2011). Unlike bulk sedimentary δ15N, which is routinely measured to reconstruct N cycling (N utilization and N-loss process), preparation of diatom-bound δ15N is more time consuming and cost intensive (Robinson et al., 2012), but does not appear to be isotopically altered by early bacterial diagenesis (e.g., Freudenthal et al., 2001).
Oxygen
The oxygen isotope (δ18O) values of calcareous marine microfossils, notably foraminifera, have been used extensively to constrain changes in ocean dynamics (such as global ice volume) over the past 70 Ma of Earth history (e.g., Zachos, 2001; Lisiecki and Raymo, 2005). Antarctic ice volume has likely played a significant role in Cenozoic deep water formation, oceanic circulation, and global-scale climate variations; however, poor preservation of calcareous fossils in high-latitude marine sediments has limited attempts to expand the geographical resolution of the benthic marine carbonate oxygen isotope record (Sarmiento and Toggweiler, 1984; Elderfield and Rickaby, 2000; Lisiecki and Raymo, 2005; Raymo et al., 2006). Unlike carbonate minerals, bSiO2 is generally well preserved in high-latitude marine sediments; therefore, the δ18O values of diatom bSiO2 from these regions have the potential to vastly expand our understanding of coupled ice-ocean dynamics (e.g., DeMaster, 2003; Swann and Leng, 2009).
Little is known about spicule δ18O systematics, although there appear to be complex biological vital effects in freshwater sponges (Matteuzzo et al., 2013). A study of a marine carnivorous sponge δ18O showed large fractionation effects and highly heterogeneous isotopic behavior within one individual (Hendry et al., 2015). One downcore study indicates that there are systematic offsets between spicule and diatom bSiO2 δ18O (Snelling et al., 2014).
Silicon
Many (at least 25) glacial-interglacial records, from marine sediment cores, of δ30Si of silicifying organisms (i.e., diatoms, sponges, radiolarians) display a similar trend: lower glacial δ30Si, higher interglacial δ30Si (see Figure 2). While this general pattern has been noted before, most of the records have been interpreted in isolation in terms of nutrient palaeo-utilization (or Si concentrations, for spicule records), and conclusions are related to local-regional nutrient supply (ocean circulation). Trace metal ratios (see section Trace Element Geochemistry of Diatoms and Sponges) may provide an additional constraint but interpretation is difficult since the marine budgets of both the elements (i.e., the trace element, together with the Si cycle) are poorly constrained.
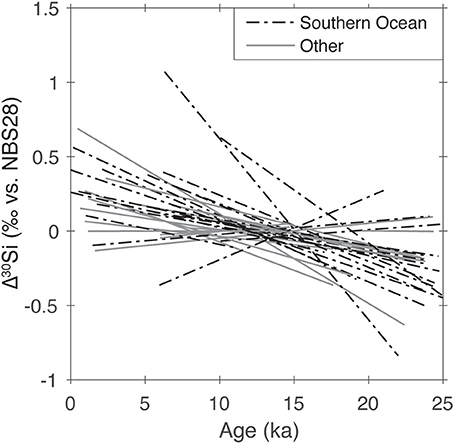
Figure 2. Least-squares regression lines fit though sedimentary δ30SibSiO2 records from 25 ka to present, standardized to mean δ30Si of the record: almost all records show an increase from the glacial to the Holocene, with an average trend of ca. 0.045‰ ka-1, suggestive of a common mechanism. Data are from Beucher et al. (2007), Brzezinski et al. (2002), De La Rocha et al. (1998), Doering et al. (2016), Ehlert et al. (2013), Ellwood et al. (2010), Hendry et al. (2010), Hendry et al. (2014), Hendry et al. (2012), Horn et al. (2011), Kim et al. (2017), Maier et al. (2013), Panizzo et al. (2014), Pichevin et al. (2012), Pichevin et al. (2009), Sutton (2011), and Xiong et al. (2015).
Diatoms
Silicon isotope ratios in diatom bSiO2 have been used widely as a proxy for marine DSi utilization in surface waters due to the preferential fractionation of the lighter isotope of Si during diatom silicification (see section Marine for more detail). The proxy works on the premise that as diatoms grow in a “parcel” of seawater and progressively use up the available DSi, both the remaining DSi and the bSiO2 produced at any instant become progressively isotopically heavier (see section Marine for detail). The fractionation can be modeled, assuming either closed or open distillation of Si isotopes, allowing the back-calculation of past utilization from downcore diatom δ30Si archives (De La Rocha et al., 1997, 1998; Varela et al., 2004). The calculation assumes a known and constant fractionation factor, denoted by epsilon (see section Stable Silicon Isotope Ratios to Study the Global Si Cycle).
Similar to marine work (Varela et al., 2004; Closset et al., 2015), lake sediment traps have shown the δ30Si signature of diatoms (δ30Sidiatom) to be resilient to dissolution and preserved through the water column into the sediment record (Panizzo et al., 2016). Accordingly, δ30Sidiatom from lacustrine sediment cores can be used to reconstruct past changes in biogeochemical cycling at the catchment/drainage basin scale. Whilst only a limited number of palaeolimnological studies have been undertaken, all demonstrate the potential for δ30Sidiatom to constrain long-term changes in the Si cycling in relation to catchment processes and/or physical limnological. For example, a record from Arctic Siberia covering the last 31 ka demonstrates that rates of DSi utilization are governed by catchment weathering and ice-cover duration together and mixing in the water column (Swann et al., 2010). Elsewhere, records from East Africa demonstrate the importance of monsoonal rainfall and their associated impact on vegetation in regulating the Si cycle. In the upper White Nile basin results reveal that higher monsoonal rainfall and associated increases in forest cover and chemical weathering increased the flux of DSi from land to rivers during the late-glacial/mid-Holocene, conditions that reversed after 5.5 ka with the emergence of dryer conditions and open vegetation/crops (Cockerton et al., 2015). Further east in Kenya measurements of δ30Sidiatom in a lake demonstrate that high levels of glacial diatom productivity were supported by increased fluxes of Si from the sparse catchment (Street-Perrott et al., 2008). Subsequent reductions in aquatic productivity then coincide with the development of grasslands and other Holocene vegetation, in response to increase monsoonal rainfall, which reduced Si transportation to the lake (Street-Perrott et al., 2008).
The potential of δ30Si is particularly evident in regions where climate change and anthropogenic lake catchment alteration (e.g., nutrient loading) are prompting limnological responses (e.g., increased duration of lake stratification and warmer surface water temperatures), which impact within lake biogeochemical cycling (Panizzo et al., 2017). For example, the approach has been demonstrated as a means to identify biogeochemical responses to climate change (Street-Perrott et al., 2008; Swann et al., 2010) and its value over more recent timescales is also stressed here as a means to address current pressures. Comprehensive contemporary limnological monitoring is also emphasized (Opfergelt et al., 2011) to enable quantitative estimations of seasonal DSi and bSiO2 cycling (via open or closed system modeling), by providing key constraints on deep-surface water δ30SiDSi exchange, Si residency times and Si uptake (Panizzo et al., 2016, 2017). These data also serve to validate conventional palaeolimnological approaches, which can be applied to constrain continental bSiO2 export from lake and reservoirs, particularly under a projected future of enhanced anthropogenic and climatic pressures on these systems.
Siliceous sponges
Unlike diatoms, there is evidence that siliceous sponges have a variable fractionation of Si isotopes with respect to seawater. A compilation of modern sponge spicules, and core top spicules, reveals that there is a statistically significant non-linear relationship between sponge δ30Si, or the difference between seawater and spicule δ30Si (denoted by Δ30Si), and ambient DSi concentrations during sponge growth (Hendry et al., 2010; Wille et al., 2010; Hendry and Robinson, 2012). This empirical calibration means that there is a great potential for spicule δ30Si in sediment cores to be used as a proxy for bottom water DSi concentrations in the past, providing key information about the supply of waters that feed diatom productivity (Hendry et al., 2010; Hendry and Robinson, 2012). The relationship is robust between laboratories (Hendry et al., 2011), and whilst the mechanism behind this relationship remains unclear, the observations are consistent with a strong growth-rate dependence on DSi uptake in sponges (Wille et al., 2010; Hendry and Robinson, 2012). The relationship between DSi and sponge δ30Si appears to hold for different filter-feeding sponges from different taxonomic groupings and ocean basins, growing in significantly different temperature, salinity and pH conditions (Hendry and Robinson, 2012). Furthermore, spicules from core top sediments also fall on the same calibration line, indicating minimal impact of early diagenetic processes (Hendry and Robinson, 2012). However, unusual biomineralization processes and potentially the internalization and isolation of the aquiferous system in highly-derived carnivorous sponges result in δ30Si values that deviate significantly from the global compilation (Hendry et al., 2015). Further investigation into different biomineralization pathways—and their impacts on isotopic fractionation—would be beneficial. Studies of isotopic fractionation by sponges grown in culture will add a great deal to our understanding of spicule geochemistry and biomineralization mechanisms.
Radiolaria
There have been fewer studies into the stable isotope composition of radiolarians compared to diatoms and sponges, and their use as palaeoceanographic archives is very much in its infancy. Radiolarian Si and oxygen isotope fractionation factors are challenging to constrain from culture studies because they are difficult to grow under laboratory conditions and field studies are limited by knowledge regarding the migration, strong biogeographic distributions and relatively sparse numbers (Suzuki and Aita, 2011). However, there have been a small number of studies that have attempted to constrain a fractionation factor for Si isotopes in radiolarians, which have estimated a similar fractionation range as for modern diatoms (ca. −0.5 to −2.1‰; Hendry et al., 2014; Abelmann et al., 2015). As such, there is potential for radiolarian δ30Si to be used, in combination with modeling efforts, to constrain mid-depth to surface DSi systematics. In short, more sediment records of δ30Si in sponges and radiolarians are needed to estimate nutrient concentrations and distribution with reorganizations in ocean circulation.
Multi-proxy Geochemical Approaches in Palaeoceanography
The Southern Ocean plays a key role in climate variability, via heat transport and atmospheric greenhouse gas changes, over glacial-interglacial timescales through both physical and biogeochemical mechanisms. One such mechanism to explain at least part of the atmospheric CO2 changes observed in ice core records is the Silicic Acid Leakage Hypothesis SALH (Brzezinski et al., 2002; Matsumoto et al., 2002). The SALH proposes that enhanced dust deposition in the Southern Ocean during glacial periods causes physiological changes in diatoms, such that they take up less DSi relative to other nutrients (i.e., N, P), resulting in the export of DSi enriched waters via AAIW (Ellwood et al., 2010; Rousseau et al., 2016). Supply of relatively DSi-rich Mode Waters at lower latitude would enhance the growth of diatoms over carbonate producers, altering ocean alkalinity leading to a reduction in atmospheric CO2 (Matsumoto and Sarmiento, 2008).
Geochemical archives locked up in bSiO2 can be used to test the SALH. A recent example, used both oxygen and Si isotopes in diatoms and radiolarians, from glacial-aged sediments from the Southern Ocean, to provide reconstructions of seasonal nutrient cycling. Data show strong variability in the mixed layer depth in the seasonal sea-ice zone, which allowed for sufficient nutrient exchange between surface and deeper waters, thereby fueling carbon drawdown in an otherwise highly stratified glacial Southern Ocean (Abelmann et al., 2015). Diatom δ30Si and organic-bound δ15N archives from the Southern Ocean have also been coupled with a spicule δ30Si record of bottom water DSi concentrations to investigate DSi utilization across the last glacial termination. By modeling the diatom Si-uptake, assuming the spicules reflect the supply of DSi, these archives demonstrate periods of intense upwelling and a greater utilization of the DSi supply in response to changes in iron availability during the deglaciation period (Horn et al., 2011).
In addition to using the relative bSiO2 accumulation rates between the Southern Ocean and lower latitudes as a test for Si export (e.g., Bradtmiller et al., 2009; Meckler et al., 2013), it is possible to use combined geochemical proxy records to test the SALH (Rousseau et al., 2016). Biogenic SiO2 archives point toward abrupt shifts in productivity and DSi utilization at glacial terminations, rather than changes on glacial-interglacial timescales (e.g., using coupled spicule, hand-picked radiolarian and diatoms from archives in the Sargasso Sea; Hendry and Brzezinski, 2014). Such observations, and other geochemical records, have led to the formulation of a new Silicic Acid Ventilation Hypothesis, which posits that sluggish glacial Southern Ocean overturning primed southern sourced Mode Waters to cause major changes in nutrient distribution during periods of abrupt climate change in low latitude regions influenced by intense ventilation (Hendry and Brzezinski, 2014; Hendry et al., 2016).
Challenges for Palaeo-Record Interpretation
Cleaning of bSiO2
The cleaning of bSiO2 remains an important challenge in δ30SibSiO2, δ18ObSiO2, δ13Cdiatom, and δ15Ndiatom reconstructions. Contamination, via the presence of residual tephras or clays can compromise the precision of reconstructions, with the introduction of significant isotopic offsets (Morley et al., 2005; Lamb et al., 2007; Brewer et al., 2008). This is often more pronounced for diatoms (and radiolarians; Abelmann et al., 2015) due to the ability for contaminant valve adherence and occlusion, over sponge spicules that can be picked. Clay δ30Si compositions are estimated between −2.95‰ and +2.5 (Douthitt, 1982; Georg et al., 2009; Opfergelt and Delmelle, 2012), which can significantly lower reported δ30Sidiatom values [published values range between −0.07 and +3.05‰; (De La Rocha et al., 2000; Cardinal et al., 2007; Sun et al., 2013; Panizzo et al., 2014, 2016)] and outside of analytical uncertainty, while high sample presence of different phases of bSiO2 can have a similar effect (e.g., δ30Sisponge signatures vary between −5.72 and +0.87‰ Douthitt, 1982; De La Rocha, 2003; Wille et al., 2010; Hendry and Robinson, 2012). As a result, purity is routinely demonstrated visually (via scanning electron microscopy) and via quantitative assessments of sample geochemistry (e.g., Al2O3) (Brewer et al., 2008; Chapligin et al., 2012). These serve, in the first instance, to demonstrate the efficiency of cleaning (SiO2:Al2O3 >1) and robustness of reconstructions. When not possible, these data permit contamination mass-balance calculations and a compositional offset correction of reported data (Brewer et al., 2008; Mackay et al., 2011, 2013; Wilson et al., 2014). There have been a number of methodological advancements following the earliest published bSiO2 cleaning protocols (Labeyrie and Juillet, 1982; Shemesh et al., 1988, 1995; Morley et al., 2004), which demonstrate that in order to address the challenges in bSiO2 cleaning, methods should be tailored depending on the characteristics of individual samples and frequent visual inspection should be carried out throughout.
Constant Silicon Isotope Fractionation Factor
Application of the δ30Si proxy for DSi utilization, assumes a constant fractionation factor (see section Stable Silicon Isotope Ratios to Study the Global Si Cycle). This has been challenged with evidence of species-dependent δ30Si fractionation variability between DSi and bSiO2 phases (Sutton et al., 2013). The work of Sutton et al. (2013) suggests that interpretation of marine sediment-based reconstructions, should include an evaluation of species composition for more accurate interpretations of δ30Si downcore variability. While these data shed light on potential species effects, each diatom strain in the Sutton et al. (2013) study was grown under its specific optimal environmental conditions in order to minimize the influence of external environmental factors (e.g., irradiance and/or temperature). The influence of environment on δ30Si composition in diatoms has yet to be shown under laboratory controlled conditions. Data from in-situ/natural settings, have not documented a species effect (e.g., Cardinal et al., 2007; Fripiat et al., 2012; Closset et al., 2015) although if anything this clearly demonstrates the need to better understand the variability of δ30Si in diatoms.
Similarly, it is essential to take any different ecological preferences of the diatoms present in the sediment core into consideration. There may be significant differences in when and where particular dominant species are growing, or if there is a pronounced preservation bias toward more heavily silicified species. For example, late season diatoms may bloom in already modified surface waters and large, deep-dwelling diatoms may opportunistically utilize DSi supplies that are hundreds of meters deep that has a different δ30SiDSi compared to near surface waters (Hendry and Brzezinski, 2014; Xiong et al., 2015). Another example is in Sea-Ice Zone of the Southern Ocean, where diatoms living within the sea-ice bear heavier δ30Si than diatoms in the surrounding waters due to a closed system where silicic acid can be limited (Fripiat et al., 2007; Panizzo et al., 2014).
Application of δ18O as a Palaeoceanographic Proxy
Despite an extensive effort by many researchers over the past five decades (e.g., Mopper and Garlick, 1971; Labeyrie, 1979; Labeyrie and Juillet, 1982; Chung-Ho and Hsueh-Wen, 1985; Leclerc and Labeyrie, 1987; Matheney and Knauth, 1989; Shemesh et al., 1992; Swann and Leng, 2009; Pike et al., 2013; Crespin et al., 2014; Abelmann et al., 2015), uncertainty associated with the bSiO2-water oxygen isotope fractionation has limited the application of bSiO2 δ18O values as a palaeoceanographic proxy. The disparate bSiO2-water fractionation factors have been attributed to two primary causes: (1) methodological biased and/or incomplete removal of hydroxyl oxygen, and (2) potential alteration of δ18O values during bSiO2 formation/diagenetic alteration of δ18O values on geologic timescales. The analytical bias has been addressed thorough an inter-laboratory comparison that showed no significant difference in δ18O values across a range of dehydration and analytical techniques (Chapligin et al., 2011); therefore, the only remaining source of uncertainty is in the bSiO2-water fractionation relationship recorded by the diatom bSiO2.
Diatoms from laboratory cultures and marine sediment traps seem to have a reproducible bSiO2-water fractionation relationship (Matheney and Knauth, 1989; Brandriss et al., 1998; Schmidt et al., 2001; Dodd et al., 2017) that is different from the bSiO2-water fractionation relationship recorded by sedimentary diatoms and quartz-water equilibrium (e.g., Leclerc and Labeyrie, 1987; Matheney and Knauth, 1989; Schmidt et al., 2001). Even within sedimentary marine diatom bSiO2 archives, there does not appear to be a single bSiO2-water fractionation relationship. Leclerc and Labeyrie (1987) published a bSiO2-water fractionation for marine diatoms from core-top sediments and surface waters (δ18Owater and T); however, within their dataset, δ18O values of diatom bSiO2 from high-latitude cores overestimated surface water temperatures by several °C. Shemesh et al. (1992) proposed the high-latitude diatoms represented a fundamentally different bSiO2-water fractionation factor; however, a recent re-examination of bSiO2-water fractionation factors over the entire range of formation temperatures (0–850°C) suggest that marine diatoms may record the same bSiO2-water fractionation relationship has high-temperature opal/quartz (Sharp et al., 2016).
A uniform bSiO2-water fractionation factor implies that living diatoms most likely precipitate bSiO2 out of isotopic equilibrium (e.g., Brandriss et al., 1998; Dodd et al., 2017); however, sedimentary (aka mature) diatom bSiO2 most likely approaches quartz-water equilibrium (Sharp et al., 2016). It is, therefore, likely that the δ18O values of diatom bSiO2 record a combination of diagenetic (e.g., sedimentary pore water) conditions. Further work is needed to establish the timing, degree, and mechanism of diagenetic changes to diatom δ18O values; however, experimental studies suggest a combination of dehydroxylation and precipitation of abiogenic SiO2 (e.g., Dodd et al., 2017).
Conclusions and Perspectives
The bio-geochemical analyses used to study the global biogeochemical cycling of Si (e.g., stable isotopes and associated trace elements), are emerging as useful tools to examine the influence that silicifying organisms have on the different reservoirs of Si (e.g., atmospheric, terrestrial, freshwater, and marine). Over the past 20 years, the ongoing development of specific low temperature bio-geochemical tools (e.g., δ30Si in silicifying organisms) has provided useful information for the reconstruction of past Si cycling and hints at the influence that environmental change can have on the global Si cycle. In addition, these tools provide knowledge on the role that silicifying organisms have on other major biogeochemical cycles (e.g., N and C). As outlined in this review, the main requirements for the future development of the bio-geochemical tools used to evaluate the global Si cycle are to:
1. Continue to produce high quality data (e.g., sample preparation, chemical procedure, mass spectrometric analysis, data analysis, the development of robust models, and the need for more data-model comparisons).
2. Use multiple bio-geochemical tools simultaneously, i.e., a single isotope system is not perfect and should be coupled with other bio-geochemical proxies (e.g., Ge/Si) and parameters (e.g., salinity, temperature).
3. Better understand what controls the fractionation factors of the various bio-geochemical tools used to study the role of siliceous organisms (e.g., sponges, diatoms, radiolaria, picophytoplankton) and globally important processes (e.g., reverse weathering, dissolution) on the global fluxes of Si.
4. Provide information on complex (though key) systems, e.g., estuaries and coastal environments, continental seas (e.g., the Arctic Ocean), sediment diagenesis (anoxic vs. bio-active), hydrothermal activity, soil systems, and particle–water interactions.
The global Si cycle starts with the chemical weathering of silicate minerals, is transformed and re-distributed into lakes, rivers, terrestrial and freshwater organisms, soils, aerosols, seawater marine organisms, and sediment, and eventually becomes a mineral once again. During these various transformations, Si interacts with numerous other major (e.g., C, N) and minor (e.g., Al, Ge, Zn) elements and, in turn, influences their biogeochemical cycles. Investigation of the movement, transformation, and fractionation of the stable isotopes and associated elements involved in the bio-geochemical cycling of Si provides knowledge not only to help constrain the distribution and behavior of the global Si fluxes and their potential variability over time, but also provides knowledge on the mechanisms of Si biomineralization. This information is essential for understanding the short- and long-term variation in the range of these data, which are used to evaluate the utility of these bio-geochemical tools for palaeoceanographic interpretation.
Author Contributions
JS and KH conceived of the project. JS drafted the manuscript with substantial contributions to the work by all authors. The manuscript was edited and approved by all authors.
Funding
The work by JS was supported by the “Laboratoire d'Excellence” LabexMER (ANR-10-LABX-19) and co-funded by a grant from the French government under the program “Investissements d'Avenir,” and by a grant from the Regional Council of Brittany (SAD programme). DJC was partially supported by the Knut and Alice Wallenberg Foundation (KAW Wallenberg Scholar) and the Swedish Research Council. This review article has benefited from funding by the European Union Seventh Framework Programme under grant agreement n°294146 (project MuSiCC, Marie Curie CIG to DC). GdS is supported by a Marie Skłodowska-Curie Research Fellowship under EU Horizon2020 (GA #708407). JuD was supported by the American Chemical Society Petroleum Research Fund (Grant # 53798-DNI2). CE acknowledges financial support by the Institute for Chemistry and Biology of the Marine Environment (Oldenburg, Germany) and the Max Planck Institute for Marine Microbiology (Bremen, Germany). KH is funded by The Royal Society (UF120084) and the European Research Council (ERC-2015-StG - 678371_ICY-LAB). PG acknowledges funding by the Collaborative Research Centre 754 “Climate-Biogeochemistry interactions in the Tropical Ocean” (www.sfb754.de), supported by the Deutsche Forschungsgemeinschaft (DFG).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling editor declared a shared affiliation, though no other collaboration, with one of the authors, JS.
Acknowledgments
We thank the organizers of Silicamics and the Silica Group (UMR 6539 CNRS/UBO/IRD/Ifremer, LEMAR - IUEM - 29280 - Plouzané, France) for providing the initiative to write this review. We also thank J. Martinez, of GraphicProtype, for his conceptual and illustrative help with the main figure (Figure 1).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/feart.2017.00112/full#supplementary-material
References
Abelmann, A., Gersonde, R., Knorr, G., Zhang, X., Chapligin, B., Maier, E., et al. (2015). The seasonal sea-ice zone in the glacial Southern Ocean as a carbon sink. Nat. Commun. 6:8136. doi: 10.1038/ncomms9136
Alleman, L. Y., Cardinal, D., Cocquyt, C., Plisnier, P.-D. P.-D., Descy, J.-P. J.-P., Kimirei, I., et al. (2005). Silicon isotope fractionation in Lake Tanganyka and its main tributaries. J. Great Lakes Res. 31, 509–519. doi: 10.1016/S0380-1330(05)70280-X
Aller, R. C. (2014). “Sedimentary diagenesis, depositional environments, and benthic fluxes,” in Treatise on Geochemistry, Second Edition, Vol. 8, eds H. D. Holland and K. K. Turekian (Oxford: Elsevier), 293–334.
Altabet, M. A., Deuser, W. G., Honjo, S., and Stienen, C. (1991). Seasonal and depth-related changes in the source of sinking particles in the North Atlantic. Nature 354, 136–139. doi: 10.1038/354136a0
Andersen, M. B., Vance, D., Archer, C., Anderson, R. F., Ellwood, M. J., and Allen, C. S. (2011). The Zn abundance and isotopic composition of diatom frustules, a proxy for Zn availability in ocean surface seawater. Earth Planet. Sci. Lett. 301, 137–145. doi: 10.1016/j.epsl.2010.10.032
André, L., Cardinal, D., Alleman, L. Y., and Moorbath, S. (2006). Silicon isotopes in ~ 3. 8 Ga West Greenland rocks as clues to the Eoarchaean supracrustal Si cycle. Earth Planet. Sci. Lett. 245, 162–173. doi: 10.1016/j.epsl.2006.02.046
Baars, O., and Croot, P. L. (2011). The speciation of dissolved zinc in the Atlantic sector of the Southern Ocean. Deep Sea Res. II Top. Stud. Oceanogr. 58, 2720–2732. doi: 10.1016/j.dsr2.2011.02.003
Baines, S. B., Twining, B. S., Brzezinski, M. A., Nelson, D. M., and Fisher, N. S. (2010). Causes and biogeochemical implications of regional differences in silicification of marine diatoms. Glob. Biogeochem. Cyc. 24:GB4031.
Barão, L., Vandevenne, F., Clymans, W., Frings, P., Ragueneau, O., Meire, P., et al. (2015). Alkaline-extractable silicon from land to ocean: a challenge for biogenic silicon determination. Limnol. Oceanogr. Methods 13, 329–344. doi: 10.1002/lom3.10028
Bareille, G., Labracherie, M., Mortlock, R. A., Maier-Reimer, E., and Froelich, P. N. (1998). A test of (Ge/Si)(opal) as a paleorecorder of (Ge/Si)(seawater). Geology 26, 179–182.
Barker, P. A., Hurrell, E. R., Leng, M. J., Plessen, B., Wolff, C., Conley, D. J., et al. (2013). Carbon cycling within an East African lake revealed by the carbon isotope composition of diatom silica: A 25-ka record from Lake Challa, Mt. Kilimanjaro. Quat. Sci. Rev. 66, 55–63. doi: 10.1016/j.quascirev.2012.07.016
Baronas, J. J., Hammond, D. E., Berelson, W. M., McManus, J., and Severmann, S. (2016). Germanium–silicon fractionation in a river-influenced continental margin: The Northern Gulf of Mexico. Geochim. Cosmochim. Acta 178, 124–142. doi: 10.1016/j.gca.2016.01.028
Bernard, C. Y., Laruelle, G. G., Slomp, C. P., and Heinze, C. (2010). Impact of changes in river fluxes of silica on the global marine silicon cycle: a model comparison. Biogeosciences 7, 441–453. doi: 10.5194/bg-7-441-2010
Berner, R. A., Lasaga, A. C., and Garrels, R. M. (1983). The carbonate-silicate geochemical cycle and its effect on atmospheric carbon dioxide over the past 100 million years. Am. J. Sci. 283, 641–683. doi: 10.2475/ajs.283.7.641
Beucher, C. P., Brzezinski, M. A., and Crosta, X. (2007). Silicic acid dynamics in the glacial sub-Antarctic: Implications for the silicic acid leakage hypothesis. Global Biogeochem. Cycles 21:GB3015. doi: 10.1029/2006GB002746
Beucher, C. P., Brzezinski, M. A., and Jones, J. L. (2008). Sources and biological fractionation of Silicon isotopes in the Eastern Equatorial Pacific. Geochim. Cosmochim. Acta 72, 3063–3073. doi: 10.1016/j.gca.2008.04.021
Beucher, C. P., Brzezinski, M. A., and Jones, J. L. (2011). Mechanisms controlling silicon isotope distribution in the Eastern Equatorial Pacific. Geochim. Cosmochim. Acta 75, 4286–4294. doi: 10.1016/j.gca.2011.05.024
Bidle, K. D., and Azam, F. (1999). Accelerated dissolution of diatom silica by marine bacterial assemblages. Nature 397, 508–512. doi: 10.1038/17351
Bidle, K. D., Manganelli, M., and Azam, F. (2002). Regulation of oceanic silicon and carbon preservation by temperature control on bacteria. Science 298, 1980–1984. doi: 10.1126/science.1076076
Bouchez, J., Von Blanckenburg, F., and Schuessler, J. A. (2013). Modeling novel stable isotope ratios in the weathering zone. Am. J. Sci. 313, 267–308. doi: 10.2475/04.2013.01
Bradtmiller, L. I., Anderson, R. F., Fleisher, M. Q., and Burckle, L. H. (2009). Comparing glacial and Holocene opal fluxes in the Pacific sector of the Southern Ocean. Paleoceanography 24, 1–20. doi: 10.1029/2008PA001693
Brady, P. V., and Carroll, S. A. (1994). Direct effects of CO2 and temperature on silicate weathering: Possible implications for climate control. Geochim. Cosmochim. Acta 58, 1853–1856. doi: 10.1016/0016-7037(94)90543-6
Brandriss, M. E., O'Neil, J. R., Edlund, M. B., and Stoermer, E. F. (1998). Oxygen isotope fractionation between diatomaceous silica and water. Geochim. Cosmochim. Acta 62, 1119–1125.
Brewer, T. S., Leng, M. J., Mackay, A. W., Lamb, A. L., Tyler, J. J., and Marsh, N. G. (2008). Unravelling contamination during MIS-10 to 12: Implications for atmospheric CO2. Clim. Past 4, 333–344. doi: 10.5194/cp-4-333-2008
Brümmer, F. (2003). “Living inside a glass box – silica in diatoms,” in Progress in Molecular and Subcellular Biology, ed W. E. G. Müller (Berlin; Heidelberg: Springer-Verlag), 3–9.
Brunner, E., Gröger, C., Lutz, K., Richthammer, P., Spinde, K., and Sumper, M. (2009). Analytical studies of silica biomineralization: towards an understanding of silica processing by diatoms. Appl. Microbiol. Biotechnol. 84, 607–616. doi: 10.1007/s00253-009-2140-3
Brzezinski, M. A., and Jones, J. L. (2015). Coupling of the distribution of silicon isotopes to the meridional overturning circulation of the North Atlantic Ocean. Deep Sea Res. II Top. Stud. Oceanogr. 116, 79–88. doi: 10.1016/j.dsr2.2014.11.015
Brzezinski, M. A., Pride, C. J., Franck, V. M., Sigman, D. M., Sarmiento, J. L., Matsumoto, K., et al. (2002). A switch from Si(OH)(4) to NO3- depletion in the glacial Southern Ocean. Geophys. Res. Lett. 29, 1–4. doi: 10.1029/2001GL014349
Buesseler, K. O. (1998). The decoupling of production and partinulate export in the surface ocean. Global Biogeochem. Cycles 12, 297–310. doi: 10.1029/97GB03366
Cardinal, D., Alleman, L. Y., Dehairs, F., Savoye, N., Trull, T. W., and Andre, L. (2005). Relevance of silicon isotopes to Si-nutrient utilization and Si-source assessment in Antarctic waters. Global Biogeochem. Cycles 19:GB2007. doi: 10.1029/2004GB002364
Cardinal, D., Gaillardet, J., Hughes, H. J., Opfergelt, S., and André, L. (2010). Contrasting silicon isotope signatures in rivers from the Congo Basin and the specific behaviour of organic-rich waters. Geophys. Res. Lett. 37:L12403. doi: 10.1029/2010GL043413
Cardinal, D., Savoye, N., Trull, T. W., Dehairs, F., Kopczynska, E. E., Fripiat, F., et al. (2007). Silicon isotopes in spring Southern Ocean diatoms: Large zonal changes despite homogeneity among size fractions. Mar. Chem. 106, 46–62. doi: 10.1016/j.marchem.2006.04.006
Carey, J. C., and Fulweiler, R. W. (2012). The terrestrial silica pump. PLoS ONE 7:e52932. doi: 10.1371/journal.pone.0052932
Cavagna, A.-J., Fripiat, F., Dehairs, F., Wolf-Gladrow, D., Cisewski, B., Savoye, N., et al. (2011). Silicon uptake and supply during a Southern Ocean iron fertilization experiment (EIFEX) tracked by Si isotopes. Limnol. Oceanogr. 56, 147–160. doi: 10.4319/lo.2011.56.1.0147
Cermeño, P., Falkowski, P. G., Romero, O. E., Schaller, M. F., and Vallina, S. M. (2015). Continental erosion and the Cenozoic rise of marine diatoms. Proc. Natl. Acad. Sci. U.S.A. 112, 4239–4244. doi: 10.1073/pnas.1412883112
Cha, J. N. N., Shimizu, K., Zhou, Y., Christiansen, S. C. C., Chmelka, B. F. F., Stucky, G. D. D., et al. (1999). Silicatein filaments and subunits from a marine sponge direct the polymerization of silica and silicones in vitro. Proc. Natl. Acad. Sci. U.S.A. 96, 361–365, doi: 10.1073/pnas.96.2.361
Chakrabarti, R., Knoll, A. H., Jacobsen, S. B., and Fischer, W. W. (2012). Si isotope variability in Proterozoic cherts. Geochimica et Cosmochimica Acta 91, 187–201. doi: 10.1016/j.gca.2012.05.025
Chapligin, B., Leng, M. J., Webb, E., Alexandre, A., Dodd, J. P., Ijiri, A., et al. (2011). Inter-laboratory comparison of oxygen isotope compositions from biogenic silica. Geochim. Cosmochim. Acta 75, 7242–7256. doi: 10.1016/j.gca.2011.08.011
Chapligin, B., Meyer, H., Swann, G. E. A., Meyer-Jacob, C., and Hubberten, H. W. (2012). A 250 ka oxygen isotope record from diatoms at Lake El'gygytgyn, far east Russian Arctic. Clim. Past 8, 1621–1636. doi: 10.5194/cp-8-1621-2012
Chung-Ho, W., and Hsueh-Wen, Y. (1985). Oxygen isotopic compositions of DSDP Site 480 diatoms: Implications and applications. Geochim. Cosmochim. Acta 49, 1469–1478. doi: 10.1016/0016-7037(85)90296-0
Claquin, P., Martin-Jezequel, V., Kromkamp, J. C., Veldhuis, M. J. W., and Kraay, G. W. (2002). Uncoupling of silicon compared with carbon and nitrogen metabolisms and the role of the cell cycle in continuous cultures of Thalassiosira Pseudonana (Bacillariophyceae) under light, nitrogen, and phosphorus control. J. Phycol. 38, 922–930. doi: 10.1046/j.1529-8817.2002.t01-1-01220.x
Closset, I., Cardinal, D., Bray, S. G., Thil, F., Djouraev, I., Rigual-Hernández, A. S., et al. (2015). Seasonal variations, origin, and fate of settling diatoms in the Southern Ocean tracked by silicon isotope records in deep sediment traps. Glob. Biogeochem. Cycles 29, 1495–1510f. doi: 10.1002/2015GB005180
Clymans, W., Struyf, E., Govers, G., Vandevenne, F., and Conley, D. J. (2011). Anthropogenic impact on amorphous silica pools in temperate soils. Biogeosciences 8, 2281–2293. doi: 10.5194/bg-8-2281-2011
Cockerton, H. E., Street-Perrott, F. A., Barker, P. A., Leng, M. J., Sloane, H. J., and Ficken, K. J. (2015). Orbital forcing of glacial/interglacial variations in chemical weathering and silicon cycling within the upper White Nile basin, East Africa: stable-isotope and biomarker evidence from Lakes Victoria and Edward. Quat. Sci. Rev. 130, 57–71. doi: 10.1016/j.quascirev.2015.07.028
Cockerton, H. E., Street-Perrott, F. A., Leng, M. J., Barker, P. A., Horstwood, M. S. A., and Pashley, V. (2013). Stable-isotope (H, O, and Si) evidence for seasonal variations in hydrology and Si cycling from modern waters in the Nile Basin: Implications for interpreting the quaternary record. Quat. Sci. Rev. 66, 4–21. doi: 10.1016/j.quascirev.2012.12.005
Conley, D. J. (2002). Terrestrial ecosystems and the global biogeochemical silica cycle. Global Biogeochem. Cycles 16, 1–8. doi: 10.1029/2002GB001894
Conley, D. J., Frings, P. J., Fontorbe, G., Clymans, W., Stadmark, J., Hendry, K. R., et al. (2017). Biosilicification drives a decline of dissolved Si in the oceans through geologic time. Front. Mar. Sci. 4:397. doi: 10.3389/fmars.2017.00397
Conley, D. J., Schelske, C. L., and Stoermer, E. F. (1993). Modification of the biogeochemical cycle of silica with eutrophication. Mar. Ecol. Prog. Ser. 101, 179–192. doi: 10.3354/meps101179
Coogan, L. A., and Dosso, S. (2012). An internally consistent, probabilistic, determination of ridge-axis hydrothermal fluxes from basalt-hosted systems. Earth Planet. Sci. Lett. 323–324, 92–101. doi: 10.1016/j.epsl.2012.01.017
Cornelis, J. T., Delvaux, B., Cardinal, D., André, L., Ranger, J., and Opfergelt, S. (2010). Tracing mechanisms controlling the release of dissolved silicon in forest soil solutions using Si isotopes and Ge/Si ratios. Geochim. Cosmochim. Acta 74, 3913–3924. doi: 10.1016/j.gca.2010.04.056
Crespin, J., Yam, R., Crosta, X., Mass,é, G., Schmidt, S., Campagne, P., et al. (2014). Holocene glacial discharge fluctuations and recent instability in East Antarctica. Earth Planet. Sci. Lett. 394, 38–47. doi: 10.1016/j.epsl.2014.03.009
Crosta, X., and Shemesh, A. (2002). Reconciling down core anticorrelation of diatom carbon and nitrogen isotopic ratios from the Southern Ocean. Paleoceanography 17, 10-1–10-8. doi: 10.1029/2000PA000565
De La Rocha, C. L. (2003). Silicon isotope fractionation by marine sponges and the reconstruction of the silicon isotope composition of ancient deep water. Geology 31, 423–426. doi: 10.1130/0091-7613(2003)031<0423:SIFBMS>2.0.CO;2
De La Rocha, C. L., Brzezinski, M. A., and DeNiro, M. J. (1997). Fractionation of silicon isotopes by marine diatoms during biogenic silica formation. Geochim. Cosmochim. Acta 61, 5051–5056. doi: 10.1016/S0016-7037(97)00300-1
De La Rocha, C. L., Brzezinski, M. A., and DeNiro, M. J. (2000). A first look at the distribution of the stable isotopes of silicon in natural waters. Geochim. Cosmochim. Acta 64, 2467–2477. doi: 10.1016/S0016-7037(00)00373-2
De La Rocha, C. L., Brzezinski, M. A., DeNiro, M. J., and Shemesh, A. (1998). Silicon-isotope composition of diatoms as an indicator of past oceanic change. Nature 395, 680–683. doi: 10.1038/27174
de Souza, G. F., Reynolds, B. C., Johnson, G. C., Bullister, J. L., and Bourdon, B. (2012a). Silicon stable isotope distribution traces Southern Ocean export of Si to the eastern South Pacific thermocline. Biogeosciences 9, 4199–4213. doi: 10.5194/bg-9-4199-2012
de Souza, G. F., Reynolds, B. C., Rickli, J., Frank, M., Saito, M. A., Gerringa, L. J. A., et al. (2012b). Southern Ocean control of silicon stable isotope distribution in the deep Atlantic Ocean. Global Biogeochem. Cycles 26, 1–13. doi: 10.1029/2011GB004141
de Souza, G. F., Slater, R. D., Dunne, J. P., and Sarmiento, J. L. (2014). Deconvolving the controls on the deep ocean's silicon stable isotope distribution. Earth Planet. Sci. Lett. 398, 66–76. doi: 10.1016/j.epsl.2014.04.040
de Souza, G. F., Slater, R. D., Hain, M. P., Brzezinski, M. A., and Sarmiento, J. L. (2015). Distal and proximal controls on the silicon stable isotope signature of North Atlantic Deep Water. Earth Planet. Sci. Lett. 432, 342–353. doi: 10.1016/j.epsl.2015.10.025
Dellinger, M., Gaillardet, J., Bouchez, J., Calmels, D., Louvat, P., Dosseto, A., et al. (2015). Riverine Li isotope fractionation in the Amazon River basin controlled by the weathering regimes. Geochim. Cosmochim. Acta 164, 71–93. doi: 10.1016/j.gca.2015.04.042
Delstanche, S., Opfergelt, S., Cardinal, D., Elsass, F., André, L., and Delvaux, B. (2009). Silicon isotopic fractionation during adsorption of aqueous monosilicic acid onto iron oxide. Geochim. Cosmochim. Acta 73, 923–934. doi: 10.1016/j.gca.2008.11.014
Delvaux, C., Cardinal, D., Carbonnel, V., Chou, L., Hughes, H. J., and André, L. (2013). Controls on riverine δ30Si signatures in a temperate watershed under high anthropogenic pressure (Scheldt - Belgium). J. Mar. Syst. 128, 40–51. doi: 10.1016/j.jmarsys.2013.01.004
Delvigne, C., Cardinal, D., Hofmann, A., and André, L. (2012). Stratigraphic changes of Ge/Si, REE+Y and silicon isotopes as insights into the deposition of a Mesoarchaean banded iron formation. Earth Planet. Sci. Lett. 355–356, 109–118. doi: 10.1016/j.epsl.2012.07.035
Delvigne, C., Opfergelt, S., Cardinal, D., Delvaux, B., and André, L. (2009). Distinct silicon and germanium pathways in the soil-plant system: evidence from banana and horsetail. J. Geophys. Res. 114, 1–11. doi: 10.1029/2008JG000899
Delvigne, C., Opfergelt, S., Cardinal, D., Hofmann, A., and André, L. (2016). Desilication in Archean weathering processes traced by silicon isotopes and Ge/Si ratios. Chem. Geol. 420, 139–147. doi: 10.1016/j.chemgeo.2015.11.007
Demarest, M. S., Brzezinski, M. A., and Beucher, C. P. (2009). Fractionation of silicon isotopes during biogenic silica dissolution. Geochim. Cosmochim. Acta 73, 5572–5583. doi: 10.1016/j.gca.2009.06.019
DeMaster, D. J. (1981). The supply and accumulation of silica in the marine-environment. Geochim. Cosmochim. Acta 45, 1715–1732. doi: 10.1016/0016-7037(81)90006-5
DeMaster, D. J. (2003). “The diagenesis of biogenic silica: chemical transformations occurring in the water column, seabed and crust,” in Treatise on Geochemistry, Vol. 7, eds H. D. Holland and K. K. Turekian (Oxford: Elsevier), 87–98. doi: 10.1016/B0-08-043751-6/07095-X
Derry, L. A., Kurtz, A. C., Ziegler, K., and Chadwick, O. A. (2005). Biological control of terrestrial silica cycling and export fluxes to watersheds. Nature 433, 728–731. doi: 10.1038/nature03299
Des Combes, H. J., Esper, O., De La Rocha, C. L., Abelmann, A., Gersonde, R., Yam, R., et al. (2008). Diatom δ13C, δ15N, and C/N since the last glacial maximum in the Southern Ocean: potential impact of species composition. Paleoceanography 23, 1–12. doi: 10.1029/2008PA001589
Ding, T. P., Gao, J. F., Tian, S. H., Wang, H. B., and Li, M. (2011). Silicon isotopic composition of dissolved silicon and suspended particulate matter in the Yellow River, China, with implications for the global silicon cycle. Geochim. Cosmochim. Acta 75, 6672–6689. doi: 10.1016/j.gca.2011.07.040
Ding, T. P., Ma, G. R., Shui, M. X., Wan, D. F., and Li, R. H. (2005). Silicon isotope study on rice plants from the Zhejiang province, China. Chem. Geol. 218, 41–50. doi: 10.1016/j.chemgeo.2005.01.018
Ding, T. P., Tian, S. H., Sun, L., Wu, L. H., Zhou, J. X., and Chen, Z. Y. (2008a). Silicon isotope fractionation between rice plants and nutrient solution and its significance to the study of the silicon cycle. Geochim. Cosmochim. Acta 72, 5600–5615. doi: 10.1016/j.gca.2008.09.006
Ding, T. P., Zhou, J. X., Wan, D. F., Chen, Z. Y., Wang, C. Y., and Zhang, F. (2008b). Silicon isotope fractionation in bamboo and its significance to the biogeochemical cycle of silicon. Geochim. Cosmochim. Acta 72, 1381–1395. doi: 10.1016/j.gca.2008.01.008
Ding, T., Wan, D., Wang, C., and Zhang, F. (2004). Silicon isotope compositions of dissolved silicon and suspended matter in the Yangtze River, China. Geochim. Cosmochim. Acta 68, 205–216. doi: 10.1016/S0016-7037(03)00264-3
Dixit, S., Van Cappellen, P., and Van Bennekom, A. J. (2001). Processes controlling solubility of biogenic silica and pore water build-up of silicic acid in marine sediments. Mar. Chem. 73, 333–352. doi: 10.1016/S0304-4203(00)00118-3
Dodd, J. P., Wiedenheft, W., and Schwartz, J. M. (2017). Dehydroxylation and diagenetic variations in diatom oxygen isotope values. Geochim. Cosmochim. Acta 199, 185–195. doi: 10.1016/j.gca.2016.11.034
Doering, K., Ehlert, C., Grasse, P., Crosta, X., Fleury, S., Frank, M., et al. (2016). Differences between mono-generic and mixed diatom silicon isotope compositions trace present and past nutrient utilisation off Peru. Geochim. Cosmochim. Acta 177, 30–47. doi: 10.1016/j.gca.2015.12.029
Douthitt, C. B. (1982). The geochemistry of the stable isotopes of silicon. Geochim. Cosmochim. Acta 46, 1449–1458. doi: 10.1016/0016-7037(82)90278-2
Egan, K. E., Rickaby, R. E. M., Hendry, K. R., and Halliday, A. N. (2013). Opening the gateways for diatoms primes Earth for Antarctic glaciation. Earth Planet. Sci. Lett. 375, 34–43. doi: 10.1016/j.epsl.2013.04.030
Egan, K. E., Rickaby, R. E. M., Leng, M. J., Hendry, K. R., Hermoso, M., Sloane, H. J., et al. (2012). Diatom silicon isotopes as a proxy for silicic acid utilisation: a Southern Ocean core top calibration. Geochim. Cosmochim. Acta 96, 174–192. doi: 10.1016/j.gca.2012.08.002
Ehlert, C., Doering, K., Wallmann, K., Scholz, F., Sommer, S., Grasse, P., et al. (2016). Stable silicon isotope signatures of marine pore waters – Biogenic opal dissolution versus authigenic clay mineral formation. Geochim. Cosmochim. Acta 191, 102–117. doi: 10.1016/j.gca.2016.07.022
Ehlert, C., Grasse, P., Mollier-vogel, E., Bo, T., Franz, J., Souza, G. F., Frank, M., et al. (2012). Factors controlling the silicon isotope distribution in waters and surface sediments of the Peruvian coastal upwelling. Geochim. Cosmochim. Acta 99, 128–145. doi: 10.1016/j.gca.2012.09.038
Ehlert, C., Grasse, P., and Frank, M. (2013). Changes in silicate utilisation and upwelling intensity off Peru since the Last Glacial Maximum–insights from silicon and neodymium isotopes. Quat. Sci. Rev. 72, 18–35. doi: 10.1016/j.quascirev.2013.04.013
Eiler, J. M., Bergquist, B., Bourg, I., Cartigny, P., Farquhar, J., Gagnon, A., et al. (2014). Frontiers of stable isotope geoscience. Chem. Geol. 372, 119–143. doi: 10.1016/j.chemgeo.2014.02.006
Elderfield, H., and Rickaby, R. (2000). Oceanic Cd/P ratio and nutrient utilization in the glacial Southern Ocean. Nature 405, 305–310. doi: 10.1038/35012507
Ellwood, M. J., and Hunter, K. A. (1999). Determination of the Zn/Si ratio in diatom opal: a method for the separation, cleaning and dissolution of diatoms. Mar. Chem. 66, 149–160. doi: 10.1016/S0304-4203(99)00037-7
Ellwood, M. J., and Hunter, K. A. (2000). The incorporation of zinc and iron into the frustule of the marine diatom Thalassiosira pseudonana. Limnol. Oceanogr. 45, 1517–1524. doi: 10.4319/lo.2000.45.7.1517
Ellwood, M. J., Kelly, M., Neil, H., and Nodder, S. D. (2005). Reconstruction of paleo-particulate organic carbon fluxes for the Campbell Plateau region of southern New Zealand using the zinc content of sponge spicules. Paleoceanography 20:PA3010. doi: 10.1029/2004PA001095
Ellwood, M. J., Kelly, M., Nodder, S. D., and Carter, L. (2004). Zinc/silicon ratios of sponges: a proxy for carbon export to the seafloor. Geophys. Res. Lett. 31:L12308. doi: 10.1029/2004GL019648
Ellwood, M. J., Wille, M., and Maher, W. (2010). Glacial silicic acid concentrations in the Southern Ocean. Science 330, 1088–1091. doi: 10.1126/science.1194614
Engström, E., Rodushkin, I., Ingri, J., Baxter, D. C., Ecke, F., Österlund, H., et al. (2010). Temporal isotopic variations of dissolved silicon in a pristine boreal river. Chem. Geol. 271, 142–152. doi: 10.1016/j.chemgeo.2010.01.005
Engström, E., Rodushkin, I., Öhlander, B., Ingri, J., and Baxter, D. C. (2008). Silicon isotopic composition of boreal forest vegetation in Northern Sweden. Chem. Geol. 257, 250–259. doi: 10.1016/j.chemgeo.2008.10.004
Escoube, R., Rouxel, O. J., Edwards, K., Glazer, B., and Donard, O. F. (2015). Coupled Ge/Si and Ge isotope ratios as geochemical tracers of seafloor hydrothermal systems: case studies at Loihi Seamount and East Pacific Rise 9 50′ N. Geochimica et Cosmochimica Acta 167, 93–112. doi: 10.1016/j.gca.2015.06.025
Fontorbe, G., De La Rocha, C. L., Chapman, H. J., and Bickle, M. J. (2013). The silicon isotopic composition of the Ganges and its tributaries. Earth Planet. Sci. Lett. 381, 21–30. doi: 10.1016/j.epsl.2013.08.026
Fontorbe, G., Frings, P. J., De La Rocha, C. L., Hendry, K. R., and Conley, D. J. (2016). A silicon depleted North Atlantic since the Palaeogene: Evidence from sponge and radiolarian silicon isotopes. Earth Planet. Sci. Lett. 453, 67–77. doi: 10.1016/j.epsl.2016.08.006
Fontorbe, G., Frings, P. J., De La Rocha, C. L., Hendry, K. R., and Conley, D. J. (2017). Enrichment of dissolved silica in the deep Equatorial Pacific during the Eocene-Oligocene. Paleoceanography 32, 848–863. doi: 10.1002/2017PA003090
Freudenthal, T., Wagner, T., Wenzhöfer, F., Zabel, M., and Wefer, G. (2001). Early diagenesis of organic matter from sediments of the eastern subtropical Atlantic: evidence from stable nitrogen and carbon isotopes. Geochim. Cosmochim. Acta 65, 1795–1808. doi: 10.1016/S0016-7037(01)00554-3
Frings, P. J., Clymans, W., Fontorbe, G., De La Rocha, C. L., and Conley, D. J. (2016). The continental Si cycle and its impact on the ocean Si isotope budget. Chem. Geol. 425, 12–36. doi: 10.1016/j.chemgeo.2016.01.020
Frings, P. J., Clymans, W., Fontorbe, G., Gray, W., Chakrapani, G., Conley, D. J., et al. (2015). Silicate weathering in the Ganges alluvial plain. Earth Planet. Sci. Lett. 427, 136–148. doi: 10.1016/j.epsl.2015.06.049
Frings, P. J., Clymans, W., Jeppesen, E., Lauridsen, T. L., Struyf, E., and Conley, D. J. (2014a). Lack of steady-state in the global biogeochemical Si cycle: Emerging evidence from lake Si sequestration. Biogeochemistry 117, 255–277. doi: 10.1007/s10533-013-9944-z
Frings, P. J., De La Rocha, C., Struyf, E., van Pelt, D., Schoelynck, J., Hudson, M. M., et al. (2014b). Tracing silicon cycling in the Okavango Delta, a sub-tropical flood-pulse wetland using silicon isotopes. Geochim. Cosmochim. Acta 142, 132–148. doi: 10.1016/j.gca.2014.07.007
Fripiat, F., Cardinal, D., Tison, J. L., Worby, A., and Andre, L. (2007). Diatom-induced silicon isotopic fractionation in Antarctic sea ice. J. Geophys. Res. 112:G02001. doi: 10.1029/2006JG000244
Fripiat, F., Cavagna, A.-J., Dehairs, F., Speich, S., André, L., and Cardinal, D. (2011). Silicon pool dynamics and biogenic silica export in the Southern Ocean inferred from Si-isotopes. Ocean Sci. 7, 533–547. doi: 10.5194/os-7-533-2011
Fripiat, F., Cavagna, A. J., Dehairs, F., De Brauwere, A., André, L., and Cardinal, D. (2012). Processes controlling the Si-isotopic composition in the Southern Ocean and application for paleoceanography. Biogeosciences 9, 2443–2457. doi: 10.5194/bg-9-2443-2012
Froelich, H., and Barthel, D. (1997). Silica uptake of the marine sponge Halichondria panicea in Kiel Bight. Mar. Biol. 128, 115–125. doi: 10.1007/s002270050075
Froelich, P. N., and Andreae, M. O. (1981). The marine geochemistry of Germanium: Ekasilicon. Science 213, 205–207. doi: 10.1126/science.213.4504.205
Froelich, P. N., Blanc, V., Mortlock, R. A., Chillrud, S. N., Dunstant, W., Udomkit, A., et al. (1992). River fluxes of dissolved silica to the ocean were higher during the glacials: Ge/Si in diatoms, rivers and oceans. Palaeoceanography 7, 739–768. doi: 10.1029/92PA02090
Froelich, P. N., Mortlock, R. A., and Shemesh, A. (1989). Inorganic germanium and silica in the Indian ocean: biological fractionation during (Ge/Si)opal formation. Global Biogeochem. Cycles 3, 79–88. doi: 10.1029/GB003i001p00079
Gao, S., Wolf-Gladrow, D. A., and Volker, C. (2016). Simulating the modern δ30Si distribution in the oceans. Glob. Biogeochem. Cycles 30, 120–133. doi: 10.1002/2015GB005189
Garcia, H. E., Locarnini, R. A., Boyer, T. P., Antonov, J. I., Baranova, O. K., Zweng, M. M., et al. (2014). “World Ocean Atlas 2013, Volume 4: dissolved inorganic nutrients (phosphate, nitrate, silicate),” in NOAA Atlas NESDIS, ed S. Levitus (Silver Spring, MD: NOAA-NESDIS), 25.
Gehlen, M., Beck, L., Calas, G., Flank, A. M., Van Bennekom, A. J., and Van Beusekom, J. E. E. (2002). Unraveling the atomic structure of biogenic silica: evidence of the structural association of Al and Si in diatom frustules. Geochim. Cosmochim. Acta 66, 1601–1609. doi: 10.1016/S0016-7037(01)00877-8
Geilert, S., Vroon, P. Z., Keller, N. S., Gudbrandsson, S., Stefánsson, A., and van Bergen, M. J. (2015). Silicon isotope fractionation during silica precipitation from hot-spring waters: evidence from the Geysir geothermal field, Iceland. Geochim. Cosmochim. Acta 164, 403–427. doi: 10.1016/j.gca.2015.05.043
Geilert, S., Vroon, P. Z., Roerdink, D. L., Van Cappellen, P., and van Bergen, M. J. (2014). Silicon isotope fractionation during abiotic silica precipitation at low temperatures: Inferences from flow-through experiments. Geochim. Cosmochim. Acta 142, 95–114. doi: 10.1016/j.gca.2014.07.003
Georg, R. B., Reynolds, B. C., Frank, M., and Halliday, A. N. (2006). Mechanisms controlling the silicon isotopic compositions of river waters. Earth Planet. Sci. Lett. 249, 290–306. doi: 10.1016/j.epsl.2006.07.006
Georg, R. B., Reynolds, B. C., West, A. J., Burton, K. W., and Halliday, A. N. (2007). Silicon isotope variations accompanying basalt weathering in Iceland. Earth Planet. Sci. Lett. 261, 476–490. doi: 10.1016/j.epsl.2007.07.004
Georg, R. B., Zhu, C., Reynolds, B. C., and Halliday, A. N. (2009). Stable silicon isotopes of groundwater, feldspars, and clay coatings in the Navajo Sandstone aquifer, Black Mesa, Arizona, USA. Geochim. Cosmochim. Acta 73, 2229–2241. doi: 10.1016/j.gca.2009.02.005
Grasse, P., Brzezinski, M., Cardinal, D., de Souza, G. F., Andersson, P., Closset, I., et al. (2017). GEOTRACES inter-calibration of the stable silicon isotope composition of dissolved silicic acid in seawater. J. Anal. At. Spectrom. 32, 562–578. doi: 10.1039/C6JA00302H
Grasse, P., Ehlert, C., and Frank, M. (2013). The influence of water mass mixing on the dissolved Si isotope composition in the Eastern Equatorial Pacific. Earth Planet. Sci. Lett. 380, 60–71. doi: 10.1016/j.epsl.2013.07.033
Hammond, D. E., McManus, J., and Berelson, W. M. (2004). Oceanic germanium/silicon ratios: Evaluation of the potential overprint of temperature on weathering signals. Paleoceanography 19:PA2016. doi: 10.1029/2003PA000940
Hammond, D. E., McManus, J., Berelson, W. M., Meredith, C., Klinkhammer, G. P., and Coale, K. H. (2000). Diagenetic fractionation of Ge and Si in reducing sediments: the missing Ge sink and a possible mechanism to cause glacial/interglacial variations in oceanic Ge/Si. Geochim. Cosmochim. Acta 64, 2453–2465. doi: 10.1016/S0016-7037(00)00362-8
Hawkings, J. R., Wadham, J. L., Benning, L. G., Hendry, K. R., Tranter, M., Tedstone, A., et al. (2017). Ice sheets as a missing source of silica to the polar oceans. Nat. Commun. 8:14198. doi: 10.1038/ncomms14198
Heck, P. R., Huberty, J. M., Kita, N. T., Ushikubo, T., Kozdon, R., and Valley, J. W. (2011). SIMS analyses of silicon and oxygen isotope ratios for quartz from Archean and Paleoproterozoic banded iron formations. Geochim. Cosmochim. Acta 75, 5879–5891. doi: 10.1016/j.gca.2011.07.023
Henchiri, S., Gaillardet, J., Dellinger, M., Bouchez, J., and Spencer, R. G. M. (2016). Riverine dissolved lithium isotopic signatures in low-relief central Africa and their link to weathering regimes. Geophys. Res. Lett. 43, 4391–4399. doi: 10.1002/2016GL067711
Hendry, K. R., and Andersen, M. B. (2013). The zinc isotopic composition of siliceous marine sponges: investigating nature's sediment traps. Chem. Geol. 354, 33–41. doi: 10.1016/j.chemgeo.2013.06.025
Hendry, K. R., and Brzezinski, M. A. (2014). Using silicon isotopes to understand the role of the Southern Ocean in modern and ancient biogeochemistry and climate. Quat. Sci. Rev. 89, 13–26. doi: 10.1016/j.quascirev.2014.01.019
Hendry, K. R., Georg, R. B., Rickaby, R. E. M., Robinson, L. F., and Halliday, A. N. (2010). Deep ocean nutrients during the Last Glacial Maximum deduced from sponge silicon isotopic compositions. Earth Planet. Sci. Lett. 292, 290–300. doi: 10.1016/j.epsl.2010.02.005
Hendry, K. R., Gong, X., Knorr, G., Pike, J., and Hall, I. R. (2016). Deglacial diatom production in the tropical North Atlantic driven by enhanced silicic acid supply. Earth Planet. Sci. Lett. 438, 122–129. doi: 10.1016/j.epsl.2016.01.016
Hendry, K. R., Leng, M. J., Robinson, L. F., Sloane, H. J., Blusztjan, J., Rickaby, R. E. M., et al. (2011). Silicon isotopes in Antarctic sponges: an interlaboratory comparison. Antarct. Sci. 23, 34–42. doi: 10.1017/S0954102010000593
Hendry, K. R., Marron, A. O., Vincent, F., Conley, D. J., Gehlen, M., Ibarbalz, F. M., et al. (2018). Competition between silicifiers and non-silicifiers in the past and present ocean and its evolutionary impacts. Front. Mar. Sci. doi: 10.3389/fmars.2018.00022
Hendry, K. R., and Rickaby, R. E. M. (2008). Opal (Zn/Si) ratios as a nearshore geochemical proxy in coastal Antarctica. Paleoceanography 23, 1–12. doi: 10.1029/2007PA001576
Hendry, K. R., and Robinson, L. F. (2012). The relationship between silicon isotope fractionation in sponges and silicic acid concentration: Modern and core-top studies of biogenic opal. Geochim. Cosmochim. Acta 81, 1–12. doi: 10.1016/j.gca.2011.12.010
Hendry, K. R., Robinson, L. F., Meredith, M. P., Mulitza, S., Chiessi, C. M., and Arz, H. (2012). Abrupt changes in high-latitude nutrient supply to the Atlantic during the last glacial cycle. Geology 40, 123–126. doi: 10.1130/G32779.1
Hendry, K. R., Robinson, L. F., McManus, J. F., and Hays, J. D. (2014). Silicon isotopes indicate enhanced carbon export efficiency in the North Atlantic during deglaciation. Nat. Commun. 5:3107. doi: 10.1038/ncomms4107
Hendry, K. R., Swann, G. E. A., Leng, M. J., Sloane, H. J., Goodwin, C., Berman, J., et al. (2015). Technical Note: Silica stable isotopes and silicification in a carnivorous sponge Asbestopluma sp. Biogeosciences 12, 3489–3498. doi: 10.5194/bg-12-3489-2015
Heureux, A. M. C., and Rickaby, R. E. M. (2015). Refining our estimate of atmospheric CO2 across the eocene-Oligocene climatic transition. Earth Planet. Sci. Lett. 409, 329–338. doi: 10.1016/j.epsl.2014.10.036
Hildebrand, M. (2008). Diatoms, biomineralization processes, and genomics. Chem. Rev. 108, 4855–4874, doi: 10.1021/cr078253z
Hildebrand, M., Volcani, B. E., Gassmann, W., and Schroeder, J. I. (1997). A gene family of silicon transporters. ature 385, 688–689
Hodson, M. J., Parker, A. G., Leng, M. J., and Sloane, H. J. (2008). Silicon, oxygen and carbon isotope composition of wheat (Triticum aestivum L.) phytoliths: implications for palaeoecology and archaeology. J. Quat. Sci. 23, 331–339. doi: 10.1002/jqs.1176
Honjo, S., Manganini, S. J., Krishfield, R. A., and Francois, R. (2008). Particulate organic carbon fluxes to the ocean interior and factors controlling the biological pump: a synthesis of global sediment trap programs since 1983. Prog. Oceanogr. 76, 217–285. doi: 10.1016/j.pocean.2007.11.003
Horn, M. G., Beucher, C. P., Robinson, R. S., and Brzezinski, M. A. (2011). Southern ocean nitrogen and silicon dynamics during the last deglaciation. Earth Planet. Sci. Lett. 310, 334–339. doi: 10.1016/j.epsl.2011.08.016
Hughes, H. J., Bouillon, S., André, L., and Cardinal, D. (2012). The effects of weathering variability and anthropogenic pressures upon silicon cycling in an intertropical watershed (Tana River, Kenya). Chem. Geol. 308–309, 18–25. doi: 10.1016/j.chemgeo.2012.03.016
Hughes, H. J., Sondag, F., Cocquyt, C., Laraque, A., Pandi, A., André, L., et al. (2011). Effect of seasonal biogenic silica variations on dissolved silicon fluxes and isotopic signatures in the Congo River. Limnol. Oceanogr. 56, 551–561. doi: 10.4319/lo.2011.56.2.0551
Hughes, H. J., Sondag, F., Santos, R. V., André, L., and Cardinal, D. (2013). The riverine silicon isotope composition of the Amazon Basin. Geochim. Cosmochim. Acta 121, 637–651. doi: 10.1016/j.gca.2013.07.040
Iler, R. K. (1979). The Chemistry of Silica: Solubility, Polymerization, Colloid and Surface Properties, and Biochemistry. New York, NY: Wiley.
John, S. G., Geis, R. W., Saito, M. A., and Boyle, E. A. (2007). Zinc isotope fractionation during high-affinity and low-affinity zinc transport by the marine diatom Thalassiosira oceanica. Limnol. Oceanogr. 52, 2710–2714. doi: 10.4319/lo.2007.52.6.2710
Kidder, D. L., and Tomescu, I. (2016). Biogenic chert and the Ordovician silica cycle. Palaeogeogr. Palaeoclimatol. Palaeoecol. 458, 29–38. doi: 10.1016/j.palaeo.2015.10.013
Kim, S., Khim, B.-K., Ikehara, K., Itaki, T., Shibahara, A., and Yamamoto, M. (2017). Millennial-scale changes of surface and bottom water conditions in the northwestern Pacific during the last deglaciation. Glob. Planet. Change 154, 33–43. doi: 10.1016/j.gloplacha.2017.04.009
King, S. L., Froelich, P. N., and Jahnke, R. A. (2000). Early diagenesis of germanium in sediments of the Antarctic South Atlantic: in search of the missing Ge sink. Geochim. Cosmochim. Acta 64, 1375–1390. doi: 10.1016/S0016-7037(99)00406-8
Koning, E., Epping, E., and Van Raaphorst, W. (2002). Determining biogenic silica in marine samples by tracking silicate and aluminium concentrations in alkaline leaching solutions. Aquat. Geochem. 8, 37–67. doi: 10.1023/A:1020318610178
Koning, E., Gehlen, M., Flank, A. M., Calas, G., and Epping, E. (2007). Rapid post-mortem incorporation of aluminum in diatom frustules: evidence from chemical and structural analyses. Mar. Chem. 106, 208–222. doi: 10.1016/j.marchem.2006.06.009
Kurtz, A. C., Derry, L. A., and Chadwick, O. A. (2002). Germanium-silicon fractionation in the weathering environment. Geochim. Cosmochim. Acta 66, 1525–1537. doi: 10.1016/S0016-7037(01)00869-9
Labeyrie, L., and Juillet, A. (1982). Oxygen isotopic exchangeability of diatom valve silica; interpretation and consequences for paleoclimatic studies. Geochim. Cosmochim. Acta 46, 967–975. doi: 10.1016/0016-7037(82)90052-7
Labeyrie, L. D. (1979). La composition isotopique de l‘oxygène de la silica des valves de diatomées. Mise au point d'une nouvelle méthode de paléoclimatologie quantitative. Thesis, Université de Paris.
Lal, D., Charles, C., Vacher, L., Goswami, J. N., Jull, A. J. T., McHargue, L., et al. (2006). Paleo-ocean chemistry records in marine opal: implications for fluxes of trace elements, cosmogenic nuclides (10Be and 26Al), and biological productivity. Geochim. Cosmochim. Acta 70, 3275–3289. doi: 10.1016/j.gca.2006.04.004
Lamb, A. L., Brewer, T. S., Leng, M. J., Sloane, H. J., and Lamb, H. F. (2007). A geochemical method for removing the effect of tephra on lake diatom oxygen isotope records. J. Paleolimnol. 37, 499–516. doi: 10.1007/s10933-006-9034-5
Laruelle, G. G., Roubeix, V., Sferratore, A., Brodherr, B., Ciuffa, D., Conley, D. J., et al. (2009). Anthropogenic perturbations of the silicon cycle at the global scale: Key role of the land-ocean transition. Global Biogeochem. Cycles 23, 1–17. doi: 10.1029/2008GB003267
Laws, E. A. Popp, B. N., Bidigare, J. R. R., Kennicutt, M. C., and Macko, S. (1995). Dependence of phytoplankton carbon isotopic composition on growth rate and [CO2]aq: theoretical considerations and experimental results. Geochim. Cosmochim. Acta 59, 1131–1138. doi: 10.1016/0016-7037(95)00030-4
Leclerc, A. J., and Labeyrie, L. (1987). Temperature dependence of the oxygen isotopic fractionation between diatom silica and water. Earth Planet. Sci. Lett. 84, 69–74. doi: 10.1016/0012-821X(87)90177-4
Lewis, B. L., Froelich, P. N., and Andreae, M. O. (1985). Methylgermanium in natural waters. Nature 313, 303–305. doi: 10.1038/313303a0
Lewis, B. L., Andreae, M. O., Froelich, P. N., and Mortlock, R. A. (1988). A review of the biogeochemistry of germanium in natural waters. Science Total Environ. 73, 107–120. doi: 10.1016/0048-9697(88)90191-X
Lewis, B. L., Andreae, M. O., and Froelich, P. N. (1989). Sources and sinks of methylgermanium in natural waters. Mar. Chem. 27, 179–200. doi: 10.1016/0304-4203(89)90047-9
Loucaides, S., Michalopoulos, P., Presti, M., Koning, E., Behrends, T., and Van Cappellen, P. (2010). Seawater-mediated interactions between diatomaceous silica and terrigenous sediments: results from long-term incubation experiments. Chem. Geol. 270, 68–79. doi: 10.1016/j.chemgeo.2009.11.006
Lisiecki, L. E., and Raymo, M. E. (2005). A Pliocene-Pleistocene stack of 57 globally distributed benthic δ 18 O records. Paleoceanography 20:PA1003. doi: 10.1029/2004PA001071
Ma, J. F., Miyake, Y., and Takahashi, E. (2001). “Silicon as a beneficial element for crop plants,” in Silicon in Agriculture, eds L. E. Datnoff, G. H. Snyder, and G. H. Korndorfer (Amsterdam: Elsevier), 17–39.
Ma, J. F., Tamai, K., Yamaji, N., Mitani, N., Konishi, S., Katsuhara, M., et al. (2006). A silicon transporter in rice. Nature 440, 688–691. doi: 10.1038/nature04590
Mackay, A. W., Swann, G. E. A., Brewer, T. S., Leng, M. J., Morley, D. W., Piotrowska, N., et al. (2011). A reassessment of late glacial - Holocene diatom oxygen isotope record from Lake Baikal using a geochemical mass-balance approach. J. Quat. Sci. 26, 627–634. doi: 10.1002/jqs.1484
Mackay, A. W., Swann, G. E. A., Fagel, N., Fietz, S., Leng, M. J., Morley, D., et al. (2013). Hydrological instability during the Last Interglacial in central Asia: a new diatom oxygen isotope record from Lake Baikal. Quat. Sci. Rev. 66, 45–54. doi: 10.1016/j.quascirev.2012.09.025
Mackenzie, F., and Garrels, R. (1966). Chemical mass balance between rivers and oceans.pdf. Am. J. Sci. 264, 19. doi: 10.2475/ajs.264.7.507
Madella, M., Aldexandre, A., and Ball, T. (2005). international code for phytolith nomenclature 1.0. Ann. Bot. 96, 253–260. doi: 10.1093/aob/mci172
Maher, W. A., Ellwood, M. J., Kelly, M., and De Deckker, P. (2006). Germanium incorporation into sponge spicules: development of a proxy for reconstructing inorganic germanium and silicon concentrations in seawater. Geochim. Cosmochim. Acta 70, A384–A384. doi: 10.1016/j.gca.2006.06.777
Maldonado, M., Carmona, M. C., Uriz, M. J., and Cruzado, A. (1999). Decline in Mesozoic reef-building sponges explained by silicon limitation. Nature 401, 785–788. doi: 10.1038/44560
Maier, E., Chapligin, B., Abelmann, A., Gersonde, R., Esper, O., Ren, J., et al. (2013). Combined oxygen and silicon isotope analysis of diatom silica from a deglacial subarctic Pacific record. J. Quat. Sci. 28:571 doi: 10.1002/jqs.2649
Maldonado, M., Carmona, C., Velasquez, Z., Puig, A., Cruzado, A., Lopez, A., et al. (2005). Siliceous sponges as a silicon sink: an overlooked aspect of benthopelagic coupling in the marine silicon cycle. Limnol. Oceanogr. 50, 799–809. doi: 10.4319/lo.2005.50.3.0799
Maldonado, M., Navarro, L., Grasa, A., Gonzalez, A., and Vaquerizo, I. (2011). Silicon uptake by sponges: a twist to understanding nutrient cycling on continental margins. Sci. Rep. 1, 30. doi: 10.1038/srep00030
Mann, S., and Perry, C. C. (1986). “Structural aspects of biogenic silica,” in Silicon Biochemistry, CIBA Foundation Symposium 121, eds D. Evered and M. O'Connor (New York, NY: Wiley), 40–58.
Marin-Carbonne, J., Robert, F., and Chaussidon, M. (2014). The silicon and oxygen isotope compositions of Precambrian cherts: a record of oceanic paleo-temperatures? Precambrian Res. 247, 223–234. doi: 10.1016/j.precamres.2014.03.016
Martin-Jezequel, V., Hildebrand, M., and Brzezinski, M. A. (2000). Silicon metabolism in diatoms: implications for growth. J. Phycol. 36, 821–840. doi: 10.1046/j.1529-8817.2000.00019.x
Matheney, R. K., and Knauth, L. P. (1989). Oxygen-isotope fractionation between marine biogenic silica and seawater. Geochim. Cosmochim. Acta 53, 3207–3214. doi: 10.1016/0016-7037(89)90101-4
Matsumoto, K., and Sarmiento, J. L. (2008). A corollary to the silicic acid leakage hypothesis. Paleoceanography 23:PA2203. doi: 10.1029/2007PA001515
Matsumoto, K., Sarmiento, J. L., and Brzezinski, M. A. (2002). Silicic acid leakage from the Southern Ocean: a possible explanation for glacial atmospheric pCO(2). Global Biogeochem. Cycles 16, 1–23. doi: 10.1029/2001GB001442
Matteuzzo, M. C., Alexandre, A., Varajão, A. F. D. C., Volkmer-Ribeiro, C., Almeida, A. C. S., Varajão, C. A. C., et al. (2013). Assessing the relationship between the δ18O signatures of siliceous sponge spicules and water in a~tropical lacustrine environment (Minas Gerais, Brazil). Biogeosci. Discuss. 10, 12887–12918. doi: 10.5194/bgd-10-12887-2013
McManus, J., Hammond, D. E., Cummins, K., Klinkhammer, G. P., and Berelson, W. M. (2003). Diagenetic Ge-Si fractionation in continental margin environments: further evidence for a nonopal Ge sink. Geochim. Cosmochim. Acta 67, 4545–4557. doi: 10.1016/S0016-7037(03)00385-5
Meckler, A. N., Sigman, D. M., Gibson, K. A., François, R., Martínez-García, A., Jaccard, S. L., et al. (2013). Deglacial pulses of deep-ocean silicate into the subtropical North Atlantic Ocean. Nature 495, 495–498. doi: 10.1038/nature12006
Michalopoulos, P., and Aller, R. C. (1995). Rapid clay mineral formation in amazon delta sediments: reverse weathering and oceanic elemental cycles. Science 270, 614–617. doi: 10.1126/science.270.5236.614
Michalopoulos, P., and Aller, R. C. (2004). Early diagenesis of biogenic silica in the Amazon delta: alteration, authigenic clay formation, and storage. Geochim. Cosmochim. Acta 68, 1061–1085. doi: 10.1016/j.gca.2003.07.018
Milligan, A. J. (2004). Dynamics of silicon metabolism and silicon isotopic discrimination in a marine diatom as a function of pCO2. Limnol. Oceanogr. 49, 322–329. doi: 10.4319/lo.2004.49.2.0322
Misra, S., and Froelich, P. N. (2012). Lithium isotope history of Cenozoic seawater: changes in silicate weathering and reverse weathering. Science 335, 818–823. doi: 10.1126/science.1214697
Montoya, J. P., and McCarthy, J. J. (1995). Isotopic fractionation during nitrate uptake by phytoplankton grown in continuous culture. J. Plankton Res. 17, 439–464. doi: 10.1093/plankt/17.3.439
Mopper, K., and Garlick, G. (1971). Oxygen isotope fractionation between biogenic silica and ocean water. Geochim. Cosmochim. Acta 35, 1185–1187. doi: 10.1016/0016-7037(71)90032-9
Moran, S. B., and Moore, R. M. (1988). Evidence from mesocosm studies for biological removal of dissolved aluminium from sea water. Nature 335, 706–708. doi: 10.1038/335706a0
Morley, D. W., Leng, M. J., Mackay, A. W., and Sloane, H. J. (2005). Late glacial and Holocene environmental change in the Lake Baikal region documented by oxygen isotopes from diatom silica. Glob. Planet. Change 46, 221–233. doi: 10.1016/j.gloplacha.2004.09.018
Morley, D. W., Leng, M. J., Mackay, A. W., Sloane, H. J., Rioual, P., and Battarbee, R. W. (2004). Cleaning of lake sediment samples for diatom oxygen isotope analysis. J. Paleolimnol. 31, 391–401. doi: 10.1023/B:JOPL.0000021854.70714.6b
Mortlock, R. A., Charles, C. D., Froelich, P. N., Zibello, M. A., Saltzman, J., Hays, J. D., et al. (1991). Evidence for lower productivity in the Antarctic Ocean during the last glaciation. Nature 351, 220–223. doi: 10.1038/351220a0
Mortlock, R. A., and Froelich, P. N. (1987). Continental weathering of germanium - Ge/Si in the global river discharge. Geochim. Cosmochim. Acta 51, 2075–2082. doi: 10.1016/0016-7037(87)90257-2
Mortlock, R. A., and Froelich, P. N. (1996). Determination of germanium by isotope dilution hydride generation inductively coupled plasma mass spectrometry. Anal. Chim. Acta 332, 277–284. doi: 10.1016/0003-2670(96)00230-9
Mortlock, R. A., Froelich, P. N., Feely, R. A., Massoth, G. J., Butterfield, D. A., and Lupton, J. E. (1993). Silica and germanium in pacific-ocean hydrothermal vents and plumes. Earth Planet. Sci. Lett. 119, 365–378. doi: 10.1016/0012-821X(93)90144-X
Muller, W. E. G., Li, J. H., Schroder, H. C., Qiao, L., and Wang, X. H. (2007). The unique skeleton of siliceous sponges (Porifera; Hexactinellida and Demospongiae) that evolved first from the Urmetazoa during the Proterozoic: a review. Biogeosciences 4, 219–232. doi: 10.5194/bg-4-219-2007
Murnane, R., Leslie, B., Hammond, D., and Stallard, R. (1989). Germanium geochemistry in the Southern California Borderlands. Geochim. Cosmochim. Acta 53, 2873–2882. doi: 10.1016/0016-7037(89)90164-6
Needoba, J., Waser, N., Harrison, P., and Calvert, S. (2003). Nitrogen isotope fractionation in 12 species of marine phytoplankton during growth on nitrate. Mar. Ecol. Prog. Ser. 255, 81–91. doi: 10.3354/meps255081
Nelson, D. M., Treguer, P., Brzezinski, M. A., Leynaert, A., and Queguiner, B. (1995). Production and dissolution of biogenic silica in the ocean - revised global estimates, comparison with regional data and relationship to biogenic sedimentation. Global Biogeochem. Cycles 9, 359–372. doi: 10.1029/95GB01070
Oelze, M., von Blanckenburg, F., Bouchez, J., Hoellen, D., and Dietzel, M. (2015). The effect of Al on Si isotope fractionation investigated by silica precipitation experiments. Chem. Geol. 397, 94–105. doi: 10.1016/j.chemgeo.2015.01.002
Oelze, M., von Blanckenburg, F., Hoellen, D., Dietzel, M., and Bouchez, J. (2014). Si stable isotope fractionation during adsorption and the competition between kinetic and equilibrium isotope fractionation: Implications for weathering systems. Chem. Geol. 380, 161–171. doi: 10.1016/j.chemgeo.2014.04.027
Opfergelt, S., Burton, K. W., Pogge von Strandmann, P. A. E., Gislason, S. R., and Halliday, A. N. (2013). Riverine silicon isotope variations in glaciated basaltic terrains: Implications for the Si delivery to the ocean over glacial-interglacial intervals. Earth Planet. Sci. Lett. 369–370, 211–219. doi: 10.1016/j.epsl.2013.03.025
Opfergelt, S., Cardinal, D., André, L., Delvigne, C., Bremond, L., and Delvaux, B. (2010). Variations of δ30Si and Ge/Si with weathering and biogenic input in tropical basaltic ash soils under monoculture. Geochim. Cosmochim. Acta 74, 225–240. doi: 10.1016/j.gca.2009.09.025
Opfergelt, S., Cardinal, D., Henriet, C., Andre, L., and Delvaux, B. (2006a). Silicon isotope fractionation between plant parts in banana: in situ vs. in vitro. J. Geochemical Explor. 88, 224–227. doi: 10.1016/j.gexplo.2005.08.044
Opfergelt, S., Cardinal, D., Henriet, C., Draye, X., Andre, L., and Delvaux, B. (2006b). Silicon isotopic fractionation by banana (Musa spp.) grown in a continuous nutrient flow device. Plant Soil 285, 333–345. doi: 10.1007/s11104-006-9019-1
Opfergelt, S., and Delmelle, P. (2012). Silicon isotopes and continental weathering processes: assessing controls on Si transfer to the ocean. Comptes Rendus Geosci. 344, 723–738. doi: 10.1016/j.crte.2012.09.006
Opfergelt, S., Delvaux, B., André, L., and Cardinal, D. (2008). Plant silicon isotopic signature might reflect soil weathering degree. Biogeochemistry 91, 163–175. doi: 10.1007/s10533-008-9278-4
Opfergelt, S., Eiriksdottir, E. S. S., Burton, K. W. W., Einarsson, A., Siebert, C., Gislason, S. R. R., et al. (2011). Quantifying the impact of freshwater diatom productivity on silicon isotopes and silicon fluxes: Lake Myvatn, Iceland. Earth Planet. Sci. Lett. 305, 73–82. doi: 10.1016/j.epsl.2011.02.043
Opfergelt, S., Georg, R. B., Delvaux, B., Cabidoche, Y. M., Burton, K. W., and Halliday, A. N. (2012). Silicon isotopes and the tracing of desilication in volcanic soil weathering sequences, Guadeloupe. Chem. Geol. 326–327, 113–122. doi: 10.1016/j.chemgeo.2012.07.032
Panizzo, V., Crespin, J., Crosta, X., Shemesh, A., Massé, G., Yam, R., et al. (2014). Sea ice diatom contributions to Holocene nutrient utilization in East Antarctica. Paleoceanography 29, 328–343. doi: 10.1002/2014PA002609
Panizzo, V. N., Swann, G. E. A., Mackay, A. W., Vologina, E., Alleman, L., André, L., et al. (2017). Constraining modern-day silicon cycling in Lake Baikal. Global Biogeochem. Cycles 31, 556–574. doi: 10.1002/2016GB005518
Panizzo, V. N., Swann, G. E. A., Mackay, A. W., Vologina, E., Sturm, M., Pashley, V., et al. (2016). Insights into the transfer of silicon isotopes into the sediment record. Biogeosciences 13, 147–157. doi: 10.5194/bg-13-147-2016
Perry, C. C., Belton, D., and Shafran, K. (2003). “Studies of biosilicas; structural aspects, chemicalprinciples, model studies and the future,” in Silicon Biomineralization: Biology—Biochemistry—Molecular Biology—Biotechnology, ed W. E. G. Müller (Berlin; Heidelberg: Springer), 269–299.
Pichevin, L., Ganeshram, R. S., Reynolds, B. C., Prahl, F., Pedersen, T. F., Thunell, R., et al. (2012). Silicic acid biogeochemistry in the Gulf of California: insights from sedimentary Si isotopes. Paleoceanography 27:PA2201. doi: 10.1029/2011PA002237
Pichevin, L., Reynolds, B., Ganeshram, R., Cacho, I., Pena, L., Keefe, K., et al. (2009). Enhanced carbon pump inferred from relaxation of nutrient limitation in the glacial ocean. Nature 459, 1114–1117. doi: 10.1038/nature08101
Pike, J., Swann, G. E. A., Leng, M. J., and Snelling, A. M. (2013). Glacial discharge along the west Antarctic Peninsula during the Holocene. Nat. Geosci. 6, 199–202. doi: 10.1038/ngeo1703
Pogge von Strandmann, P. A. E., Opfergelt, S., Lai, Y. J., Sigfússon, B., Gislason, S. R., and Burton, K. W. (2012). Lithium, magnesium and silicon isotope behaviour accompanying weathering in a basaltic soil and pore water profile in Iceland. Earth Planet. Sci. Lett. 339–340, 11–23. doi: 10.1016/j.epsl.2012.05.035
Pokrovski, G. S., and Schott, J. (1998). Experimental study of the complexation of silicon and germanium with aqueous organic species: implications for germanium and silicon transport and Ge/Si ratio in natural waters. Geochim. Cosmochim. Acta 62, 3413–3428. doi: 10.1016/S0016-7037(98)00249-X
Pokrovsky, O. S., Reynolds, B. C., Prokushkin, A. S., Schott, J., and Viers, J. (2013). Silicon isotope variations in Central Siberian rivers during basalt weathering in permafrost-dominated larch forests. Chem. Geol. 355, 103–116. doi: 10.1016/j.chemgeo.2013.07.016
Presti, M., and Michalopoulos, P. (2008). Estimating the contribution of the authigenic mineral component to the long-term reactive silica accumulation on the western shelf of the Mississippi River Delta. Cont. Shelf Res. 28, 823–838. doi: 10.1016/j.csr.2007.12.015
Qin, T., Wu, F., Wu, Z., and Huang, F. (2016). First-principles calculations of equilibrium fractionation of O and Si isotopes in quartz, albite, anorthite, and zircon. Contrib. Mineral. Petrol. 171:91. doi: 10.1007/s00410-016-1303-3
Rabouille, C., Mackenzie, F. T., and Ver, L. M. (2001). Influence of the human perturbation on carbon, nitrogen, and oxygen biogeochemical cycles in the global coastal ocean. Geochim. Cosmochim. Acta 65, 3615–3641. doi: 10.1016/S0016-7037(01)00760-8
Ragueneau, O., Tréguer, P., Leynaert, A., Anderson, R., Brzezinski, M., DeMaster, D., et al. (2000). A review of the Si cycle in the modern ocean: recent progress and missing gaps in the application of biogenic opal as a paleoproductivity proxy. Glob. Planet. Change 26, 317–365. doi: 10.1016/S0921-8181(00)00052-7
Rahman, S., Aller, R. C., and Cochran, J. K. (2016). Cosmogenic 32 Si as a tracer of biogenic silica burial and diagenesis: major deltaic sinks in the silica cycle. Geophys. Res. Lett. 43, 7124–7132. doi: 10.1002/2016GL069929
Rau, G. H., Riebesell, U., and Wolf-Gladrow, D. (1996). A model of photosynthetic 13C fractionation by marine phytoplankton based on diffusive molecular CO2 uptake. Mar. Ecol. Prog. Ser. 133, 275–285. doi: 10.3354/meps133275
Rau, G. H., Riebesell, U., and Wolf-Gladrow, D. (1997). CO 2aq -dependent photosynthetic 13 C fractionation in the ocean: a model versus measurements. Global Biogeochem. Cycles 11, 267–278. doi: 10.1029/97GB00328
Raymo, M. E., Lisiecki, L. E., Nisancioglu, K. H., Hemispheres, S., and Instead, O. (2006). Plio-Pleistocene ice volume, Antarctic climate, and the Global. Science 313, 492–496. doi: 10.1126/science.1123296
Reincke, T., and Barthel, D. (1997). Silica uptake kinetics of Halichondria panicea in kiel bight. Mar. Biol. 129, 591–593. doi: 10.1007/s002270050200
Ren, H., Brunelle, B. G., Sigman, D. M., and Robinson, R. S. (2013). Diagenetic aluminum uptake into diatom frustules and the preservation of diatom-bound organic nitrogen. Mar. Chem. 155, 92–101. doi: 10.1016/j.marchem.2013.05.016
Reynolds, B. C. (2011). “Silicon isotopes as tracers of terrestrial processes,” in Handbook of Environmental Isotope Geochemistry, ed M. Baskaran (Berlin: Springer-Verlag), 87–104. doi: 10.1007/978-3-642-10637-8_6
Reynolds, B. C., Aggarwal, J., André, L., Baxter, D., Beucher, C., Brzezinski, M. A., et al. (2007). An inter-laboratory comparison of Si isotope reference materials. J. Anal. At. Spectrom. 22, 561–568. doi: 10.1039/B616755A
Reynolds, B. C., Frank, M., and Halliday, A. N. (2006). Silicon isotope fractionation during nutrient utilization in the North Pacific. Earth Planet. Sci. Lett. 244, 431–443. doi: 10.1016/j.epsl.2006.02.002
Rickert, D., Schlüter, M., and Wallmann, K. (2002). Dissolution kinetics of biogenic silica from the water column to the sediments. Geochim. Cosmochim. Acta 66, 439–455. doi: 10.1016/S0016-7037(01)00757-8
Robert, F., and Chaussidon, M. (2006). A palaeotemperature curve for the Precambrian oceans based on silicon isotopes in cherts. Nature 443, 969–972. doi: 10.1038/nature05239
Robinson, R. S., Brunelle, B. G., and Sigman, D. M. (2004). Revisiting nutrient utilization in the glacial Antarctic: evidence from a new method for diatom-bound N isotopic analysis. Paleoceanography 19:PA3001. doi: 10.1029/2003PA000996
Robinson, R. S., Kienast, M., Luiza Albuquerque, A., Altabet, M., Contreras, S., De Pol Holz, R., et al. (2012). A review of nitrogen isotopic alteration in marine sediments. Paleoceanography 27:PA4203 doi: 10.1029/2012PA002321
Robinson, R. S., and Sigman, D. M. (2008). Nitrogen isotopic evidence for a poleward decrease in surface nitrate within the ice age Antarctic. Quat. Sci. Rev. 27, 1076–1090. doi: 10.1016/j.quascirev.2008.02.005
Robinson, R. S., Sigman, D. M., DiFiore, P. J., Rohde, M. M., Mashiotta, T. A., and Lea, D. W. (2005). Diatom-bound 15N/14N: new support for enhanced nutrient consumption in the ice age subantarctic. Paleoceanography 20, 1–14. doi: 10.1029/2004PA001114
Roerdink, D. L., van den Boorn, S. H. J. M., Geilert, S., Vroon, P. Z., and van Bergen, M. J. (2015). Experimental constraints on kinetic and equilibrium silicon isotope fractionation during the formation of non-biogenic chert deposits. Chem. Geol. 402, 40–51. doi: 10.1016/j.chemgeo.2015.02.038
Rousseau, J., Ellwood, M. J., Bostock, H., and Neil, H. (2016). Estimates of late quaternary mode and intermediate water silicic acid concentration in the Pacific Southern Ocean. Earth Planet. Sci. Lett. 439, 101–108. doi: 10.1016/j.epsl.2016.01.023
Rouxel, O. J., and Luais, B. (2017). Germanium isotope geochemistry. Rev. Mineral. Geochem. 82, 601–656. doi: 10.2138/rmg.2017.82.14
Rude, P. D., and Aller, R. C. (1994). Fluorine uptake by Amazon continental shelf sediment and its impact on the global fluorine cycle. Cont. Shelf Res. 14, 883–907. doi: 10.1016/0278-4343(94)90078-7
Sarmiento, J. L., Gruber, N., Brzezinski, M. A., and Dunne, J. P. (2004). High-latitude controls of thermocline nutrients and low latitude biological productivity. Nature 427, 56–60. doi: 10.1038/nature02127
Sarmiento, J. L., and Toggweiler, J. R. (1984). A new model for the role of the oceans in determining atmospheric PCO2. Nature 308, 621–624. doi: 10.1038/308621a0
Schmidt, M., Botz, R., Rickert, D., Bohrmann, G., Hall, S. R., and Mann, S. (2001). Oxygen isotopes of marine diatoms and relations to opal-A maturation. Geochim. Cosmochim. Acta 65, 201–211. doi: 10.1016/S0016-7037(00)00534-2
Schröder, H. C., Natalio, F., Shukoor, I., Tremel, W., Schloßmacher, U., Wang, X., et al. (2007). Apposition of silica lamellae during growth of spicules in the demosponge Suberites domuncula: biological/biochemical studies and chemical/biomimetical confirmation. J. Struct. Biol. 159, 325–334. doi: 10.1016/j.jsb.2007.01.007
Schuessler, J. A., and von Blanckenburg, F. (2014). Testing the limits of micro-scale analyses of Si stable isotopes by femtosecond laser ablation multicollector inductively coupled plasma mass spectrometry with application to rock weathering. Spectrochim. Acta Part B At. Spectrosc. 98, 1–18. doi: 10.1016/j.sab.2014.05.002
Sharp, Z. D., Gibbons, J. A., Maltsev, O., Atudorei, V., Pack, A., Sengupta, S., et al. (2016). A calibration of the triple oxygen isotope fractionation in the SiO2–H2O system and applications to natural samples. Geochim. Cosmochim. Acta 186, 105–119. doi: 10.1016/j.gca.2016.04.047
Shemesh, A., Burckle, L. H., and Hays, J. D. (1995). Late Pleistocene oxygen isotope records of biogenic silica from the Atlantic sector of the Southern Ocean. Paleoceanography 10, 179–196. doi: 10.1029/94PA03060
Shemesh, A., Charles, C. D., and Fairbanks, R. G. (1992). Oxygen isotopes in biogenic silica: global changes in ocean temperature and isotopic composition. Science 256, 1434–1436. doi: 10.1126/science.256.5062.1434
Shemesh, A., Macko, S. A., Charles, C. D., and Rau, G. H. (1993). Isotopic evidence for reduced productivity in the glacial Southern Ocean. Science 262, 407–410. doi: 10.1126/science.262.5132.407
Shemesh, A., Mortlock, R. A., Smith, R. J., and Froelich, P. N. (1988). Determination of Ge/Si in marine siliceous microfossils - separation, cleaning and dissolution of diatoms and radiolaria. Mar. Chem. 25, 305–323. doi: 10.1016/0304-4203(88)90113-2
Siever, R. (1991). The silica cycle in the Precambrian. Geochim. Cosmochim. Acta 56, 3265–3272. doi: 10.1016/0016-7037(92)90303-Z
Sigman, D. M., Altabet, M. A., Francois, R., McCorkle, D. C., and Gaillard, J.-F. (1999). The isotopic composition of diatom-bound nitrogen in Southern Ocean sediments. Paleoceanography 14, 118–134. doi: 10.1029/1998PA900018
Singh, S. P., Singh, S. K., Bhushan, R., and Rai, V. K. (2015). Dissolved silicon and its isotopes in the water column of the Bay of Bengal: internal cycling versus lateral transport. Geochim. Cosmochim. Acta 151, 172–191. doi: 10.1016/j.gca.2014.12.019
Smetacek, V., Klaas, C., Strass, V. H., Assmy, P., Montresor, M., Cisewski, B., et al. (2012). Deep carbon export from a Southern Ocean iron-fertilized diatom bloom. Nature 487, 313–319. doi: 10.1038/nature11229
Snelling, A. M., Swann, G. E. A., Pike, J., and Leng, M. J. (2014). Pliocene diatom and sponge spicule oxygen isotope ratios from the Bering Sea: isotopic offsets and future directions. Clim. Past 10, 1837–1842. doi: 10.5194/cp-10-1837-2014
Stallard, R. F., and Edmond, J. M. (1983). Geochemistry of the Amazon: 2. The influence of geology and weathering environment on the dissolved load. J. Geophys. Res. Ocean. 88, 9671–9688. doi: 10.1029/JC088iC14p09671
Steinhoefel, G., Breuer, J., von Blanckenburg, F., Horn, I., Kaczorek, D., and Sommer, M. (2011). Micrometer silicon isotope diagnostics of soils by UV femtosecond laser ablation. Chem. Geol. 286, 280–289. doi: 10.1016/j.chemgeo.2011.05.013
Stoll, H. M., Mendez-Vicente, A., Abrevaya, L., Anderson, R. F., Rigual-Hernández, A. S., and Gonzalez-Lemos, S. (2017). Growth rate and size effect on carbon isotopic fractionation in diatom-bound organic matter in recent Southern Ocean sediments. Earth Planet. Sci. Lett. 457, 87–99. doi: 10.1016/j.epsl.2016.09.028
Street-Perrott, F. A., Barker, P. A., Leng, M. J., Sloane, H. J., Wooller, M. J., Ficken, K. J., et al. (2008). Towards an understanding of late quaternary variations in the continental biogeochemical cycle of silicon: multi-isotope and sediment-flux data for Lake Rutundu, Mt Kenya, East Africa, since 38 ka BP. J. Quat. Sci. 23, 375–387. doi: 10.1002/jqs.1187
Struyf, E., and Conley, D. J. (2012). Emerging understanding of the ecosystem silica filter. Biogeochemistry 107, 9–18. doi: 10.1007/s10533-011-9590-2
Struyf, E., Smis, A., Van Damme, S., Garnier, J., Govers, G., Van Wesemael, B., et al. (2010). Historical land use change has lowered terrestrial silica mobilization. Nat. Commun. 1:129. doi: 10.1038/ncomms1128
Struyf, E., Smis, A., Van Damme, S., Meire, P., and Conley, D. J. (2009). The global biogeochemical silicon cycle. Silicon 1, 207–213. doi: 10.1007/s12633-010-9035-x
Struyf, E., Van Damme, S., and Meire, P. (2004). Possible effects of climate change on estuarine nutrient fluxes: a case study in the highly nutrified Schelde estuary (Belgium, The Netherlands). Estuar. Coast. Shelf Sci. 60, 649–661. doi: 10.1016/j.ecss.2004.03.004
Sun, L., Wu, L. H., Ding, T. P., and Tian, S. H. (2008). Silicon isotope fractionation in rice plants, an experimental study on rice growth under hydroponic conditions. Plant Soil 304, 291–300. doi: 10.1007/s11104-008-9552-1
Sun, X., Andersson, P., Humborg, C., Gustafsson, B., Conley, D. J., Crill, P., et al. (2011). Climate dependent diatom production is preserved in biogenic Si isotope signatures. Biogeosciences 8, 3491–3499. doi: 10.5194/bg-8-3491-2011
Sun, X., Andersson, P. S., Humborg, C., Pastuszak, M., and Mörth, C. M. (2013). Silicon isotope enrichment in diatoms during nutrient-limited blooms in a eutrophied river system. J. Geochemical Explor. 132, 173–180. doi: 10.1016/j.gexplo.2013.06.014
Sun, X., Higgins, J., and Turchyn, A. V. (2016). Diffusive cation fluxes in deep-sea sediments and insight into the global geochemical cycles of calcium, magnesium, sodium and potassium. Mar. Geol. 373, 64–77. doi: 10.1016/j.margeo.2015.12.011
Sutton, J. (2011). Germanium/Silicon and Silicon Isotope Fractionation by Marine Diatoms and Sponges and Utility as Tracers of Silicic Acid Utilization. Thesis, Australian National University.
Sutton, J., Ellwood, M. J., Maher, W. A., and Croot, P. L. (2010). Oceanic distribution of inorganic germanium relative to silicon: Germanium discrimination by diatoms. Global Biogeochem. Cycles 24:GB2017. doi: 10.1029/2009GB003689
Sutton, J. N., Varela, D. E., Brzezinski, M. A., and Beucher, C. P. (2013). Species-dependent silicon isotope fractionation by marine diatoms. Geochim. Cosmochim. Acta 104, 300–309. doi: 10.1016/j.gca.2012.10.057
Suzuki, N., and Aita, Y. (2011). Radiolaria: achievements and unresolved issues: taxonomy and cytology. Plankt. Benthos Res. 6, 69–91. doi: 10.3800/pbr.6.69
Swann, G. E. A., and Leng, M. J. (2009). A review of diatom δ18O in palaeoceanography. Quat. Sci. Rev. 28, 384–398. doi: 10.1016/j.quascirev.2008.11.002
Swann, G. E. A., Leng, M. J., Juschus, O., Melles, M., Brigham-Grette, J., and Sloane, H. J. (2010). A combined oxygen and silicon diatom isotope record of late quaternary change in Lake El'gygytgyn, North East Siberia. Quat. Sci. Rev. 29, 774–786. doi: 10.1016/j.quascirev.2009.11.024
Swann, G. E. A., and Snelling, A. M. (2015). Photic zone changes in the north-west Pacific Ocean from MIS 4-5e. Clim. Past 11, 15–25. doi: 10.5194/cp-11-15-2015
Takahashi, E., Ma, J. F., and Miyake, Y. (1990). The possibility of silicon as an essential element for higher plants. Comments Agric. Food Chem 2, 99–122.
Tatzel, M., von Blanckenburg, F., Oelze, M., Schuessler, J. A., and Bohrmann, G. (2015). The silicon isotope record of early silica diagenesis. Earth Planet. Sci. Lett. 428, 293–303. doi: 10.1016/j.epsl.2015.07.018
Thamatrakoln, K., and Hildebrand, M. (2008). Silicon uptake in diatoms revisited: a model for saturable and nonsaturable uptake kinetics and the role of silicon transporters. Plant Physiol. 146, 1397–1407. doi: 10.1104/pp.107.107094
Thamatrakoln, K., and Kustka, A. B. (2009). When to say when: can excessive drinking explain silicon uptake in diatoms? BioEssays 31, 322–327. doi: 10.1002/bies.200800185
Tortell, P. D., and Morel, F. M. M. (2002). Sources of inorganic carbon for phytoplankton in the eastern Subtropical and Equatorial Pacific Ocean. Limnol. Oceanogr. 47, 1012–1022. doi: 10.4319/lo.2002.47.4.1012
Tréguer, P. J., and De La Rocha, C. L. (2013). The world ocean silica cycle. Ann. Rev. Mar. Sci. 5, 477–501. doi: 10.1146/annurev-marine-121211-172346
Twining, B. S., Baines, S. B., Fisher, N. S., Maser, J., Vogt, S., Jacobsen, C., et al. (2003). Quantifying trace elements in individual aquatic protist cells with a synchrotron X-ray fluorescence microprobe. Anal. Chem. 75, 3806–3816. doi: 10.1021/ac034227z
Uriz, M. J., Turon, X., Becerro, M. A., and Agell, G. (2003): Siliceous spicules skeleton frameworks in sponges: origin, diversity, ultrastructural patterns, biological functions. Microsc. Res. Tech. 62, 279–299. doi: 10.1002/jemt.10395
van Bennekom, A. J., and van der Gaast, S. J. (1976). Possible clay structures in frustules of living diatoms. Geochim. Cosmochim. Acta 40, 1149–1150. doi: 10.1016/0016-7037(76)90150-2
Vandevenne, F. I., Delvaux, C., Hughes, H. J., André, L., Ronchi, B., Clymans, W., et al. (2015). Landscape cultivation alters δ30Si signature in terrestrial ecosystems. Sci. Rep. 5:7732. doi: 10.1038/srep07732
Varela, D. E., Brzezinski, M. A., Beucher, C. P., Jones, J. L., Giesbrecht, K. E., Lansard, B., et al. (2016). Heavy silicon isotopic composition of silicic acid and biogenic silica in Arctic waters over the Beaufort shelf and the Canada Basin. Global Biogeochem. Cycles 30, 804–824. doi: 10.1002/2015GB005277
Varela, D. E., Pride, C. J., and Brzezinski, M. A. (2004). Biological fractionation of silicon isotopes in Southern Ocean surface waters. Global Biogeochem. Cycles 18, 1–8. doi: 10.1029/2003GB002140
Wetzel, F., de Souza, G. F., and Reynolds, B. C. (2014). What controls silicon isotope fractionation during dissolution of diatom opal? Geochim. Cosmochim. Acta 131, 128–137. doi: 10.1016/j.gca.2014.01.028
Wheat, C. G., and McManus, J. (2005). The potential role of ridge-flank hydrothermal systems on oceanic germanium and silicon balances. Geochim. Cosmochim. Acta 69, 2021–2029. doi: 10.1016/j.gca.2004.05.046
White, A. F., and Blum, A. E. (1995). Effects of climate on chemical weathering in watersheds. Geochim. Cosmochim. Acta 59, 1729–1747. doi: 10.1016/0016-7037(95)00078-E
Wiederhold, J. G. (2015). Metal stable isotope signatures as tracers in environmental geochemistry. Environ. Sci. Technol. 49, 2606–2624. doi: 10.1021/es504683e
Wille, M., Sutton, J., Ellwood, M. J., Sambridge, M., Maher, W., Eggins, S., et al. (2010). Silicon isotopic fractionation in marine sponges: a new model for understanding silicon isotopic variations in sponges. Earth Planet. Sci. Lett. 292, 281–289. doi: 10.1016/j.epsl.2010.01.036
Wilkinson, C. R., and Garrone, R. (1980). Ultrastructure of siliceous spicules and microsclerocytes in the marine sponge Neofibularia irata N. SP. J. Morphol. 166, 51–63. doi: 10.1002/jmor.1051660105
Wilson, K. E., Leng, M. J., and Mackay, A. W. (2014). The use of multivariate statistics to resolve multiple contamination signals in the oxygen isotope analysis of biogenic silica. J. Quat. Sci. 29, 641–649. doi: 10.1002/jqs.2729
Wollast, R., and Mackenzie, F. T. (1989). “Global Biogeochemical Cycles and Climate,” in Climate and Geo-Sciences: A Challenge for Science and Society in the 21st Century, eds A. Berger, S. Schneider, and J. C. Duplessy (Dordrecht: Springer), 453–473.
Xiong, Z., Li, T., Algeo, T., Doering, K., Frank, M., Brzezinski, M. A., et al. (2015). The silicon isotope composition of Ethmodiscus rex laminated diatom mats from the tropical West Pacific: implications for silicate cycling during the last glacial maximum. Paleoceanography 30, 803–823. doi: 10.1002/2015PA002793
Zachos, J. (2001). Trends, rhythms, and aberrations in global climate 65 Ma to Present. Science 292, 686–693. doi: 10.1126/science.1059412
Zhao, Y., Vance, D., Abouchami, W., and De Baar, H. J. W. (2014). Biogeochemical cycling of zinc and its isotopes in the Southern Ocean. Geochim. Cosmochim. Acta 125, 653–672. doi: 10.1016/j.gca.2013.07.045
Keywords: C – N – O - Si isotopes, biogenic silica, element/Si ratios, biogeochemical cycles, silicon
Citation: Sutton JN, André L, Cardinal D, Conley DJ, de Souza GF, Dean J, Dodd J, Ehlert C, Ellwood MJ, Frings PJ, Grasse P, Hendry K, Leng MJ, Michalopoulos P, Panizzo VN and Swann GEA (2018) A Review of the Stable Isotope Bio-geochemistry of the Global Silicon Cycle and Its Associated Trace Elements. Front. Earth Sci. 5:112. doi: 10.3389/feart.2017.00112
Received: 29 May 2017; Accepted: 21 December 2017;
Published: 30 January 2018.
Edited by:
Brivaela Moriceau, Centre National de la Recherche Scientifique (CNRS), FranceReviewed by:
Peter Croot, National University of Ireland Galway, IrelandAnne-Julie Cavagna, Vrije Universiteit Brussel, Belgium
Copyright © 2018 Sutton, André, Cardinal, Conley, de Souza, Dean, Dodd, Ehlert, Ellwood, Frings, Grasse, Hendry, Leng, Michalopoulos, Panizzo and Swann. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jill N. Sutton, amlsbC5zdXR0b25AdW5pdi1icmVzdC5mcg==